Embed Size (px)
Citation preview

of June 19, 2018.This information is current as
Mycobacterium tuberculosisAntibacterial Activity Against Virulent Macrophage-Like Cells with Increased IL-10 Converts Human Dendritic Cells into
Daniela Förtsch, Martin Röllinghoff and Steffen Stenger
http://www.jimmunol.org/content/165/2/978doi: 10.4049/jimmunol.165.2.978
2000; 165:978-987; ;J Immunol
Referenceshttp://www.jimmunol.org/content/165/2/978.full#ref-list-1
, 37 of which you can access for free at: cites 69 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2000 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on June 19, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on June 19, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

IL-10 Converts Human Dendritic Cells into Macrophage-LikeCells with Increased Antibacterial Activity Against VirulentMycobacterium tuberculosis1
Daniela Fortsch, Martin Rollinghoff, and Steffen Stenger2
Dendritic cells (DC) are unique in their ability to initiate a primary immune response by the presentation of soluble Ags to T cells.Recent studies have shown that DC also phagocytose particulate Ags including the intracellular pathogenMycobacterium tuber-culosis. However, it is not known whether DC contain the growth of intracellular organisms or allow unlimited replication. Toaddress this question, we infected human DC with a virulent strain ofM. tuberculosisand monitored the intracellular growth. Thebacteria grew two orders of magnitude within 7 days of culture. Among cytokines known to modulate mycobacterial growthparticularly in murine macrophages (TNF-a, IFN-g, TGF-b, IL-4), only IL-10 modulated the growth in human DC. This effectwas specific for immature dendritic cells, as IL-10 did not induce growth inhibition in human macrophages. In searching for themechanism of growth inhibition, we found that IL-10 induces the down-regulation of the DC marker CD1, while the macrophagemarker CD14 was up-regulated. Functionally, IL-10-treated cells had a reduced capacity to induce an alloresponse, but phagocyticuptake of M. tuberculosiswas more efficient. We also show that DC are inferior to macrophages in containing mycobacterialgrowth. These findings show that IL-10 converts DC into macrophage-like cells, thereby inducing the growth inhibition of anintracellular pathogen. At the site of a local immune response, such as a tuberculous granuloma, IL-10 might therefore participatein the composition of the cellular microenvironment by affecting the maturity and function of DC. The Journal of Immunology,2000, 165: 978–987.
Mycobacterium tuberculosis, the causative agent of tuber-culosis, is a facultative intracellular bacterium that hasevolved sophisticated mechanisms to allow its survival
inside host mononuclear phagocytes. AlthoughM. tuberculosiswas discovered by Koch over a century ago (1), the precise resis-tance mechanisms to this pathogen have not yet been resolved. Anefficient cellular immune response is critical for preventing thedevelopment of clinically overt tuberculosis (2). One major medi-ator of protective immunity is IFN-g, as mice or humans that lackcomponents of the IFN-g signaling pathway are highly susceptibleto mycobacterial infection (3). Nevertheless, in vitro studies thatinvolve human macrophages fail to demonstrate the activation ofantimycobacterial effects by IFN-g (4, 5). Thus, the activation ofmacrophages by IFN-g is necessary, but not sufficient, for eradi-cation of intracellular microorganisms. Therefore, additionalmechanisms must be involved in the clearance of these organisms,particularly at the site of primary infection.
M. tuberculosisinfects humans primarily via the respiratoryroute. Inhaled particles that are small enough to gain access to theterminal alveoli (,5mm) are phagocytosed, processed, and pre-sented mainly by alveolar macrophages (6). Another cell type
present in the bronchoalveolar space, the airway epithelium, andlung parenchyma are dendritic cells (DC)3 (7, 8). Immature pul-monary DC are strategically located in pulmonary airways anddistal alveoli, where they may function as sentinels for inhaledpathogens. Recent reports indicate an increased trafficking of DCinto mucosal tissues in response to local bacterial stimuli such asMoraxella,Bordatellaor Mycobacterium bovisbacillus Calmette-Guerin (BCG) (9–11). This property is likely to be central to therole of mucosal DC in surveillance of these front-line tissues forincoming microbial pathogens.
More recently, it has been discovered that DC can internalizelive pathogens including BCG (12),M. tuberculosis(13, 14),Bor-datella bronchosepticum(15), Chlamydia trachomatis(16, 17),Borrelia burgdorferi(18), Trypanosoma cruzi(19), andLeishma-nia major (20). Although the in vivo interaction of intracellularmicroorganisms and lung DC is essential for priming naive T cells,the ability of bacteria to survive in DC has not been elucidated.Human DC infected in vitro withM. tuberculosisor BCG undergoactivation and maturation (13, 21, 22) that presumably enhancetheir potential to stimulate T cells. Also, murine DC infected withBCG and injected into the foot pad of mice can induce a specificT cell response and provide protection against a subsequent aero-sol challenge withM. tuberculosis(23). These findings support thehypothesis that DC strengthen the cellular immune responseagainst mycobacteria.
We designed experiments to investigate whether DC also par-ticipate in protective immunity by mediating direct antimycobac-terial activity. Growth of intracellular bacteria was investigated inthe absence or presence of cytokines, which had previously beenshown to modulate mycobacterial growth in murine macrophages.Among a variety of cytokines tested, only IL-10, a product of
Institut fur Klinische Mikrobiologie, Immunologie und Hygiene der Friedrich-Alex-ander Universitat Erlangen-Nurnberg, Erlangen, Germany
Received for publication December 7, 1999. Accepted for publication May 2, 2000.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby markedadvertisementin accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 S.S. is supported by the Aids-Stipendienprogramm of the Bundesministerium fu¨rForschung und Technologie, Germany, and the German Research Foundation (DFG925 1-1).2 Address of correspondence and reprint requests to Dr. Steffen Stenger, Friedrich-Alexander Universitat Erlangen-Nurnberg, Institut fuer Klinische Mikrobiologie, Im-munologie und Hygiene, Wasserturmstrasse 3, D-91054 Erlangen, Germany. E-mailaddress: [email protected]
3 Abbreviations used in this paper: DC, dendritic cell(s); MOI, multiplicity of infec-tion; BCG, bacillus Calmette-Guerin; rhu, recombinant human.
Copyright © 2000 by The American Association of Immunologists 0022-1767/00/$02.00
by guest on June 19, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

macrophages with normally immunosuppressive effects, decreasedthe intracellular growth ofM. tuberculosis. In parallel, IL-10-treat-ment reduced the ability of DC to present mycobacterial lipid Agsto CD1-restricted T cells. This differential activity of IL-10 on DCmay provide a mechanism to maintain the balance between a pro-tective immune response and excessive cellular activation in themicroenvironment of a tuberculous granuloma.
Materials and MethodsCell culture reagents
Cells were cultured in RPMI 1640 (Biochrom KG, Berlin, Germany) sup-plemented with 10% heat-inactivated FCS (Sigma, St. Louis, MO), glu-tamine (2 mM, Sigma), 10 mM HEPES, 13 mM NaHCO3, 100 mg/mlstreptomycin, and 60mg/ml penicillin (all purchased from Biochrom). Ex-periments involving the infection of cells withM. tuberculosiswere per-formed in the absence of antibiotics, and FCS was replaced by pooledhuman serum (generated from the blood of healthy volunteers) to optimizethe phagocytosis of the bacteria.
Cytokines and Abs
The following cytokines and Abs were used:recombinant human (rhu)IL-2 (Proleukin; Chiron, Ratingen, Germany), rhu IL-4 (Strathmann Bio-tech, Hannover, Germany), rhu TNF-a, rhu IL-10 (both purchased fromEndogen, Woburn, MA), purified TGF-b1, anti IL-10 (R&D Systems,Wiesbaden, Germany), and rhu GM-CSF (kindly supplied by Novartis,Vienna, Austria).
The following Abs were used for flow cytometry and immunostaining:CD83-PE, CD56 (both from Immunotech, Marseilles, France), CD19,MHC class II-FITC, CD14-PE, CD86-PE, CD40-FITC (all from CymbusBiotechnology, Chandlers Ford, U.K.), CD3 (clone OKT3, obtained fromAmerican Type Culture Collection, Manassas, VA), CD1a-FITC (Serotec,Oxford, U.K.), goat-anti mouse-FITC (Jackson ImmunoResearch, WestGrove, PA), Texas Red-conjugated goat-anti-mouse Ab (Caltag, Burlin-game, CA), CD1b (clone bCD1b3.1, kindly provided by S. Porcelli, Bos-ton, MA), CD1c (clone 10C3, kindly provided by W. Knapp, Vienna, Aus-tria), lipoarabinomannan (kindly provided by J. Belisle, Fort Collins, CO).Isotype controls were all purchased from Cymbus Biotechnology.
Growth ofM. tuberculosis
M. tuberculosis(virulent strain H37Rv) was grown in suspension withconstant, gentle rotation in roller bottles containing Middlebrook 7H9 broth(Becton Dickinson, Heidelberg, Germany) supplemented with 1% glycerol(Roth, Karlsruhe, Germany), 0.05% Tween 80 (Sigma), and 10% Middle-brook oleic acid albumin dextrose catalase (OADC) enrichment (BectonDickinson). Aliquots from logarithmically growing cultures were frozen inPBS containing 10% glycerol, and representative vials were thawed andenumerated for viable CFU on Middlebrook 7H11 plates. Staining of bac-terial suspensions with fluorochromic substrates differentiating betweenlive and dead bacteria (BacLight, Molecular Probes, Leiden, The Nether-lands) revealed a viability of the bacteria above 90%. Because clumping ofmycobacteria is a common problem that can influence the validity andreproducibility of the experiments, we undertook several precautions tominimize clumps. First, culture conditions (rotation, Tween) were chosento support the growth of single cell suspensions. Second, before in vitroinfectionM. tuberculosisbacilli were sonicated to disrupt small aggregatesof bacteria. Third, the multiplicity of infection (MOI) was selected suchthat there were only two to three bacilli per DC.
