Embed Size (px)
Citation preview

BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofit publishers,academic institutions, research libraries, and research funders in the common goal of maximizing access to critical research.
Identification of Bloodmeal Sources and Trypanosoma cruziInfection in Triatomine Bugs (Hemiptera: Reduviidae) fromResidential Settings in Texas, the United StatesAuthor(s): Sonia A. Kjos , Paula L. Marcet , Michael J. Yabsley , Uriel Kitron ,Karen F. Snowden , Kathleen S. Logan , John C. Barnes , and Ellen M. DotsonSource: Journal of Medical Entomology, 50(5):1126-1139. 2013.Published By: Entomological Society of AmericaURL: http://www.bioone.org/doi/full/10.1603/ME12242
BioOne (www.bioone.org) is a nonprofit, online aggregation of core research in thebiological, ecological, and environmental sciences. BioOne provides a sustainable onlineplatform for over 170 journals and books published by nonprofit societies, associations,museums, institutions, and presses.
Your use of this PDF, the BioOne Web site, and all posted and associated content indicatesyour acceptance of BioOne’s Terms of Use, available at www.bioone.org/page/terms_of_use.
Usage of BioOne content is strictly limited to personal, educational, and non-commercialuse. Commercial inquiries or rights and permissions requests should be directed to theindividual publisher as copyright holder.

VECTOR/PATHOGEN/HOST INTERACTION, TRANSMISSION
Identification of Bloodmeal Sources and Trypanosoma cruzi Infectionin Triatomine Bugs (Hemiptera: Reduviidae) From Residential
Settings in Texas, the United States
SONIA A. KJOS,1,2,3 PAULA L. MARCET,1 MICHAEL J. YABSLEY,4,5 URIEL KITRON,6
KAREN F. SNOWDEN,7 KATHLEEN S. LOGAN,7 JOHN C. BARNES,8 AND ELLEN M. DOTSON1
J. Med. Entomol. 50(5): 1126Ð1139 (2013); DOI: http://dx.doi.org/10.1603/ME12242
ABSTRACT The hostÐvectorÐparasite interactions in Chagas disease peridomestic transmissioncycles in the United States are not yet well understood. Trypanosoma cruzi (Kinetoplastida: Trypano-somatidae) infection prevalence and bloodmeal sources were determined for adult and immaturetriatomine (Hemiptera: Reduviidae) specimens collected from residential settings in central Texas.Sequenced cytochrome b DNA segments obtained from triatomine digestive tract identiÞed ninevertebrate hosts and one invertebrate host in four triatomine species (Triatoma gerstaeckeri, Triatomaindictiva, Triatoma protracta, and Triatoma sanguisuga). The broad range of wild and domestic hostspecies detected in triatomine specimens collected from residential sites indicates high host diversityand potential movement between the sylvatic and peridomestic settings. Domestic dogs appear to bekey in the maintenance of the peridomestic transmission cycle as both a blood host for the triatominevectors and a potential reservoir for the parasite. The high rate of T. cruzi infection among triatominespecimens that were collected from inside houses, outside houses, and dog kennels (69, 81, and 82%,respectively) suggests a current risk for Chagas disease vector-borne transmission for humans anddomestic animals in residential settings in Texas because of overlap with the sylvatic cycle.
KEY WORDS triatomine, Trypanosoma cruzi, host association, bloodmeal analysis
Chagas disease, caused by the hemoßagellate proto-zoan parasite, Trypanosoma cruzi (Kinetoplastida:Trypanosomatidae), is the most important parasiticdisease on the American continents, accounting for anestimated 8 million human infections, with 56,000 newcases and 12,000 deaths annually occurring in LatinAmerican countries (PAHO 2006, World Health Or-ganization [WHO] 2008). Hematophagous triatominebugs (Hemiptera: Reduviidae: Triatominae) are thebiological arthropod vectors of T. cruzi, transmittingthe parasite in infectious feces deposited on or nearthe host during or shortly after the bug feeds. Theparasite enters the vertebrate host when the feces
contaminate the bite site or mucous membranes. Al-though vector-borne transmission continues to be theprimary route of infection, alternative modes of trans-mission include blood transfusion, organ transplanta-tion, congenital, and oral ingestion of contaminatedfoods or drinks (Coura and Dias 2009). Enzootic trans-mission cycles are endemic across the southern UnitedStates, where 24 wildlife mammalian species havebeen identiÞed as reservoirs, acute and chronic infec-tions have been documented in at least 48 domesticdog breeds, and triatomine vectors have been re-ported from 28 states (Bern et al. 2011).
Of the 141 currently recognized triatomine species,11 are native to the United States and all are consid-ered potential vectors of Chagas disease (SchoÞeldand Galvao 2009). Although 8 of the 11 species havebeen associated with human bites in the United States(Packchanian 1940, Wood 1942, Arnold and Bell 1944,Ryckman 1981, Klotz et al. 2010), human T. cruziinfection because of autochthonous vector-bornetransmission has been rarely reported. Between 1955and 2006, seven human cases were reported in twoadults and Þve young children from the states of Texas(n� 4), CA (n� 1), Tennessee (n� 1), and Louisiana(n � 1) (Woody and Woody 1955, Anonymous 1956,Schifßer et al. 1984, Ochs et al. 1996, Herwaldt et al.2000, Dorn et al. 2007, Kjos et al. 2009). With theexception of one adult case from Louisiana, all in-
1 Division of Parasitic Diseases and Malaria, Entomology Branch,Centers for Disease Control and Prevention, 1600 Clifton Rd., MSG49, Atlanta, GA 30329.
2 Current address: Department of Biology, University of Minnesota,207 Swenson Science Bldg, 1035 Kirby Dr., Duluth, MN, 55812-3004.
3 Corresponding author, e-mail: [email protected] Southeastern Cooperative Wildlife Disease Study, Department of
Population Health, College of Veterinary Medicine, University ofGeorgia, Athens, GA 30602.
5 Daniel B. Warnell School of Forestry and Natural Resources,University of Georgia, Athens, GA 30602.
6 Department of Environmental Studies, Emory University, 400Dowman Dr., E511 Math and Science Center, Atlanta, GA 30322.
7 Department of Veterinary Pathobiology, College of VeterinaryMedicine and Biomedical Sciences, Texas A&M University, CollegeStation, TX 77843-4467.
8 Southwest Texas Veterinary Medical Center, Uvalde, TX 78801.

fected individuals sought medical attention because ofacute symptoms. Since 2007, voluntary testing of do-nated blood forT. cruzi antibodies in the United Stateshas identiÞed �1,600 conÞrmed seropositive dona-tions (AABB 2012). Although most seropositive do-nors are Latin American immigrants who likely ac-quired the infection before relocating to the UnitedStates, a subset of these seropositive donors were U.S.-born with no travel history to endemic areas (Bern etal. 2008). A recent study of conÞrmed seropositive U.S.blood donors identiÞed 16 additional autochthonouscases, all asymptomatic chronic infections, presum-ably acquired by vector-borne transmission in theUnited States (Cantey et al. 2012). The predominantrisk factors included living in a southern state wheretriatomine vectors or infected mammalian reservoirshad been reported and participation in outdoor ac-tivities. Documentation of humanÐtriatomine encoun-ters in the United States predates the discovery of theT. cruzi vector-borne transmission cycle (Leconte1855, Kimball 1894), and these encounters continue tobe reported today primarily in association with aller-gic reactions to triatomine bites (Klotz et al. 2010,Reisenman et al. 2010). The frequency of humanÐtriatomine encounters in the United States is consid-erably lower than in areas with high vector-bornetransmission rates in Latin America, presumably be-cause of the uncommon occurrence of triatomine col-onization of U.S. houses. However, the compellingevidence for vector-borne transmission in newly di-agnosed chronic cases among blood donors suggestsvector-borne transmission may occur more frequentlyin the United States than previously recognized, par-ticularly because acute infection is very mild orasymptomatic in most immunocompetent individuals(Rassi et al. 2010).
Whereas autochthonous human T. cruzi infectionsin the United States are rarely reported, acute andchronic cases among domestic dogs have been docu-mented widely across the southern states, beginningwith Texas in 1972 (Williams et al. 1977, Tomlinson etal. 1981, Barr et al. 1991, Duprey et al. 2006, Kjos et al.2008, Rowland et al. 2010). The disparity in reportedinfection prevalence between humans and dogs in theUnited States is likely because of differences in vectorexposure levels and transmission routes, as well as anincreasing awareness among veterinarians of the po-tential risk of infection for dogs and therefore in-creased testing (Kjos et al. 2008) in contrast to the lackof awareness of human risk among physicians (Stim-pert and Montgomery 2010, Verani et al. 2010). Thepresence of infected domestic animals in peridomesticand domestic settings, particularly dogs, is a signiÞcantcontributor to human infection in certain areas ofSouth America, with dogs serving both as blood hostsfor the triatomine vectors and as parasite reservoirs(Gurtler et al. 1986, 1996, 2007). Dogs housed in prox-imity to two U.S. human autochthonous cases testedpositive for T. cruzi antibodies, providing some evi-dence of a potential role for dogs in human risk in theUnited States (Navin et al. 1985, Herwaldt et al. 2000).Recent studies of T. cruzi infection in triatomine spec-
imens collected in and around houses in the UnitedStates report prevalence rates of 19% (n � 139) inCalifornia (Hwang et al. 2010), 42% (n � 164) inArizona (Reisenman et al. 2010), 53% (n � 137)in Texas (Kjos et al. 2009), and 56% (n � 18) inLouisiana (Dorn et al. 2007), indicating continuedpresence of infected vectors in peridomestic and do-mestic domains across a wide geographic area. Theinteractions of hosts, parasite, and vectors in theUnited States in the peridomestic and domestic do-mains, where triatomine species are not habituallydomiciliated, are not yet well understood.
Bloodmeal analysis has proven effective for eluci-dating vectorÐhost interactions and pathogen trans-mission dynamics in vector-borne disease systems, in-cluding, malaria, Lyme disease (Pichon et al. 2003),leishmaniasis (Haouas et al. 2007), African trypano-somiasis (Muturi et al. 2011), several arboviral diseases(Molaei et al. 2006, Scott et al. 2006), and Chagasdisease (Zarate et al. 1980). Originally based on sero-logical methods, bloodmeal analysis in arthropod vec-tors has largely shifted to DNA-based techniques thatoffer greater accessibility by eliminating the need forhost-derived antisera, and increased speciÞcity in hostidentiÞcation (Mukabana et al. 2002, Kent 2009). Theuse of bloodmeal analysis in triatomine vectors hasbeen informative in determining host preferences andseasonal changes in host selection (Gurtler et al.1997), describing disparities in T. cruzi infection ratesamong triatomine populations (Villalobos et al. 2011),analyzing movement of triatomine vectors betweendifferent habitats (Pizarro and Stevens 2008), andassessing the vector potential of a species (Sandoval etal. 2004). The application of bloodmeal analysis to U.S.triatomine species has only recently been reported ina study of 13 specimens collected using light traps insylvatic settings in California in which rodent, canine,avian, porcine, and human DNA were detected (Ste-vens et al. 2012). The aim of the current study was todetermine hosts that serve as bloodmeal sources andto quantify T. cruzi infection levels of triatomine spe-cies found in residential settings in Texas, particularlythose associated with domestic dogs. IdentiÞcation ofthe sylvatic and domestic hosts used by triatominespecimens collected in this study will also provideinformation on the components of the transmissioncycle in this area as well as triatomine movementbetween the sylvatic and domestic settings. Our over-all objective was to better understand the currentvector-borne transmission risk in this Chagas diseaseendemic region of the United States.
