Embed Size (px)
Citation preview

Eur. J. Biochem. 116, 397-402 (1981) <c FEBS 1981
Identification of Homologous Ribosomal Proteins in HeLa Cells and in Tetrahymena pyrzfiormis A Study of Proteins Binding 5-S RNA and Acidic Proteins Released from 40-S Subunits by EDTA
Adelaide J . SVOBODA and Edwin H McCONKEY
Department of Molecular, Cellular, and Developmental Biology, University of Colorado
(Received July 10/December 2, 1980)
Tetruhymena pyriformis 60-S ribosomal subunits treated with EDTA release a 7-S particle containing 5-S RNA and a 36000-M,. protein that is similar to mamnialian 5-S-RNA-binding protein L5 in molecular weight, in two-dimensional acrylamide gel mobility, and in peptide pattern as generated by a simple, one-dimensional acrylamide gel technique. Human and T. pyriformis 40-S ribosomal subunits, treated with buffers lacking magnesium or containing EDTA, release varying amounts of two large acidic proteins. We have identified these released proteins by two-dimensional gel electrophoresis.
Ciliates of the genus Tetrahymena represent one of the most primitive groups of eukaryotes known. Evolutionary relationships, derived from the amino acid sequences of cyto- chrome c, suggest that Tetruhymenu pyriformis diverged from a common eukaryotic ancestor earlier than plants or yeast [I], and the histone IV sequence of T. thermophilu is sub- stantially different from the other eukaryotic histone IV se- quences thus far examined [2]. Recently the efficient isolation of active ribosomes from T. pyriformis and the two-dinien- sional gel identification of its ribosomal proteins have been accomplished [3,4]. Therefore, in an effort to study the relationships between ribosomal proteins froin two very distantly related eukaryotes, we have studied selected human and T. pyriformis ribosomal proteins that share similar proper- ties. Some fundamental similarities are expected between ciliate and mammalian ribosomal components because T. pyriforormis 40-S subunits added to rat liver 60-S subunits can translate poly(U) in vitro [5].
A 7-S particle containing 5-S RNA and protein is released from EDTA-treated 60-S subunits from rat liver [6]. The protein moiety exists in a 1 : 1 ratio with 5-S RNA and has been identified by two-dimensional gel electrophoresis as protein L5 [7] using the standard nomenclature for mam- malian ribosomal proteins [S]. A protein complexed with 5-S RNA is also released from yeast 60-S subunits by EDTA [9,10]. We report here that a similar 5-S-RNA . protein com- plex is released from EDTA-treated 60-S subunits from T. pyrifbrmis. Both a two-dimcnsional gel comparison and a peptide comparison of the T. pyriformis 5-S-RNA-associated protein and HeLa protein L5 are presented.
Eukaryotic 40-S subunits unfold into slower sedimenting particles when they are treated with EDTA [ll] . After re- suspension in buffer containing 2.5 mM EDTA, 40-S subunits
Dt$niiion. A Z ~ O unit, the amount of material in 1 ml of a aolution that has an absorbance of 1 when 260-nm light is passcd through a l-cm- pathlength cell.
Enzvmt7s. RNase A (K 3.1.27.5), RNase TI (EC 3.1.27.3), chymo- trypsin (EC 3.4.21.1); Sfuphylococcus uu~cus VX protease (EC 3.4.21.19), papain (EC 3.4.12.2).
~~
from rabbit reticulocytes release two high and two low- molecular-weight ribosomal proteins that are visible on a one-dimensional sodium dodecyl sulfate gel but have not been identified by two-dimensional gel electrophoresis 1121. We have treated 40-S subunits from HeLa cells and T.pyri- fbrrnis with buffers lacking magnesium or containing 0.01 mM or 1 .O mM EDTA. Using two-dimensional gels that display both acidic and basic ribosomal proteins, we have identified the proteins released from the unfolded small ribosomal sub- units. The unique mRNA-binding protein S1 of Eschrrichiu coli is selectively released from 30-S subunits in buffers similar to those used in our study [13].