Generation of DC and macrophages
PBMCs from healthy donors were isolated from buffy coats obtained fromthe Institute for Transfusion Medicine (University of Erlangen, Erlangen,Germany) by Ficoll-Hypaque (Pharmacia, Freiburg, Germany) density gra-dient centrifugation. Cells were allowed to adhere to Nunclon culture flasks(Nunc, Roskilde, Denmark) in RPMI 1640 plus 10% FCS. After 2 h at37°C, the nonadherent cells were removed by vigorous washing with PBS.In control experiments, cells were detached by incubation with Mg21- andCa21-free PBS containing 1 mM EDTA at 37°C for 10 min and harvestedfor flow cytometry. Cell-surface staining showed that the adherent popu-lation contained.95% monocytes. Adherent cells were incubated in cul-ture medium plus 10% FCS supplemented with GM-CSF (1000 U/ml) andIL-4 (1000 U/ml). Cytokines and 50% of the culture medium were replacedafter 3 days of culture. After 7 days, nonadherent (.85% of the totalpopulation) cells were harvested and used as the starting population for thefollowing experiments. This method is widely used becasue it yields sub-
stantial and pure populations of immature DC (24–26). To induce matu-ration of immature DC, cultures were supplemented with TNF-a (10 ng/ml) and incubated for an additional 48 h. Macrophages were generated byincubating PBMC in a culture flask for 1–2 h. Nonadherent cells wereremoved by three thorough washing steps. The adherent cells were thencultured in culture medium for 7 days before harvesting. The purity of themacrophage population was confirmed by FACS staining (CD141,CD11b1, CD192, CD562, CD3 , 2%) and was above 98% in allexperiments.
Infection of DC
DC were infected with single-cell suspensions ofM. tuberculosis. After 4 hof incubation at 37°C, DC were harvested (slightly adherent cells weredetached by vigorous pipetting) and centrifuged at 800 rpm for 8 min. Thislow-speed centrifugation selectively spins down DC while extracellularbacteria remain in the supernatant. After three cycles of centrifugation, themajority of extracellular bacteria were removed as determined by aura-mine-rhodamine stain (TB-fluor, Merck, Darmstadt, Germany). Infectedcells were then plated at a concentration of 13 106 cells/ml in a 24-wellplate in culture medium without antibiotics plus 10% human serum. Theefficiency of infection, as quantitated by staining of control cultures onPermanox chamber slides (Nunc) in every experiment was dependent onthe MOI. The microscopic evaluation of infected macrophages under thefluorescence microscope confirmed the absence of any mycobacterial ag-gregates. Cell viability of infected DC was determined by trypan blueexclusion and was.99% in all experiments.
Cytokine treatment of cell cultures
Dendritic cells or macrophages were harvested, washed, and plated intotissue cultures plates. After the pulse infection with mycobacteria, cyto-kines or anti IL-10 were added. The cytokines were not renewed (with theexception of one experimental series) and were present throughout theincubation period.
Confocal laser microscopy
Mycobacteria were incubated with an Ab directed against lipoarabinoman-nan (1:1 dilution of pure hybridoma supernatant) diluted in PBS/1% BSA/20%FCS for 30 min at room temperature. Lipoarabinomannan was labeledby incubation of a 1:50 dilution (PBS/1% BSA/20%FCS) of a Texas Red-conjugated goat-anti-mouse Ab for 30 min. After an additional washingstep, stained mycobacteria were resuspended and used for infection of DC,which were cultured on Permanox chamber slides. After 4 h, nonphago-cytosed bacteria were removed by washing with PBS, and infected cellswere stained with a FITC-conjugated Ab recognizing CD1a (1:20 dilution,30 min, room temperature). Finally, cells were fixed with 4% paraformal-dehyde (Sigma), mounted (Aquatex, Merck), and analyzed using a confo-cal microscope (Leica, Solms, Germany).
Quantification of mycobacterial growth
To ensure the reliable quantification of intracellularM. tuberculosisweemployed three independent methods. The first method used acid-fast stain(auramine-rhodamine). The second method employed colony forming units(CFU). After various time points of incubation, cells were lysed with 0.3%saponin (Sigma) to release intracellular bacteria. At all time points, analiquot of unlysed, infected cells was harvested and counted. This allowedan exact quantification of cells as well as the determination of cellularviability by trypan blue exclusion. Recovery of cells was.80% in allexperiments, with cell viability regularly exceeding 90% of total cells.Lysates of infected cells were resuspended vigorously, transferred intoscrew caps, and sonicated in a preheated (37°C) waterbath sonicator (Elma,Singen, Germany) for 5 min. Aliquots of the sonicate were diluted 10-foldin 7H9 medium. Four dilutions of each sample were plated in duplicates on7H11 agar plates and incubated at 37°C and 5% CO2 for 21 days. The thirdmethod involved [3H]uracil incorporation. Incorporation of [3H]-labeleduracil into the mycobacterial RNA was determined following the methodpublished by Rook and Rainbow (27) with several modifications. First, 13106 infected DC were cultured in duplicates as described above. At the endof the incubation period, cells were lysed using 0.3% saponin, resuspendedvigorously, and transferred into screw caps. Lysates were centrifuged in anaerosol-tight microfuge (3000 rpm, 20 min) and resuspended in 100ml of7H9 to allow optimal growth of the released mycobacteria. Lysates werethen transferred into 96-well round-bottom plates (Nunc) and incubated inthe presence of 3mCi [3H]uracil (Amersham-Pharmacia, Freiburg, Ger-many). After 24 h, mycobacteria were killed by treatment with parafor-maldehyde (final concentration, 4%) for 30 min. The mycobacteria wereharvested onto glass fiber filters (Inotech, Dottikon, Switzerland), and
979The Journal of Immunology
by guest on June 19, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
[3H]uracil incorporation was measured in a beta counter (Berthold,Munchen, Germany). Background radioactivity in uninfected cells was be-low 500 cpm in all experiments. To document the suitability of our quan-tification methods to detect antimycobacterial activity, we performed someexperiments in the presence of the mycobactericidal drug rifampicin(Sigma).
Flow cytometry
A total of 3 3 105 cells were resuspended in 100ml staining buffer (2%FCS, 1% NaN3, PBS without Mg21/Ca21) and incubated with unconju-gated or conjugated Abs for 30 min on ice. Samples were washed twice instaining buffer and if necessary incubated for an additional 30 min on icewith goat anti mouse-FITC Abs (1:500). Cells were then fixed in 2% para-formaldehyde and stored at 4°C until analysis in a FACScan flow cytom-eter. Data were analyzed using CellQuest software (Becton Dickinson).
Mixed lymphocyte reaction
DC were irradiated with 30 Gy (137Cs source). Subsequently, graded num-bers of these stimulator cells were mixed with a fixed amount of purifiedperipheral blood CD41 cells (5 3 104) and seeded into 96-well round-bottom tissue culture plates in a final volume of 200ml. CD41 cells wereenriched by indirect immunomagnetic depletion of cells expressing CD8,CD14, CD19, and CD56 (sheep anti mouse Dynabeads, Dynal, Oslo, Nor-way) from PBMC prepared by Ficoll-Hypaque density centrifugation. Pu-rity, as determined by flow cytometry, proved to be.95%. Controls in-cluded DC alone, responder cells alone, and responder cells cultured in thepresence of 100 U/ml IL-2. All cultures were set up as triplicates. After 5days of culture, 0.5mCi [3H]thymidine (Amersham-Pharmacia) was addedto each well, and thymidine incorporation was measured 18 h later using acell harvester and a betaplate counter.
CD1-restricted cell lines
CD1-restricted cell lines were generated as described earlier (28). DN7 wasderived from a healthy, purified protein derivative-positive donor. DN7 isTCRab1, CD42, CD82, and specifically recognizes purified lipoarabino-mannan fromM. tuberculosisin the context of the nonclassical MHC mol-ecule CD1b. To measure the Ag-specific response, DN7 (13 104) wasincubated in the presence of IL-10-treated or control DC (13 104) andlipoarabinomannan (1mg/ml) for 3 days. The last 4 h of incubation wereperformed in the presence of 0.5mCi [3H]thymidine. Incorporated radio-activity was measured after harvesting cells onto glass fiber filters in a betacounter.
Statistical analysis
Data are presented as mean value6 SEM except where stated otherwise.Student’st test was used to determine statistical significance between twodifferentially treated cultures. Differences were considered significant ifp , 0.05.
ResultsInfection of human DC with virulentM. tuberculosis
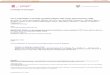
To monitor the growth ofM. tuberculosisin DC, it was necessaryto determine the MOI, which allowed us to culture infected cellsfor at least 7 days. Therefore, we infected DC with an increasingMOI. Four hours later, extracellular bacteria were removed by dif-ferential centrifugation, and the number of infected cells was de-termined by acid-fast stain (Fig. 1A). MOIs of 2.5 or 12.5 resultedin a high infection efficiency of 62% and 93%, respectively. How-ever, the high bacterial burden of the cells (Fig. 1B) resulted in celldeath within 48 h (data not shown). A MOI of 0.5 resulted ininfection of 24%6 0.4 of the cells, whereby each cell harbored anaverage of 2.26 0.3 bacteria. This infection rate did not result inrapid cell death and was chosen for studying the intracellulargrowth of M. tuberculosisin DC for 7 days. To ensure that wewere studying the growth of intracellular, but not extracellular,mycobacteria, we double-stained infected DC with Abs directedagainst mycobacteria and DC. Bacteria were labeled with an anti-lipoarabinomannan Ab and a Texas Red-conjugated secondary Abbefore infection of DC. Four hours after infection, DC werestained with an Ab recognizing the CD1a-FITC glycoprotein onthe cell surface of DC. Confocal microscopy revealed that over
FIGURE 1. Infection of DC withM. tuberculosis. Human DC were in-fected with an increasing number ofM. tuberculosis. After 4 h of incuba-tion, cells were fixed and stained with auramine-rhodamine. The cells wereexamined under a fluorescence microscope (wavelength, 594 nm), and thepercentage of infected cells (A) as well as the bacterial load of infectedcells (B) was determined. For each MOI, at least 300 cells were examinedfor intracellular acid-fast rods. The graphs show the average results of eightindependent experiments using DC derived from different donors.C, DCwere infected with stainedM. tuberculosis(anti-lipoarabinomannan, Texas-Red) and then labeled with anti CD1a (FITC). Cells were analyzed usingconfocal microscopy. The panel shows a representative overlay of the red(mycobacteria) and green (DC) fluorescence (1000-fold magnification).
