Embed Size (px)
Citation preview

VIROLOGY 231, 231– 238 (1997)ARTICLE NO. VY978517
Identification of Regions in HIV-1 Nef Required for EfficientDownregulation of Cell Surface CD4
Jian Hua, Wade Blair, Ray Truant, and Bryan R. Cullen1
Howard Hughes Medical Institute and Department of Genetics, Duke University Medical Center, Durham, North Carolina 27710
Received January 9, 1997; accepted February 25, 1997
Downregulation of cell surface CD4 is a characteristic property of all lentiviral Nef proteins. We have used mutationalanalysis to define regions within HIV-1 Nef that are critical for this biological activity. Two discontinuous regions in Nef,extending approximately from residues 96 to 144 and from residues 175 to 186, are reported to be essential for efficientCD4 downregulation. Interestingly, these sequences coincide with two conserved regions of the Nef protein that arejuxtaposed to form a single surface on the known structure of Nef. A third, more amino terminal conserved region in Nef,previously reported to be important for Nef enhancement of virion infectivity, was found to be largely dispensable for CD4downregulation. These data raise the possibility that Nef may contain two structurally distinct functional domains, only oneof which contributes to the CD4 downregulation phenotype. q 1997 Academic Press
INTRODUCTION this downregulation remains to be established, loss ofsurface CD4 does block viral superinfection and may
Although the Nef protein encoded by the primate len- facilitate progeny virion release from infected cells (Ben-tiviruses is essential for efficient viral replication and son et al., 1993). While CD4 downregulation is a highlypathogenesis in vivo (Kestler et al., 1991; Jamieson et al., conserved property of both human and simian Nef iso-1994), its role(s) in the viral life cycle remains unclear lates (Anderson et al., 1993; Benson et al., 1993; Mariani(Reviewed by Cullen, 1994). Effective analysis of Nef and Skowronski, 1993), the mechanism of downregula-function has been made more difficult by the general tion remains unclear. It also remains uncertain whetherobservation that this viral gene product is dispensable enhanced virion infectivity, CD4 downregulation, and thefor efficient replication of both human immunodeficiency reported effects of Nef on the activation state of cellsvirus type 1 (HIV-1) and simian immunodeficiency virus (Bandres and Ratner, 1994; Baur et al., 1994; Du et al.,(SIV) in culture. More recently, evidence has been pre- 1995) are aspects of a single biological activity or insteadsented by several groups demonstrating that Nef can reflect mechanistically distinct functions of Nef.enhance the infectivity of virions released from Nef-ex- One way to address this latter issue is to mutationallypressing cells under some experimental conditions (De define regions within Nef that are essential for the vari-Ronde et al., 1992; Zazopoulos and Haseltine, 1992; ous reported biological activities of Nef. Here, we reportChowers et al., 1994; Miller et al., 1994; Aiken and Trono, that two distinct regions of Nef are critical for CD4 down-1995). However, this effect is generally modest and ap- regulation. Of interest, these two regions are closely jux-pears to vary significantly with the source and/or activa- taposed in the recently published structure of the HIV-1tion state of the cells used in the replication assay (Ryan- Nef protein and may therefore form a single functionalGraham and Peden, 1995). For example, the nef gene domain.expressed by the SIVmac239 proviral clone, while clearlybiologically active in infected primates, proved unable to
MATERIALS AND METHODSenhance viral replication in either primary simian lympho-cytes or macrophages in culture (Kestler et al., 1991). Molecular clones
A second, readily reproducible property of Nef is down-regulation of cell-surface CD4 receptor expression (Gar- The full-length nef gene present in the NL-43 HIV-1cia and Miller, 1991; Benson et al., 1993; Aiken et al., proviral clone was amplified by polymerase chain reac-1994; Rhee and Marsh, 1994). Although the reason for tion (PCR) and then inserted into the expression plasmid
pBC12/CMV (Cullen, 1986) to generate pcNef/NL-43. Mu-tated forms of the NL-43 nef gene, encoding the mutant1 To whom correspondence and reprint requests should be ad-Nef proteins indicated in Fig. 1, were generated by PCRdressed at Department of Genetics, Duke University Medical Center,mutagenesis using Pfu DNA polymerase (Stratagene).Box 3025, CARL Building, Research Drive, Durham, NC 27710. Fax:
(919) 681-8979. E-mail: Internet, [email protected]. The identity of the resultant Nef mutants was confirmed
2310042-6822/97 $25.00Copyright q 1997 by Academic PressAll rights of reproduction in any form reserved.