Materials and Methods
Experimental Evaluation of Bloodmeal AnalysisPolymerase Chain Reaction Assays. The feasibility ofusing previously published polymerase chain reaction(PCR) assays for bloodmeal analysis was evaluated intriatomine bugs. Two species of laboratory-rearedbugs, Triatoma dimidiata and Rhodnius prolixus,werefed deÞbrinated blood from two blood hosts, humanand rabbit, and tested at different time intervals using
September 2013 KJOS ET AL.: BLOODMEAL AND T. cruzi ANALYSIS IN U.S. TRIATOMINE BUGS 1127

a mammal-speciÞc assay (Molaei et al. 2006) and avertebrate-speciÞc assay (Boakye et al. 1999) (Table1). Bugs were fed rabbit blood at all previous feedings,with the last meal before the experiment occurringduring the previous instar. Bugs were used for theexperiment 2Ð4 wk after their last molt. Bugs were fedto repletion on a single source of blood using an ar-tiÞcial membrane feeding system (Pennington et al.2004). Seven fourth-instar T. dimidiata and eight Þfth-instar R. prolixus nymphs were fed human blood, andone specimen of each species was tested on days 0, 5,10, 20, 31, 40, 55, and 69. Nine fourth-instarT.dimidiataand nine Þfth-instar R. prolixus nymphs were fed rab-bit blood, and one specimen of each species was testedon days 4, 9, 16, 20, 25, 30, 42, 60, and 71.Specimen Collection and Processing. Specimens
were collected during the summer and fall of 2008 ateight residential sites in Þve counties located in centralTexas (Uvalde County, sites 1Ð4; Zavala County, site5; Comal County, site 6; Bexar County, site 7; GonzalesCounty, site 8). Property sizes were either �3 acresand located in an urban setting (site 6), or were �10acres and located in rural settings (all other sites).Dogs were present at seven residences (sites 1Ð7),three of which had a history of canine Chagas disease(sites 3, 5, and 6). Other animals housed within theperidomestic setting at the sites include cats (sites 1Ð4and 8), cattle (sites 2, 3, and 5), horses (sites 1 and4Ð7), goats (sites 3 and 4), poultry (sites 2,4, and 7),and pigs (site 7). All residents reported previous pres-ence of triatomines on the site, four reported presenceinside houses (sites 3, 4, 7, and 8), and two reportedhuman bites (sites 7 and 8). Manual inspection of eachsite was conducted from 7 July 2008 to 19 August 2008for 2Ð4 d during both daylight and nighttime hours,including inside houses and surrounding areas, dogkennels, outbuildings, wood and rock piles, tree cav-ities, and yard lights. Woodrat nests located at threerural sites in Uvalde County (sites 1, 3, and 4) weredismantled and searched. A universal black light fun-nel trap (BioQuip Products, Rancho Dominguez, CA)was placed in a natural ßyway protected from yard andhouse lights on each property for two to three nights.Captured bugs were placed in separate ventilated con-tainers and cooled on ice until processed. Additionaltriatomine specimens collected by residents of thesites from 23 June 2008 to 20 November 2008 weresubmitted directly to the laboratory.
Location, date, stage, sex (adults only), and condi-tion (live/dead) were recorded at the time of collec-tion. Adult specimens were identiÞed to the specieslevel using the taxonomic key of Lent and Wygodz-insky (1979). Nymphs were identiÞed to the specieslevel using molecular methods described later in thetext. Qualitative nutritional status of adult specimenswas determined by direct observation of the backlitanterior midgut and classiÞed as either starved, or withscarce, good, or large volume of blood (Ceballos et al.2005, VazquezÐProkopec et al. 2006). For nymphs, aqualitative nutritional classiÞcation of either starvedor not starved was assigned based on presence orabsence of abdominal distention. Specimens that were
Tab
le1
.D
escr
ipti
onof
PC
Ras
says
Tar
get
Pri
mer
nam
eSequen
ce(5
�-3�
)P
roduct
bp
Cycl
ing
con
dit
ion
s
Refe
ren
ceD
en
atura
tion
An
neal
ing
Ext
en
sion
No.
cycl
es
Blo
odm
eal
anal
ysi
sV
ert
ebra
tem
tDN
Acytb
BM
1C
CC
CT
CA
GA
AT
GA
TA
TT
TG
TC
CT
CA
358
95�C
,30
s57
�C,50
s72
�C,40
s36
(Boak
ye
et
al.19
99)
BM
2C
CA
TC
CA
AC
AT
CT
CA
GC
AT
GA
TG
AA
AM
amm
alia
nm
tDN
Acytb
Mam
mal
ian
aF
CG
AA
GC
TT
GA
TA
TG
AA
AA
AC
CA
TC
GT
T77
295
�C,30
s55
�C,45
s72
�C,1.
5m
in36
(Mola
ei
et
al.20
06)
Mam
mal
ian
aR
TG
TA
GT
T(A
/G)T
C(A
/T)G
GG
TC
(A/C
/T)C
CT
AA
via
nm
tDN
Acytb
L15
557
GA
CT
GT
GA
CA
AA
AT
CC
C(G
/A/T
/C)
TT
CC
A50
895
�C,30
s57
�C,50
s72
�C,40
s36
(Cic
ero
and
Joh
nso
n20
01)
H16
065
GG
TC
TT
CA
TC
T(C
/T)
(C/T
/A)G
G(T
/C)T
TA
CA
AG
AC
Cri
cket
mtD
NA
CO
IC
1-J-
2183
CA
AC
AT
TT
AT
TT
TG
AT
TT
TT
TG
G72
494
�C,30
s52
�C,30
s72
�C,1
min
30(S
imon
et
al.19
94,
Gra
yet
al.20
08)
TL
2-N
-301
4T
CC
AA
TG
CA
CT
AA
TC
TG
CC
AT
AT
TA
Trypanosomacruzi
dete
ctio
nn
DN
Asa
tellit
ere
peat
TC
Z1
CG
AG
CT
CT
TG
CC
CA
CA
CG
GG
TG
CT
188
94�C
,20
s57
�C,10
s72
�C,30
s35
(Mose
ret
al.19
89)
TC
Z2
CC
TC
CA
AG
CA
GC
GG
AT
AG
TT
CA
GG
Min
icir
cle
kD
NA
Tc1
21A
AA
TA
AT
GT
AC
GG
G(T
/G)
GA
GA
TG
CA
TG
A33
095
�C,30
s58
�C,30
s72
�C,1
min
35(W
inck
er
et
al.19
94)
Tc1
22G
GT
TC
GA
TT
GG
GG
TT
GG
TG
TA
AT
AT
AT
riat
om
ine
speci
es
iden
tiÞ
cati
on
mtD
NAcytb
cytb
7432
FG
GA
CG
(AT
)G
G(A
T)
AT
TT
AT
TA
TG
GA
TC
663
94�C
,30
s47
�C,30
s72
�C,1
min
35(M
on
teir
oet
al.20
03)
cytb
7433
RG
C(A
T)
CC
AA
TT
CA
(AG
)G
TT
A(A
G)T
AA
cytb
-in
tF(C
/T)C
TA
CT
AT
CC
GC
GG
TT
CC
TT
A44
395
�C,30
s51
�C,50
s72
�C,40
s36
this
pap
er
cytb
-in
tRA
TA
CT
AT
TG
CA
AT
TA
CT
CC
TC
CT
A28
SR
NA
D2
D2F
GC
GA
GT
CG
TG
TT
GC
TT
GA
TA
GT
GC
AG
633
94�C
,1
min
50�C
,2
min
72�C
,2
min
25(F
itzp
atri
cket
al.20
08)
D2R
TT
GG
TC
CG
TG
TT
TC
AA
GA
CG
GG
1128 JOURNAL OF MEDICAL ENTOMOLOGY Vol. 50, no. 5

completely desiccated at the time of processing wereexcluded. For specimens collected during Þeld stud-ies, the digestive tract was removed (Garcia and DeAzambuja 1997) using sterile forceps and scalpel, andplaced in 50 �l phosphate-buffered saline. Sampleswere ßash-frozen in an ethanol � dry ice bath andstored on dry ice until transported to the laboratory,where they were stored at �70�C until processed. Forspecimens submitted directly to the laboratory, thelower abdomen was detached with a sterile disposablescalpel and homogenized with a 0.25-inch sterile ce-ramic bead for 20 s using the BIO101/Savant FastPrepFP120 (Qbiogene, Inc., Carlsbad, CA). DNA extrac-tion was performed using the QIAmp DNA Blood MiniKit (QIAGEN, Inc., Valencia, CA), according to themanufacturerÕs instructions.Molecular Analysis.The primer sequences, cycling
parameters, and references for the PCR assays areprovided in Table 1. All PCR products were visu-alized using electrophoresis on 1.5% agarose gels,stained with ethidium bromide. PCR products sub-ject to sequencing were puriÞed using the MilliporeMultiScreenHTS PCR 96-well plate system (Milli-pore Corp., Billerica, MA), and sequenced using theBigDye Terminator Ready Reaction Kit v1.1 andABI 3130xl Genetic Analyzer (Applied Biosystems,Foster City, CA). To minimize the risk of cross-contamination among samples, DNA extraction,PCR setup, and electrophoresis were conducted inseparate dedicated areas of the laboratory.Triatomine Species Identification of Nymphs. Stan-
dard PCR methods were used to amplify fragments ofthe mitochondrial cytochrome b (cytb) gene (Mon-teiro et al. 2003) or D2 section of the nuclear 28Sribosomal RNA gene (Fitzpatrick et al. 2008) (Table1). Sequences that were unreadable using the cytbassay were ampliÞed and sequenced using an internalsegment of the cytb fragment (Table 1). Sequenceswere compared with previously generated sequencesfrom morphologically identiÞed adult specimens todetermine species.T. cruziDetection.TwoDNAtargetswereampliÞed
to determine infection status:T. cruzi-speciÞc TCZ1/2nuclear satellite repeat (Moser et al. 1989) and Tc121/122 minicircle fragment (Wincker et al. 1994) (Table1). A positive result as determined by visualization ofthe expected band size on an agarose gel for bothassays was required for a specimen to receive a pos-itive T. cruzi infection status.Bloodmeal Analysis. Three different cytb targets
were used in three separate PCR assays to identifybloodmeal hosts belonging to mammalian, avian, orvertebrate animal groups. Two of the targets, primersets Mammalian a F/R (Molaei et al. 2006) andL15557/H16065 (Cicero and Johnson 2001) (Table 1),speciÞcally amplify mammal and bird DNA, respec-tively. The third target, primer set BM1/BM2 (Boakyeet al. 1999) (Table 1), ampliÞes all vertebrate DNA,including reptiles and amphibians (Meece et al. 2005).Positive DNA controls were used in each assay asfollows: macaw (avian), dog (mammalian and verte-brate), and ground skink (vertebrate). Sterile water
was used as a negative control in all assays. The prod-ucts of all positive reactions were sequenced and com-pared with DNA nucleotide sequences in NCBIBLAST using the megablast query. Query resultsachieving �95% maximum identity and Expect (E)value �0 were considered matches. Samples matchingÞeld cricket DNA were subjected to a fourth PCRassay targeting a Þeld cricket-speciÞc mitochondrialcytochrome c oxidase subunit one (COI) fragment(Simon et al. 1994, Gray et al. 2008) (Table 1).Statistical Analysis. All statistical tests were per-
formed in SAS 9.2 software (SAS Institute Inc., Cary,NC). Chi-square tests were used to assess differencesin probability of obtaining a bloodmeal identiÞcationby nutritional status (starved vs. not starved) and stateat dissection (live vs. dead). The nutritional status ofadult specimens was converted to either starved or notstarved (with scarce, good, or large volume of blood)to allow for the combination of adults and nymphs inthis analysis. Specimens with unreadable or undeter-mined sequences only were removed from the �2
analysis.