980 IL-10 LIMITS THE GROWTH OFM. tuberculosisIN DENDRITIC CELLS
by guest on June 19, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
90% of the bacteria were localized intracellularly (Fig. 1C). Toconfirm the intracellular localization ofM. tuberculosis, DC wereharvested and centrifuged at 800 rpm. At this low speed, cells arepreferentially spun down, while the majority of bacteria will re-main in the supernatant. Plating of these supernatants revealed that,0.1% of the initial inoculum remained in the extracellular com-partment (not shown).
Growth ofM. tuberculosisin human DC
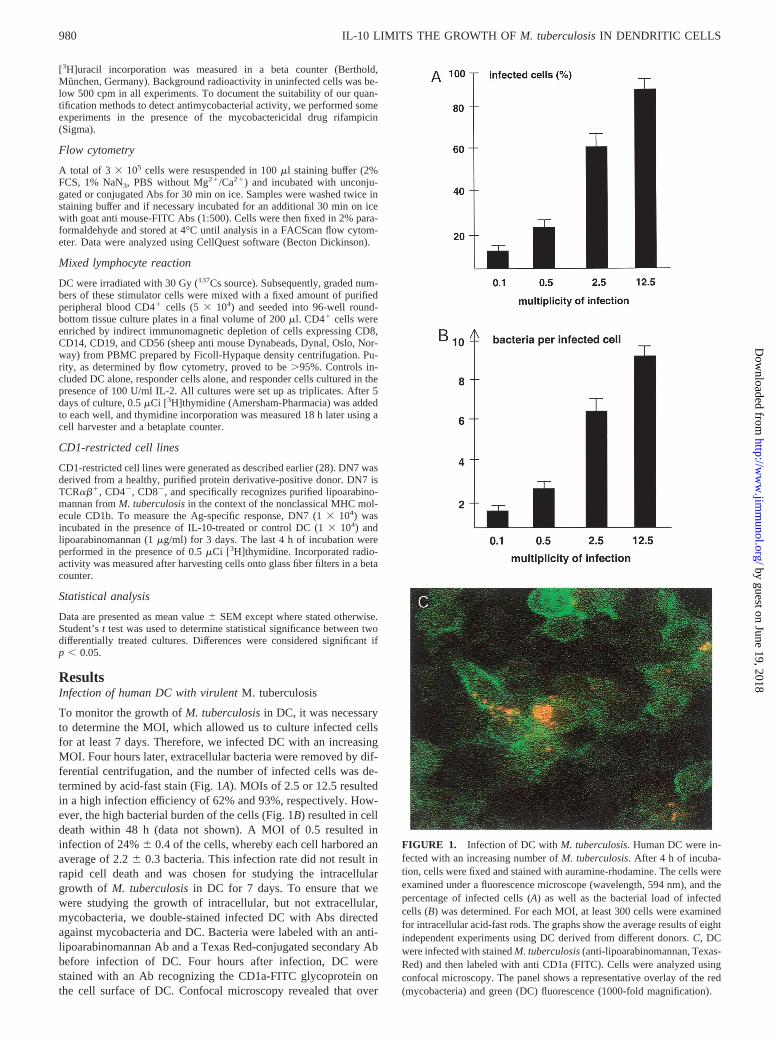
Using this model of low-dose infection (MOI 0.5), we found thatafter an initial quiescent phase of 24 h the number ofM. tubercu-losis increased 4-fold within 1 day and continued to multiplythroughout the observation period (Fig. 2). Total growth of thebacteria was two orders of magnitude within 7 days, which isequivalent to an average generation time ofM. tuberculosisinhuman DC of 26 h. The replication of bacteria resulted in an in-creased bacterial burden of individual cells, as well as a highernumber of infected cells (Table I). Even though this finding indi-cates that some cells must have been lysed to allow spreading ofthe mycobacteria, the number of viable cells after 7 days of infec-tion was above 90% of the initial inoculum (data not shown). Toverify that we were not studying the growth of extracellular bac-teria that had been released into the culture medium by dying cells,we centrifuged representative lysates at 800 rpm. These superna-tants contained,0.5% of the total bacteria in the culture plate.
Even though there was a considerable variabilty in the ability ofthe DC of individual donors to phagocytoseM. tuberculosis(21–32%), the generation time remained similar. An extension of theobservation period beyond 7 days was not informative as cell vi-ability declined drastically.
Influence of IFN-g and TNF-a on the growth ofM. tuberculosis
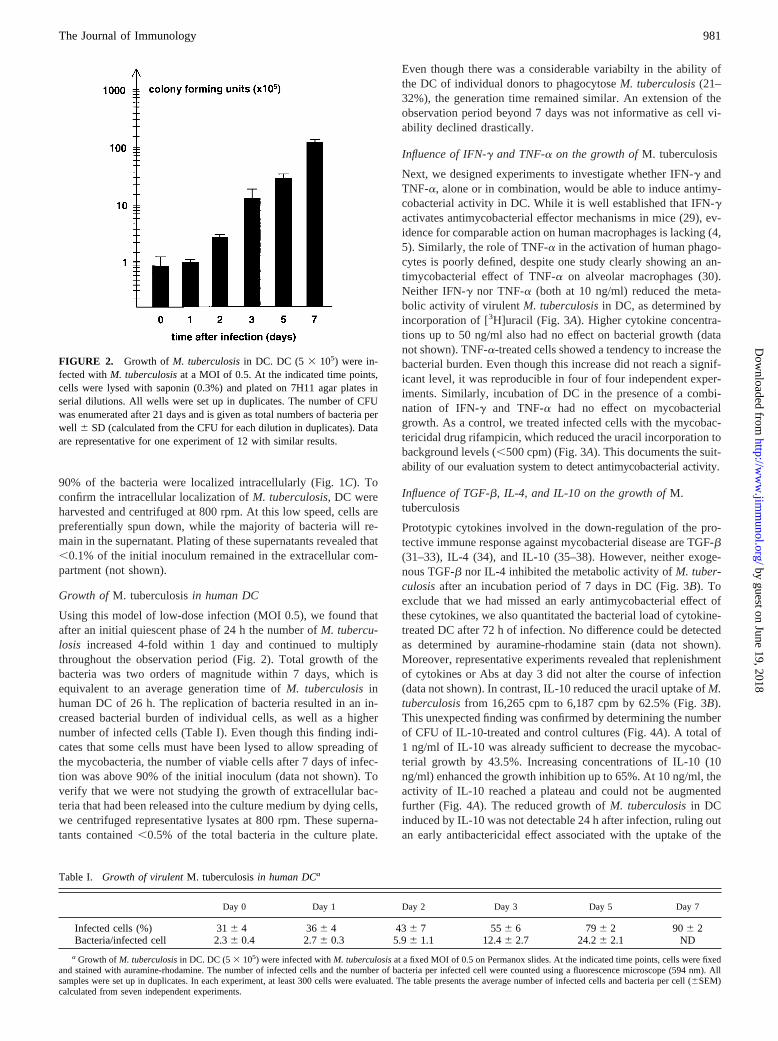
Next, we designed experiments to investigate whether IFN-g andTNF-a, alone or in combination, would be able to induce antimy-cobacterial activity in DC. While it is well established that IFN-gactivates antimycobacterial effector mechanisms in mice (29), ev-idence for comparable action on human macrophages is lacking (4,5). Similarly, the role of TNF-a in the activation of human phago-cytes is poorly defined, despite one study clearly showing an an-timycobacterial effect of TNF-a on alveolar macrophages (30).Neither IFN-g nor TNF-a (both at 10 ng/ml) reduced the meta-bolic activity of virulentM. tuberculosisin DC, as determined byincorporation of [3H]uracil (Fig. 3A). Higher cytokine concentra-tions up to 50 ng/ml also had no effect on bacterial growth (datanot shown). TNF-a-treated cells showed a tendency to increase thebacterial burden. Even though this increase did not reach a signif-icant level, it was reproducible in four of four independent exper-iments. Similarly, incubation of DC in the presence of a combi-nation of IFN-g and TNF-a had no effect on mycobacterialgrowth. As a control, we treated infected cells with the mycobac-tericidal drug rifampicin, which reduced the uracil incorporation tobackground levels (,500 cpm) (Fig. 3A). This documents the suit-ability of our evaluation system to detect antimycobacterial activity.
