Embed Size (px)
Citation preview

II. REVIEW OF LITERATURE

15
2.1. Antennae and Olfactory Sensilla
VJhen considering antennae of different species,
the V8riation in form is striking. To understar:d
the evolution:; ry development of variations i'h size
1 nd shnpe one must consider all of the different
functions the gntennae may serve (e.g. olfaction,
t3ste, touch, perception of air-movement, heat and
C02 ), which vary considerably between species
(5 chne:l der, 1964) • Antennae with feather like
sl-1apes (e.g., in saturniids, lymantriids) seem most
advanced as 'odor filters'. The surL>ce is greatly
increased by thousands of long vertically oriented
hair se:--1silla on the antennal branches. The large
surface area and characteristic geometry enable the
antennae to sieve out odorous molecules effectively
from the air pa :- :"ing through it ( Kaissling, 1971) •
Olfactory sensilla have been classified on the.
b:isis of size and shape as sensilla trichodea, s.
basiconic3, s. placodea (Schneider and Steinbrecht,
1968). In l_epidotera particularly, in moths ani
butter flies the s ensil1 a t ri chodea, Sen sill a
Ch1etica, ?ensil:::.a b8siconica ::md Sensil. a Coelonica
have been sho•m elect rophysiologicall y re sponci to odor

16
{Schneider ~Jnd steinb;-> ··cht, 1968). In males of
Bombyx II!..Q..:d-_, "the s en.sil::_a tr:i.chodea are known to be
extremely s ensi ti ve to the female sex. pheromone
( S c hn ej_ d e r, 1 9 57 ) •
Sensil~.a _qr~a_tics: are similar to sensilla trichodea
but can be distingushed only by a cuticular collar
gt, the base and have sli;;ht longitudinal ridges at
·the base. They oc ·-.::urmainly on the out erolst eral
:;md later:1l portions of each flae;elJar segment. Most
of them are closed type tips.
§_~n~.i-~l.:_q_s;_~~l_Q_conica_: ar~~ receptors in. vvhich the
sonsory portion lies j_n- a pit or depression of-..the
cuticle. Thece pits are always surrounded by cuti
cular hairs. They :.~relocated mostly ·on the
dis tal half of s egmerrc s and groups o.f 3 or 4 are
frequently found on diot nl edge of the segment.
S ensil)_a ___ ~~lo coni ca: are relatively large and occur
ventr1lly on the distal porti6n of the flagellar
segments.· Although they may be lacking on certain
segments but are concentrated on the 3pi90l two
thirds of the antenna.
Seni?..i.J_j_(!_ll~r.:h_ci@c~: are named by jefferson et al..,
( 1970) • They o ccour in relat:. iv ely large number on
the antenna of noctuids.

17
Sensi_-1:)-..£._td:_c_.t_o_de~: do not have cuti cu.lar collar at
the base, short an~i long Sensillae are found. The
short i -:3 type 1, 17 pm long and having blunt tip
·with annular ridge along the whole length. \rw'ith
scanning election microscope three t~'pes II,III and
IV long Sensillae have been distinguished. Type II
have spiral ridge which appears somewhat annular
n,:;ar the tip of the hair. Type Ill have also spiral
ridge but less evident than tho~\e of Type II. The
tip of Type III appear to be smooth. Type IV have
annular ridge near the base. Th::: tip of Type IV is
blunt and distgl portion of these hair.s is carrugated.
There is a row of type IV" on the proximal border of
e:Jch flag·-:=1:-:._ar se6m=:n t .but th :y also occur elsewhere on
the segments.
In Lepidopter8 the SensilJa trichodea have boen
identified as the sex pheromone l"'eCeptors {Albert
;t 81., 1974). lVl.ore thFm one type of sensillum tri
chodc;i..nn is foun::l on the ant enn.a of Trichoplusi8 ni
(Jefferson et al., 1970). Four morphologically distinct
LyJY?S oJ' sensilla trichodea have been identifi ~~d. on the
'·n tenn a of Cho ri stan eura fun:h.,ferana . (Albert and
Seabrook, 1973), althoJ.gi1 only one or.' these. (ty.[)e IV)
h8 s sho ,ffi elr;ctrophysiolo.::;ically to resp·:Jnd to the ·

18
sex phe romon2 ( .1.\l bert et al , 19 74) • Sexual dia morphism
is present in some lepidopteran's antennae (Kais:::;ling,
1971; Steinbrecht, 1973) and many lcpido1Jteran females /
do not posses sensill.a trichodea.
In Bo_~_!_~a_L~ mo ri (schneider· and Kai :=;:line;, 19 57) ,
197"3) Sensil·: ::~ tricbod,~a are present on the anten.'l8e
of both sexes; however it hgs not been established
"Jhether or not, th2 Sensilla trichodea of the ferrnle
responds to its own sex pheromone.
There is a considerable diversity in the number
of sex pheromone sensitive sensilla trichodea present
on the anten'1ae of v::n:·ious species of lepi:h_?ter~.
This is rel8t ed primarily to the form of the antennae.
The larg? plumose, nntenng of BomJ2.yx mor.:_i. posseses
upto 16,000 sex pheromone sensills trichodea.
(Schneider, 1970) where as the small filamen-vous
::JnJ~ ~nr1a of Chari stoneur:J fumi f ~.t:.:~n~l posses es only
2300 sex ph2romon0 sr;;;nsil:..a •rrichod•29 ( ~lbert and
seabro k, 1973) of which only 300 are responied Lo :.:.he
.se~~ pheromone (1\lbert et 81., 1974). The size of the t
antenna ;:md the number of t ri chcd ':8 r(:;fle ct UIJtO the
sen:;::itivity of the moth to its sex pheromone. The
large gntenna h.3s a more efficient catchment areg

19
( K.qi s:3li ng, 1 971 } and trap to odorant molecules of
• much lol'ler concentrations. Hovvever, it does not
r2fl(~ct upon the sensitivity of the individual recep-
tors that appesr to be similar in their threshold
sensitivities. sexpheromone Sensilla trichodea are
i.nnerv8l:.ed by a number of primary sensory neurons.
In Bo_.:_~ mori ( Steinbrecht, 1973) two neurons inner----- ..
vat.· the sex phe~ romone sen sill a t ri chodea. El ectroply-
siolu.gi c:::l Lly 3 neurons respond in Argrot eni3 v el~~i-~~na,
although O' Connell ( 1972) sho~vPd that only 2 of' these
are chemosensory.
In light microscope, five neurons ap:)ear to
innervated the type r'v. Se;1silla trichodewn of
Chodst_~'fl~~ra- funif~:r.~G3. (Albert, 1972), althouc3h not
all of these r(-'spon:l to the sex pheromone (Albert et
al., 1974.). Other types were later described in a
nurnber· o.f' sp!~Ci'38 ( Altner and PriL .. inger, 1980;
Steinbrecht and lVlull.er, 1976}. The first two types,
s. trichodea and s. basiconic.g, are hair-shaped
sensiF.a. The s. coeloconica type, also called a
pit-peg, consists of a peg embedded in a cavity of
the cuticle which opens to the surface. s. placodea,
or poreplGtes: consits of a circular
structure 'J~hi.ch is perforated by pores. Hovoever, n

20
on the basis of exL. emal mo rpholo0y. Internal
morpholo:;,-ic31 properties revesled by studies of
fine strJcture do not alvvays correlate with the
outer shape. For example, S· tri.chodea and .S.
b3siconica usuaLLy differ in their long an·3. short
h~irs resp2ctively, but not al~Jys, ani in this C8Se
th" char.1ct<?r of their fine structure is the prlncip1l
·lifference between the two types (Schneid·-,r nnd
S"c.einb.cecht, 1963). Furthermo.re, intermin<~;led wi-'c.h
S· trichodec1 and S. basiconica in B. mori ar·e found I
sensilJa which .sho''" a fine structure of an int.ermeC:.i'J~e
type (Steinbrecht and l"luller, 1971). 'l'he limited vo1ue
o.:.· ...lsi ng the ext ernsl morphology for cla ssi lying
sensil' a is discus sed by Altner and Pril.l_inger ( 19d0).
One sensil iJ.m type for wh.!. c l1 t !w fine st .cu.ct ur•:: is
well studied is S. trichodea of the silk mo'ch ~· mor:,i_
(StP.inbrecht and ~1uller, 1971; Steinbrecht, 19dO).
This h;;:Ji r-shr:iped sensillum ( Fig.1) can· be ch~r13cteri ?:ed
gsa thin-walled (0.) pm at half len~th) cuticular
protr.uston (10 p.rn long, dianeter ca.2 pm). The
vvsll is pPnertr'ted by numorous pores (ca.250C;) over
the entire suxf'.ice, e:-ce:ept 3t th2 most proxirn'J1 part
of the h3i r. The h:.dr is inL ervated by one or more
sensory cells, vvith dendrites penetrating the hair

·· ······· dendrtte branch
.......• pore tubulus
-------- receptor lymph
------ culicuiar wall
··----outer dendrite segment
-------cuticle
------ tormogene cell
Fig. ! : Scheme of a hair~shaped sensillum, showing one sensory cell, embru.ced by the three auxiliary cells. The dendrite of the sensor'y'~dl is divided into a branched outer segment and an unbranched inner segment (containing'Nl'itochondria). The conduction system for the odor molecules (pore and pore tubules) ma'kcs·c011t~ct with the dendrilir. membr.Euw.

21
lumPn ;:md re:::~ching out to the tip. Each dendrite is
divided into an inner and outer s::.egment, separated by
a speci;d cilia r structure which is. short and 'neck-like'
::Jnd contains the typic8l concentric doublets of tubules
found in other sensory cilia. It is only the outer
dendrh. e s egt:J ent which resides v~i thin the hair lumen.
The tvw sc:>grr.ents also differ with respect to the
contents of cell organ~lies. Whereas the outer
s egwent s m::J i nly contain mi c rot ubul e s and no mit :co chondri a,
the latter are abundant -,vithin the inner segment. The
outer segment is usually divided longitudinall)' into
br·anche•o, rC'~lat.iv;:-:ly fevJ inS. trichodea, but up to
50 in others, e.g. S. basiconica (Fig.2). The inner
segment and the cell bodies are surrounded by thTee
auxiliary cells (the the chogen, to nnogen and trichogen)
wh~- c h form a sheath around the sensory cells. Thus an
olfactor:,; sensillum consists of at least four cells.
The auxiliary cells, ~~hich carry out an import ant
:-ecretory function during the ontogenetic development
of the sr:>nsilJ um,' forming h;::~ir cuticle and the microtubles
(Ernst, 197?), may have another important function
during adult life. Th~y apJ,arently msintain the
electric:-:11 potential (ca. + 50 mV) between thE! haemolymph
"lild the receptor lymph (Kaissling and Thorson, 1930).

r·~-"'l''"-...,.,.~·····-·· k .: •
r~-
. Fig: 2 : Transmission electron microgrJphs of Bomhyx mori sensWa trichodea (above, x 20 000 and 40 000) and s. basiconica (below, x 20 000). Fixation by frer.:e substitution results in almost round cross-sectional profiles of the dendrites, cor:taining a teguiM array of microtubules. Pore tubules are well fixed in the outer parts of the hair: .. ~ (courtesy of Steinbrecht, l980).

