Embed Size (px)
Citation preview

IDENTIFICATION AND CHARACTERIZATION OF CUCUMBER MOSAIC AND BEAN YELLOW MOSAIC
VIRUSES AFFECTING CHICKPEA (CICER ARlETlNUM L.) IN INDIA
Thesis submitted to the Andhra Pradesh Agricultural University in part fulfilment of the requirements
for the award of the degree of
DOCTOR OF PHILOSOPHY
by
T. VENKATA CHALAM, M. Sc. (Ag.)
Department of Plant Pathology College of Agriculture Andhra Pradesh Agricultural University Rajendranagar, Hyderabad 600 030
Pulse Pathology Subprogram ICRISAT Patancheru P.O. 6 0 2 324 Andhra Pradesh India
November 1982

IDENTIFICATION AND CHARACTERIZATION OF
CUCUMBER MOSAIC AND BEAN YELLOW MOSAIC VIRUSES AFFECTING CHICKPEA (CICER ARlETlNUM L.) IN INDIA
Thesis submitted to the Andhra Pradesh Agricultural University in part fulfilment of the requirements
for the award of the degree of
DOCTOR OF PHILOSOPHY
by
T. VENKATA CHALAM, M . Sc. (Ag.)
Department of Plant Pathology College of Agriculture Andhra Pradesh Agricultural University Rajendranagar, Hyderabad 5 0 0 0 3 0
Pulse Pathology Subprogram ICRISAT Patancheru P.O. 5 0 2 324 Andhra Pradesh India
November 1982

CERTIF ICATE
Shr i T. Venkata Chalam has s a t i s f a c t o r i l y prosecuted t h e
course of research and t h a t t h e t h e s i s e n t i t l e d " I d e n t i f i c a t i o n and
cha rac t e r i za t i on o f cucumber mosaic and bean yellow mosaic v i r u s e s
a f f e c t i n g chickpea (Cicer , . ari,etinum L.) i n Indi,a," submitted i s t h e
r e s u l t o f o r i g i n a l research w o ~ k and d s of s u f f i c i e n t l y h igh s tandard
t o warrant i t s presentaf ion t o t h e examination. We,also c e r t i f y
t h a t t h e t h e s i s o r p a r t i t h e r eof has*?gt. peen previous ly submitted
by him f o r a degree of any Univers i ty .
Co- Chairman
Y.L. NENE Pr inc ipa l P l an t Pa thologis t and Leader, Pulses Improvement Program I CRISAT Patancheru P.O. 502 324 Andhra Pradesh India
Chairman
J . SUBBAYYA Professor and Head Department of P l an t Pathology S. V. Agr i cu l tu ra l College T i rupa t i 517 502 Andhra Pradesh India

CERTIFICATE
This i s t o c e r t i f y t h a t the t h e s i s e n t i t l e d " I d e n t i f i c a t i o n
and c h a r a c t e r i za t ion of cucumber mosaic and bean ye1 low mosaic
v i r u s e s a f f e c t i n g chickpea (cicer arietinum L .) i n India" submit ted
i n p a r t i a l f u l f i l m e n t of t h e requirements f o r t h e degree of Doctor
of Philosophy i n Agr icu l tu re , i n t h e major s u b j e c t o f P lan t Pathology
of t h e Andhra Pradesh A g r i c u l t u r a l Univers i ty , Hyderabad, i s a record
of the bond fide r e s e a r c h work c a r r i e d o u t by Mr. T. Venkata Chalam
under our guidance and ruperv i s ion . The s u b j e c t o f t h e t h e s i s has
been approved by t h e S t u d e n t ' s Advisory Committee.
No p a r t o f t h e t h e s i s has been submit ted f o r any o t h e r
degree o r diploma.
A l l a s s i s t a n c e and h e l p received dur ing t h e course o f t h e
i n v e s t i g a t i o n s have been duly acknowledged by him.
7 ' . -Y 1 7. \ +, .;..4 -.
- . _ - _. _. ... .- -- Y . L . NENE Co-Chairman Advisory Committee
- G -3
-:. 2
J . SUBBAYYA Chairman Advisory Committee
The t h e s i s approved by t h e Advisory Committee. - -'
Chairman (J. SUBBAYYA) 2
Co- Chairman (Y . L . NENE)
Member (S.P.S. BENIWAL)
Member (N. GOVINDA RAO)
Member (A. NARAYANAN)
Member (V. ARUNACHALAM)

CONTENTS
Page No,
List of i l l u s t r a t i o n s
L i s t of t a b l e s
Acknowl edgnent
Abstract
List o f symbols and abbrevia t ions
I . INTRODUCTION
11. REVIEW OF LITERATURE
111. MATERIALS AND METHODS
General
I d e n t i f i c a t i o n o f v i ruses by serology
Host range and symptoms
Physical p roper t i e s of v i ruses i n crude sap
Insect transmission
P u r i f i c a t i o n of v i ruses
Elect ron microscopy
Physico-chemical p roper t i e s of p u r i f i e d CMV
Production of antiserum t o CMV
b e t e r k n a t ~ o n o f t h e serological r e l a t i o n s h i p between t h e p resen t s t r a i n of 6n and o ther known st r a i n s
Screening f o r d i sease r e s i s t a n c e
I V . RESULTS
Symptomatology i n chickpea
Host range and symptoms
I n vi tro proper t i e s
Insec t transmission - P u r i f i c a t i o n o f v i r u s e s
Elect ron microscopy
I d e n t i f i c a t i o n of v i ruses by serology
Physico-chemical p roper t i e s o f p u r i f i e d CMV
Production o f antiserum t o CMV
v i i
v i i i

Serological r e l a t i o n s h i p between presen t s t r a i n . . of CMV and o ther known s t r a i n s
Screening f o r d i sease r e s i s t a n c e . . V. DISCUSSION AND CONCLUSION . .
V I . SUMMARY . . LITERATURE CITED D .
APPEND1 X . . VITA . .
Page No,
60

List of I l l u s t r a t i o n s
Fig.No. I l l u s t r a t i o n
1. Symptoms produced by CMV on chickpea . O
2. Symptoms produced by BYMV on chickpea 0 .
3 . Local l e s i o n s produced by CMV on Chenopodium . . m a n t i c o Z o r , broad bean, Phuseolus vulgaris v a r . Pinto , and P. vulgaris var. P o r r i l o
4 . Local l e s i o n s produced by CMV on P. vulgaris . . v a r . Lavica, Vigna mguiculata var . Pusa Dophasli and V. wtguiculata var. Barsa t i mutant
5 . Systemic symptoms produced by CMV on . . Nicotiana rus t i ca , cucumber and peas
6. Systemic symptoms produced by BYMV on . . P. vuZgaris
7 . Light s c a t t e r i n g zone o f CMV i n d e n s i t y . . grad ien t tube ( a ) , uv absorpt ion and i n f e c t i v i t y o f f r a c t i o n s of sucrose dens i ty g rad ien t tube @), and uv absorp- t i o n spectrum o f p u r i f i e d CMV (c)
8 . Elect ron photomicrograph of p u r i f i e d CMV . . 9. Electron photomicrograph of l e a f d i p . .
prepara t ion o f BYMV
10. Length d i s t r i b u t i o n histogram of BYMV . . 11. Elect ron photomicrograph o f p u r i f i e d BYMV . . 12. U l t r a t h i n s e c t i o n s of BYMV i n f e c t e d . .
chickpea l e a f
13. Agar double-di f fus ion t e s t f o r i d e n t i - . . f i c a t i o n o f CMV and BW
14. Enzyme- l i k e d immunosorbent assay f o r . . i d e n t i f i c a t i o n o f CMV
15. Schle i ren p a t t e r n o f p u r i f i e d CMV i n an . . a n a l y t i c a l u l t r a c e n t r i f u g e
16. UV absorpt ion and d e n s i t i e s o f C s C l gradient . . column (CMV)
17. Polyacrylamide gel e l ec t rophores i s of CMV . . coat p r o t e i n (a) and (;MY RNAs (b)
18. Photometric scanning o f CMV coat p r o t e i n ( a ) , . . marker p r o t e i n s (b), CMV coat p r o t e i n and markers (c) and re la t ic in between migra- t i o n d i s t ance and l o g mol .wt. o f markers (d)
Page No.
4 7b
4 7d
4 7f

Lis t of Tables
S.No. Table - - 1. Randomization o f t r ea tments t o determine
thermal i n a c t i v a t i o n p o i n t o f CMV
2 . Randomization o f t r ea tments t o determine d i l u t i o n end p o i n t of CMV
3. Host range and symptomatology of CMV and BY MV
4 . Inf luence o f hea t ing t h e s t andard e x t r a c t o f CMV a t d i f f e r e n t temperatures f o r 10 min on i n f e c t i v i t y
5. Influence of d i l u t i n g t h e s t andard e x t r a c t o f CMV on i n f e c t i v i t y
6. Inf luence o f s to rage o f t h e s t andard e x t r a c t of CMV a t room temperature on i n f e c t i v i t y
7 . Inf luence of hea t ing t h e s t andard e x t r a c t o f BYMV a t d i f f e r e n t temperatures f o r 10 min on i n f e c t i v i t y
8 . Inf luence o f d i l u t i n g t h e s t andard e x t r a c t o f BYMV on i n f e c t i v i t y
9. In f luence of s to rage o f t h e s tandard e x t r a c t of BYMV a t room temperature on i n f e c t i v i t y
10. Aphid t ransmiss ion o f CMV and BYMV
11. Sero log ica l a f f i n i t y o f p resen t CMV s t r a i n wi th o t h e r known s t r a i n s o f CMV
Page No,
34

ACKNOWLEDGMENT
1 de& hmende p t e a s w ~ e i n e ~ p h e b b i n g my h c a e 2hanh6 and deep senbe 0 6 g m f i d e la Dh. Y . 1 , N w e , P h i n c i p d P h n t Pa&o&ogAt and l e a d a , P u b e s lmpmvement P m g m , ICRlSAT, doh buggu. t ing t h e pmblem and doh kib enwwuzgement and guidance dwLing t h e c o w e 0 6 t k i d i n v e b t i g a t i a n , and in p / t e p W o n 0 6 .the manub+t.
1 am k i g M y h k 6 d ;to Dh. 3. Subbayya, Pkodedbofi and H a d , Dept. 0 6 PlanX P a t h o b g y , S . V. A g h i d W CoUege, T A W , doh kis v a l u a b l e buggebtLon6 m d doh b m v i n g M ChlLiRrnan 0 6 Xhe Advbotry Conanittee.
1 am g m t e d d RD Dh. M.V. Reddy, Chickpea PaZhotog-bt, lCARVA, S y ~ a , doh h d p i n g me ach ieve p a d e c f i o n in t h e 6iel .d 0 6 plant v h L o g y .
7 am aedo ; t hank id b Dh. S.P.S. Beniwd, S e n i o t P l a n t PadhoLogAist ( P u b e s I , ICRISAT, doh kis h d p dwLing A%Q h.W phA ad my wotrk, 60tr go ing t h o u g h & manuc&ipt, and 604 acting ad Membeh od isthe AdvAohy CormLittee.
T h a n k m e aedo due ;to Dh, D. V. R. Reddy, P ~ i c i p d V h t o g A t , ICRlSAT, doh kis v d u a b l e s u g g u k i o n b .
1 am thank6u.l RD D u . N , Govinda Rao, A. Nuhayanan and V. Atrunachdeam doh acaXng as membcm od t h e AdvAorry ComnuXee, a3 Dh. R. JambunaLhan, P a n c L p a t 12iochemist, 1CRlSAT; M u b ~ . Y. 8 b h , S.K. M a n a h , S.M. Raju, and b.tad6 0 6 P&e Pathology, ICRlSAT doh 2%eih b h c e h e h d p .
1 w h h ib t o k e f&.6 p /L i vdkge b exptredb y prto60Wld senbe 0 6 h e s p e d and Love b my s A & , Mu. T. V. S u b b m and my b m t h m Mk. T . Venkac teaodu, who bo tdey ltook up t h e b w d e n 0 6 my d u d o n eve4 b h c e t h e demide ad my dathm.
1 bpeciaeey Zbnh my w i d e S a v W , doh h m w o p , W n and LnbpjrLLttion d u g my Ph.D. prrogham,
The & U o u u k i p and 6acLlLiAa p w v i & d by ZCRISAT dwLing .the c o m e 0 6 a%& invesa%jat ian m e ghateduiQ acknavledged.
7 w h h a% euplle6b my appke-n Xu to. R. N a h s h g Rm, do t typ ing Xki.4 manttbchipt and to h. H.S. kis w U a g u ~ 604 pbhgrrapkic wohh.
. . T . VENKATA CHALAM

ABSTRACT
Author : T. Venkata Chalam
T i t l e of the t h e s i s : I d e n t i f i c a t i o n and charac te r i za t ion o f cucumber m s a i c and bean yellow m s a i c v i ruses a f f e c t i n g chickpea (Cicer m i e t i n t m L . ) i n India
Degree : Ph.D.
Facul ty : Plan t Pathology
Guides : Y.L. Nene (ICRISAT) J . Subbayya (APAU)
Universi ty : Andhra Pradesh Agricul tura l Universi ty
Year of submission : 1982
Two mechanically t r ansmiss ib le v i ruses ; v iz . , cucumber mosaic
v i r u s (CMV) and bean yellow mosaic v i r u s (BYMV) were found t o n a t u r a l l y
i n f e c t chickpeas. These were i d e n t i f i e d by studying t h e i r hos t range,
biophysi ca l p roper t i e s , i n s e c t transmission, e l ec t ron microscopy and
serology. Also e l i t e chickpea germplasm access ions were screened f o r
r e s i s t a n c e t o these v i r u s e s .
Both the v i ruses i n i t i a l l y produced twis t ing o f terminal bud
i n chickpea. Subsequently, CMV e i t h e r produced w i l t i n g and death of
p l a n t s o r p r o l i f e r a t i o n and bushiness of branches bearing very small
green leaves and very small pods, whereas BYMV produced p r o l i f e r a t i o n
of branches with narrow o r f i l i f o r m leaves, bearing very small pods.
The (51V could i n f e c t 13 p lan t species belonging t o f i v e
fami l i e s , whereas t h e hos t range o f BYMV was r e s t r i c t e d to Leguminosae
only. The r e s u l t s on i n vitro p r o p e r t i e s ind ica ted t h a t CMV was more
s t a b l e than BYMV. Both t h e v i ruses could be t ransmit ted by two aphid
species ; v i z . , Aphis maccivora and t$jzus perskae i n a non-pers is tent
manner.
An e f f i c i e n t p u r i f i c a t i o n procedure developed for CMV yielded
80-100 mg o f v i r u s per kg of p lan t t i s s u e . The CMV p a r t i c l e s were

s p h e r i c a l and measured about 30 nm i n d iameter . The p u r i f i e d CMV had
a buoyant d e n s i t y and sedimentat ion c o e f f i c i e n t o f 1.325 g/ml and 104S,
r e s p e c t i v e l y . Using t h e polyacrylamide ge l e l e c t r o p h o r e s i s (PAGE),
t h e molecular weight o f CMV coat p r o t e i n was determined t o be 25,000
da l tons . The PAGE o f CMV WAS revea l ed t h e presence o f f o u r RNA s p e c i e s ,
The p r e s e n t s t r a i n of CMV was r e l a t i v e l y more c l o s e l y r e l a t e d t o CMV-C
pool, fo l lowed by CMV-Ix and M-CMV. The ant i serum produced t o CMV had
a t i t r e o f 1/1024 a s determined by agar double-d i f fus ion t e s t .
A s a t i s f a c t o r y p u r i f i c a t i o n procedure was developed f o r BYMV.
The p a r t i c l e s of BYMV measured 750 nm i n l e n g t h and 15 nm i n width.
Pinwheels and l amina t e agg rega t e s t y p i c a l o f po tyv i ruses were observed
i n t h e cytoplasm of BYMV-infected ch ickpea c e l l s . S ince t h e v i r u s
p repa ra t ion f r e e o f hos t components cou ld n o t be obta ined , t he physico-
chemical p r o p e r t i e s o f BYMV were n o t determined.
E l i t e chickpea germplasm acces s ions were screened f o r r e s i s -
tance t o both CMV and BYMV, and sou rces o f r e s i s t a n c e were i d e n t i f i e d
f o r both t h e v i r u s e s .