Results
Evaluation of Bloodmeal Analysis PCR Assays. Thelaboratory-based feeding experiments resulted in dif-ferent PCR detection limits for the two tested assaysby both blood source and triatomine species (Table2). The vertebrate assay detected human blood for alonger period postfeeding (up to day 55) comparedwith the mammalian assay (up to day 40), and for anequal number of days in the two triatomine species.
Table 2. Comparison of PCR assays in laboratory-reared tri-atomine bugs fed human or rabbit blood
Bloodhost
Dayspostfeeding
Triatoma dimidiata Rhodnius prolixus
Stage PCRm PCRv Stage PCRm PCRv
Humana 0 N4 � � N5 � �5 N4 � � N5 � �
10 N4 � � N5 � �20 N4 � � N5 � �31 N4 � � Adult � �40 N4 � � Adult � �55 N4 � � Adult � �69 � � � Adult � �
Rabbitb 4 N4 � � N5 � �9 N4 � � N5 � �
16 N4 � � N5 � �20 N4 � � N5 � �25 N4 � � Adult � �30 N4 � � Adult � �42 N4 � � Adult � �60 N5 � � Adult � �71 N5 � � � � �
PCRm, mammalian a F/R; PCRv, vertebrate BM1/BM2.All T. dimidiata and R. prolixus were fed as fourth (N4)- and Þfth
(N5)-instar nymphs, respectively.One live specimen of each species was tested at each time point.a Seven T. dimidiata and 8 R. prolixus nymphs were fed and tested;
four R. prolixus specimens molted before testing.bNineT. dimidiata andR. prolixusnymphs were fed and all but one
R. prolixus specimen survived to be tested; two T. dimidiata and fourR. prolixus specimens molted before testing.
September 2013 KJOS ET AL.: BLOODMEAL AND T. cruzi ANALYSIS IN U.S. TRIATOMINE BUGS 1129

The assays performed the same for specimens fedrabbit blood, with a longer period postfeeding detect-able in T. dimidiata compared withR. prolixus.Rabbitblood was detectable the greatest number of dayspostfeeding (to at least day 71) compared with humanblood (up to day 55).Triatomine Specimen Collections by Site. In total,
153 triatomine specimens were collected and pro-cessed (site 1, n� 50; site 2, n� 2; site 3, n� 10; site4, n� 13; site 5, n� 9; site 6, n� 53; site 7, n� 10; site8, n � 6) (Table 3). Inspection of 59 woodrat nestsyielded 60 specimens of two species (T. protracta,adults, n� 4, nymphs, n� 45; T. gerstaeckeri, nymphs,n� 11) from nine nests at two of the three sites wherewoodrat nests were searched (Table 3). All triatominespecimens were collected by manual search methodsor submitted by residents; none were captured usingthe black light funnel trap. The specimens were pri-marily collected from woodrat nests (n� 60, 39%) anddog kennels (n � 59, 39%), with the remaining spec-imens collected from around (n� 18, 12%) and insidehouses (n � 16, 10%).
Specimens of Þve triatomine species were col-lected, and 78% of specimens tested were infectedwith T. cruzi (Table 3). The most commonly collectedspecies was T. gerstaeckeri (n � 70, 46%). The DNAtarget of the two PCR assays used for triatomine spe-cies identiÞcation did not provide enough sequencedifferentiation between the morphologically identi-Þed T. sanguisuga and T. indictiva adult specimens,and no sequences for T. indictiva were available inGenBank for comparison. Thus we were unable tomake a deÞnite species determination for 21 Þfth-instar nymphs, which could be either T. sanguisuga orT. indictiva. Because the two species are sympatric in
our study area, these specimens were designated as T.sanguisuga/indictiva for the analyses. Sequences gen-erated for species identiÞcation have been submittedto GenBank (KF188579-663).Identification of Bloodmeal Hosts. A bloodmeal
host was identiÞed for 63% (n� 96) of the specimens(Table 4). IdentiÞcation of a bloodmeal host was sig-niÞcantly higher in specimens not starved (83/99,84%) compared with specimens that were starved(12/36, 33%; �2 � 32.30, df � 1, P 0.0001). Identi-Þcation of a bloodmeal host was signiÞcantly higher inspecimens alive at the time of dissection (55/70, 79%)compared with specimens dead at the time of dissec-tion (41/67, 61%; �2 � 4.93, df � 1, P � 0.026).
Nine vertebrate and one invertebrate host specieswere identiÞed in four species of triatomine bugs (Ta-ble 4). Dog DNA was detected in three triatominespecies and was collected from the greatest number ofsites (n � 4). Woodrat was the most frequently de-tected vertebrate species (n � 47), followed by dog(n� 19), cat (n� 6), cow (n� 5), and one each forturkey vulture, black vulture, human, raccoon, andpig. One invertebrate species, Þeld cricket, was de-tected in 15 nymphs using the vertebrate assay. Nineof the 15 samples produced amplicons using the Þeldcricket-speciÞc COI assay. No reptile or amphibianhosts were identiÞed. The greatest variety of hostspecies was identiÞed in T. gerstaeckeri (nine), includ-ing mammal, bird, and insect species. Woodrat was theonly host species identiÞed in T. protracta specimensthat were exclusively collected from woodrat nests.One mixed bloodmeal was identiÞed in an adult T.sanguisuga (dog and turkey vulture) collected in Co-mal County (site 6).
Table 3. Triatomine specimens by species and stage
Triatoma species and stagea No. Trypanosoma cruzi �/tested (%) Bloodmeal ID �/tested (%) Collection siteb Countyc
gerstaeckeri 70 40/57 (70) 33/70 (47) dk,ih,oh,wn B,C,G,U,ZN1 2 0/2 (0) 0/2 (0) wn UN2 5 3/5 (60) 5/5 (100) wn UN3 1 0/1 (0) 0/1 (0) wn UN4 5 1/2 (50) 3/5 (60) dk,wn C,UN5 7 1/3 (33) 2/7 (29) dk,ih,wn C,U,ZAdult 50 35/44 (80) 23/50 (46) dk,ih,oh B,C,G,U,Zindictiva 3 1/1 (100) 2/3 (67) dk C
Adult 3 1/1 (100) 2/3 (67) dk Clecticularia 1 1/1 (100) 0/1 (0) oh U
Adult 1 1/1 (100) 0/1 (0) oh Uprotracta 49 43/49 (88) 40/49 (82) wn U
N1 4 3/4 (75) 4/4 (100) wn UN2 2 2/2 (100) 2/2 (100) wn UN3 2 2/2 (100) 2/2 (100) wn UN4 18 15/18 (83) 14/18 (78) wn UN5 19 17/19 (90) 17/19 (90) wn UAdult 4 4/4 (100) 1/4 (25) wn Usanguisuga 9 4/6 (67) 6/9 (67) dk,ih,oh B,C
Adult 9 4/6 (67) 6/9 (67) dk,ih,oh B,Csanguisuga/indictiva 21 not tested 15/21 (71) dk C
N5 21 not tested 15/21 (71) dk CTotal 153 89/114 (78) 96/153 (63)
aN, nymph, no. refers to instar.b dk, dog kennel; ih, inside house; oh, outside house; wn, woodrat nest.c B, Bexar; C, Comal; G, Gonzales; U, Uvalde; Z, Zavala.
1130 JOURNAL OF MEDICAL ENTOMOLOGY Vol. 50, no. 5

The vertebrate and mammalian assays differed inresults by species detected and frequency of detec-tion. The vertebrate assay detected pig and raccoonDNA in a single specimen each, but the mammalianassay was negative for these same specimens. Themammalian assay detected woodrat DNA in 47 spec-imens, but the vertebrate assay detected woodratDNA in only six of these specimens. The mammalianassay also more frequently detected dog and cat DNAcompared with the vertebrate assay (18 vs. 3, 6 vs. 1,respectively). The vertebrate and avian assays dif-fered in results with the vertebrate assay only detect-ing one of the two bird species detected by the avianassay. Seventeen successfully sequenced PCR prod-ucts (n� 8, vertebrate assay; n� 9, avian assay) failedto match with at least 95% identity to an availablereference sequence in GenBank, so they were con-sidered “undetermined.” Twenty-seven PCR productshad unreadable sequences (n � 25, vertebrate assay;n � 2, mammalian assay). The vertebrate assay am-pliÞed human DNA in 21 specimens, of which onlyone was conÞrmed with the mammalian assay.Whereas seven of these samples produced negativePCR results by the other assays, 13 produced a dif-ferent host species. From these, 10 specimens wereidentiÞed as containing woodrat DNA by the mam-malian assay. These specimens were all nymphs col-lected from woodrat nests, which makes it highly un-likely that they had fed on humans because of theirlimited mobility (wingless in the immature stages) anddistant location from houses. Because of this fact andthe lack of agreement between assays, all human re-sults obtained only with the vertebrate assay weredisregarded. PCR and sequencing results by assay foreach specimen are provided in Appendix A.
Discussion
Bloodmeal hosts were identiÞed in four species ofChagas disease vectors collected in central Texas andincluded seven mammalian, two avian, and one inver-tebrate species. Four of the seven groups of domesticanimals (dogs, cats, cattle, and pigs) known to behoused at the study sites were detected through
bloodmeal analysis. Woodrat DNA was the only hostdetected in specimens collected from woodrat nests.Raccoon and both vulture species identiÞed in spec-imens are commonly found in the study area (Bensonand Arnold 2001, Schmidly 2004). Human DNA wasdetected in one adult specimen, T. gerstaeckeri, col-lected from a bedroom in Bexar County (site 7),where residents had reported recent triatomine bites.
Based on the laboratory feeding experiments, thesuccess of host identiÞcation postfeeding using thevertebrate and mammalian assays ranged from 20 d toat least 71 d, varying with both triatomine and hostspecies. These results were similar to other studies inwhich postfeeding detection limits using a cytbmarkerranged from 30 to 70 d (Bosseno et al. 2006, Mota etal. 2007). Although determination of assay sensitivityin triatomine bugs was beyond the scope of this study,we observed that the vertebrate assay proved to bemuch more sensitive in detecting human DNA in thelaboratory feeding experiments, particularly in spec-imens tested at greater number of days postfeedingand in specimens that had molted. The shorter am-plicon produced by the vertebrate primers comparedwith the mammalian primers may make ampliÞcationmore successful in bloodmeals where the DNA hasdegraded with time and digestive processing or con-tain only trace amounts. These results suggest thatassay sensitivity can vary by both host and triatominespecies, as well as by postfeeding period, possiblybecause of variation in bloodmeal processing timeamong triatomine species and stages and differencesin blood components among host species. Further-more, Þeld specimens with higher nutritional status(evidence of meal in digestive tract) and those aliveat the time of processing were more likely to have abloodmeal host identiÞed using these methods.
