Embed Size (px)
Citation preview

of February 16, 2018.This information is current as
Lineagec-Fms and Diverts the Cells to Dendritic CellBone Resorption through Downregulation of IL-3 Inhibits Human Osteoclastogenesis and
Pote, Gyan C. Mishra and Mohan R. WaniTomar, Rupesh K. Srivastava, Supinder Kour, Satish T. Navita Gupta, Amruta P. Barhanpurkar, Geetanjali B.
ol.1000015http://www.jimmunol.org/content/early/2010/07/19/jimmun
published online 19 July 2010J Immunol
MaterialSupplementary
5.DC1http://www.jimmunol.org/content/suppl/2010/07/20/jimmunol.100001
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. All rights reserved.1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on February 16, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on February 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

The Journal of Immunology
IL-3 Inhibits Human Osteoclastogenesis and Bone Resorptionthrough Downregulation of c-Fms and Diverts the Cells toDendritic Cell Lineage
Navita Gupta, Amruta P. Barhanpurkar, Geetanjali B. Tomar, Rupesh K. Srivastava,
Supinder Kour, Satish T. Pote, Gyan C. Mishra, and Mohan R. Wani
IL-3 is an important cytokine that regulates hematopoiesis and functions as a link between the immune and the hematopoietic system.
In this study, we investigated the role andmechanism of IL-3 action on human osteoclast formation and bone resorption using PBMCs.
PBMCs differentiate into functional osteoclasts in the presence of M-CSF and receptor activator of NF-kB ligand as evaluated by 23c6
expression and bone resorption.We found that IL-3 dose-dependently inhibited formation of 23c6-positive osteoclasts, bone resorption
and C-terminal telopeptide of type I collagen, a collagen degradation product. The inhibitory effect of IL-3 on bone resorption was
irreversible. To investigate the mechanism of IL-3 action, we analyzed the effect of IL-3 on the receptor activator of NF-kB and c-Fms
receptors and c-Fos, PU.1, NFAT cytoplasmic 1, and RelB transcription factors essential for osteoclastogenesis. IL-3 significantly
inhibited c-Fms and downregulated both PU.1 and c-Fos at both mRNA and protein level. Furthermore, IL-3–treated cells showed
increased expression of dendritic cell markers CD1a and CD80 and decreased expression of monocyte/macrophage marker CD14.
Interestingly, IL-3 inhibited formation of human osteoclasts derived from blood monocytes and bone marrow cells of osteoporotic
individuals. Thus, IL-3 may have therapeutic potential as an antiosteolytic agent in treatment of osteoporosis. The Journal of Immu-
nology, 2010, 185: 000–000.
The coordination between bone-forming osteoblasts andbone-resorbing osteoclasts is very crucial in maintainingthe bone homeostasis and structural integrity of the skele-
ton. Important bone diseases, such as osteoporosis, rheumatoid ar-thritis, Paget’s disease, and bone metastasis of breast and prostatecancers, are caused by increased osteoclast number and activity(1, 2). Osteoclasts, the multinuclear cells, differentiate from hema-topoietic precursors of the monocyte/macrophage lineage thatalso give rise to macrophages or dendritic cells (3, 4). Two criticalmolecules, M-CSF and receptor activator of NF-kB (RANK) ligand(RANKL) expressed by osteoblasts, are essential and sufficient forthe differentiation of hematopoietic cells to form functional osteo-clasts (5, 6). Like mouse osteoclasts, RANKL is a critical factor forhuman osteoclastogenesis, and in the presence of M-CSF, it inducesformation of 23c6-positive human osteoclasts and bone resorptionfrom adherent PBMCs, CD14+ monocytes, and nonadherent bone
marrow cells (7–10). The phenotypes of RANKL-induced humanosteoclasts are largely similar to those formed in vivo.M-CSF and RANKL promote osteoclastogenesis by acting through
their receptors c-Fms and RANK, respectively (11). M-CSF supportssurvival and proliferation of myeloid progenitors and promotes gen-eration of osteoclast precursors that express RANK (5, 11). RANKLby binding to RANK on osteoclast precursors activates downstreamsignaling pathways that control osteoclast differentiation and boneresorption. M-CSF as well as RANKL-deficient mice show severeosteopetrosis (5, 6, 12). In addition to these molecules, several tran-scription factors, such as NF-kB, NFAT cytoplasmic 1 (NFATc1),micropthalmia transcription factor (MITF), PU.1, and c-Fos, are vitalfor osteoclast formation (13–15). PU.1 is essential for development ofmyeloid cells. PU.1- and MITF-deficient mice show a failure in mac-rophage differentiation (15, 16) and inhibition of osteoclastogenesis atearly stage of differentiation (13). Mice deficient in c-Fos exhibitosteopetrosis because of osteoclast differentiation defect, whereasthe number of macrophages increases, indicating that c-Fos acts afterPU.1 activation in osteoclast differentiation (14). Also, mice deficientin NFATc1 and NF-kB (RelB) are osteopetrotic despite normal orincreased number of monocytes/macrophages (15).Osteoclastogenesis is regulated by immune cell-derived cytokines
(1, 17). IL-3, a cytokine secreted by Th cells, stimulates proliferation,differentiation, and survival of pluripotent hematopoietic stem cells(18–20). The receptor for IL-3 consists of a heterodimer of the IL-3–specific a-chain and a common b-chain that is shared with GM-CSFand IL-5 (21). We have previously demonstrated that IL-3 inhibitsRANKL (22) and TNF-a–induced osteoclast differentiation inpurified mouse osteoclast precursors (23). Recently, we have alsodemonstrated that IL-3 potently and irreversibly inhibits TNF-a–induced bone resorption in vitro and prevents development ofinflammatory arthritis as well as cartilage and bone loss in mice(24). These results indicated the potent inhibitory nature of IL-3 onmouse osteoclast differentiation. However, the role of IL-3 in humanosteoclast differentiation and bone resorption is still unknown.
National Center for Cell Science, University of Pune Campus, Pune, India
Received for publication January 5, 2010. Accepted for publication June 16, 2010.
This work was supported by the Department of Biotechnology, Government of India.N.G. and G.T. received Senior Research Fellowships from the University Grant Com-mission (New Delhi, India). A.P.B. received a Senior Research Fellowship from theIndian Council of Medical Research (New Delhi, India). R.K.S. is the recipient ofa Senior Research Fellowship from the Council for Scientific and Industrial Research(New Delhi, India).
Address correspondence and reprint requests to Dr. Mohan R. Wani, National Centerfor Cell Science, University of Pune Campus, Pune 411 007, India. E-mail address:[email protected]
The online version of this article contains supplemental material.
Abbreviations used in this paper: BMD, bone mineral density; CAII, carbonic anhy-drase II; CT, threshold cycle; CTR, calcitonin receptor; CTX-I, C-terminal telopeptideof type I collagen; FRA-1, Fos related Ag-1; MITF, micropthalmia transcriptionfactor; NFATc1, NFAT cytoplasmic 1; PI, propidium iodide; RANK, receptoractivator of NF-kB; RANKL, receptor activator of NF-kB ligand.
Copyright� 2010 by The American Association of Immunologists, Inc. 0022-1767/10/$16.00
www.jimmunol.org/cgi/doi/10.4049/jimmunol.1000015
Published July 19, 2010, doi:10.4049/jimmunol.1000015 by guest on February 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

In this study, we investigated the role and mechanism(s) of IL-3action on human osteoclast differentiation and bone resorption in-duced by RANKL in peripheral blood monocyte cultures fromhealthy donors and patients with osteoporosis. We found thatIL-3 inhibited human osteoclast formation and bone resorptionby acting directly on early osteoclast precursors. This inhibitory ef-fect of IL-3 on osteoclast formation was through downregulationof c-Fms receptor and PU.1 and c-Fos transcription factors. IL-3–treated cells showed increased expression of dendritic cell markersCD1a and CD80 and decreased monocyte/macrophage differen-tiation marker CD14. Interestingly, IL-3 also inhibited osteoclastdifferentiation and bone resorption in human monocytes and bonemarrow cultures of osteoporotic individuals. Thus, we conclude thatIL-3, in addition to its effects on mouse osteoclastogenesis, is a po-tent inhibitor of human osteoclast formation and bone resorption andmay act as a therapeutic antiresorptive agent in skeletal diseases,such as osteoporosis and metastatic bone disease.
Materials and MethodsSample collections
Human blood and bone marrow of healthy (age, 20–35 y) and clinicallyproven osteoporotic patients (age, 55–65 y) were obtained from localhospitals in Pune, India, after approval of the institutional ethics com-mittee. Informed consent was obtained from all the individuals. In someexperiments, the buffy coat used was obtained from blood transfusiondepartment of a local medical college.
Isolation of human osteoclast precursors and in vitro osteoclastdifferentiation and bone resorption
As model systems of human osteoclastogenesis, we used adherent humanPBMCs, CD14+ cells purified from PBMCs, and nonadherent human bonemarrow cells. All these cells differentiate into osteoclast-like cells in thepresence of RANKL and M-CSF (7–10). Briefly, human PBMCs were sep-arated by centrifugation over Ficoll-Paque (Sigma-Aldrich, St. Louis, MO)and seeded in 96-well plates on bone slices at a density of 5 3 105 cells/well or in 25-cm2 culture flasks at a density of 1.5 3 107 cells/flask. Afterincubation for 2 h, nonadherent cells were discarded, loosely adherent cellswere removed by thorough washing, and remaining adherent cells were usedas osteoclast precursors. These adherent PBMCs showed .95% expressionof CD14 by flow cytometry (Fig. 1A). Alternatively, CD14+ cells werepurified using negative selection kit (StemCell Technologies, Vancouver,British Columbia, Canada) and plated on bone slices in 96-well plates ata density of 5 3 104 cells/well. Nonadherent bone marrow cells were re-moved from 3-d-old culture of human bone marrow cells and plated ata density of 23 105 cells/well on bone slices in a 96-well plate. All cultureswere maintained in a-MEM (Sigma-Aldrich) containing 10% FBS (LifeTechnologies, Rockville, MD). For osteoclastic differentiation and boneresorption, cells were incubated for 21 d in the presence of human M-CSF(25 ng/ml; R&D Systems,Minneapolis, MN) and humanRANKL (40 ng/ml;Insight Biotechnology, Wembley, U.K.). When indicated, cultures were trea-ted with recombinant human IL-3 (R&D Systems). IL-3 was added to thecultures simultaneously with RANKL plus M-CSF on day 0 or when in-dicated, after 7 and 14 d. Cultures were fed every third day by replacing halfculture medium with an equal quantity of fresh medium and cytokines.