MATERIALS AND METHODS
Cell Culture and Ribosomal Subunit Prepurution
The amicronucleate, GL strain of Tetruhymena pyrifi,rmi.v, was grown as described [14] using 30 mg/l Sequestrine (Ciba- Geigy) instead of ferric citrate in the growth medium. Cell harvest conditions and ribosomal subunit isolation were slight modifications of the procedures of Rodrigucs-Pousada and Hayes [4]. Cells from a 1-1 culture at 240000 cells/ml were pelleted and resuspended in 10 ml cold lysis buffer: 0.25 M KCI, 2.5 mM MgC12, 20mM Tris/HCl, pH 7.5, 6 mM 2-mercaptoethanol. All procedures were performed at 4 C unless otherwise noted. Cells were broken in an Omni-mixer, and the lysate was spun at 10000 x g for 30 min. The super- natant was removed and layered over a 2-1111, 1.75 M sucrose cushion (made in lysis buffer) in each of two 10-ml tubes. Polysomes were pelleted at 110000xg for 16 h. The clear polysomal pellet was resuspended to 70 A2h0 units of poly- somesiml with lysis buffer. Puromycin was added to 0.2 mM and GTP to 0.2 mM before the solution was incubatcd at 28 "C for 30 min. The 40-S and 60-S subunits were separated using 5 - 20 '%; linear sucrose gradients made in lysis buffer. HcLa cells were grown and polysomes prepared as dcscribcd [15], and ribosomal subunits were prepared using the puro- mycin procedurc of Blobel and Sabatini 1161.

Release of 7-S Particles The 60-S subunit pellets from both 7: pyrijormis and HeLa
cells were resuspended at 50 A260 units/ml in 10 mM EDTA, were loaded onto separate 5-20 ”/, linear sucrose gradients made in 10 mM KCI and 10 mM Tris/HCl, pH 7.4, and were centrifuged for 37 h at 96000 x g. Fractions containing 7-S particles were pooled and the particles were precipitated overnight at - 20 “C with two volumes of ethanol containing 15 mM MgC12. The remaining cores of the Tpyriformis 60-S subunits, which pelleted in the gradient, were collected and the protein extracted [17].
Identification of the 7-S Particle R N A and Protein
1 A260 unit of T.pyriformis 7-S particles (one-fourth of the total yield from 200 A260 units of T.pyriformis 60-S sub- units) was phenol extracted. The final aqueous phase was ethanol precipitated. Electrophoresis of the RNA was con- ducted according to Mirault and Scherrer [18] using a 2.5- 20% exponential acrylamide gel. The gel was stained with methylene blue.
1 A260 unit of ethanol-precipitated 7-S particles was dis- solved in 0.1 ml 3 M urea, 10 mM EDTA, 10 mM Tris/HCl, pH 7.5. 5 pg RNase A and 200 units RNase TI were added before sample incubation at 37°C for 2 h. Urea was added to 8 M, glycerol to lo%, concentrated acetic acid to 4 % and 2-mercaptoethanol to 5 % in the sample before electrophoresis on a Mets and Bogorad [19] first-dimension gel and a Laemmli [20] second-dimension slab gel containing 15
Peptide Analysis
A culture of T. pyriformis at 25 000 cells/ml was grown for 7 h at 37 “C in a defined basal medium [21] containing one-eighth the specified methionine concentration and 3 pCi/ ml [35S]methionine. HeLa cells at 3 x lo5 cells/ml were grown for 24 h in fresh Eagle’s minimum essential medium con- taining 50 % of the standard methionine concentration, 10 % calf serum, and 2 pCi/ml [35S]methionine. Protein was extracted from the isolated 60-S subunits of each organism [I71 and the specific activity of 35S was equalized in the two samples by the addition of unlabeled 60 - S subunit protein. The samples were run on two-dimensional gels as above. The gels were stained for 15 min in 0.005 Coomassie blue in water before the protein-containing gel spots for peptide analysis were excised and stored at - 20 “C. Thawed gel pieces were loaded directly into sodium dodecyl sulfate gel wells and were treated with either chymotrypsin, Staphylococcus aureus V8 protease, or papain, as described by Cleveland et al. [22]. The gels were stained with Coomassie blue and fluorographed [23] using Kodak XR-5 film.
acrylamide.
