Embed Size (px)
Citation preview

Eur. J. Biochem. 90,463-469 (1978)
Identification and Characterization of Ribosomal Proteins Phosphorylated in Vaccinia-Virus-Infected HeLa Cells Margret KAERLEIN and Ivan HORAK
Institut fur Virologie und Immunbiologie der Universitat Wurzburg
(Received June 5,1978)
Two-dimensional analysis of 32P-labelled ribosomal proteins revealed three proteins which are phosphorylated in vaccinia-virus-infected HeLa cells. All three proteins belong to the 40-S ribosomal subunits and were identified as S2, S6 and S16. The ribosomal protein S6 is phosphorylated also in uninfected HeLa cells. Phosphoserine was detected in all three proteins, phosphothreonine only in the protein S2. Phosphorylation of these ribosomal proteins in infected cells is dependent on the multiplicity of the viral infection and increases during the first six hours of infection. All three proteins are also phosphorylated in virus-infected cells treated with cycloheximide and in cells infected with ultraviolet-irradiated virus. This suggests that the phosphorylation reaction involves a vaccinia virion-associated protein kinase.
Vaccinia-virus-infected cells have been chosen as a model system in which to investigate changes in the host cell translational apparatus resulting from virus infection. Both the switch-off of host cell protein synthesis and the temporal regulation of 'early' and 'late' vaccinia mRNA translation in infected cells suggest some degree of translational regulation (for review, see [l, 21).
Recently, we have reported that ribosomal pro- teins of HeLa cells are modified after infection with vaccinia virus [3]. The phosphorylation of the ribo- somal protein S6, which is also phosphorylated in un- infected cells, is enhanced in infected cells, and two additional proteins, P2 and P3, become specifically phosphorylated in the infected cells. The enhanced phosphorylation of the ribosomal protein S6 has also been found in cells infected with adenovirus [4] and poliovirus (unpublished results).
The aim of the present study is to identify the ribosomal phosphoproteins P2 and P3 and to further characterize the phosphorylation of ribosomal pro- teins in vaccinia-infected HeLa cells.
MATERIALS AND METHODS
Buffers
Buffer A : 20 mM Tris-HC1 pH 7.5, 100 mM KC1, 5 mM MgC12, 5 mM 2-mercaptoethanol. Buffer B: 20 mM Tris-HC1 pH 7.5, 500 mM KC1, 3 mM MgC12, 5 mM 2-mercaptoethanol.
Cells and Virus
HeLa S3 cells were grown in suspension as de- scribed [5]. Vaccinia WR virus (stock was kindly provided by Dr B. Moss) was propagated on L cells and purified by the procedure of Joklik [6].
Ultraviolet Irradiation and Heat Inactivation of Vaccinia Virus
Purified virus was irradiated with ultraviolet light as described [7]. Heat inactivation of the virus was carried out at 60 "C for 60 min. The residual infectivity of the virus preparations was checked by a plaque assay on mouse L cells.
Labelling Conditions
Vaccinia-virus-infected and mock-infected HeLa cells were suspended after 15 min of virus adsorption in phosphate-free Eagle's medium (Gibco) which was supplemented with 5 % of dialyzed horse serum, and 50 pCi/ml of carrier-free (32P)orthophosphate were added immediately. After a period of incubation (see legends to figures) in suspension at 37 "C, the cells were cooled, centrifuged and washed twice with cold phosphate-buffered saline. All subsequent operations were carried out between 0 and 4 T.
Isolation of Ribosomal Particles
HeLa cells were homogenized in buffer A con- taining 0.5 % nonionic detergent NP-40 (Shell Co.).

464 Ribosomal Proteins Phosphorylated in Vaccinia-Virus-Infected Cells
The ribosomal subunits were separated by sucrose gradient centrifugation in buffer B [ 5 ] .
Extraction of Ribosomal Pvotciii.\
Ribosomal proteins were extracted, dialyzed and lyophilized according to the procedure of Sherton and Wool [8].