Although the PCR assays used in this study provedsuccessful in achieving our goals and have been citedextensively in bloodmeal analysis in other insect vec-tors (Ngo and Kramer 2003, Kilpatrick et al. 2006,Molaei et al. 2006, Richards et al. 2006, Savage et al.2007, Hamer et al. 2008), limitations were noted.Mixed bloodmeals were detected in 1% (1/153) ofspecimens in this study, compared with 6.4% (11/64)
Table 4. Triatomine specimens with DNA sequence bloodmeal identifications
Host identiÞcation No.Trypanosoma cruzi Triatoma speciesa Collection
sitebCountyc
�/tested (%) g i p sd s/i
Bos taurus (cow) 5 3/4 (75) 5 0 0 0 0 dk C,ZCanis lupus familiaris (dog) 19 9/12 (75) 10 2 0 6 1 dk,ih,oh B,C,U,ZCathartes aura (turkey vulture) 1 1/1 (100) 0 0 0 1 0 dk CCoragyps atratus (black vulture) 1 Not tested 1 0 0 0 0 oh CFelis catus (cat) 6 5/5 (100) 6 0 0 0 0 dk,ih,oh C,G,UGryllus texensis/rubens (Þeld cricket) 15 Not tested 1 0 0 0 14 dk CHomo sapiens (human) 1 0/1 (0) 1 0 0 0 0 ih BNeotoma micropus (woodrat) 47 39/47 (83) 7 0 40 0 0 wn UProcyon lotor (raccoon) 1 1/1 (100) 1 0 0 0 0 dk CSus scrofa (pig) 1 1/1 (100) 1 0 0 0 0 dk C
a g, gerstaeckeri; i, indictiva; p, protracta; s, sanguisuga; s/i, sanguisuga or indictiva.b dk, dog kennel; ih, inside house; oh, outside house; wn, woodrat nest.c B, Bexar; C, Comal; G, Gonzales; U, Uvalde; Z, Zavala.dOne specimen was positive for both dog and turkey vulture DNA.
September 2013 KJOS ET AL.: BLOODMEAL AND T. cruzi ANALYSIS IN U.S. TRIATOMINE BUGS 1131

and 40.7% (33/81) in specimens collected from per-idomestic settings in Mexico and Bolivia, respectively,using other nondirect sequencing PCR-based host de-tection methods (Bosseno et al. 2006, Pizarro andStevens 2008). The ability to detect multiple hostspecies in a single specimen was limited by directsequencing of a single product from each assay. Inaddition, 18% (27/153) of specimens had a PCR pos-itive result but unreadable sequences, which could bebecause in part of the presence of DNA from multiplehost species. Cloning and sequencing multiple sam-ples from individual specimens may have increasedour ability to detect multiple hosts (Stevens et al.2012), but were not conducted because of cost andtime limitations. DNA sequencing using the cytb gene,which has extensive taxonomic coverage in GenBank,was determined to be the optimal approach for thisstudy because the host range for the bug species ofinterest is unknown. Even so, 17 samples producedhigh-quality sequences but did not satisfactorilymatch with any published material in GenBank. WithdeÞnitive knowledge of a set of host species (10 spe-cies identiÞed through our current methods), consid-erations for future studies in this geographic area mayinclude adaptations to enhance detection of multiplebloodmeal sources and reduce costs for large samplesizes through the use of alternative DNA-based meth-ods such as terminal restriction fragment length poly-morphism, heteroduplex, or high-resolution meltingassays (Meece et al. 2005, Buitrago et al. 2012, Pena etal. 2012).
The disparity in results from the bloodmeal identi-Þcation assays may be because of differences in sen-sitivity or PCR bias, which has been reported in othergene markers (Polz and Cavanaugh 1998, Acinas et al.2005). Differences between the vertebrate and themammalian assays, including the identiÞcation of hu-man DNA by the vertebrate assay but not the mam-malian assay in the same samples, have been previ-ously reported (Murdock et al. 2010). We questionedthe validity of human positive results obtained by thevertebrate assay because of the lack of agreementbetween the vertebrate and mammalian results andthe fact that most of these human results came fromnymphs captured in woodrat nests, which have a lowdispersal probability, particularly as Þrst and secondinstars. Whereas little data have been reported on theactive dispersal capabilities of triatomine nymphs, Þfth-instar nymphs collected from light traps were estimatedto have traveled up to 42 m from the nearest knowninfested sites in a study in Argentina (VazquezÐProko-pec et al. 2004). In our study, the distance from thewoodrat nests to houses was considerable: 46Ð114 m atone site and 109 m at the other site. The lack of a bloodhost in the nest does not appear to be a reason fordispersal because other specimens collected from thesame nests were positive for woodrat DNA and thesehuman positive specimens by the vertebrate assaywere positive for woodrat DNA by the mammalianassay. In addition, the woodrat nests from which thespecimens were collected had signs of recent activityand woodrats were noted to be present at the sites
(Charles et al. 2013), further corroborating the wood-rat DNA-positive results. If an adult triatomine had fedon a human host and then returned to the woodratnest, it is plausible that the nymphs ingested humanblood while feeding on the engorged adult (Wood andWood 1941, Ryckman 1951, Hays 1965, Alves et al.2011). In this study, no adult specimens collected fromwoodrat nests or any other location at these two siteswere positive for human DNA. Contamination couldbe considered a potential factor, even though all DNAextraction and PCR negative controls were negativeand DNA extractions and PCR setup were performedin separate designated areas. The errant detection ofhuman DNA under carefully controlled laboratoryconditionshasbeenreported inPCR-basedbloodmealstudies in other arthropods (Humair et al. 2007).
The detection of invertebrate DNA (Þeld cricket)by the vertebrate assay indicates the occurrence ofnonspeciÞc binding, which has been reported previ-ously in this marker (Murdock et al. 2010). IdentiÞ-cation of Þeld cricket DNA was conÞrmed using theÞeld cricket-speciÞc COI assay. The lack of COI am-pliÞcation in 6 of the 15 specimens may be because oflower sensitivity of the COI assay compared with thevertebrate cytb assay. Although we were unable todistinguish between the closely related Þeld cricketspecies, Gryllus texensis and Gryllus rubens, based onsequence comparisons in GenBank, the species is mostlikelyG. texensis, the Texas Þeld cricket, based on theknown geographic distribution of the twoGryllus spe-cies (Gray et al. 2008) and the collection site of oursamples. The 15 Þeld cricket-positive specimens wereamong 28 fourth- and Þfth-instar nymphs collectedfrom beneath a concrete slab located in a dog kennel(site 6). The practice of triatomines feeding on thehemolymph of other arthropods (hemolymphagy),including cockroaches and spiders, has been previ-ously reported (Miles et al. 1981, Sandoval et al. 2004).The collection site of the Þeld cricket-positive nymphswas within 3 m of kenneled dogs. One other nymphcollected with the Þeld cricket-positive specimens atthis same site was positive for dog DNA. The stage(Þfth instar) and species (T. sanguisuga and T. indic-tiva) of the dog-positive nymph were also the pre-dominant species and stage of the Þeld cricket-posi-tive nymphs. It appears this triatomine species is ableto readily switch between vertebrate and invertebratehosts in the immature stages, but the circumstancesthat inßuence the selection of hosts are unknown.Because the assays used in this study were designed toidentify vertebrate blood host species, we cannot ruleout the possibility of the presence of other inverte-brate blood host species in these specimens.
The high rate of T. cruzi infection among triatominespecimens collected from inside houses, outsidehouses, and in dog kennels (69, 81, and 82%, respec-tively) suggests a current risk for Chagas disease vec-tor-borne human and domestic animal transmission inresidential settings in central Texas. The methods usedin this study do not allow us to directly associate thebloodmeal host species with the infection source of T.cruzi of a given specimen because we could, presum-
1132 JOURNAL OF MEDICAL ENTOMOLOGY Vol. 50, no. 5

ably, only identify the most recent bloodmeal. Infec-tion could have occurred during a previous meal,which may have been from a different host species. Inaddition, we are not able to conclude host preferencesfor a particular triatomine species because host spe-cies abundance and availability at the time of feedingare unknown for most host species. However, we canconclude that the triatomine species collected in thisstudy willingly feed on the hosts identiÞed, and thehigh T. cruzi prevalence represents a risk for infectionto the susceptible host species. Data on the prevalenceof T. cruzi infection in wild mammals are availablefrom a related study for three of the study sites (1, 3,and 4) plus one additional site in Uvalde County(Charles et al. 2013). Brießy, a highT. cruzi seropreva-lence was detected in woodrats (50/104, 48%), skunks(four-fourths, 100%), raccoons (18/20, 90%), andother rodents (5/28, 18%) sampled at the sites in 2008and 2010. Combined, these results indicate that T.cruzi is currently circulating in both sylvatic and per-idomestic cycles in this area of Texas.
Domestic dogs, particularly those conÞned to out-door kennels, appear to be key in the maintenance ofthe peridomestic transmission cycle in Texas as botha blood host for the triatomine vectors, as determinedin this study, and potentially a reservoir for the par-asite. Previous research indicates that T. cruzi infec-tion among dogs in Texas occurs more frequently inthe sporting and working breeds, which are morelikely to be housed in outdoor kennels (Kjos et al.2008). Two of the four sites where dog DNA wasdetected in triatomine specimens had a known historyof canine Chagas disease. The dogs at these sites (Lab-rador Retriever, German Shorthaired Pointer, andPointer) were used for bird hunting and were housedin conÞned outside kennels. Dogs at the remainingtwo dog DNA-positive sites were housed outside, un-conÞned, indicating that sleeping outdoors (conÞnedor unconÞned) puts dogs at risk for encounters withtriatomine vectors. Although the risk of Chagas dis-ease has been reported to be greater for dogs sleepingoutdoors compared with indoors in a study in urbanMexico (JimenezÐCoello et al. 2010), other factorsincluding triatomine species, T. cruzi prevalence, anddog behavior may impact infection risk among dogsthat sleep outside.
Although both dogs and cats have been implicatedin peridomestic and domestic transmission cycles inSouth America as both a blood host and parasite res-ervoir (Gurtler et al. 1996, 2007), data are lacking onthe role of cats in U.S. transmission cycles. To ourknowledge, this is the Þrst report of U.S. triatominespecies using cats as a bloodmeal host. Additionalresearch on infectivity of dogs and cats to triatominevectors and pathogenicity of strains in triatomine vec-tors collected in residential settings in the UnitedStates would be beneÞcial to further understandingthe dynamics and potential threat of Chagas diseasevector-borne transmission cycles operating in prox-imity to humans.
