Embed Size (px)
Citation preview

JOURNAL OF PATHOLOGY, VOL. 178: 21-29 (1996)
IDENTIFICATION OF THE ANTIGEN RECOGNIZED BY THE MONOCLONAL ANTIBODY BU31 AS LAMINS
A AND C PHILIP J. COATES*, R. CARL HOBBS~, JOHN CROCKER~, DAVID c. ROWLANDS§, PAUL MURRAY$, ROY Q U I N L A N I I AND
PETER A. HALL* *Department of Pathology, University of Dundee, Ninewells Hospital and Medical School, Dundee, DDI SSY, U.K.; ?Department
of Histopathology, UMDS, St. Thomas's Hospital, Lambeth Palace Road, London SEI 7EH, U. K.; f Department of Histopathology, East Birmingham Hospital, Birmingham B9 5ST, U. K.; $Department of Pathology, University of Birmingham,
Birmingham BI5 2TT, U. K.; IlDepartment of Biochemistry, University of Dundee, Dundee, U. K.
SUMMARY The murine monoclonal antibody BU31 binds to the nuclear membrane of many cell types. The expression of the BU31 antigen has
previously been shown to have an inverse correlation with the proliferative index in lung tumours, defined by Ki67 staining. The distribution of BU31-positive cells is now shown to parallel the distribution of non-dividing cells in a range of normal human and rat tissues, although neuroendocrine cells and germ cells in the testis show no reactivity. Cells grown in culture and induced to undergo growth arrest show a higher level of labelling with BU31 than their proliferating counterparts. Confocal laser scanning microscopy reveals that the BU31 antigen is distributed predominantly along the nuclear lamina, with occasional internal foci. This distribution is very similar to that of the nuclear membrane proteins lamin A and lamin C, suggesting that the BU31 antigen and lamins A and C could be one and the same. Lmmunoblotting using recombinant lamin proteins confirmed this proposal. Moreover, a monoclonal antibody to the non-proliferation-associated antigen, statin, also recognizes lamins A and C. These data indicate that the demonstration of lamins A and C can be used to provide information on the proliferative activity of normal and neoplastic tissues. These data also suggest a role for nuclear lamins A and C during cellular quiescence, possibly through the reorganization and maintenance of nuclear structure, or more directly through interactions with the retinoblastoma gene product or related proteins.
KEY WORDS-mOnOClOna~ antibodies; nuclear lamina; cell cycle; growth arrest; lamin 4 lamin c
INTRODUCTION
Cellular proliferation is a complex process involving the coordinated expression of a large number of genes. One of the main determinants of the rate of cellular proliferation in adult tissues occurs at the transition from the quiescent state (Go) to the G, phase of the cell cycle.' Molecular analyses of growth control have focused largely on the identification and characteriz- ation of molecules which stimulate cell replication, are induced during the Go to G , transition, or are involved in the subsequent stages of the cell In addition to our rapidly increasing knowledge of the mechanisms governing the cell cycle, there is now a wide variety of methods for assessing cell proliferation both in vitro and in vivo, including the immunocytochemical demonstra- tion of a variety of proliferation-associated antigen^.^-^
In contrast, we know relatively little about the mech- anisms which control cellular quiescence. A number of genes have been identified on the basis that they are expressed at higher levels in non-proliferating cells, although the function of these genes is unclear. The products of the tumour suppressor genes Rb and p53 are also implicated in the negative regulation of cell g r o ~ t h . ~ , ~ Finally, a number of inhibitors of the cyclin-
Addressee for correspondence: P. J. Coates, Department of Pathology, University of Dundee, Ninewells Hospital and Medical School, Dundee DDl 9SY, Scotland, U.K.
CCC 0022-341 7/96/010021-09 0 1996 by John Wiley & Sons, Ltd.
dependent kinases have recently been identified, which may negatively regulate cell-cycle progre~sion.~ To date, however, only one well-characterized immunocyto- chemical marker of non-proliferating cells, termed statin, has been identified. This antigen was discovered through the production of monoclonal antibodies against a cytoskeleton-enriched fraction of senescent fibroblasts.'O A variety of studies have shown that expression of this nuclear membrane antigen is associ- ated with the process of growth arrest's'2 and correlates inversely with the proliferative rate of tumours. I 3 How- ever, we have recently shown that statin is expressed at low levels in some proliferating cells. l4
Besides antibodies to statin, the novel monoclonal antibody BU31 was recently shown to be a marker of non-proliferating cells in a series of lung tumours.15 Like antibodies to statin, BU31 also detects a nuclear membrane-associated antigen. We were interested to characterize further the relationship between immuno- cytochemical staining with BU3 1 and the proliferative activity of cells and tissues. We also wanted to determine any similarities between antibodies to statin and BU31 and to identify the antigens recognized by these reagents, in order to enhance the application of immunocytochemical markers of cellular quiescence. Furthermore, the study of proteins involved in cellular quiescence is important for a fuller understanding of the mechanisms by which cells control their ability to proliferate.
Received 4 January 199.5 Accepted 26 June 199.5

22 P. J. COATES ET AL.
MATERIALS AND METHODS
Zmmunocytochemistry Human and rat tissues were frozen in liquid nitrogen
and stored at - 70°C. Cryostat sections were collected onto silane-coated glass slides, dried for at least 1 h, and stored at - 70°C. Tissue sections or cell cytospins were allowed to warm to room temperature, were fixed in acetone at room temperature for 10 min, and allowed to air-dry. They were rinsed with 0-1 M phosphate-buffered saline, pH 7.4 (PBS) before applying BU31 diluted 1/50 or 1/200 for 45 min at room temperature. Reactive sites were detected by indirect immunoperoxidase, indirect immunofluorescence, or the alkaline phosphatase/anti- alkaline phosphatase technique, using appropriate reagents (Dako Ltd., Bucks, U.K.).