Influence of TGF-b, IL-4, and IL-10 on the growth ofM.tuberculosis
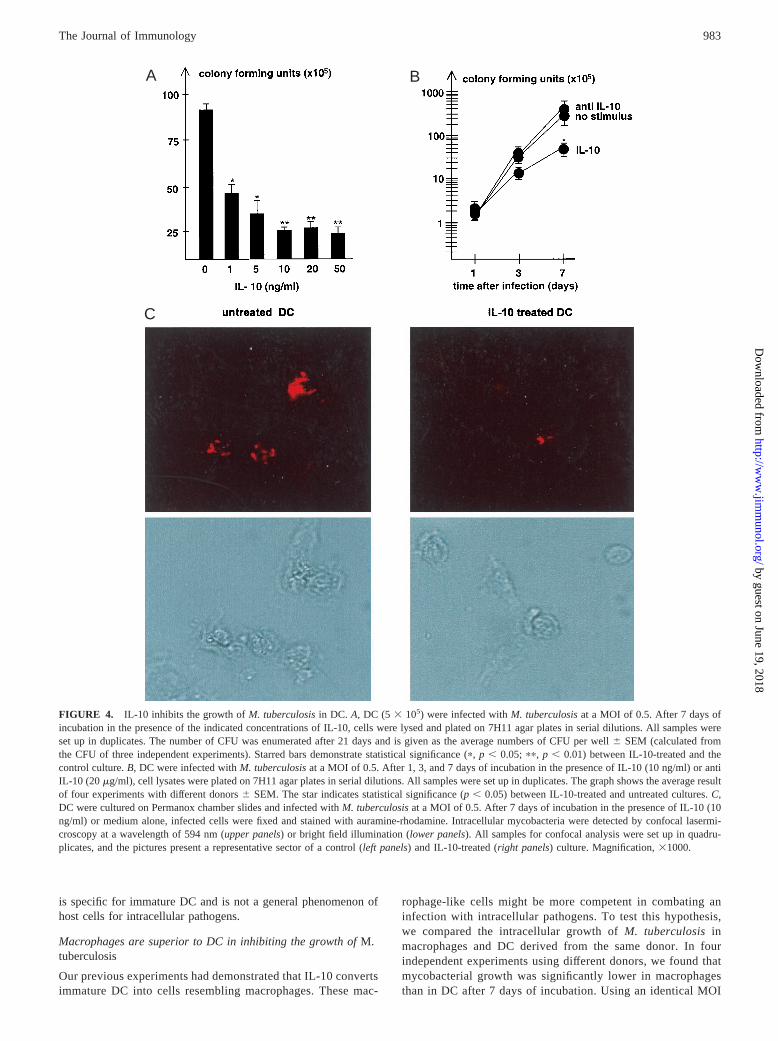
Prototypic cytokines involved in the down-regulation of the pro-tective immune response against mycobacterial disease are TGF-b(31–33), IL-4 (34), and IL-10 (35–38). However, neither exoge-nous TGF-b nor IL-4 inhibited the metabolic activity ofM. tuber-culosisafter an incubation period of 7 days in DC (Fig. 3B). Toexclude that we had missed an early antimycobacterial effect ofthese cytokines, we also quantitated the bacterial load of cytokine-treated DC after 72 h of infection. No difference could be detectedas determined by auramine-rhodamine stain (data not shown).Moreover, representative experiments revealed that replenishmentof cytokines or Abs at day 3 did not alter the course of infection(data not shown). In contrast, IL-10 reduced the uracil uptake ofM.tuberculosisfrom 16,265 cpm to 6,187 cpm by 62.5% (Fig. 3B).This unexpected finding was confirmed by determining the numberof CFU of IL-10-treated and control cultures (Fig. 4A). A total of1 ng/ml of IL-10 was already sufficient to decrease the mycobac-terial growth by 43.5%. Increasing concentrations of IL-10 (10ng/ml) enhanced the growth inhibition up to 65%. At 10 ng/ml, theactivity of IL-10 reached a plateau and could not be augmentedfurther (Fig. 4A). The reduced growth ofM. tuberculosisin DCinduced by IL-10 was not detectable 24 h after infection, ruling outan early antibactericidal effect associated with the uptake of the
FIGURE 2. Growth of M. tuberculosisin DC. DC (5 3 105) were in-fected withM. tuberculosisat a MOI of 0.5. At the indicated time points,cells were lysed with saponin (0.3%) and plated on 7H11 agar plates inserial dilutions. All wells were set up in duplicates. The number of CFUwas enumerated after 21 days and is given as total numbers of bacteria perwell 6 SD (calculated from the CFU for each dilution in duplicates). Dataare representative for one experiment of 12 with similar results.
Table I. Growth of virulentM. tuberculosisin human DCa
Day 0 Day 1 Day 2 Day 3 Day 5 Day 7
Infected cells (%) 316 4 36 6 4 43 6 7 55 6 6 79 6 2 90 6 2Bacteria/infected cell 2.36 0.4 2.76 0.3 5.96 1.1 12.46 2.7 24.26 2.1 ND
a Growth ofM. tuberculosisin DC. DC (53 105) were infected withM. tuberculosisat a fixed MOI of 0.5 on Permanox slides. At the indicated time points, cells were fixedand stained with auramine-rhodamine. The number of infected cells and the number of bacteria per infected cell were counted using a fluorescence microscope (594 nm). Allsamples were set up in duplicates. In each experiment, at least 300 cells were evaluated. The table presents the average number of infected cells and bacteria per cell (6SEM)calculated from seven independent experiments.
981The Journal of Immunology
by guest on June 19, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
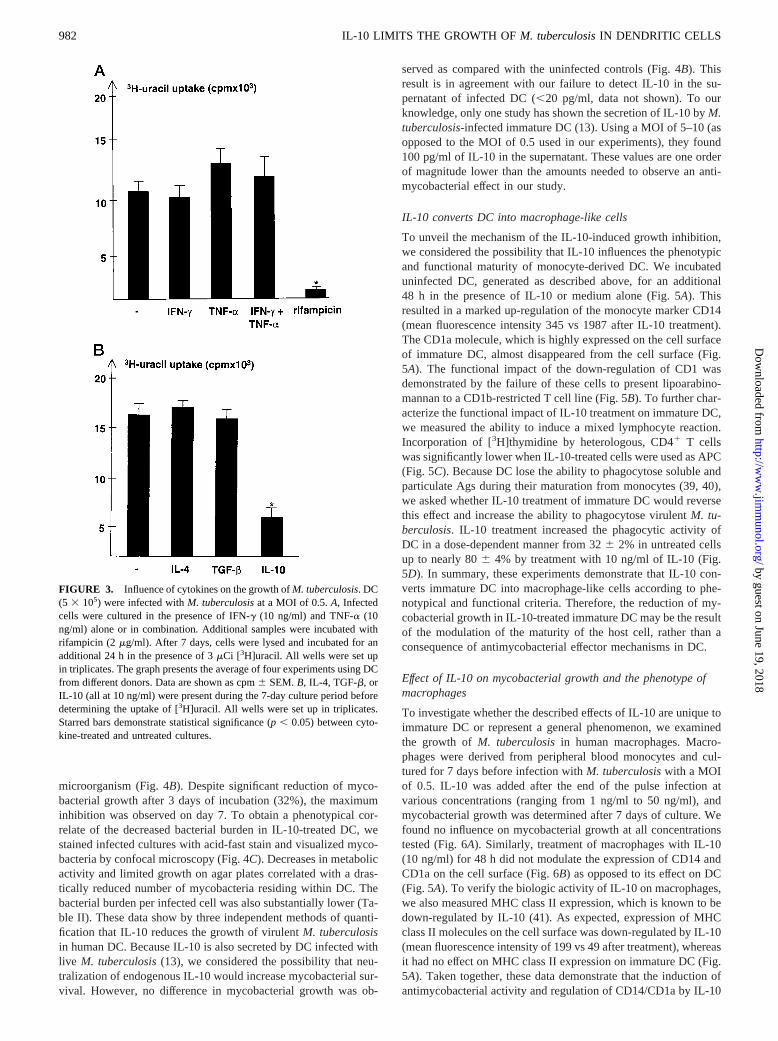
microorganism (Fig. 4B). Despite significant reduction of myco-bacterial growth after 3 days of incubation (32%), the maximuminhibition was observed on day 7. To obtain a phenotypical cor-relate of the decreased bacterial burden in IL-10-treated DC, westained infected cultures with acid-fast stain and visualized myco-bacteria by confocal microscopy (Fig. 4C). Decreases in metabolicactivity and limited growth on agar plates correlated with a dras-tically reduced number of mycobacteria residing within DC. Thebacterial burden per infected cell was also substantially lower (Ta-ble II). These data show by three independent methods of quanti-fication that IL-10 reduces the growth of virulentM. tuberculosisin human DC. Because IL-10 is also secreted by DC infected withlive M. tuberculosis(13), we considered the possibility that neu-tralization of endogenous IL-10 would increase mycobacterial sur-vival. However, no difference in mycobacterial growth was ob-
served as compared with the uninfected controls (Fig. 4B). Thisresult is in agreement with our failure to detect IL-10 in the su-pernatant of infected DC (,20 pg/ml, data not shown). To ourknowledge, only one study has shown the secretion of IL-10 byM.tuberculosis-infected immature DC (13). Using a MOI of 5–10 (asopposed to the MOI of 0.5 used in our experiments), they found100 pg/ml of IL-10 in the supernatant. These values are one orderof magnitude lower than the amounts needed to observe an anti-mycobacterial effect in our study.