AID VY 8517 / 6a33$$$121 04-11-97 13:09:26 vira AP: Virology

232 HUA ET AL.
by DNA sequencing using the Sequenase version 2.0 were highlighted, from its Brookhaven PDB coordinatesusing RasMac v2.5-ucb1.3 software.kit (Amersham). A cDNA encoding full-length wild-type
human CD4 was generated by PCR and expressed usingthe pBC12/CMV vector (pcCD4). The expression plasmid RESULTSpIL-2Ra, encoding the full-length human IL-2 receptor achain, has been described (Cullen et al., 1988). The HIV-1 Nef protein is myristoylated at its amino
terminus and is associated with cytoplasmic membranesCD4 downregulation assay in expressing cells (Hammes et al., 1989). Analysis of
protein sequence conservation in Nef isolates obtainedCD4 downregulation assays were performed usingfrom a large number of infected patients has identifiedtransiently transfected 293T cell cultures (35 mm). Thefive conserved regions within Nef (Shugars et al., 1993).transfection cocktail contained 14 ml of lipofectamine re-These are the short, amino-terminal myristoylation signalagent (Life Technologies, Inc.), 3 mg of pcCD4, 4 mg ofand the four sequence blocks, termed A through D, indi-the IL-2 receptor a-chain expression plasmid pIL-2Ra,cated in Fig. 1. Little is currently known about criticaland 7.5 mg of the relevant Nef expression plasmid. Thesequence motifs in Nef. However, four proline residuesparental pBC12/CMV plasmid served as a negative con-located between position 69 and 78 in Nef, as well as atrol. At 85 hr posttransfection, the transfected cells wereproline located at position 147, have been reported toharvested as a single-cell suspension and washed twiceform an SH3 motif that is important in mediating an inter-with cold PBS containing 4 mM EDTA and 2% calf serum.action between Nef and cellular tyrosine kinases (Sak-The cells were then stained using a 1:100 dilution of bothsela et al., 1995). In addition, two arginines located ata fluorescein (FITC) conjugated anti-human IL-2 receptorpositions 105 and 106 in Nef have been reported to bea-chain antiserum (Pharmingen) and a phycoerythrincritical for the interaction of Nef with a cellular serine/(PE) conjugated anti-human CD4 antiserum (Phar-threonine kinase (Sawai et al., 1995). The functional im-mingen) for 30 min on ice. After two washes with coldportance of these kinase interactions remains of unclearPBS, the stained cells were fixed with 4% paraformalde-significance, although mutation of either of these twohyde and surface CD4 and IL-2 receptor a-chain expres-interaction sites does result in a loss of the enhancedsion quantified by two-color fluorocytometric analysis us-virion infectivity phenotype of Nef (Goldsmith et al., 1995;ing FACScan (Becton Dickinson). Surface IL-2 receptorSaksela et al., 1995; Wiskerchen and Cheng-Mayer,a-chain expression was not affected by Nef and thus1996).served as a marker for transfected cells.
To begin to map regions within Nef that are im-portant for CD4 downregulation by Nef, we introducedWestern blot analysis17 scattered mutations into the 206-amino-acid NL-43
Human 293T cells (35-mm cultures) were transfected Nef protein (Fig. 1). Each mutation introduced eitherusing lipofectamine using 4 mg of the wild-type or mutant a single or a double missense mutation. These muta-Nef expression plasmids. The parental pBC12/CMV vec- tions are named according to the location and identitytor served as a negative control. At 72 hr after transfec- of the mutated amino acids and the substituted resi-tion, cells were harvested and extracted for 30 min on dues. Thus, SK6/7LA indicates that the serine and ly-ice using 200 ml of lysis buffer [50 mM Tris – HCl, pH 8.0, sine residues found at positions six and seven in Nef100 mM NaCl, 0.5% NP-40, 2 mM phenylmethylsulfonyl have been mutated to leucine and alanine, respec-fluoride, 100 mM leupeptin, and 2 mg/ml of aprotinin (Boe- tively. The introduced mutations were designed nothringer Mannheim)]. Extracts were clarified by centrifu- only to address the functional significance of Nef resi-gation for 5 min at 14,000 g in a microcentrifuge and then dues proposed to be involved in mediating proteinsubjected to electrophoresis on 12% SDS/polyacrylamide interactions but also to be relatively evenly spacedgels. The resolved proteins were transferred onto nitro- throughout the length of the protein. Specific muta-cellulose membranes (Amersham) and then incubated tions were targeted at the Nef myristoylation signalwith a 1:500 dilution of ascites fluid containing the anti- (G2A), the proposed SH3 sequence (P69L, P72L, VP74/Nef monoclonal antibody EH1 (a gift of James Hoxie) in 75LA and P147L), and the arginine residues at posi-PBS containing 0.1% Tween 20 and 1% nonfat dry milk. tions 105 and 106 (RR105/106LL, RQ106/107LA).After extensive washing, bound antibodies were de- The assay used to measure the efficiency of CD4tected using a horseradish peroxidase conjugated goat downregulation by the various Nef mutants is a tran-anti-mouse antiserum and enhanced chemilumines- sient assay in the human cell line 293T, performedcence (Amersham). essentially as previously described (Goldsmith et al.,
1995). The 293T cells were transfected with plasmidsNef structure visualization
expressing wild-type human CD4, wild-type or mutantHIV-1 Nef proteins, and the cell surface human IL-2RaThe published solution structure of HIV-1 Nef (Grzesiek
et al., 1996) was visualized, and appropriate domains receptor. The IL-2Ra protein is not subject to downreg-
AID VY 8517 / 6a33$$$121 04-11-97 13:09:26 vira AP: Virology

233FUNCTIONAL DOMAINS OF HIV-1 Nef
FIG. 1. Primary sequence of mutants of HIV-1 Nef. The primary sequence and location of mutations introduced into the 206-amino-acid Nefprotein of the NL-43 HIV-1 isolate are indicated. Names of mutants reflect the location and identity of mutated residues. Blocks of conservedresidues in Nef (Shugars et al., 1993) are indicated.