A range of histologically normal human and rat tissues were studied, as well as ten human non- Hodgkin’s lymphomas, comprising five low-grade and five high-grade tumours. In these tumours, BU3 1 imniunoreactivity was assessed by counting a total of 1000 cells for each tumour. In some tissues, sections were additionally stained with a 1/50 dilution of the monoclonal antibody Ki67 (Dako), to assess proliferat- ing cells. Sections of human pancreas and tonsil, and of adult rat testis, were also stained with a monoclonal antibody, S44, which recognizes statin, using avidin- biotin imniunoperoxidase methods as described previ- 0us1y.l~ Control sections and cytospins were stained without application of the primary antibodies or with an anti-rotavirus monoclonal antibody (diluted 1/50 or 11100).
For assessment of bromodeoxyuridine (BrdU) incor- poration, cytospin preparations were fixed in 10 per cent formalin for 10 min. Cells were hydrolised in 1.0 M HCl at 60°C for 10 min, washed well in water, and digested with 0.01 per cent (w/v) protease type XXIV (Sigma Chemical Co. Ltd., Dorset, U.K.) in PBS for 10 min at 37°C. After washing in PBS, a monoclonal antibody to BrdU (Dako) was applied, diluted 1/50 for 30 min at room temperature; reactive sites were revealed by the indirect immunoperoxidase technique.
Cell culture
Swiss 3T3 murine fibroblasts, Rat-1 fibroblasts or HeLa human cervical carcinoma cells were grown in Dulbecco’s modified Eagle’s medium (DMEM) contain- ing 10 per cent fetal calf serum (FCS; ICN Biomedicals Ltd., Thame, Oxfordshire, U.K.). In initial experiments, rapidly growing cells (approximately 20-30 per cent confluence) were compared with cells which had become confluent and had been maintained in reduced serum (0.5 per cent FCS) for a further 48-72 h prior to analysis.
In subsequent experiments, replica flasks of cells were seeded at a density of 10 per cent confluency and maintained in medium containing 10 per cent FCS for up to 12 days, with the medium being changed every 24 h. Alternatively, cells were allowed to grow for 48 h before the serum concentration was reduced to 0.5 per cent. Cells were maintained for 72 h under reduced
serum conditions before serum was reintroduced to 10 per cent. In each case, cells were incubated with I O ~ M BrdU (Sigma) for 1 h before harvesting.
Cells were collected by trypsinization. The cells were resuspended at a concentration of approximately 5 x lo5 cells/ml and 200 pl of the suspension was centrifuged onto glass slides using a centrifuge (Shandon Scientific Ltd., Runcorn, Cheshire, U.K.) operating at 800 rpm for 3 min. Slides were allowed to dry for 30 min at room temperature, wrapped in aluminium foil, and stored at -70°C. For quantitation of the number of BrdU- or BU31-positive cells, at least 500 cells were counted for each time point.
Confocal microscopy
Rat-1 cells at approximately 50 per cent confluence were trypsinized and spun onto glass coverslips using a cytocentrifuge operating at low speed (500 rpm). Cells were air-dried, fixed in acetone, and stained with BU31 as before, but using FITC-labelled rabbit anti-mouse immunoglobulins (Dako) to identify reactive sites. Coverslips were mounted onto glass slides and viewed with a Biorad MRC 600 confocal laser scanning microscope.
Zmmunoblotting
Bacterially expressed recombinant human lamins A, B, and C were produced by expressing PET vectors containing the appropriate cDNA sequence in Escherichia coli. After induction of protein expression, the lamins were purified by ion-exchange chromatogra- phy.16 Lamins were separated by SDS-PAGE, using either a 7.5-15 per cent gradient gel, or 8 or 10 per cent polyacrylamide resolving gels, l 7 and transferred onto nitrocellulose or PVDF membranes and blots were stained with 0.3 per cent Ponceau S in 3 per cent trichloroacetic acid to ensure adequate transfer of pro- teins. The membranes were rinsed in PBS and blocked for 2 h at room temperature in 5 per cent non-immune rabbit serum (NRS) in PBS. The membranes were incubated overnight at room temperature in primary antibodies diluted in PBS containing 5 per cent NRS. BU31 was used at a dilution of 1/50 and S44 was used diluted 1/200. After extensive washing with PBS, peroxidase-conjugated rabbit anti-mouse immunoglobu- lin (Dako) diluted 1/500 in PBS containing 1 per cent bovine serum albumin was applied for 1 h at room temperature. Immunoreactive bands were identified with 3,3’-diaminoben~idine’~ or with the ECL chemilumines- cent detection system (Amersham, Little Chalfont, Bucks, U.K.). As a control the primary antibody was omitted.