IL-10 converts DC into macrophage-like cells
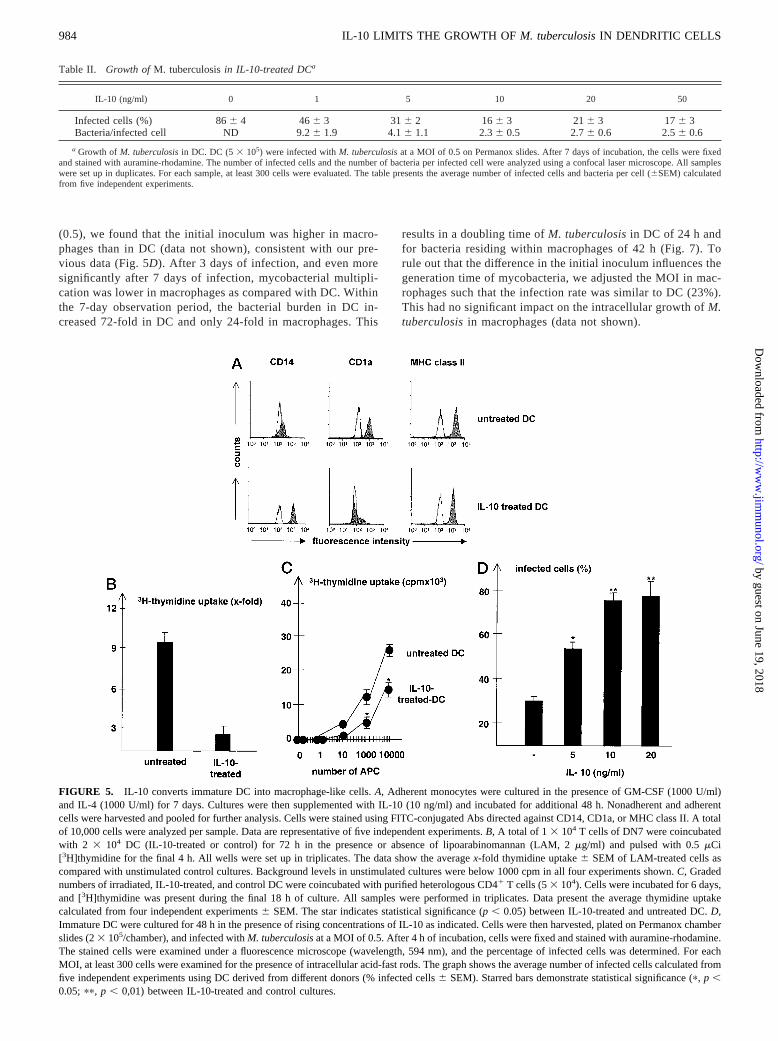
To unveil the mechanism of the IL-10-induced growth inhibition,we considered the possibility that IL-10 influences the phenotypicand functional maturity of monocyte-derived DC. We incubateduninfected DC, generated as described above, for an additional48 h in the presence of IL-10 or medium alone (Fig. 5A). Thisresulted in a marked up-regulation of the monocyte marker CD14(mean fluorescence intensity 345 vs 1987 after IL-10 treatment).The CD1a molecule, which is highly expressed on the cell surfaceof immature DC, almost disappeared from the cell surface (Fig.5A). The functional impact of the down-regulation of CD1 wasdemonstrated by the failure of these cells to present lipoarabino-mannan to a CD1b-restricted T cell line (Fig. 5B). To further char-acterize the functional impact of IL-10 treatment on immature DC,we measured the ability to induce a mixed lymphocyte reaction.Incorporation of [3H]thymidine by heterologous, CD41 T cellswas significantly lower when IL-10-treated cells were used as APC(Fig. 5C). Because DC lose the ability to phagocytose soluble andparticulate Ags during their maturation from monocytes (39, 40),we asked whether IL-10 treatment of immature DC would reversethis effect and increase the ability to phagocytose virulentM. tu-berculosis. IL-10 treatment increased the phagocytic activity ofDC in a dose-dependent manner from 326 2% in untreated cellsup to nearly 806 4% by treatment with 10 ng/ml of IL-10 (Fig.5D). In summary, these experiments demonstrate that IL-10 con-verts immature DC into macrophage-like cells according to phe-notypical and functional criteria. Therefore, the reduction of my-cobacterial growth in IL-10-treated immature DC may be the resultof the modulation of the maturity of the host cell, rather than aconsequence of antimycobacterial effector mechanisms in DC.
Effect of IL-10 on mycobacterial growth and the phenotype ofmacrophages
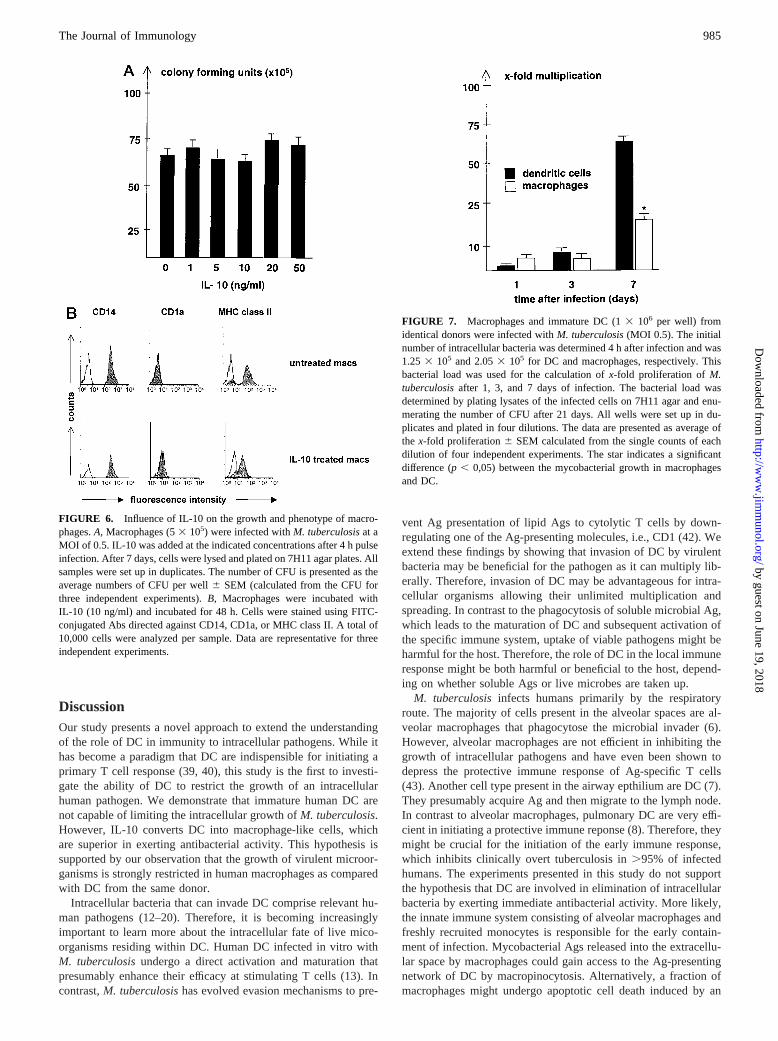
To investigate whether the described effects of IL-10 are unique toimmature DC or represent a general phenomenon, we examinedthe growth of M. tuberculosisin human macrophages. Macro-phages were derived from peripheral blood monocytes and cul-tured for 7 days before infection withM. tuberculosiswith a MOIof 0.5. IL-10 was added after the end of the pulse infection atvarious concentrations (ranging from 1 ng/ml to 50 ng/ml), andmycobacterial growth was determined after 7 days of culture. Wefound no influence on mycobacterial growth at all concentrationstested (Fig. 6A). Similarly, treatment of macrophages with IL-10(10 ng/ml) for 48 h did not modulate the expression of CD14 andCD1a on the cell surface (Fig. 6B) as opposed to its effect on DC(Fig. 5A). To verify the biologic activity of IL-10 on macrophages,we also measured MHC class II expression, which is known to bedown-regulated by IL-10 (41). As expected, expression of MHCclass II molecules on the cell surface was down-regulated by IL-10(mean fluorescence intensity of 199 vs 49 after treatment), whereasit had no effect on MHC class II expression on immature DC (Fig.5A). Taken together, these data demonstrate that the induction ofantimycobacterial activity and regulation of CD14/CD1a by IL-10
FIGURE 3. Influence of cytokines on the growth ofM. tuberculosis. DC(5 3 105) were infected withM. tuberculosisat a MOI of 0.5.A, Infectedcells were cultured in the presence of IFN-g (10 ng/ml) and TNF-a (10ng/ml) alone or in combination. Additional samples were incubated withrifampicin (2 mg/ml). After 7 days, cells were lysed and incubated for anadditional 24 h in the presence of 3mCi [3H]uracil. All wells were set upin triplicates. The graph presents the average of four experiments using DCfrom different donors. Data are shown as cpm6 SEM.B, IL-4, TGF-b, orIL-10 (all at 10 ng/ml) were present during the 7-day culture period beforedetermining the uptake of [3H]uracil. All wells were set up in triplicates.Starred bars demonstrate statistical significance (p, 0.05) between cyto-kine-treated and untreated cultures.
982 IL-10 LIMITS THE GROWTH OFM. tuberculosisIN DENDRITIC CELLS
by guest on June 19, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
is specific for immature DC and is not a general phenomenon ofhost cells for intracellular pathogens.
Macrophages are superior to DC in inhibiting the growth ofM.tuberculosis
Our previous experiments had demonstrated that IL-10 convertsimmature DC into cells resembling macrophages. These mac-
rophage-like cells might be more competent in combating aninfection with intracellular pathogens. To test this hypothesis,we compared the intracellular growth ofM. tuberculosisinmacrophages and DC derived from the same donor. In fourindependent experiments using different donors, we found thatmycobacterial growth was significantly lower in macrophagesthan in DC after 7 days of incubation. Using an identical MOI
C
BA
FIGURE 4. IL-10 inhibits the growth ofM. tuberculosisin DC. A, DC (5 3 105) were infected withM. tuberculosisat a MOI of 0.5. After 7 days ofincubation in the presence of the indicated concentrations of IL-10, cells were lysed and plated on 7H11 agar plates in serial dilutions. All samples wereset up in duplicates. The number of CFU was enumerated after 21 days and is given as the average numbers of CFU per well6 SEM (calculated fromthe CFU of three independent experiments). Starred bars demonstrate statistical significance (p,p , 0.05; pp, p , 0.01) between IL-10-treated and thecontrol culture.B, DC were infected withM. tuberculosisat a MOI of 0.5. After 1, 3, and 7 days of incubation in the presence of IL-10 (10 ng/ml) or antiIL-10 (20 mg/ml), cell lysates were plated on 7H11 agar plates in serial dilutions. All samples were set up in duplicates. The graph shows the average resultof four experiments with different donors6 SEM. The star indicates statistical significance (p, 0.05) between IL-10-treated and untreated cultures.C,DC were cultured on Permanox chamber slides and infected withM. tuberculosisat a MOI of 0.5. After 7 days of incubation in the presence of IL-10 (10ng/ml) or medium alone, infected cells were fixed and stained with auramine-rhodamine. Intracellular mycobacteria were detected by confocal lasermi-croscopy at a wavelength of 594 nm (upper panels) or bright field illumination (lower panels). All samples for confocal analysis were set up in quadru-plicates, and the pictures present a representative sector of a control (left panels) and IL-10-treated (right panels) culture. Magnification,31000.