Two-Dimensional Polyacrylamide Gel Electrophoresis
Ribosomal proteins were separated by a slightly modified procedure of Kaltschmidt and Wittmann [9], as described [ 5 ] . The acrylamide concentration was 5 % in the first dimension and 18 in the second dimension. The gels were stained with amido black or Coomassie blue, The destained gels were incubated 30 min in 7.5 trichloroacetic acid at 90 "C and washed overnight in 5 % acetic acid to remove any residual contamination. This treatment did not remove any radioactivity spots from radioautographed gels.
Dodt~cylsulfate/Polyucrylamide Gel Electrophoresis
Dodecylsulfate slab-gel electrophoresis was per- formed according to Laemmli [ 101. The electrophoresis of separated protein spots from two-dimensional gels on dodecylsulfate/polyacrylamide gels, 'third-dimen- sion', was described earlier [5 ] .
Determination of Phosphoamino Acids Fig. 1. Two-dime17sionirl unui,vsis of 40-S ribosomal subunit proteins from vacciriia-viru.P-iizf~cled HeLQ cclls. Ribosomal Droteins from
stained protein spots were cut of the gels, dried in vacuo and then hydrolyzed in 1 ml of 2 M or 6 M HCl at 110 "C for 6 h in vacuo. The hydroly-
3zP-labelled, virus-infected cells were prepared and analyzed as described in Methods. Labelling time was 2.5 h. (A) Coomassie- blue-stained gel, (B) autoradiograph
sates were analyzed for 32P incorporated into phospho- serine and phosphothreonine on columns of Dowex 5OW-X8 as described [ l l ] .
Nomenclature of Ribosomal Proteins
Ribosomal proteins from HeLa cells were num- bered according to the system of Sherton and Wool [8] as described earlier [15].
Radioactivity Determination
Stained gels were covered with Saran wrap and exposed to X-ray film (Kodak X-omat) between two plates of plexiglass for 3 - 14 days at 4 "C. Radioac- tivity in individual ribosomal proteins cut out of the gels was determined as described [12]. RESULTS
Localization of the Ribosomal Phosphoproteins
The analysis of the 80-S ribosomes in our previous study [3] did not permit us to determine whether the P3 phosphoprotein is associated with the small or the large ribosomal subunit. For this reason, 40-S and 60-S subunits from 32P-labelled vaccinia-virus- infected HeLa cells were separated and their proteins analyzed by two-dimensional polyacrylamide gel elec- trophoresis and autoradiography (Fig. 1 ). This anal-
Labelling of Ribosomal Proteins in vitro
Protein extracted from 40-S ribosomal subunits was labelled by reductive alkylation with ['4C]formal- dehyde [I31 and analyzed as described [12].
Protein Dete~minution
This was done by Lowry's method [14].
M. Kaerlein and I. Horak 465
800. + -
A - 0
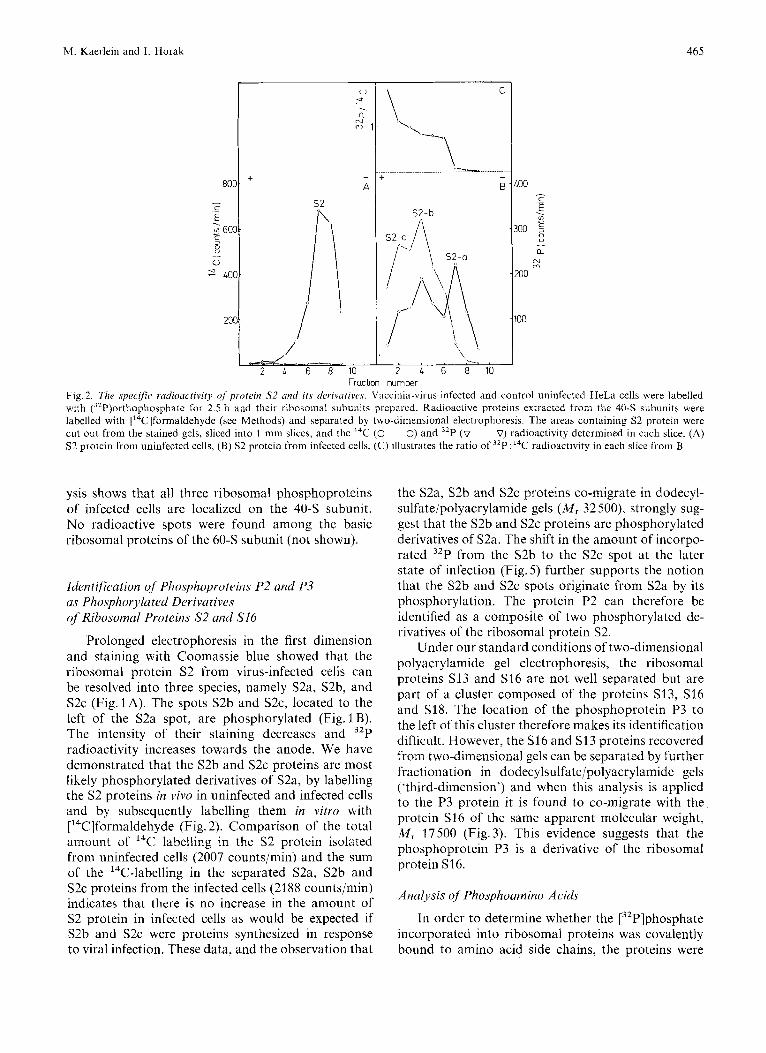