The broad range of wild and domestic host speciesdetected in triatomine specimens collected from res-
idential sites indicates high host diversity and poten-tial movement between the sylvatic and peridomesticsettings. In addition to the 10 host species describedhere, analysis of additional triatomine specimens fromTexas using the same methods included detection ofchimney swift in T. sanguisuga (Brazos County) andnine-banded armadillo and striped skunk in T. ger-staeckeri (Bexar and Nueces County, respectively)(S.A.K., unpublished data). Flexibility in host selec-tion could complicate efforts to control or eliminatethe bugs from residential property in the study areaand subsequently pose a risk for T. cruzi transmission.At one site in Comal County, 7 of 15 dogs housed onthe property were serologically positive for T. cruzi atthe time of the study. The previous summer, the acutedeath of a 6-yr-old Pointer belonging to the residentswas attributed to T. cruzi infection based on histo-pathologic evidence, making it the Þrst known caninecase in the kennel. It was also the Þrst year the resi-dents noticed the presence of triatomine bugs on theirproperty, although they had resided and housed dogson the property for 13 yr. The residence is located ina rapidly growing urban area north of the city of SanAntonio (population 1,327,407). A comparison of aer-ial photographs taken the Þrst year the residents oc-cupied the residence and the year of the study (13 yrlater) shows pervasive human development in thesurrounding region over that period. The displace-ment of wild vertebrate hosts in the area because ofhabitat changes may have facilitated the movement oftriatomine bugs to the residence, where the kenneleddogs provided an easily accessible alternative blood-meal source. Human alteration of natural habitat andsubsequent migration of triatomine bugs to residentialsettings has been reported forT. protracta in California(Wood and Wood 1964) as well as for several speciesin deforested areas of Central and South America(Romana et al. 2003, Aguilar et al. 2007). The con-Þrmed persistence of T. cruzi and low speciÞcity oftriatomine host selection in this area of Texas, coupledwith the dynamic nature of residential settings, pres-ent a challenge for both predicting and preventing T.cruzi transmission among humans and their compan-ion animals.
Acknowledgments
We thank Jillian Gembler for help with Þeld collectionsand the many Texas residents who submitted specimens orpermitted access to their properties for sampling. We alsothank Gena Lawrence for assistance with bloodmeal PCRassay optimization and the Centers for Disease Control andPrevention (CDC) Biotechnical Core Facility for providingthe oligonucleotides. Funding for S.A.K. was providedthrough an American Society for Microbiology/CDC post-doctoral fellowship. Support for M.J.Y. was provided by Na-tional Institutes of Health (R15 AI067304-02). Use of tradenames is for identiÞcation purposes only and does not implyendorsement of the Centers for Disease Control and Pre-vention, the Public Health Service, or the U.S. Health andHuman Services.
September 2013 KJOS ET AL.: BLOODMEAL AND T. cruzi ANALYSIS IN U.S. TRIATOMINE BUGS 1133

References Cited
AABB. 2012. ChagasÕ biovigilance network, pp. Reportsthrough 04/18/2012. (http://www.aabb.org/programs/biovigilance/Pages/chagas.aspx). Accessed 18 April 2012.
Acinas, S. G., R. Sarma–Rupavtarm, V. Klepac–Ceraj, andM. F. Polz. 2005. PCR-induced sequence artifacts andbias: insights from comparison of two 16S rRNA clonelibraries constructed from the same sample. Appl. Envi-ron. Microbiol. 71: 8966Ð8969.
Aguilar, H. M., F. Abad–Franch, J. C. Dias, A. C. Junqueira,and J. R. Coura. 2007. Chagas disease in the Amazonregion. Mem. Inst. Oswaldo Cruz 102 (Suppl. 1): 47Ð56.
Alves, C. L., R. N. Araujo, N. F. Gontijo, and M. H. Pereira.2011. Importance and physiological effects of hemo-lymphagy in triatomines (Hemiptera: Reduviidae).J. Med. Entomol. 48: 372Ð381.
Anonymous. 1956. Found: two cases of Chagas disease. Tex.Health Bull. 9: 11Ð13.
Arnold, H. L., and D. B. Bell. 1944. Kissing bug bites. J.Allergy Clin. Immunol. 74: 436Ð442.
Barr, S. C., V. A. Dennis, and T. R. Klei. 1991. Serologic andblood culture survey of Trypanosoma cruzi infection infour canine populations of southern Louisiana. Am. J. Vet.Res. 52: 570Ð573.
Benson, K.L.P., and K. A. Arnold. 2001. The Texas breedingbird atlas. Texas A&M University System, Corpus Christi,TX. (http://txtbba.tamu.edu/). Accessed 30 March 2012.
Bern, C., S. P. Montgomery, L. Katz, S. Caglioti, and S. L.Stramer. 2008. Chagas disease and the US blood supply.Curr. Opin. Infect. Dis. 21: 476Ð482.
Bern, C., S. Kjos, M. J. Yabsley, and S. P. Montgomery. 2011.Trypanosoma cruzi and chagasÕ disease in the UnitedStates. Clin. Microbiol. Rev. 24: 655Ð681.
Boakye, D. A., J. Tang, P. Truc, A. Merriweather, and T. R.Unnasch. 1999. IdentiÞcation of bloodmeals in haema-tophagous Diptera by cytochrome B heteroduplex anal-ysis. Med. Vet. Entomol. 13: 282Ð287.
Bosseno, M. F., L. S. Garcia, F. Baunaure, E. M. Gastelum,M. S. Gutierrez, F. L. Kasten, E. Dumonteil, and S. F.Breniere. 2006. IdentiÞcation in triatomine vectors offeeding sources and Trypanosoma cruzi variants by het-eroduplex assay and a multiplex miniexon polymerasechain reaction. Am. J. Trop. Med. Hyg. 74: 303Ð305.
Buitrago, R., S. Depickere, M. F. Bosseno, E. S. Patzi, E.Waleckx, R. Salas, C. Aliaga, and S. F. Breniere. 2012.Combination of cytochrome b heteroduplex-assay andsequencing for identiÞcation of triatomine blood meals.Infect. Genet. Evol. 12: 21Ð27.
Cantey, P. T., S. L. Stramer, R. L. Townsend, H. Kamel, K.Ofafa, C. W. Todd, M. Currier, S. Hand, W. Varnado, E.Dotson, et al. 2012. The United States Trypanosomacruzi infection study: evidence for vector-borne trans-mission of the parasite that causes Chagas disease amongUnited States blood donors. Transfusion 52: 1922Ð1930.
Ceballos, L. A., G.M. Vazquez–Prokopec,M.C. Cecere, P. L.Marcet, and R. E. Gurtler. 2005. Feeding rates, nutri-tional status and ßight dispersal potential of peridomesticpopulations of Triatoma infestans in rural northwesternArgentina. Acta. Trop. 95: 149Ð159.
Charles, R. A., S. Kjos, A. E. Ellis, J. C. Barnes, and M. J.Yabsley. 2013. Southern plains woodrats (Neotoma mi-cropus) from southern Texas are important reservoirs oftwo genotypes of Trypanosoma cruzi and host of a puta-tive novel Trypanosoma species. Vector Borne ZoonoticDis. 13: 22Ð30.
Cicero,C., andN.K. Johnson. 2001. Higher-level phylogenyof new world vireos (aves: vireonidae) based on se-
quences of multiple mitochondrial DNA genes. Mol. Phy-logenet. Evol. 20: 27Ð40.
Coura, J. R., and J. C.Dias. 2009. Epidemiology, control andsurveillance of Chagas disease: 100 years after its discov-ery. Mem. Inst. Oswaldo Cruz 104 (Suppl. 1): 31Ð40.
Dorn, P. L., L. Perniciaro, M. J. Yabsley, D. M. Roellig, G.Balsamo, J. Diaz, and D. Wesson. 2007. Autochthonoustransmission of Trypanosoma cruzi, Louisiana. Emerg. In-fect. Dis. 13: 605Ð607.
Duprey,Z.H., F. J. Steurer, J. A.Rooney,L.V.Kirchhoff, J. E.Jackson, E. D. Rowton, and P. M. Schantz. 2006. Caninevisceral leishmaniasis, United States and Canada, 2000Ð2003. Emerg. Infect. Dis. 12: 440Ð446.
Fitzpatrick, S., M. D. Feliciangeli, M. J. Sanchez–Martin,F. A. Monteiro, andM. A. Miles. 2008. Molecular genet-ics reveal that silvatic Rhodnius prolixus do colonise ruralhouses. PLoS Negl. Trop. Dis. 2: e210.
Garcia, E. S., and P. De Azambuja. 1997. Infection of tri-atomines with Trypanosoma cruzi, pp. 146Ð155. In J. M.Crampton, C. B. Beard, and C. Louis (eds.), MolecularBiology of Disease Vectors. Chapman & Hall, New York,NY.
Gray, D. A., H. Huang, and L. L. Knowles. 2008. Molecularevidence of a peripatric origin for two sympatric speciesof Þeld crickets (Gryllus rubens andG. texensis) revealedfrom coalescent simulations and population genetic tests.Mol. Ecol. 17: 3836Ð3855.
Gurtler,R.E.,N.D.Solard,M.A.Lauricela,A. S.Haedo, S.M.Pietrokovski, A. A. Alberti, and C. Wisnivesky–Colli.1986. Dynamics of transmission of Trypanosoma cruzi ina rural area of Argentina. III. Persistence of T. cruziparasitemia among canine reservoirs in a two-year follow-up. Rev. Inst. Med. Trop. Sao Paulo. 28: 213Ð219.
Gurtler,R.E.,M.C.Cecere,D.P.Vazquez,R.Chuit, and J.E.Cohen. 1996. Host-feeding patterns of domiciliary Tri-atoma infestans (Hemiptera: Reduviidae) in NorthwestArgentina: seasonal and instar variation. J. Med. Entomol.33: 15Ð26.
Gurtler, R. E., J. E. Cohen,M.C.Cecere, andR.Chuit. 1997.Shifting host choices of the vector of Chagas diseaseTriatoma infestans in relation to the availability of hosts inhouses in north-west Argentina. J. Appl. Ecol. 34: 699Ð715.
Gurtler,R.E.,M.C.Cecere,M.A.Lauricella,M.V.Cardinal,U.Kitron, and J. E.Cohen. 2007. Domestic dogs and catsas sources of Trypanosoma cruzi infection in rural north-western Argentina. Parasitology 134: 69Ð82.
Hamer, G. L., U. D. Kitron, J. D. Brawn, S. R. Loss, M. O.Ruiz, T. L. Goldberg, and E. D. Walker. 2008. Culexpipiens (Diptera: Culicidae): a bridge vector of West Nilevirus to humans. J. Med. Entomol. 45: 125Ð128.
Haouas, N., B. Pesson, R. Boudabous, J. P. Dedet, H. Babba,andC. Ravel. 2007. Development of a molecular tool forthe identiÞcation of Leishmania reservoir hosts by bloodmeal analysis in the insect vectors. Am. J. Trop. Med. Hyg.77: 1054Ð1059.
Hays, K. L. 1965. The frequency and magnitude of intras-peciÞc parasitism in Triatoma sanuguisuga (Leconte)(Hemiptera). Ecology 46: 875Ð877.
Herwaldt,B.L.,M. J.Grijalva,A.L.Newsome,C.R.McGhee,M. R. Powell, D. G. Nemec, F. J. Steurer, and M. L.Eberhard. 2000. Use of polymerase chain reaction todiagnose the Þfth reported US case of autochthonoustransmission of Trypanosoma cruzi, in Tennessee, 1998.J. Infect. Dis. 181: 395Ð399.
Humair, P. F., V. Douet, F. Moran Cadenas, L. M. Schouls,I. Van De Pol, and L. Gern. 2007. Molecular identiÞca-
1134 JOURNAL OF MEDICAL ENTOMOLOGY Vol. 50, no. 5

tion of bloodmeal source in Ixodes ricinus ticks using 12SrDNA as a genetic marker. J. Med. Entomol. 44: 869Ð880.
Hwang,W. S., G. Zhang, D. Maslov, and C.Weirauch. 2010.Infection rates of Triatoma protracta (Uhler) withTrypanosoma cruzi in Southern California and molecularidentiÞcation of trypanosomes. Am. J. Trop. Med. Hyg. 83:1020Ð1022.