22
. The receptor lymph fills the hair lumen around the
outer segment of the d~ndrites and is separated from
the ha emolymph by tight junctions formed between the
au.xillary cells jupt belo1\ the ciliary structure.
ThP pot entic;l of the receptor lymph, which has
relr:Jtively hi,r;_;h K+ contr•nt, is vrobably maintained
. . + uy an elect~lCJgenl.c K -vum]J locc3Led iu thr; folded
mr-•n,branr"s of the trichogen 8nd tormogen cells. The
membrane fol>lds are only present on the side facinG thE.:
receptor lymph. The sensory and auxiliary cell. s are
loc8ted .dthin the epithelium which is separated from
the haemolymph by the bas;:!l membrane. An axon (0.~ pm)
leading from a sensory cell to the brain is s~rrounded
by gli8l cells, and does not interconnect with other
gxonr, either by fusions or synapses, before re.schin[;
th~· br::1in (Stf~inbrecht' 196?).
The co ndu.ctio n system t~1rough ··~hi ch t hr-: orlo r rr.ol ,c.-
cul es reach the dendrite membrj n e or the sensory .;; ·-~L:
ClC ttl' in t1·JO pri.ncip.1lly difi'erent types (Steinbrr·'·:ht
• l·::>qdins from lon_;itudin3l grooves on the h:-lir 3 urf·,c·.::
to '1n in·-:. er cylinder that contains the dr.:>nd~it es
(Steinbrecht and. £Viulter, 1976). However, the conduction

23
system hAs been rnainl y studied in the ot~.tel~ type
(Steinbrecht and 1v1uller, 1971). Here, the molecules
a 1~e adsorb e::l. on the cuticle of the hair w:Jl:. (FiG· 3)
and diffuse through the pore openings ( fun:1el 100 nm
;vide) vvhich oft en l.ead into 8 1i ne ( S run) anJ
,hort ( 25 nm.) pore charr:1el. This 1videns j_n'.::.o a
cavity ( 50-10,! nm). 'l'h:::~ inside of the cavit.y faces
the r'"'CetJtor lymph of the liquor ch-~nnel (200-40!) nrr1
in leoght) ·which penetrates the remaind<'r of the
inside of the cavity 3-7
fine porR tubules pass through the liquor chan:1el
and the hair lumen to the dendrite membrane.
"©.c:>ct.rJn microscopical st.J.dies have est~blished
th~t contact exists between the_ 0 e tubules and
t.he dendrite membrone (Stej_nbrecht and Muller,
1971). Furthermora, Ernst (1969) h9s shown th8t
tr:;cer subst1nceQ penetrate from the outc-ide
through thF: pores into the pore tu.bulPs, g rout:>
by 1'>lh~- ch odorous rnolPcul e::: c3n evidently re:1ch the
dendrite meulJr.cme. Calculations by Ad~m and
Delbruc;_<:: ( 1968)- sug .est a two-dimetts.i.on:-Jl surt'3ce
di ff'..J.sio.:. of the odorous molecules throu0h the
pore-tubule conduction systEm~
•

cutlcu\ar waH
receptor ceU 'dendrite
lli.a.. ___ 3 :
surrounding air . ______ pore
---------- __ . -- pore kettle
1000 A
·internal fluid
Schematic diagram of culticular wall and dendritic branches of a sensillum basiconi.cum of carr ion beetle, l'lecropho~.

24
'!'he specificity of insect olf::1ctory rec~ptors
.has bnpn studied in details by O'Con_ell ( 1975).
This is ab,.olutely relC:ited on the perception of
sex pheromone:;. ln T-GAGS recording from the antenna of
Bomb_E m..?_:ri_ Schneider ( 19&2) found that there were
no qualitative differences in the respone to the
odor of phe-romone~ gl3nds of various saturnid moths.
The differences were fo0.nd only in the rela"cive
gmplitud·-"s of the response::-;. Schneider et al ...
(1Q64) reported by using extra c~llular r~cording
techniques that 2 to 3 neurons were co nstr1ntly
firing as background activity in s ensil2.a t ri cho·dea
of _A_nthe1:~ea and these celLs responded by an incre:Jsed
r1"c.e of dischar,;e to stimulation by the se:xpheromone.
'1'hey further noted that in the s snsill3 basi coni ca
the longer the hair, the fev-ver the compound.:, th·1t
~>t:."i.:nul3ted it. These finJin~~ led to the concr-f't
of oJor generalist neuruns (cel.i.s) and odor ~pc-cL~Jli:;0
neurons (cells)(Boeckh et al., 1965).
Odor generalists art=; neurons of the S8me sen
sillum type in one individual, which h~ve different
but partly overlap~Jing reaction patterns j_n aJ 1

25
re1cting cells. Examples of such odor journalis;,:,s.
1rr.; sensil i_a basiconica.
Odor specialists are olfactory neurons belonging
to 8 rc:;cognizable type of sep.silJ.um, giving sterotyped
responses to 8 seri<os of compounds. Example8 of
odor spr:>ci~lists a.rf~ thp, cRc:::ptors for sex attra·ct::mts.
Schneider et al., ( 1Q64) noted in Antherea that of
3 cells in comnon sensill u.m basiconicum, one may
be:: excited one m'1'Y be inhibited. CX.YJ.d one may show
no response to a common olf3 cto ry stimulus. A
sirnila r phPnomenon has been found in the sex pheromo)')e
r~·c epto r s ens ill urn of Tri chop~~sia ni.
Kais~:u_ing and Renner ( 1968) by using single
unit recordings ident,ified tvvo cell types respui1ding to
the pheromone of the honeybee. One cell res:Jonded
to scent from Nasanov gland. Vnreschi ( 1971) further
·separated 7 cell types with no overla}> in B}Jecifi-
city; ho•~ever, there was some vari:lt:.ion of speci:.:'i city
within e3ch of these cP.LI types. SchnE:ider 0t 31.,
( 1()64) notod th:::lt gt=?omc>tric isomer·s of bombykol ~'>~ere
u ltO 1000 times l e:;s pffectiv e at the bombykol re c optcr
th;m thP true phenomon~ ( bomhykol) • The sa?:ne rule
>p;Jlied to Tr~.SJ2QE.l!!.:?J.2 ni '•lhere :)8yue et c.ll. ( 1q73)

26
reported th?.t. the more a parapheramone vnries in
its structure from the structurc-; of the true pherommone, " the greater the amount needed to elicit a significant EAG.
This infonnation indicates a considerable det;ree
of receptor cell specificity to compounds of closely
related chemic.'1l structures and 1 end support to the
concept of odor-specialist nPurons. O'Con:·1ell ( 1975)
by using computer an::llysis of single-unit neuronal
rt:;spon!::Oes in sex pheromone receptor sensil.._a of the
red bonded leaf roller has indicated that the two
cells responsive to the sex pheromone and number of
its anolot;ues do not respond in an identical fashion
to each of these va2ious compounds. He found that
the response freqw:"ncy to the pheromone and to six
other behaviorally active compounds varied be:::.ween
the tv-vo cells and cL3ims that these diL'r:;rences in
rP.sponse were due to intrisic factors within the
neurons. He argues that the encoding of odor quality
i ~'not simply in the pr1'~seric e or ab~ence of activity
in anyone neuro, but is encoded by a pattern that may
vary across an ensemble of receptor neurons to produce
~ unique total response.· 0' Connell ( 1 97 5) states that
·as some neurons can respond to 8 out of 9 b~;haviorally

27
active compounds, there could be as many as 8 fun
ction:ll :_y dif1 p,·~nt receptor sites. He further
concludes ·chat because one cell responds prefe.Len-
t;ially to cis-11-tetrAdecenyl acctat;e (the pherGmone)
and the second cell respond. s preferentially to
tt3ns isomer (1n inhlbitor), there mu:t be ~t lP1St '
tvvo function~::lLLy distinct rt:;C~~ltor sites. Phercrnone
receptors do reSJ:Jond to the isomers r:md analo~ues
of Jche ap,.'ropriate pheromone. This indicates that
gl·chough the receptor may be more sensitive to
the true pher·omone it does r(~spond in a ~vesker
fashion, to the=e ot~e~ compounds (Scnneider et al.,
1C164; Pgyne, 1971). Kaissling ( 1976) has propo:3ed;
t rnt the 8p 9ro p ri ate molecules for a given a c <' epto r,
r·.::2ult in large, i rregulr-J rl:; timed elementary
gene rAt or pot enti:~l~3 • Er~ch potenti:.:Jl co rr:; S.fJOnds to
an individu<Jl stimv..latory molecule encounter with
the ap rep ri 3t ~' acceptor.
On the othc r- hand, other molecules, wit:1 some
structur8l relationship to the correct molecule,
res:..1lt in smooth receptor potential::; wi:,i<.;h aric.e from
the surnmatio;1 of ·many smaLi.er ele1nentary 'generator
potenti:;ls.

28
Seabrook's ( 1977) study showed th~•t the neurons
of the sex phpromone rr:::ceptor of male Choris~~neur~
£~~!!_i_(c~_r:r!Jl·~ r ·;.Jpond to both its sex pheromone, a
mixture of cis and trans-11-tetradecenal (sanders
and "1-v2atherstone, 1976) and 'co trans-11-tetradecenyl)
8cr~tntr; 8 kno•vn inhib:Ltor of the beh~vioral r~~ponse
to the s:::xphrrornone (Sanders et al., 1972). Seabrook
( ·197'7) demon::,trat ed in his cro'ss adoptation exp,eriments
th3'c both the aldehyde and the acct ate are inter
acting with the same dendrite acceptor site. Boeckh
et aL (1965) shovved the same interaction in the honeybee.
Th~ pheromonl?- receptor when a:.tapted to the
phr->romone :~1 ~::'0, is 8dapted to caproic acid. Payne
and Dickens { 1976) have shov~n the aggregation
ph-=:romone of TJend~.J;_~o-~~ £rontali~,inacted with
the acceptor for all other cofnpou.nds tested.
Thus it lhGY be a:::;r_;umE-~d that as an acceptor may
respond differenti31 Ly to certgin stimulatory
molecc...les, a specialist rec·?ptor sensu stricto
does not exist.
Here, the minor pheromone components are phero
mone inhibitors ;=uri synergists. Pheromone inhibitors

are those compounds which reduce the tra ppi_ng
efficiency of a· ph2romone or rt~duce the behavioral
t~. sponse o.t' 8n insect to its sex pheromone \vhen -
compou.nd is rele1sed along ;,vith the pheromone.
Pheromone synergists are those compounds which
enhance the rPsponse of the male insect to the
p hero mo n e so uc c e.
Inhibitors and synergists c~n operate at
Pxtremely lo1v concentrations and are often isomers
of the rna jor component.
Inhibitors have be en described for a number
of insect species (Hathaway et al., 1974). In
some instances, the inhi.b:i."c.or is produced by the·
s~1me insect gs is the pheromone and masks the
effect of the pheromone (Casida et al., 1963;
Shorey and Gaston, 1967).
The inhibitor may serve to chemically isolate
tvvo species with a common pheromone and with over-
lapping geographicGl ranges (sower ~tal., 1971+).
There are 3 potent~al neurological mechanisms
for the perception of inhibitors.
1. .rtec:eption by separate sensory neurons.
(MaclaYiughlin et al., 1974) and thus the information
is car,·i(·;d by di::;crete information channels to the
olfactory lobe or deutocerebrum o.r the brain.
2. rtec epti on \;y ac.eeptor molecules, lo cat -=;d on a
common neuron (in this instrance, the pattern

JO
of action pot ::'ntinl dis charge vmul d code the
signal) and
3. neception by a common acceptor in which C3se
the pattern of the acti!l>n potential sequence
co,ld code the information or a binding between
pher.o:none 3nd inhibitor could occur.
sov.Jec et al., (1g7h) have shown that, the males
of Plodia ~-0~ er~)gn_§_t. e~l~ and Cadr§_ cautella can
be ln bi tu.r; ted to inhi bi tors of the sex pheromone
respc5ns e while subseyuently showing a response to
thE; SE~X pheromon·' plus the inhibit-or. A 3 minutes
pre-exposure to the inhibitor trans-trans-9-12-
tetradecadien-1-01 acetate produced larger increased
rP.sponses in both species on subsequent exposure
to the pheromone (trens-tis) 9, ·12-tetradecadien-1-01
than the response of individuals that m d not been
pre-treated. Here the main interpretation is that
the pheromone and inhibitor were being perceived by
-separate sensors. The male of Trichoplusia ni
eli cited EAGS by expo sure to the sex pheromone
inhi bi tor t ra ns-7-dodec en-1-0 1 vvhich were similar
to. EA.GS resui ti ng from exposure of the antenna to
the pheromone trans-7-dodecer'l-1-01 acetate (iVi'ayer,
1973). Trichoplusi a ni and E;:;eudoplu.§..ia ~~ludens

Jl
upe the same pheromone. M.aclahughlin et al. ,. 1S7ld
found that the inhibitor did not interfer with sex
pheromone behavior L1 Ghat males which showed an
upwind flight behavior and clasper ext ens ion when
exposed to the sex pheromone. If did, however,
p rev .:-:nt the moths from o t~i enting to a lo cu.s of
at-.::.raction when the inhib].tor source W<:;s wh,hin
30 em of the pheromone sou r·c e.
Above conclusion is that the inhibitor is
perceived by a sensory neuron separate from the
pheromone receptor neuron, and that modification of
pheromone-elicited behavior is not. peripheral but a
central phenowenon. ·rhis phenomenon is also sho.m
by Knis~ling (1071).
R:::Jelofs and Com~au, ( 196~ 1971) reported that
compounds structurally related to the sex att.ract:mt
for the red-banded leaf roller moth cis-11-tetradecany-
lacota·._,e can have either a synergistic or inhibitory
ef.Pct on attract::mcy in field studies. Thr=y proposed
~that th8se structurally relat8d molecules are intPr-t
ncting wtth the bindin~: sites on the similrJr rr-,ceptors,
vvitll v::1rying dc-;::;rees of affinity.
In single rect=·ptor recondings from the pheromone
rPc eptor of .1\.rgyrota eni a v el.Jtinana, 0 1 Connell ( 19 72)