Bone resorption assay
After 21 d, bone slices were immersed in 4% sodiumhypochlorite for 15minand washed thoroughly to remove the cells. After drying, bone slices weremounted onto glass slides and then sputter coated (SC500 sputter coater;Emscope Laboratories, Ashford, U.K.) with gold. Bone slices were exam-ined by reflected light microscopy, and bone resorption was quantified usingan eyepiece graticule.
MTT assay
Adherent PBMCs were cultured in 96-well plates in the presence of differ-ent concentrations of IL-3 alone or with or without M-CSF (25 ng/ml) andRANKL (40 ng/ml). Cultures were fed on day 3, and after day 7, mediumwas removed. MTT (0.5 mg/ml; Sigma-Aldrich) was added, and after 4 h,the MTT was replaced with 100 ml DMSO, the formazan crystals weredissolved, and absorbance was measured at 570 nm.
Flow cytometric analysis
Adherent cells were harvested from culture flasks using cell dissolution buffer(Life Technologies), and few remaining cells were dislodged by gentle scrapingon ice. Cell surface staining was performed by incubating∼105 cells in 100 mlPBS with the indicated Abs. Expression of monocyte/macrophage/dendriticcells markers was assessed using PE-conjugated anti–IL-3Ra (CD123; BDBiosciences, Mountain View, CA), c-Fms (Santa Cruz Biotechnology, SantaCruz,CA),RANK(Imgenex,SanDiego,CA),CD205,CD86,HLA-DR,CD1a,and CD80 (eBioscience, San Diego, CA), and FITC-conjugated CD44 (BDBiosciences), CD83, CD11c, andCD14 (eBioscience)mAbs. Nonspecific fluo-rescencewas assessed by incubation with relevant isotype-matched conjugatedmAbs. Flow cytometric analyses were performed by FACSCalibur (BD Bio-sciences).
Cytochemical staining
Staining for coexpression of CD14 and IL-3Ra was carried out usingprimary mouse anti-human CD14 (BD Biosciences) and rabbit anti-humanIL-3Ra (Santa Cruz Biotechnology) mAbs, followed by anti-mouse PE(BD Biosciences) and anti-rabbit FITC (Bangalore Genei, Bangalore, In-dia) secondary mAbs. For c-Fms and RANK coexpression, goat anti-human RANK (R&D Systems) and rat anti-human c-Fms (Santa CruzBiotechnology) were used as primary Abs, followed by rabbit anti-goatAlexa Fluor 647 (Molecular Probes, Eugene, OR) and rabbit anti-rat FITC(DakoCytomation, Glostrup, Denmark) as respective secondary Abs. Glasscoverslips were mounted using DAPI-containing mounting medium (SantaCruz Biotechnology). Similarly, 21-d-old osteoclast cultures were incu-bated with primary mouse anti-human 23c6 (a gift from Dr. M. Horton,University College London, London, U.K.) and phallodin FITC (Sigma-Aldrich) mAbs. Goat anti-mouse PE-cy5 (Molecular Probes) was used assecondary Ab, and nuclei were labeled using propidium iodide. The cover-slips were viewed with a Zeiss LSM 510 confocal microscope equippedwith argon and helium lasers (Zeiss, Oberkochen, Germany). Staining forPU.1 and c-Fos was carried out using primary rabbit anti-human PU.1(Santa Cruz Biotechnology) and c-Fos (Cell Signaling Technology, Bev-erly, MA) mAbs, followed by secondary goat anti-rabbit FITC Ab. Cellswere permeabilized using 0.1% Triton X-100.
Immunocytochemistry
After 21 d, cells on plastic were washed twice with PBS, fixed with freshlyprepared 3.7% paraformaldehyde, and blocked with 10% human serum(Sigma-Aldrich) (all steps were performed at 4˚C). Cells were then in-cubated overnight with mouse anti-human 23c6 Ab, and cells were washedwith PBS and labeled with anti-mouse HRP Ab (Bangalore Genei) andincubated with diaminobenzidine solution (Invitrogen, Carlsbad, CA).23c6-positive cells were stained dark brown.
CrossLaps ELISA
Bone resorption activity in cell culture supernatants was determined byquantifying the degradation products of collagen type I, C-terminal telopep-tide of type I collagen (CTX-I) (25), using CrossLaps ELISA kit (NordicBioscience Diagnostics, Herlev, Denmark). One-step ELISA was per-formed according to manufacturer’s instructions.
RNA extraction and analysis by RT-PCR and quantitativereal-time PCR
Expression of b3 integrin, carbonic anhydrase II (CAII), cathepsin K,calcitonin receptor (CTR), IL-3R, and b-actin was assessed by RT-PCRanalysis. The primer sequences and the PCR conditions used are summa-rized in Table I. RNA was isolated from IL-3–treated and control cellsusing the TRIzol reagent (Invitrogen). Total RNA was used for synthesisof cDNAs by reverse transcription (cDNA synthesis kit; Invitrogen). ThecDNAs were amplified using PCR for 30–35 cycles. Each cycle consistedof 30 s of denaturation at 94˚C and 30 s of annealing and 30 s of extensionat 72˚C. b-Actin was used as the internal control. Densitometry of bandswas performed using gene tool software (Syngene, Cambridge, U.K.) usingb-actin as a reference gene.
For real-time PCR, a 10-ml reaction mixture containing SYBR Green IQsupermix (Bio-Rad, Hercules, CA) and 10 pmol each of human GAPDH,c-Fms, c-Fos, RANK, PU.1, RelB, and NFATc1 primers were used. PCRwas set using Eppendorf Realplex (Eppendorf, Hamburg, Germany). Theamplification was performed using 1 cycle of 95˚C for 3 min and 40 cyclesof 94˚C for 30 s, 55–58˚C for 20–25 s, and 72˚C for 30 s, followed bymelt curve analysis. The primer sequences (Integrated DNATechnologies,Coralville, Iowa) and the PCR conditions used are summarized in Table I.
2 IL-3 HAS POTENTIAL AS AN ANTIOSTEOLYTIC AGENT
by guest on February 16, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

The changes in the threshold cycle (CT) values were calculated by theequation DCT = CT (target) 2 CT (endogenous control). The fold induction wascalculated as follows: fold induction = 22D(DCT). M-CSF group was takenas input.
Isolation of mature osteoclasts from giant cell tumors of bone
Osteoclastoma tissue was used as a positive control for osteoclast-specificprimer standardization. Mature human osteoclasts were isolated from osteo-clastoma tissue as described previously (26).
Statistical analysis of data
The results were evaluated by using ANOVAwith subsequent comparisonsby Student t test for paired or nonpaired data, as appropriate. Statisticalsignificance was defined as p# 0.05. Values are reported as mean 6 SEM.
ResultsEffect of IL-3 on number of human osteoclast precursorsexpressing IL-3Ra
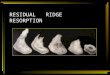
Adherent PBMCs were removed by cell dissociation buffer and an-alyzed for CD14 expression by flow cytometry. These cells showed.95% expression of CD14 and were referred to as osteoclast pre-cursors (Fig. 1A). We first examined the expression of IL-3Ra onosteoclast precursors by flow cytometry after incubating cellswith M-CSF (25 ng/ml) and RANKL (40 ng/ml) for 1, 3, 7, 14, and21 d. Human osteoclast precursors showed 89.9% expression ofIL-3Ra on day 1, 62.3% expression on day 3, and 16.4%expression on day 7. Mature osteoclasts on days 14 and 21 showedno expression of IL-3Ra (Fig. 1B). These cells also showed mRNAexpression of IL-3Ra on days 1 and 3 (Fig. 1C). By immuno-fluorescence, CD14+ cells showed coexpression of IL-3R on day 3(Fig. 1D). We then checked the effect of IL-3 on the number ofosteoclast precursors by MTT assay by incubating cells for 7 d withdifferent concentrations of IL-3 in the absence or presence ofM-CSFand RANKL. We found that IL-3 dose-dependently increased thenumber of osteoclast precursors in both the culture conditions (Fig.1E). The increase in viable cell number even at high concentrationsof IL-3 rules out its any toxic effect on osteoclast precursors.
IL-3 inhibits RANKL-induced human osteoclastogenesis
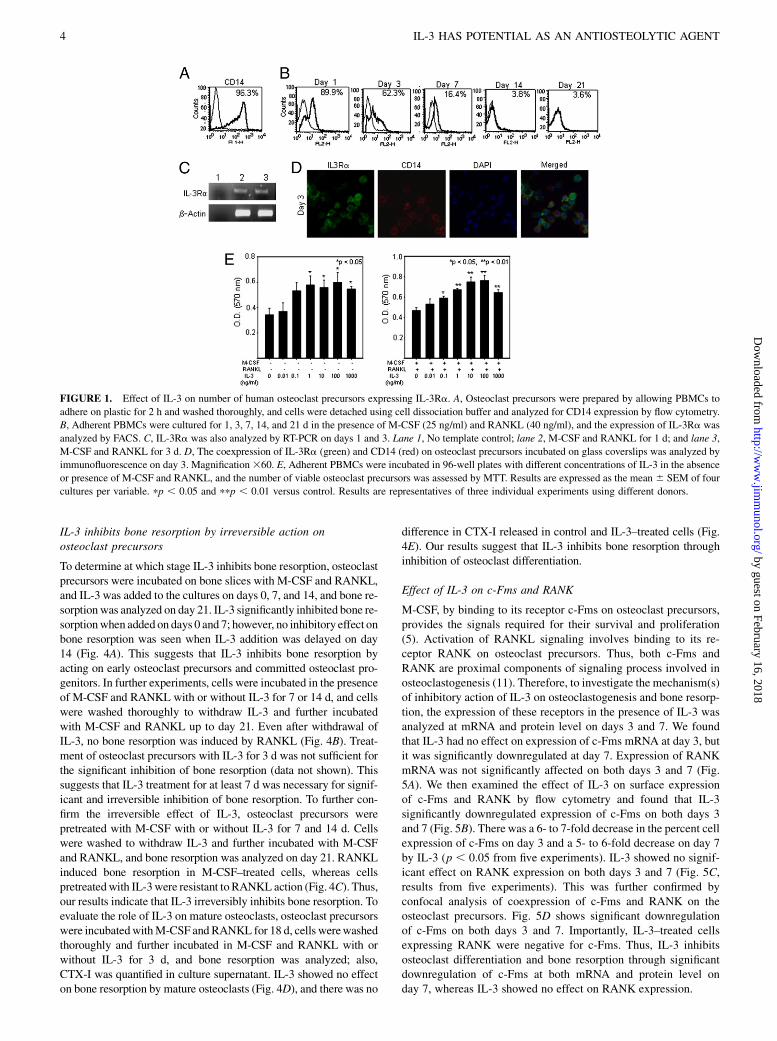
To investigate the role of IL-3 on osteoclast differentiation, osteo-clast precursors were incubated for 21 d with M-CSF and RANKLin the absence or presence of increasing concentrations of IL-3.RANKL induced the formation of multinucleated giant cells withruffled borders that were strongly positive for human avb3 integrin23c6, which stained dark brown. Photomicrographs in Fig. 2A showthe characteristic morphology of IL-3–treated cells at day 21. In thepresence of IL-3, the majority of cells were mononuclear, and fewmultinuclear cells were also present that were round, small in size,and without ruffled borders. IL-3 in a dose-dependent manner in-
hibited formation of multinuclear osteoclasts. The number of 23c6-positive cells decreased in the presence of IL-3, and significant in-hibition of osteoclast formation was seen from 0.1 ng/ml (Fig. 2B).This inhibition was further confirmed by immunofluorescence stain-ing of cells incubated in the presence of IL-3 (100 ng/ml) using 23c6Ab and phallodin F-actin. Cells treated with M-CSF and RANKLweremultinuclear and coexpressed 23c6 and prominent F-actin ring,whereas cells treated with IL-3 were mononuclear and stained onlyfor F-actin without a ring (Fig. 2C).We further analyzed the effect ofIL-3 on osteoclast-specific genes. Osteoclastoma and Saos-2 cellswere used as positive and negative controls, respectively, for osteo-clast genes. RANKL-induced human osteoclasts strongly expres-sed osteoclast-specific genes, such as integrin b3, CTR, cathepsinK, and CAII, which were downregulated by IL-3 (Fig. 2D). Theseresults suggest that IL-3 inhibits RANKL-induced human osteo-clast differentiation.