RESULTS BU31 immunoreactivity is distributed in non-proliferating zones of many tissues
In acetone-fixed sections from all tissues examined, BU31 stained the cell nucleus exclusively, with a pre- dominance of staining in the nuclear membrane and

BU31 ANTIGEN IS LAMINS AIC 23
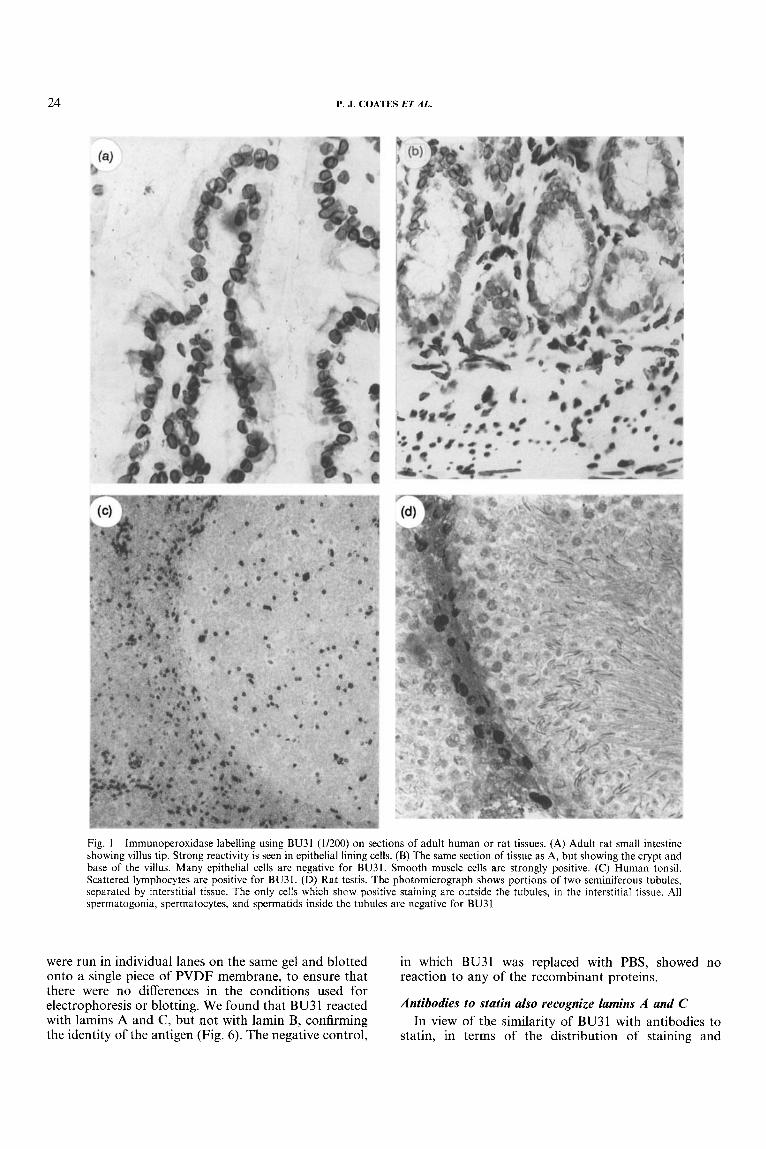
with lower intensity in the nucleus itself. In sections of normal small or large intestine stained with a 1/200 dilution of BU31, occasional cells towards the base of the epithelial crypts were negative for BU3 1. Other cells in this area were often weakly stained. Epithelial cells higher up the crypt and on the surface of the lumen were consistently strongly positive for BU3 1. Stromal cells, smooth muscle cells, and endothelial cells were also strongly BU31-positive (Figs 1A and 1B). These patterns of staining are the inverse of those seen with markers of proliferation. Increasing the concentration of BU3 I resulted in fewer negative cells being seen in the crypts.
In human tonsil, approximately 5 per cent of follicle centre cell nuclei were positive. These cells had the appearance of small lymphocytes. In addition, about 5 per cent of scattered lymphoid cells outside the follicles were also stained with BU31 (Fig. IC). In contrast, over 95 per cent of stromal cell nuclei, endothelial cell nuclei, and vascular smooth muscle cell nuclei were positive. In sections containing surface squamous epithelial cells, it was noticed that occasional basal and parabasal cells were negative, whilst the remaining cells were positive.
In sections from normal adult human thyroid, kidney, liver, lung, skeletal muscle, brain, adrenal and salivary gland, greater than 95 per cent of all cells were BU31- positive. In sections of pancreas, the majority of acinar and ductal cells were stained with BU31, as were endothelial cells and vascular smooth muscle cells. One striking feature of this tissue was the lack of staining in the majority of cells in the islets of Langerhans. In all of these tissues, essentially identical results were obtained using BU31 diluted 1/50 or 11200.
Very similar patterns of staining were seen in sections from a variety of adult rat tissues stained with BU3 1. In sections of skin, occasional cells at the base of the epithelium were negative for BU31, whilst all keratino- cytes in the squamous layer were strongly positive. Dermal fibroblasts and endothelium were strongly BU3 1 -positive. Similarly, in sections of oesophagus, the majority of the epithelial lining cells were positive, with a minority of cells at the base of the epithelium being negative. Again, fibroblasts and endothelium were con- sistently BU3 1-positive. In each of these tissues, increas- ing the concentration of BU31 led to a decreased number of negative cells in the proliferative compart- ment of the epithelium. In sections of brain, liver, lung, and kidney, approximately 95 per cent of all cells showed positive staining, including endothelial cells, smooth muscle cells, and fibroblasts. In a section of adult testis, however, the majority of cells within the testicular tubules were negative for BU3 1, including all spermatocytes, spermatids, and sperm. Leydig cells and other cells outside the tubules were positive for BU31 (Fig. 1D). In these tissues, the results were unchanged by using an increased concentration of BU3 1.
In a preliminary study of ten non-Hodgkin’s lympho- mas divided into high-grade and low-grade groups, the number of BU3 1 -positive cells varied from 1 1.6 to 18.4 per cent in high-grade tumours and from 56.3 to 8 1.4 per cent in low-grade tumours. These values are the inverse of measurements of cell proliferation for these groups of tumours.”
BU31 staining relates to proliferative activity in vitro Growth arrest of 3T3 murine fibroblasts, induced by
contact inhibition and serum deprivation for 48 h, resulted in a dramatic increase in the percentage of BU31-positive cells, from 30.2 to 98.5 per cent. Simi- larly, the proportion of HeLa cells which showed immunostaining with BU3 1 increased from 9.2 per cent in rapidly growing cultures to 90.5 per cent after contact inhibition and serum deprivation. These results suggested that the level of the BU31 antigen relates to the proliferative activity of the cell cultures.