983The Journal of Immunology
by guest on June 19, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
(0.5), we found that the initial inoculum was higher in macro-phages than in DC (data not shown), consistent with our pre-vious data (Fig. 5D). After 3 days of infection, and even moresignificantly after 7 days of infection, mycobacterial multipli-cation was lower in macrophages as compared with DC. Withinthe 7-day observation period, the bacterial burden in DC in-creased 72-fold in DC and only 24-fold in macrophages. This
results in a doubling time ofM. tuberculosisin DC of 24 h andfor bacteria residing within macrophages of 42 h (Fig. 7). Torule out that the difference in the initial inoculum influences thegeneration time of mycobacteria, we adjusted the MOI in mac-rophages such that the infection rate was similar to DC (23%).This had no significant impact on the intracellular growth ofM.tuberculosisin macrophages (data not shown).
Table II. Growth ofM. tuberculosisin IL-10-treated DCa
IL-10 (ng/ml) 0 1 5 10 20 50
Infected cells (%) 866 4 46 6 3 31 6 2 16 6 3 21 6 3 17 6 3Bacteria/infected cell ND 9.26 1.9 4.16 1.1 2.36 0.5 2.76 0.6 2.56 0.6
a Growth of M. tuberculosisin DC. DC (53 105) were infected withM. tuberculosisat a MOI of 0.5 on Permanox slides. After 7 days of incubation, the cells were fixedand stained with auramine-rhodamine. The number of infected cells and the number of bacteria per infected cell were analyzed using a confocal laser microscope. All sampleswere set up in duplicates. For each sample, at least 300 cells were evaluated. The table presents the average number of infected cells and bacteria per cell (6SEM) calculatedfrom five independent experiments.
FIGURE 5. IL-10 converts immature DC into macrophage-like cells.A, Adherent monocytes were cultured in the presence of GM-CSF (1000 U/ml)and IL-4 (1000 U/ml) for 7 days. Cultures were then supplemented with IL-10 (10 ng/ml) and incubated for additional 48 h. Nonadherent and adherentcells were harvested and pooled for further analysis. Cells were stained using FITC-conjugated Abs directed against CD14, CD1a, or MHC class II. A totalof 10,000 cells were analyzed per sample. Data are representative of five independent experiments.B, A total of 1 3 104 T cells of DN7 were coincubatedwith 2 3 104 DC (IL-10-treated or control) for 72 h in the presence or absence of lipoarabinomannan (LAM, 2mg/ml) and pulsed with 0.5mCi[3H]thymidine for the final 4 h. All wells were set up in triplicates. The data show the averagex-fold thymidine uptake6 SEM of LAM-treated cells ascompared with unstimulated control cultures. Background levels in unstimulated cultures were below 1000 cpm in all four experiments shown.C, Gradednumbers of irradiated, IL-10-treated, and control DC were coincubated with purified heterologous CD41 T cells (53 104). Cells were incubated for 6 days,and [3H]thymidine was present during the final 18 h of culture. All samples were performed in triplicates. Data present the average thymidine uptakecalculated from four independent experiments6 SEM. The star indicates statistical significance (p, 0.05) between IL-10-treated and untreated DC.D,Immature DC were cultured for 48 h in the presence of rising concentrations of IL-10 as indicated. Cells were then harvested, plated on Permanox chamberslides (23 105/chamber), and infected withM. tuberculosisat a MOI of 0.5. After 4 h of incubation, cells were fixed and stained with auramine-rhodamine.The stained cells were examined under a fluorescence microscope (wavelength, 594 nm), and the percentage of infected cells was determined. For eachMOI, at least 300 cells were examined for the presence of intracellular acid-fast rods. The graph shows the average number of infected cells calculated fromfive independent experiments using DC derived from different donors (% infected cells6 SEM). Starred bars demonstrate statistical significance (p, p ,0.05; pp, p , 0,01) between IL-10-treated and control cultures.
984 IL-10 LIMITS THE GROWTH OFM. tuberculosisIN DENDRITIC CELLS
by guest on June 19, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
DiscussionOur study presents a novel approach to extend the understandingof the role of DC in immunity to intracellular pathogens. While ithas become a paradigm that DC are indispensible for initiating aprimary T cell response (39, 40), this study is the first to investi-gate the ability of DC to restrict the growth of an intracellularhuman pathogen. We demonstrate that immature human DC arenot capable of limiting the intracellular growth ofM. tuberculosis.However, IL-10 converts DC into macrophage-like cells, whichare superior in exerting antibacterial activity. This hypothesis issupported by our observation that the growth of virulent microor-ganisms is strongly restricted in human macrophages as comparedwith DC from the same donor.
Intracellular bacteria that can invade DC comprise relevant hu-man pathogens (12–20). Therefore, it is becoming increasinglyimportant to learn more about the intracellular fate of live mico-organisms residing within DC. Human DC infected in vitro withM. tuberculosisundergo a direct activation and maturation thatpresumably enhance their efficacy at stimulating T cells (13). Incontrast,M. tuberculosishas evolved evasion mechanisms to pre-
vent Ag presentation of lipid Ags to cytolytic T cells by down-regulating one of the Ag-presenting molecules, i.e., CD1 (42). Weextend these findings by showing that invasion of DC by virulentbacteria may be beneficial for the pathogen as it can multiply lib-erally. Therefore, invasion of DC may be advantageous for intra-cellular organisms allowing their unlimited multiplication andspreading. In contrast to the phagocytosis of soluble microbial Ag,which leads to the maturation of DC and subsequent activation ofthe specific immune system, uptake of viable pathogens might beharmful for the host. Therefore, the role of DC in the local immuneresponse might be both harmful or beneficial to the host, depend-ing on whether soluble Ags or live microbes are taken up.
M. tuberculosisinfects humans primarily by the respiratoryroute. The majority of cells present in the alveolar spaces are al-veolar macrophages that phagocytose the microbial invader (6).However, alveolar macrophages are not efficient in inhibiting thegrowth of intracellular pathogens and have even been shown todepress the protective immune response of Ag-specific T cells(43). Another cell type present in the airway epthilium are DC (7).They presumably acquire Ag and then migrate to the lymph node.In contrast to alveolar macrophages, pulmonary DC are very effi-cient in initiating a protective immune reponse (8). Therefore, theymight be crucial for the initiation of the early immune response,which inhibits clinically overt tuberculosis in.95% of infectedhumans. The experiments presented in this study do not supportthe hypothesis that DC are involved in elimination of intracellularbacteria by exerting immediate antibacterial activity. More likely,the innate immune system consisting of alveolar macrophages andfreshly recruited monocytes is responsible for the early contain-ment of infection. Mycobacterial Ags released into the extracellu-lar space by macrophages could gain access to the Ag-presentingnetwork of DC by macropinocytosis. Alternatively, a fraction ofmacrophages might undergo apoptotic cell death induced by an
FIGURE 6. Influence of IL-10 on the growth and phenotype of macro-phages.A, Macrophages (53 105) were infected withM. tuberculosisat aMOI of 0.5. IL-10 was added at the indicated concentrations after 4 h pulseinfection. After 7 days, cells were lysed and plated on 7H11 agar plates. Allsamples were set up in duplicates. The number of CFU is presented as theaverage numbers of CFU per well6 SEM (calculated from the CFU forthree independent experiments).B, Macrophages were incubated withIL-10 (10 ng/ml) and incubated for 48 h. Cells were stained using FITC-conjugated Abs directed against CD14, CD1a, or MHC class II. A total of10,000 cells were analyzed per sample. Data are representative for threeindependent experiments.
FIGURE 7. Macrophages and immature DC (13 106 per well) fromidentical donors were infected withM. tuberculosis(MOI 0.5). The initialnumber of intracellular bacteria was determined 4 h after infection and was1.25 3 105 and 2.053 105 for DC and macrophages, respectively. Thisbacterial load was used for the calculation ofx-fold proliferation ofM.tuberculosisafter 1, 3, and 7 days of infection. The bacterial load wasdetermined by plating lysates of the infected cells on 7H11 agar and enu-merating the number of CFU after 21 days. All wells were set up in du-plicates and plated in four dilutions. The data are presented as average ofthe x-fold proliferation6 SEM calculated from the single counts of eachdilution of four independent experiments. The star indicates a significantdifference (p, 0,05) between the mycobacterial growth in macrophagesand DC.
985The Journal of Immunology
by guest on June 19, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

overwhelming bacterial burden. These apoptotic bodies could thenbe taken up by DC, which present mycobacterial Ags to T cells.While direct evidence for the uptake of regurgitated mycobacterialproducts by DC is lacking (44), DC that have taken up Ags derivedfrom apoptotic bodies have been shown to activate CD81 cytolyticT cells (45). CD81 T cells have been suggested to play a specialrole in the human immune response toM. tuberculosisby injectinganti-mycobacterial effector molecules such as granulysin into thetarget cell (46). The special role of DC in Ag presentation is un-derscored by their unique ability to present nonprotein Ags to Tcells via the nonclassical MHC molecules CD1a, -b, and -c (47).Because our experiments demonstrate that DC are poor antibacte-rial effector cells (Figs. 2 and 7) and are clearly inferior to mac-rophages in this regard, their prominent function is more likely tolink the innate and acquired immune response by recruiting andactivating Ag-specific T cells.
IL-10 was first detected based on its cytokine synthesis inhibi-tory activity mainly on macrophages (48–50). Recently, evidenceis accumulating that DC are another major target for the action ofthis immunosuppressive cytokine. IL-10 was reported to inhibit theAg-presenting capacity of DC (51–55), to reduce the expression ofAg-presenting and costimulatory molecules (56–59), and to inter-fere with the maturation of monocytes to DC (60–62). Taken to-gether, these studies suggest an overall picture in which IL-10prevents the differentiation of monocytes to mature DC but pro-motes their maturation to macrophages (63). Recent findings dem-onstrate that maturation of monocytes to DC also occurs in vivo(64, 65). These findings point out that the development of mono-cytic cells into either mature tissue-macrophages or DC dependson the local microenvironment. The opposite development of im-mature DC into macrophages as shown here in an in vitro culturesystem has not been formally proven in vivo. However, the studiesdiscussed above demonstrate the potential of immature cells todevelop differentially in vivo. This may serve to pave the way forthe maturation of cells optimally equipped to meet the functionalrequirements of the local immune response. Specifically, in thecase of a tuberculous granuloma, the local cytokine microenviron-ment may be dominated by IL-10 in certain instances (36), therebysupporting the development of macrophage-like cells. These willthen complement effector mechanisms of the protective cellularimmune response and contribute to the eradication ofM.tuberculosis.