40-S Subunit Dialysis and the Identification of Released Proteins
The 40-S subunits obtained from sucrose gradients were pelleted by centrifugation and resuspended in 50 mM KCI, 10 mM Tris/HCl, pH 7.5, at a concentration of 35 A260 units/ml. All procedures were carried out at 4°C. A 0.5-ml aliquot containing 17 A260 units of 40-S subunits was dia- lyzed against bufferA (150mM KCl, 10mM TrisiHCI, pH 7.5), a second aliquot against buffer B (50 mM KCl, 10 mM Tris/HCl, pH 7.5, 0.01 mM EDTA), and a third against buffer C (50 mM KCI, 10 mM Tris/HCl, pH 7.5, 1.0 mM EDTA). The subunits were dialyzed for 7 h against 500-ml volumes of buffer that was changed after 1, 3 and 5 h.
Each sample was loaded onto a 17-ml 5 -40 linear sucrose gradient made in 50 mM KC1, 10 mM Tris/HCI, pH 7.5. The gradients were centrifuged for 16 h at 75000 x g before collection by continuous flow through a Gilford spectro- photometer. The top 3 ml of each gradient were pooled and supplemented with 30 pg bovine serum albumin as carrier protein. After overnight dialysis against 0.1 M acetic acid, the samples were lyophilized. Fractions containing the major peaks within the sucrose gradients (at approximately 25 S in the HeLa gradients and 20 S and 32 S in T.pyriformis gradients) were collected and pelleted by centrifugation. Protein was acetic acid extracted from the pellets [17] and was run on one-dimensional sodium dodecyl sulfate gels [20] using a 10 - 18 ”/, acrylamide exponential gradient in the slab gel. Two-dimensional gel electrophoresis was conducted using a Mets and Bogorad[19] first-dimension gel and a Laemmli [20] second-dimension, 15 ”/, acrylamide slab gel.
RESULTS
Treatment of Tetrahymena pyriformis 6 0 8 ribosomal sub- units with 10 mM EDTA results in the release of an RNA-
Fig. 1, Two-dimensional gel analysis of the protein associated with T. pyriformis 7-S particles. Protein from RNase-treated Tpyriformis 7-S particles supplemented with 20 Fg 60-S-subunit proteins is shown on a Mets and Bogorad [I91 first-dimension gel and a Laemmli [20] second- dimension slab gel in (A). Portions of gels run concomitantly with the gel in (A) are shown for proteins (30 pg/gel) from untreated T.pyriformi.7 60-S subunits (B), proteins remaining with EDTA-treated T.pyriformis 60-S subunits (C) , and proteins from untreated HeLa 6 0 3 subunits (D). The identification of HeLa 60-S-subunit proteins is based on Madjar el al. [24] and that of T. pyriformis 60-S-subunit proteins on Kristiansen and Kriiger [ 3 ] . HeLa protein L3 in (D) should not be confused with T. pyriformis L3 in (B)

390
Fig. 2. Comparison of partial proteolyric products j rom T. pyriformis proteins L40 and L3 Ltith HrLa L5. Two-dimcnsional gel pieces containing 35S-labeled protein were protease digested by the method of Cleveland et al. [22] on sodium dodecyl sulfate gels using the following enzyme concen- trations: S. aureus V8 protease, 0.001 kg in lane e, 0.01 pg in lanes a and c, 0.05 pg in lanes b, d and f; papain, 0.001 pg in lanes g, i and k, 0.01 pg in lanes h, j and I ; chymotrypsin, 1.0 pg in lanes m, o and q, 2.5 pg in lanes n, p and r. Each of lanes s and t contains ''C-labeled protein and 0.2 pg chymotrypsin. (b) The undigested protein; numbers to the left of the figure indicate approximate molecular weight x The patterns shown are autoradiograms. T = Tpyriformis, H = HeLa
containing particle that is seen as a single, dicrete peak at the same relative position in a sucrose gradient as the 7-S particle released from EDTA-treated HeLa 60-S subunits run on a separate gradient. Phenol extraction of the 7-S particle from T. pyriformis and gel electrophoresis of its RNA component reveal that the particle contains 5-S RNA (data not shown).