ulation by Nef (Goldsmith et al., 1995) and therefore regulate cell-surface CD4 expression. As shown inFig. 3, these mutants showed widely variable levelsserves as an important internal control for transfection
efficiency. of activity ranging from inactive (e.g., RF184/185LA) tofully active (e.g., HP166/167LA). Nef mutants lackingA representative FACS analysis of a CD4 downregu-
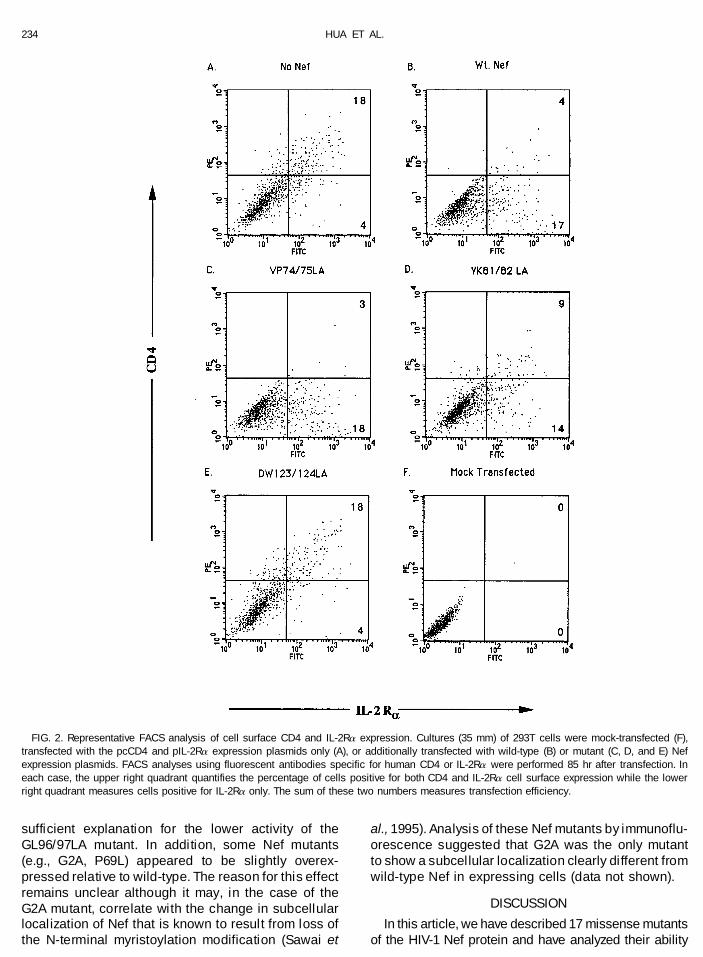
lation assay performed in 293T cells is shown in Fig. a functional myristoylation signal (G2A) or arginine106 (RR105/106LL and RQ106/107LA) showed low but2. Intensity of cell surface staining for IL-2Ra is shown
on the x axis while CD4 expression is analyzed on the detectable levels of CD4 downregulation. In contrast,Nef proteins bearing mutations that affect the pro-y axis. The upper right quadrant therefore gives the
percentage of cells that are both IL-2Ra- and CD4- posed proline-rich tyrosine kinase binding site (P69L,P72L, VP74/75LA, P147L) all displayed close to wild-positive while the lower right quadrant measures the
cells that are IL-2Ra-positive but express little or no type activity in this CD4 downregulation assay. Unfor-tunately, none of the inactive Nef mutants shown inCD4. The sum of these two numbers gives the overall
transfection efficiency. Fig. 3 proved able to inhibit the ability of wild-type Nefprotein to downregulate CD4 expression when ex-In the absence of Nef (Fig. 2A), 18% of the cells were
found to be double-positive while 4% were IL-2Ra-posi- pressed in trans (data not shown).An important question that arises in any mutationaltive only. In contrast, upon cotransfection of a wild-type
Nef expression plasmid (Fig. 2B), the level of double- analysis is whether the stability, and hence the steady-state expression level, of the mutated protein is com-positive cells decreased to 4% of the total while the IL-
2Ra single-positive percentage increased to 17%. Thus, promised. To address this issue, we performed aWestern analysis on 293T cells transfected with thewhile the level of IL-2Ra-positive cells was unchanged