Jimenez–Coello,M., E. Guzman–Marin, A. Ortega–Pacheco,and K. Y. Acosta–Viana. 2010. Serological survey ofAmerican trypanosomiasis in dogs and their owners froman urban area of Merida Yucatan, Mexico. Transbound.Emerg. Dis. 57: 33Ð36.
Kent, R. J. 2009. Molecular methods for arthropod blood-meal identiÞcation and applications to ecological andvector-borne disease studies. Mol. Ecol. Resour. 9: 4Ð18.
Kilpatrick,A.M., P.Daszak,M. J. Jones, P. P.Marra, andL.D.Kramer. 2006. Host heterogeneity dominates West Nilevirus transmission. Proc. Biol. Sci. 273: 2327Ð2333.
Kimball, B. M. 1894. Conorhinus sanguisuga, its habits andlife history. Kansas Acad. Sci. 14: 128Ð131.
Kjos, S. A., K. F. Snowden, T. M. Craig, B. Lewis, N. Ronald,and J. K. Olson. 2008. Distribution and characterizationof canine Chagas disease in Texas. Vet. Parasitol. 152:249Ð256.
Kjos, S. A., K. F. Snowden, and J. K. Olson. 2009. Biogeog-raphy and Trypanosoma cruzi infection prevalence ofChagas Disease Vectors in Texas, USA. Vector BorneZoonotic Dis. 9: 41Ð50.
Klotz, J. H., P. L. Dorn, J. L. Logan, L. Stevens, J. L. Pinnas,J. O. Schmidt, and S. A. Klotz. 2010. “Kissing bugs”: po-tential disease vectors and cause of anaphylaxis. Clin.Infect. Dis. 50: 1629Ð1634.
Leconte, J. L. 1855. Remarks on two species of AmericanCimex. Proc. Acad. Nat. Sci. Phila. 7: 404.
Lent, H., and P.Wygodzinsky. 1979. Revision of the Triato-minae (Hemiptera: Reduviidae) and their signiÞcance asvectors of Chagas disease. Bull. Am. Mus. Nat. Hist. 163:123Ð520.
Meece, J. K., C. E. Reynolds, P. J. Stockwell, T. A. Jenson, J. E.Christensen, and K. D. Reed. 2005. IdentiÞcation ofmosquito bloodmeal source by terminal restriction frag-ment length polymorphism proÞle analysis of the cyto-chrome B gene. J. Med. Entomol. 42: 657Ð667.
Miles, M. A., A. A. de Souza, and M. Povoa. 1981. ChagasÕdisease in the Amazon basin III. Ecotopes of ten triato-mine bug species (Hemiptera: Reduviidae) from the vi-cinity of Belem, Para State, Brazil. J. Med. Entomol. 18:266Ð278.
Molaei, G., T. G. Andreadis, P.M. Armstrong, J. F. Anderson,and C. R. Vossbrinck. 2006. Host feeding patterns ofCulex mosquitoes and West Nile virus transmission,northeastern United States. Emerg. Infect. Dis. 12: 468Ð474.
Monteiro, F. A., T. V. Barrett, S. Fitzpatrick, C. Cordon–Rosales, D. Feliciangeli, and C. B. Beard. 2003. Molec-ular phylogeography of the Amazonian Chagas diseasevectors Rhodnius prolixus and R. robustus.Mol. Ecol. 12:997Ð1006.
Moser, D. R., L. V. Kirchhoff, and J. E. Donelson. 1989.Detection of Trypanosoma cruzi by DNA ampliÞcationusing the polymerase chain reaction. J. Clin. Microbiol.27: 1477Ð1482.
Mota, J., J. C. Chacon, A. E. Gutierrez–Cabrera, V. Sanchez–Cordero, R. A. Wirtz, R. Ordonez, F. Panzera, and J. M.Ramsey. 2007. IdentiÞcation of blood meal source andinfection with Trypanosoma cruzi of Chagas disease vec-tors using a multiplex cytochrome b polymerase chainreaction assay. Vector Borne Zoonotic Dis. 7: 617Ð627.
Mukabana, W. R., W. Takken, and B. G. Knols. 2002. Anal-ysis of arthropod bloodmeals using molecular geneticmarkers. Trends Parasitol. 18: 505Ð509.
Murdock, C. C., K. J. Olival, and S. L. Perkins. 2010. Mo-lecular identiÞcation of host feeding patterns of snow-melt mosquitoes (Diptera: Culicidae): potential implica-tions for the transmission ecology of Jamestown Canyonvirus. J. Med. Entomol. 47: 226Ð229.
Muturi, C. N., J. O. Ouma, Malele, I. I., R. M. Ngure, J. J.Rutto, K.M.Mithofer, J. Enyaru, andD. K.Masiga. 2011.Tracking the feeding patterns of tsetse ßies (Glossinagenus) by analysis of bloodmeals using mitochondrialcytochromes genes. PLoS One 6: e17284.
Navin, T. R., R. R. Roberto, D. D. Juranek, K. Limpakarn-janarat, E. W. Mortenson, J. R. Clover, R. E. Yescott, C.Taclindo, F. Steurer, and D. Allain. 1985. Human andsylvatic Trypanosoma cruzi infection in California. Am. J.Public Health 75: 366Ð369.
Ngo, K. A., and L. D. Kramer. 2003. IdentiÞcation of mos-quito bloodmeals using polymerase chain reaction (PCR)with order-speciÞc primers. J. Med. Entomol. 40: 215Ð222.
Ochs, D. E., V. S. Hnilica, D. R. Moser, J. H. Smith, and L. V.Kirchhoff. 1996. Postmortem diagnosis of autochtho-nous acute chagasic myocarditis by polymerase chainreaction ampliÞcation of a species-speciÞc DNA se-quence of Trypanosoma cruzi.Am. J. Trop. Med. Hyg. 54:526Ð529.
Packchanian, A. 1940. Natural infection of Triatoma heide-manni with Trypanosoma cruzi in Texas. Public HealthRep. 55: 1300Ð1306.
PAHO. 2006. Estimacio�n cuantitativa de la enfermedad deChagas en las Americas OPS/HDM/CD/425Ð06. Orga-nizacio�n Panamericana de la Salud, Washington, DC.
Pena, V. H., G. J. Fernandez, A. M. Gomez–Palacio, A. M.Mejia–Jaramillo, O. Cantillo, and O. Triana–Chavez.2012. High-resolution melting (HRM) of the cyto-chrome B Gene: a powerful approach to identify blood-meal sources in Chagas disease vectors. PLoS Negl. Trop.Dis. 6: e1530.
Pennington, P.M.,C. B. Beard, and J. Anderson. 2004. Care,maintenance, and handling of infected triatomines, pp.717Ð725. In W. C. Marquardt (ed.), Biology of DiseaseVectors. Elsevier Academic, Burlington, MA.
Pichon,B.,D.Egan,M.Rogers, and J.Gray. 2003. Detectionand identiÞcation of pathogens and host DNA in unfedhost-seeking Ixodes ricinus L. (Acari: Ixodidae). J. Med.Entomol. 40: 723Ð731.
Pizarro, J. C., and L. Stevens. 2008. A new method for fo-rensic DNA analysis of the blood meal in chagas diseasevectors demonstrated using Triatoma infestans fromChuquisaca, Bolivia. PLoS ONE 3: e3585.
Polz, M. F., and C. M. Cavanaugh. 1998. Bias in template-to-product ratios in multitemplate PCR. Appl. Environ.Microbiol. 64: 3724Ð3730.
Rassi, A., Jr., A. Rassi, and J. A. Marin–Neto. 2010. Chagasdisease. Lancet 375: 1388Ð1402.
Reisenman, C. E., G. Lawrence, P. G. Guerenstein, T. Greg-ory, E. Dotson, and J. G. Hildebrand. 2010. Infection ofkissing bugs with Trypanosoma cruzi, Tucson, Arizona,USA. Emerg. Infect. Dis. 16: 400Ð405.
Richards, S. L., L. Ponnusamy, T. R. Unnasch, H. K. Hassan,andC.S.Apperson. 2006. Host-feedingpatternsofAedesalbopictus (Diptera: Culicidae) in relation to availabilityof human and domestic animals in suburban landscapes ofcentral North Carolina. J. Med. Entomol. 43: 543Ð551.
Romana, C. A., D. Brunstein, A. Collin–Delavaud, O. Sousa,and E. Ortega–Barria. 2003. Public policies of develop-
September 2013 KJOS ET AL.: BLOODMEAL AND T. cruzi ANALYSIS IN U.S. TRIATOMINE BUGS 1135

ment in Latin America and ChagasÕ disease. Lancet 362:579.
Rowland,M.E., J.Maloney, S.Cohen,M. J. Yabsley, J.Huang,M. Kranz, A. Green, J. R. Dunn, L. R. Carpenter, T. F.Jones, et al. 2010. Factors associated with Trypanosomacruzi exposure among domestic canines in Tennessee. J.Parasitol. 96: 547Ð551.
Ryckman, R. E. 1951. Recent observations of cannibalism inTriatoma (Hemiptera: Reduviidae). J. Parasitol. 37: 433Ð434.
Ryckman, R. E. 1981. The kissing bug problem in westernNorth America. Bull. Soc. Vector Ecol. 6: 167Ð169.
Sandoval, C.M., R.Duarte, R.Gutierrez, S. RochaDda,V.M.Angulo, L. Esteban, M. Reyes, J. Jurberg, and C. Galvao.2004. Feeding sources and natural infection of Belminusherreri (Hemiptera, Reduviidae, Triatominae) fromdwellings in Cesar, Colombia. Mem. Inst. Oswaldo Cruz99: 137Ð140.
Savage, H. M., D. Aggarwal, C. S. Apperson, C. R. Katholi, E.Gordon, H. K. Hassan, M. Anderson, D. Charnetzky, L.McMillen, E. A. Unnasch, et al. 2007. Host choice andWest Nile virus infection rates in blood-fed mosquitoes,including members of the Culex pipiens complex, fromMemphis and Shelby County, Tennessee, 2002Ð2003.Vector Borne Zoonotic Dis. 7: 365Ð386.
Schiffler, R. J., G. P. Mansur, T. R. Navin, and K. Limpa-karnjanarat. 1984. Indigenous ChagasÕ disease (Ameri-can trypanosomiasis) in California. JAMA 251: 2983Ð2984.
Schmidly, D. J. 2004. The mammals of Texas. University ofTexas Press, Austin, TX.
Schofield, C. J., and C. Galvao. 2009. ClassiÞcation, evolu-tion, and species groups within the Triatominae. ActaTrop. 110: 88Ð100.
Scott, T.W., A. K.Githeko, A. Fleisher, L.C.Harrington, andG. Yan. 2006. DNA proÞling of human blood inanophelines from lowland and highland sites in westernKenya. Am. J. Trop. Med. Hyg. 75: 231Ð237.
Simon, C., F. Frati, A. Beckenbach, B. Crespi, H. Liu, and P.Flook. 1994. Evolution, weighting, and phylogeneticutility of mitochondrial gene sequences and a compila-tion of conserved polymerase chain reaction primers.Ann. Entomol. Soc. Am. 87: 651Ð701.
Stevens, L., P. L. Dorn, J. Hobson, N. M. de la Rua, D. E.Lucero, J. H. Klotz, J. O. Schmidt, and S. A. Klotz. 2012.Vector blood meals and Chagas disease transmissionpotential, United States. Emerg. Infect. Dis. 18: 646Ð649.