A sodium dodecyl sulfate gel of the protein extracted from the sucrose gradient pellet containing the cores of the EDTA-treated T. pyriformis large subunits shows a deficiency in one ribosomal protein band (data not shown). A protein identification technique, using an excess of the 5-S RNA binding protein against a faint background of added 6 0 3 subunit protein, facilitates the identification (Fig. 1 A) of the single overloaded protein spot on a two-dimensional gel as protein L3 by the nomenclature of Kristiansen and Kriiger [3]. A comparison of untreated T.pyriformis 60-S subunit protein with the protein remaining on the 7-S-particle-depleted 60-S subunit shows only a small amount of protein L3 remaining with the EDTA-treated subunits (Fig. 1 B and C). Similarly, nearly all of the 5-S RNA is removed from EDTA-treated T.pyriformis 60-S subunits as judged by the fraction of the total 260-nm-light-absorbing material on the gradient re- presented by the 7-S peak (data not shown). This fraction is equivalent for both HeLa and T. pyriformis RNA.
Fig. 1 D shows the upper section of a two-dimensional gel of HeLa 60-S ribosomal subunit protein run at the same time as the T.pyriformis 60-S subunit protein shown in Fig. 1 B. In both organisms the 5-S-RNA-binding protein is the fourth largest protein of the 60-S subunit. By sodium dodecyl sulfate gel analysis we have determined that the molecular weight of T.pyrijbrmis L3 is near 36000 and that of HeLa L5 is near 38 000. Both HeLa L5 and T. pyriformis L3 display a minor protein shadow below the main protein spot (Fig. 1 B and D). This shadow has been reported below rat liver L5 [7] and probably represents a proteolytic derivative of this protein. The susceptibility of HeLa L5 and T. pyriformis L3 to modifi- cation suggests a similarity in the structure of these proteins.
Peptide patterns generated using the Cleveland technique [22] of partial proteolysis are shown for T. pyrzformis proteins
L3 and L40 and HeLa protein L5 in Fig. 2. Proteins L40 (see Fig. 1 B) serves as a control, representing a protein not likely to be homologous to the other two proteins in this com- parison. Two major fragments of approximately 34000 and 24000 molecular weight are common to the Sfuphylococcus aureus V8 protease peptide patterns of T. pyriformis L3 and HeLa L5 (Fig.2, lanes c-f). Similar fragments (with respect to molecular weight) are seen in the papain digest of either protein (Fig. 2, lanes i - 1). The digestion pattern produced from these two proteins by chymotrypsin is characterized by a tight group of large fragments (Fig.2, lanes 0-r). Com- parison of the peptide maps of T. pyriformis L40 with T. pyri- formis L3 and HeLa L5 in Fig.2 demonstrates the dissimi- larity between L40 and the 5-S-RNA-binding proteins in this analysis.
As an additional example of the use of the Cleveland gel technique, peptide patterns produced by chymotrypsin are shown for HeLa protein L3 and T. pyriformis L2 (Fig. 2, lanes s and t). These proteins are the second largest proteins of their respective 60-S subunit (Fig. 1 B and D). There is a similar distribution of doublet fragments (more obviously doublets in the original autoradiogram) in the two peptide patterns, and neither pattern resembles the pattern from T. pyriformis L3 or HeLa L5.