in these two cultures (22% versus 21%), the level of cell- various Nef expression plasmids in order to assesswhether the steady-state level of Nef protein expres-surface CD4-positive cells was dramatically reduced
(18% versus 4%). No positive cells were observed using a sion was significantly affected by any introduced mu-tation. As shown in Fig. 4, all the Nef mutants weremock-transfected culture (Fig. 2F). This figure also shows
results obtained using an essentially fully active Nef mu- expressed at readily detectable levels in transfected293T cells. Two mutants, GL96/97LA and HP166/tant (VP74/75LA, Fig. 2C), a mutant of intermediate activ-
ity (YK81/82LA, Fig. 2D), and an essentially inactive mu- 167LA, were observed to be expressed at a two- tothreefold lower level than wild-type. While the GL96/tant (DW123/124LA, Fig. 2E). In no case was a significant
effect on the level of cell surface expression of the IL- 97LA mutant was indeed found to be compromisedfor efficient CD4 downregulation, the HP166/167 LA2Ra receptor observed.
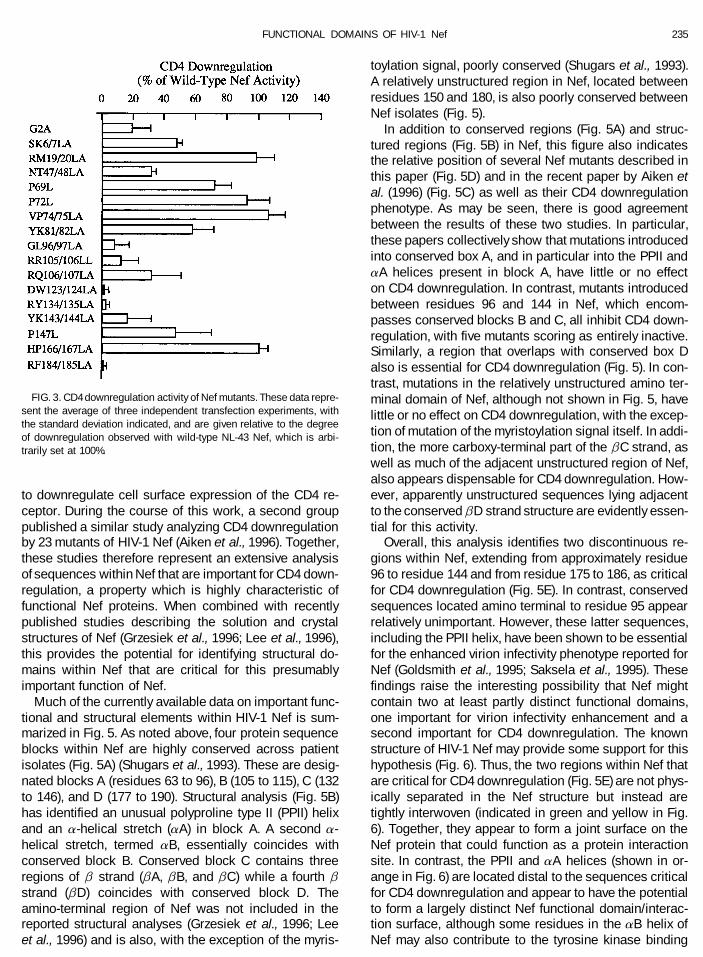
This assay was next used to measure the ability of mutant was essentially fully active (Fig. 3). Therefore,this reduced level of expression is apparently not aeach of the Nef mutants described in Fig. 1 to down-
AID VY 8517 / 6a33$$$121 04-11-97 13:09:26 vira AP: Virology
234 HUA ET AL.
FIG. 2. Representative FACS analysis of cell surface CD4 and IL-2Ra expression. Cultures (35 mm) of 293T cells were mock-transfected (F),transfected with the pcCD4 and pIL-2Ra expression plasmids only (A), or additionally transfected with wild-type (B) or mutant (C, D, and E) Nefexpression plasmids. FACS analyses using fluorescent antibodies specific for human CD4 or IL-2Ra were performed 85 hr after transfection. Ineach case, the upper right quadrant quantifies the percentage of cells positive for both CD4 and IL-2Ra cell surface expression while the lowerright quadrant measures cells positive for IL-2Ra only. The sum of these two numbers measures transfection efficiency.