List o f symbols and abbrevia t ions
A absorbance
absorbance a t 260 nm
absorbance a t 280 nm
A max absorbance maximum in a absorption spectrum
A min
APAU
BY MV
cm
am cv
DEP
absorbance minimum i n a absorption spectrum
Andhra Pradesh Agricul tura l Universi ty
bean yellow mosaic v i r u s
centimeter ( s )
cucumber m s a i c v i r u s
c u l t i v a r
d i l u t i o n end point
E an a l t e r n a t i v e symbol f o r A
s p e c i f i c ex t inc t ion coef f i c i en t = the absorbance value of a 0.1% so lu t ion i n a cuvet te wi th an o p t i c a l path length o f t h e l i g h t o f 10 mm
: idem, but a t a wave length of 280 nm
E DTA : ethyl enediaminete t raacet ic acid, sodium s a l t
EL I SA : enzyme-linked immunosorbent assay ( se ro log ica l 5
I . C q t e s t ) . .
g : gram(s) (a u n i t symbol of mass)
g ' : i n i t i a l acce le ra t ion i n f r e e f a l l . Values i n u l t r acen t r i fuga t ion a r e given in t h i s g r a v i t y u n i t
I CC
ICRISAT
: hydrochloric a c i d
: hour ( s )
: In te rna t iona l Center k r A g r i c u l t u r a 1 Research i n t h e Dry Areas
: ICRISAT chickpea
: In te rna t iona l Crops Research I n s t i t u t e f o r t h e Semi -Arid Tropics
: potassium chlor ide
: potassium phosphate, m n o b a s i c

mm
ml . w t .
N
NaCl
Na2CO3 NaHC03
NaH2POq
Na2HP04
NaOH
nm
OD PAGE
P B ~ '
PEG
P04 p s i
PSV
PVP
BNA
RNAs
RNase
rpm S
longevity i n vitm molar concentrat ion i n grams per l i t r e
m i l l i molar concentration = 0.001M
milligram = 0.001 g
m i l l i ampere
minute( s )
m i l l i l i t e r = 0.001 l i t r e
mil l imeter = 0.001 meter
molecular weight
normality
sodium ch lo r ide
sodium carbonate
sodium b i carbonate
sodium phosphate, monobasic
sodium phosphate, d ibas ic '
sodium hydroxide -9 nanometre = 10 m (formerly mp was used)
a l t e r n a t i v e symbol f o r A
polyacrylamide gel e lec t rophores i s
phosphate buffered s a l i n e
polyethylene glycol
hydrogen ion exponent ( a u n i t symbol f o r t he degree of a l k a l i n i t y o r a c i d i t y )
phosphate
per square inch
peanut s t u n t v i r u s
polyvinyl pyrrolidone
r ibonucle ic ac id
r ibonucle ic ac ids
r ibonuclease
revolu t ions per minute (cent r i fuga t ion)
svedberg un i t s , 10-l3 sec. Values o f sedime: t a t i o n c o e f f i c i e n t s
: sedimentation c o e f f i c i e n t a t 2 0 ' ~ i n water
: sodium dodecyl s u l f a t e

TAV
TEME D
TIP
T r i s
Tween-20
W
tomato aspermy v i r u s
N , N , N t , N ' t e t r amethy l e thy lene diamine
thermal i n a c t i v a t i o n p o i n t
t r i s (Hydroxymethyl) Aminomethane
polyoxyethylene s o r b i t a n mono1 a u r a t e
u l t r a v i o l e t
v a r i e t y
volume/volume
weight/volume
weight /weight
microgram = g
m i c r o l i t r e = l om6 l i t r e
d i f f e r e n c e i n t h e d i s t a n c e t r a v e l l e d between any two t ime i n t e r v a l s
: d i f f e r e n c e between any two fime i n t e r v a l s
: r o t o r ' s angu la r v e l o c i t y
: mean o f t h e d i s t a n c e s t r a v e l l e d

INTRODUCTION
Chickpea (Cicer a r i e t i m L . ) is an important g ra in legume crop of dry-
land a g r i c u l t u r e i n Asia, Africa and Central and South America. The
t o t a l c u l t i v a t e d a r e a o f chickpea i n the world i s about 9 . 3 mil l ion
hec ta res with annual production of about 4 .8 mi l l ion tonnes (FAO, 1980)
The average y i e l d s pe r hectare a r e es t imated t o be around 700 kg.
Chickpea i s known by o t h e r names such a s Bengal gram, gram, Egyptian
pea, Spanish pea, Chestnut bean ( a l l Engl ish) , Pois chiche (French),
chana, (Hindi) , homos (Arabic), grao-de-bico (Portuguese), garbanzo o r
garavance (Spanish), e t c .
About 50 pathogens have so f a r been repor ted on chickpea
from d i f f e r e n t p a r t s of t h e world (Nene, 1980). While many r e p o r t s
a r e mere records o f occurrence, severa l d i seases a r e widespread and a
few a r e devasta t ing. A survey o f t h e l i t e r a t u r e r e v e a l s t h a t on ly a
few d i seases have been inves t iga ted i n d e t a i l (Nene e t aZ . , 1978b) .
h r r g chickpea diseases, w i l t and r o o t r o t s a r e considered
t o be of ,major importance. Considerable confusion e x i s t e d i n t h e p a s t
i n diagnosing var ious chickpea d i so rders , p a r t i c u l a r l y w i l t s and r o o t
r o t s (Nene e t a t . , 1978a). A t ICRISAT ( In te rna t iona l Crops Research
I n s t i t u t e f o r t h e Semi-Arid Tropics), a p r o j e c t was i n i t i a t e d i n 1974
on ' w i l t complex1. During the course of i n v e s t i g a t i o n s i n t o t h e
e t i o l o g y o f so -ca l l ed w i l t c o q l e x , two v i ruses ; v i z . , a l f a l f a mosaic
v i r u s and pea l e a f r o l l v i r u s were i d e n t i f i e d and found t o be t h e compo-
n e n t s o f wilt complex (Nene e t a1 ., 1978a). Chickpea p l a n t s a f f e c t e d
by t h e s e two v i r u s e s o f t e n d ie prelpaturely and hence were wrongly
considered t o be a f f e c t e d by %wilt1'.
Present s t u d i e s were i n i t i a t e d t o know i f o t h e r v i ruses

a f f e c t e d chickpeas under natura l condit ions i n India, and t h e i r c o n t r i -
but ion, i f any, t o the " w i l t complexw.
With the exception of b r i e f r epor t s , very l i t t e has been
published on the i d e n t i t y o f v i ruses a f f e c t i n g chickpeas i n na tu re .
Viral d iseases of chickpea may have escaped detection i n many coun t r i e s
s ince the macroscopic symptoms produced i n such p l a n t s a r e o f t e n n o t
conspicuous. Because o f premature death of these p lan t s , symptoms a r e
f requent ly confused with those o f w i l t o r r o o t r o t s . I t i s therefore ,
necessary t o observe t h e suspected p l a n t s ca re fu l ly and the v i r a l
na tu re needs t o be confirmed by s p e c i f i c procedures (Kaiser and Danesh,
197 l a ) .
The present inves t igat ions were undertaken with t h e o b j e c t i v e
o f i s o l a t i n g v i ruses a f f e c t i n g chickpeas in nature . Wo mechanically
t ransmiss ible v i ruses ; v i z . , cucumber mosaic v i rus (CMV) and bean ye l low
mosaic v i r u s (BYMV) were i so la ted and t h e i r i d e n t i t y was confirmed by
procedures such a s host range, biophysical proper t ies , i n s e c t t r a n s - . . mis$ion, serolbgy, and e lec t ron, microscopy. I n addi t ion, production
of antiserum and screening of germplasm l i n e s f o r d i sease r e s i s t a n c e
were a l s o attempted.

REVIEW OF LITERATURE
I. MICKPEA VIRUSES
S i x v i ruses have been reported t o i n f e c t chickpea from
d i f f e r e n t p a r t s of the world (Nene, 1980). These were: a l f a l f a mosaic
v i r u s from Algeria, India , Iran, Morocco and U.S .A. ; bean yellow mosaic
v i r u s from I ran and U.S.A.; cucumber mosaic v i rus from Colombia, India ,
I ran and USSR; l e t t u c e n e c r o t i c yellows v i r u s from Austra l ia ; pea l e a f
r o l l v i r u s from Algeria, Bangladesh, Ethiopia, India, I ran , Lebanon,
b r o c c o , New Zealand, Pakistan, Sudan, Syria, Tunisia, and Turkey;
pea enat ion mosaic v i r u s from U.S.A.
I n India, four v i ruses have been reported t o n a t u r a l l y occur
on chickpea; v i z . , a l f a l f a m s a i c v i rus (Nene e t at., 1978a), pea l e a f
r o l l v i r u s (Nene and Reddy, 1976; Nene et at. , 1978a; ICRISAT, 1980),
cucumber mosaic v i r u s (Dhingra e t aZ., 1979) and an un iden t i f i ed v i r u s w
causing l e a f d i s t o r t i o n and k s a i c (Mali and Vyanjane, 1980).
11. CUCUMBER MOSAIC VIRUS (CMV)
A: F i r s t Description
The v i r u s was f i r s t described by Dool i t t l e (1916) and Jagger
(1916) i n cucumber. The natura l occurrence of W on chickpea was
f i r s t repor ted by Kaiser e t a2 . (1968). In India i ts na tu ra l occurrence
on chickpea was repor ted by Dhingra e t at. (1979). -
B, Symptoms on Chickpea
Kaiser and Danesh (1971a) repor ted t h a t W caused bushiness
and shortening of internodes i n chickpea. Dhingra e t aZ. (1979) a l s o

observed diseased p lan t s both infected natura l ly and experimentally,
developed ( i n 3-4 weeks a f t e r infect ion) excessive a x i l l a r y shoots
bearing very small, narrow, s l i g h t l y chlorot ic , and deeply denta te
leaves. Affected p lan t s remained stunted giving a bushy appearance.
Phloem of roo t s and c o l l a r region stem developed s l i g h t necros is i n
severe cases of infect ion. Diseased plants bore fewer flowers and
pods than t h a t of the healthy, and plants infected a t a very ea r ly
stage of growth died prematurely, Similar symptoms were observed i n
p lan t s a r t i f i c i a l l y inoculated with the causal v i rus .
C. Host Range
Host range of CMV a s reported by Pr ice (1940), Komuro (1958,
1973), Smith (1972), and Douine e t aZ. (1979) i s given below:
A 1 ZOACEAE
APOCYNACFAE . 9 '
ASCLEPIADACEAE
BALSAMINACEAE
BOUGINACEAE
BROMELIACEAE
CAPPARI DACEAE -
CARRY OPHY LLACEAE
CHENOPODIACEAE
: Mesmhyanthwn crystQZZinwn L . , TetragoniQ e;.cpansa Mum.
: W a n t h u s caudatus L., b a n t h u s r e t r o f l m L.
A. tricoZor L. , Celosia argent& L.
: Vinca minor L. , V. rosea L.
: ~ s c ~ e p i a s ' &iaca L . : Imptiens baZsamina L .
: Anchus sp. , CynogZosswn d i Z e Stapf and Drummond . , BeZiotropiun corynboswn Rui z and Pav . , H. perwiunun L , , Uyosotie syluatioa Hoffm.
: Amnas cornsus Merr.
: Lobetia cardiml is L.
: CZeome spinosa L . , Polanisia trcrchyspemna Torr and
Gray.
: Lychnis aZ& M i l l . , L. chakedonica L. , L. visoaria L.
: Beta vulgaris L . , Chempdiwn atbun L., C. mmZe L. ,
Spimcicr olemoea L.
: Cometinu commmis L., C. errsctu L . , C . nudiflora L . ,
lbadeeoanticr sp., Zebrinu p d u Z a Schni zl .

COMPOS I TAE : hnbrosia e ta t ior L . , CaZenduZa of f ic inaZis L . ,
C a l l i s k p h w chinensis Nees., Centaurea moschata
L . , Chrysanthemum sp. , Cichoriwn endivia L.,
&ilia sagi t t a ta (vahl) Dc., Helianthus annuus L . , H. debi l i s Nutt . , Helichrysm bmc teatwn Andr . , Lactwa satiuz L., Tagetes erecta L . , T. ptula L. , Zinnia ezegans Jacq.
COWOLVULACEAE : CuscutQ subinclusa Durr., C. cazifornica Hook 6 Am.,
Ipmoea batatus Lam., I. pwpurea Lam.
CRUCIFERAE : B m s s h a mpa L . , LobuZaria mritim Desv.,
Nasturtiwn of f ic inaze R. Br.
CUC URB I TAC EAE : Benincasa cerifera Cogn,, B. hispida Cogn., B ~ y o n k
aZba L., B. dwica L . , Bryonopsis lacimosa
Naudin., Chayota edulis Jacq., CitruZZus vulgaris Schrad., C w d s angwia L . , C. f i c i - f o l k Bouche., C. flellruosus Naudin., C. grossu-
h k e f o Z m i 6 Hort., C. melo L . , C. m t u z i f e r w -6
May., C. odoratissirnus Moensch., C. sativus L.,
C. uti l issima Roxbg. Cucmbita marima Duchesne.,
C . .mschata Duchesne., C. p e p L . , EcbQZZiwn' . .
e l a t e r i m A. Rich., Kedrostis foetidissima Cogn.,
Lugenaria leucantha Rusby., L. vulgaris Sch.,
Luffa acutanguk Roxb., L. cyZi&ica Roem.,
MeZothria scabra Naudin., ~ c n m r p e l i s l o b a h
(Mich.) Greene., Memrdica balsamina L . , M.
charantia L . , M. involwrata E. Meyer . , Physedm
barteri Cogn., Sicyos angutatus L., bichosanthes
- anguim L .
DIPSACEAE : Dipsacus sp,, Scabwsa atropurpurea L. , S. japonka
Miq . EUPHORB I ACEAE : hcphorbiu sptendens Bajer.
GERANI ACEAE : Cemniun cumlinianwn L., Pehgoniun hortorwn Bailey.