Our second series of experiments concerns similarities in the behavior of certain HeLa and T.pyrjformis small ribo- somal subunits proteins. Fig. 3 shows a one-dimensional gel of the proteins removed from both HeLa and Tpyrifbrmis 40-S subunits after dialysis against buffers A, B or C (see Materials and Methods). Banding patterns of the proteins remaining on the subunits after treatment are also displayed in Fig. 3. Small amounts of low-molecular-weight proteins were removed by these treatments (particularly in buffer A) but were not identified on two-dimensional gels owing to our primary interest in the more extensively removed, larger proteins. The same large proteins are released in varying amounts from HeLa 40-S subunits in each of the treatments (Fig.3, lanes b, d and 0. These large proteins appear to be the same ones seen on a one-dimensional gel of proteins
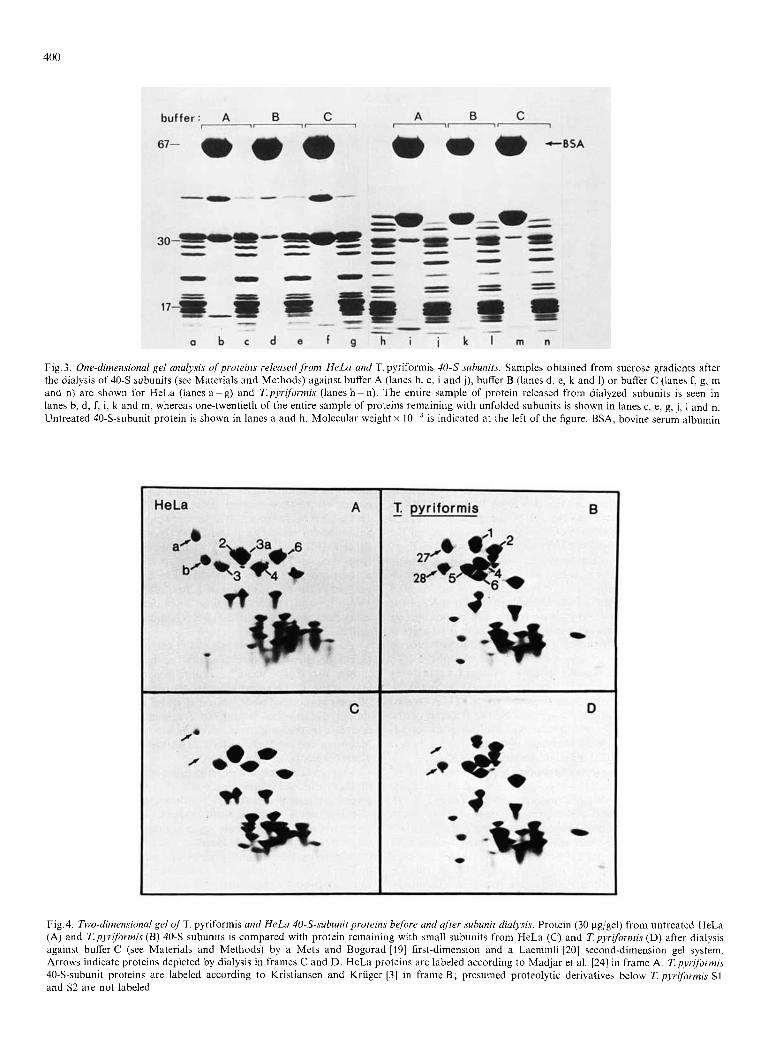
400
Fig. 3. One-dimensional gel analysis of proteins relmsed,from HeLa and T. pyriformis 40-S subunits. Samples obtained from sucrose gradients after the dialysis of 4 0 3 aubunits (see Materials and Methods) against buffer A (lanes b, c, i and j), buffer B (lanes d, e, k and 1) or buffer C (lanes f, g, m and n) are shown for HeLa (lanes a-g) and Tpyriformis (lanes 11-n). The entire sample of protein released from dialyzed subunits is seen in lanes b, d, f, i, k and m, whereas one-twentieth of the entire sample of proteins remaining with unfolded subunits is shown in lanes c, e, g, j, 1 and n. Untreated 40-S-subunit protein is shown in lanes a and h. Molecular weight x is indicated at the left of the figure. BSA, bovine serum albumin
Fig. 4. Two-dimensional gel of T. pyriformis and HeLa 40-S-subunit proteins bcfore and after subunit dialyyis. Protein (30 pg/gel) from untreated HeLa (A) and T.pyriformi.7 (B) 40-S subunits is compared with protein remaining with small subunits from HeLa (C) and T.pyriformis (D) after dialysis against buffer C (see Materials and Methods) by a Mets and Bogorad [I91 first-dimension and a Laemmli [20] second-dimension gel system. Arrows indicate proteins depleted by dialysis in frames C and D. HeLa proteins are labeled according to Madjar et al. [24] in frame A. Tpyr~formis 40-S-subunit proteins are labeled according to Kristiansen and Kriiger [3] in frame B; presumed proteolytic derivatives below T. pyrifi,rmis S1 and S 2 are not labeled

40 1