32
reported that the same two neurons responded to
the sex pheromone cis-11-tetradecenyl acetate
to ·an inhibitor (trans-11-tetradecenyl acetate)
and synergist (dodecyl acetate). The response of
the sensory neurons to st,imulation by a mixture of
cis-11-tetradecenyl acetate+ trr:Jns-11-tetradeccnyl
acetate was d~creased in the activity of one
neuron when compared to stimulation by an equial ent
8mount of the pure cis-11-tetradecenyl acetate.
The response to cis-11-tetradecenyl acetate
+ the synergist (dodpcyl acetate) 'was an incrn;;~se
in neur':"-Jl activity. In a subsequent study, differential
r ponsPs of the tv~o chemosensory cel~s to the
sex pheromone 3nd 7 other beh8vio rally active com-
t)''tl;!d..J led O'Con.'ell ( 1975) to conclude thst
s•:?parJte acceptor:::. are present for the cis-11-
'JC~et~Jte. Roelofs et al. \1975) shovved th·'lt all
thrr.~'- com. ounds :1 re pre~~ ent in the c~1 1 in<.:.~ fe:n·1l e.
by usi.n;; :S;\G.S, conformed O'Con,le.~.i. 's vir.:>'" t:h:n:. the
two j_somc'7rs are pl?rcei.ved by two fun~L..i.cJn:.~lly different
acceptor sites.

33
A similar situation vJas found in Adoxop_J::!Y_es Q_£_ana,
in ~vhich one of the 2 sensory neurons responded to
ci~3-9-vetradec:enyl acetr1te and 2nd neurone responsed
to cis-11-Letradecenyl acetate, which_comprise the
moth's pheromone in a 9:1 ratio. No neurone w8s
turned to this ratio. Thus the determin3tion of
the opti;:mm blend is D central nervou~:: system pheno-
menon (Den Otter, 1976).
Adoxophyes orana uses tr~ns-9-and ~r3n-11-Ce~radecenyl . -----.,..---- -·--:- ---
a c et 8t e, t. he geomet rl cal is orne r.s of its pher·om.one as
i'cs inhibitors. EtiG responses to mixtures of the
pheromone and its inhibitor showed no aLi.1itJ.Ve
effects vvhen comps red •1i'c h EAG !\-; spons es to the
pheromone aione (IVIinks et nl·, 1974).
of the sex ph.:;rornone trans-11-tetradecanal and the
inhibitor trans-11-tetradecenyl acetate are perceived
by the same neurons and react with the same acceptor
molecules (Seabrook 1977). lt has been suggested that
the n -,u ron ad.apts more to the inhi bi tor than to
the pheromone.
This differP.nti<-1l ad8ptation of the nrouron b:; the
two compoun,ts indicates a clii'fet~ent affinity of each
of t.he·~e mol ecul2,':' for the ccmn:.on acceptor ( ,3eabro·)k,

34
1977) and sUj)i.JOi~ts the view of Roelofs and corneau
( 1971) and KaiS'sling ( 1976) for the responses of a
neuron to structurallysimilar compound.
It is of int.erest to speculate on the potential
beh?.vioral role of the so Cc.llled inhibitor compounds
· tha'c supress the activity of sPnsory n·=.,urons. It may
b~ thg",; th~y do not inh.~bit the beh::~viorol rP::-:-ponse
to Lhe pher'Gmone but that they may act as flight
nr:~est::mt,_,, initiating the short range activity between
the male and fan.sl·e moth. This has been demonstrgted
behaviorally ( C"'rde et al., 1975) by the a1j.dition of
do~lecyl al cohc-:1 to the IJheromone blend for the o ri ential
fr:.:it moth. 'fhe:::e com.i:Jou.nds are often present in small
3mourr'cs in the pheromone blend (Roelofs et gl., 1975)
and therefo ce \1\;'0uld only reach a b ehaviral threshold
at di~:;tances very close to the emitting female. In
Choristoneura fumiferan_? the sensory neurons are able
to perceive c0ncentration dif_,_'erences only during
th-:: first 200 ~(lsce of the exposure to it::; pheromone
(;:;e::~brook, 107·/) and ther~fore are not lik~ly to
be able to percei,(e a concentr!3tion t5radient or to
determine their pro}:irnity to 8 female based on the
cone entration of the mnjor pheromone component •

35
Recently more attention has be en devoted
to the study of minor components of the pheromone
system. Thes (= may occ:our as a part of the insects
pheromone system {Young et al., 1973 ·; Minks et al.,
1974; and Roelofs et al., 1975). In addition,
there rn3y be synthetic structural analogue of
the pheromone (Eoelofs and Comeau, 1968, 1971;
Co;rde <:.t al. ~ 197 5} , t hs t when mixed i'Vi th the
pheromone, increased the behCJvio ral response
of the insect.
R.o r-::lofs and Come om ( 1968) reported that
dodecyl :1cetate w.gs str'Ong;ly synergh>tic in HiBluHe
in traping tests with the Red banded leaf roller
moth and they subsequently li~ted other 11 - and 12 -
carbon acetates as synergistic to RiBLURE (Roelofs
ana. (o.me3u 1971) • Similarities in molecular structure,
they sug,;ested that these componds may be reacting
with the same receptor site as does the pheromone.
Little el~ctro-phy~3iological evidence is avail:Jble
on the perception of the minor pheromone components
and synergists. O'Connell (1972) reported th1t
dodecyl acetate alone had little effect on pheromone
receptor activity in the Red banded leaf roller moths.

2.3: Mo.l::EhQ.~'?..gzL_aQ.d Histology of sexpheromone glands of Female .fuloth:
J6
The c; bdomint.ll tip of female moth containing 7th, Sth and
9th segments, is densely closed with scales and Mirs.' Setae
are al ivays on the posterior rna rgin of the 8th t ergum and on
the valves and minute spines cover both scl erotized ana
unscl erotized a rea. Normally, the Sth and 9th segments are
telescoped into the 7th segment. They are·retracted by longi
tudinal muscles which hnve their origin in the 7th segment and
are inserted on the valves, the common eviduct, the apophyses
anteriores, and the apophyses posteriores. T·wo sets of
protractor mu8cles are present. One set h~s its origin on
the posterior mar;in of the 7th tergum and is inserted
on th:! anterior end of the apophyses anteriores, and ·the
other set ori6ina~es on the posterior margin of the 9th
.ter;um nnd is secreated on the anterior en~ of the
apophyses posteriores. The gland which produces the female
sex pheromone is Yeversible of glandular epithelium
situated dorsoposterior and ventral region of the inter
segmental mel.llbrane bet~veen the ·8th and 9th segments. When •
the 8th and 9th segment are extruded the gland ap~ears
as a white sac or vesicle above the valves -which is res-
ponsible to produce sex pheromone. VJhen the abdominal
tip i~ in its ext ended calling position, the longitudinal
muscl'."!s are relr:~x.=>c:l. and cuticUlar oblique ones contracted
the distances between the strB~t\tions of the longitudim 1

37
and cu ti cul ar oblique muscles ext end strongly for
their st3tes of relaxation and contraction respectively •
.Uternating relaxation and contraction of longitudi-
nal and circular oblique mu~3cles are essential to.
the rhythmic protraction and retraction of the
abodominal tip during c·alling.
For the identification of the sexpheromone glands,
the external morphology of the concerned part is shown
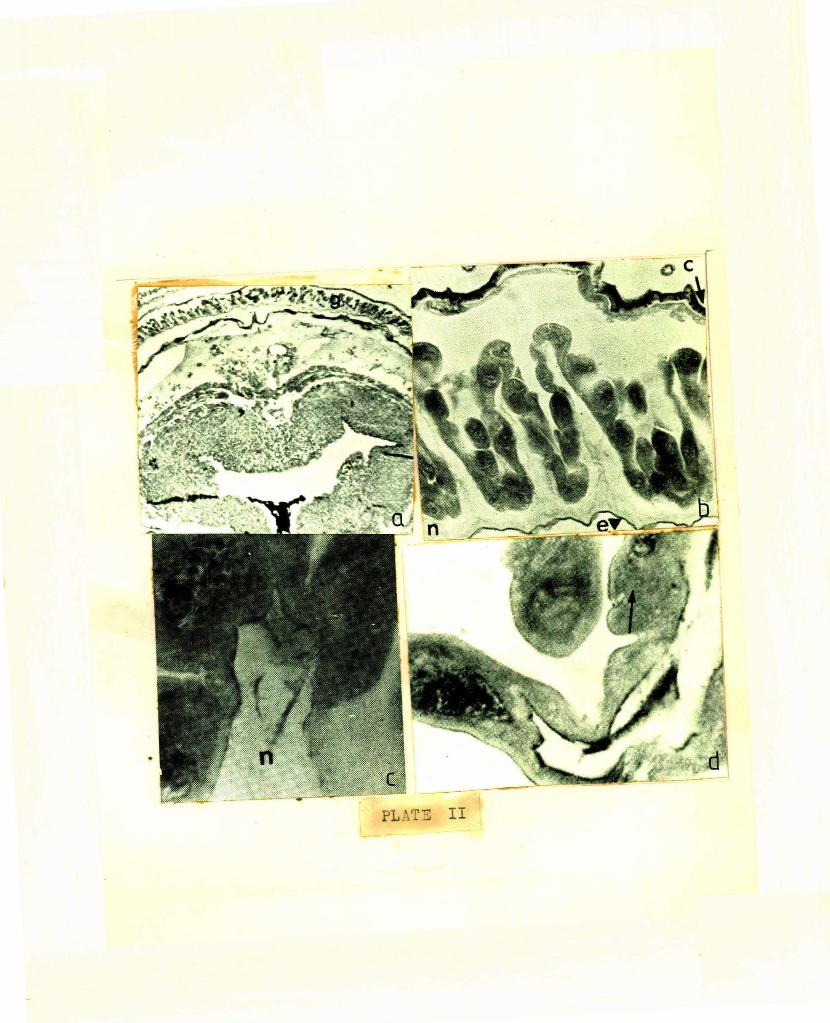
under sc8n11ing elect·"'nmicroscope (PLATEI b). The
dorsal vievv of the intersegmental meubrane shows a
well defined a rea containg a highly convoluted
i nt egumenta ry st rlil.cture which seems to be strongly
indicative of its glandular nature ( PL A.TEIG) .
The ventral aspects of the intersegmental mernbeane
between the 8th and 9th abodominal segments is also
convoluted (PLAT13': I d). The lriteral aspects of
the membrane betlfjeen the 8th and 9th segments desc-ribe
as smooth in comparision with the topography of the
dorsal or ventral aspects. Histologically, in the
c~.:iLl.i.nt; fanal e moth, the rhythmical protraction
and retraction of the abdominal se;9U mts reveals the
presence of a modified intersegmental membrane. The
modified area is located dornally between the 8th and
9th abodominal segments (PLA'l'E II a). ' sex pheromooo

J8
See PLATE I a,b,c,d

PLATE I:( \.1. Hollsnder, 1932, J.Insect physiol, 28)
a: Lateral view of extended abdominal tip of female moth. With Sth &. 9th abdominal segments (8 \S .&, 9AS) arrl the intersegmental membrgne (LVI) in between (scale bar 1 mm) .
b: scanning electronmicrograph of extended 3bdomin31 tip. sho.-Ji ng dorsal and ventral cuticular convolutions (arrows) of sexpheromone gland. (Scale ba1:.. = 0·4'5 mm).
c: Scanning electron micrograph of cuticular convolutions (arrow) of the sex pre romone gland located on the dorsal aspect of the. intercegmental membrane between the 8th and 9th abdominal segments (Scale bar= 0•5 mrn).
d: Scanning electron micrograph of cuticular convolutions (arro·w) of the sex pheromone gland located on the ventral
. aspect of the int ersegrnental membrane (Scale bar= 0•5 mm} •
•