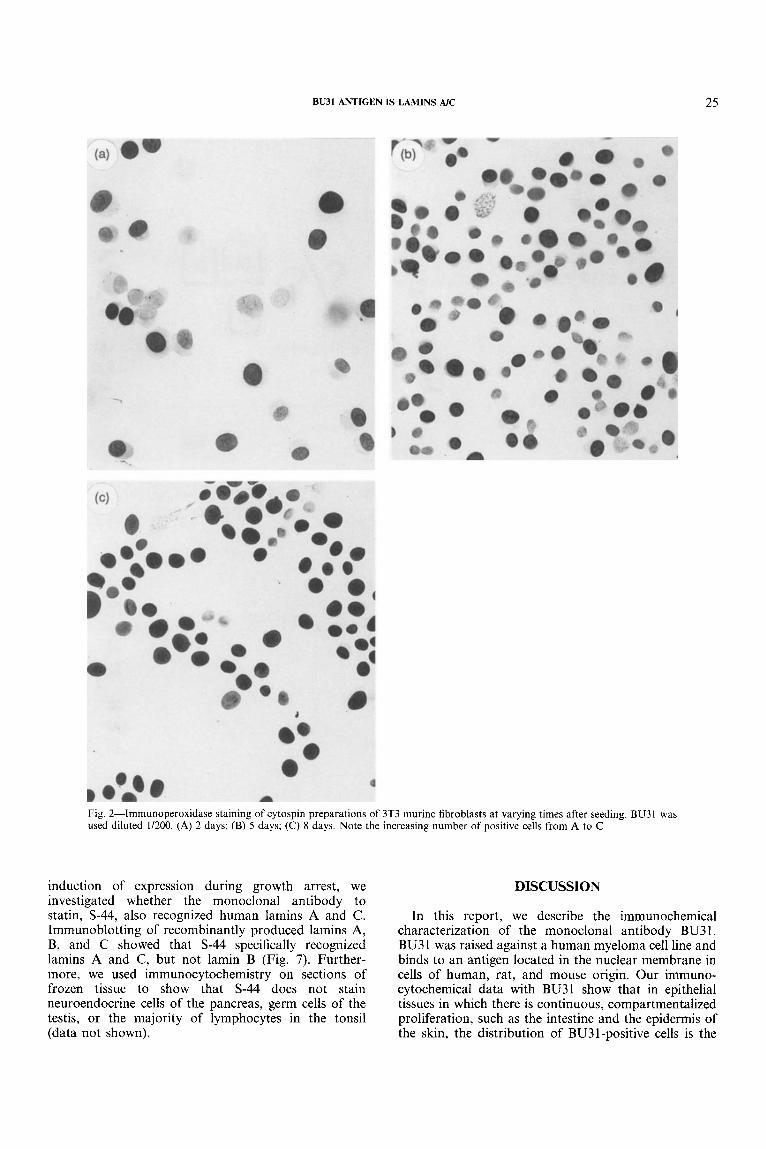
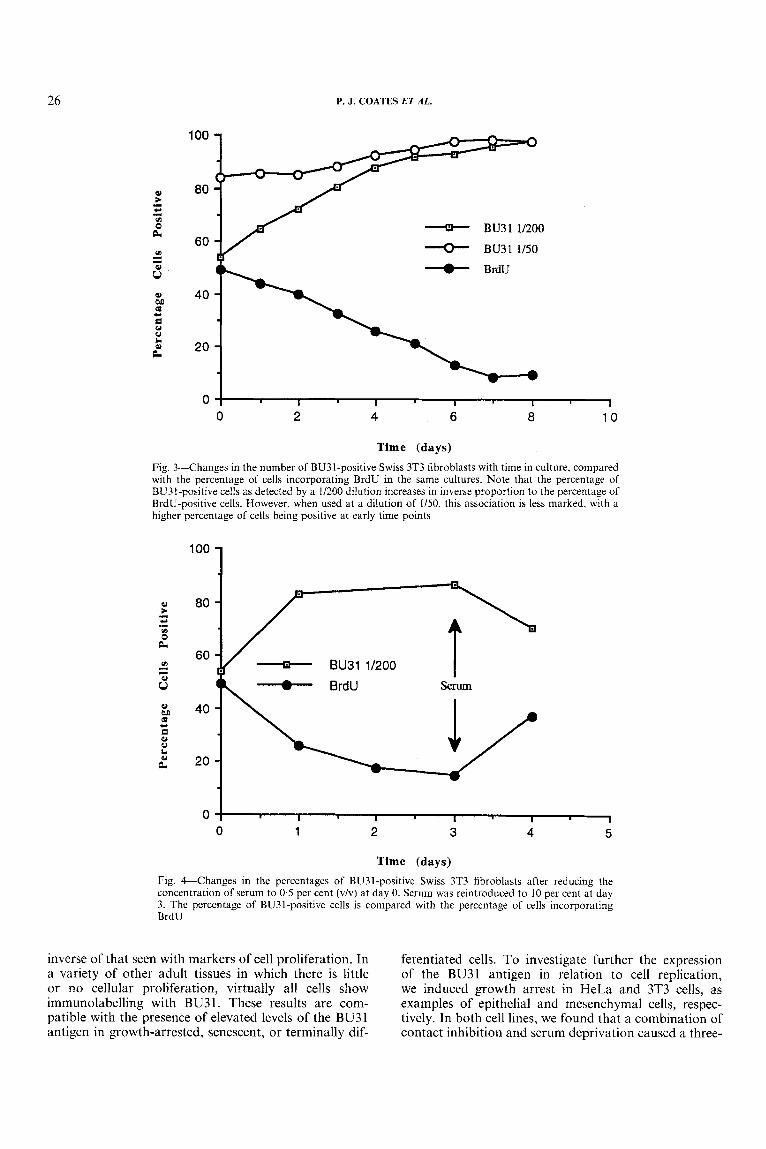
To define further the relationship between BU3 1 staining and proliferative activity, we examined the time course of induction of BU31-positive nuclei in 3T3 fibroblasts and compared this with the extent of prolif- eration as measured by the incorporation of BrdU. In the first experiment, cells were seeded at a density of 10 per cent confluency and allowed to grow in the presence of 10 per cent serum. We found that the percentage of BU3 1 -positive nuclei increased gradually throughout the course of this experiment, coinciding with a gradual decrease in the percentage of nuclei which contained BrdU (Figs 2 and 3). In the second experi- ment, we investigated the effect of reduced serum con- centration on the induction of BU31 -positive cells. We found that the number of BU31-positive nuclei was increased within 24 h of reducing the concentration of serum from 10 to 0.5 per cent. The percentage of BU3 1 -positive cells in serum-deprived cultures remained high for 72 h and then decreased within 24 h after re-introducing serum. This pattern mirrors the pro- liferative activity of the cultures, as measured by the percentage of nuclei which have incorporated BrdU (Fig. 4).