GRAMINEAE
HY DROPHY LL ACEAE
LAB I ATAE
LEGUMINOSAE
LILIACEAE
LOBELIACEAE
LOGAN I ACEAE
LY mRAC Ep . MALVACEAE
. t
MUSACEAE '
PASSIFLORACEAE
PHYTOLACCACEAE
POLEMONIACEAE
POLYGONACEAE -
P ORTUL AC AC EAE
RANUNCULACEAE
S CROP ULAR I AC EAE
: EuchZuena m e a m Schrad., HoZcus sorghwn L ., SeoaZe cereale L. , Ditiown aestivwn L , , Zea
m y s L. : PhaceZia tanacetifoziu Benth., P. whitlavia Gray,
: CoZeus blwneri Benth., Nepetu cataria L. , Salt&
spZendens Ker . : Cicer arietinun L . , CrotaZariu intermedia Kotschy.,
Lupinus angwtifoZius L. , L. leteus L . , Phaseolus
a m u s Roxb., P. lumtus L . , P. vulgarw L . ,
Piswn sativun L. , PifoZiwn incarmtmn L., V i c i a
faba I,. , Vigna sinensis End1 . : A Z Z i w n c e p L . , Hyacinthus orientalis L., LiZiwn
awrztwn Lindl ., L. candidun., L. formosanwn
Stapf., L. crocewn Chaix., L. Z0ngifLort.m Thunb.,
L. testacewn Lindl. , L. hrzrfisii Carr., L. tri-
grinwn Ker-Gawl . , filip gesneriana L .
: b b e l i a cardimzis L . , L. erinua L . , L. tenuitor
R . Br.
: Buddleia sp. : L y t h u n salicaria L.
: Hibiscus rianihot L . , Lcrtxrtem tmhestr is L .
: Musa cavendishii Lamb. M. paradisiuca Kuntze.,
M, sapbntun Cowan., M. t e x t i l i s Nees.
: &ssi f lom edulis Sims.
: Phytohca decandra L.
: Cilia capituta b u g l . , G. l ini f lora Benth ., PhZm d r m n d i i ~ o o k .
: Fagopyrun esculentta Gaertn., Rheun rhaponticm L.,
Rwnex crispus L.,
: B~tulaoa g m n d i f l m Hook.
: AqrciZegb camdensis L., Detphimwn consolida L.
: Antim?tinwn mjus L. , CymhaLark muralis Gaertn.,
Mey and Scherb, M - h . d ~ 8 mo8cham bug1 . ,

SOLANACEAE
Nemesk strwnosa Benth., Pentstenron sp. , Verbascwn phoenicewn L., Vemnica longifolia L., ZaZuzian-
skya vi l losa Schmlt.
: Atropo belladonna L., BromZZia specwsa Hook.,
Capsicwn annuwn L , , C. frutescens L , , Dztura mebtoides hnal. , D. metal L., D. stmmoniwn L., Hyo6cyamus niger L., Lycopersicon escuzenhm
M i l l . , Nicandra physaZoides (L.) Pers,, Nicotiunu g k w a Grah., N. glutinosa L. , N. b igelovi i Wats.,
N . rust ica L . , N, sanderae Sander, N. sy tves t r i s
Spegaz and Comes, N. tabacwn L. , Petunia hybrida V i l m . , P. v iohcea Lindl . , Physalis alkukengi
L . , P. anguIsrta L, , P. heterophylk Nees., P.
Isrgasoae R . and S. , P. peruviana L . , P. pubescens L., P. s u b g k h t a MacKenzie and Bush., Salpi- gZossis sinwzta Ruiz and Pav., S c h i m t h u a
wisetonemis Low., Solanwn aviculare Frost . ,
S. carolinense L. , S. melongem L. , S. nigrwn
L. , S. n o d i f l o r n Desv., S. tuberoswn L .
THYMELAEACF , : h p h rnezerewn . . L .
TROPAEO LACEAE : Tr.apaeoZun m jua L. , T. p e r e p i m L . ,
UMBELL IFERAE : Anethwn gmveolens L., Anthriscus cerefoziwn Hoffm.,
Apiwn p v e o l e n s L., Azucus camta L. , FoenicuZun
vulgare H i 1 1, Pas t i m a satiua L . , PetroseZinm hortense Hoffm.
V IOLACAEA : V i o k cornuta L.
D.- 1n v i m Properties
The The has thermal inact ivat ion point between 60' and ~o'c, the d i l u t i o n end point between 1:1000 and 1:10,000 and the longevity
i n v i t ro between 3 and 4 days (Gibbs and Harrison, 1970) . However,
d i f f e r e n t i s o l a t e s of t h e v i r u s d i f f e r i n t h e i r i n vitro proper t ies .
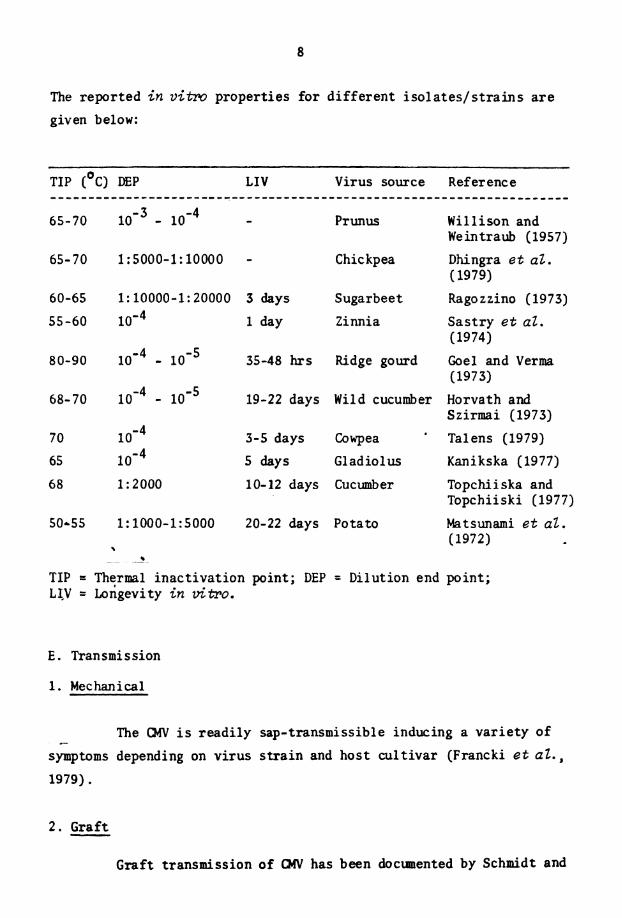
The reported i n v i t m propert ies fo r d i f f e r en t is01 a t e s / s t r a i n s a r e
given below:
TIP (OC) DEP L I V Virus source Reference . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 65-70 10" - 10 - 4 - P r unus Willison and
Weintraub (1957)
65-70 1 : 5000-1: 10000 - Chickpea Dhingra e t aZ. (1979)
60-65 1: 10000-1: 20000 3 days Sugarbeet Ragozzino (1973)
55-60 lom4 1 &Y Zinnia Sas t ry e t aZ. (1974)
80-90 1 0 ' ~ - 10" 35-48 hrs Ridge gourd Gael and Verma (1973)
68-70 lom4 - 10" 19-22 days Wild cucumber Horvath and Szirmai (1973)
7 0 3-5 days Cowpea Talens (1979)
65 l o m 4 5 days Gladiolus Kanikska (1977)
6 8 1 : 2000 10-12 days Cucumber Topchiiska and Topchiiski (1977)
50-55 1: 1000-1:5000 20-22 days Potato Matsunami e t aZ. (1972)
3
TIP = Therml inac t iva t ion point; DEP = Dilution end point; LI,V = I,ongevity i n vitro.
E. Transmission
1. Mechanical
- The UdV i s readi ly sap-transmissible inducing a var ie ty of
symptoms depending on v i rus s t r a i n and host cu l t i va r (Francki e t aZ.,
1979).
2. Graft - Graft transmission of UW has been documented by Schmidt and
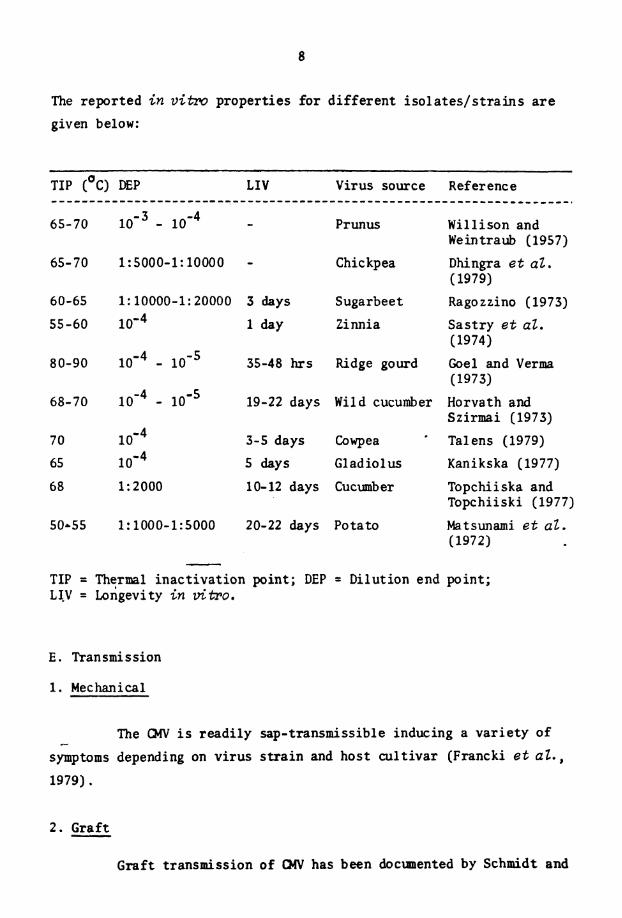
The reported i n v i h w propert ies fo r d i f f e r en t i s o l a t e s / s t r a i n s a r e
given below:
TIP (OC) DEP LIV Virus source Reference
65-70 lom3 - 10 '~ - Prunus Willison and Weintraub (1957)
65-70 1: 5000-1: 10000 - Chickpea Dhingra e t aZ. (1979)
60-65 1: 10000-1: 20000 3 days Sugarbeet Ragozzino (1973)
55-60 10 '~ 1 day Zinnia Sas t ry e t aZ. (1974)
80-90 - 35-48 hrs Ridge gourd Goel and Verma (1973)
68-70 lo'4 - lo1' 19-22 days Wild cucumber Horvath and Szirmai (1973)
70 3-5 days Cowpea Tal ens (1979)
65 5 days Gladiolus Kanikska (1977)
6 8 1: 2000 10-12 days Cucumber Topchiiska and Topchiiski (1977)
50-55 1: 1000-1: 5000 20-22 days Potato Matsunami e t aZ. (1972)
- TIP = Thermal inac t iva t ion point; DEP = Dilution end point; LJV = Longevity i n uitro.
E . Transmission
1. Mechanical
- The i s readi ly sap-transmissible inducing a va r i e ty of
symptoms depending on v i rus s t r a i n and host cu l t i va r (Francki e t at . ,
1979) .
2 . Graf t - Graft transmission of C3.N has been docmented by Schmidt and

Karl (1970) i n case of sann hemp, Jones (1976) i n case o f wineberry,
and Dhingra e t a t . (1979) i n case o f chickpea.
3. Seed - The CMV has not been reported seed t ransmiss ible i n chickpea.
However, seed transmission occurs t o varying degrees i n 18 spec ies
(Neergaard, 1977), including some weed species. The hosts i n which
CMV is seed t ransmiss ible a r e given below with the transmission percent-
ages i n parentheses.
Benincasa hispi& ( I%) , Capsicwn a n n m ( I % ) , Cemstiwn holo-
s t e o 2 e s ( 2 % ) , Cucwnis me20 and C u d s sativus ( t r a c e ) , Ctdc~crbitu
moschata (0.7%), Cuctirbitz pepo and Echinocystis Zobata (9%), h i m
pmpirrewn (4%), Lupinus Zuteus (14%), ~ y c o ~ e r s i c o i escuzentwn (0.2%),
Phaseolus aureus ( 5 % ) , P. vuZgaris ( 7 % ) , SperguZa arwnsis (2%),
Stel laria media (21-40%), Vi'igna cyZindrica cv. k n a r c h , V . sesquipedalb,
V , tolguiculatu (4-28%).
4 . Dodder
Schmel zer (1957) obtained pos i t ive transmission o f CMV through
8 species of Cuscuta, v i z . , C. cqrnpestris, C. cali fomica, C. subinc'lusa,
C. gronovii, C. americuna, C. e m p e a , C. epithymwn, and C . ZupiZi- formis. In chickpea, p o s i t i v e transmission of the v i r u s through
C. r e f l a was obtained by Dhingra e t aZ. (1979).
5 . Insec t - CMV is t ransmiss ible i n a non-persistent manner by a number
o f aphid species . The aphid species involved i n t h e transmission a r e
1 is ted below :

Aphid species
Amphorophoro rhododendri
A. rubi A. sonchi Aphis cardui
A. craccivora
A. euoymi A. fabae A. gossypii
Brevicoryrre brassicae Rzc tyno tus hen& h i
Qsaphis era taegi
~ c h i m c y s t i s lobata
~is-ke+on&z*sktariae
k ros iphwn b m i M.' caZifornicwn M. duphnidis
M. euphorbiae
M. gei M. p i s i M , rosae M. sokmi fo l i i M. sonchifolii &aocaZZis asclepiadia
Virus source
Gladiolus
Raspberry
Gladiolus
Gladiolus
Capsicm amuwn
Chickpea
Pumpkin
Cucumber
Cherry
Cantaloups
Cucumber
Pumpkin
- Cucumber
Cantaloups
Cantaloups
Cherry
Cucumber
Cucumber
- Cantaloups
Gladiolus
Pumpkin
Gladiolus
Cucumber
Gladiolus
Reference
Swenson and Nelson (1959)
Harrison (1959)
Swenson and Nelson (1959)
Swenson and Nelson (1959)
Szalay-Marzso and Solymossy (1961)
Dhingra e t aZ. (1979)
Forghani e t aZ. (1966)
McClanahan and Guyer (1964)
S z a l a y - b r z s o and Solymossy (1961)
Swenson and Marsh (1967)
Goudriet (1962)
Heinze (1959)
Forghani e t aZ. (1966)
Karl (1971)
McClanahan and Guyer (1964)
Goudriet (1962)
Goudriet (1962)
Swenson and Marsh (1967)
Heinze (1959)
McClanahan and Guyer (1964)
Vovk (1956)
Goudriet (1962)
Swenson and Nelson (1959)
Forghani e t aZ. (1966)
Swenson and Nelson (1959)
McClanahan and Guyer (1964)
Swenson and Nelson (1959)