PLI\TS I I : (J. _; . Percy , 197 1 , Can. f.nl.. . 103}
a: cro ~s section of 8th a bdominal s e sznent of f Amale moth showi ng dors Rl cresc ent shn p ed . S ex ph e romone gland ( g ) x 35.
b: Cross section t h rou.gh s.ex pheromone gland to s how the U sh~ped arrangement of the modified eqiderma~ cells. e, epicuticle; n, endocuti cle; c. cuticle of Sth abdomina l s egme.t x ~ 50.
c : Br.3nching of endocuticle in neck of epi d ermal c el ls of setr pheromone gland X 1000.
d: Goblet shape d glandul ·· r c el l . The arro w i ndica te an indi s tinct cell me.mbrc.: ne x 1300 •
•

39
See PLATE II a, b, c,d
' ..
I . · ~
. . , • f
PL . T3 II

40
glarrls th:,t have been studied in the female lepidop
tera are located in the intersegmental membrane
between the 8th and 9th abdominal segments. Generally
in the moth, the glandular area is cr~scent shaped
( PL :\ TE II a) •
Stnined sActi :)ns of abdominal tips reveal tmt ·
the epicuticle ave:r•laying the gland is smooth and
devoid of scales, and the enQ.ocuticle is considerably
thicker th:3n tmt a overlaying · normal membrane
{PL.AT1l: II b). As in PLATE II c &. d, the modified
epidermis of the glandular intersegmental membrane
ccnsists of goblet shaped cells, but columnar and
cuboidal cells are also present. The nuclei of the
cells' are i-rregular in outline and usually contain
one nucleolus. The cytoplasm contains many tiny
vacuoles (PL~TE II c). The number of cells increases
by a U shaped arcangement and the endocuticle pro
truders into the folds created b:;r.this arranf:;ement.
The endocuticle also penetrates the necks of the
cells wrere it gives branches further. These small
branches appear to be' closely associated with the
cell membranes of the necks {PLATE II c)".

41
The communi cation system in .the moth as
shovm schematically in Fig.4 illustrates the extent
of the p ~·es~nt und er·s'ca ndi ng of the different fun
cL.ions involved. No atL empt will be made here to
prPsent a complete picture, but rather to emphasize
sCJme functions and to present some examples.
Some time after emergence f.rom the pupa, the vir
gin fe.nal e moth ~""pands hey sexual attractant glands.
The perform::mc e of thi-s "calling" behavior 1 Fi·~-5 .)
depends upon a nwaber of factors. .As a rule, cglling
is only o bs~rved before copul3 tio n. In the female
of the domesticat·.?d silkmoth Bomb)%, calling is more
or le~s continuous, .vhile in mo:-t other moths s~::..uclied
C'Jl:J.ing depends upon a diel rhythm 111hich ap,•ears to
be correlated l.Vith the activity rhythm oi' the m0le
(1:"lrtell, 1963; Traynier, 1970; Shorey, 1973;
Fatzinger, 1973). In the 'ivell studied cabba ~e
lo .. per moth Trichoplusi3 ni , it was fot:.nd that the
ciuratic-n of calling and th~~ amOtlnt of attract~nt
ewi tt ed depe nd8d upon the ,>Ji nd speed ( Ka a e and
Shorey, 1972). This ph,:,.nomeno ::1 can be understoud
in relation to the distribution of tre luring odor.

Fig, 4:
Ph.R. C II Mol .
Functional scheme of the pheromone corrununication systera between a female and a male moth. Arrow indicates the di.:rection of the information flux. Small symbol s (circles, triangles, squares, pentagons) symbolize odor rnoh;cules (filled) or respective precursors (open). Open niches in the receptor cells represent the acceptors (receptor sites). Ci,iS = Centra 1 nervous sys tern; ~"~1ot(!f) = expansion rn0chanism for the lure gland; Mot (d') ::=motor system (wings and legs) for approaches to the female; Mot. PhoGl = Motor system
. to expand the · male hair pencil gland j Ph.Gl = Pheromone gland ( cfY and !;? ) ; Ph.R,C. I and II ~ 0') receptor cells for two female sex attractants; Ph.R,C. (fl.) :::: rec~Rtor Cl'~ lls for two fern ale sex attractants; P l.R .c. ( <;f. ) -= Plant odor receptor ce ll.

Fc·Jll:t l ,. i'.YI':;y Ill<> ill (./.<ftll.rJJI.t·r.r d.i::J>.tr)
\vil.il l't:p:tndc·d lurl' l',io'lll<l. i"llgt.ll or In c:1JI Ln1'. Jll>::iti()ll l'l1e [orCI-'ing ·- !0 P:n.

L~2
In some cgses (Shorey, 1973), tha f'Amale moths I
fly to the l-3J~val food plant before they start
c·l.Ling. A particul3rly striking food-pL:mt odor
depc~ndency was reported in hnther~ea ES?l:Jphanus
( Ri dai ford, 19 67; Ridcd. fo r\.i and williams., 1967) •
The f~male of t·his moth only called in the presence
of the odor of the preferred food plant; oak. The
compound 1.., t-2-hexenal, v~hich is found in most
greRn leves. Pure hexenal brings about the S3me
e.i:'.':'ect .as oak leaves, while other leaves- il). spite
of the fact that they contain hexenal - clo not
induce c2lling. 'l'he odor signal is perc~i'f' 0 d vvith
"Jntc:mr1Jl r,·.,c ~pto es. With regard to the failure
01 leaves such as birch to elicit calling, one is led
to 8S sume th-, t le·w es of plr.Jnt s other than oak con-
tain, in addition to the omnipresent h 8..X enal,
some other compounci s .YVhich prevent calling. The ex-
periments oi' Riddiford and Williams ( 1971) h;'ve
enabled us to advance a t"ontative func1:ional scheme
011 thE> basis of e:xp(~rimr-;nts vvith AntheraerJ an<1 other
ac~.-ivate neurosecretory br.::..in cells, the Tr.:ons ol"
which run to the corpora cardia ca, wb ere t h0ir

odor rt>ceptors on an\(L"nna
brain neurosecrel. cells
~- cardiaca
C. alia\ a
ganglion motoneur..ons
lure gland rtttracled
and el'lponded
Functional scheme of the olfactory, nourohormonal, hormonal, andneural control of the calling behavior of the female giant silk moth, ~-~-~~ uolyphemt22..

43
neuros~·cretion subsequently activates. local hormone
cel.ls. The latter prodi.J.ce a '•calling hormone" ~"hich
circulc;tes in the' hemolymph and is the prerequistite
for expansion oi' the lure gland• This gland is
shifted outw:::Jrd by hemolymph pre8sure as the result
of n1nsrular contractio sin the:; 8bdo111Prl, and can
be r"-'trnct,ed by a retractor mus~le. Since hormonal
effects are ~lov~ and lasting, quick expansion and
rstr8ction of the glands arP presumably control] ed
·through neural pathvvays.
'I'he glt3nds which produce the female attrsctant
pheromones aY'e composed of spPcialized intersegmental
2pidermis cells vJith deeply fluted distal cell
menbr3nes, underlying a cuticle with some pores
( S':..einbrecht 1 1964; Jefferson and Rubin, 1973). In
'I'he gypsy moth Lymant ria dispa r for instance the
attnctant is ;:m epoxide (disparlure) which app;,rently
synthesized from its olefinic }!recursor (bierl et al.,
1972; Garde et al., 1973; K~sang (.,tal., 1971~). ln
tbi:.~ ~:pr,ci c-s, as well as in !3om()yx, whrere the 1-1n~curso r
of the ~~lcoholi c pheromone mit;ht be a l'att,y acid,
the ';vei:;ht ratio of prec:n.·::;or; ~Jheromone j s tD the
order of. 1 () : 1 • 'l'ransforYr1E.ticn of the p1.·ecur::~or

44
to the final produc"c mit;ht t'-3ke place in the cell
or even in the cuticle. The pheromone '-lu3nt.ity per
gbnd (Li1e so C!:lllecl female equivalent) is of the
order· of 1 yg ( apiJroximately 101 5 molecules) in
~.orne spec:.i.es (Steinbr~cht, 1964b; Shorey and Gaston,
1q6r;;), b·.1t l'::s:c: in L-,he majority of cases. The-number ;
of pheromone mol~cul es emanating fr.~)m the glan:i sur-
fc-1c e ~as e ::;tim:::t eel in J3om_Q.y_JS_ to be of the order of
10 1 1 molecules/sec, corresponding to a pheromone ,g .·
molecule density of 10 /cm3 in a moderate airstream
paG8ing over·the female gland (Kaissling and Friesner,
1970; Pr:i.8sner, 1973).
The trGnsfer m ... dium is air, \vhich carci.es the
phr.~l~omone to the recc·ptors of th~; male moth anten·ta.
Repo1~t~~ on e:i'l'ective attr;:Jction dist.'·mc es vary from
:;;everal Kilometers to a fe~v llleter (Pri esneri' 1973),
dependtng on the pheromone output rate, the physicgl
trans fer· condition, and eventually on the male moth's
behavior threshold, .. .,.hich in .Bombvx was found to be
rP::JChed at 103 bombykol molecules/cm3 of air
(Kaissling and Friesner, 1970).
On the r('C ei vi ng side, the signal mol ec 11les are
fi.rst adsorbed on the cuticle of the r~ceptor hair
and then migr::rte by surface diffusion through pores

45
::~nd tubules to the membr:>ne of the recentor cell
d.~ncirite, ,Jh,~re they interact with postulated acceptors
(.receptor.s). While the surface· diffusio::-1 and penet-
- tati0n of the cuticle by the pheromone is well documen.:.
.ted (K3s8ns, 1971, 19T3; ~asang and Kaissling, 1972;
.Steinbre.cht and K'lsang, 1972) •; Pheromone molecules
h::1ve for technical reasons not yet been seen in
fl.3 .. ~ranti on th2 dendritic membrane or in the tubules
le3ding to the membrane. The result··of this so •
far postul~3t ed signal-acceptor interaction can be
seen as a depolarization of the dendritic membra~1e '
(the receptor potential of a single cell, or, ih
8n ovP.rall r~::cording, the electtroantennogr.3rn, EAG),
l eadin,; to the elicita-cion of nerve impulses. The
amplitude of the r'?Ceptor potential and consequently I
th2 frequency of the impuls3S are an e;{ponentiu.l
function of the stimulus strc~ngth. In Bomby~ (Kais::;ling
and Priesner, 1970) and presumably in other moths
al·::;o, sir11~l e pheromone molecules elicit ~Jingle impulses
but, the .behAvior threshold of the mGle insect is not
r"'1Chej b"'.'fore seversl hundred cells are ;:;ctivated.
The· nerve :fibers (axons) run ~vithout branching throuGh
106?) where they synaptically cont~~ct seccnd817

46
olf-:1ctory cPlls. As io. the vertebr3tes, the to1~r1l
number or s·-3condary olfactory celLs -in insects is
less than the number of receptor cells(Pareto,
1Q72; Bosckh, 197l,).
The number of olf8ctory r(~cepto.r cells in each
~mt enna of the ~9.m~yx _male is B bout 50,000 of 1·1hich
mor~ th8n 30,000 are specialized for the female
pheromone (Schneider and KRissling, 1957; Schneider
and Steinbrecht, 1965; K,:Jissling. and Friesner, 1970;
3teinbrecht, 1970, 1973); j_n some ~vild silkmoths,
these figures are more '.:.han tvm fold higher (3oeckh
·~t al., 1960). While the specificity of the pheromone
receptor eel" s itt the males of B~b)?.C_ and ~anJ~.J.:L~
is quite advanced, and a very little of the specificity
o1 the oth2r odor r·~C<j~Jtor cells. The=e cel.1s which
do not respond to th2 phe1~omone have b..:;en found in
3n earli:""r study ·i:,o be present in rnther 1:-.Jr::;e
numbers of physiological cell types with dif.r. erent,
but ov,~rluppin6, l~espon oe spectr8 for general odors
(,:)chneider .c.:t al., 1964). HerP. it r1-:main3 to be seen
wheth-~r the~3e cells respond to as yet unkno•·-ln odorc.mts
l"ihi ch are biologically sL;;nifi cant.
Odor molc~cules must for a number of r<::a3ons
inter3ct dir8ctly with the proposed acc,~ptors. The

47
ceL:_'s (or· cell type.'s) specificity is defined by the
v<holc set of comrounds to which the cell responds,
3ni neces~~rily 3lso.by the respective stimulus
r"-'2 1Jonse char::Jcteris.tics (Kais<::ling, 1971). Since
sorEe odor r~=~ceptor C\?-lls seem to have more tlnn one
tyu~:: of '~cc-·~:_.;~or in th::ir mn11br·.ne, th2 r(~spective
sp::~dl'icity of on!'; acceptor is only part of the cell's
specificity (K.<:Jfka et al._, 1973). Interaction probably
i:3 by wc·ak physiC8l, (polar and dispersion) forces
(Kafka, 1970).
The specificity of the o<;ior receptors gener3ll y
i'S rr~ther high bec.suse the effectiveness of the "wrong''
geometrical isomor is in some cases only 1/10()
1/1000 of that of the pheromone, and even chir3lly
different molecules elicit different cell responses
in the receptor cell and can be distinguished behavio
r.L'._y (l\gfka et.al., 1973; Riley et al., 1974). iVletabc
lie processes which chr:~n.::..e the odor molecule in
JntP.nn;:.Jl tis2.ue are probably not ,fast enough to be
either directly rel nt ed to the transduction or to
irru,lecliate inactiv.Jtion of the~ odor moL;c~le (;<.as<'~ng,
1971, 1973; F'erkovich et al., 1973).