Fraction number Fig. 2. The specific radioactivity of protein S2 und its derivurives. Vaccinia-vii-us-infected and control uninfected HeLa cells were labelled with (32P)orthophosphate for 2.5 h and their ribosomal subunits prepared. Radioactive proteins extracted from the 40-S subunits were labelled with ['4C]formaldehyde (see Methods) and separated by two-dimensional electrophoresis. The areas containing S2 protein were cut out from the stained gels, sliced into 1 -mm slices, and the I4C (0-0) and 32P (v- ~ --V) radioactivity determined in each slice. (A) S2 protein Urom uninfected cells, (B) S2 protein from infected cells, (C) illustrates the ratio of 3LP:14C radioactivity in each slice tiom B
ysis shows that all three ribosomal phosphoproteins of infected cells are localized on the 40-S subunit. No radioactive spots were found among the basic ribosomal proteins of the 60-S subunit (not shown).
Identification of Phosphoproteins P2 and P3 as Phosphorylated Derivutives of Ribosomal Proteins S2 and S16
Prolonged electrophoresis in the first dimension and staining with Coomassie blue showed that the ribosomal protein S2 froin virus-infected cells can be resolved into three species, namely S2a, S2b, and S2c (Fig. 1 A). The spots S2b and S ~ C , located to the left of the S2a spot, are phosphorylated (Fig. 1 €3). The intensity of their staining decreases and 32P radioactivity increases towards the anode. We have demonstrated that the S2b and S2c proteins are most likely phosphorylated derivatives of S2a, by labelling the S2 proteins in vivo in uninfected and infected cells and by subsequently labelling them in vitro with ['4C]formaldehyde (Fig. 2). Comparison of the total amount of I4C labelling in the S2 protein isolated from uninfected cells (2007 counts/min) and the sum of the ''C-labelling in the separated S2a, S2b and S2c proteins from the infected cells (2188 counts/min) indicates that there is no increase in the amount of S2 protein in infected cells as would be expected if S2b and S2c were proteins synthesized in response to viral infection. These data, and the observation that
the S2a, S2b and S2c proteins co-migrate in dodecyl- sulfate/polyacrylamide gels ( M , 32 500), strongly sug- gest that the S2b and S2c proteins are phosphorylated derivatives of S2a. The shift in the amount of incorpo- rated 32P from the S2b to the S2c spot at the later state of infection (Fig. 5 ) further supports the notion that the S2b and S2c spots originate from S2a by its phosphorylation. The protein P2 can therefore be identified as a composite of two phosphorylated de- rivatives of the ribosomal protein S2.
Under our standard conditions of two-dimensional polyacrylamide gel electrophoresis, the ribosomal proteins S13 and S16 are not well separated but are part of a cluster composed of the proteins S13, S16 and S18. The location of the phosphoprotein P3 to the left of this cluster therefore makes its identification difficult. However, the S16 and S13 proteins recovered from two-dimensional gels can be separated by further fractionation in dodecylsulfate/polyacrylamide gels ('third-dimension') and when this analysis is applied to the P3 protein it is found to co-migrate with the protein S16 of the same apparent molecular weight, M , 17500 (Fig.3). This evidence suggests that the phosphoprotein P3 is a derivative of the ribosomal protein S16.