BU31 binds to the nuclear lamina, with occasional internal foci of staining
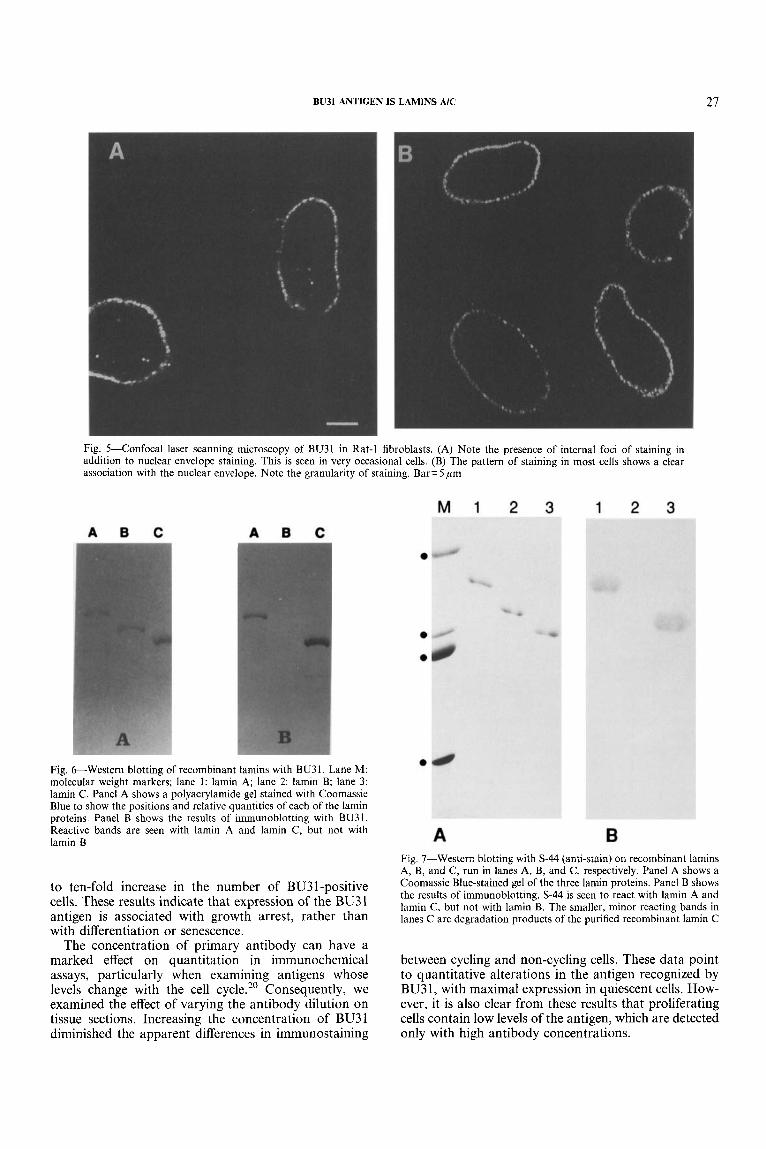
We used confocal laser scanning microscopy to ana- lyse the subcellular distribution of the BU31 antigen. In non-synchronously proliferating Rat- 1 fibroblasts, we identified two distinct patterns of staining. A small proportion of cells showed the presence of dots within the body of the nucleus, as well as staining at the nuclear membrane (Fig. 5A). To confirm that these dots were present inside the nucleus, a three-dimensional recon- struction was created from a series of images taken in the z-plane. This clearly showed that the foci of staining were contained in the body of the nucleus and were not simply invaginations of the nuclear membrane (data not shown). The vast majority of cells showed a clear nuclear membrane distribution of staining, which often appeared quite granular (Fig. 5B).