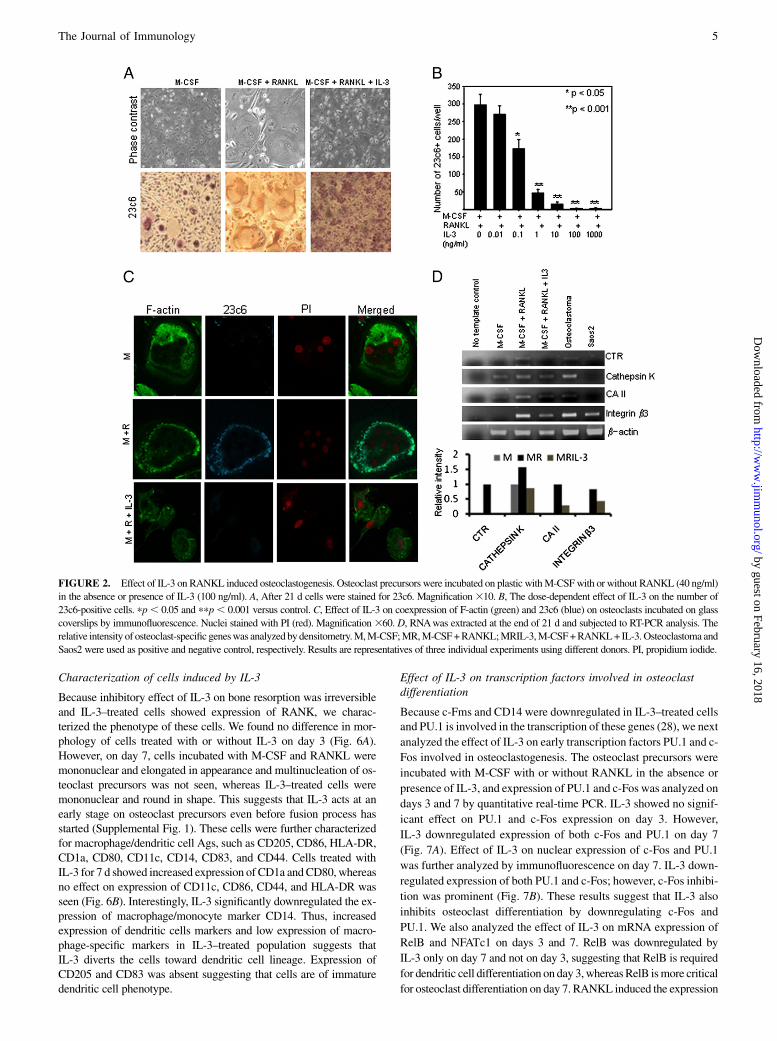
IL-3 inhibits bone resorption and collagen degradation productCTX-I
We further assessed whether the decrease in osteoclast differentia-tion observed in IL-3–treated cultures was accompanied witha decrease in resorptive capacity of these cells. For this osteoclast,precursors were incubated for 21 d on bone slices with M-CSF andRANKL in the absence or presence of different concentrations ofIL-3. RANKL induced formation of several resorption pits, andIL-3 in a dose-dependent manner inhibited RANKL-induced boneresorption (Fig. 3A). Significant inhibition of bone resorption wasseen from 1 ng/ml. We also used purified CD14+ cells, and IL-3dose-dependently inhibited bone resorption in these cells, too. Sig-nificant inhibition was seen from 0.1 ng/ml (Fig. 3B). Althoughthere was variation in percent bone resorption by PBMCs of differ-ent donors, we observed that IL-3 consistently inhibited bone re-sorption in all the donors. As reported previously by others (27), wealso observed that percent bone resorption in PBMCs of normalfemales was much less as compared with normal males (data notshown); therefore, for bone resorption studies, we used PBMCsderived from normal males. During bone resorption, type I collagenis degraded by osteoclasts, and a small degradation product,CTX-I, is released into the medium (25). The inhibitory effect ofIL-3 on bone resorptionwas further confirmed by assessing its effecton CTX-I. It was found that IL-3 decreases CTX-I levels in a dose-dependent manner (Fig. 3C). This quantitative assay confirms thatIL-3 inhibits RANKL-induced bone resorption. Because in most ofthe experiments complete inhibition of bone resorption by IL-3 wasseen at 100 ng/ml, we used this concentration in all further experi-ments. Photomicrographs in Fig. 3D shows the dose-dependent ef-fect of IL-3 on formation of resorption pits.
Table I. PCR primers
Gene Sense Primer Antisense Primer Annealing Temperature (˚C) Product Size (bp)
IL-3Ra 59-GGAGAATCTGACCTGCTGGA-39 59-ACTTTGAGAACCGCTGGAGA-39 56 217Integrin b3 59-GCAATGGGACCTTTGAGTGT-39 59-ACATGCAGGTGTCAGTACGC-39 55 354Cathepsin K 59-ACCGGGGTATTGACTCTGAA-39 59-GAGGTCAGGCTTGCATCAAT-39 58 190CTR 59-ACTGCTGGCTGAGTGTGGAAAC-39 59-GAAGCAGTAGATGGTCGCAAC-39 64 317CAII 59-CTGAAGCCCCTGTCTGTTTC-39 59-TCCATCAAGTGAACCCCAGT-39 54 177PU.1 59-CAGAAGACCTGGTGCCCTAT-39 59-GGAGCTCCGTGAAGTTGTTC-39 59 171c-Fms 59-CAGAGCCAGGACTATCAATGC-39 59-CTGGCTGAGCACACGATCT-39 56 170RANK 59-CGTAGACCACGATGATGTCG-39 59-GTCTGGAAGCTCCCCTGGT-39 60 122c-Fos 59-CTGGCGTTGTGAAGACCA T-39 59-TCCCTTCGGATTCCTTTT-39 60 109RelB 59-AGCATCCTTGGGGAGAGC-39 59-GAGGCCAGTCCTTCCACAC-39 60 121NFATc1 59-GTCCTGTCTGGCCACAACTT-39 59-GGTCAGTTTTCGCTTCCATC-39 58 100GAPDH 59-GAAGGTGAAGGTCGGAGT-39 59-GAAGATGGTGATGGGATTTC-39 60 180b-Actin 59-CGGGAAATCGTGCGTGACAT-39 59-ATCTTCATTGTGCTGGGTGCC-39 58 390
The Journal of Immunology 3
by guest on February 16, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
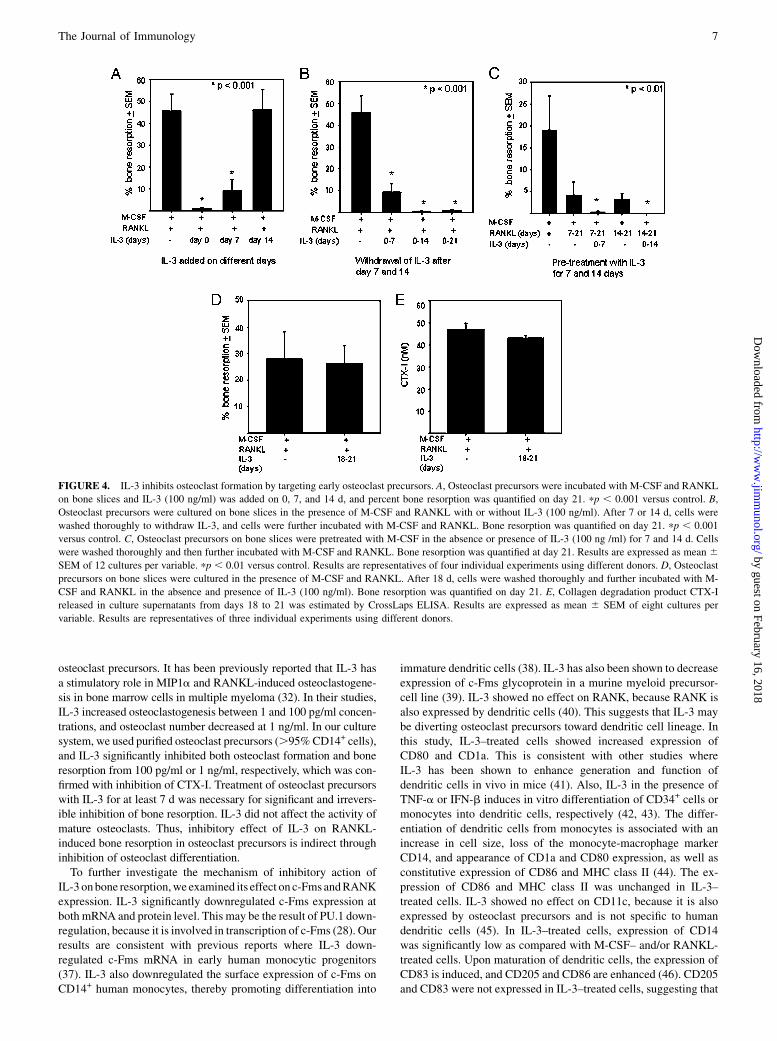
IL-3 inhibits bone resorption by irreversible action onosteoclast precursors
To determine at which stage IL-3 inhibits bone resorption, osteoclastprecursors were incubated on bone slices with M-CSF and RANKL,and IL-3 was added to the cultures on days 0, 7, and 14, and bone re-sorptionwas analyzed on day 21. IL-3 significantly inhibited bone re-sorptionwhen added on days 0 and 7; however, no inhibitory effect onbone resorption was seen when IL-3 addition was delayed on day14 (Fig. 4A). This suggests that IL-3 inhibits bone resorption byacting on early osteoclast precursors and committed osteoclast pro-genitors. In further experiments, cells were incubated in the presenceof M-CSF and RANKL with or without IL-3 for 7 or 14 d, and cellswere washed thoroughly to withdraw IL-3 and further incubatedwith M-CSF and RANKL up to day 21. Even after withdrawal ofIL-3, no bone resorption was induced by RANKL (Fig. 4B). Treat-ment of osteoclast precursors with IL-3 for 3 d was not sufficient forthe significant inhibition of bone resorption (data not shown). Thissuggests that IL-3 treatment for at least 7 d was necessary for signif-icant and irreversible inhibition of bone resorption. To further con-firm the irreversible effect of IL-3, osteoclast precursors werepretreated with M-CSF with or without IL-3 for 7 and 14 d. Cellswere washed to withdraw IL-3 and further incubated with M-CSFand RANKL, and bone resorption was analyzed on day 21. RANKLinduced bone resorption in M-CSF–treated cells, whereas cellspretreatedwith IL-3were resistant toRANKL action (Fig. 4C). Thus,our results indicate that IL-3 irreversibly inhibits bone resorption. Toevaluate the role of IL-3 on mature osteoclasts, osteoclast precursorswere incubatedwithM-CSF and RANKL for 18 d, cells werewashedthoroughly and further incubated in M-CSF and RANKL with orwithout IL-3 for 3 d, and bone resorption was analyzed; also,CTX-I was quantified in culture supernatant. IL-3 showed no effecton bone resorption by mature osteoclasts (Fig. 4D), and there was no
difference in CTX-I released in control and IL-3–treated cells (Fig.4E). Our results suggest that IL-3 inhibits bone resorption throughinhibition of osteoclast differentiation.