sufficient explanation for the lower activity of the al., 1995). Analysis of these Nef mutants by immunoflu-orescence suggested that G2A was the only mutantGL96/97LA mutant. In addition, some Nef mutants
(e.g., G2A, P69L) appeared to be slightly overex- to show a subcellular localization clearly different fromwild-type Nef in expressing cells (data not shown).pressed relative to wild-type. The reason for this effect
remains unclear although it may, in the case of theDISCUSSIONG2A mutant, correlate with the change in subcellular
localization of Nef that is known to result from loss of In this article, we have described 17 missense mutantsof the HIV-1 Nef protein and have analyzed their abilitythe N-terminal myristoylation modification (Sawai et
AID VY 8517 / 6a33$$$121 04-11-97 13:09:26 vira AP: Virology
235FUNCTIONAL DOMAINS OF HIV-1 Nef
toylation signal, poorly conserved (Shugars et al., 1993).A relatively unstructured region in Nef, located betweenresidues 150 and 180, is also poorly conserved betweenNef isolates (Fig. 5).
In addition to conserved regions (Fig. 5A) and struc-tured regions (Fig. 5B) in Nef, this figure also indicatesthe relative position of several Nef mutants described inthis paper (Fig. 5D) and in the recent paper by Aiken etal. (1996) (Fig. 5C) as well as their CD4 downregulationphenotype. As may be seen, there is good agreementbetween the results of these two studies. In particular,these papers collectively show that mutations introducedinto conserved box A, and in particular into the PPII andaA helices present in block A, have little or no effecton CD4 downregulation. In contrast, mutants introducedbetween residues 96 and 144 in Nef, which encom-passes conserved blocks B and C, all inhibit CD4 down-regulation, with five mutants scoring as entirely inactive.Similarly, a region that overlaps with conserved box Dalso is essential for CD4 downregulation (Fig. 5). In con-trast, mutations in the relatively unstructured amino ter-
FIG. 3. CD4 downregulation activity of Nef mutants. These data repre- minal domain of Nef, although not shown in Fig. 5, havesent the average of three independent transfection experiments, with little or no effect on CD4 downregulation, with the excep-the standard deviation indicated, and are given relative to the degree
tion of mutation of the myristoylation signal itself. In addi-of downregulation observed with wild-type NL-43 Nef, which is arbi-tion, the more carboxy-terminal part of the bC strand, astrarily set at 100%.well as much of the adjacent unstructured region of Nef,also appears dispensable for CD4 downregulation. How-ever, apparently unstructured sequences lying adjacentto downregulate cell surface expression of the CD4 re-
ceptor. During the course of this work, a second group to the conserved bD strand structure are evidently essen-tial for this activity.published a similar study analyzing CD4 downregulation
by 23 mutants of HIV-1 Nef (Aiken et al., 1996). Together, Overall, this analysis identifies two discontinuous re-gions within Nef, extending from approximately residuethese studies therefore represent an extensive analysis
of sequences within Nef that are important for CD4 down- 96 to residue 144 and from residue 175 to 186, as criticalfor CD4 downregulation (Fig. 5E). In contrast, conservedregulation, a property which is highly characteristic of
functional Nef proteins. When combined with recently sequences located amino terminal to residue 95 appearrelatively unimportant. However, these latter sequences,published studies describing the solution and crystal
structures of Nef (Grzesiek et al., 1996; Lee et al., 1996), including the PPII helix, have been shown to be essentialfor the enhanced virion infectivity phenotype reported forthis provides the potential for identifying structural do-
mains within Nef that are critical for this presumably Nef (Goldsmith et al., 1995; Saksela et al., 1995). Thesefindings raise the interesting possibility that Nef mightimportant function of Nef.