Anulysis of Phosphoumino Acids
In order to determine whether the [32P]phosphate incorporated into ribosomal proteins was covalently bound to amino acid side chains, the proteins were
466 Ribosomal Proteins Phosphorylated in Vaccinia-Virus-Infected Cells
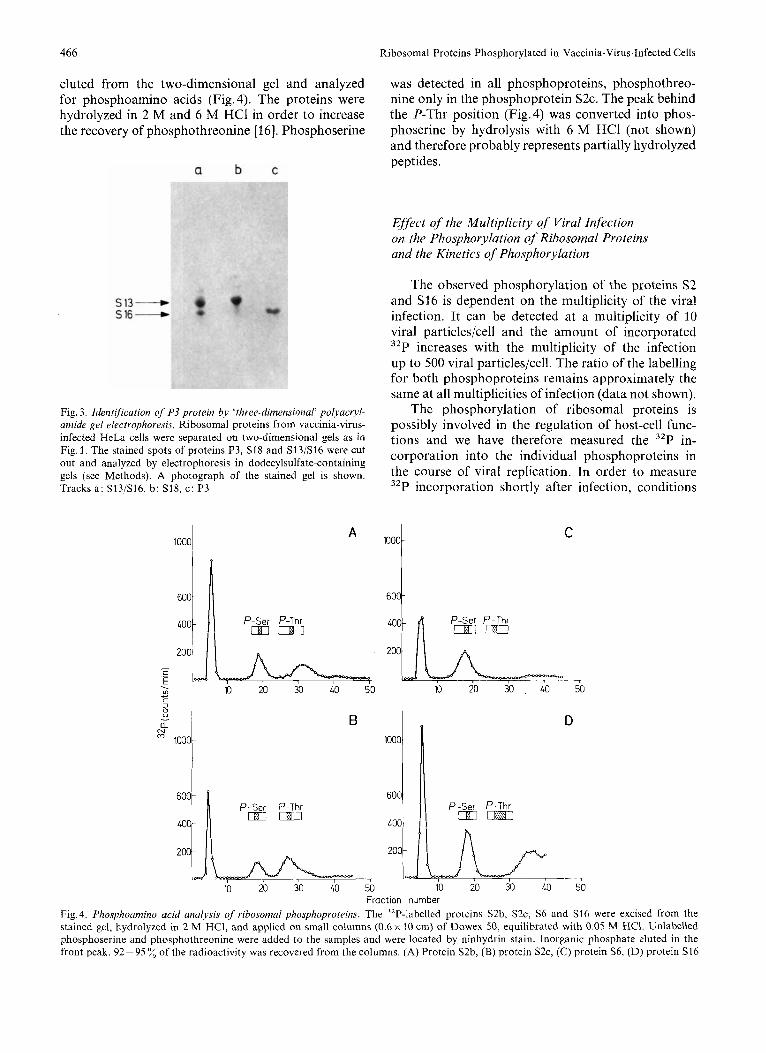
eluted from the two-dimensional gel and analyzed for phosphoamino acids (Fig. 4). The proteins were hydrolyzed in 2 M and 6 M HCl in order to increase the recovery of phosphothreonine [16]. Phosphoserine
Fig. 3. Identification of P3 protein by ’three-dimensional‘ polyacryl- amide gel electrophoresis. Ribosomal proteins from vaccinia-virus- infected HeLa cells were separated on two-dimensional gels as in Fig. 1. The stained spots of proteins P3, S18 and S13/S16 were cut out and analyzed by electrophoresis in dodecylsulfate-containing gels (see Methods). A photograph of the stained gel is shown. Tracks a: S13/S16, b: S18, c: P3
,000~ A I
was detected in all phosphoproteins, phosphothreo- nine only in the phosphoprotein S2c. The peak behind the P-Thr position (Fig. 4) was converted into phos- phoserine by hydrolysis with 6 M HCl (not shown) and therefore probably represents partially hydrolyzed peptides.