Stimpert, K. K., and S. P. Montgomery. 2010. Physicianawareness of Chagas disease, USA. Emerg. Infect. Dis. 16:871Ð872.
Tomlinson, M. J., W. L. Chapman, Jr., W. L. Hanson, andH. S. Gosser. 1981. Occurrence of antibody to Trypano-soma cruzi in dogs in the southeastern United States.Am. J. Vet. Res. 42: 1444Ð1446.
Vazquez–Prokopec, G. M., L. A. Ceballos, U. Kitron, andR. E. Gurtler. 2004. Active dispersal of natural popu-lations of Triatoma infestans (Hemiptera: Reduviidae)in rural northwestern Argentina. J. Med. Entomol. 41:614Ð621.
Vazquez–Prokopec, G.M., L. A. Ceballos, P. L.Marcet,M.C.Cecere, M. V. Cardinal, U. Kitron, and R. E. Gurtler.2006. Seasonal variations in active dispersal of naturalpopulations of Triatoma infestans in rural north-westernArgentina. Med. Vet. Entomol. 20: 273Ð279.
Verani, J. R., S. P.Montgomery, J. Schulkin, B.Anderson, andJ. L. Jones. 2010. Survey of obstetrician-gynecologists inthe United States about Chagas disease. Am. J. Trop. Med.Hyg. 83: 891Ð895.
Villalobos, G., F.Martinez–Hernandez, P. de la Torre, J. P.Laclette, and B. Espinoza. 2011. Entomological indi-ces, feeding sources, and molecular identiÞcation ofTriatoma phyllosoma (Hemiptera: Reduviidae) one ofthe main vectors of Chagas disease in the Istmo deTehuantepec, Oaxaca, Mexico. Am. J. Trop. Med. Hyg.85: 490Ð497.
(WHO)WorldHealthOrganization. 2008. The global bur-den of disease: 2004 update. World Health Organization,Geneva, Switzerland.
Williams, G. D., L. G. Adams, R. G. Yaeger, R. K. McGrath,W. K. Read, and W. R. Bilderback. 1977. Naturally oc-curring trypanosomiasis (ChagasÕ disease) in dogs. J. Am.Vet. Med. Assoc. 171: 171Ð177.
Wincker, P., C. Britto, J. B. Pereira, M. A. Cardoso, W.Oelemann, and C. M. Morel. 1994. Use of a simpliÞedpolymerase chain reaction procedure to detect Trypano-soma cruzi in blood samples from chronic chagasic pa-tients in a rural endemic area. Am. J. Trop. Med. Hyg. 51:771Ð777.
Wood, S. F. 1942. Observations on vectors of ChagasÕ dis-ease in the United States. I. California. Bull. South Calif.Acad. Sci. 41: 61Ð69.
Wood, F. D., and S. F. Wood. 1941. Present knowledge ofthe distribution ofTrypanosoma cruzi in reservoir animalsand vectors. Am. J. Trop. Med. 21: 335Ð345.
Wood, S. F., andF.D.Wood. 1964. Nocturnal aggregationand invasion of homes in southern California by insectvectors of ChagasÕ disease. J. Econ. Entomol. 57: 775Ð776.
Woody, N. C., and H. B. Woody. 1955. American Trypano-somiasis (ChagasÕ disease): Þrst indigenous case in theUnited States. JAMA 159: 676Ð677.
Zarate, L. G., R. J. Zarate, C. H. Tempelis, and R. S. Gold-smith. 1980. The biology and behavior of Triatoma bar-beri (Hemiptera: Reduviidae) in Mexico. I. Blood mealsources and infection with Trypanosoma cruzi. J. Med.Entomol. 17: 103Ð116.
Received 23 October 2012; accepted 11 July 2013.
1136 JOURNAL OF MEDICAL ENTOMOLOGY Vol. 50, no. 5
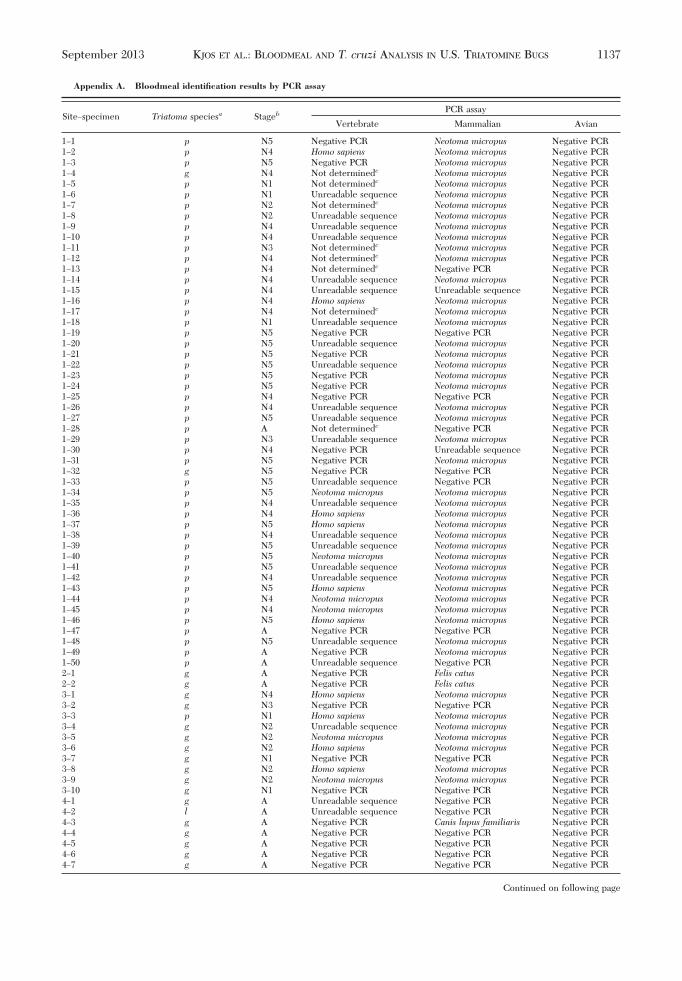
Appendix A. Bloodmeal identification results by PCR assay
SiteÐspecimen Triatoma speciesa StagebPCR assay
Vertebrate Mammalian Avian
1Ð1 p N5 Negative PCR Neotoma micropus Negative PCR1Ð2 p N4 Homo sapiens Neotoma micropus Negative PCR1Ð3 p N5 Negative PCR Neotoma micropus Negative PCR1Ð4 g N4 Not determinedc Neotoma micropus Negative PCR1Ð5 p N1 Not determinedc Neotoma micropus Negative PCR1Ð6 p N1 Unreadable sequence Neotoma micropus Negative PCR1Ð7 p N2 Not determinedc Neotoma micropus Negative PCR1Ð8 p N2 Unreadable sequence Neotoma micropus Negative PCR1Ð9 p N4 Unreadable sequence Neotoma micropus Negative PCR1Ð10 p N4 Unreadable sequence Neotoma micropus Negative PCR1Ð11 p N3 Not determinedc Neotoma micropus Negative PCR1Ð12 p N4 Not determinedc Neotoma micropus Negative PCR1Ð13 p N4 Not determinedc Negative PCR Negative PCR1Ð14 p N4 Unreadable sequence Neotoma micropus Negative PCR1Ð15 p N4 Unreadable sequence Unreadable sequence Negative PCR1Ð16 p N4 Homo sapiens Neotoma micropus Negative PCR1Ð17 p N4 Not determinedc Neotoma micropus Negative PCR1Ð18 p N1 Unreadable sequence Neotoma micropus Negative PCR1Ð19 p N5 Negative PCR Negative PCR Negative PCR1Ð20 p N5 Unreadable sequence Neotoma micropus Negative PCR1Ð21 p N5 Negative PCR Neotoma micropus Negative PCR1Ð22 p N5 Unreadable sequence Neotoma micropus Negative PCR1Ð23 p N5 Negative PCR Neotoma micropus Negative PCR1Ð24 p N5 Negative PCR Neotoma micropus Negative PCR1Ð25 p N4 Negative PCR Negative PCR Negative PCR1Ð26 p N4 Unreadable sequence Neotoma micropus Negative PCR1Ð27 p N5 Unreadable sequence Neotoma micropus Negative PCR1Ð28 p A Not determinedc Negative PCR Negative PCR1Ð29 p N3 Unreadable sequence Neotoma micropus Negative PCR1Ð30 p N4 Negative PCR Unreadable sequence Negative PCR1Ð31 p N5 Negative PCR Neotoma micropus Negative PCR1Ð32 g N5 Negative PCR Negative PCR Negative PCR1Ð33 p N5 Unreadable sequence Negative PCR Negative PCR1Ð34 p N5 Neotoma micropus Neotoma micropus Negative PCR1Ð35 p N4 Unreadable sequence Neotoma micropus Negative PCR1Ð36 p N4 Homo sapiens Neotoma micropus Negative PCR1Ð37 p N5 Homo sapiens Neotoma micropus Negative PCR1Ð38 p N4 Unreadable sequence Neotoma micropus Negative PCR1Ð39 p N5 Unreadable sequence Neotoma micropus Negative PCR1Ð40 p N5 Neotoma micropus Neotoma micropus Negative PCR1Ð41 p N5 Unreadable sequence Neotoma micropus Negative PCR1Ð42 p N4 Unreadable sequence Neotoma micropus Negative PCR1Ð43 p N5 Homo sapiens Neotoma micropus Negative PCR1Ð44 p N4 Neotoma micropus Neotoma micropus Negative PCR1Ð45 p N4 Neotoma micropus Neotoma micropus Negative PCR1Ð46 p N5 Homo sapiens Neotoma micropus Negative PCR1Ð47 p A Negative PCR Negative PCR Negative PCR1Ð48 p N5 Unreadable sequence Neotoma micropus Negative PCR1Ð49 p A Negative PCR Neotoma micropus Negative PCR1Ð50 p A Unreadable sequence Negative PCR Negative PCR2Ð1 g A Negative PCR Felis catus Negative PCR2Ð2 g A Negative PCR Felis catus Negative PCR3Ð1 g N4 Homo sapiens Neotoma micropus Negative PCR3Ð2 g N3 Negative PCR Negative PCR Negative PCR3Ð3 p N1 Homo sapiens Neotoma micropus Negative PCR3Ð4 g N2 Unreadable sequence Neotoma micropus Negative PCR3Ð5 g N2 Neotoma micropus Neotoma micropus Negative PCR3Ð6 g N2 Homo sapiens Neotoma micropus Negative PCR3Ð7 g N1 Negative PCR Negative PCR Negative PCR3Ð8 g N2 Homo sapiens Neotoma micropus Negative PCR3Ð9 g N2 Neotoma micropus Neotoma micropus Negative PCR3Ð10 g N1 Negative PCR Negative PCR Negative PCR4Ð1 g A Unreadable sequence Negative PCR Negative PCR4Ð2 l A Unreadable sequence Negative PCR Negative PCR4Ð3 g A Negative PCR Canis lupus familiaris Negative PCR4Ð4 g A Negative PCR Negative PCR Negative PCR4Ð5 g A Negative PCR Negative PCR Negative PCR4Ð6 g A Negative PCR Negative PCR Negative PCR4Ð7 g A Negative PCR Negative PCR Negative PCR