BU31 recognizes lamins A and C With the results obtained thus far, we thought that
BU31 may recognize the nuclear lamin proteins. To identify the antigen, we used immunoblotting techniques against the three major human lamins, expressed as recombinant proteins in bacteria. Lamins A, B, and C
24 P. J. COATES ET AL.
Fig. I-Immunoperoxidase labelling using BU31 (11200) on sections of adult human or rat tissues. (A) Adult rat small intestine showing villus tip. Strong reactivity is seen in epithelial lining cells. (B) The same section of tissue as A, but showing the crypt and base of the villus. Many epithelial cells are negative for BU31. Smooth muscle cells are strongly positive. (C) Human tonsil. Scattered lymphocytes are positive for BU31. (D) Rat testis. The photomicrograph shows portions of two seminiferous tubules, separated by interstitial tissue. The only cells which show positive staining are outside the tubules, in the interstitial tissue. All spermatogonia, sperrnatocytes, and spermatids inside the tubules are negative for BU3 1
were run in individual lanes on the same gel and blotted onto a single piece of PVDF membrane, to ensure that there were no differences in the conditions used for electrophoresis or blotting. We found that BU3 1 reacted with lamins A and C , but not with lamin B, confirming the identity of the antigen (Fig. 6). The negative control,
in which BU31 was replaced with PBS, showed no reaction to any of the recombinant proteins.
Antibodies to Statin a h f'ecognize lamins A and c In view of the similarity of BU31 with antibodies to
statin, in terms of the distribution of staining and
BU31 ANTIGEN I S LAMINS AIC 25
Fig. 2-Imrnunoperoxidase staining of cytospin preparations of 3T3 murine fibroblasts at varying times after seeding. BU3 1 was used diluted 1/200. (A) 2 days; (B) 5 days; (C) 8 days. Note the increasing number of positive cells from A to C
induction of expression during growth arrest, we investigated whether the monoclonal antibody to statin, S-44, also recognized human lamins A and C. Immunoblotting of recombinantly produced lamins A, B, and C showed that S-44 specifically recognized lamins A and C, but not lamin B (Fig. 7). Further- more, we used immunocytochemistry on sections of frozen tissue to show that S-44 does not stain neuroendocrine cells of the pancreas, germ cells of the testis, or the majority of lymphocytes in the tonsil (data not shown).
DISCUSSION
In this report, we describe the immunochemical characterization of the monoclonal antibody BU31. BU3 1 was raised against a human myeloma cell line and binds to an antigen located in the nuclear membrane in cells of human, rat, and mouse origin. Our immuno- cytochemical data with BU31 show that in epithelial tissues in which there is continuous, compartmentalized proliferation, such as the intestine and the epidermis of the skin, the distribution of BU31-positive cells is the
26 P. J. COATES ET AL.
100
80
60
40
20
0
Y BU31 1/200
BU31 1/50 BrdU
I 0 2 4 6 8 10
I I I I I
Time (days) Fig. 3--Changes in the number of BU31-positive Swiss 3T3 fibroblasts with time in culture, compared with the percentage of cells incorporating BrdU in the same cultures. Note that the percentage of BU31-positive cells as detected by a 1/200 dilution increases in inverse proportion to the percentage of BrdU-positive cells. However, when used at a dilution of 1/50, this association is less marked, with a higher percentage of cells being positive at early time points
loo 1 8 t - c *I
v) 0 &
v) - I s Q) M co C 8 u L
c
z
I I I I I
0 1 2 3 4 5
Time (days) Fig. &Changes in the percentages of BU31-positive Swiss 3T3 fibroblasts after reducing the concentration of serum to 0.5 per cent ( v h ) at day 0. Serum was reintroduced to 10 per cent at day 3. The percentage of BU31-positive cells is compared with the percentage of cells incorporating BrdIJ
inverse of that seen with markers of cell proliferation. In a variety of other adult tissues in which there is little or no cellular proliferation, virtually all cells show immunolabelling with BU3 1. These results are com- patible with the presence of elevated levels of the BU31 antigen in growth-arrested, senescent, or terminally dif-
ferentiated cells. To investigate further the expression of the BU31 antigen in relation to cell replication, we induced growth arrest in HeLa and 3T3 cells, as examples of epithelial and mesenchymal cells, respec- tively. In both cell lines, we found that a combination of contact inhibition and serum deprivation caused a three-
BU31 ANTIGEN IS LAMINS AIC 27
Fig. 5-Confocal laser scanning microscopy of BU31 in Rat-1 fibroblasts. (A) Note the presence of internal foci of staining in addition to nuclear envelope staining. This is seen in very occasional cells. (B) The pattern of staining in most cells shows a clear association with the nuclear envelope. Note the granularity of staining. Bar= 5 ,um
Fig. &-Western blotting of recombinant lamins with BU31. Lane M: molecular weight markers; lane 1: lamin A; lane 2: lamin B; lane 3: lamin C . Panel A shows a polyacrylamide gel stained with Coomassie Blue to show the positions and relative quantities of each of the lamin proteins. Panel B shows the results of immunoblotting with BU31. Reactive bands are seen with lamin A and lamin C, but not with lamin B
to ten-fold increase in the number of BU3 1 -positive cells. These results indicate that expression of the BU31 antigen is associated with growth arrest, rather than with differentiation or senescence.
The concentration of primary antibody can have a marked effect on quantitation in immunochemical assays, particularly when examining antigens whose levels change with the cell cycle.20 Consequently, we examined the effect of varying the antibody dilution on tissue sections. Increasing the concentration of BU3 1 diminished the apparent differences in immunostaining
Fig. 7-Western blotting with S-44 (anti-stain) on recombinant lamins A, B, and C, run in lanes A, B, and C, respectively. Panel A shows a Coomassie Blue-stained gel of the three lamin proteins. Panel B shows the results of immunoblotting. S-44 is seen to react with lamin A and lamin C, but not with lamin B. The smaller, minor reacting bands in lanes C are degradation products of the purified recombinant lamin C
between cycling and non-cycling cells. These data point to quantitative alterations in the antigen recognized by BU31, with maximal expression in quiescent cells. How- ever, it is also clear from these results that proliferating cells contain low levels of the antigen, which are detected only with high antibody concentrations.