48
The even::.u::1l outcome of g)_l these processes in
the pht>romone receptor cells, which send t·hPir messages
Lo th~ br~in vi8 their axons in the nerves of both
8nten''JSe, is '3 motor reaction of the male insect.
The domesticgted and flightless nl'ale silkmoth Bombyx,
when stimul·,ted with the .female sexual attractant,
.ficst lifts its anten iae and then start::, marchj_ng
upwir1d, vvhile vibrating its anten:-"ae and fluttering
it:::; .vi~c';f, (Schvvincl<, 1954., 1955). Wild moths only
re::Jct if they are stimulated at cert3in hours of the
d::-Jy; they then lift their antenuae like~ and
:,oono.r o c l3t er fly upwind in the a erial pheromone
tr:::il or odor plume. If the¥ lose it, they may start
' . .mdirected circling, searching flights, as do other
insect::: (Steiner, 1953). Systematic studies of all
factors involved in the final appros ch of the male to
the female in the field h8ve not bec:n conducted.
Since odor alons does not t;ive the anim:::~l a directional
clue, it presumably uses its optomotor system to
~teer upwind (Kennedy ::md I·JlJr·sh, 1974). Bui.", wingless
moths ar·-' able to follow a terrestri-al ph~~romone
trial like ants and termites ( Shor!~Y :md Farka3, 1973). 1t.
Theoretic3l consid•3ra~vions of the composition of
f3Uch an odor· plume l e8d to unexpected prf::dictions,

4-9
.1t 1P8st in an ideali?;Pd case (Wright. 195t'i; Boss~rt
1nd l.Jil.:o n, 1963; ~\Tilson and Bossert, 1963). The
gr·;di ent of uecrr-;asing odor concentration is very
steep near the odor source when there is a reasonable·
air flovv speed. Next to the source the air may contain
up to 1 Od ph:c< romone molecules/ cm3, while at a few to
30 meters distance (as a rr-:sult of air turbulence) the
cone ent, r"''ti.') n rangr-; is 1 o3 -104 molecules/ cm3. The
odor plume then extends with little reduction of
concentr3tio11. (dependj_ng upon the rate of air flow
:::riJ. d.i..LJ.tic)n, sower et al., 1973) to more than 1QO
meters .. Effective luring 8t longer distances is
only possible if '~i ther the source is more effective
than is knov,rn at present, or the pheromone catching
rr-1t e of the m.~Jl (?. ant en<~a i :' better than in Bombyx.
The s'2Ylsitivity of the attractant receptor cell
reaches its theor(~tic3l physical limitis· and the
:.112u l~al noi s espontaneous firing of the odor receptor
cel.Ls - (Ka~ss.ling and Priesner, 1970), 0.1 impulses/
nee., is r3ther low for such a sensitive system.
However, theoretically, a reduction of this spontaneous
activity by a factor of ten vwuld allow th2 r(~c.::p-:.:.or
system to r2spond to a greater dilution range o.f
the pheromone ::Jn:J al]_od the nBle to api;roach from

50
In th(! lic:;ht of this theoretical background, it
i interesting to nots> th~?t Trich<?_P.lus~~ ni females
rel3te their pheromone-rdr;ssing behavior to the
wind velocity (Kaae and Shorey, 1972).. A"'v lovv wind
speed, ·111hen the odor i~3 persistent and dilution is
low, ~:.he f enwl es call for longer p(~ riod s. In adui tion
most fem8les vibrate their wings (fan"'ning) 'v'!hen
c::LI ing 8t low wind spe::ds and do not at high wind
spe ''OS.
ri' his com pl et es the story with respect to c h emi c gl
info rmstio n transfer from. the female to the male which
eventu~lly mates with her. The message is biologically
hi;;;hly meaningful; the male moth underst':lnds the
11 1 a ngu 3 g e 11 o f hi :3 rna t e •
While thi.s flmc'donal cycle seems in some moths
tc be completed with the male's approach and inst:YQ ___ 't
m:tin:; :::~fter he r:3.sches the fem3le, some groups of
moths do have male pheromones, ·.tJhich are apparently
desi,gned for short distance effects. In fact, male
scent scales anJ scent brushes or hair pencils
(androconia and coremata) have been described in
m3ny lepidoptera. Many male Noctuids, Sphingids and
Arctiids h:~ve lar:~e hai r_pencils which they can
Pxp:~ nd for the dis serninatio n of an odor.

51
2 ·5· Beh<wio cal Rr;sponse
The functions of the olfactory organs can be
:,~L ;~ '·~d j nr.lir~~ctJ.y through the observ::.1tions of
b<2ha vio ral responses of many moths species.
In sim~les~ case, an application of a
:ccAnt-sLirnulur-5 rr~lA3Ses a rAflex-li.ke response.
For example: l'~lale moth responds to the sexual . '
3ttract8nts or sex pheromones by showing wing vib-
rations and ant ennal elvations and abdominal move-
r.i2nts. Thc-y cannot fly even and start walking to
the source. Such type of response i;.:. called walking
up wind ('tve anemot8xis) . . This response has been
reported in Bo~x ~g_ri ( Schwinck, 19 54, 1955), in
Por~h_Q..t_ri~ dispa.x (Ivlarsh et al.,· 1978) and in
Gra~h_olit~ molesta (Kuenen and Baker, 1982).
When strong stimulation comes from the source,
m:1le moths sho,, a spiral or circling pattern of
the body with movements of the abdomen. This
pattern has been found in ~ombJ!?C mori (Sch,Jinck,
1 q 54-, 19 55) , in T ri c_£!o...£_~ ~~i:3 ni (Hen~ eb e rry and
HovJl:md, 1966) in Diory~triaab:!:_~!-_~ll9_ (Fatzinger
and AshPr, 1970, 1971 ) , 0 stinia nubilalis ( Klun and
no bi n~"i:' n, 1970) and in ~h e~r::h.~ di~J2J!£ (Marsh
!ct al.' 1973).

52
The IIIJings flutt ~ring or vibrations ccm be
m,:;asure 1 mechanically and can be used for determi
W:lt:Lorl of the m8l es olfactorjr threshold when
stintulatc-;d \IIJith sex pheromones or other chemical
compounds (Schneider et al., 1967; Kaissli ng and
p;·iesnerl1970; priesner, 1979b; and RoelCifs, 1979).
La.boratory experiments usuC:Jlly allovJ a beti~er
yualifi cation of the odor stimulus. Male moths
arc often made to select between the odor free
and odor containing arms of "Y" ma~e or "Y" tube
olfactometer. hey and mills ( 1968); J.Vu1ller ( 1968);
Toh"-J et 3l, ( 1963); Brady ( 1969) and Rejesus ani
R'qnolds ( 1070) succeded to measure the attracti-
.._,. en e? s and repellent response of male moths and
properties of chemical c·:)mpounds in thr:ir olfactometer
studies.
The olfactory organs play two major role during
male moth's orientati.:-m to an odor source. First
is to detect the presence of odor and 2nd is the
quality and intensity of an odor.
Most works h:::1ve been d·esi ._ned to me~.l sure or
yo evaluate the levc:ls of responsivene:·s oi' males.
(Trayni•~r, 1968; Bartell and shorey, 196g; Brady
and Finery, 1969; Fatzinger, 1970; Klun and