EJfect qf the Multiplicity of Viral Infection on the Phosphorylation of Ribosomal Proteins and the Kinetics of Phosphorylation
The observed phosphorylation of the proteins S 2 and S16 is dependent on the multiplicity of the viral infection. It can be detected at a multiplicity of 10 viral particles/cell and the amount of incorporated 32P increases with the multiplicity of the infection up to 500 viral particles/cell. The ratio of the labelling for both phosphoproteins remains approximately the same at all multiplicities of infection (data not shown).
The phosphorylation of ribosomal proteins is possibly involved in the regulation of host-cell func- tions and we have therefore measured the 32P in- corporation into the individual phosphoproteins in the course of viral replication. In order to measure 32P incorporation shortly after infection, conditions
lO0OC C
400 P-Ser P-Thr
200
- -
_I
u D - a. mN 1000
600
LOO
200
P-Ser P-Thr m m
- 7 10 20 30 40 50 10 20 30 40 50
Fraction number
Fig. 4. Phosphoamino acid analysis of ribosomal phosphoprotrins. The 32P-labelled proteins S2b, S ~ C , S6 and S16 were excised from the stained gel, hydrolyzed in 2 M HCI, and applied on small columns (0.6 x 10 cm) of Dowex 50, equilibrated with 0.05 M HC1. Unlabelled phosphoserine and phosphothreonine were added to the samples and were located by ninhydrin stain. Inorganic phosphate eluted in the front peak. 92-95 ”/, of the radioactivity was recovered from the columns. (A) Protein S2b, (B) protein S ~ C , (C) protein S6, (D) protein S16
M. Kaerlein and I. Horak 461
. In
3 6 9
t . L In
3 6 9 Time post- infection (h)
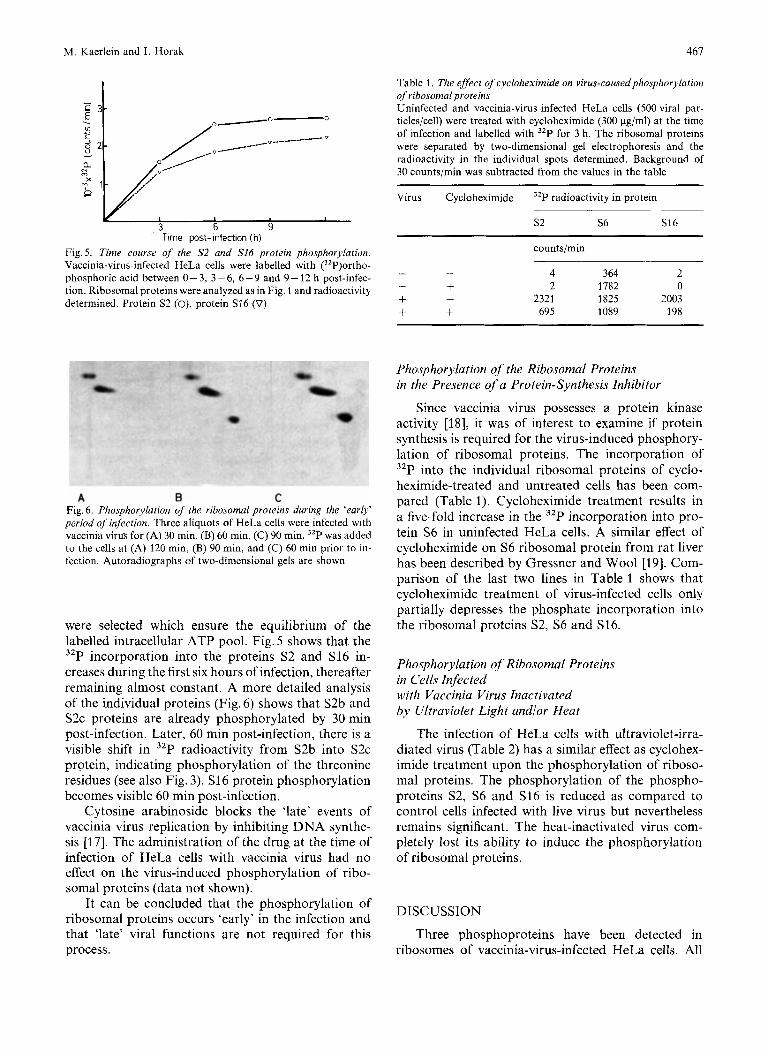
Fig.5. Time course of the S2 and S16 protein phosphorylation. Vaccinia-virus-infected HeLa cells were labelled with (3zP)ortho- phosphoric acid between 0 - 3, 3 - 6, 6 -9 and 9 - 12 h post-infec- tion. Ribosomal proteins were analyzed as in Fig. 1 and radioactivity determined. Protein S2 (O), protein S16 (0)
Table 1. The effect of cycloheximide on virus-caused phosphorylation of ribosomal proteins Uninfected and vaccinia-virus-infected HeLa cells (500 viral par- ticles/cell) were treated with cycloheximide (300 pg/ml) at the time of infection and labelled with 3zP for 3 h. The ribosomal proteins were separated by two-dimensional gel electrophoresis and the radioactivity in the individual spots determined. Background of 30 counts/min was subtracted from the values in the table
Virus Cycloheximide ”P radioactivity in protein ~~
s 2 S6 S16
counts/min
- - 4 364 2 - + 2 1782 0 + - 2321 1825 2003 + + 695 1089 198