28 P. J. COATES ET AL.
A number of factors led us to suspect that BU31 may be recognizing lamins A and C, or a closely associated molecule. Firstly, lamins A and C are major proteins of the nuclear membrane. They share an identical amino acid sequence, with lamin A possessing additional amino acids at its C-terminus.21 Secondly, the subcellular dis- tribution of immunostaining with BU3 1 is remarkably similar to the distribution of lamins A and C, including the presence of internal structures in occasional nuclei.16*22 Thirdly, the lack of staining of BU31 in germ cells of the testis and in the majority of lymphocytes may relate to the lack of lamins A and C in these Finally, the unusual tissue distribution of staining with BU31 in normal adult tissues, and the lack of staining of neuroendocrine cells of the pancreas, is similar to that recently described using polyclonal antibodies against lamins A and C.25 That BU31 does indeed recognize lamins A and C is demonstrated by immunoblotting of recombinant proteins. The specificity of this reaction is shown by the lack of binding to lamin B.
In view of the similarities between BU31 and statin, which are both selective for the nuclear membrane of non-proliferating cells, we investigated whether anti- bodies to statin recognize lamins A and c. Using the S-44 antibody to statin, we showed by immunoblotting that this antibody also recognizes lamins A and C. Our finding differs from other reports, which have suggested that statin is a 57 kD protein, distinct from the nuclear lamins.’0326 Moreover, a putative gene coding for statin has been identified which bears no obvious homology to lamins A and C.27 However, antibodies to statin bind to a variety of protein species in cells, as evidenced by the multiple and variable reactive bands seen on Western blots probed with S-44.26 In view of the nuclear localiz- ation and tissue distribution of staining with antibodies to tati in,'^^'^^'^ it . is likely that the immunocytochemical detection of statin reactivity is due to the reaction with lamins, rather than to any other protein.
lmmunocytochemical data with BU3 1 and antibodies to statin thus indicate that the levels of lamins A and C are increased in quiescent cells. This suggestion provides an alternative explanation for the differences in expres- sion of lamins A and C previously observed in the context of cellular differentiati~n.~~-~’ Lamins A and C are thought to be involved in the regulation of gene expression, through attachments with ~ h r o r n a t i n . ~ ~ These interactions help to control the three-dimensional organization of the nucleus, which is believed to be important for regulating gene e x p t - e s ~ i o n . ~ ~ - ~ ~ Thus, our results suggest that low levels of lamins A and C are characteristic of cells which have a lesser degree of organized structure to their nucleus, reflecting the need of these cells to alter gene expression rapidly during the cell cycle, and to remain adaptable and able to differen- tiate along any of a variety of pathways. In contrast, higher levels of lamins A and C reflect the stability of differentiated, non-proliferating cells, which require only limited changes in gene expression.
A more direct mechanism by which the levels of lamins A and C may influence the cell cycle is through interactions with the retinoblastoma gene product (Rb) or related proteins. It has recently been shown that
hy ophosphorylated Rb associates with lamins A and
with phosphorylation leading to release from the lamina and cell-cycle progression. Hypophosphorylated Rb is known to bind to and inactivate molecules involved in cell-c cle progression, including the transcription factor E2F. Therefore, one consequence of Rb binding to the lamins would be the sequestration of key regulatory proteins on the nuclear lamina. The importance of Rb-lamin interactions in regulating the cell cycle is strengthened by observations indicating that Rb may bind to lamins A and C at the same site as Rb-E2F interactions
In conclusion, we have described the characterization of the monoclonal antibody BU3 1, which selectively binds to the nuclear membrane of non-proliferating cells, and have shown that the antibody recognizes the lamins A and C. Similarly, antibodies to the previously described quiescence-associated protein statin also rec- ognize lamins A and C. Thus, the use of antibodies to lamins A and C provides a mean for identifying non- proliferating cells in normal and neoplastic tissues and should assist with the subdivision of cell populations in histological material.
C. P5 This form of Rb predominates during early GI ,
J
ACKNOWLEDGEMENTS
We are grateful to N. Ling for providing the mono- clonal antibody BU31 and E. Wang for s-44 anti-statin. We wish to thank the Cancer Research Campaign for generous financial support (Grant SP2148 to PAH and Grant SP2060 to RQ).
REFERENCES
1.
2.
3.
4. 5.
6.
7.
8.
9.
10.
I I .
12.
13.
14.
Pardee AB. G I events and regulation of cell proliferation. Science 1989; 246 603-608. Murray AW, Hunt T. The Cell Cycle: An Introduction. Oxford: Oxford University Press, 1993. Hall PA, Coates PJ. Assessment of cell proliferation in pathology. What next? ffistc~pafhu/ogy 1995; 2 6 105-1 12. Sherr CJ. Mammalian G, cycliiis. CrN 1993; 7 3 1059--1065. Hall PA, Levison DA, Wright NA (Eds). Assessment of Cell Proliferation in Clinical Practice. London: Springer-Verlag. I99 I . Gerdes J, Schwab U, Lemke H, Stein H. Production of monoclonal antibody reactive with a human nuclear antigen associated with cell proliferation. Int J Cancer 1983; 31: 13-20, Hall PA, Levison DA, Woods AL, ef a/ . Proliferating cell nuclear antigen (PCNA) immunolocalization in paraffin sections: an index of cell prolifer- ation with evidence of deregulated expression in some neoplasms. J fathol 1990; 162 285-294. Garrido MC, Cordell JL, Becker MHG. c( a/. Monoclonal antibody JCI: new reagent for studying cell proliferation. J Clin Puthol 1992; 45: 860-865. Sampson SA, Kreipe H, Gillett CE, rt ill KiSl--ii novel monoclonal antibody which recognizes proliferating cells: evaluation of its relationship to prognosis in mammary carcinoma. J frrthol 1992; 168: 179-185. Wang E. A 57 000-mol-wt protein uniquely present in nonproliferating cells and senescent human fibroblasts. J C d B i d 1YX5; 100: 545--551. Wang E. Rapid disappearance of statin, a iionproliferating and senescent cell-specific protein, upon reentering the process of cell cycling. J (‘el/ B id 1985; 101: 1695-1701. Wang EA, Kreuger JG. Application of a unique monoclonal antibody as a marker for nonproliferating subpopulations o f cells of some tissues. J Histocl?r.m C’tochem 1985; 3 3 587-594. Tsanaclis AMC, Brem SS, Gately S, Schipper HM, Wang E. Statin immunolocalization in human brain tumours. Detection of noncycling cells using a novel marker of cell quiescence. C m c w 1991; 6 8 786-792. Ansari B, Dover R, Gillmore CP. Hall PA. Expression of the nuclear membrane protein statin in cycling cells. J fafliol 1993; 169 391-396.