53
Robinson,.1970; T~kahashi and Kitamura, 1970; Brady
:1ru TunilinfOn 1971; Fatzinger and Asher~ 1971;
Dater.11an, 1972). Following are the behavioral steps
depend upon the use of labortary techni4ues that
l~>·~~ay r1 p::lJ~tic~~lar type of respont>e.
1 • ~.r:z J?..b.~-r:~~~ix!.c}:.':!:.~~-ci_§_~y uen\~e of behavior
!\C:.:onll.ng to Tr<:lynier ( 196S) observ3tions with
the males of Anagasta kuhniella were released in a
wind tunnel, downwind of the calling females
fehv upvvind. If they lost the sex pheromone scent,
m:jde the cross wind and located the female by
p? rc piving the scent.
Takaha ;:;hi and Kutamure ( 1970) r:::.port ed that
the sex phenomone rele3sed by the female Bomb~
~<2_ri, indue ed the male for matine; purpose's.
r~l8le shovv~"d the irrmedir-Jte response tovvards
virgin fem3le and nttr~ct::;d to the source of
pheromone. Olfactometer studies of Rejesus and
f\.Pynolds (1970) showed that the male of Bu~_C'_!-~,'"Jt~ix
~~~_r.::_l~~!::.~fJ..:.:.~~ responded to the C'"lling ferrrJ. e
shovving the typical performance of wint; vibration,
ant ennol movement, but 100% att ra ctio n s vJ er:; not
tound though the virgiu females were in c0lJing state.

fAmales vverr:~ reported by Fatiznger ( 1970) and.
FstiznLel' and Asher ( 1971 .) in the olfactometer
study by pa ~sing air through a chamber of virgin
females into a chamber containing males. Enit'cing
,'herornone:~ ~vere rPspon:::;j_ble to behavior8l activities
of t,he m:.Jles. Attr3ction studies experiments were
concmcteci by Bsrtell and shorey ( 1969) on male and
' female of :92_i_p_hy_as Postvitan~ using straight ,glass
'cube· Olfactometer Li the laboratory. ,Male showed
irrrnediate respon<c2 to the scent emitting from female
and stR:rt ed orientating to the source of phe.romon e.
GOtz ( 1930) studied the choice experi ent 011 Labesia
bo rt1n8 males. In the tests, males wer·e released --·-----freely and Given a choice between 1 and 3 virgin
f·9males. Mo,~t attraction of males w::1s found tow:Jrds
the scr~nt source, 8!lln::.ting fr)m•3 females. Qw1ntit8tively
the pheromone3 from 3 females were more attractive
to male~:; than one. Klun and Robinson (1970) succed.ed
to see the attr:=Jcti 111 respon:.:;e of male moth of
started from the virgin female, the males got excited
and started orienting to the source of pheromone.
Rem<JrkGble studies were carried out by schwnick ( 1953),

55
Brady and Smith ( 1968), Brady and Finery (1969) and
Br~.dy and Tumlinson; '( 1971) on pheromone indue ed
behaviour in male Pl<2_~i:'.! inte q~u:r:!.:t ~l_la. lVlale was
'·lui ,,e e:xcit ed by the scent of virgin .t'em8l e. 70 perc:-;nt
an.:i above attr·;ction:,; wer·r,; observed.
The fir~.t rPcogni7able step in the seyuence c:f
re·wons e is the activation of a resting male such
activity is represented by elevation of the antennae,
grooming of the antennae and wing vibration. \'Then
a resting male perceives pheromone at a level above
its bc;h':i·Vioral thrt~shold it is stimulated to '<valk
or fly. This study is being supported by Baker
et al.~(1976),&Flint et al., (1977).
I). Orientative beh;::;vior - --- ---------------·-Orient8tive behSJvior rPsult:-:; iD the male
locating the pheromone ~-;outce: the conspectific
femole. Uf cour·se, the analy:-::is of insect orien-
tative beh8vior is not rt~stricted to ori.e~·1tation
to pheromones, nor even to other chemical cu-es. A
V3St library of cherni c<~l kno\~l edge is now available.
It should serve as a verstila tool for the st,udy
'"~nd the manipulation of orientative behrwiot~. Two

56
}lrincip3l m•"'Ch-1ni:;ms have:; be m aug .•·sted whereby
f lying insects could orientate to pheromone source
cheiHotaxis (chemical st ree ring) ane;Hbtoxis (·wind
~-~ t ~ r: ri n g ) .
Farkas and shorr-;y ( 1972) h?.ve r2 por·Gecl ti1at
ms l es o f th2 [email protected]_t)_r.!2.P.l:!~.rr:!. g_os·~2i.§.~_l~ i s stillair
rr-~m3ined ca pable of orienting along ~an o dor plume
t:.h?t the ins ects wer e ca pable of gAini ng chemo
tact ic .orientatio:1 cues from th e st?tionary plume
along which thtO} moths . progressed in the former
upvvind di r ection . Kennedy and March ( 1974), hovJever,
chgll.enged this vievv in their experiments _with Pllodia
int ec.2_~l'!_c_·~_ella th-'3t the insects maintain ed up~r;1nJ
orienta tidn by mPanG of anemota Yis, r egul8ted by
optomotor respons es to the visugl surr'Ounding and
m0dul.1t "' d by the phe r' ·fllOn e flowing do wnvdnd. They
concluded' th!'lt althouJ;h the me ch:::nism ofcxior r t:gu
lsted 01j tornotor anernotaxis rerroins lart;ely unanaly -
. ~ed it is still · the mo s t r.>lausibl~ gu.idance rn:c.hr..nism
for thernoths studied so far. Farkas et :::~1 · (197~. )
lo0k0d more closely at cnvironm .:~ntal factors ~Ji'J'P.ctin~
the aeri::1l odor trial follo;,ving of mal-:=s of E_2ct_i_nophora
gg_~~YJ)iolla. 'fhey found that the behavior or" the

57
males was regulated by the concentration (at source)
of the pheromone, t·he spatial disposition and intensity
of an illumina Ling light source and visual cues
from nearby objPcts. Increa-·ing pheromone
concentrati'Jn tended to reduce the forward air
spr~ -:::d of the rr~spond.in,; m~:lle, both 8lon[; th"'? zig,.,ag
flight path and the axis of the trail. Th-:: reduction
in speed occurs in part, as a result of r-~duction
in wingbeat frequensy and represents a nagative
ortho kin r::si s.
(Oef: speed or feyuency of locomotion is
dependent on the intensity of Stimulat:..on visual
cur.;s and the concentrations of the pheromone appe~~r
to 1,vork jointly in stimulating the slowin;_; of flight
and. landing) •
4. !IE_~~~l'!d __ a_n_e:!!!_o_~~~-xis: The ,dnd flow is a suL .. icient
magnitude for a moth to detect i-cs direction, for
locating a chemical source in such wind. such
orienteition i.s called upwind an·111otaxis. This process
h"-Js been studied in vdnd tunnels in several.moth
species ( Trn yni er, 1C! 68; F'a rka s ;-md sho r',Y, 1072;
K8rk::Js et al., 1974; Ke.Jt1edy and l1ilarsh_,1074; Kennedy
et :::Jl·)19f~0; 1nt31, Marsh et Al., 1978 Ca1.~d~ and

.58
Hug"lm:;n, 1 q7q; Sanders et r1l . , 1981 ; Baker and
Ku~nen, 198~-2; Kuenen and Baker, 1982).
In mo::;t of these ex~mples the flight trsck
::3lon,;; the ;::J:Xis of the pheromone plume has been
charceterized as 8 zig:;>;ag, along an upwind tra-
j~ctory. Th"-" expl3nations of kennedy and Marsh
( 107h) for the zigzag behaviour was that. the
' successive tu.ms were initiated after the loss of
the scent, the turn carried the moth back towards
the direction where the scent was lost sensed. (the
direction to\·Jards the source, however, would be
detected by the optomotor response). Complimentary
evidence that such turns are initiated by the loss
of the plume come from Traynier' s ( 19 68) wirrl '
tun:1el observations with the moth Ana gust§_ KUh't'li ella
in homogeneous pheromone clouds; moths were reputed
to fly upwind in a straight path. Tests designed
to compare the flight track of males in narrow
discrete plumes V.S. homogeneous clouds of phero
mones in Adox~2he£ orana (Kennedy et al., 1980,
1981)' unlike Tray'nier' s ( 1968) observations W:.'ls
shovm that zigzag path occured under both conditions
in contradiction to the mechanism proposed·by \
I

59
~ennedy and M<Jrsh ( 1974). Lateral revers~1s, tlH.m,
seem to occur in the presence of windbor:Q.e pheromone
and thus represent on internally generated pattern
0 f fl~ght.
The wide lateral excursion characteristic of the
zigzag subject an air borne moth to increase aggular
deflections of path rdative to the upwind direction
over the deflectiun of track that would be. incun~ed
when the heading is aimed continuaLLy upwind.
A zigzag flight track offers an alternating
series of comparisons of deflection and elevated
th~"' navigational inform ?t tion available to the
insect. The upwind could be set by adju:::ting the
heading betiNeen reversals to msi ntain a flo vJ of the
ground pattern that ::Jltern:=:ttes on the right and
left l~?gs of the zigza.s at the s:1me drift angle
between the body aixs (course hegdin!;) anci track.
In the gypsy moth, the position of the moth .1s
it fliPs along the zi~zag is such that tbe moth's
body axif, is ::Jli,~nr~d with the coursr~ he::)J.ing
(body axis). The ad·vantat;e of this str:1t·~t-~ern, .<:Js
coutrasted with mr-Jintaining <.m upwind body axis
heading along the zigzag course, be rela"Ged to an
improvement in flight efficiency.
/

6o
The visu;l effects generAted in .both situatipns
are similar im>:::rms of the amount of angular deflec
tion, althou:):1 th_e path of the visual field across
the eyes >llould be quite dil'l'(::rent. If the insectbody
is aligned vvith the course direction, although
the psth of the visual field across the eyes 1vould be
quite different. If·th' Irlsect,body i:. ali.....;n---d ,.,:i.th
the COUrse di rP.Ction, the fl01tJ Of the Visual field
beneath the eyes, alternates at an angle relative
to the head-body axis that is ~qual to the drift·
angle (Marsh et al., 1978).
Another explanation for the zigzag 'path J the
moth WAs allowed to fly up in plume by sequential
sampling of the pheromone plume's spatial structure,
~vben the· vvind drop1Jed below the anemotactic threshold.
Another consideration W8S thRt· a zig7,ag pClth
(noting both the lateral and thP. vertical cotnponents)
increased t be pro ba bili ty rc co uta cti ng chemic a l
stimulus after loss of the scent (Kennedy, 19?8;
David et al., 1982).
Tbe effecls of increase~ in pheromone concc~nt.
rntion upon the zig zag flight of Grc:pholi ta
ffiolesta were to increase.

61'
1) 'rhe frequ~ncy in turns
2) Decrease the airspeed (and therefore the
distance across the plwp.e's axis between
turns). The an~les between turns, however,
rewain relatively constant ( Kuenen and
B:::tkP.r, 1982). rrhP resultant effect is to
provide a. narrower zigzag path and slow the
speed of progress up the plume as the moth
approaches the pheromone source.
The; pr::>cision and success of Grapholi ta mol esta
o ri ent&tion to pheromone was quite dependent on
a narrow range of optimal. pheromone co nc ent ration
{C8rde et al., 10?5; Baker and Garde 1979; Baker
3nd Roelofs, 1981) and this fact of their behavior
differed from most other moths Lymantria dispar:_ which
oriented to a ·rather bro::1d range of stimulus
concentr8tions.
As pheromcne concent ratir_,n increased the
di stance t r.'3vell ed ':m the directly upvvi nd cornpon ent
o1' the flit_:;ht track on each zigzag decr2ased as did
the ground spe t:~d. In contrast of Gr~1 El~o_J}_t_a_ u;_o_l_~_~t-~
j_n the :!:oY 1Jsy n1oth, t..he ir1terle~ angles narro~~ sligh~.-ly
at increased phecomon~~ levels· and the distance across
the plume bet"1een the turns rf'.rn-.ined relatively constant.

When the plumes removed sud,lenly, the gypsy moths
were allowed to cast into a wide homegAneous cloud of
pheromone, the zigza.;::;ing upwind flight resumed, but
with some alternations: the width of the zigza.::;; p3th
v~ 8 s ap:Jreciably increased as the velocity along each
leg, compared to a na rr'Ow pheromone plum e. These trends
in fli;ht behsvior in plume Vs. Cloud remained simi.lar
over rou -:.:~hly 100-i'old pheromonf~ ranges and so seem
at-::.ribJ.t3ble to the spatial structu ,··~ of the chemical
sticomlus rather than pheromone flux.
These observations compliment those with
Adoxop}~es oran~- ( Ken_:v=:dy et al., 1981) in that zigzagging
was an internal J.y ,gener:~ted beh8viour (pre-programned),
but in ad_~_ ition, they shovved that the zig7.8g path
of Lyma~~r:.~'! di spar was influenced by the external.
information about the spatial structure of the
pheromone stimulus.
Tr::qnier ( 1968) reported that the pyralid moth
Anag8~t8_j<·uh~:b_el~'! in a cloud of pheromone, flied
directly up~vind in straight (or at least not zigzag;)
path over A m,_,ter distance.

2.6. S8x Pheromone s Production and Release A.g.~=q):_the_[_~ma-le motiiaruf its Effect
on the I.Vlale moth ---·--·------·-
l'~lany insect groups produce sex pheromones and