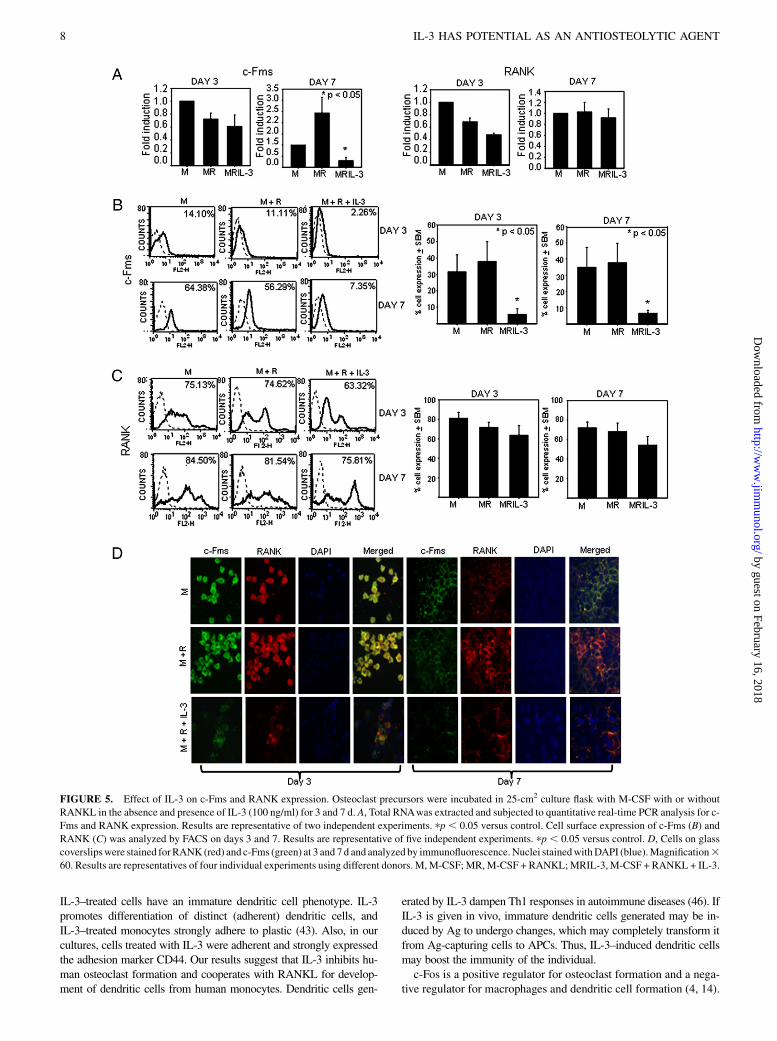
Effect of IL-3 on c-Fms and RANK
M-CSF, by binding to its receptor c-Fms on osteoclast precursors,provides the signals required for their survival and proliferation(5). Activation of RANKL signaling involves binding to its re-ceptor RANK on osteoclast precursors. Thus, both c-Fms andRANK are proximal components of signaling process involved inosteoclastogenesis (11). Therefore, to investigate the mechanism(s)of inhibitory action of IL-3 on osteoclastogenesis and bone resorp-tion, the expression of these receptors in the presence of IL-3 wasanalyzed at mRNA and protein level on days 3 and 7. We foundthat IL-3 had no effect on expression of c-Fms mRNA at day 3, butit was significantly downregulated at day 7. Expression of RANKmRNA was not significantly affected on both days 3 and 7 (Fig.5A). We then examined the effect of IL-3 on surface expressionof c-Fms and RANK by flow cytometry and found that IL-3significantly downregulated expression of c-Fms on both days 3and 7 (Fig. 5B). There was a 6- to 7-fold decrease in the percent cellexpression of c-Fms on day 3 and a 5- to 6-fold decrease on day 7by IL-3 (p , 0.05 from five experiments). IL-3 showed no signif-icant effect on RANK expression on both days 3 and 7 (Fig. 5C,results from five experiments). This was further confirmed byconfocal analysis of coexpression of c-Fms and RANK on theosteoclast precursors. Fig. 5D shows significant downregulationof c-Fms on both days 3 and 7. Importantly, IL-3–treated cellsexpressing RANK were negative for c-Fms. Thus, IL-3 inhibitsosteoclast differentiation and bone resorption through significantdownregulation of c-Fms at both mRNA and protein level onday 7, whereas IL-3 showed no effect on RANK expression.
FIGURE 1. Effect of IL-3 on number of human osteoclast precursors expressing IL-3Ra. A, Osteoclast precursors were prepared by allowing PBMCs to
adhere on plastic for 2 h and washed thoroughly, and cells were detached using cell dissociation buffer and analyzed for CD14 expression by flow cytometry.
B, Adherent PBMCs were cultured for 1, 3, 7, 14, and 21 d in the presence of M-CSF (25 ng/ml) and RANKL (40 ng/ml), and the expression of IL-3Ra was
analyzed by FACS. C, IL-3Ra was also analyzed by RT-PCR on days 1 and 3. Lane 1, No template control; lane 2, M-CSF and RANKL for 1 d; and lane 3,
M-CSF and RANKL for 3 d. D, The coexpression of IL-3Ra (green) and CD14 (red) on osteoclast precursors incubated on glass coverslips was analyzed by
immunofluorescence on day 3. Magnification360. E, Adherent PBMCs were incubated in 96-well plates with different concentrations of IL-3 in the absence
or presence of M-CSF and RANKL, and the number of viable osteoclast precursors was assessed by MTT. Results are expressed as the mean 6 SEM of four
cultures per variable. pp , 0.05 and ppp , 0.01 versus control. Results are representatives of three individual experiments using different donors.
4 IL-3 HAS POTENTIAL AS AN ANTIOSTEOLYTIC AGENT
by guest on February 16, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
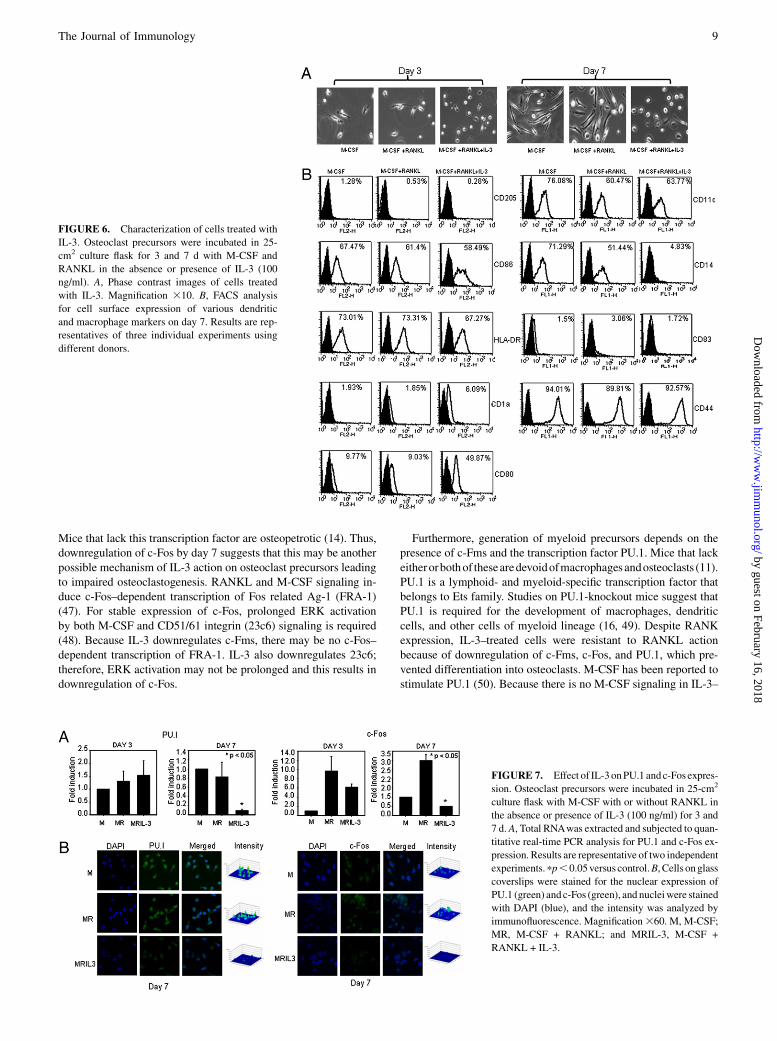
Characterization of cells induced by IL-3
Because inhibitory effect of IL-3 on bone resorption was irreversibleand IL-3–treated cells showed expression of RANK, we charac-terized the phenotype of these cells. We found no difference in mor-phology of cells treated with or without IL-3 on day 3 (Fig. 6A).However, on day 7, cells incubated with M-CSF and RANKL weremononuclear and elongated in appearance and multinucleation of os-teoclast precursors was not seen, whereas IL-3–treated cells weremononuclear and round in shape. This suggests that IL-3 acts at anearly stage on osteoclast precursors even before fusion process hasstarted (Supplemental Fig. 1). These cells were further characterizedfor macrophage/dendritic cell Ags, such as CD205, CD86, HLA-DR,CD1a, CD80, CD11c, CD14, CD83, and CD44. Cells treated withIL-3 for 7 d showed increased expression of CD1a andCD80, whereasno effect on expression of CD11c, CD86, CD44, and HLA-DR wasseen (Fig. 6B). Interestingly, IL-3 significantly downregulated the ex-pression of macrophage/monocyte marker CD14. Thus, increasedexpression of dendritic cells markers and low expression of macro-phage-specific markers in IL-3–treated population suggests thatIL-3 diverts the cells toward dendritic cell lineage. Expression ofCD205 and CD83 was absent suggesting that cells are of immaturedendritic cell phenotype.