Much of the currently available data on important func- contain two at least partly distinct functional domains,one important for virion infectivity enhancement and ational and structural elements within HIV-1 Nef is sum-
marized in Fig. 5. As noted above, four protein sequence second important for CD4 downregulation. The knownstructure of HIV-1 Nef may provide some support for thisblocks within Nef are highly conserved across patient
isolates (Fig. 5A) (Shugars et al., 1993). These are desig- hypothesis (Fig. 6). Thus, the two regions within Nef thatare critical for CD4 downregulation (Fig. 5E) are not phys-nated blocks A (residues 63 to 96), B (105 to 115), C (132
to 146), and D (177 to 190). Structural analysis (Fig. 5B) ically separated in the Nef structure but instead aretightly interwoven (indicated in green and yellow in Fig.has identified an unusual polyproline type II (PPII) helix
and an a-helical stretch (aA) in block A. A second a- 6). Together, they appear to form a joint surface on theNef protein that could function as a protein interactionhelical stretch, termed aB, essentially coincides with
conserved block B. Conserved block C contains three site. In contrast, the PPII and aA helices (shown in or-ange in Fig. 6) are located distal to the sequences criticalregions of b strand (bA, bB, and bC) while a fourth b
strand (bD) coincides with conserved block D. The for CD4 downregulation and appear to have the potentialto form a largely distinct Nef functional domain/interac-amino-terminal region of Nef was not included in the
reported structural analyses (Grzesiek et al., 1996; Lee tion surface, although some residues in the aB helix ofNef may also contribute to the tyrosine kinase bindinget al., 1996) and is also, with the exception of the myris-
AID VY 8517 / 6a33$$$121 04-11-97 13:09:26 vira AP: Virology
236 HUA ET AL.
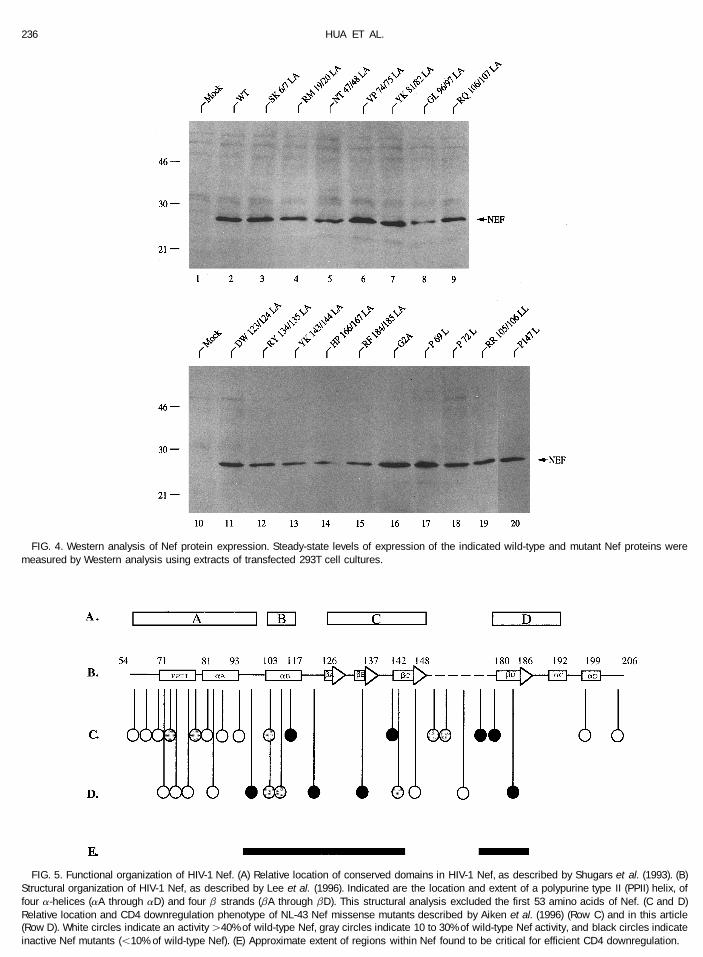
FIG. 4. Western analysis of Nef protein expression. Steady-state levels of expression of the indicated wild-type and mutant Nef proteins weremeasured by Western analysis using extracts of transfected 293T cell cultures.
FIG. 5. Functional organization of HIV-1 Nef. (A) Relative location of conserved domains in HIV-1 Nef, as described by Shugars et al. (1993). (B)Structural organization of HIV-1 Nef, as described by Lee et al. (1996). Indicated are the location and extent of a polypurine type II (PPII) helix, offour a-helices (aA through aD) and four b strands (bA through bD). This structural analysis excluded the first 53 amino acids of Nef. (C and D)Relative location and CD4 downregulation phenotype of NL-43 Nef missense mutants described by Aiken et al. (1996) (Row C) and in this article(Row D). White circles indicate an activity ú40% of wild-type Nef, gray circles indicate 10 to 30% of wild-type Nef activity, and black circles indicateinactive Nef mutants (õ10% of wild-type Nef). (E) Approximate extent of regions within Nef found to be critical for efficient CD4 downregulation.