Aphid species Virus source Reference
M. Zythri Cherry Swenson and Marsh (1967)
M, persicae Cherry Swenson and Marsh (1967)
Pen taZonia nigroner uosa Banana Medeiros (1963)
Phorodon cannubis Sann hemp Schmidt and Karl (1970)
RhophaZosiphwn m i d i s Cantaloups Goudriet (1962)
R. pisi Gladiolus Swenson and Nelson (1959)
F. P u r i f i c a t i o n
Several methods f o r p u r i f i c a t i o n of CMV have been used most
of which a r e modifications o f t h e one devised by S c o t t (1963). The
d i f f e r e n t methods used a r e summarized below:
1. The method of S c o t t (1963)
Infected leaves were homgenized i n 0.5 M sodium c i t r a t e ,
pH' 6.5, conta ining 5 mM EDTA and 0.5% th iog lyco l l i c a c i d . Af te r
s q u e e ~ i n p th,rqugh cheesecloth. t h e e x t r a c t was emulsified with ch lo ro l
form. The aqueous phase obtained a f t e r low speed c e n t r i f u g a t i o n was
d ia lysed aga ins t 0.005M borate buffer , pH 9.0 and subjected to t h r e e
cycles of d i f f e r e n t i a l centr i fugat ion.
Tremaine (1968) reported more s a t i s f a c t o r y preparat ion of Y
s t r a i n o f CMV by using S c o t t ' s method.
- Takanami and Tomaru (1969) improved S c o t t ' s method i n which
sephadex gel f i l t r a t i o n and EDTA were used t o avoid aggregation of the
v i r u s .
2 . The method of Lot e t aZ. (19721
They modified Sco t t ' s method i n which v i r u s was p r e c i p i t a t e d

by polyethylene glycol (10%) which was added to the aqueous phase,
obtained a f t e r chloroform emulsification. After incubat ion, it was
centrifuged a t 12,000 g f o r 10 min and the sediment was resuspended
i n 5mM sodium borate buffer , 0.5mM EDTA, pH 9.0, t o which Tri ton X-100
was added t o 2% and s t i r r ed . I t was then subjected t o three cycles
of d i f f e r en t i a l centrifugation. The v i rus was fur ther pur i f ied by
centr i fugat ion i n 5-25% sucrose density gradients (Francki, 1972).
3. The method of Tomlinson e t aZ. (1958)
Infected leaves were homogenized i n O.SM potassium phosphate
buffer , pH 7.5, containing 0.3% thioglycol l ic acid, Clar i f ica t ion of
the f i l t e r e d sap was done by 8.5% n-butanol. The v i rus was recovered
by one cycle of d i f f e r en t i a l centrifugation, resuspending the high
speed p e l l e t s i n 0,05M potassium phsophate buffer , pH 7.5.
Tomlinson e t aZ. (1959) used the same method a s above, but
fu r the r pur i f ied the virus by prec ip i ta t ion with 10% a c e t i c acid
followeQ by fur ther d i f f e r en t i a l centrifugation.
4. The method of Shohara and Osaki (1975)
They purif ied CMV by repeated prec ip i ta t ions with 8% PEG
and 0.2M NaC1, followed by PEG s o l u b i l i t gradient (0-6% PEG s t ab i l i zed
i n 35-5% sucrose) centrifugation.
5. Other methods
Other methods f o r CMV pur i f ica t ion (Morris e t aZ., 1978;
Mossop e t aZ., 1976; Watemorth and Povish, 1975) involved s l i g h t
modifications of one o r the other of the above methods.
Virus yields varied according to the virus s t r a i n and host

p l a n t used, but over 500 mg virus/kg o f l ea f material have been obtained
i n some ins tances (Francki e t at., 1979).
G . Proper t ies of Virw P a r t i c l e s
1. S t ruc tu re
The CMV p a r t i c l e s are isometric, about 28-30 nm i n diameter
(Francki e t ale , 1966, 1979; Debrot e t a l . , 1974; Betto e t aZ., 1964).
Waterworth and Povish (1975) reported t h a t a s t r a i n of CMV from Ixora
p lan t s (CMV-IX) d i f fe red i n having p a r t i c l e s of 24 nm i n diameter r a t h e r
than 28 nm reported f o r o the r CMV s t r a i n s . P a r t i c l e s of a s t r a i n of
CMV from Prunus measured 35 w (Willison and Weintraub, 1957). Matsunami
e t aZ, (1972) observed CMV p a r t i c l e s from potato to be 33 m i n d ia -
meter. The p a r t i c l e s of CMV a r e known t o d i s rup t when mounted unfixed
i n neutra l phosphotungstate fo r e l ec t ron m i c r o s c o ~ (Francki e t aZ.,
1979). However, p a r t i c l e s remain i n t a c t and contras t well i n uranyl
ace ta te , pH 4.5 (Francki e t aZ., 1966). .+-
2. G e d i d n t a f i ~ n coef f i c i en t ($~OW)
a The p a r t i c l e s of CMV sediment a s a s ing le component with
S20W (Swedbergs) of 98.6 (Francki e t aZ., 1966). A s l i g h t l y higher
sedimentation coef f i c i en t (101s) was reported by Tomlinson e t aZ.
(1959).
0 1% 3 . Extinction coef f i c i en t (E260
- I t has been determined as 5.0 (Francki e t aZ., 1966).
4. Az6o/Az80 r a t i o
I t has been determined a s about 1.7 (Francki e t aZ ., 1979).

5. Buoyant densi ty
I t has been determined a s 1.361 and 1.359 g/ml f o r CMV-S and
CMV-D, respec t ive ly (Lot and Kaper, 1976).
6 . Nucleic ac id
The CMV p a r t i c l e s contain single-stranded r ibonuc le ic ac id ,
about 18% of p a r t i c l e weight (Kaper and Re, 1974). The f a c t t h a t CW
contains a func t iona l ly divided genome has been revealed by Peden and
Symons (1973), who detected four species of RNA.
The exact molecular weights of t h e CMV RNAs i s a s u b j e c t o f
debate. Values, ranging between 1.01-1.35, 0.89-1.21, 0.68-0.93 and
0.33-0.38 x106 da l tons have been obtained f o r RNAs 1, 2, 3 and 4,
r espec t ive ly (Kaper and West, 1972; Peden and Symons, 1973; Marchoux
e t aZ., 1973). However, e lec t rophore t i c mobi l i t i e s o f RNA species
may d i f f e r s l i g h t l y between s t r a i n s (Hanada and Tochihara, 1980;
b s s o p , unpublished da ta c i t e d by Francki e t aZ., 1979).
. . A . sqal l , r ep l i ca t ing , single-stranded s a t e l l i t e RNA o f ab&t 5 1 x10 dal tons has been i s o l a t e d i n some s t r a i n s o f CMV (Mossop and
Francki, 1977; Could e t aZ., 1978). One of these, c a l l e d CARNA 5
(cucumber mosaic v i r u s associa ted RNA 5) appeares t o be responsible
f o r l e t h a l necros i s o f tomato (Kaper and Waterworth, 1977).
7 . Pro te in
- The CMV can be read i ly disrupted i n t h e presence of sodium
dodecyl sulphate (SDS) (Kaper, 1975). Analysis of SDS-treated v i r a l
p ro te in i n polyacrylamide gel e lec t rophores i s revealed t h a t t h e
p r o t e i n consis ted o f a s i n g l e polypeptide o f 24,500 da l tons (Habil i and
Francki, 1974) t o 25,200 da l tons (van Regenmortel e t a t . , 1972).

H. Serology
The immunogenicity o f t h e v i r u s is poor but , can be enhanced
by f i x a t i o n wi th formaldehyde (Francki and Habi l i , 1972; Habi l i and
Francki, 1975). The v i r u s p r e c i p i t a t e s on exposure t o physiological
sa l t so lu t ions and, therefore , se ro log ica l t e s t s a r e usua l ly done by
agarose double di f fus ion t e s t s i n buf fe r s o f low molar i ty o r i n water
(Francki e t aZ., 1966; Sco t t , 1968). Unstabil ized v i r u s may give one
o r two p r e c i p i t i n l i n e s , one near t h e ant igen wel l , caused by more o r
1 ess i n t a c t v i r u s and a second one near t h e antiserum well caused by
degraded v i r u s (Scott , 1968; Devergne and Cardin, 1970).
In i t s physical and chemical proper t ies , CMV has a f f i n i t i e s
with tomato aspenny v i r u s (TAV) and peanut s t u n t v i r u s (PSV) (Francki
e t aZ., 1979). More re l i ance has been placed on t h e r e s u l t s o f
se ro log ica l t e s t s than any o ther s ing le c r i t e r i o n t o t r a c e the degree
of r e l a t i o n s h i p s among the th ree v i ruses (Habil i and Francki, 1975) . Thorough serological ana lys i s o f CMV s t r a i n s indicated t h e presence of
a t l e a s t two serogroups both of which a r e se ro log ica l ly r e l a t e d t o
sohe' st;aina b f TAV and PSV (Devergne and Cardin, 1975). In c o n t r a i t ,
Hab i l i and Francki (1975) f a i l e d t o de tec t any serological r e l a t i o n s h i p
between t h e Q-CMV (Queensland s t r a i n of CMV) and V-TAV (Vic to r ia s t r a i n
of TAV). The lack of agreement on the serological r e l a t i o n s h i p between
TAV and W has been probably due t o the use o f d i f f e r e n t s t r a i n s o f
t h e v i ruses which appear t o be very var iable , and t o the l ack o f h igh
t i t r e d a n t i s e r a due t o the poor immunogenicity of t h e v i ruses (Rao,
1982). However, Rao (1982) showed by agar double-diffusion t e s t and
ELISA, a d i s t a n t serological r e l a t i o n s h i p between s i x s t r a i n s o f CMV
and two of TAV.
I . S t r a i n s
Many symptom var ian t s o f W a r e known t o occur making t h e

v i r u s o f ten d i f f i c u l t t o i d e n t i f y from symptoms alone. S t r a i n s well
character ized by t h e i r b iological proper t ies include t h e Y s t r a i n
which i n f e c t s cowpea systemically (Price, 1934), t h e spinach s t r a i n
(Bhargava, 1951) i s o l a t e d from ch loro t i c and deformed leaves o f
spinach, which produces l o c a l l e s ions i n several hosts , and M s t r a i n
o r i g i n a l l y i so la ted from Pr ice ' s (1934) yellow s t r a i n which induces
severe ch lo ros i s i n several p lan t species (Mossop e t aZ., 1976) . The
(5N s t r a i n s characterized physico-chemically include the Y s t r a i n
(Kaper e t aZ., 1965), the Q s t r a i n (Francki e t aZ., 1966; Habi l i and
Francki, 1974) and the S s t r a i n (van Regenmrtel e t aZ, , 1972).
However, o the r s t r a i n s reported a re :
r CMV-Ixora (Watemorth and Povish, 1975) which d i f fe red
from o ther s t r a i n s i n not inducing f e r n l e a f symptoms i n
Lycopersicon escuZentwn cv. Rutgers. I t a l s o produced a
few la rge ch lo ro t i c spots i n C u c k s s a t i v u s r a t h e r than
in fec t ing systemically.
r CMV-Comelina s t r a i n (Dool i t t le and Webb, 1960).
CMV-Cowpea s t r a i n (Anderson, 1955a) .
r CMV-Easter l i l y s t r a i n (Brierley and Travis, 1958).
CMV-Prunus s t r a i n (Willison and Weintraub, 1957)
a. Yield Losses
Kaiser e t aZ. (1968) i n a f i e l d inoculation t r i a l i n chickpea
wi th CMV reported a y i e l d l o s s of 52% due t o in fec t ion . 'Ihe y ie ld
losses depended on the s t age of p lan t in fec t ion . The v i rus caused
about 98% and 87% decrease i n chickpea y ie lds when p lan t s were inocu-
l a t e d a t 4 weeks (pre-bloom) and 9 weeks ( f u l l bloom) a f t e r p lant ing,
respect ively (Kaiser and Danesh, 1971b) .

K. Screening o f Germplasm
There is no repor t i n l i t e r a t u r e regarding screening o f
chickpea germplasm agains t CMV.
111. BE&' YELLOW MOSAIC VIRUS (BYMV)
A. F i r s t Description of the Virus
Pierce (1933), while working on the r e l a t i v e r e s i s t a n c e o f
bean v a r i e t i e s t o common bean mosaic (Bean v i rus I ) , i s o l a t e d another
v i r u s from a p lan t o f t h e Red Valentine va r ie ty . I t s s p e c i f i c i t y was
f i r s t suspected no t only from the f a c t t h a t it produced symptoms
d i s t i n c t from t h a t of the common bean mosaic v i rus , but a l s o because
it a f fec ted the v a r i e t i e s - Corbet Refugee, Great 'Northern U1 No.1
and Robust, which a re r e s i s t a n t t o common mosaic. The v i rus was
designated a s bean v i rus 2 (Bean yellow mosaic v i r u s ) . Occurrence o f
BYMV on chickpea which caused yellowing was f i r s t reported by Erwin
(1958), and Erwin and Snyder (1958) . However, they a t t r i b u t e d t h e
diskage t o othkr v i ruses a l s o ; . The f i r s t authent ic evidence on t h e
na tu ra l occurrence of t h i s v i rus on chickpea was provided by Kaiser
and Danesh (1971a). In India, the re i s no repor t on the na tu ra l
occurrence of BYMV on chickpea.
B . Symptoms on Chickpea
..- Kaiser and Danesh (1971a) repor ted two s t r a i n s of BYMV on
chickpea; a v i r u l e n t s t r a i n causing wi l t ing and yellowing, and a l e s s
v i r u l e n t s t r a i n inducing feathery and deformed leaves.
The sca t t e red incidence o f aphid-transmitted v i r u s yellowing
has been reported by Erwin and Snyder (1958) i n the causation o f which

the bean yellow mosaic virus is s t a ted t o be a dominant f ac to r , though
o the r aphid-transmitted viruses, including pea enation mosaic and a l f a l f a
mosaic v i ruses a l s o i n f e c t t h i s host i n Cal i fornia .
?he b r i g h t yellow color o f the leaves and discolourat ion of
the stem t i s s u e s i n the phloem o f p lan t s a f fec ted with a virus-induced
yellows disease was a t t r i b u t e d t o bean yellow m s a i c and poss ibly o the r
v i ruses (Erwin, 1958).
C. Host Range
BYMV, i n i t i a l l y thought t o be r e s t r i c t e d t o leguminous p lan t s ,
i s now known t o i n f e c t several non-leguminous p l a n t s (Bos, 1970a). Host
range o f the v i rus under d i f fe ren t famil ies i s given below,
A 1 ZOACEAE : Tetrugonia expansa Murr . AMARANTHACEAE : Gomphrena globma Murr.
CHENOPODIACEAE : Chenopodiwn m a n t i c o Z o r Coste e t Reyn ., C. quinoa Willd. . .
LEGUMINQSAE : AstragaZus s inkus Thunb . , Cassia t o m L. , Cicer crrietinwn L. , CrotoZaria spec tab i lb Roth., Glycine
nux Merr., Lathyrus odoratus L., Lars esculenta Moench., Lespedeza capitutu Michx., Lwinus aZbus L., L. Zuteus L . , L. pilosus Walt., MeliZotus alba Desr., M. dentatua Pers., M. o f f i c i m l i s Lam.,
PhaseoZus trulgaris L . , P. acutifoziuc Gray, P.
a m u s Roxb., P . coccineus L . , P . c a l w m t u s Roxb.,
Piswn satitrum L., Difo2it.m arvense L., T. compestre
Schreb., T . bbridwn L., T . incarmtwn L. , T.
micheZimrwn Gaud. ,T. prcrtense L. , T. repens L. ,
T. v e r s i c u l o m Savi. , V i C k faba L. , V. hirsutu
Gray, V. narbonenszk L., V . ViZZosa Roth.,
Vim sesquipedalis Wight.