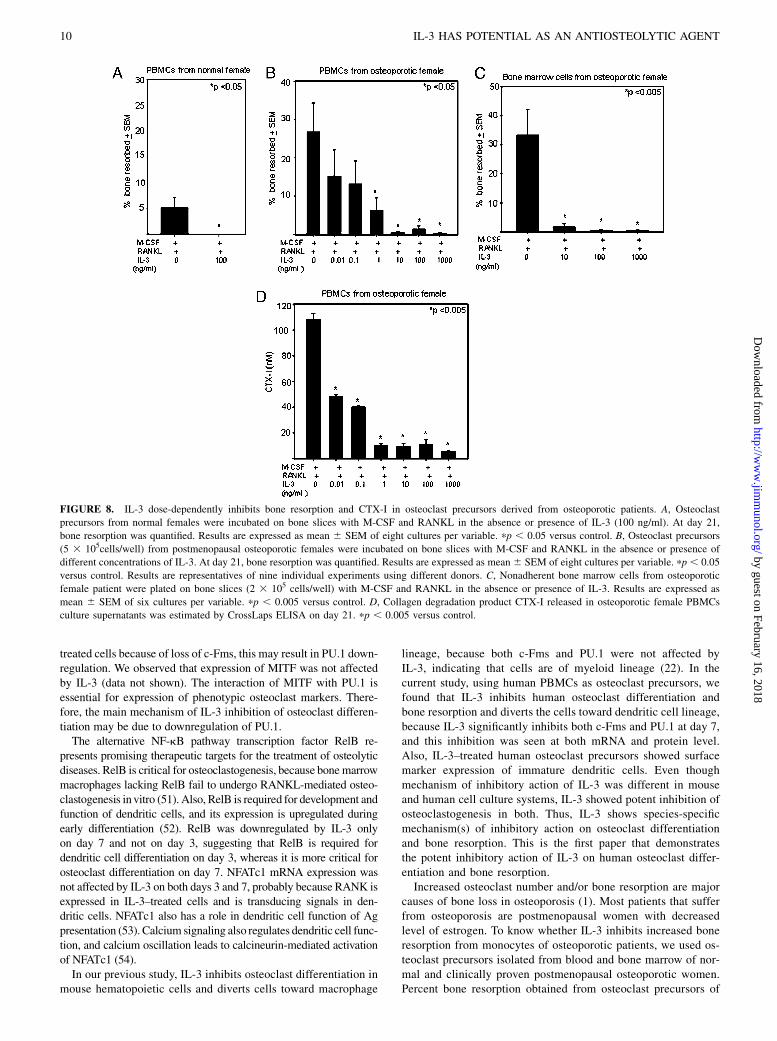
Effect of IL-3 on transcription factors involved in osteoclastdifferentiation
Because c-Fms and CD14 were downregulated in IL-3–treated cellsand PU.1 is involved in the transcription of these genes (28), we next
analyzed the effect of IL-3 on early transcription factors PU.1 and c-
Fos involved in osteoclastogenesis. The osteoclast precursors were
incubated with M-CSF with or without RANKL in the absence or
presence of IL-3, and expression of PU.1 and c-Fos was analyzed on
days 3 and 7 by quantitative real-time PCR. IL-3 showed no signif-
icant effect on PU.1 and c-Fos expression on day 3. However,
IL-3 downregulated expression of both c-Fos and PU.1 on day 7
(Fig. 7A). Effect of IL-3 on nuclear expression of c-Fos and PU.1
was further analyzed by immunofluorescence on day 7. IL-3 down-
regulated expression of both PU.1 and c-Fos; however, c-Fos inhibi-
tion was prominent (Fig. 7B). These results suggest that IL-3 also
inhibits osteoclast differentiation by downregulating c-Fos and
PU.1. We also analyzed the effect of IL-3 on mRNA expression of
RelB and NFATc1 on days 3 and 7. RelB was downregulated by
IL-3 only on day 7 and not on day 3, suggesting that RelB is required
for dendritic cell differentiation on day 3,whereasRelB ismore critical
for osteoclast differentiation on day 7. RANKL induced the expression
FIGURE 2. Effect of IL-3 on RANKL induced osteoclastogenesis. Osteoclast precursors were incubated on plastic withM-CSFwith or without RANKL (40 ng/ml)
in the absence or presence of IL-3 (100 ng/ml). A, After 21 d cells were stained for 23c6. Magnification310. B, The dose-dependent effect of IL-3 on the number of
23c6-positive cells. pp, 0.05 and ppp, 0.001 versus control. C, Effect of IL-3 on coexpression of F-actin (green) and 23c6 (blue) on osteoclasts incubated on glass
coverslips by immunofluorescence. Nuclei stained with PI (red). Magnification360. D, RNAwas extracted at the end of 21 d and subjected to RT-PCR analysis. The
relative intensity of osteoclast-specific geneswas analyzed bydensitometry.M,M-CSF;MR,M-CSF+RANKL;MRIL-3,M-CSF+RANKL+ IL-3.Osteoclastoma and
Saos2 were used as positive and negative control, respectively. Results are representatives of three individual experiments using different donors. PI, propidium iodide.
The Journal of Immunology 5
by guest on February 16, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
of NFATc1 on both days 3 and 7. However, there was no effect ofIL-3 on NFATc1 mRNA expression (Supplemental Fig. 2A, 2B).
IL-3 inhibits bone resorption in osteoclast precursors derivedfrom osteoporotic patients
Our results indicated that IL-3 is a potent inhibitor of bone resorp-tion. Therefore, in further investigation, we examined whetherIL-3 inhibits bone resorption in osteoclast precursors isolatedfrom osteoporotic patients. Osteoclast precursors obtained fromnormal and postmenopausal osteoporotic females were used to an-alyze the effect of IL-3 on bone resorption. As mentioned earlier,few bone resorption pits were observed in M-CSF– and RANKL(∼5%)-treated cells from normal females, and no bone resorptionwas seen in IL-3–treated cells (Fig. 8A). Monocytes obtained fromnormal females do not give very high bone resorption values ascompared with males or post menopausal osteoporotic femalesbecause of increased secretion of TNF-a and IL-1 by monocytesof postmenopausal osteoporotic females (27, 29). In osteoclast pre-cursors of postmenopausal osteoporotic females, there was ∼35%bone resorption in control, and IL-3 dose-dependently inhibited boneresorption, and significant inhibition of bone resorption was seenfrom 1 ng/ml (Fig. 8B). We also evaluated the effect of IL-3 on boneresorption using osteoclast precursors isolated from bone marrowof osteoporotic females. In control cultures there was ∼45% boneresorption, and IL-3 dose-dependently inhibited bone resorption inthis culture system also, and significant effect was seen from 10ng/ml (Fig. 8C). This inhibitory effect of IL-3 on bone resorptionwas further confirmed by assessing its effect on CTX-I. IL-3decreases CTX-I levels in a dose-dependent manner in osteoporo-tic PBMCs, and significant effect was seen even at 0.01 ng/ml(Fig. 8D). All these results suggest that IL-3 is a potent inhibitor ofbone resorption in clinical conditions also and may serve as a poten-tial antiosteolytic agent to treat osteoporosis and other diseases ofskeletal system.
DiscussionCytokines play an important role in the etiology and pathogenesisof bone-related diseases (1). Our previous studies in mouse
hematopoietic cells suggested that IL-3 is a potent inhibitor of
osteoclast differentiation and has potential to prevent pathological
bone resorption and inflammatory arthritis in mice (22–24). How-
ever, the role of IL-3 in human osteoclast formation and bone
resorption is not known. IL-3 has been shown to induce human
osteoclast differentiation (30–32). These effects of IL-3 on human
osteoclastogenesis are reported in the culture systems that consist
of a heterogeneous population of cells with different culture con-
ditions. In this study, we investigated the role and mechanism(s)
of IL-3 action on human osteoclast differentiation and bone re-
sorption using CD14+ (.95% pure) monocytes. IL-3 exerts its
biological activities by binding to specific high-affinity receptors,
which are expressed by human hematopoietic stem cells, endothe-
lial cells, and monocytes, and its expression is donor dependent
(33, 34). We report in this paper for the first time that monocyte-
derived osteoclast precursors express IL-3Ra at both mRNA and
protein level, and its expression decreases when cells move toward
maturation. IL-3 dose-dependently increased the number of oste-
oclast precursors and also suggested that high concentration of
IL-3 is not toxic to osteoclast precursors. In our culture system,
IL-3 inhibited RANKL-induced osteoclast differentiation by di-
rectly acting on osteoclast precursors. Tartrate-resistant acid phos-
phatase is a marker of osteoclast but is not specific for human
osteoclasts (35). Therefore, we determined the expression of 23c6,
an epitope of the vitronectin receptor (CD51/61), which is specif-
ically expressed by human osteoclasts (36). IL-3 significantly de-
creased the number of 23c6-positive cells and also inhibited
expression of osteoclast-specific genes, such as CTR, CAII, cathep-
sin K, and integrin b3. Importantly, IL-3 inhibited RANKL-induced
bone resorption and bone degradation product CTX-I in human
FIGURE 3. Effect of IL-3 on RANKL-induced bone resorption and CTX-I. Osteoclast precursors (5 3 105 cells/well) (A) or purified CD14+ cells (5 3104 cells/well) (B) were incubated on bone slices in 96-well plates with M-CSF and RANKL in the absence or presence of different concentration of IL-3.
After day 21, percent bone resorption was quantified. Results are expressed as mean 6 SEM of eight cultures per variable. pp , 0.001 and ppp , 0.05
versus control. Results are representatives of six individual experiments using different donors. C, Collagen degradation product CTX-I released in culture
supernatants at day 21 was estimated by CrossLaps ELISA. Results are expressed as mean 6 SEM of eight cultures per variable. pp , 0.05 versus control.
Results are representatives of two individual experiments using different donors. D, Phase contrast images of resorption pits at different concentrations of
IL-3. Magnification 310.