AID VY 8517 / 6a33$$8517 04-11-97 13:09:26 vira AP: Virology

237FUNCTIONAL DOMAINS OF HIV-1 Nef
FIG. 6. Correlation of Nef structure with function. The indicated structure for the HIV-1 Nef protein was reported by Grzesiek et al. (1996). Theregions extending from residue 96 to 144 (green) and 174 to 185 (yellow), which are critical for efficient CD4 downregulation, are highlighted.Conserved Nef sequence block A, which appears to play no role in CD4 downregulation, is indicated in orange.
motif in the membrane-proximal CD4 cytoplasmic domain. Cell 76,activity displayed by the Nef PPII helix (Lee et al., 1996).853–864.In the future, it will be important to correlate the data
Aiken, C., Krause, L., Chen, Y.-L., and Trono, D. (1996). Mutationalreported here and previously by Aiken et al. (1996) with analysis of HIV-1 Nef: Identification of two mutants that are tempera-more extensive mutational analyses of other Nef pheno- ture-sensitive for CD4 downregulation. Virology 217, 293 – 300.
Aiken, C., and Trono, D. (1995). Nef stimulates human immunodefi-types than have as yet been reported. Based on thisciency virus type 1 proviral DNA synthesis. J. Virol. 69, 5048– 5056.preliminary analysis, it clearly appears possible that Nef
Anderson, S., Shugars, D. C., Swanstrom, R., and Garcia, J. V. (1993).serves at least two distinct functions in the viral life cycleNef from primary isolates of human immunodeficiency virus type 1
and that these functions are, at least in part, performed suppresses surface CD4 expression in human and mouse T cells.by distinct regions in Nef (Fig. 6). Once fully characterized J. Virol. 67, 4923–4931.
Bandres, J. C., and Ratner, L. (1994). Human immunodeficiency virusmutants of these potentially distinct Nef functional do-type 1 Nef protein down-regulates transcription factors NF-kB andmains are available, it should prove possible to measureAP-1 in human T cells in vitro after T-cell receptor stimulation. J.the relative contribution of different Nef activities to theVirol. 68, 3243– 3249.
observed Nef phenotype of efficient viral replication in Baur, A. S., Sawai, E. T., Dazin, P., Fantl, W .J., Cheng-Mayer, C., andinfected primates. Peterlin, B. M. (1994). HIV-1 Nef leads to inhibition of activation of T
cells depending on its intracellular localization. Immunity 1, 373–384.ACKNOWLEDGMENTS
Benson, R. E., Sanfridson, A., Ottinger, J. S., Doyle, C., and Cullen, B. R.The authors thank Ted Benson for technical help, Michael Cook (1993). Downregulation of cell-surface CD4 expression by simian
for assistance with flow cytometry, Sharon Goodwin for secretarial immunodeficiency virus Nef prevents viral super infection. J. Exp.assistance, and James Hoxie for the anti-Nef monoclonal antibody. This Med. 177, 1561– 1566.work was supported by NIH Grant R01 AI35372 from the National Chowers, M. Y., Spina, C. A., Kwoh, T. J., Fitch, N. J. S., Richman, D. D.,Institute of Allergy and Infectious Diseases and by funds from the and Guatelli, J. C. (1994). Optimal infectivity in vitro of human immuno-Howard Hughes Medical Institute. deficiency virus type 1 requires an intact nef gene. J. Virol. 68, 2906–
2914.Cullen, B. R. (1986). Trans-activation of human immunodeficiency virusREFERENCES
occurs via a bimodal mechanism. Cell 46, 973– 982.Aiken, C., Konner, J., Landau, N. R., Lenburg, M. E., and Trono, D. (1994). Cullen, B. R. (1994). The role of Nef in the replication cycle of the human
and simian immunodeficiency viruses. Virology 205, 1– 6.Nef induces CD4 endocytosis: Requirement for a critical dileucine
AID VY 8517 / 6a33$$$121 04-11-97 13:09:26 vira AP: Virology

238 HUA ET AL.
Cullen, B. R., Podlaski, F. J., Peffer, N. J., Hosking, J. B., and Greene, Crystal structure of the conserved core of HIV-1 Nef complexed witha Src family SH3 domain. Cell 85, 931– 942.W. C. (1988). Sequence requirements for ligand binding and cell sur-
Mariani, R., and Skowronski, J. (1993). CD4 down-regulation by nefface expression of the Tac antigen, a human interleukin-2 receptor.alleles isolated from human immunodeficiency virus type 1-infectedJ. Biol. Chem. 263, 4900–4906.individuals. Proc. Natl. Acad. Sci. USA 90, 5549–5553.De Ronde, A., Klaver, B., Keulen, W., Smit, L., and Goudsmit, J. (1992).