emit than during their entire life span begining at
the time of emergence. I~1any others do riot attain
sexu.ql maturity until they reach at a certain age
or sometimes natural death occurs due to a cease
of pheromone production. Girnila rly t~e· responding
sex may or may not be sexually mature of emergence.
r'emal t~ nun moths Porthetria monacha were most . -------
attr.:~ctive vJhen 1-2 days old ( Jascentkovaski 1932).
Hanno ( 1939) and tun bros ( 19 40) claimed that newly
emerged f emales are only weakly attractive, the
att.raction becomes stron~er in female 1-2 days old;
8t0raction almost disappears af~er mating and com-
pl Pt ely gone by oviposition. The female continues
to attrsct males for about 12 days ( Hanno 1939).
Richards and Tl1ornson ( 1932) sho~~ed that females
or' Pl0_'l.ia interp1!_n.~~ella_ are ready for f!lating ~
very shor c ti!lle after em c~ rt!;enc e. L ehmensi ck and
Liebers ( 1938) showed that females are not -attractive
until they are at least 3 hour.s' old, and male do
not respond until they are at least 50 minutes old.

64
One day old males placed in a dish in which a 1 day
old female was kept, imrni ediately beg:-1n a court ship
dane e and attempted to copulate. ..U though 5 days
old feltlaJes wen! ~ot attracttve, a we·,k male re..-;-
ponse was evoked by 1 5 days old females. According
to Brady and Smithwick (196:1) newly emerged femr?les
contained a concentration of attraction, equal to
5 days old adults. Virgin female released attractant
almost continuosuly during their adult life.
Vohringer ( 1934) reported that Cfall~ria m2llon.::;lla
begins to -relea~3e attractant 12 hours old after
emergence, copul:1tion was observed ~etween 1i·-2
hou1~s old males anl 12 hours old females. Roller
et al.' ( 19 63) showed th8b the males develop a
ch·1 r<1ct erstic odor when the<J are 2-24 hours old;
this odor researches a maximum during the evenlng
hours.
Labor:::tory experirner1ts showed thul:J a 2 d:1y
old female par.slobesi:3 viteana imnediat.ely exceited, -- ----·-- ----when ::3 rne1l e w~s brou[:';ht to her ( GOtz 193q). One
d'1Y old females of Lobesi8 botrgna ami Paralob,o;!"'ia ---·-- ---- . .........,...---- ---!_i_t_~~E.~ are mdlre atl,ractive to males th:::m are
2 day old females although virgin females rPifl3in

attractive for their normal life span (9-13} days
o 1 d ( GOt z , 1 939 ) •
65
M8kino et gl (1956) and Jones&. Jacabson (1964) reported
that male and female g,omb~ mori do not fly but. eager to mate
immedi3tely after emergence. Female ·remains attractive in
the last days of its pupal stage. Perez & Long ( 1964) reported
that females of Diatraea ~~s_haralis begin to emit their
attr:3 ctant soon after emergence and are most attractive
during the 1st 3 days o~ life then attractiveness decrease·
with agee Most Zeadiatrea grandiosell.a. females began to
attract males on the night of emergence. Attractancy drop-
ped sharply after the4th day and was completely absent in
8 d8y old females (Davis & Henderson, 1967). George &
Ho'nerd (1968) found out that virgin female of Grapholitha . molesta increased in attraactiveness with the age atleast
until their 5th day of 'life and Grapholitha funebrana
female shov~ed the great est degree when 1-3 days old
(Saringer et al., 1968). The sensitivity of male almond
moth Cadra £3_UtelJ.a to the female sex pheromone increases
gradu:=illy with age upto the 7th dRy after emergence.
Maximum s ensi ti vtty occurs at 3 days· of age. The phero
mone is already detectable in female which have begun
eclosion: it increases in amount upto a maximum of 3 days
after emergence and maintains this 1 evel until death if it
does not mate (Kuwaharct et al., 1968).

2:1. Structur~~--'?__~t_iv_~t_Y. __ r!~_l_<?~G-~Q_~sh~J2..s in SE?~ --- -- -?tt::_r:_c:.~_c_~_::J_'Q_t _ _R_h_erorr~~nes
66
i3ec su.s e of the relat i v.e sirnplici ty of thE! chemical
::;~ructt.lT·~; of fP-male sex-att~r~-:1ctant pheromones; a
consider8b1e number of pheromone analogues have br:~en
synthesiz"'d and their biologic activity evalua~ed
1vith var·L.JUS species. Generally any slight changes·
in the chemical structure of pheromones ro2sult in
.1 pronounc~· .. l r•::>du.c:t"i.on in activity. The ap,•rent
biologic activity of pheromone analogues differs
'1ccocJ.in~:; to which assay method is used. The follovJing
.tn•''tho; s :·.in:;ly or in combin~tion de·Germin~ biolo;:;ic
ac'.::.lvity: Field tests for msle attrsction.
Laboratory .tests for sexual ·excitement or
short-range orientation behaviour and EAG responses.
Out of thes"~ 3 methods, the highest specificity in
structure activity relationships is usually obs2rved
by field tests and lovvest by EAG response as sho\vn
in Tri~h~lu~~~ ni. (Gaston et al., 1971) and the
a no. the gypsy moth (Adler et al·, 1972) •
The low specj_ficity of the EAG m<S.thoq for
detennining struct-.J.re-activity relationships is shown
with doubly unsat.ur.:.::ted sex pheromones and their
::malogi.les. For Ex: In the case o~ the sex

phPromone of the almond moth Cadra cauJella Cis-9 1
t.r8ns 12-Getr<-)decadienyl acetate, there were strong
E\G responses with the monounsaturated tetradecenyl
.'3cP.tAtes having the cis-9 or the trans-12 double
bond (Roelofs and Comeau, 1971). The lovv specificity
in the EAG mothod has been successfully utilized for
the structur:"3l elucidation of the sex pheromone of
the codJ_ing moth, trans-8 trans-10 dodecadien-1-0J I
( Ho el o f s et a 1 . , 19 71 ) •
Laboratory assays on a considerable number of
pheromone analogues revealed that some of these
synthetic compounds elicit s exuF.Jl behaviour from
m3le moths such ~s the cabbage looper moth
(,Tacobson et al., 1968; Toba et al., 1968 and
Gaston et al., 1971) , the southern army wo nn moth
(Toba et al., 1968), the codling moth (Butt et al.,
1968); the gypsy moth (Sarmiento et al., 1972) and
thc-' almond moth ( Takaha s:hi et al . , 1971) • lVlo st of
the compounds found to be active in laboratory
bioassays, hovJever, were in active in the field for
at t, ra ctio n of males.
Fi-"'ld tes~s v.Jith various pheromone ,analogues
show that a minor structural change of a pheromone
resi;lts in a · considr.>rable d;>crerlse in attractive
pot en.(; y • /

6g
The first electophysiological recordings from
olfactory receptor cells in the male of Bombyx mori
were performed by schn.eider .( 19 57) who obtained a
slovJ pot entinl!:-'J, g;rsd.pd. potentir:1ls anc1 !"'.Urnu:atc~u
receptor potentials or electroantenogram (EAG).
In 1962 , Bo eckh reported the 1st quantitative
recordin;;s from single olL'lctory CE.:lls (Sensilla
basiconica) in the corrion beetle Necrophorus
vespilloides .Tungsten micro-electrode with a tip
diameter of about 1-10 JU (different electrode) v~AS
inserted in the cuticle in the vacinity of one of
the olfactory sensilJa. .A 2nd broad~.r,. electrode
(Indifferen:t electrode) w~s inserted into the antenrtal
h-;emolymph. 1/Jith excitatory stimuli, the different
electrode usually b"'came-vely charged by the receptor
potenti:Jls. The nerve impulses, hov.Jever, usually
Rt<'earerl. 8!:'. thP. deflections. This tP.chniciue turned
out Lo be well-suited for recordings from sent~illa
of short hairs of coeloconic and placoc..aic type in
several species. Mor'" suitable technique for record-
ins from single cells ( Sensilla t ri chodea) of long
hair was cut off and. a glass capillary micro-electrode
was brought into contact with the sensory cell by

covering the cut end of the hair with the electrode
tip. In that manner, it was possible to record impul-
ses and the slovv receptor potential of the in!lervating
cells. Recording from a single, sensillum, impulses
from one or more cells may be obtained. In the later
cases, the spikes of varioUs cells are distrint;:;uished
by difference in amplitudes. The stim_ulation techniq_ues
chosen for studies of insect olfaction have to be
considered \~ith respect to the small quantities of
phsromone compounds that are sometimes available.
One technique employed i:!'"l stu?ies of lepidopteran
pheromone, is the use of cartridges (Kaissling and
Pr.iesner, 1970). A specific amount of a compound: is
applied on filter paper inside a glass tube (cartridge)
through which a constant air stream (Puff) is
blown over the antenna. A stream of pure air is
blovm over the antenna between each stimulaticn:n
pP.riod. In other techniques, the odor is introduced
as a puff into an air stream, contineously blo,>ing
over the antenna. (Roelofs and Comeau, 1971).
2 .~.1: Responses of Olfacto :ry Receptor cell to general odors
The receptor cells are either activated, inhibited
or 'unaffected Vl'hen an odor :is blown over the antennae.
I

70
When activated a negative slow deflection is recorded
between the receptor lymph of a sincle s ensillum and
the 11aemolywph (Kaissling and Thorson, 1980). '1'his
. defl~:~ction· is assumed to be the summated receptor
potenti<Jl of the innervated olfactory cells of a
particular sensillum. The sum of deflections from
all. olfactory s ensilla is as swned to constitute the
EAG recol~ded with relatively large diameter glass
C"'Jpillary electrod<>s. When recording from a single
sensili.um with glass capillary micro-electrodes,
dy.ring stimulation \'lith a. low pheromone concentr.3tion,
small elementary potentials (0-5 mV) are observed
10-40 ms befor·e a spike occurs. These potentinls
have been interpreted as .the result of opening of
single ion channels involved in the transduction
process. The spikes of extracellular r2cording
co nf',i st of an initial po si ti ve deflect ion ( 2ms)
follovpd by n'"gative one (Kaissling. and thorson, 1960 ) .
ln some ca~es, responses to odors may also be
recorded 8s an inhibition by receptor celLs, when
they shOV'J a slo·,v hyperj..>olarization and reducr:?d the
firing rate ( Bo eckh, 1967a).

71
Th9 specificity of an olfactory cell reflects the
specificity of membrane. receptors. also called acceptors:
which are located in the dendritic membranes r The
possibility that the specificity might be confined
to the conduction system, e.g.; microtubules w3s
excluded by the observation th:Jt two sensorJ ceLLs
innervating the s~me sensilLum, and sharing the same
conduction system, could be activated by different
'key' subtances.
In activating cell, the odor moleculB is thought
to bind to a membrane receptor protein (Hansen, 1978)
wherc.>by a conformational ch:1nge to an active sta.ge of
the receptor occurs, leading to the opening of ion
channels and depol a ri zatio n of the cell membrane
( Kaisding and Thorson, 19 SO). So far, different
olfactory cells have been shovm to process varying
degrer~s of specialization, some respondirg only to
one of the test compounds while others may respond
to IilCJ!lY difl'~rc-!nt odors. Varying coruposltton of
·ctiffArent types of membrune rr•CPIJtors within eoch I.
cell. h;·1s bf'en postulated as a moclel to explr-3in the
d9gr~e of specialization possessed by different
olf'3ctorj cells (Kafka, 1974).

72
In an early study in the saturniid moth,
\ntherea pern;y:h, 'specialists' cells are those
c elj. s which rEspond to phe ramon e compounds while
'generalist cells are those cells which respond
to 1na,1y dif;'erent host compo...cndc (Schneider et al.,
1961~). 'I'h~ eel LS activated by phr-;romones are
specir.1lized for one compound of a pheromone mi~~ture
(Kais~;ling, 1979; priesner, 1979a; Den Otter, 197'1;
T>'iust8pa l~t- , 1979; Angst, 1981 ; Wadhams et al.,
1982). On the other hand, odors with other
functions such as food attractants usually seem to
activate cells which h8Ve broader response spect,ra.
HovJever, the turm "generalists" can be misleadirJ6 . sine e it may inean either cells which all resporrl
difi'er·.~ntly (individual rr-:sponse spectr;=l) or cells
which br::.lone; to a certain response group: where
th('? r(~:>ponse spectra within a group is identical
( v :~ res chi 1 C? 7 ·1 ) • Further mo re , t h es e g ro ups rna y