PEDALIACEAE : SesumunindicwnL.
SOLANACEAE : Nicotiana benthamiam Domin.
D. In vitro Proper t i e s
In vi tro p r o p e r t i e s depend g r e a t l y on v i r u s source, t e s t
p l a n t and t e s t condi t ions , The thermal i n a c t i v a t i o n po in t i s u s u a l l y
between 55 and 6 0 ' ~ though i t may range between 50 and 70°c, t h e
d i l u t i o n end po in t usual ly - l om4 , and longevi ty i n v i tro normally
1-2 b u t some times over 7 days (Bos, 1970a).
I n vitro proper t i e s reported f o r some of t h e i s o l a t e s l s t r a i n s
o f t h e v i r u s a r e given below:
TIP(OC) DEP LIV Virus source Reference .-------------------------------------------.
60-70 - 2 days Red c lover Kme e t aZ. (1970)
55-60 lo-' - 2-3 days White beans Tu (1980)
60-62 , - l ow3 1 day Broad bean Shagrun (1973)
1 day Broad bean El-Attar e t a l . (1971)
50-55 1: 3000-1:5000 French bean Tosso and Vidal (19 76)
55-60 1:500-1:1000 1-2 days Broad bean Tho t tapp i l ly e t aZ. (1976)
TIP = Thermal i n a c t i v a t i o n point; DEP = Dilut ion end point ; L I V = Longevity in uitro.
E . Transmission
1. Mechanical
The v i r u s i s r e a d i l y mechanically t r ansmiss ib le (Bos, 1970a)

2 . Graft
No g r a f t transmission of t he v i rus has been reported i n t he
1 i t e r a t u r e .
3. Dodder
Schmelzer (1957) f a i l e d t o transmit BYMV through e i g h t
Cuscuta spp., which could transmit CMV.
4. Seed - The v i rus is not seed-borne i n French bean, but t ransmit ted
t o a small percentage i n Lqinus albus, L. luteus, MeZiZotus atba, Piswn sativwn and V i c k fQba (Bos, 1970a; Neergaard, 1977) .
5. Insect
More than 20 aphid spp. can transmit t he v i rus i n a non-
pe r s i s t en t manner (Kennedy e t a t . , 1962). Different aphid spp. \
re&rted t o 4)ansrnit the viru.s a r e given below:
Aphid species
Acyrthosiphon pis ton
A . sesbaniae
Amphorphom sp.
A . sonchi
Anuraphis t u Z i p e A: baker-i
Aph i s oardui
A . orac&wm
A . f a k e A . gossypi i
Virus source
Chickpea
Chickpea
Broad bean
Broad bean
Broad bean
White clover
Broad bean
Chickpea
Broad bean
Broad bean
Reference
Kaiser and Danesh (1971b)
Kaiser and Danesh (1971b)
Swenson (1957)
Swenson (1957)
Swenson (1957)
Manglitz and Kreitlow (1960)
Swenson (1957)
Kaiser and Danesh (1971b)
Kaiser e t aZ. (1971)
Kaiser e t aZ. (1971)

Aphid species
A. helianthi A. medimginis
Capi tophor us f iagaefotii HyaZopterus a tr ipz ic i s hhcrosiphwn barri
M. g r a m r i m M . rosae M. e,>ZanifoZii
Megowrz d c i u e Myzus c e m s i M. certus
M. persicae M . sotani
Virus source
Broad bean
Lupins
Broad bean
Broad bean
Broad bean
Broad bean
Broad bean
Broad bean
Broad bean
Broad bean
Broad bean
Broad bean
Broad bean
Reference
Kaiser e t aZ. (1971)
Corbett (1958)
Swenson (1957)
Swenson (1957)
Swenson (1957)
Swenson C1957)
Swenson (1957)
Swenson (1957)
Tapio (1964)
Swenson (1957)
Swenson (1957)
Swenson (1954)
Swenson (1954)
&zus persicae was found t o be the most e f f i c i e n t vector
(Swenson, 1957; Aldez, 1959), followed by Mzcrosiphwn p is i and
~ n k z ~ h i s tuZipae (Swenson, 1957) . Anuraphis sonchi was found t o be
very i n e f f i c i e n t vector whereas Brev-icorp bmssicae f a i l e d t o
transmit , t h i s v i rus (Frandsen, 1952; Swenson, 1954; Swenson, 1957) .
F. Pur i f i ca t ion
Viruses of the potato Y group, to which BYMV belongs, a r e
c h a r a c t e r i s t i c a l l y d i f f i c u l t t o manipulate i n vi tro because o f t h e i r
tendency t o aggregate, both end-to-end and l a t e r a l l y (Shepherd and
Po-md, 1960). However, various methods have been followed s a t i s f a c t o r i l y
f o r pur i f i ca t ion of BYMV which a r e given below:
1. The method of Uyeda e t aZ. (1975)
They described pur i f i ca t ion of i s o l a t e no. 30 of BYMV
propagated i n broad bean. Homogenization of frozen leaves was done

i n 0.1M Tris-HC1 buffer, pH 7.0, containing 0.05M EDTA and 1% 2-Mercapto-
ethanol. After passing through cheesecloth, the e x t r a c t was emulsified
with 4 volume of carbon te t rachlor ide . 'Ihe v i rus was recovered from
aqueous phase by precipi ta t ion with PEG (4%), followed by d i f f e r e n t i a l
centr i fugat ion. The v i r u s pe l l e t s were suspended i n 0.01M phosphate
buffer . To avoid aggregation, 0.5M urea was added t o t h e suspending
medium. Further pur i f i ca t ion was achieved by 10-40% sucrose dens i ty
gradient centr i fugat ion.
2, The method of Huttinga (1973)
He pur i f ied BYMV by applying moderate centr i fugal fo rces .
Infected pea leaves were homogenized i n 0.1M t r i s - t h i o g l y c o l l i c ac id ,
pH 9.0, along with carbon te t rachlor ide and chloroform (40 m l each
f o r 150 m l of buf fe r ) . The virus was concentrated from aqueous phase
by applying 2 cycles of d i f f e r e n t i a l centr i fugat ion (1.5 h r a t 26,500 g
and 10 min a t 8000 g) . The virus was suspended i n 0.lM tris-HC1 buf fe r U'
pH 9.0. The low centr i fugal forces yielded unaggregated v i r u s with
neg l ig ib le breakage. On the contrary, higher centr i fugal fo rces
proguced %consi$erable amounts of broken p a r t i c l e s . * . .
3.- The method of Granett and Prowident i (1975)
They pur i f ied two s t ra ins , v i i . , BYMV' and BYMY' by using
a modification of Hut t ingats method. Each gram of c h i l l e d leaves was
homozenized i n 7.5 ml of 0.1M tris buffer , 2 ml each of carbon t e t r a -
chlor ide and chloroform. The pH of the e x t r a c t was adjus ted with
th iog lyco l l i c a c i d t o 6.5 f o r BYW' and 8 .5 fo r BYMV'. After breaking - the emulsion, the v i r u s was recovered from the aqueous phase by c e n t r i -
fugat ion a t 15,000 rpm f o r 1 hr. The p e l l e t was suspended i n ex t rac t ing
buffer and c l a r i f i e d a t 5000 r p m f o r 5 min. Higher cen t r i fuga l fo rces
during e i t h e r i n i t i a l c l a r i f i c a t i o n o r pe l l e t ing increased p a r t i c l e
aggregation and breakage, and reduced v i rus y ie ld .

4. The method of Uyemoto e t aZ. (1972)
They pur i f i ed BYMV and BCMV by the method followed by
Damirdagh and Shepherd (1970) f o r pur i f i ca t ion of tobacco e tch v i r u s .
One gram of infected t i s s u e was homogenized with 2 ml of 0.5M potassium
phosphate buffer , pH 7.0, containing 1% 2-Mercaptoethanol . After
s t r a i n i n g through cheesecloth the homogenate was s to red overnight a t
8 ' ~ i n butanol (8 m l per 100 ml). After c l a r i f i c a t i o n by c e n t r i -
fugation a t 8,500 rpm f o r 10 min, the v i rus was recovered from super-
natant with 4% PEG followed by d i f f e r e n t i a l centr i fugat ion (28,000 rpm
f o r 1 .S hr; 8,500 rpm f o r 10 min, suspending the v i r u s i n 0.025M
phosphate buffer , pH 7.0 containing O.5M urea and 0.1% 2-Mercaptoethanol)
Further pur i f i ca t ion was achieved by 10-40% sucrose dens i ty g rad ien t
centr i fugat ion.
G. Proper t ies of Virus P a r t i c l e s
1. Structure
P a r t i c l e s a r e elongated and flexuous, 750nm long and 1Snm \
wid; XBrandes 'and Quant z, 1955;. Brandes, 1964). P a r t i c l e s of g r e a t e r
length have a l s o been repor ted . Bos (1970b), using tobacco mosaic
v i rus a s an in te rna l s i z e standard, found p a r t i c l e s of 660 nm length .
Taylor and Smith (1968) and Bos (1970b) found t h a t p a r t i c l e s from
Chenopodiun amvanticolor were 50-60 nm longer than those from legumes.
Contrar i ly , no influence of host on p a r t i c l e morphology could be
noticed by Moghal and Francki (1981).
2 r Sedimentation coef f i c i en t (S*-jW)
The sedimentation c o e f f i c i e n t of 5 i s o l a t e s of BYMV ranged
between 140 and 145s (Huttinga, 1975).

0 1% 3. Ex t inc t ion c o e f f i c i e n t (E260 limm)
I t was determined as 2.4 (Uyeda e t at., 1975).
4 . A260/A280 r a t i o
The r a t i o f o r several preparat ions , corrected f o r l i g h t
s c a t t e r i n g var ied from 1.24 t o 1.31 wi th an average o f 1.28 (Uyeda
e t aZ., 1975).
5 . Nucleic acid
BYMV has a s i n g l e molecule o f s ingle-s t randed RNA (Matthews, 6 1979) wi th a presumed molecular weight o f about 3.0 x 10 a s f o r
t u r n i p mosaic, tobacco e tch and maize dwarf mosaic v i r u s e s ( H i l l and
Benner, 1976), and pea mosaic v i r u s (Abu-Samah and Randles, 1981) . Lisa e t aZ. (1981) es t imated t h e molecular weight of RNA of a poty-
v i r u s , which caused mosaic on ~Cucurbitu pepo and which showed s l i g h t
s e r o l o g i c a l r e l a t i o n wi th BYMV, t o be 2.93 x lo6 d a l t o n s .
6 . P r o t e i n
Huttinga (1975) determined t h e molecular weight o f c o a t
p r o t e i n of 5 i s o l a t e s of BYMV a s 34,000 daltons. Moghal and Francki
(1976) repor ted a molecular weight o f 33,000 da l tons f o r 3 i s o l a t e s
of BYMV. A s l i g h t l y higher molecular weight of 35,000 da l tons has
been repor ted f o r t h e BYMV i s o l a t e I of D r . D.J . Hagedorn, Univers i ty
o f Wisconsin (Uyemoto e t a t . , 1972). Y i l m z (1979) determined t h e
d e c u l a r weight of coat p r o t e i n of BYMV from broad bean a s 33,350 - 33,820 d a l t o n s .

H . Serology
Antisera t o BYMV with a t i t r e o f 112048 have been produced
(Bos, 1970a). These could be f u r t h e r increased by chemically concen-
t r a t i n g the globul in f r a c t i o n o f t h e se ra giving t i t r e s up t o 1/256,000
(Bercks, 1960; Bercks, 1961), but t h i s i s exceptional. Uyeda e t aZ.
(1975) obtained a n t i s e r a t o 3 s t r a i n s of BYMV with t i t r e s ranging from
1024-2048.
Microprecipit in and p r e c i p i t i n tube t e s t s a r e mostly used
because i n t a c t v i r u s p a r t i c l e s do no t d i f fuse i n agar g e l s . However,
t h e d i f fus ion r a t e of filamentous v i ruses through agar could be
increased by var ious detergents (Hamilton, 1964; Gooding and Bing,
1970; Purci ful and Batchelor, 1977) o r physical treatments (Tomlinson
and Wal key, 1967) which allowed degradation o f v izus p a r t i c l e s i n t o
r e a d i l y d i f f u s a b l e un i t s .
Granett and Provvidenti (1975) found t h a t 0.3% (w/v) o f SDS
i s ' good f o r degradation of BYMV i n t o d i f fusab le u n i t s than pyrrolidone
o r ethanblamiqe.
BYMV i s se ro log ica l ly d i s t a n t l y r e l a t e d t o bean common
mosaic v i r u s (Beemster and van der Want, 1951; Moghal and Francki,
1976), soyabean mosaic, sugar beet mosaic, watermelon mosaic v i ruses ,
and pota to v i r u s Y (Bercks, 1960) .
I . S t r a i n s - Various s t r a i n s d i f f e r i n g i n t h e symptoms they produce i n
c e r t a i n p l a n t species have been recognised. They a re :
Pea mosaic s t r a i n : Pea mosaic v i r u s ( D o o l i t t l e and
Jones, 1925) was f o r long considered a separa te v i r u s ,

d i f f e r i n g from BYMV by the b r i g h t yellow symptoms it causes
i n pea and broad bean, and by i t s i n a b i l i t y t o i n f e c t most
v a r i e t i e s of PhaseoZus vuZgaris (Schroeder and Provviden ti,
1966; Cousin, 1969). However, based on t h e i r c l o s e se ro-
l og i ca l r e l a t i onsh ip , i t was concluded t h a t pea mosaic
v i r u s was a s t r a i n of BYMV (Taylor and Smith, 1968).
Cowpea s t r a i n of Anderson (1955b) and Br i e r ly and Smith
(1962), i s o l a t e d from and produces systemic symptoms i n
V C g m sinemi4 which i s no t i n fec t ed by common s t r a i n o f
the v i r u s . The cowpea s t r a i n g ives severe systemic yellow
spo t t i ng i n Chenopodim amranticolor.
Red c lover necros i s s t r a i n of Zaumeyer and Goth (1963),
produced necros i s i n red c lover and pea, l oca l and systemic
necros i s i n C. amant-icoZor, death of DifoZiwn incarnatwn, T. subterranewn and Vicia faba-minor.
Pod d i s t o r t i n g s t r a i n of Grogan and Walker (1948), i s o l a t e d
from Idaho Refugee bean and o the r v a r i e t i e s r e s i s t a n t t~
common bean mosaic v i r u s .
Black r o o t o r X-disease s t r a i n o f McWhorter and Boyle
(1946) which produces purpl ing of t h e l e a f bases o f t h e
lower leaves . This purpl ing i s accompanied by dying of
the t i s s u e s within and o f t en leads t o prena ture dea th of
t he p l an t .
Necrotic l e s i o n s t r a i n : A l o c a l 1 esion-producing v i r u s
i s o l a t e d from in fec t ed bean pods by Zaumeyer and Fisher
(1953) . Top necros i s , produced by t h e pod d i s t o r t i n g and
X-disease s t r a i n s seldom develops with t h i s s t r a i n :
PhaseoZus Zunatus, V'igna sesquipedaZis and V. s inens i s

are suscept ib le t o the nec ro t i c l e s ion s t r a i n , but a r e
r e s i s t a n t t o o ther s t r a i n s of BYMV.
Severe yellow mosaic s t r a i n of Rex and Zaumeyer (1953)
which causes strong epinasty, ch lo ros i s and ve ina l
necros is of inoculated primary leaves of P. vulgaris .
These symptoms are followed by top necros is and death
of c e r t a i n bean v a r i e t i e s and severe yellow mosaic i n
o thers . No infect ion was obtained ou t s ide t h e Leguminosae,
Other s t r a i n s of BYMV reported are : BYMV-Q from Canna sp .
i n Queensland (Moghal and Francki, 1976), BYMV-G i s o l a t e d from
GZadwZw connntazis i n South Austral ia (Lee e t aZ., 1979) and BYMV-S
from V i c k faba i n South Austral ia (Randles e t aZ., 1980).
J. Yield Losses
Kaiser e t aZ., (1968), i n a f i e l d inocula t ion t r i a l s wi th
BYMV r e p ~ r t e d a y ie ld l o s s of 77-92% due to in fec t ion i n chickpeas. . . .
' Kaiser and Danesh (1971b) reported about 92-100% and 84-85%
decrease i n chickpea y ie lds when p lan t s were inoculated a t 4 weeks
(pre-bloom) and 9 weeks ( f u l l bloom) a f t e r planting, r e spec t ive ly .
K. Screening of Germplasm
There i s no r e p o r t i n the l i t e r a t u r e on the screening of
chickpea germplasm f o r r e s i s t ance t o BYMV.