6 IL-3 HAS POTENTIAL AS AN ANTIOSTEOLYTIC AGENT
by guest on February 16, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
osteoclast precursors. It has been previously reported that IL-3 hasa stimulatory role in MIP1a and RANKL-induced osteoclastogene-sis in bone marrow cells in multiple myeloma (32). In their studies,IL-3 increased osteoclastogenesis between 1 and 100 pg/ml concen-trations, and osteoclast number decreased at 1 ng/ml. In our culturesystem, we used purified osteoclast precursors (.95%CD14+ cells),and IL-3 significantly inhibited both osteoclast formation and boneresorption from 100 pg/ml or 1 ng/ml, respectively, which was con-firmed with inhibition of CTX-I. Treatment of osteoclast precursorswith IL-3 for at least 7 d was necessary for significant and irrevers-ible inhibition of bone resorption. IL-3 did not affect the activity ofmature osteoclasts. Thus, inhibitory effect of IL-3 on RANKL-induced bone resorption in osteoclast precursors is indirect throughinhibition of osteoclast differentiation.To further investigate the mechanism of inhibitory action of
IL-3 onbone resorption,we examined its effect on c-Fms andRANKexpression. IL-3 significantly downregulated c-Fms expression atboth mRNA and protein level. This may be the result of PU.1 down-regulation, because it is involved in transcription of c-Fms (28). Ourresults are consistent with previous reports where IL-3 down-regulated c-Fms mRNA in early human monocytic progenitors(37). IL-3 also downregulated the surface expression of c-Fms onCD14+ human monocytes, thereby promoting differentiation into
immature dendritic cells (38). IL-3 has also been shown to decreaseexpression of c-Fms glycoprotein in a murine myeloid precursor-cell line (39). IL-3 showed no effect on RANK, because RANK isalso expressed by dendritic cells (40). This suggests that IL-3 maybe diverting osteoclast precursors toward dendritic cell lineage. Inthis study, IL-3–treated cells showed increased expression ofCD80 and CD1a. This is consistent with other studies whereIL-3 has been shown to enhance generation and function ofdendritic cells in vivo in mice (41). Also, IL-3 in the presence ofTNF-a or IFN-b induces in vitro differentiation of CD34+ cells ormonocytes into dendritic cells, respectively (42, 43). The differ-entiation of dendritic cells from monocytes is associated with anincrease in cell size, loss of the monocyte-macrophage markerCD14, and appearance of CD1a and CD80 expression, as well asconstitutive expression of CD86 and MHC class II (44). The ex-pression of CD86 and MHC class II was unchanged in IL-3–treated cells. IL-3 showed no effect on CD11c, because it is alsoexpressed by osteoclast precursors and is not specific to humandendritic cells (45). In IL-3–treated cells, expression of CD14was significantly low as compared with M-CSF– and/or RANKL-treated cells. Upon maturation of dendritic cells, the expression ofCD83 is induced, and CD205 and CD86 are enhanced (46). CD205and CD83 were not expressed in IL-3–treated cells, suggesting that
FIGURE 4. IL-3 inhibits osteoclast formation by targeting early osteoclast precursors. A, Osteoclast precursors were incubated with M-CSF and RANKL
on bone slices and IL-3 (100 ng/ml) was added on 0, 7, and 14 d, and percent bone resorption was quantified on day 21. pp , 0.001 versus control. B,
Osteoclast precursors were cultured on bone slices in the presence of M-CSF and RANKL with or without IL-3 (100 ng/ml). After 7 or 14 d, cells were
washed thoroughly to withdraw IL-3, and cells were further incubated with M-CSF and RANKL. Bone resorption was quantified on day 21. pp , 0.001
versus control. C, Osteoclast precursors on bone slices were pretreated with M-CSF in the absence or presence of IL-3 (100 ng /ml) for 7 and 14 d. Cells
were washed thoroughly and then further incubated with M-CSF and RANKL. Bone resorption was quantified at day 21. Results are expressed as mean 6SEM of 12 cultures per variable. pp , 0.01 versus control. Results are representatives of four individual experiments using different donors. D, Osteoclast
precursors on bone slices were cultured in the presence of M-CSF and RANKL. After 18 d, cells were washed thoroughly and further incubated with M-
CSF and RANKL in the absence and presence of IL-3 (100 ng/ml). Bone resorption was quantified on day 21. E, Collagen degradation product CTX-I
released in culture supernatants from days 18 to 21 was estimated by CrossLaps ELISA. Results are expressed as mean 6 SEM of eight cultures per
variable. Results are representatives of three individual experiments using different donors.
The Journal of Immunology 7
by guest on February 16, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
IL-3–treated cells have an immature dendritic cell phenotype. IL-3promotes differentiation of distinct (adherent) dendritic cells, andIL-3–treated monocytes strongly adhere to plastic (43). Also, in ourcultures, cells treated with IL-3 were adherent and strongly expressedthe adhesion marker CD44. Our results suggest that IL-3 inhibits hu-man osteoclast formation and cooperates with RANKL for develop-ment of dendritic cells from human monocytes. Dendritic cells gen-
erated by IL-3 dampen Th1 responses in autoimmune diseases (46). IfIL-3 is given in vivo, immature dendritic cells generated may be in-duced by Ag to undergo changes, which may completely transform itfrom Ag-capturing cells to APCs. Thus, IL-3–induced dendritic cellsmay boost the immunity of the individual.c-Fos is a positive regulator for osteoclast formation and a nega-
tive regulator for macrophages and dendritic cell formation (4, 14).
FIGURE 5. Effect of IL-3 on c-Fms and RANK expression. Osteoclast precursors were incubated in 25-cm2 culture flask with M-CSF with or without
RANKL in the absence and presence of IL-3 (100 ng/ml) for 3 and 7 d. A, Total RNAwas extracted and subjected to quantitative real-time PCR analysis for c-
Fms and RANK expression. Results are representative of two independent experiments. pp , 0.05 versus control. Cell surface expression of c-Fms (B) and
RANK (C) was analyzed by FACS on days 3 and 7. Results are representative of five independent experiments. pp , 0.05 versus control. D, Cells on glass
coverslipswere stained forRANK (red) and c-Fms (green) at 3 and 7 d and analyzed by immunofluorescence.Nuclei stainedwithDAPI (blue).Magnification360. Results are representatives of four individual experiments using different donors. M,M-CSF;MR,M-CSF + RANKL;MRIL-3, M-CSF + RANKL + IL-3.
8 IL-3 HAS POTENTIAL AS AN ANTIOSTEOLYTIC AGENT
by guest on February 16, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
Mice that lack this transcription factor are osteopetrotic (14). Thus,downregulation of c-Fos by day 7 suggests that this may be anotherpossible mechanism of IL-3 action on osteoclast precursors leadingto impaired osteoclastogenesis. RANKL and M-CSF signaling in-duce c-Fos–dependent transcription of Fos related Ag-1 (FRA-1)(47). For stable expression of c-Fos, prolonged ERK activationby both M-CSF and CD51/61 integrin (23c6) signaling is required(48). Because IL-3 downregulates c-Fms, there may be no c-Fos–dependent transcription of FRA-1. IL-3 also downregulates 23c6;therefore, ERK activation may not be prolonged and this results indownregulation of c-Fos.
Furthermore, generation of myeloid precursors depends on thepresence of c-Fms and the transcription factor PU.1. Mice that lackeitherorbothof thesearedevoidofmacrophagesandosteoclasts (11).PU.1 is a lymphoid- and myeloid-specific transcription factor thatbelongs to Ets family. Studies on PU.1-knockout mice suggest thatPU.1 is required for the development of macrophages, dendriticcells, and other cells of myeloid lineage (16, 49). Despite RANKexpression, IL-3–treated cells were resistant to RANKL actionbecause of downregulation of c-Fms, c-Fos, and PU.1, which pre-vented differentiation into osteoclasts. M-CSF has been reported tostimulate PU.1 (50). Because there is no M-CSF signaling in IL-3–
FIGURE 6. Characterization of cells treated with
IL-3. Osteoclast precursors were incubated in 25-
cm2 culture flask for 3 and 7 d with M-CSF and
RANKL in the absence or presence of IL-3 (100
ng/ml). A, Phase contrast images of cells treated
with IL-3. Magnification 310. B, FACS analysis
for cell surface expression of various dendritic
and macrophage markers on day 7. Results are rep-
resentatives of three individual experiments using
different donors.
FIGURE 7. Effect of IL-3onPU.1 andc-Fos expres-
sion. Osteoclast precursors were incubated in 25-cm2
culture flask with M-CSF with or without RANKL in
the absence or presence of IL-3 (100 ng/ml) for 3 and
7 d. A, Total RNAwas extracted and subjected to quan-
titative real-time PCR analysis for PU.1 and c-Fos ex-
pression. Results are representative of two independent
experiments.pp,0.05versus control.B, Cells onglass
coverslips were stained for the nuclear expression of
PU.1 (green) andc-Fos (green), and nucleiwere stained
with DAPI (blue), and the intensity was analyzed by
immunofluorescence. Magnification360. M, M-CSF;
MR, M-CSF + RANKL; and MRIL-3, M-CSF +
RANKL + IL-3.