released from EDTA-treated rabbit reticulocyte 40-S subunits [12j. The identification of these proteins as Sa and Sb (see the nomenclature of Madjar et al. [24])is made by acomparison of two-dimensional gels of proteins associated with the 40-S subunit before and after the removal of specific proteins by dialysis (Fig.4A and C). The removal of Sa is partial in all three treatments, whereas Sb is partially removed in buffers A and B (Fig.3, lanes b and d) but completely removed in buffer C (Fig.4C). Both proteins Sa and Sb are acidic pro- teins, i.e. they are negatively charged near neutral pH [24]. The remaining large proteins of the 40-S subunit are basic proteins. The molecular weights of Sa and Sb appear to be near 43000 and 32000 respectively on a sodium dodecyl sulfate gel (Fig. 3).
The proteins released from T. pyrlformis 40-S subunits during dialysis and the proteins remaining with the treated subunits are also shown in Fig.3. The released proteins are identified as proteins S27 and S28 (Fig.4B and D) by the nomenclature of Kristiansen and Kriiger [3] , who used a two-dimensional gel system comparable to ours. The molec- ular weight of S27 is near 37000 and that of S28 is near 30000 on a sodium dodecyl sulfate gel (Fig.3). The identification of S28 as the smaller protein removed was verified by running the released proteins alone on a two-dimensional gel (data not shown). As in HeLa subunits, the two major proteins released from dialyzed 7: pyr$m?qis small subunits are the two largest acidic protcins. Protein S27 is completely removed in buffers A, B and C (in Fig. 3, note the depletion in the largest protein band in lanes j, 1 and n, compared with lane h). Protein S28 is much more resistant to removal in all three buffers (Fig. 3, lanes i, k and m).
There is a marked decrease in the sedimentation rate of both HeLa and T.pyriformis small subunits in sucrose gra- dients after dialysis against buffer A, B or C compared with the normal 40-S sedimentation value (data not shown). Our observation that the greatest unfolding, i.e. the greatest de- crease in sedimentation value of HeLa small subunits, occurs in buffer C (resulting in 25-S particles) is consistent with the finding that the greatest protein release occurs in this buffer (Fig. 3, lane f). Small subunits from T. pyriformis are equally unfolded by each buffer treatment (to particles of 20 S or 32 S) and release nearly the same amounts of the large acidic proteins in each case (Fig.3, lanes i, k and m).