A B C Fig. 6. Phosphorylation of the ribosomal proteins during the ‘early’ period of infection. Three aliquots of HeLa cells were infected with vaccinia virus for (A) 30 min, (B) 60 min, (C) 90 min. 32P was added to the cells at (A) 120 min, (B) 90 min, and (C) 60 min prior to in- fection. Autoradiographs of two-dimensional gels are shown
were selected which ensure the equilibrium of the labelled intracellular ATP pool. Fig. 5 shows that the 32P incorporation into the proteins S2 and S16 in- creases during the first six hours of infection, thereafter remaining almost constant. A more detailed analysis of the individual proteins (Fig. 6) shows that S2b and S2c proteins are already phosphorylated by 30 min post-infection. Later, 60 min post-infection, there is a visible shift in 32P radioactivity from S2b into S2c protein, indicating phosphorylation of the threonine residues (see also Fig. 3). S16 protein phosphorylation becomes visible 60 min post-infection.
Cytosine arabinoside blocks the ‘late’ events of vaccinia virus replication by inhibiting DNA synthe- sis [17]. The administration of the drug at the time of infection of HeLa cells with vaccinia virus had no effect on the virus-induced phosphorylation of ribo- somal proteins (data not shown).
It can be concluded that the phosphorylation of ribosomal proteins occurs ‘early’ in the infection and that ‘late’ viral functions are not required for this process.
Phosphorylation of the Ribosomal Proteins in the Presence of a Protein-Synthesis Inhibitor
Since vaccinia virus possesses a protein kinase activity [18], it was of interest to examine if protein synthesis is required for the virus-induced phosphory- lation of ribosomal proteins. The incorporation of 32P into the individual ribosomal proteins of cyclo- heximide-treated and untreated cells has been com- pared (Table 1). Cycloheximide treatment results in a five-fold increase in the 32P incorporation into pro- tein S6 in uninfected HeLa cells. A similar effect of cycloheximide on S6 ribosomal protein from rat liver has been described by Gressner and Wool [19]. Com- parison of the last two lines in Table 1 shows that cycloheximide treatment of virus-infected cells only partially depresses the phosphate incorporation into the ribosomal proteins S2, S6 and S16.
Phosphorylation of Ribosomal Proteins in Cells Infected with Vaccinia Virus Inactivated by Ultraviolet Light and/or Heat
The infection of HeLa cells with ultraviolet-irra- diated virus (Table 2) has a similar effect as cyclohex- imide treatment upon the phosphorylation of riboso- mal proteins. The phosphorylation of the phospho- proteins S2, S6 and S16 is reduced as compared to control cells infected with live virus but nevertheless remains significant. The heat-inactivated virus com- pletely lost its ability to induce the phosphorylation of ribosomal proteins.