The Journal of Immunology 9
by guest on February 16, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
treated cells because of loss of c-Fms, this may result in PU.1 down-regulation. We observed that expression of MITF was not affectedby IL-3 (data not shown). The interaction of MITF with PU.1 isessential for expression of phenotypic osteoclast markers. There-fore, the main mechanism of IL-3 inhibition of osteoclast differen-tiation may be due to downregulation of PU.1.The alternative NF-kB pathway transcription factor RelB re-
presents promising therapeutic targets for the treatment of osteolyticdiseases. RelB is critical for osteoclastogenesis, because bonemarrowmacrophages lacking RelB fail to undergo RANKL-mediated osteo-clastogenesis invitro (51). Also, RelB is required for development andfunction of dendritic cells, and its expression is upregulated duringearly differentiation (52). RelB was downregulated by IL-3 onlyon day 7 and not on day 3, suggesting that RelB is required fordendritic cell differentiation on day 3, whereas it is more critical forosteoclast differentiation on day 7. NFATc1 mRNA expression wasnot affected by IL-3 on both days 3 and 7, probably because RANK isexpressed in IL-3–treated cells and is transducing signals in den-dritic cells. NFATc1 also has a role in dendritic cell function of Agpresentation (53). Calcium signaling also regulates dendritic cell func-tion, and calcium oscillation leads to calcineurin-mediated activationof NFATc1 (54).In our previous study, IL-3 inhibits osteoclast differentiation in
mouse hematopoietic cells and diverts cells toward macrophage
lineage, because both c-Fms and PU.1 were not affected byIL-3, indicating that cells are of myeloid lineage (22). In thecurrent study, using human PBMCs as osteoclast precursors, wefound that IL-3 inhibits human osteoclast differentiation andbone resorption and diverts the cells toward dendritic cell lineage,because IL-3 significantly inhibits both c-Fms and PU.1 at day 7,and this inhibition was seen at both mRNA and protein level.Also, IL-3–treated human osteoclast precursors showed surfacemarker expression of immature dendritic cells. Even thoughmechanism of inhibitory action of IL-3 was different in mouseand human cell culture systems, IL-3 showed potent inhibition ofosteoclastogenesis in both. Thus, IL-3 shows species-specificmechanism(s) of inhibitory action on osteoclast differentiationand bone resorption. This is the first paper that demonstratesthe potent inhibitory action of IL-3 on human osteoclast differ-entiation and bone resorption.Increased osteoclast number and/or bone resorption are major
causes of bone loss in osteoporosis (1). Most patients that sufferfrom osteoporosis are postmenopausal women with decreasedlevel of estrogen. To know whether IL-3 inhibits increased boneresorption from monocytes of osteoporotic patients, we used os-teoclast precursors isolated from blood and bone marrow of nor-mal and clinically proven postmenopausal osteoporotic women.Percent bone resorption obtained from osteoclast precursors of
FIGURE 8. IL-3 dose-dependently inhibits bone resorption and CTX-I in osteoclast precursors derived from osteoporotic patients. A, Osteoclast
precursors from normal females were incubated on bone slices with M-CSF and RANKL in the absence or presence of IL-3 (100 ng/ml). At day 21,
bone resorption was quantified. Results are expressed as mean 6 SEM of eight cultures per variable. pp , 0.05 versus control. B, Osteoclast precursors
(5 3 105cells/well) from postmenopausal osteoporotic females were incubated on bone slices with M-CSF and RANKL in the absence or presence of
different concentrations of IL-3. At day 21, bone resorption was quantified. Results are expressed as mean 6 SEM of eight cultures per variable. pp , 0.05
versus control. Results are representatives of nine individual experiments using different donors. C, Nonadherent bone marrow cells from osteoporotic
female patient were plated on bone slices (2 3 105 cells/well) with M-CSF and RANKL in the absence or presence of IL-3. Results are expressed as
mean 6 SEM of six cultures per variable. pp , 0.005 versus control. D, Collagen degradation product CTX-I released in osteoporotic female PBMCs
culture supernatants was estimated by CrossLaps ELISA on day 21. pp , 0.005 versus control.
10 IL-3 HAS POTENTIAL AS AN ANTIOSTEOLYTIC AGENT
by guest on February 16, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

normal females was much less than that obtained from osteopo-rotic females, probably because of secretion of TNF-a and IL-1by osteoporotic monocytes, which are mediators of bone resorp-tion (27, 29). Indeed, IL-3 inhibited increased bone resorption inboth osteoclast precursors derived from both normal and osteo-porotic females. In conclusion, IL-3 may have therapeutic poten-tial to inhibit increased osteoclast number and bone resorption inosteoporosis.Aspirin has been shown to have a positive effect on bone mineral
density (BMD) in the aged population (55), but the exactmechanismof aspirin action is still not known. Aspirin has also been shown toinhibit the activity of the enzyme cyclooxygenase in macrophagesresulting in overproduction of leukotrienes, which are capable ofstimulating IL-3 production in vitro (56). Thus, we hypothesize thatthe beneficial effect of aspirin on BMD may be due to overproduc-tion of IL-3 by monocytes, which may result in increased BMDbecause of inhibition of osteoclastogenesis. Also, expression ofIL-3 has been suggested to be important in the etiology of osteo-porosis (57). They observed that mRNA for IL-3 was less fre-quently present in osteoblasts derived from osteoporotic bonecompared with nonosteoporotic age-matched subjects and nonos-teoporotic younger subjects. Low-dose aspirin induces IL-3 pro-duction by monocytes, which completely abrogate manifestationsof experimental antiphospholipid syndrome (58). Mice deficient inIL-3 and GM-CSF are more susceptible to autoimmune diseases(59). Since IL-3 is already in use in clinics to treat different statesof bone marrow failure and hematologic malignancies and tosupport engraftment after bone marrow transplantation (19), wesuggest that IL-3 may have therapeutic potential to act as an anti-osteolytic agent to reduce bone loss in osteoporosis and otherbone diseases.
AcknowledgmentsWe thank Hemangini Shikhare for FACS analysis and Ashwini Atre for
confocal microscopy. We also thank Dr. Mike Horton, University College
(London, England), for providing anti-human 23c6 Ab. We also thank Col.
Sarkar, Department of TrasfusionMedicine, Armed ForcesMedical College
(Pune, India) for providing buffy coat samples.
DisclosuresThe authors have no financial conflicts of interest.
References1. Rodan, G. A., and T. J. Martin. 2000. Therapeutic approaches to bone diseases.
Science 289: 1508–1514.2. Rivollier, A., M. Mazzorana, J. Tebib, M. Piperno, T. Aitsiselmi, C. Rabourdin-
Combe, P. Jurdic, and C. Servet-Delprat. 2004. Immature dendritic celltransdifferentiation into osteoclasts: a novel pathway sustained by therheumatoid arthritis microenvironment. Blood 104: 4029–4037.
3. Akagawa, K. S., N. Takasuka, Y. Nozaki, I. Komuro, M. Azuma, M. Ueda,M. Naito, and K. Takahashi. 1996. Generation of CD1+RelB+ dendritic cells andtartrate-resistant acid phosphatase-positive osteoclast-like multinucleatedgiant cells from human monocytes. Blood 88: 4029–4039.
4. Miyamoto, T., O. Ohneda, F. Arai, K. Iwamoto, S. Okada, K. Takagi,D. M. Anderson, and T. Suda. 2001. Bifurcation of osteoclasts and dendritic cellsfrom common progenitors. Blood 98: 2544–2554.
5. Tanaka, S., N. Takahashi, N. Udagawa, T. Tamura, T. Akatsu, E. R. Stanley,T. Kurokawa, and T. Suda. 1993. Macrophage colony-stimulating factor is in-dispensable for both proliferation and differentiation of osteoclast progenitors. J.Clin. Invest. 91: 257–263.
6. Kong, Y. Y., H. Yoshida, I. Sarosi, H. L. Tan, E. Timms, C. Capparelli,S. Morony, A. J. Oliveira-dos-Santos, G. Van, A. Itie, et al. 1999. OPGL is a keyregulator of osteoclastogenesis, lymphocyte development and lymph-node or-ganogenesis. Nature 397: 315–323.
7. Sarma, U., and A. M. Flanagan. 1996. Macrophage colony-stimulating factorinduces substantial osteoclast generation and bone resorption in human bonemarrow cultures. Blood 88: 2531–2540.
8. Fujikawa, Y., J. M. Quinn, A. Sabokbar, J. O. McGee, and N. A. Athanasou.1996. The human osteoclast precursor circulates in the monocyte fraction. En-docrinology 137: 4058–4060.
9. Nicholson, G. C., M. Malakellis, F. M. Collier, P. U. Cameron, W. R. Holloway,T. J. Gough, C. Gregorio-King, M. A. Kirkland, and D. E. Myers. 2000.Induction of osteoclasts from CD14-positive human peripheral bloodmononuclear cells by receptor activator of nuclear factor kB ligand (RANKL).Clin. Sci. 99: 133–140.
10. Mangashetti, L. S., S. M. Khapli, and M. R. Wani. 2005. IL-4 inhibits bone-resorbing activity of mature osteoclasts by affecting NF-kB and Ca2+ signaling.J. Immunol. 175: 917–925.
11. Arai, F., T. Miyamoto, O. Ohneda, T. Inada, T. Sudo, K. Brasel, T. Miyata,D. M. Anderson, and T. Suda. 1999. Commitment and differentiation of osteo-clast precursor cells by the sequential expression of c-Fms and receptor activatorof nuclear factor kB (RANK) receptors. J. Exp. Med. 190: 1741–1754.
12. Begg, S. K., J. M. Radley, J. W. Pollard, O. T. Chisholm, E. R. Stanley, andI. Bertoncello. 1993. Delayed hematopoietic development in osteopetrotic (op/op) mice. J. Exp. Med. 177: 237–242.
13. Tondravi, M. M., S. R. McKercher, K. Anderson, J. M. Erdmann, M. Quiroz,R. Maki, and S. L. Teitelbaum. 1997. Osteopetrosis in mice lacking haemato-poietic transcription factor PU.1. Nature 386: 81–84.
14. Grigoriadis, A. E., Z. Q. Wang, M. G. Cecchini, W. Hofstetter, R. Felix,H. A. Fleisch, and E. F. Wagner. 1994. c-Fos: a key regulator of osteoclast-macrophage lineage determination and bone remodeling. Science 266: 443–448.
15. Ivashkiv, L. B. 2009. Cross-regulation of signaling by ITAM-associated recep-tors. Nat. Immunol. 10: 340–347.
16. Anderson, K. L., K. A. Smith, H. Perkin, G. Hermanson, C. G. Anderson,D. J. Jolly, R. A. Maki, and B. E. Torbett. 1999. PU.1 and the granulocyte- andmacrophage colony-stimulating factor receptors play distinct roles in late-stagemyeloid cell differentiation. Blood 94: 2310–2318.
17. Lorenzo, J., M. Horowitz, and Y. Choi. 2008. Osteoimmunology: interactions ofthe bone and immune system. Endocr. Rev. 29: 403–440.
18. Niemeyer, C. M., C. A. Sieff, B. Mathey-Prevot, J. Z. Wimperis, B. E. Bierer,S. C. Clark, and D. G. Nathan. 1989. Expression of human interleukin-3 (multi-CSF) is restricted to human lymphocytes and T-cell tumor lines. Blood 73: 945–951.
19. Schrader, J. W. 2003. Interleukin-3. In The Cytokine Handbook, 2nd Ed., A. W.Thomson, and M. T. Lotze, eds. Academic, London, p. 201–225.
20. Bryder, D., and S. E. Jacobsen. 2000. Interleukin-3 supports expansion of long-term multilineage repopulating activity after multiple stem cell divisions in vitro.Blood 96: 1748–1755.
21. de Groot, R. P., P. J. Coffer, and L. Koenderman. 1998. Regulation of pro-liferation, differentiation and survival by the IL-3/IL-5/GM-CSF receptor family.Cell. Signal. 10: 619–628.
22. Khapli, S. M., L. S. Mangashetti, S. D. Yogesha, and M. R. Wani. 2003. IL-3 actsdirectly on osteoclast precursors and irreversibly inhibits receptor activator ofNF-kB ligand-induced osteoclast differentiation by diverting the cells to macro-phage lineage. J. Immunol. 171: 142–151.
23. Yogesha, S. D., S. M. Khapli, and M. R. Wani. 2005. Interleukin-3 and gran-ulocyte-macrophage colony-stimulating factor inhibits tumor necrosis factor(TNF)-a–induced osteoclast differentiation by downregulation of expression ofTNF receptors 1 and 2. J. Biol. Chem. 280: 11759–11769.