Continued on following page
September 2013 KJOS ET AL.: BLOODMEAL AND T. cruzi ANALYSIS IN U.S. TRIATOMINE BUGS 1137
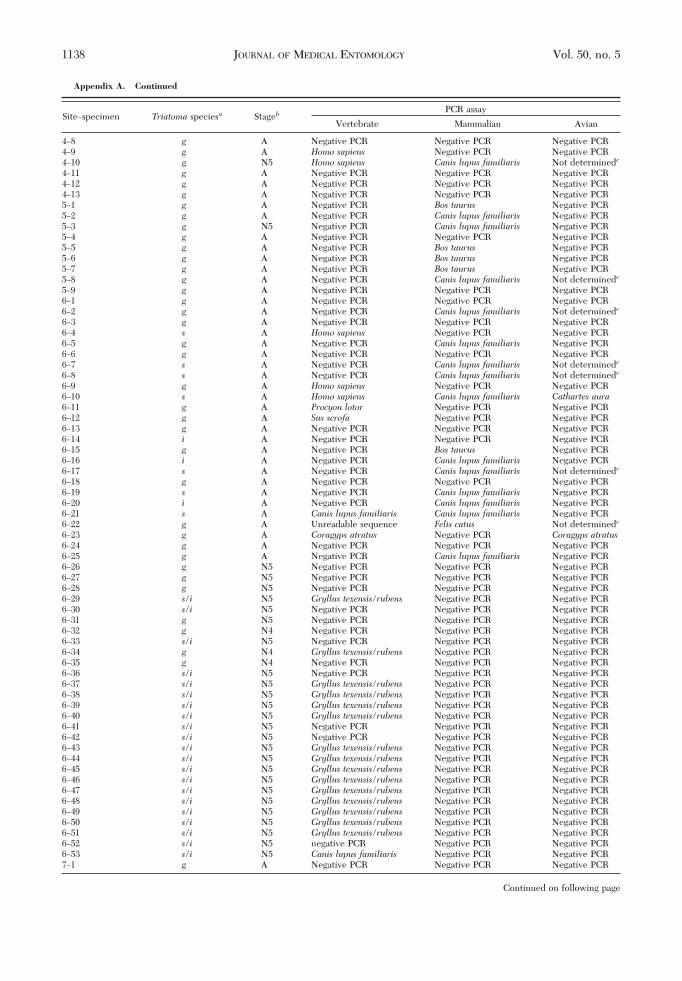
Appendix A. Continued
SiteÐspecimen Triatoma speciesa StagebPCR assay
Vertebrate Mammalian Avian
4Ð8 g A Negative PCR Negative PCR Negative PCR4Ð9 g A Homo sapiens Negative PCR Negative PCR4Ð10 g N5 Homo sapiens Canis lupus familiaris Not determinedc
4Ð11 g A Negative PCR Negative PCR Negative PCR4Ð12 g A Negative PCR Negative PCR Negative PCR4Ð13 g A Negative PCR Negative PCR Negative PCR5Ð1 g A Negative PCR Bos taurus Negative PCR5Ð2 g A Negative PCR Canis lupus familiaris Negative PCR5Ð3 g N5 Negative PCR Canis lupus familiaris Negative PCR5Ð4 g A Negative PCR Negative PCR Negative PCR5Ð5 g A Negative PCR Bos taurus Negative PCR5Ð6 g A Negative PCR Bos taurus Negative PCR5Ð7 g A Negative PCR Bos taurus Negative PCR5Ð8 g A Negative PCR Canis lupus familiaris Not determinedc
5Ð9 g A Negative PCR Negative PCR Negative PCR6Ð1 g A Negative PCR Negative PCR Negative PCR6Ð2 g A Negative PCR Canis lupus familiaris Not determinedc
6Ð3 g A Negative PCR Negative PCR Negative PCR6Ð4 s A Homo sapiens Negative PCR Negative PCR6Ð5 g A Negative PCR Canis lupus familiaris Negative PCR6Ð6 g A Negative PCR Negative PCR Negative PCR6Ð7 s A Negative PCR Canis lupus familiaris Not determinedc
6Ð8 s A Negative PCR Canis lupus familiaris Not determinedc
6Ð9 g A Homo sapiens Negative PCR Negative PCR6Ð10 s A Homo sapiens Canis lupus familiaris Cathartes aura6Ð11 g A Procyon lotor Negative PCR Negative PCR6Ð12 g A Sus scrofa Negative PCR Negative PCR6Ð13 g A Negative PCR Negative PCR Negative PCR6Ð14 i A Negative PCR Negative PCR Negative PCR6Ð15 g A Negative PCR Bos taurus Negative PCR6Ð16 i A Negative PCR Canis lupus familiaris Negative PCR6Ð17 s A Negative PCR Canis lupus familiaris Not determinedc
6Ð18 g A Negative PCR Negative PCR Negative PCR6Ð19 s A Negative PCR Canis lupus familiaris Negative PCR6Ð20 i A Negative PCR Canis lupus familiaris Negative PCR6Ð21 s A Canis lupus familiaris Canis lupus familiaris Negative PCR6Ð22 g A Unreadable sequence Felis catus Not determinedc
6Ð23 g A Coragyps atratus Negative PCR Coragyps atratus6Ð24 g A Negative PCR Negative PCR Negative PCR6Ð25 g A Negative PCR Canis lupus familiaris Negative PCR6Ð26 g N5 Negative PCR Negative PCR Negative PCR6Ð27 g N5 Negative PCR Negative PCR Negative PCR6Ð28 g N5 Negative PCR Negative PCR Negative PCR6Ð29 s/i N5 Gryllus texensis/rubens Negative PCR Negative PCR6Ð30 s/i N5 Negative PCR Negative PCR Negative PCR6Ð31 g N5 Negative PCR Negative PCR Negative PCR6Ð32 g N4 Negative PCR Negative PCR Negative PCR6Ð33 s/i N5 Negative PCR Negative PCR Negative PCR6Ð34 g N4 Gryllus texensis/rubens Negative PCR Negative PCR6Ð35 g N4 Negative PCR Negative PCR Negative PCR6Ð36 s/i N5 Negative PCR Negative PCR Negative PCR6Ð37 s/i N5 Gryllus texensis/rubens Negative PCR Negative PCR6Ð38 s/i N5 Gryllus texensis/rubens Negative PCR Negative PCR6Ð39 s/i N5 Gryllus texensis/rubens Negative PCR Negative PCR6Ð40 s/i N5 Gryllus texensis/rubens Negative PCR Negative PCR6Ð41 s/i N5 Negative PCR Negative PCR Negative PCR6Ð42 s/i N5 Negative PCR Negative PCR Negative PCR6Ð43 s/i N5 Gryllus texensis/rubens Negative PCR Negative PCR6Ð44 s/i N5 Gryllus texensis/rubens Negative PCR Negative PCR6Ð45 s/i N5 Gryllus texensis/rubens Negative PCR Negative PCR6Ð46 s/i N5 Gryllus texensis/rubens Negative PCR Negative PCR6Ð47 s/i N5 Gryllus texensis/rubens Negative PCR Negative PCR6Ð48 s/i N5 Gryllus texensis/rubens Negative PCR Negative PCR6Ð49 s/i N5 Gryllus texensis/rubens Negative PCR Negative PCR6Ð50 s/i N5 Gryllus texensis/rubens Negative PCR Negative PCR6Ð51 s/i N5 Gryllus texensis/rubens Negative PCR Negative PCR6Ð52 s/i N5 negative PCR Negative PCR Negative PCR6Ð53 s/i N5 Canis lupus familiaris Negative PCR Negative PCR7Ð1 g A Negative PCR Negative PCR Negative PCR
Continued on following page
1138 JOURNAL OF MEDICAL ENTOMOLOGY Vol. 50, no. 5
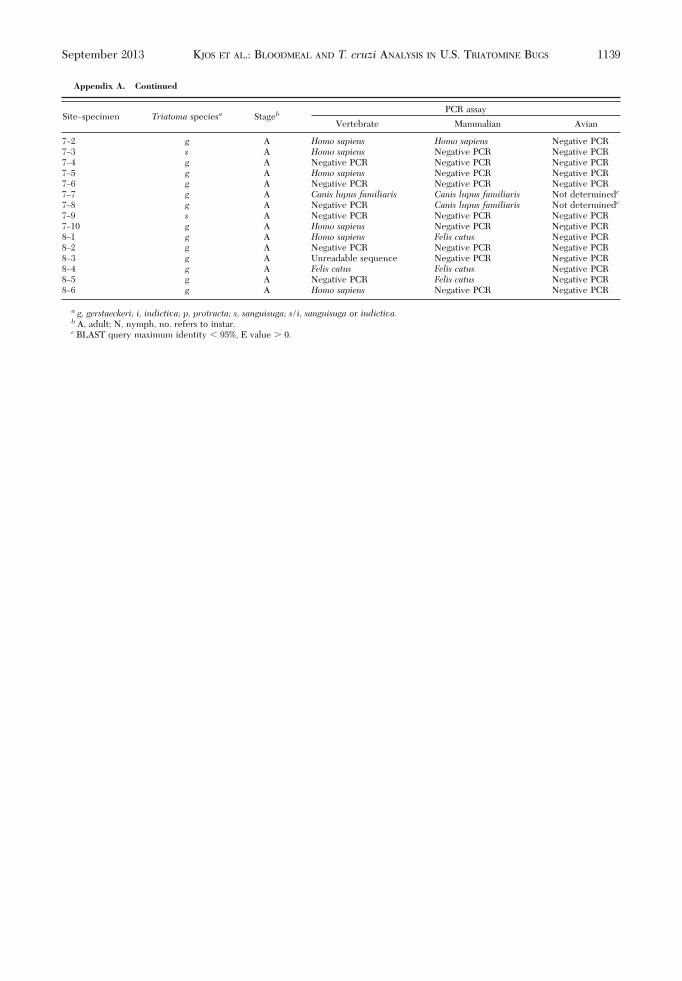
Appendix A. Continued
SiteÐspecimen Triatoma speciesa StagebPCR assay
Vertebrate Mammalian Avian
7Ð2 g A Homo sapiens Homo sapiens Negative PCR7Ð3 s A Homo sapiens Negative PCR Negative PCR7Ð4 g A Negative PCR Negative PCR Negative PCR7Ð5 g A Homo sapiens Negative PCR Negative PCR7Ð6 g A Negative PCR Negative PCR Negative PCR7Ð7 g A Canis lupus familiaris Canis lupus familiaris Not determinedc
7Ð8 g A Negative PCR Canis lupus familiaris Not determinedc
7Ð9 s A Negative PCR Negative PCR Negative PCR7Ð10 g A Homo sapiens Negative PCR Negative PCR8Ð1 g A Homo sapiens Felis catus Negative PCR8Ð2 g A Negative PCR Negative PCR Negative PCR8Ð3 g A Unreadable sequence Negative PCR Negative PCR8Ð4 g A Felis catus Felis catus Negative PCR8Ð5 g A Negative PCR Felis catus Negative PCR8Ð6 g A Homo sapiens Negative PCR Negative PCR
a g, gerstaeckeri; i, indictiva; p, protracta; s, sanguisuga; s/i, sanguisuga or indictiva.b A, adult; N, nymph, no. refers to instar.c BLAST query maximum identity 95%, E value � 0.
September 2013 KJOS ET AL.: BLOODMEAL AND T. cruzi ANALYSIS IN U.S. TRIATOMINE BUGS 1139





