DISCUSSION
The homology of HeLa ribosomal protein LS and Tctru- hymmn pyriformis protein L3, suggested by their independent existence in a complex with 5-S RNA and by their similar mobilities in gel electrophoresis, is corroborated by a simi- larity between the peptide patterns generated from these two proteins by the Cleveland technique [22]. The physical param- eters governing the probability of proteolytic cleavage events at specific points along a protein in the sodium-dodecyl-sulfate- containing Cleveland gel system have not been described [22]. Although unique peptide patterns are produced from a given protein by each of the proteases tested (Staphylococcus aureus V8 protease cleaves preferentially at Asp and Glu, papain at Arg and Lys, and chymotrypsin at Trp, Tyr, Phe and Leu), similarities seen between the S. aureus V8 protease and papain proteolytic fragment patterns of T. pyriformis and HeLa 5-S- RNA-binding proteins (Fig. 2), as well as markedly similar high-molecular-weight peptide bands seen among the peptide patterns produced by each of the three proteases from mam-
malian ribosomal protein S3a [2S], suggest that protease digestion can be directed by both the structure and the amino acid sequence of the protein substrate. Protein secondary structure present in sodium dodecyl sulfate [26] is sufficient to maintain protease activity in the Cleveland gel system (no 2-mercaptoethanol is used in this system [22]). Substrate protein secondary structure present during the Cleveland procedure may result in the vulnerability of certain protein regions to protease attack. Therefore, similar Cleveland gel peptide patterns from homologous proteins may be duc both to conserved protein structure and to a degree of amino acid sequence homology in protein regions vulnerable to protease attack.
The significant amino acid homology (from 30 7; to 50 yCi) found among partial sequences from the 5-S-RNA-binding proteins of organisms as diverse as Esclierichiu cd i , the archaebacterium Halobacterium cutirubrum, and yeast [I01 is greater than the homology expected among 5-S-RNA-binding proteins of these species when one makes a calculation using the average evolutionary rate of prokaryotic ribosomal pro- teins [27] as a standard evolutionary rate for ribosomal pro- teins of all organisms. These findings suggest that the S-S- RNA-binding proteins are among the most conserved ribo- somal proteins and suggest that clear amino acid sequence homology should exist between 5-S-RNA-binding proteins of even distantly releated eukaryotes.
The release of the two largest acidic proteins from un- folded small ribosomal subunits of both HeLa and T.pyr;- formis suggests the evolutionary relationship of these proteins in the two organisms. These proteins may also be related to E. coli protein S1, an acidic protein that is the largest protein (68000 M,) of the 30-S ribosomal subunit and a protein essen- tial for the recognition of mRNA by the 30-S subunit [2X]. The S1 protein is selectively removed from 30-S subunits unfolded in 1 m M Tris [29] or in buffers with a high ratio of monovalent cations to magnesium cations [I 31. Functional studies are needed to determine whether HeLa Sa and Sb or T.pyriformis S27 and S28 share the mRNA recognition and mRNA unwinding functions of E. coli S1 [30].
There is evidence to suggest the homology of other 40-S- subunit proteins from T.pyriformis and HeLa cells. HeLa protein S 3 a has been crosslinked to the 3’ end of 18-S RNA by a periodate oxidation procedure 1311 and proteins S2X and S5 of T. pyriformis have been crosslinked to 17s R N A in 7: pyr!formi.s 40-S subunits by the periodate procedure [25]. The large and basic HeLa S3a and T.pyriformis S5 proteins (Fig. 4) have similar peptide patterns when analyzed by three separate proteases in the Cleveland procedure 1251 and may represent homologous ribosomal proteins. As protein ho- mologs are identified among many eukaryotic ribosomes, information learned about one eukaryotic translational system can increase our understanding of translational systems in other eukaryotes.
We thank Tom Cech for advice on the culture of T.p.vriji~rmi.s. This research was supported by National Institutes of- Health grant GM21749.
REFERENCES
1. Schwartz, R. M. & Dayhoff, M. 0. (1978) in Atlas uf’ Protein Se- quence and Structure (Dayhoff, M. O., ed.) suppl. 3, pp. 29-44, Natl Biomed. Res. Found., Washington, DC.
2. Glover, C. V. C. & Gorovsky, M. A. (1979) Proc. Nut1 Acad. Sci. USA, 76, 585-589.
3 . Kristiansen, K. & Kruger, A. (1978) Biochirn. Biophys. Actu, 5 2 / , 435-451.