Miller, M. D., Warmerdam, M. T., Gaston, I., Greene, W. C., and Fein-Natural HIV-1 Nef accelerates virus replication in primary humanberg, M. B. (1994). The human immunodeficiency virus-1 nef genelymphocytes. Virology 188, 391–395.product: A positive factor for viral infection and replication in primaryDu, Z., Lang, S. M., Sasseville, V. G., Lackner, A. A., Ilyinskii, P. O., Dan-lymphocytes and macrophages. J. Exp. Med. 179, 101– 113.iel, M. D., Jung, J. U., and Desrosiers, R. C. (1995). Identification of a
Rhee, S. S., and Marsh, J. W. (1994). Human immunodeficiency virusnef allele that causes lymphocyte activation and acute disease intype 1 Nef-induced downmodulation of CD4 is due to rapid internal-macaque monkeys. Cell 82, 665– 674.ization and degradation of surface CD4. J. Virol. 68, 5156– 5163.Garcia, J. V., and Miller, A. D. (1991). Serine phosphorylation-indepen-
Ryan-Graham, M. A., and Peden, K. W. C. (1995). Both virus and hostdent downregulation of cell-surface CD4 by nef. Nature 350, 508–components are important for the manifestation of a Nef0 phenotype511.in HIV-1 and HIV-2. Virology 213, 158–168.Goldsmith, M. A., Warmerdam, M. T., Atchison, R. E., Miller, M. D., and
Saksela, K., Cheng, G., and Baltimore, D. (1995). Proline-rich (PxxP)Greene, W. C. (1995). Dissociation of the CD4 downregulation andmotifs in HIV-1 Nef bind to SH3 domains of a subset of Src kinasesviral infectivity enhancement functions of human immunodeficiencyand are required for the enhanced growth of Nef/ viruses but notvirus type 1 Nef. J. Virol. 69, 4112 –4121.for downregulation of CD4. EMBO J. 14, 484–489.Grzesiek, S., Bax, A., Clore, G. M., Gronenborn, A. M., Hu, J.-S., Kaufman,
Sawai, E. T., Baur, A. S., Peterlin, B. M., Levy, J. A., and Cheng-Mayer,J., Palmer, I., Stahl, S. J., and Wingfield, P. T. (1996). The solutionC. (1995). A conserved domain and membrane targeting of Nef fromstructure of HIV-1 Nef reveals an unexpected fold and permits delin-HIV and SIV are required for association with a cellular serine kinase
eation of the binding surface for the SH3 domain of Hck tyrosineactivity. J. Biol. Chem. 270, 15307 –15314.protein kinase. Nature Struct. Biol. 3, 340– 345.
Shugars, D. C., Smith, M. S., Glueck, D. H., Nantermet, P. V., Seillier-Hammes, S. R., Dixon, E. P., Malim, M. H., Cullen, B. R., and Greene, Moiseiwitsch, F., and Swanstrom, R. (1993). Analysis of human imu-
W. C. (1989). Nef protein of human immunodeficiency virus type 1: nodeficiency virus type 1 nef gene sequences present in vivo. J. Virol.Evidence against its role as a transcriptional inhibitor. Proc. Natl. 67, 4639– 4650.Acad. Sci. USA 86, 9549–9553. Spina, C. A., Kwoh, T. J., Chowers, M. Y., Guatelli, J. C., and Richman,
Jamieson, B. D., Aldrovandi, G. M., Planelles, V., Jowett, J. B. M., Gao, D. D. (1994). The importance of nef in the induction of human immu-L., Bloch, L. M., Chen, I. S. Y., and Zack, J. A. (1994). Requirement of nodeficiency virus type 1 replication from primary quiescent CD4human immunodeficiency virus type 1 nef for in vivo replication and lymphocytes. J. Exp. Med. 179, 115–123.pathogenicity. J. Virol. 68, 3478– 3485. Wiskerchen, M., and Cheng-Mayer, C. (1996). HIV-1 Nef association
Kestler, H. W., III, Ringler, D. J., Mori, K., Panicali, D. L., Sehgal, P. K., with cellular serine kinase correlates with enhanced virion infectivityDaniel, M. D., and Desrosiers, R. C. (1991). Importance of the nef and efficient proviral DNA synthesis. Virology 224, 292– 301.gene for maintenance of high virus loads and for development of Zazopoulos, E., and Haseltine, W. A. (1992). Mutational analysis of theAIDS. Cell 65, 651– 662. human immunodeficiency virus type 1 Eli Nef function. Proc. Natl.
Acad. Sci. USA 89, 6634–6638.Lee, C.-H., Saksela, K., Mirza, U. A., Chait, B. T., and Kuriyan, J. (1996).
AID VY 8517 / 6a33$$$121 04-11-97 13:09:26 vira AP: Virology





