MATERIALS A N D METHODS
I . GENERAL
A. Virus Source
Cucumber mosaic v i rus (CMV) was i so la ted from f i e l d s of
chickpea genetic resources uni t , ICRISAT, from the p lan ts showing t i p
necrosis . Bean yellow mosaic v i rus (BYMV) was i so la ted from f i e l d s
of the Agricultural College Farm, Parbhani, Maharashtra from p lan t s
showing the narrow leaf symptoms.
B. Maintenance of Viruses
Both the viruses were maintained i n the glasshouse on chick-
pea by periodic mechanical inoculations. A kabuli cu l t i va r obtained
from the loca l market and a kabuli germplasm accession, ICC-162, were
used f o r the maintenance of CMV and BYMV, respect ively. Besides chick-
pea, W was maintained on Nicoticzm rustica var . White pathar ,
obtained' f rop She Central Tobacco Research I n s t i t u t e , Rajahmundry,
Andhra Pradesh, India, and BYMV on PhaseoZus vuZgaris var . Dwarf
obtained from the local market.
C. Plants
A l l chickpea and other t e s t plants were raised i n a mixture
of Alf isol and farmyard manure (1:4, w/w) i n 15 cm pots i n the glass-
&e (temperature range 25-30'~).
D. Inoculum and Inoculations
The inoculum was prepared by grinding the infected chickpea
leaves w i t h a pre-chi l led pest le and mortar i n 0.05 M potassium

phosphate buffer , pH 7.5, containing 0.01M sodium s u l p h i t e and 0.01M
sodium d i e t h y l dithiocarbamate (4 m l per gram t i s s u e ) . For determining
physical p r o p e r t i e s of t h e v i ru ses , t he standard e x t r a c t was prepared
by grinding t h e i n f ec t ed chickpea leaves i n t h e same bu f f e r a s above
(1: 1, w/v) but without any add i t i ve s . C e l i t e ( I%, w/v) was added t o
t he e x t r a c t and rubbed onto t h e leaves of t e s t p l a n t s wi th t h e broad
end of t he p e s t l e , Inoculated p l a n t s were thoroughly r i n sed with t a p
water . All chickpea p l a n t s were inoculated when 10 days o ld , whereas
o t h e r p l a n t s were inocula ted when young.
11. IDENTIFICATION OF VIRUSES BY SEROLOGY
The v i r u s e s under s tudy were i d e n t i f i e d by serology employing
double- d i f f u s i o n t e s t i n agar (Ball , 1974) and enzyme-linked imrnuno-
sorbent a s say (ELISA) ( L i s t e r , 1978; Reddy e t a Z . , 1981).
A: lbublc+ Diffusion i n Agar
Agarose 0.5% (Sigma chemicals, U.S,A.) was prepared i n 0.01M
potassium phosphate bu f f e r , pH 7.0, t o which 0.15M sodium c h l o r i d e was
added. I t was then autoclaved a t 15 l b p s i f o r 15 minutes and 0.05%
Mer th io la te (Ethyl mercu r i t h io sa l i cy l i c ac id) was added t o a r r e s t
microbial contamination. Eight m l o f agar were poured i n t o p l a s t i c P e t r i
p l a t e s (5.3 cm diameter) . Af te r s o l i d i f i c a t i o n , w e l l s o f 7 mm
diameter were c u t with a cork borer . The p a t t e r n cons i s t ed of 4
pe r iphe ra l we l l s around a c e n t r a l well (5 nun d i s t ance from c e n t r a l w e l l ) .
Ant i sera t o W and BYMV were obtained from Dr. H.A, Sco t t , Univers i ty
o f Arkansas, U.S.A., and Dr. N. Conti, Laboratorio F i tov i ro log ia , Torino,
I t a l y , r e spec t ive ly . P u r i f i e d v i r u s and sap from in fec t ed chickpea
were used a s an t igens . Healthy sap and bu f f e r were kept a s c o n t r o l s .
The p l a t e s were incubated i n a mist chamber (des icca tor w i th moist

cot ton i n the bottom) a t room temperature (25-28'~) and observed d a i l y
f o r r eac t ion .
I n the case of BYMV, t he v i r u s was t r e a t e d wi th 0.3% sodium
dodecyl s u l f a t e before incorporating i n t o wel ls , t o break t h e v i r u s
i n t o d i f fusab le u n i t s (Purc i fu l l and Batchelor, 1977) .
B. Enzyme-Linked Immunosorbent Assay (ELISA)
1. Extract ion of y-globulins
One m l of antiserum was d i l u t e d with 2 ml o f phosphate
buffered s a l i n e (PBS) pH 7 . 4 (0.8% NaC1, 0.005M Na2HP04, 0.001M KH2P04,
0.02% K C 1 , 0.02% osdium azide) and mixed gent ly .
To t h e above, 3 m l of 36% sodium s u l f a t e were added, mixed
gen t ly and centr i fuged a t 5000 rpm f o r 15 min.
The p e l l e t was dissolved i n 4 m l of 18% sodium s u l f a t e , and
centr i fuged gt 5000 rpm f o r 15 .minutes.
The p e l l e t was resuspended i n 2 m l of PBS.
Dia lys i s was done agains t PBS i n r e f r i g e r a t o r a t about 4 ' ~
g iving two changes of 1 l i t r e each ( f i r s t d i a l y s i s was f o r 4 hours and
second overnight ) . The y-globulin so lu t ion was remove from t h e d i a l y s i s bag.
The concentrat ion of y-globulins was adjus ted t o 1 mg/ml, assuming an
Oo1' = 1.6). ex t inc t ion c o e f f i c i e n t of 1.6 OD (AZIO
2. Con jugat ion of y-globulins with a l k a l i n e phosphatase enzyme
The enzyme a l k a l i n e phosphatase (Sigma P-4502, type VII,

5 mg/ml) was p r e c i p i t a t e d from enzyme so lu t ion by cen t r i fug ing a t
5000 rpm f o r 15 minutes, and dissolved i n y-globulin s o l u t i o n (1.5 mg
o f enzyme per 1 nil of 1 mg/ml y-globulin so lu t ion) .
Dia lys is of conjugated y-globulins was done a g a i n s t PBS with
two changes of 1 l i t r e each ( f i r s t d i a l y s i s f o r 4 hours and second
overnight) .
Conjugated y-globulins were t r e a t e d with g lutara ldehyde by
d ia lys ing aga ins t PBS containing 0.06% glutaraldehyde f o r 1 hour a t
room temperature.
Dia lys is bags were t r ans fe r red t o PBS and dia lysed overnight ,
a t ~ O C , giving two changes of 1 l i t r e each.
Conjugated y-globul ins were remved and s to red i n g l a s s tubes
a t doc.
3. Preparation of p l a t e s \
a I ' . .
Micro t i t r e p l a t e s (Dynatech labs . , England) were demarcated
according t o the var ious d i l u t i o n s o f t e s t so lu t ions , l eav ing o u t
per iphera l we l l s .
y-globul in d i l u t i o n s were made i n coat ing buf fe r (0.02M
Na2C03, 0,02M NaHC03, 0.02% sodium azide , pH 9.6) and 0.2 m l was
added t o each well o f t h e p l a t e . Water was added t o the pe r iphera l
v e l l s and t h e p l a t e s were incubated a t 3 7 ' ~ f o r 2% hours.
y-globulins were poured o f f and t h e we l l s were washed t h r i c e
with PBS containing 0.05% Tween-20 (PBS-Tween) leaving the b u f f e r i n
w e l l s f o r 3 minutes f o r each washing.

Dilutions of t e s t samples (healthy and infected) were made by
grinding the t i s s u e i n antigen ext ract ion buffer (PBS-Tween containing
2% polyvinyl pyrrolidone (PVP) of mol. w t . 40,000) and s t r a i n i n g t h e
e x t r a c t through cheesecloth.
Test samples (0 .2 ml) were added t o each well and water t o
per iphera l wells . P la tes were incubated overnight a t 4 ' ~ .
Test samples were poured o f f and t h e wells were washed a s
e a r l i e r i n PBS-Tween.
Dilutions o f conjugated y-globulins were made i n conjugate
buf fe r (PBS-Tween containing 2% PVP; 0.2% ovalbumin).
Conjugated y-globulins (0.2 ml) were added t o each well and
water t o peripheral we l l s , P la tes were incubated a t 3 7 ' ~ f o r 3 hours.
y-globulins were poured o f f and t h e wel ls were washed i n
PBS- ween a s e a r l i e r .
The s u b s t r a t e (0.2 ml), p-nitrophenyl phosphate (1 mg/ml),
prepared i n subs t ra te buf fe r (9.7% aqueous diethanolamine, pH 9.8)
was added t o each well. P la tes were incubated f o r 30-45 minutes a t
room temperature.
One drop of 3M NaOH was added t o each well t o s t o p f u r t h e r
r eac t ion . A p o s i t i v e react ion was detected by the development of
yellow colour i n the subs t ra te .
111. HOST RANGE AND SYMPTOMS
Host range s tud ies were c a r r i e d out f o r CTW and BYMV, by
mechanical inocula t ion of d i f f e r e n t p lant species belonging t o the

families: Aizoaceae, Amaranthaceae, Chenopodiaceae, Compositae, Cucur-
b i taceae , Leguminosae and Solanaceae ra i sed under i n s e c t - f r e e glasshouse
condi t ions . Four p lan t s were inoculated with each v i r u s and an equal
number of p l a n t s were kept a s controls . P lan t s were observed d a i l y and
symptoms recorded a s and when they appeared.
IV. PHYSICAL PROPERTIES IN CRUDE SAP
A. Cucumber Mosaic Virus
Physical p roper t i e s of CMV were determined i n the standard
e x t r a c t following the procedure described by Bos e t aZ. (1960).
Chenopodiun m a n t i c o l o r was used a s an assay host .
1. Thermal inac t iva t ion point
Five m l samples o f the standard e x t r a c t i n t e s t tubes were
subjected separa te ly f o r 10 minutes t o heat treatments s t a r t i n g from
45' and 9 0 ' ~ with an i n t e r v a l of 5 ' ~ . Immediately a f t e r heat treatment,
the. sap Gas cooled by passing . t a p water over the outer surface o f t h e
tube. To t h i s was added c e l i t e (1%) and rubbed on the leaves o f the
assay host . Ten C. amrant icolor plants , each with 6 leaves were used
r e p l i c a t i n g each treatment 6 times (Table 1) . Observations were
recorded 3 days a f t e r inocula t ion, when the loca l l e s ions were conspi-
cuous i n control treatment.
2 . Dilution end m i n t
The standard e x t r a c t was d i lu ted t o ge t d i l u t i o n s varying
from 10-I through lom7. After adding c e l i t e to each, they were
separa te ly rubbed on the leaves of the assay host . Eight C. amrant i -
coZor plan t s , each w i t h 6 leaves were used and each treatment was
r e p l i c a t e d 6 times (Table 2 ) . Observations were recorded 3 days a f t e r
inocula t ion.

TABLE 1. Randomization of treatments t o determine thermal inac t iva t ion point (TIP)
Plant No.
1 2 3 4 5 6 7 8 9 1 0 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
6 l a g c i e j f b h d
& s 2 b h d j f a g c i e 0
N 0 ~ 3 c i e a g b h d j f +
j 8 4 d j f b h c i e a g c o
a = control ; b = 45'~; c = SOOC; d = 55'~; e = 60'~; f = 65'~; g = 70'~; h = 75'~; i = 80°c; j = 85'~.
TABLE 2 . Randomization of treatments t o determine d i l u t i o n end point , (DEP)
Plant No.
8 = control (undiluted); b = 1 0 c = d = 10'~; e =
f = 10-5; = 10-6; t, =

3 . Longevity i n tritro
The standard e x t r a c t was s tored i n a 50 m l beaker a t room
temperature (22'- 24'~) and was inoculated d a i l y on assay host , u n t i l
the sap was confirmed t o be non-infectious.
B . Bean Yellow Mosaic Virus
The preparation o f the standard e x t r a c t and the treatments
were s imi la r a s described f o r CMV. Since no loca l l e s ion host could
be found f o r t h i s v i rus , the assay was done on kabuli chickpea
(ICC-162), taking 10 seedlings f o r each treatment.
V. INSECT TRANSMISSION
Two aphid species, v i z . , Aphis crvrccivora and 4 jzus persicae,
were obtained from f i e l d grown pigeonpea and tobacco, r e spec t ive ly
and healthy cqlonies were ra i sed before using them i n transmission
s tud ies .
Aphids were fas ted i n P e t r i p l a t e s f o r an hour and then
allowed an aqu i s i t ion feeding period of one minute on vi rus- infected
chickpea leaves. Thereafter they were t r ans fe r red onto the healthy t e s t
p l a n t s f o r inocula t ion feeding period of 12-14 hours. Thereafter , the
insec t s were k i l l e d by spraying Rogor (0.02%). Five t o 10 aphids were
used per p lan t and 10-day-old healthy chickpea seedlings (10) were used
a s t e s t p lants .
V I . PURIFICATION OF VIRUSES
A. Cucumber Mosaic Virus
Chickpea was used a s a source p lan t s ince it supported a

high v i r u s concentrat ion. Plants were harvested 7-10 days a f t e r inocu-
l a t i o n , when they showed terminal drooping. The e n t i r e p lan t , except
roo t s , was used for pur i f i ca t ion . The p u r i f i c a t i o n procedure developed
by Tomlinson e t a'L. (1958) was used with modifications which i s given
below. A l l the s teps of pur i f i ca t ion procedure were c a r r i e d out a t
about 4 ' ~ by using pre-chi l led mater ia ls and keeping the prepara t ions
i n an i c e bucket.
Infected chickpea p lan t s were homogenized using a pre-chi l led
Waring blendor i n 0.5M potassium phosphate buffer , pH 7.5 containing
0.1% sodium th iog lyco l l i c ac id and ImM EDTA (4 ml/g t i s s u e ) . The sap
was s t ra ined through 2 l aye rs of cheesecloth. To the f i l t r a t e 8.5%
(v/v) of n-butanol was added dropwise with constant s t i r r i n g . S t i r -
r i n g was done f o r another 15 minutes. Then the mixture was c e n t r i -
fuged a t 5000 rpm f o r 10 minutes i n SS-34 r o t o r i n Sorval l centr i fuge.
The supernatant was saved t o which 8% polyethylene glycol (PEG) mol.
w t . 6000 (Carbowax, Fisher S c i e n t i f i c Company, U. S .A. ) and 0.2M sodium
chlor ide were added and dissolved f o r 30 minutes and incubated f o r 90
minutes a t 4 ' ~ . I t was then centrifuged a t 10,000 rpm f o r 10 minutes
and t h e ' r e s u l t i n g p e l l e t was resuspended i n hal f the o r ig ina l volume
of 0.025M potassium phosphate buffer pH 7.5, containing 1mM EDTA ( P E ) .
The p e l l e t was thoroughly dissolved using a vortex mixer and c e n t r i - t
fuged a t 5000 rpm fo r 10 m i n ~ e s . The supernatant was saved and c e n t r i -
fuged a t 30,000 rprn f o r 2 hours i n R-35 r o t o r i n a Beckman prepara t ive
u l t r acen t r i fuge (model L5-50) a t 4 ' ~ . The p e l l e t was resuspended i n
PE and centrifuged a t 5000 rpm f o r 5 minutes. The supernatant was
col lec ted and designated a s p a r t i a l l y p u r i f i e d v i rus .
Further pur i f i ca t ion of the v i r u s was achieved by r a t e - zonal
dens i ty gradient centr i fugat ion i n sucrose. Sucrose gradients were
prepared by layering 9, 9, 9 and 6 ml o f 40, 30, 20 and 10% sucrose
prepared i n PE, respect ively . me gradients were kept a t 4 ' ~ f o r
16 hours before use. Four m l of the p a r t i a l l y p u r i f i e d v i rus preparation
were layered over the gradients and centrifuged i n SW-27 ro to r f o r