DISCUSSION
Three phosphoproteins have been detected in ribosomes of vaccinia-virus-infected HeLa cells. All

468 Ribosomal Proteins Phosphorylated in Vaccinia-Virus Infected Cells
Table 2. Phosphorylation .f ribosomal proteins in cellv infrcted wirh vaccinia virus inaciivated by ultraviolet light and/or heat Three aliquots of HeLa cells were infected with 500 viral particles/ cell of (a) live vaccinia virus, (b) ultraviolet-irradiated virus (496 pJ/ mm’), (c) heat-inactivated virus (60 ‘C, 60 min), and labelled with 3zP for 3 h. The radioactivity in electrophoretically separated pro- teins was determined as described in Methods. The infectivity of the virus preparations was checked by plaque assay on L cell mono- layers and corresponded to (a) 25 p.f.u./cell, (b) 5 x p.f.u./ cell, (c) 2 x lo-’ p.f.u./cell (p.f.u. = plaque-forming units). Back- ground of 30 counts/min was subtracted from the values in the table
Protein 3zP radioactivity in ribosomal protein from HeLa cells infected with
a b C
- ~ - ~~~~
s2 S6 S16
counts/min
4472 3437 3196 2778 2993 440
-~ __ 41
1821 8
three phosphoproteins are located on the 40-S ribo- somal subunit and meet the criteria for ribosomal phosphoproteins as suggested by Eil and Wool [20]. The ribosomal protein S6 is also phosphorylated in uninfected cells. The two additionally phosphorylated proteins have been identified as S2 and S16.
Our results indicate that the phosphorylation of the S2 protein in vaccinia-infected HeLa cells proceeds in two steps to give rise to two phosphorylated forms of protein S2, namely S2b and S2c. The protein S2b is phosphorylated only on the serine residues, while the protein S2c contains phosphoserine and phospho- threonine. The time-lag in the phosphorylation of proteins S2c and S16 could reflect the requirement for some conformational change introduced by the phosphorylation of the protein S2b.
Changes in the phosphorylation of protein S6 are commonly observed when the cellular metabolism is altered. An increase of S6 protein phosphorylation has been reported in situations of stimulated [12] as well as depressed [19] protein synthesis. The phospho- rylation of the S6 protein increases during the infec- tion of HeLa cells with vaccinia virus [3] and adeno- virus [4]. It has been suggested [19] that changes of the intracellular cAMP concentration might be a common modulator of S6 phosphorylation. However, some contradictory results have recently been reported. An increase of the intracellular cAMP concentration alone does not lead to the enhancement of S6 phos- phorylation [21,22], and conversely, increased phos- phorylation of S6 protein has been reported under conditions when cAMP concentration was not ele- vated [23].
Cyclohexiniide treatment increases the phospho- rylation of the S6 protein in uninfected HeLa cells,
but decreases its phosphorylation in vaccinia-virus- infected cells (Table 1). Also, the phosphorylation of the proteins S2 and S16 is in some part dependent upon protein synthesis, but not entirely; for S2 and S16, as well as for S6, there remains a significant level of virus-induced phosphorylation even when the pro- tein synthesis was inhibited over 99 %,. These two ob- servations are consistent with our notion that the pro- tein kinase from the inoculated virus particles plays a part in the phosphorylation of the ribosomal pro- teins S2, S6 and S16. The partial dependence on protein synthesis might reflect the synthesis of the new viral protein kinase which should be an ‘early enzyme’ (conclusion from experiments with cyto- sine arabinoside). The data presented do not ex- clude the possible activation of a pre-existing cellu- lar protein kinase by a structural component of the vaccinia virion. However, this seems to be less likely because our preliminary experiments show that the proteins S2, S6 and Sl6 can also be phosphorylated in vitro with vaccinia protein kinase and ( Y - ~ ~ P ) ATP.
We do not have any direct evidence on the func- tional significance of the virus-induced modifications of the host-cell ribosomes. However, our results in- dicate an interesting correlation between the phos- phorylation of the ribosomal proteins and the switch- off of the host cell protein synthesis. First, the time course of the phosphorylation correlates well with the time course of the protein synthesis inhibition in infected cells and, second, ultraviolet-irradiated virus, which shuts off host protein synthesis [7], is still active in phosphorylation of the ribosomal pro- teins.