One mechanism of infected cells to eliminate intracellularpathogens is to undergo apoptosis, thereby exposing the microbesto the extracellular environment. IL-10 has been shown to inhibitapoptosis of cells infected with intracellular bacteria including my-cobacteria (66, 67). However, we did not observe apoptosis inuntreated or IL-10-treated immature DC as determined by annexinV staining (data not shown). The failure to detect apoptotic cellswas most likely a consequence of the low MOI, which was chosento allow a 7-day observation period of mycobacterial growth. Also,if IL-10 would inhibit apoptosis of infected DC, bacterial growthwould more likely be increased, rather than inhibited as observedin this study. Therefore, growth inhibition of mycobacteria in im-mature DC by IL-10 is unlikely to be mediated by the modulationof apoptosis.
In vivo studies revealed that IL-10-expressing cells tend to ac-cumulate in patients suffering from an unfavorable outcome ofdisease (35, 68). However, the data from murine studies argueagainst a critical, nonredundant role of IL-10 in immunity to tu-berculosis because IL-10-deficient mice are not or are only par-tially protected from tuberculosis (38, 69).
This study exemplifies the versatility of the immune system withan armamentarium of effector cells, each having specific and spe-
cialized functions in immunity to microorganisms. In the setting ofa local cellular immune response, as is typical for a tuberculousgranuloma, the differential effects of IL-10 on macrophages andDC may contribute to the fine-tuned balance that must provideprotection from the pathogen, as well as limitation of excessivetissue destruction.
AcknowledgmentsWe acknowledge the expert technical assistance of Kirstin Castiglione andNives Schwerdtner. We thank Dr. Heinrich Korner for critical reading ofthe manuscript, and Dr. Klaus Schroppel for helping us with the amexinstaining.
References1. Koch, R. 1882. Die Aetiologie der Tuberkulose.Berl. Klin. Woch. 15:428.2. Stenger, S., and R. L. Modlin.1999. T-cell-mediated immunity toMycobacterium
tuberculosis.Curr. Opin. Immun. 2:89.3. Jouanguy, E., R. Doffinger, S. Dupuis, A. Pallier, F. Altare, and J. L. Casanova.
1999. IL-12 and IFN-g in host defense against mycobacteria and salmonella inmice and men.Curr. Opin. Immunol. 3:346.
4. Douvas, G. S., D. L. Looker, A. E. Vatter, and A. J. Crowle. 1985. Gammainterferon activates human macrophages to become tumoricidal and leishmani-cidal but enhances replication of macrophage-associated mycobacteria.Infect.Immun. 50:1.
5. Rook, G. A., J. Steele, L. Fraher, S. Barker, R, Karmali, J. O’Riordan, andJ. Stanford. 1986. Vitamin D3,g interferon, and control of proliferation ofMy-cobacterium tuberculosisby human monocytes.Immunology 57:159.
6. Hocking, W. G., and D. W. Golde. 1979. The pulmonary-alveolar macrophage.N. Engl. J. Med. 301:639.
7. Sertl, K. T., T. Takemura, E. Tschachler, V. J. Ferrans, M. A. Kaliner, andE. M. Shevach. 1986. Dendritic cells with antigen presenting capability reside inairway epithelium, lung parenchyma and visceral pleura.J. Exp. Med. 163:436.
8. Maarten, J., W. van Haarst. H. C. Hoogsteden, H. J. de Wit, G. T. Verhoeven,C. E. G. Havenith, and H. A. Drexhage. 1994. Dendritic cells and their precursorsisolated from human bronchoalveolar lavage: immunocytologic and functionalproperties.Am. J. Respir. Cell Mol. Biol. 11:344.
9. Havenith, C .E. G., A. J. Breedijk, and E. C. M. Hoefsmit. 1992. Effect of BacilleCalmette Guerin inoculation on numbers of dendritic cells in bronchoalveolarlavage of rats.Immunobiology 184:336.
10. McWilliam, A. S., D. Nelson, J. A. Thomas, and P. G. Holt. 1994. Rapid den-dritic cell recruitment is a hallmark of the acute inflammatory response at mu-cosal surfaces.J. Exp. Med. 179:1331.
11. McWilliam, A. S., S. Napoli, A. M. Marsh, F. L. Pemper, D. J. Nelson,C. L. Pimm, P. A. Stumbles, T. N. C. Wells, and P. G. Holt. 1996. Dendritic cellsare recruited to the airway epithelium by a broad spectrum of stimuli.J. Exp.Med. 184:2429.
12. Inaba, K., M. Inaba, M. Naito, and R. M. Steinman. 1993. Dendritic cell pro-genitors phagocytose particulates, including Bacillus Calmette-Guerin organisms,and sensitize mice to mycobacterial antigens in vivo.J. Exp. Med. 178:479.
13. Henderson, R. A., S. C. Watkins, and J. L. Flynn. 1997. Activation of humandendritic cells following infection withMycobacterium tuberculosis.J. Immunol.159:635.
14. Larsson, M., M. Majeed, J. D. Ernst, K.-E. Magnussen, O. Stendahl, andU. Forsum. 1997. Role of annexins in endocytosis of antigens in immature den-dritic cells. Immunology 92:501.
15. Guzman, C. A., M. Rhode, M. Bock, and K. N. Timmis. 1994. Invasion andintracellular survival ofBordetella bronchisepticain mouse dendritic cells. In-fect. Immun. 62:5528.
16. Su, H., R. Messer, W. Whitmire, E. Fischer, J. C. Portis, and H. D. Caldwell.1998. Vaccination against genital tract infection after immunization with den-dritic cells pulsed ex vivo with nonviableChlamydia.J. Exp. Med. 188:809.
17. Ojcius D. M., Y. B. de Alba, J. M. Kanellopoulos, R. A. Hawkins,K. A. Kelly R. G. Rank, and A. Dautry-Varsat. 1998. Internalization ofChla-mydiaby dendritic cells and stimulation ofChlamydia-specific T-cells.J. Immu-nol. 160:1297.
18. Filgueira, L., F. O. Nestle, M. Rittig, H. I. Joller, and P. Groscurth. 1996. Humandendritic cells phagocytose and processBorrelia burgdorferi.J. Immunol. 157:2998.
19. Van Overtvelt, L., N. Vanderheyde, V. Verhasselt, J. Ismaili, L. de Vos,M. Goldman, F. Willems, and B. Vray. 1999.Trypanosoma cruziinfects humandendritic cells and prevents their maturation: inhibition of cytokines, HLA-DR,and costimulatory molecules.Infect. Immun. 67:4033.
20. Blank, C., H. Fuchs, K. Rappersberger, M. Rollinghoff, and H. Moll. 1993. Par-asitism of epidermal Langerhans cells in experimental cutaneous leishmaniasiswith Leishmania major.J. Infect. Dis. 167:418.
21. Thurnherr, M., R. Ramoner, G. Gastl, C. Radmayer, G. Bock, M. Herold,H. Klocker, and G. Bartsch. 1997. Bacillus Calmette-Guerin mycobacteria stim-ulate human blood dendritic cells.Int. J. Cancer 70:128.
22. Kim, K. D., H. G. Lee, J. K. Kim, N. Park, I. S. choe, Y.-K. Choe, S. J. Kim,E. Lee, and J.-S. Lim. 1999. Enhanced antigen-presenting activity and tumornecrosis factor-a independent activation of dendritic cells following treatmentwith Mycobacterium bovisbacillus Calmette-Guerin.Immunology 97:626.
986 IL-10 LIMITS THE GROWTH OFM. tuberculosisIN DENDRITIC CELLS
by guest on June 19, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

23. Demangel, C., A. G. D. Bean, E. Martin, C. G. Feng, A. T. Kamath, and W. J,Britton.1999. Protection against aerosolMycobacterium tuberculosisinfectionusing Mycobacterium bovisbacillus Calmette Guerin-infected dendritic cells.Eur. J. Immunol. 29:1972.
24. Peters, J. H., H. Xu, J. Ruppert, D. Ostermeier, D. Friedrichs, andR. K. H. Gieseler. 1993. Signals required for differentiating dendritic cells fromhuman monocytes in vitro.Adv. Exp. Med. Biol. 329:275.
25. Romani, N., S. Gruner, D. Brang, E. Kampgen, A. Lenz, B. Trockenbacher, G.,Konwalinka, P. O. Fritsch, R. M. Steinman, and G. Schuler. 1994. Proliferatingdendritic cell progenitors in human blood.J. Exp. Med. 180:83.
26. Kiertscher, S. M., and M. D. Roth. 1996. Human CD141 leukocytes acquire thephenotype and function of antigen presenting dendritic cells when cultured inGM-CSF and IL-4.J. Leukocyte Biol. 59:208.
27. Rook, G. A. W., and S. Rainbow. 1981. An isotype incorporation assay for theantimycobacterial effects of human monocytes.Ann. Immunol. 132D:281.
28. Stenger, S, R. J. Mazzaccaro, K. Uyemura, S. Cho, P. F. Barnes, J. P. Rosat,A. Sette, M. B. Brenner, S. A. Porcelli, B. R. Bloom, and R. L. Modlin. 1997.Differential effects of cytolytic T-cell subsets on intracellular infection.Science276:1684.
29. Flesch, I., and S. H. E. Kaufmann. 1990. Activation of tuberculostatic macro-phage functions byg interferon, IL-4 and tumor necrosis factor.Infect. Immun.58:2675.
30. Hirsch, C. S., J. J. Ellner, D. G. Russell, and E. A. Rich. 1994. Complementreceptor-mediated uptake and tumor necrosis factor-a-mediated growth inhibi-tion of Mycobacterium tuberculosisby human alveolar macrophages. J. Immunol.152:743.