90 minutes a t 24,000 rpm. A l i g h t s c a t t e r i n g zone was observed 2.4 cm
below the meniscus when a narrow beam o f l i g h t was passed through the
top of the tube i n a dark room. The zone was co l l ec ted by an ISCO
dens i ty gradient f r ac t iona to r (model 185) with UA-5 absorpt ion monitor.
I t was d i lu ted with an equal volume o f PE and centr i fuged
f o r 2 hours a t 30,000 rpm i n R-35 r o t o r o r 1 hour a t 45,000 rpm i n
R-50 r o t o r . The p e l l e t was dissolved i n 0.5 t o 1 m l of PE and des ig-
nated a s p u r i f i e d v i r u s . A t d i f f e r e n t s t eps of p u r i f i c a t i o n , t h e
i n f e c t i v i t y of the v i rus was assayed by mechanical inocula t ion on
C. amranticolor.
From another gradient , one-ml f r a c t i o n s were co l l ec ted by
the f r a c t i o n a t o r and each f r a c t i o n was separa te ly assayed on C. a m a n t i - color a f t e r d i l u t i n g 10 times, t o c o r r e l a t e t h e W a b s o r p t i o n component
wi th i n f e c t i v i t y . Three leaves were used f o r each f r a c t i o n and the
average number of loca l l e s ions was obtained.
For studying biochemical p roper t i e s of t h e v i rus , f u r t h e r
p u r i f i c a t i o n yas attempted by quasi-equil ibrium dens i ty gradient
cen t r i fuga t ion . The v i r u s zone co l l ec ted a f t e r the rate-zonal d e n s i t y
gradient cen t r i fuga t ion was d i l u t e d wi th PE ( 1 : l ) and layered over
60-30% sucrose gradients , prepared by layer ing 12, 7, 7, and 7 ml of
60, 50, 40, and 30% sucrose i n PE, r e spec t ive ly . After cen t r i fuga t ion
a t 24,000 rpm f o r 2 hours i n SW-27 r o t o r t h e l i g h t s c a t t e r i n g zone
was withdrawn by a bent needle a t tached t o a syr inge. I t was d i l u t e d ,
p e l l e t e d and dissolved i n PE a s described i n t h e rate-zonal dens i ty
g rad ien t cen t r i fuga t ion .
B. Bean Yellow b s a i c Virus
Several methods (Uyeda e t aZ. , 1975; Huttinga, 1973; and
Uyemoto e t aZ., 1972) were used f o r BYMV p u r i f i c a t i o n and the one

described below was found t o be s u i t a b l e . A l l t he s t eps of p u r i f i -
ca t ion procedure were carr ied ou t a t about 4 ' ~ a s i n the case of CMV.
BYMV infected chickpea t i s s u e was homogenized i n 0.1M Tris-PO4
buffer , pH 8.5 containing 0.1% th iog lyco l l i c ac id and 0.01M EDTA (1:3,
w/v). Carbon t e t r a chloride (4 volume of the buffer) was added during
homogenization i n a Waring blendor. After s t r a i n i n g through two-layered
cheesecloth, it was centrifuged a t 5000 rpm f o r 10 minutes. The aqueous
phase was recovered to which 4% PEG was added and incubated overnight
a t 4 ' ~ . Then it was centrifuged a t 10,000 rprn f o r 10 minutes. The
r e s u l t a n t p e l l e t was dissolved i n ha l f the o r i g i n a l volume of 0.01M
Tris-PO4 buffer , pH 8.5 containing 0.2M urea (PU) and centrifuged a t
5000 rprn f o r 5 min. Twenty-five m l of supernatant were layered over
13 m l of a sucrose pad (30% sucrose i n TPU containing 4% PEG) and
centrifuged a t 24,000 rprn f o r 2 hours. The pel le t 'was dissolved i n
TPU and centrifuged a t 5000 rprn f o r 5 minutes.
Four m l of the supernatant ( p a r t i a l l y p u r i f i e d v i rus ) were
layered over the preformed sucrose densi ty gradient column prepared
i n TPU a s in -case of W and centrifuged a t 24,000 rprn f o r 90 min.
An improminent, diffused v i rus zone formed j u s t above the midspoint
was co l l ec ted with a bent needle at tached to a syringe. I t was
d i lu ted with TPU and centrifuged a t 24,000 rprn f o r 90 min. The pel le '
was dissolved i n a small amount of 0.01M Tris-PO4 buffer (pur i f ied
v i rus ) .
/I1 . ELECTRON MICROSCOPY
Electron microscopy of p u r i f i e d preparations of (=MV and BYMV
was done. In addi t ion, e lec t ron microscopy of d i p preparations and
u l t r a t h i n sect ions of BYMV-infected chickpea leaves was a l s o done.

A. CMV-Purif i e d Preparat ion
A drop of p u r i f i e d v i r u s was kept on a paraf i lm on which a
formvar and carbon-coated copper g r i d (300 mesh) was f l o a t e d f o r 2 min,
The g r i d was removed with f i n e forceps , excess s o l u t i o n wiped o f f by
t h e pointed edge of f i l t e r paper and then f l o a t e d on a drop o f 1%
uranyl a c e t a t e f o r one minute. Thereaf ter , it was taken ou t , excess
s t a i n wiped o f f , d r i ed and observed under a P h i l i p s 201C e l e c t r o n
microscope. From negat ive e l e c t r o n photo micrographs, t h e average
diameter o f the v i r u s p a r t i c l e s was ca lcu la ted by measuring 100
p a r t i c l e s , using a Bousch and Lomb magnifier.
B . BYMV-Purified Preparat ion
The procedure was s i m i l a r t o t h a t described f o r CMV, except
t h a t n e u t r a l phosphotungstic ac id was used f o r negat ive s t a i n i n g .
C. Leaf Dip Preparation
The d i p preparat ion was done with BYMV-infected chickpea
leaves s ince i t i s necessary i n case of rod-shaped v i ruses t o measure
t h e p a r t i c l e s i n crude sap. A BYMV-infected chickpea leaf was
chopped i n a drop of water on a paraf i lm. The water drop was removed
with t h e h e l p o f a pas teur p i p e t t e and used f o r t h e p repara t ion o f a
g r i d f o r t h e e l e c t r o n microscopy. The length o f 100 p a r t i c l e s was
measured, a length d i s t r i b u t i o n histogram was p l o t t e d and a modal
l eng th ca lcu la ted .
D. U l t ra th in Sect ions o f BYMV-Infected Chickpea Leaves
U l t r a t h i n sec t ions of B W - i n f e c t e d chickpea leaves were
prepared t o observe inc lus ion bodies induced due t o in fec t ion .

Leaves were c u t i n t o small p i eces of 1 mm x 1.5 nun. They were
f i xed i n 3% glutaraldehyde f o r 2 hours, washed f o r 15 h r i n 0.1M phos-
phate buffer pH 7 . 3 , containing 0.5% sucrose. They were s to red over-
n i g h t i n r e f r i g e r a t o r i n the same bu f f e r . Next day, they were f i x e d i n
2% osmium t e t r o x i d e made i n 0.1M phosphate b u f f e r pH 7 . 3 containing 5%
sucrose f o r 2.5 h r s . They were washed i n d i s t i l l e d water f o r 20 minutes
and then dehydrated i n 50, 70 and 90% acetone (20 min each) . The f i n a l
dehydrat ion was done i n 100% acetone t h r i c e , each time f o r 20 minutes.
Acetone dehydrated t i s s u e s were t r a n s f e r r e d t o 1:l mixture o f 100%
acetone and epoxy embedding r e s i n (Ladd Inc . , U.S.A.) and shaken f o r
1 h r . The t i s s u e s were drained on a paper towel and soaked i n f r e s h
r e s i n fo r 4 h r s .w i th a change of r e s i n every hour. The t i s s u e s were
drained once aga in and t r a n s f e r r e d t o f r e s h r e s i n i n s i l i c o n rubber
blocks which were kept a t 6 5 ' ~ f o r 48 hours . Sec t ions were c u t (50-60
nm t h i ck ) wi th a ' Reichert-Jung Ul t r acu t ' , s ta ined ' i n s a t u r a t e d uranyl
a c e t a t e f o r 3 min and in 5% lead c i t r a t e f o r 3 min, washed, d r i ed
and observed under t he e l ec t ron microscope.
I11 I . PHYS I ~ O - QIEMICAL PROPERTIES OF PURIFIED CMV
A. W-Absorption
The W-absorpt ion spec t r a of t h e p u r i f i e d CMV ,were recorded
i n Beckman D. B . G . T. spectrophotometer a t wave l eng ths ranging from
200 t o 300 mm. Rat ios of 2601280, 280/260, and A max/A min were
ca l cu l a t ed . The y i e l d of t h e v i r u s was est imated using an ex t inc t ion
c o e f f i c i e n t of 5.0 OD = 5.0) (Gibbs and Harrison, 1970).
B . Sedimentation Coeff ic ien t
Sedimentation c o e f f i c i e n t of the p u r i f i e d CMV was determined
i n a Spinco model E a n a l y t i c a l u l t r a c e n t r i f u g e . The p u r i f i e d v i r u s
suspended i n 0.025M potassium phosphate bu f f e r was run a t 30,000 rpm.

After reaching the des i red speed, the Schleiren p a t t e r n was photo-
graphed a t 3 min in te rva l s . The sedimentation coef f i c i en t was deter .
mined by w i n g the following formula:
C. Buoyant Density
One m l of the pur i f i ed CMV was layered over 4 m l o f 40%
cesium chlor ide (density = 1.42 g/ml) i n 5 m l polyallmoer tubes, and
centrifuged i n SW-50 r o t o r a t 40,000 rpm f o r 18-20 h r s a t 4 ' ~ . The
gradient column was f rac t iona ted by puncturing the bottom of the tube
and c o l l e c t i n g 0 . 2 rnl f r ac t ions . These f r a c t i o n s were monitored i n t h e
Beckman Spectrophotometer a t 260 nm, and the d e n s i t i e s o f t h e f r a c -
t i o n s were determined by t h e i r r e f r a c t i v e indices measured with an
Abbe refractometer.
D. ~ o l ~ a c r ~ l a m ~ d e Gel Electrophoresis (PAGE) o f CMV Coat Prote in
The PAGE of CMV coat prote in was done following the method
of Laemmli (1970).
Stock so lu t ions
The stock so lu t ions were prepared a s follows:
Solution A (3M Tris-HC1 buffer , pH 8.91
Tri s : 36.6 g
1NHC1 : 48 .0ml
TEMED : 0 .23 m l
Made up t o 100 ml with d i s t i l l e d water and adjusted pH t o 8.9 with 1N HC1

Solution B (1M Tris-HC1 buffer , pH 6.8)
Tr is : 5.98 g
1 N HC1 : 48.0 m l
TEMED : 0.46ml
Made up t o 100 m l with d i s t i l l e d water and adjus ted pH t o 6.8 with 1N HC1
Solution C (Acrylamide : Bis)
Acrylamide : 30.0 g
B i s : 0.735 g
Made up t o 100 ml with d i s t i l l e d water.
Solution D (Acrylamide : Bis)
Acrylamide : 10.0 g
B i s : 2.5 g
Made up t o 100 ml with d i s t i l l e d water.
Preparation of the Gels
Resolving (rgnplraing) gel (10%)
Solution A : 2.0 ml
Solution C : 5 .2 ml
1% Sodium dodecyl s u l f a t e (SDS) : 1.6 m l
10% Urea : 4.8111.1
D i s t i l l e d water : 1.4 m l
1% Ammonium persu l fa te : 1.0 ml
Stacking (spacer) gel (3.6%)
Solution B : 0.5 m l
Solution D : 1.4 ml
1% SDS : 0.4 m l
10% Urea : 1 . 2 ml
Ammonium persu l fa te : 0.25 ml

With t h e he lp of a long-beaked Pas teur p i p e t t e , the reso lv ing
ge l was poured i n t o Bio-rod gel tubes up t o a l eng th o f 6 cms and
allowed t o polymerize f o r 15 minutes. Over t h i s s tacking gel was
poured up t o a length of 1 cm and allowed t o polymerize f o r 30 min.
After polymerization, t h e tubes were inse r t ed i n t h e e l e c t r o p h o r e t i c
apparatus and f i l l e d with e lec t rophores i s b u f f e r (0.05M Tr i s , 0.384M
glycine , 0.1% SDS, pH 8.3).
The v i r u s was d i s soc ia ted i n 0.0625M Tris-HC1 buf fe r , pH 6.8
conta ining 1% 2-Mercaptoethanol, 1% SDS and 6M urea . I t was then
incubated a t 1 0 0 ' ~ f o r 2 min ( i n b o i l i n g water) . Ten t o 100 p1 o f
t h e d i s soc ia ted v i r u s sample was layered on each ge l . A mixture o f
s i x marker p ro te ins (Bio-Rod Chemicals, U. S .A.) containing 20-100 pg
o f each polypeptide was a l s o layered separa te ly a s well a s mixed wi th
v i r u s p ro te ins ( a - e l e c t r o p h o r e s i s ) . Af ter e l e c t r o p h o r e s i s a t 30 v o l t s
f o r 6 hours a t room temperature, t h e g e l s were taken o u t o f t h e tubes
by l u b r i c a t i n g t h e inner wal ls o f t h e tubes wi th water, using a
syr inge with a b l u n t needle.
?h'e'gels were s t a ined f o r 4 hr i n 0.2% coomassie b r i l l i a n t
b lue prepared i n 20% methanol i n 7% a c e t i c a c i d . Destaining o f t h e
g e l s was done overnight with 20% methanol i n 7% a c e t i c a c i d . The g e l s
were than scanned a t 540 nm i n a Gi l l fo rd spectrophotometer .
The m l e c u l a r weight of CMV coat p r o t e i n was determined by
t h e l i n e a r r e l a t i o n s h i p between t h e migration d i s t a n c e o f marker
p r o t e i n s and t h e logari thm o f t h e i r known m l e c u l a r weights.
E. Polyacrylamide Gel Elect rophores is of CMV WAS
The PAGE of CMV RNAs was performed using t h e method a s
described by Adesnik (1970). A combination of 2.5% acrylamide and
0.5% agarose was used.