24. Yogesha, S. D., S. M. Khapli, R. K. Srivastava, L. S. Mangashetti, S. T. Pote,G. C. Mishra, and M. R. Wani. 2009. IL-3 inhibits TNF-a–induced boneresorption and prevents inflammatory arthritis. J. Immunol. 182: 361–370.
25. Kirstein, B., U. Grabowska, B. Samuelsson, M. Shiroo, T. J. Chambers, andK. Fuller. 2006. A novel assay for analysis of the regulation of the function ofhuman osteoclasts. J. Transl. Med. 4: 45.
26. Nesbitt, S. A., and M. A. Horton. 2003. Fluorescence imaging of bone-resorbingosteoclasts by confocal microscopy. Methods Mol. Med. 80: 259–281.
27. Jevon, M., A. Sabokbar, Y. Fujikawa, T. Hirayama, S. D. Neale, J. Wass, andN. A. Athanasou. 2002. Gender- and age-related differences in osteoclast for-mation from circulating precursors. J. Endocrinol. 172: 673–681.
28. Scott, E. W., M. C. Simon, J. Anastasi, and H. Singh. 1994. Requirement oftranscription factor PU.1 in the development of multiple hematopoietic lineages.Science 265: 1573–1577.
29. Pacifici, R. 1996. Estrogen, cytokines, and pathogenesis of postmenopausal os-teoporosis. J. Bone Miner. Res. 11: 1043–1051.
30. Enelow, R. I., G. W. Sullivan, H. T. Carper, and G. L. Mandell. 1992. Inductionof multinucleated giant cell formation from in vitro culture of human monocyteswith interleukin-3 and interferon-g: comparison with other stimulating factors.Am. J. Respir. Cell Mol. Biol. 6: 57–62.
31. Fujikawa, Y., A. Sabokbar, S. D. Neale, I. Itonaga, T. Torisu, and N. A. Athanasou.2001. The effect of macrophage-colony stimulating factor and other humoralfactors (interleukin-1, -3, -6, and -11, tumor necrosis factor-a, and granulocytemacrophage-colony stimulating factor) on human osteoclast formation fromcirculating cells. Bone 28: 261–267.
32. Lee, J.W.,H.Y. Chung, L.A. Ehrlich, D. F. Jelinek, N. S. Callander, G.D.Roodman,and S. J. Choi. 2004. IL-3 expression by myeloma cells increases both osteoclastformation and growth of myeloma cells. Blood 103: 2308–2315.
33. McKinstry, W. J., C. L. Li, J. E. Rasko, N. A. Nicola, G. R. Johnson, andD. Metcalf. 1997. Cytokine receptor expression on hematopoietic stem andprogenitor cells. Blood 89: 65–71.
34. Budel, L. M., O. Elbaz, H. Hoogerbrugge, R. Delwel, L. A. Mahmoud,B. Lowenberg, and I. P. Touw. 1990. Common binding structure for granulocytemacrophage colony-stimulating factor and interleukin-3 on human acute mye-loid leukemia cells and monocytes. Blood 75: 1439–1445.
35. Hayman, A. R. 2008. Tartrate-resistant acid phosphatase (TRAP) and theosteoclast/immune cell dichotomy. Autoimmunity 41: 218–223.
The Journal of Immunology 11
by guest on February 16, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

36. Horton, M. A. 1997. The alpha v beta 3 integrin “vitronectin receptor.” Int. J.Biochem. Cell Biol. 29: 721–725.
37. Panterne, B., A. Hatzfeld, P. Sansilvestri, A. Cardoso, M. N. Monier, P. Batard,and J. Hatzfeld. 1996. IL-3, GM-CSF and CSF-1 modulate c-fms mRNA morerapidly in human early monocytic progenitors than in mature or transformedmonocytic cells. J. Cell Sci. 109: 1795–1801.
38. Suzuki, H., N. Katayama, Y. Ikuta, K. Mukai, A. Fujieda, H. Mitani, H. Araki,H. Miyashita, N. Hoshino, H. Nishikawa, et al. 2004. Activities of granulocyte-macrophage colony-stimulating factor and interleukin-3 on monocytes. Am. J.Hematol. 75: 179–189.
39. Gliniak, B. C., and L. R. Rohrschneider. 1990. Expression of the M-CSF receptoris controlled posttranscriptionally by the dominant actions of GM-CSF or multi-CSF. Cell 63: 1073–1083.
40. Anderson, D. M., E. Maraskovsky, W. L. Billingsley, W. C. Dougall,M. E. Tometsko, E. R. Roux, M. C. Teepe, R. F. DuBose, D. Cosman, andL. Galibert. 1997. A homologue of the TNF receptor and its ligand enhance T-cell growth and dendritic-cell function. Nature 390: 175–179.
41. Storozynsky, E., J. G. Woodward, J. G. Frelinger, and E. M. Lord. 1999.Interleukin-3 and granulocyte-macrophage colony-stimulating factor enhancethe generation and function of dendritic cells. Immunology 97: 138–149.
42. Caux, C., B. Vanbervliet, C. Massacrier, I. Durand, and J. Banchereau. 1996.Interleukin-3 cooperates with tumor necrosis factor a for the development ofhuman dendritic/Langerhans cells from cord blood CD34+ hematopoieticprogenitor cells. Blood 87: 2376–2385.
43. Buelens, C., E. J. Bartholome, Z. Amraoui, M. Boutriaux, I. Salmon,K. Thielemans, F. Willems, and M. Goldman. 2002. Interleukin-3 and interferonb cooperate to induce differentiation of monocytes into dendritic cells withpotent helper T-cell stimulatory properties. Blood 99: 993–998.
44. Smyth, C. M., G. Logan, R. Boadle, P. B. Rowe, J. A. Smythe, andI. E. Alexander. 2005. Differential subcellular localization of CD86 in humanPBMC-derived macrophages and DCs, and ultrastructural characterization byimmuno-electron microscopy. Int. Immunol. 17: 123–132.
45. Lader, C. S., J. Scopes, M. A. Horton, and A. M. Flanagan. 2001. Generationof human osteoclasts in stromal cell-free and stromal cell-rich cultures: differ-ences in osteoclast CD11c/CD18 integrin expression. Br. J. Haematol. 112:430–437.
46. Ebner, S., S. Hofer, V. A. Nguyen, C. Furhapter, M. Herold, P. Fritsch,C. Heufler, and N. Romani. 2002. A novel role for IL-3: human monocytescultured in the presence of IL-3 and IL-4 differentiate into dendritic cells thatproduce less IL-12 and shift Th cell responses toward a Th2 cytokine pattern. J.Immunol. 168: 6199–6207.
47. Lerner, U. H. 2004. New molecules in the tumor necrosis factor ligand and re-ceptor superfamilies with importance for physiological and pathological boneresorption. Crit. Rev. Oral Biol. Med. 15: 64–81.
48. Murphy, L. O., S. Smith, R. H. Chen, D. C. Fingar, and J. Blenis. 2002. Mo-lecular interpretation of ERK signal duration by immediate early gene products.Nat. Cell Biol. 4: 556–564.
49. Guerriero, A., P. B. Langmuir, L. M. Spain, and E. W. Scott. 2000. PU.1 is requiredfor myeloid-derived but not lymphoid-derived dendritic cells. Blood 95: 879–885.
50. Yao, Z., P. Li, Q. Zhang, E. M. Schwarz, P. Keng, A. Arbini, B. F. Boyce, andL. Xing. 2006. Tumor necrosis factor-a increases circulating osteoclast precursornumbers by promoting their proliferation and differentiation in the bone marrowthrough upregulation of c-Fms expression. J. Biol. Chem. 281: 11846–11855.
51. Vaira, S., T. Johnson, A. C. Hirbe, M. Alhawagri, I. Anwisye, B. Sammut,J. O’Neal, W. Zou, K. N. Weilbaecher, R. Faccio, and D. V. Novack. 2008. RelBis the NF-kB subunit downstream of NIK responsible for osteoclastdifferentiation. Proc. Natl. Acad. Sci. USA 105: 3897–3902.
52. Cejas, P. J., L. M. Carlson, D. Kolonias, J. Zhang, I. Lindner, D. D. Billadeau,L. H. Boise, and K. P. Lee. 2005. Regulation of RelB expression during theinitiation of dendritic cell differentiation. Mol. Cell. Biol. 25: 7900–7916.
53. Connolly, S. F., and D. J. Kusner. 2007. The regulation of dendritic cell functionby calcium-signaling and its inhibition by microbial pathogens. Immunol. Res.39: 115–127.
54. Liu, K.-J., and C. L. Chu. 2006. Current progress in dendritic cell research. J.Cancer Mol. 2: 217–220.
55. Carbone, L. D., F. A. Tylavsky, J. A. Cauley, T. B. Harris, T. F. Lang, D. C. Bauer,K. D. Barrow, and S. B. Kritchevsky. 2003. Association between bone mineraldensity and the use of nonsteroidal anti-inflammatory drugs and aspirin: impact ofcyclooxygenase selectivity. J. Bone Miner. Res. 18: 1795–1802.
56. Fishman, P., E. Falach-Vaknin, B. Sredni, P. L. Meroni, C. Rudniki, andY. Shoenfeld. 1995. Aspirin modulates interleukin-3 production: additional ex-planation for the preventive effects of aspirin in antiphospholipid antibody syn-drome. J. Rheumatol. 22: 1086–1090.
57. Walsh, C. A., M. A. Birch, W. D. Fraser, A. F. Ginty, and J. A. Gallagher. 2000.Cytokine expression by cultured osteoblasts from patients with osteoporoticfractures. Int. J. Exp. Pathol. 81: 159–163.
58. Shoenfeld, Y., and P. Fishman. 1994. Role of IL-3 in the antiphospholipid syn-drome. Lupus 3: 259–261.
59. Enzler, T., S. Gillessen, J. P. Manis, D. Ferguson, J. Fleming, F. W. Alt,M. Mihm, and G. Dranoff. 2003. Deficiencies of GM-CSF and interferon g linkinflammation and cancer. J. Exp. Med. 197: 1213–1219.
12 IL-3 HAS POTENTIAL AS AN ANTIOSTEOLYTIC AGENT
by guest on February 16, 2018http://w
ww
.jimm
unol.org/D
ownloaded from








![Parathyroid Hormone: Is It Really the Cause for Increased ... · osteoclastogenesis, root resorption, and tooth eruption[5]. Others have discussed the anabolic and catabolic p o-tential](https://img.pdfslide.us/doc/110x75/5ea93e30692ebf09fd250b5e/parathyroid-hormone-is-it-really-the-cause-for-increased-osteoclastogenesis.jpg)