Evidence to suggest that the phosphorylation reac- tions might be involved in the regulation of protein synthesis has come from the recently discovered pro- tein kinase activity associated with a protein-syn- thesis inhibitor in heme-deficient rabbit-reticulocyte lysates [24,25]. The mechanism whereby phosphor- ylation could influence the host-cell protein-synthe- sizing apparatus is, however, still obscure. Phosphor- ylation of a ribosomal protein does change its elec- trical charge and such modification may alter some functional sites on the ribosomes and as a consequence influence the interactions of the ribosomes with other components (for example, binding of an initiation factor or mRNA) of the protein-synthesizing complex. The importance of electrostatic interactions has been stressed recently by Adams et al., who tested the effect of differently charged ‘cap’ analogs on reovirus mRNA binding to wheat germ ribosomes [26].
We thank Drs E. Wecker and S. Sidell for critical reading of the manuscript and Ms 1. Sauer for technical assistance. This pro- ject was supported by the Deutschr Forscliung.sgenzrinschajt, SFB 105.

M . Kaerlein and I. Horak 469
REFERENCES
1 . Joklik, W. K . (1968) Annu. Rev. Microbiol. 22, 359-390. 2. Horak, 1. (1975) Curr. Top. Microhiol. Immunol. 69, 117-135. 3 . Kaerlein, M. & Horak, I. (1976) Naiure (Lond.) 259, 150-151. 4. Blair, G. E. & Horak, I. (1977) Biochem. Soc. Trans. 5, 660-
5 . Horak, I. & Schiffiiiann, D. (1977) Eur. J . Biochem. 79, 375-
6. Joklik, W. K. (1962) Virology, 18, 9- 18. 7. Moss, B. (1968) J . Virol. 2, 1028-1037. 8. Sherton, C. C. &Wool, 1. G. (1972) J , Biol. Chem. 247,4460-
9. Kaltschmidt, E. & Wittinann, H. G. (1970) Anal. Biochem. 36,
661,
380.
4467.
401 -402. 10. Laemmli, U. K. (1970) Nature (Lond.) 227, 680-682. 11. Olson, M. 0. J., Orrick, L. R., Jones, C. E. & Busch, H. (1974)
12. Gressner, A. M. & Wool, I . G. (1974) J . Biol. Chem. 249,
13. Rice, R. H. & Means, G. E. (1971) J . Biol. Chem. 246, 831-
J . Biol. Chem. 249, 2823 - 2827.
691 7 - 6925.
832.
14. Lowry, 0. H., Rosebrough, N. J., Farr, A. L. & Randall, R.
15. SchifTmann, D. & Horak, I. (1978) Eur. J . Biochem. 82, 91 -95. 16. Bylund, D. B. & Huang, T. S. (1976) Anat. Biochem. 73, 477-
17. Oda, K. I. & Joklik, W. K. (1967) J . Mol. B i d . 27, 395-419. 18. Paoletti, E. & Moss, B. (1972) J . Virol. 10, 417-424. 19. Gressner, A. M. & Wool, I. G. (1974) Biochem. Biophys. Res.
20. Eil, C. & Wool, 1. G. (1973) J . Biol. G e m . 248, 5122-5129. 21. Treloar, M. A., Treioar, M . E. & Kisilcvsky, R. (1977) .I. Biol.
22. Horak, I. & Koschel, K. (1977) FEBS Lett. 83, 68-70. 23. Leader, D. P., Rankine, A. D. & Coia, A. A. (1976) Biochrm.
24. Levin, D. H., Ranu, R. S., Ernst, V. & London, I. M. (1976)
25. Farrell, P. J . , Balkow, K., Hunt, T., Jackson, R. J. & Trachsel,
26. Adanis, B. L., Morgan, M., Muthukrishnan, S. , Hecht, S. M.
J. (1951)J. Bid . Chem. 193, 265-275.
485.
Commun. 60, 1482- 1490.
Chem. 252,6217-6221.
Biophys. Res. Commun. 71, 966-974.
Proc. Narl Acad. Sci. U.S.A. 73, 31 12-3116.
H. (1977) CeN, I / , 187-200.
& Shatkin, A. J. (1978) J . Biol. Chem. 253, 2589-2595.
M. Kaerlein and I. Horak*, Institut fur Virologie und Immunbiologie der Justus-Maximilians-Universit~t Wurzburg, Versbacher LandstraBe 7, D-8700 Wurzburg, Federal Republic of Germany
To whom correspondence should be addressed





