BU31 ANTIGEN IS LAMINS AIC 29
15. Rowlands DC, Bunce CM, Crocker J, el a/. Expression of a nuclear envelope protein recognized by the monoclonal antibody BU31 in lung tumours: relationship to Ki-67 antigen expression. J Pathol 1994; 173 89-96.
16. Goldman AE, Moir RD, Montag-Lowy M, Stewart M, Goldman RD. Pathway of incorporation of microinjected lamin A into the nuclear envelope. J Cell Biol 1992; 119 725-735.
17. Laemmli UK. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nalure 1970; 227: 680-685.
18. Harlow E, Lane DP. Antibodies. A Laboratory Manual. New York: Cold Spring Harbor Laboratory, 1988.
19. Hall PA, Gregory W, Richards MA, d’Ardenne AJ, Lister TA, Stansfeld AG. The prognostic significance of Ki67 immunostaining in non-Hodgkin’s lymphomas. J Pathol 1988; 154 223-235.
20. McCormick D, Yu C, Hobbs C, Hall PA. The relevance of antibody concentration to the immunohistological quantification of cell proliferation- associated antigens. His&patholugy 1993; 2 2 543-547.
21. Fisher DZ, Chaudhary N, Blobel G, cDNA sequencing of nuclear lamins A and C reveals primary and secondary structural homology to intermediate filament proteins. Proc Natl Acad Sci USA 1986; 83: 645M454.
22. Bridger JM, Kill IR, O’Farrell MO, Hutchinson CJ. Internal lamin struc- tures within G, nuclei of human dermal fibroblasts. J Cell Sci 1993; 104: 297-306.
23. Moss SB, Burnham BL, Bellve AR. The differential expression of lamin epitopes during mouse spermatogenesis. Mol Reprod Dev 1993; 3 4 164174.
24. Rober RA, Sauter H, Weber H, Osborn M. Cells of the cellular immune and hemopoietic system of the mouse lack lamins AIC: distinction versus other somatic cells. J Cell Sci 1990; 95: 587-598.
25. Cance WG, Chaudhary N, Worman HJ, Blobel G, Cordon-Cardo C. Expression of the nuclear lamins in normal and neoplastic human tissues. J Exp CIin Cuncer Res 1992; 11: 233-246.
26. Sester U, Moutsatsos IK, Wang E. A rat liver 57-kDa protein is identified to share antigenic determinants with statin, a marker for non-proliferating cells. Exp Cell Res 1989; 182 550-558.
27. Ann DK, Moutsatsos IK, Nakamura T, et al. Isolation and characterization of the rat chromosomal gene for a polypeptide (PSI) antigenically related to statin. J Biol Chem 1991; 266: 10429-10437.
28. Lebel S, Lampron C, Royal A, Raymond Y . Lamins A and C appear during retinoic acid-induced differentiation of mouse embryonal carcinoma cells. J Cell Bid 1987; 105 1099-1 104.
29. Rober RA, Weber K, Osborn M. Differential timing of nuclear lamin A/C expression in the various organs of the mouse embryo and the young animal: a developmental study. Development 1989; 105: 365-378.
30. Collard JF, Seneca1 JL, Raymond Y. Redistribution of nuclear lamin A is an early event associated with differentiation of human promyelocytic leukemia HL-60 cells. J Cell Sci 1992; 101: 657-670.
31. Yuan J, Simos G, Blobel G, Georgatos SD. Binding of lamin A to polynucleosomes. J Bid Chem 1991; 266 921 1-9215.
32. Blobel G. Gene gating: a hypothesis. Proc Nut1 Acud Sci USA 1985; 8 2 8527-8529.
33. Nigg EA. Assembly and cell cycle dynamics of the nuclear lamina. Semin Cell Biol 1992; 3: 245-253.
34. Xing Y, Johnson CV, Dobner PR, Lawrence JB. Higher level organization of individual gene transcription and RNA splicing. Science 1993; 259 1326-1330.
35. Mancini MA, Shan B, Nickerson JA, Penman S, Lee W-H. The retinoblas- toma gene product is a cell cycle-dependent, nuclear matrix-associated protein. Proc Nut1 Acad Sci USA 1994; 91: 418422.
36. Ozaki T, Saijo M, Murakami K, Enomoto H, Taya Y , Sakiyama S. Complex formation between lamin A and the retinoblastoma gene product: identification of the domain on lamin A required for its interaction. Oncogene 1994; 9 2649-2653.





