ei.ther over lap or be non-overlapping.

73
The female sex pheromones specifically activate
sensilla trichodea re·ceptor cells on the m;::;le r-mten-
n::Je and 2licit 8ttraction andsexual beh8vior in
th9. m."l es (Kromc-:T, 1973). 'rhe ph .. ~romone compotwnt,s
which ar identified by chemical an8lysis nre
str'1it;ht chgin unsatur3ted hycirocarbons vdth acetates,
alcohols ~~nd aldehyde ;:.Js on termine1l group; often in
co mv,ctio n with electro ~)hy siolo gi cal tests and field
observations. Synthetic pheromones' availability
m;;;kes this sti1L~lus rece;_.Jtor system most sditable for ' li
studying the structure-Gct.ivity relat-ionship of the
olf::Jctory system (Kaissling, 1971 1 1979; Priesn'?.r,
1970a, 197C!b; Schneider et al., 197'7; Den Otter,1977)
nncl for test:.in;:; various hypothesis on molecul''lr
receptor (acceptor) interaction (Kafk3 &:- i~eu.virt.h,
1975) gnd transduction processes (Kai3sling, 197h).
Th" m.unbc'r of sensilla trichodea SNlsOr:l C"'l2.::;
v:-:1ries from 1 to 5 and clussify A,B,C,D,E, dep~nding
on the moth species {Priesner, 1979a). In some
spociQs, the cell generated the larg0st spike amplitude
in each sensillum consistently belong to th:> sr!me
..

74'
cell tylJe; i.e. maxim3li.y responds to the same "Key
subst<mce", which i~. the major pheromone component.
This cell type "A" is generally present in all
san<;,ory tricbodea vvhile the others gre Keyed to a
minor componont s and frec1uently found. In Bombyx mori_
::ensilla tri chod.e:~ contain two cells,· one larger
tlnn th:::> oth.-.,r. From the lCJrger cell, l.1rgest
Si.Ji k -·s el c ct ted by bombykol ( Kai ssling and Tho-rson,
1 q :.30) • Similn r rels ti ons hip bet ween spike amplitudes
ana sped fi cities of sensilla t ri chod ea sensory
cells seem to exist in m3ny moths (Priesner, 1979b;
K·,is~,;]_ins;, 197g; Den Otter, 1977; ·O'connell, 1975).
Thus, even ~hou~h most species use multicomponent
pheromones, the r:::ceptor cells det.ecting the~~e
sie;nals ar~" 11 sp,~ciGlists"' for one "Key" .subst'1nce.
Other yuest,ions ot interest include th.e
sensitivity and the SlJecificity of the cells for
th8i r ro:: specti ve "Key" compounds. The pi one· ring
•":1. p c'd.tn •"nt l e.'tdi nt; to the conclusion th:'J t one of
t.-vo moleculr.:-s ol' bomby.kol hittin;::; the rece1Jto~·
cell is e:,olu;h for eliciting a spike, WJ, p~r-
fo nned usin,s el ectrophysiologi cal reco n:iin6s
combin-=·d .. vith m~~surc:nwnt::: of adsorption oi'
L1b"7c.led borubykol on the antennae of Bo~by.x n~ori

75
( Kai ssli n; and Priesner, 1970). In other species,
the rd a:_.i v e s ensit:i vit_r of the cells to t h2i r
·r·-"sp.:>ctive "Keyn compound has .been determined by
measuring the amount of t h~~ unlebc=iU ed compoun.J needed
to (~ive ;1 cert:1in respon~·e at lovJ levels. • In that
m~.; uer it v~··s found in Bombyx_ mori, /\nth~~~ polyphyemus
(l\8isslin.;, 1979) and various noctuids and--tortricid
mot.hs (Pr:;.esner, 1979a, Den Or.t.er, 1977) that
approximately the s8me degree of response of the
respective cells was el:Lcited by e4uivalent amounts
ol' thei.c 11 Key" substances.
In severt'll moths, priesner ( 1979a, 1979b) deter-
t:,lned the specificiti::::s of the various pheromone
cells -~ithin sensi u_ a t.ri chodea by measuring their
t.h~ ''Key'' compoun-::.t. The function of each cell. type
:1ppears to be ''A" is only for main pheromont;: component
"B .. :!GD" are fur interspecific inhibitors. ".E" is
for synergi~t only; and 11 C11 is unknown {Hill ~nd
Ho <.:>lofs, 19[)0; M eDonough et al . , 1980; Fa rine et al.,
1081). All ce.Li. s h8vin.; ap;Jl"'Xim::Jtely the s8rne
threshold for their "Key 11 su b.st8. nc es, also r~sponded
to other compounds but o:1ly ~:lt highr,r corne::1t.r9tions.

76
to 8}.dshyci0s And 8lcohol~~ ::m;i the old,c;lydrc: cell (D) I
len~th from th·1t of the "Key" subst8nce, gen~r1ll.y
csused a decr--a~~e of the stimulatory efl'(:!ct. HovJever,
it cioes not s'?~::J.n to be the total ch:1in le,-:gth, but
rgther the len:;th of particular section of th2 ch::Jin
is most important.
The receptors in the Noctuid and Tortricid moths
ar,_, fo ·n<i dif. er: 'The part of chain bAt\P·:n th<:· polar
alq.1l ,.rou_(; and the double bond. being most criticc:ll ~
for the svirol:J.L~t.ory efL'ect on recr::pt,or celLs in noctuid
moL;!~_, and the asher part of chain is wo~-~t criticJl I
in torticid mcchs. (Priesnsr·, 1979a, 1979b). ll~;ain
thP recc•ptor .rp:_::cificity Of 8 p:o.~rticular CPll t:;pe
gr~ner·1lly r:"mains consistent \.vithin clo~ely r.:.l3ted
sn0Ci•"'S· 'l'hu~~ Pri esner ( 1Cl79a, 1?79b) found
. J , S[)!?C1.•"':3 3S ~~·e .. .L 8S in closely r2la'c ed speci •2s. ·rhis
in contr:.st to the receptor cells of taxonomically
for which th2 specificities diff~red :,s de~3C;'ibed above

77
Th~~s·"' findin,~s of const::Jnt specii'icitL='s of phRromone
C'-::LI __ s belon··:in,; to one 'cype, diLi'er fmm thr~ results of .
sen::.:il .a tl~ichod<?a rec<=;ptor ce.:_! s viere_, each found
to be mo,;l:. sr:::n:_~iti.ve to one o::.' the ph2romone compo-
nen"c.s, (Z) 11-13: r;C 01~ (E) 11-1,+: 1\C. As but indi-
vidu.··1lly the ceLLs differed both in respect to ·absolute
~-HLl r·:'!lJ-L-i.ve sensitivie~::; for the:; pheromones and other
rel3'c :::d c-:>rr>~~ounds. Each type of pheromone cell in
moth species possesses only one membr&ne receptor type,
a ~.peciali:::-t for thr: "Key 11 substance, and thst
ph~romone constituent~ in these species can indeed be
iden~.:.ifi .,d electrophysiolo::::,i cally by scceenin:j; the
cells. Screenin~ tests h~ve been c~rried out in many
species usi~ sin:::;le' cell rGcorJin.;s as WC?LL 1s
reco -,.,di n~~s of 1i'•\G lp..,; econp r 'JO'?o., . 107q·o· K'll. c·~ll' n'~ . "" i \ .l -"- • ' ~ .. , ' :A ' ' ' ''. • . t..d
Prie:.>n0.r (1977) rns r::u-.;~nst.ed th:Jt the reC"p1~0l~s
h:=~ve undergone a stepwL:-;e ch71nge in cert;-:lin sp::ocies.
The ide1 vvcls mainly on electrophysiologic.'3l studies
ol' some moths, shovJin"; a grsdual change in r2c:;ptor
responses of male antennae with the degree of
taxonomic separation (Priesner, 1979a).

78
.Pheromone molecules are filtered out of the ait
str2am by the? 1ntenna and its br:mches anJ sensory
hEucc;. Kai~..:oslin.::; ( 1971) has demonstr.:lted "chat the
cros.c:-sP.ction'il area eyur.;l tu the outline a1·ea of
its :.c1nt en:1a.
'I'he phc~romone molecules removed frcm the
~:~irstream are aJsorbed onto th2 surf·,.'s o:~ th...,
antemw and its sensilla. It hc:~s been su;_;:_.ested
tlYlt only the.c::e moleculPs thAt m;:Jk-:; a di11.=~ct hit
on the pore kattl e are stimulatory, although
Ksissling ( 1C)60) st1tes that the entire sensi:Llum
acts as an ausoj:"bti'!e su1~fr1ce for stimulatory
mol P.cul es. l'hi ~' 8 bso qJtio n phenomenon is non- e.iJ cci fie
(Ka~.~-:lny; and K~~isslin.;;1CJ7:2) and does not contribu.L.e ' ' '
to L-hP 3pt?ci i'i.city of olL~ctory rr·-spon:.3e.
K<1san2.., 3nJ K8is~ling ( 1072) rnPntiorv·d that t.he
S!Y"CiJ'icity o:!.' the' sr~nsory respon~"! must be rel'lted
to the slJeciU.city of the acceptor complex (binuing
site). The:(' c:cc ·1Jtor contplexr::-s (binuins sitss) are
loc;~tPd on ths cc·ll manbrane of t.he denrlritcs o:i' the
s"'nsory n~uronc. (K~1isc:lint;, 1969).

79
The number of accept-or complexes for each r3en-
siL. um is L-.nknovm, althout)l' it probably at least
equals the number of pore tubules (Kaissling, 1()69).
Z3charuk (1971) has sug .. ;r>sted that the specificity
of the s ensil: urn may be a function of the fluid. of
th8 pore tubule.
Se8brook ( 1077) has sppculated that the proteineous
outer coat of the sensillum may contribute to its
specificity and that the bindin"; sites may b~
locatc;d th2re.
In stucii es on the fate of the sex pheromone
molecules of Tl~ichoplusic; ;1i cis -7-dodecen-1-01-acr->t"lte
after reaction with anten~al proteins, FerKovich
·- l (107_,) d·s vo ed· "-'n!:l.L. ev a • . ·' ) l co .Jr L, ·~v both enzym~tic 8nl non
' enzymatic bindin1s occurred. The pheromone wr-~s
degraded to i~s alcohol cis-7-dodecen-1-Ct which
is a potent i nhi bi tor of s ·"'X phenomone behavior in
this species . They sug;es t that T, he inhibitor and
pheromone bind to different pnrts of the proi...ein
or to diJ.'l'erent }Jrotein5·· These J?roteins tllal
bind and degrade the sex pherou~ne were lecated in
the heomolymvh and un the legs as well as on antenna
(i\asan~; :% Kaisslj_ng, 1972; F'erkovich et al., 1r173).

Bo
Schneider (1970) hbs sug~ested that enzymes
C'-ij)8ble of dP-gradin:; pheromones m21y play a d.ouole
role in olf~1ction on the antenna and on the bodies
and legs of insects, to prevent a buildup . of phero-
mu nP. rnol r.:cul ~->f: in these regions.
~ builrlup of these molecules on the body of
the in:.Pct \vould interfer I"Jith the chemical
co .. xuunic~01tion system. Mayer ( 1973) found that the
m::jl" antenna de;:;radeci the pheromone to alcohol
tvvice as fast as did the~ female antenna of Trich<2.2_lusia
ni 3nd that the pheromone w~1s hydrolyzed more rapidly
than six closely related isomers and analogues which
indicates some specificity in the acceptor proteins
for the sex pheromone.
K· sang (1974) calculated that 9C% of the
pheromone molecules of Bombyx mori are absorbed
noncovalently onto micromolecular structures and t~t
large amounts of the pheromone are enzymotically
converced into acids. Little is known of the fate
of the pheromone molecules one e they have reacted with
th<=> ;:mt ,:nnal prot einf. and have been enzywati c ally
dr>grnd""d. l\asaQ.£.!:: (1974), ho,.,over, reports thnt
after ~0 min of incub ~ion with Taitium-labeled
bomykol, H3 molc:c:.J.les h;,_i penetr::Jt~'l to the inner

an upt21k2 of me'cabolic by products by the dendritic
branche3 of the neuron~ •
Kaissling (1972) has proposed a two step proce3s
for the iaactiv~'Jtion of the pheromone molecules:
·1 :st stPp is earl:/ inactiv8tion of the molecule a
pro c e :::s t.hat VJould physical iy or chemic aH y reduce
the concentration. of stimul a tiry mol ec ul e.:, nea r~~r·
the ac eptor and n1ay cont:"ibute to the specificity
o t "C he s y s t em ( K a i s sl j_ ng 19'76) • 2nd step , late
inacti'<'.Jtion is a process that remove the odor
molecules or their reaction product from the sen:3ory
hair:3 ov,c. · a period of minutes. Early activation
accounts for certain characceristies of the :SAG
w;weform anu late activation account-s the rnie;ration
of bombykol molecules 'lii~hin the antenna. Six steps
'3r~ considered itl the tmnsduction of the Olfnctory
stimulus to the receptor potential (Kais::;linr.;, 1974).
l. .\dsor·ptiwn o 1 the antennal survace.
Stimulus {S) ->S '.E,·1s ---/ su rf::~c e
2. Diffusion to the ro:;ceptor moleculr.:s.
3su rf':-•" '" · · --·- + ..... • • ..1\...... \., ""
k----
("<
;:)at r2c epto r
4 ·~ctiv:=Jti·.Jn of receptor molecule
-------;;. A'J -4--·----

S· Ch:.; nge o l' membrane conduct8nc e
!\'S . 6G · lnduces --r membnmce
6 · ~~rly in8ctivation of the odor molecule
Sadsorbed )---~ Satr2coptor( .f---- S inactivated
82
It is dif.ficult to associate the slow enzymatic
d.•~gr'-ldation of bombykol (50% in 4 min) .dth wore
rapid res".Jon:~e time of theEAG whic£1 Boeckh et al.
( 1965) h.:.:Jve shown,may r·each it ~3 maximum voltage
cvithin 0-5 sec. and return to its restin;_; m~mbrane
pot enti3l within 3 sec. of t 0rmim ti on of the
stimulus.
B 1 l .... "l .. o ec,-<: 1 e~, a_.:..., ( 19 65) · ex peri men ts shovv that the
1i ni tial reset ton in the olfaction process· i ~' the
adsorrJti~::-n of thr-; odor;:mt molecule onto the anteltnal
surface. This is follov~ed by dif f·usion of the
molecules to the acceptor complex: where bindi.ng
occurs which r~sults i.l a ch3ng':3 in msrnbn;ne
conductr.JnC'"• The stimulatory molecule aCC'3(Jtor
complex i:3 inact1.v~-1ted by the process of r.arly
i !1Acti vgtion of the odor molecule.
This ol f:::Jctory process reli e.s on the stereo-
chemic~:1l fit of the odor molecule and L:.s ac,:e_titor I
complex, a process sUL:; ested by f<o elofs and
Comeau ( 1971 ) •





