402
4. Rodrigues-Pousada, C. & Hayes, D. (1978) Eur. J. Biochem. 89,
5. Martin, T. E. & Wool, I. G. (1969) J . Mol. Biol. 43, 151 - 161. 6. Blobel, G. (1971) Proc. Natl Acad. Sci. USA, 68, 1881 - 1885. 7. Terao, K., Takahashi, Y. & Ogata, K. (1975) Biochim. Biophys.
Acta, 402, 230-237. 8. McConkey, E. H., Bielka, H., Gordon, J., Lastick, S . M., Lin, A.,
Ogata, K., Reboud, J.-P., Traugh, J. A,, Traut, R. R., Warner, J. R., Welfle, H. & Wool, I. G. (1979) Mol. Gen. Genet. 169, 1-6.
10. Nazar, R. N., Yaguchi, M., Willick, G. E., Rollin, C. F. & Roy, C.
11. Tashiro, Y. & Siekevitz, P. (1965)J. Mol. Biol. 11, 149-165. 12. Freienstein, C. & Blobel, G. (1975) Proc. Natl Acad. Sci. USA, 72,
13. Laughrea, M. & Moore, P. B. (1978) J. Mol. Biol. 121, 41 1-430. 14. Kristiansen, K., Plesner, P. & Kriiger, A. (1978) Eur. J. Biochem.
15. McConkey,E. H. (1974) Proc. NatlAcad. Sci. USA, 71,1379- 1383. 16. Blobel, G. & Sabatini, D. (1971) Proc. Nut1 Acad. Sci. USA, 68,
17. Hardy, S . J. S., Kurland, C. G., Voynow, P. & Mora, G. (1969)
407 - 41 5.
9. Nazar, R. N. (1979) J . Biol. Chem. 254, 7724-7729.
(1979) Eur. J . Biochem. 102, 573-582.
3 392 - 3396.
83, 395-403.
390 - 394.
Biochemislry, 8, 2897 -2905.
18. Mirault, M.-E. & Scherrer, K. (1971) Eur. J. Biochem. 23, 372-386. 19. Mets, L. J. & Bogorad, L. (1974) Anal. Biochem. 57, 200-210. 20. Laemmli, U. K. (1970) Nature (Lond.) 227,680-685. 21. Murti, K. G. & Prescott, D. M. (1970) J . Cell Biol. 47, 460-467. 22. Cleveland, D. W., Fischer, S. G., Kirschner, M. W. & Laemmli,
23. Bonner, W. M. & Laskey, R. A. (1974) Eur. 1. Biochem. 46, 83-88. 24. Madjar, J.-J., Arpin, M., Buisson, M. & Reboud, J.-P. (1979) Mol.
25. Svoboda, Y. J. (1980) Doctoral Dissertation, University Microfilms
26. Mattice, W. L., Riser, J. M. & Clark, D. S. (1976) Biochemistry, 15,
27. Hori, H., Higo, K. & Osawa, S. (1977) J . Mol. Evolut. Y, 191 -201. 28. Van Duin, J . & Van Knippenberg, P. H. (1974) J . Mol. Biol. 84,
29. Tal, M., Aviram, M., Kanarek, A. & Weiss, A. (1972) Biochim.
30. Van Dieijen, G., Van Knippenberg, P. H. & Van Duin, J. (1976)
31. Svoboda, A. J . & McConkey, E. H. (1978) Biochem. Biophys. Res.
U. K.(1977)J. Biol. Chem.252, 3102-1106.
Gen. Genet. 171, 121 - 134.
International, Ann Arbor, Michigan.
4264- 4272.
185 - 195.
Biophys. Acta, 281, 381 - 392.
Eur. J . Biochem. 64, 511 -518.
Commun. 81, 1145-1152.
A. J. Svoboda, 319 D Hayden Street, Sayre, Pennsylvania, USA 18840
E. H. McConkey*, Department of Molecular, Cellular, and Developmental Biology, University of Colorado, Boulder, Colorado, USA 80309 _ _ _ ~
* TO whom reprint requests should be addressed.





