31. Hirsch, C., T. Yoneda, L. Averill, J. J. Ellner, and Z. Toossi. 1994. Enhancementof intracellular growth ofMycobacterium tuberculosisin human monocytes bytransforming growth factor-b1. J. Infect. Dis. 170:1229.
32. Toossi, Z., P. Gogate, H. Shiratsuchi, T. Young, and J. J. Ellner. 1995. Enhancedproduction of TGF-b by blood monocytes from patients with active tuberculosisand presence of TGF-b in tuberculous granulomatous lung lesions.J. Immunol.154:465.
33. Hirsch, C., J. J. Ellner, R. Blinkhorn, and Z. Toossi. 1997. In vitro restoration ofT-cell responses in tuberculosis and augmentation of monocyte effector functionsagainstMycobacterium tuberculosisby natural inhibitors of transforming growthfactor b. Proc. Natl. Acad. Sci. USA 94:3926.
34. Brown, M. A., and Hural, J. 1997. Functions of IL-4 and control of its expression.Crit. Rev. Immunol. 17:1.
35. Yamamura, M., K. Uyemura, R. J. Deans, K. Weinberg, T. H. Rea, B. R. Bloom,and R. L. Modlin. 1991. Defining protective responses to pathogens: cytokineprofiles in leprosy lesions.Science 254:277.
36. Gong, J., M. Zhang, R. L. Modlin, P. S,. Linsley, D. Iyer, Y. Lin, andP. F. Barnes. 1996. Interleukin-10 downregulatesMycobacterium tuberculosis-induced TH1 responses and CTLA-4 expression.Infect. Immun. 64:913.
37. Murray, P. J., L. Wang, C. Onufryk, R. I. Tepper and R. A. Young. 1997. T-cellderived IL-10 antagonizes macrophage function in mycobacterial infection.J. Im-munol. 158:315.
38. Murray, P. J., and R. A. Young. 1999. Increased antimycobacterial immunity ininterleukin-10 deficient mice.Infect Immun. 67:3087.
39. Cella, M., F. Sallusto, and A. Lanzavecchia.1997. Origin, maturation and antigenpresenting function of dendritic cells.Curr. Opin. Immunol. 9:10.
40. Banchereau, J., and R. M. Steinman. 1998. Dendritic cells and the control ofimmunity. Nature 392:245.
41. de Waal Malefyt, R., J. Haanen, H. Spits, M. G. Roncarolo, A. te Valde,C. Figdor, K. Johnson, R. Kastelein, H. Yssel, and J. E. de Vries 1991. Interleukin10 (IL-10) and viral IL-10 strongly reduce antigen-specific human T-cell prolif-eration by diminishing the antigen-presenting capacity of monocytes via down-regulation of class II major histocompatibility complex expression.J. Exp. Med.174:915.
42. Stenger S., K. R. Niazi, and R. L. Modlin. 1998. Down-regulation of CD1 onantigen-presenting cells by infection withMycobacterium tuberculosis.J. Immu-nol. 161:3582.
43. Holt, P. G., J. Oliver, N. Bilyk, C. McMenamin, P. G. McMenamin, G. Kraal, andT. Thepen. 1993. Downregulation of the antigen presenting cell function(s) ofpulmonary dendritic cells in vivo by resident alveolar macrophages.J. Exp. Med.177:397.
44. Pancholi, P., A. Mirza, V. Schauf, R. M. Steinman, and N. Bhardwaj. 1993.Presentation of mycobacterial antigens by human dendritic cells: lack of transferfrom infected macrophages.Infect. Immun. 61:5326.
45. Albert M. L., B. Sauter and N. Bhardwaj. 1998. Dendritic cells acquire antigenfrom apoptotic cells and induce class I-restricted CTLs.Nature 392:86.
46. Stenger, S., D. A. Hanson, R. Teitelbaum, P. Dewan, K. R. Niazi, C. J. Froelich,T. Ganz, S. Thoma-Uszynski, A. Melian, C. Bogdan, et al. 1998. An antimicro-bial activity of cytolytic T cells mediated by granulysin.Science 282:121.
47. Porcelli, S. A., and R. L. Modlin. 1999. The CD1 system: antigen-presentingmolecules for T-cell recognition of lipids and glycolipids.Annu. Rev. Immunol.17:297.
48. Bogdan, C., Y. Vodovotz, and C. Nathan. 1991. Macrophage deactivation byIL-10. J. Exp. Med. 174:1549.
49. Fiorentino D. F., A. Zlotnik, T. R. Mosmann, M. Howard, and A. O’Garra. 1991.IL-10 inhibits cytokine production by activated macrophages.J. Immunol. 147:3815.
50. Moore, K. W., A. O’Garra, R. de Waal Malefyt, P. Vieira, and T. R. Mosmann.1993. Interleukin-10.Annu. Rev. Immunol. 11:165.
51. Enk, A., V. Angeloni, M. Udey, and S. Katz. 1993. Inhibition of Langerhans cellantigen presenting function by IL-10.J. Immunol. 151:2390.
52. Beissert, S., J. Hosoi, S. Grabbe, A. Asahina, and R. D. Granstein. 1995. IL-10inhibits tumor antigen presentation by epidermal antigen-presenting cells.J. Im-munol. 154:1280.
53. Macatonia, S. E., T. M. Doherty, S. C. Knight, and A. O’Garra. 1993. Differentialeffect of IL-10 on dendritic cell-induced T cell proliferation and IFN-g produc-tion. J. Immunol. 150:3755.
54. Morel, A.-S., S. Quaratino, D.C. Douek, and M. Londei. 1997. Split activity ofinterleukin-10 on antigen capture and antigen presentation by human dendriticcells: definition of a maturative step.Eur. J. Immunol. 27:26.
55. Caux, C., C. Massacrier, B. Vanbervliet, C. Barthelemy, Y.-J. Liu, andJ. Banchereau. 1994. Interleukin 10 inhibits T cell alloreaction induced by humandendritic cells.Int. Immunol. 6:1177.
56. Chang, C. H., M. Furue, and K. Tamaki. 1995. B7.1 expression of Langerhanscells is upregulated by proinflammatory cytokines, and is downregulated by in-terferon-g or by interleukin-10.Eur. J. Immunol. 25:394.
57. Buelens, C., F. Willems, A. Delvaux, G. Pierard, G, J. P. Delville, T. Velu, andM. Goldman. 1995. Interleukin-10 differentially regulates B7.1 (CD80) and B7.2(CD86) expression on human peripheral dendritic cells.Eur. J. Immunol. 25:2668.
58. Thomssen, H., M. Kahan, and M. Londei. 1995. Differential effects of interleu-kin-10 on the expression of HLA class II and CD1 molecules induced by gran-ulocyte/macrophage colony stimulating factor/interleukin-4. Eur. J. Immunol. 27:1229.
59. Koch, F., U. Stanzl, P. Jennewein, K. Janke, C. Heufler, E. Kempgen, N. Romani,and G. Schuler. 1996. High-level IL-12 production by murine dendritic cells:upregulation via MHC class II and CD40 molecules and downregulation by IL-4and IL-10.J. Exp. Med. 184:747.
60. Lipscomb, M. F., C. R. Lyons, G. Nunez, E. J. Ball, P. Stastny, W. Vial, V. Lem,J. Weissler, L. M. Miller, and G. B. Toews. 1986. Human alveolar macrophages:HLA-DR-positive macrophages that are poor stimulators of a primary mixedleukocyte reaction.J. Immunol. 136:497.
61. Allavena, P., L. Piemonti, D. Longoni, S. Bernasconi, A. Stoppacciaro, L. Ruco,and A. Montovani. 1998. IL-10 prevents the differentiation of monocytes to den-dritic cells but promotes their maturation to macrophages.Eur. J. Immunol. 28:359.
62. De Smedt, T., M. Van Mechelen, G. De Becker, J. Urbain, O. Leo, and M. Moser.1997. Effect of interleukin-10 on dendritic cell maturation and function.Eur.J. Immunol. 27: 1229.
63. Buelens, C., V. Verhasselt, D. De Groote, K. Thielemanns, M. Goldmann, andF. Willems. 1997. Interleukin 10 prevents the generation of dendritic cells fromhuman peripheral blood mononuclear cells cultured with interleukin-4 and gran-ulocyte/macrophage-colony-stimulating-factor.Eur. J. Immunol. 27:756.
64. Randolph, G. J., S. Beaulieu, S. Lebecque, R. M. Steinman, and W. A. Muller.1998. Differentiation of monocytes into dendritic cells in a model of transendo-thelial trafficking.Science 283:480.
65. Randolph, G. J., K. Inaba, D. F. Robbiani, R. M. Steinman, and W. A. Muller.1999. Differentiation of phagocytic monocytes into lymph node dendritic cells invivo. Immunity 11:753.
66. Balcewicz-Sablinska M., J. Keane, H. Kornfeld, and H. G. Remold. 1998. Patho-genicMycobacterium tuberculosisevades apoptosis of host macrophages by re-lease of TNF-R2, resulting in inactivation of TNF-a. J. Immunol. 161:2636.
67. Rojas, M., M. Olivier, P. Gros, L. Barrera, and L. F. Garcia. 1999. TNF-a andIL-10 modulate the induction of apoptosis by virulentMycobacterium tubercu-losis in murine macrophages.J. Immunol. 162:6122.
68. Zhang, M., J. Gong, D. V. Iyer, B. E. Jones, R. L. Modlin, and P. F. Barnes. 1994.T cell cytokine responses in persons with tuberculosis and human immune defi-ciency virus infection.J. Clin. Invest. 94:2435.
69. R. J. North. 1998. Mice incapable of making IL-4 or IL-10 display normal re-sistance to infection withMycobacterium tuberculosis.Clin. Exp. Immunol.113:55.
987The Journal of Immunology
by guest on June 19, 2018http://w
ww
.jimm
unol.org/D
ownloaded from