Stock solut ions
The stock solut ions were prepared a s follows:
Solution A (15% acrylamide)
Acrylamide : 7 .1 g
B i s : 0.4 g
Dissolved i n d i s t i l l e d water and made up t o 50 m l .
Solution B (15 x buffer , pH 7.5)
Tri s : 0.4M
NaH2P04 : 0.3M
E DTA : 0.01M
Solution C
10% ammonium persu l fa te (prepared j u s t before use)
Preparation of gels
. Mix
Solution A 5 .0 ml
15 x buffer 2 . 0 m l
D i s t i l l e d w a t e r 7 .7ml
TEMED 0.03 m l
1% agarose 15.0 ml
10% ammonium 0.3 m l (added a t t he end) per sul f a t e
The gels were immediately ca s t in to t he Bio-rod gel tubes
with the help of a long-beaked Pasteur p ipe t te . lbo drops of
d i s t i l l e d water were layered over the gel surface with the help of
a syringe and needle t o obtain a sharp meniscus. The gel tubes were
in se r t ed in to an electrophoret ic apparatus and f i l l e d with e lec t ro-
phoresis buffer (1 x buffer) t o which 0.05% die thyl pyrocarbonate was

added t o avoid poss ib le contamination with ribonudease (RNase) . Before
loading the samples, a pre-run was given a t 5 mA/gel f o r + h r . Virus
suspension was disrupted i n 1.5 x buf fe r conta ining 10M urea, 4 % SDS,
1% 2-Mercaptoethanol, 2mM EDTA, t o which 0.001% bromophenol b lue was
added a s a tracking dye.
The disrupted v i r u s containing about 50 t o 100 pg of nuc le ic
ac id was layered on the ge l s . Some ge l s were loaded with t h e d i s rup ted
v i r u s t h a t was t r e a t e d with RNase a t the r a t e of 1 pg/ml, and incubated
a t 2 5 ' ~ f o r 1 hour.
Electrophoresis was done a t 5mA/gel a t room temperature
u n t i l the t racking dye reached near the bottom of t h e g e l (about 3
hours) . The ge l s were removed from the tubes a s described i n p ro te in
gels . They were suspended i n 1 M a c e t i c ac id f o r 15-30*minutes, s t a i n e d
i n 0.2% to lu id ine blue, prepared i n 0.4M a c e t a t e b u f f e r , pH 4 . 7 f o r
15-30 min.
Destaining of the g e l s was done i n severa l changes of
d i s t i l l e d water i n order t o loca te RNA bands.
IX. PRODUCTION OF ANTISERUM TO CMV
An Albino rabb i t , weighing about 2 kg was used f o r immuni-
za t ion. One ml o f p u r i f i e d v i r u s (2-3 mg/ml) was emulsif ied with an
equal volume o f Fruend's incomplete adjuvant and i n j e c t e d i n t o the
rabb i t intramuscularly i n t h e th igh muscle of hind l e g . A t o t a l of
four i n j e c t i o n s were given a t 10-day-intervals. Ten days a f t e r t h e
l a s t in jec t ion , the r a b b i t a f t e r 16 h r s t a r v a t i o n was bled through a
marginal e a r vein by making an i n c i s i o n wi th a s t e r i l i z e d razor . The
blood was c o l l e c t e d i n a t e s t tube, allowed t o c l o t f o r one hour a t
room temperature and kept a t 4 ' ~ overnight. The serum was drawn with

a Pasteur p i p e t t e and centr i fuged a t 3000 rpm f o r 10 min i n a c l i n i c a l
cen t r i fuge t o separa te the serum from t h e blood plasma. Sodium azide (0.1%) was added a s a preservat ive t o the serum.
Ten-fold d i l u t i o n s of the antiserum were made i n normal
s a l i n e and t h e t i t r e was determined by employing t h e Ouchterlony agar
d o u b l e d i f fus ion t e s t .
X. DETERMINATION OF ?HE SEROLOGICAL RELATIONSHIP BETWEEN THE PRESENT S W I N OF CMV WITH OTHER KNOWN STRAINS
The se ro log ica l a f f i n i t y of t h e present s t r a i n of CMV with
o t h e r s t r a i n s was determined employing t h e Ouchterlony agar Quble-
d i f fus ion t e s t . Ant isera t o e igh t s t r a i n s of MV, obtained from
Dr. H . A . S c o t t (U.S.A.), Dr. H.E. Waterworth (U.S.A.), and Dr. R.I.B.
Francki (Aus t ra l i a ) , along with t h e homologous antiserum were made t o
form p r e c i p i t i n bands with pur i f ied vi rus . Thickness o f bands formed
was taken a s a method f o r determining se ro log ica l a f f i n i t y .
X I . SCREENING FOR DISEASE RESISTANCE
Chickpea germplasm l i n e s t h a t were repor ted t o be r e s i s t a n t
t o o t h e r d i seases ; v iz . , w i l t , r o o t r o t s , Ascochyta b l i g h t and s tun t
(pea l e a f r o l l v i rus ) were screened f o r r e s i s t a n c e t o CMV and BYMV.
For each gemplasm l i n e , 10 seeds were sown i n each 20 cm
p l a s t i c po t . Ten-day-old seedlings were separa te ly rub-inoculated
with t h e standard e x t r a c t of the two v i ruses . Observations on the
number of p l a n t s infected were recorded f o r each l i n e .

RESULTS
I. SYMPTOMATOLOGY IN CHICKPEA
A. Symptoms Produced by CMV on Chickpea
Two types of symptoms were produced by CMV i n chickpea. In
c e r t a i n v a r i e t i e s , t w i s t i n g o f terminal bud occu ed 4 - 5 days a f t e r ii i nocu la t i on (F ig . l a ) which was followed by n e c r o s i s o f te rmina l and
a x i l l a r y buds and w i l t i n g o f t h e p l a n t . P l a n t s d i ed 9-10 days a f t e r
i n o c u l a t i o n (Fig. lb ) . In o t h e r v a r i e t i e s , i nocu la t ed p l a n t s a f t e r
showing t w i s t i n g and n e c r o s i s o f terminal bud, d i d not w i l t , b u t showed
c h l o r o s i s followed by p r o l i f e r a t i o n of branches. The new branches
remained green which bore very small l eaves and very few, small pods.
Affec ted p l a n t s appeared b u h y (Fig. l c ) .
B. Symptoms Produced by BYMV on Chickpea
' h i s t i n g o f t h e te rmina l bud occurred 6-7 days a f t e r inocu-
l a t i o n which-was very s l i g h t a s compared wi th t h a t by CMV i n f e c t i o n .
This was followed by i n i t i a t i o n o f very narrow l eaves from new buds
and was more ev ident 15 days a f t e r i nocu la t i on (F ig . 2a). As t h e time
advanced, p r o l i f e r a t i o n of bo th terminal and a x i l l a r y branches occurred
r e s u l t i n g i n t o long and t h i n branches c o n s i s t i n g o f very narrow o r
f i l i form leaves . The leaves below t h e pro1 i f e r a t e d branches e x h i b i t e d
yellowing o r i n t e r v e i n a l c h l o r o s i s o r mosaic depending on t h e genotype
t e s t e d . The o v e r a l l he igh t of t h e p l a n t was reduced. Such p l a n t s
produced very few d i s t o r t e d f lowers t h a t were converted t o very small
pods (F ig . 2b) . The seeds from i n f e c t e d p l a n t s were b l ack , small and
very much s h r i v e l l e d (Fig. 2c) .
11. HOST RANGE
The r e a c t i o n s of d i f f e r e n t p l an t s p e c i e s t o CMV and BYMV a r e
p re sen ted i n Table 3 and Figures 3, 4 , 5 and 6 .
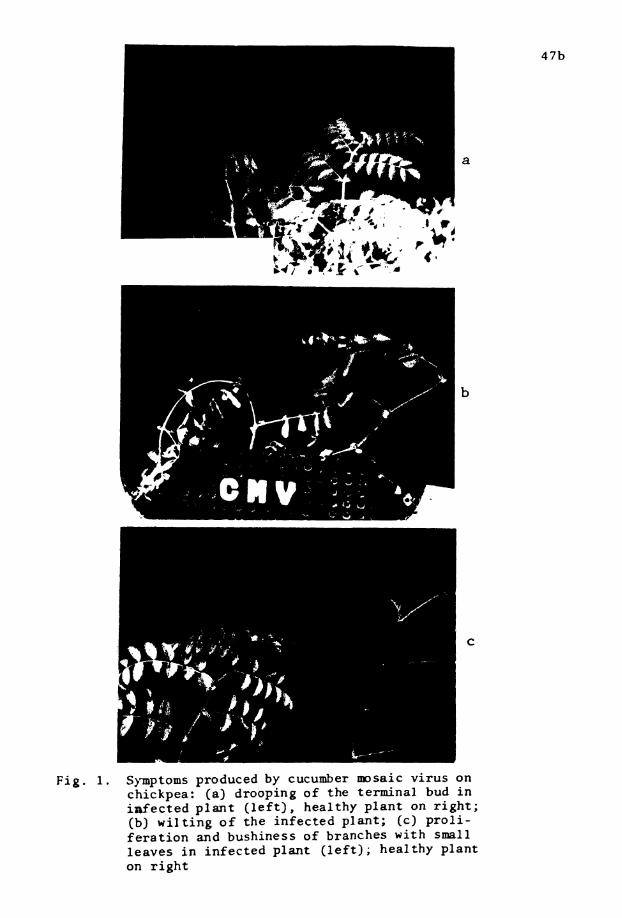
F i g . Symptoms produced by cucumber mosaic v i r u s on chickpea: ( a ) drooping of the terminal bud i n i a f e c t e d p l a n t ( l e f t ) , hea l thy p l a n t on r i g h t ; (b) w i l t i n g of t h e i n f e c t e d p l a n t ; (c) p r o l i - f e r a t i o n and bushiness of branches wi th small l e a v e s i n i n f e c t e d p l a n t ( l e f t ) ; hea l thy p l a n t on r i g h t

F i g . 2 . Symptoms produced by bean yellow mosaic v i r u s on chickpea: (a) an i n f e c t e d p l a n t wi th narrow o r f i l i f o r m leaves ; (b) showing p r o l i f e r a t i o n o f a branch with small pods ( r i g h t ) , h e a l t h y branch on l e f t ; (c) d iscoloured, deformed and s h r i v e l l e d chickpea seeds from i n f e c t e d p l a n t s ( l e f t ) , seeds from hea l thy p lan t on r i g h t .
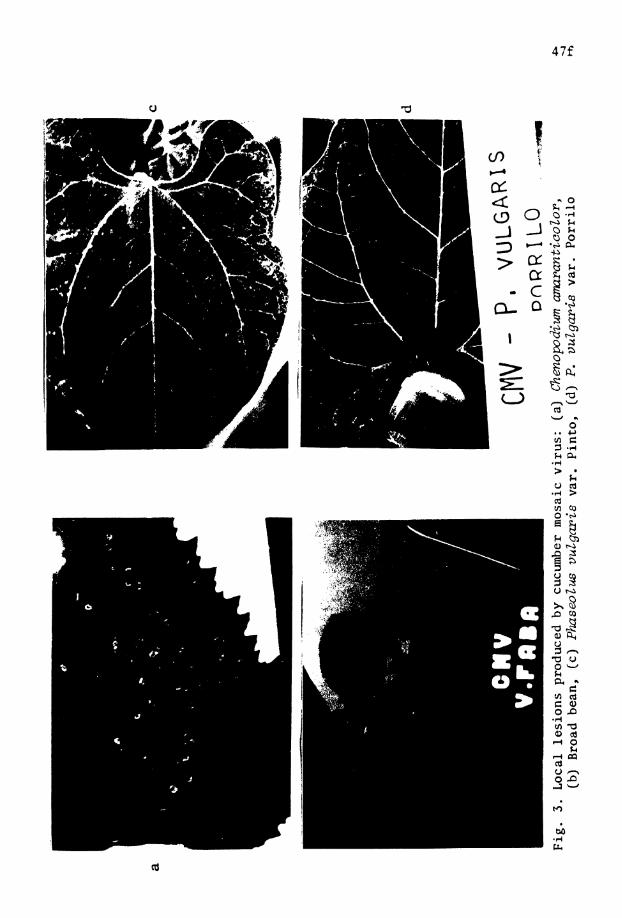
Fig. 3 . Local l es ions produced by cucumber mosaic virus: (a) Chenopodiwn amaranticoZor, (b) Broad bean, (c) PhuseoZus ~uZgnris var. Pinto, (d) P. vuZgaris var. Porrilo
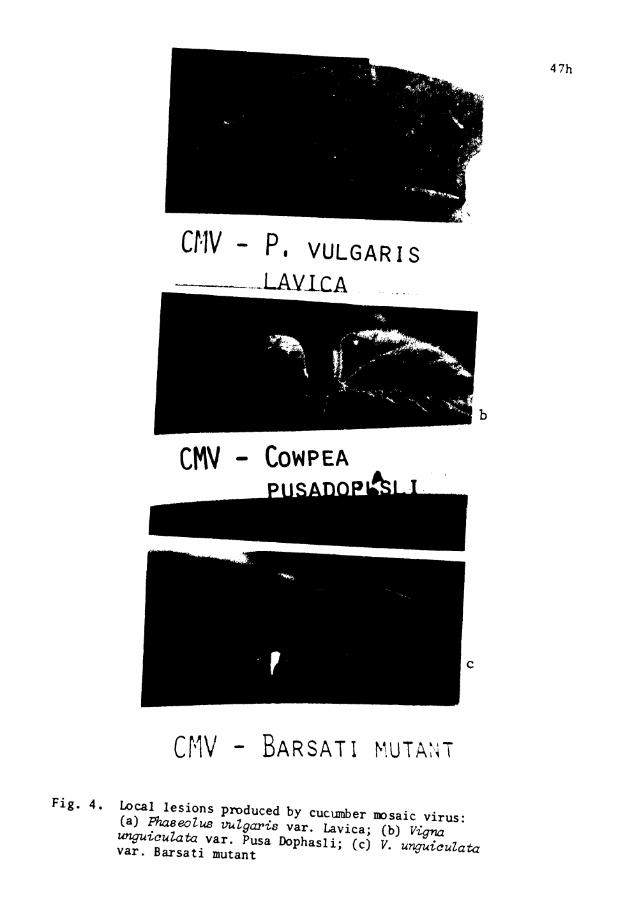
CMV - P a VULGAR IS
CMV - BAR SAT^ M U T A : ; ~
Fig. 4 . Local l e s i o n s produced by cucumber m s a i c v i r u s : (a) PRPBeoZw m 2 g a l . i ~ va r . Lavica; (b) V i g w u n g u i c u b t a v a r . Pusa Dophasli; (c ) V. urguiculata v a r . B a r s a t i mutant

CMV - CUCUMBER LONG F l g . 5 . Systemic symptoms produced by cucumber mosaic v irus on: (a) Nicotiana rustics; (b) cucumber
and (c ) peas
F i g . 6 . Systemic symptoms produced by bean yellow mosaic virus on P. vuZgaris

TABLE 3 . Host range and symptomatology of cucumber mosaic and bean yellow mosaic viruses
Symptoms Host CMV BYMV
A 1 ZOACEAE
Tetragoniu expansa Murr . AMARAN lMACEAE
Gomphrena globosa Murr . CHENOPODIACEAE
Chenopodiwn mant-LcoZor Coste e t NLL Ryne
COW 0s I TAE
Chry sun themwn sp . CUCURBI TACEAE
Systemic, inducing - large ch loro t ic patches
Cucurbitu pepo L . CLL - LEGUMINOSAE
Arachis hypogaea L. - - Detailed symptoms are described i n the t e x t
Cajanus cajan ( L . ) Millsp. NLL - CanavaZia ensi fomis D.C. - Mos, f r b t l .
&crotyZoma m i f l o r m (Lam.) Verdc. - - PhQseoZus vuLgaris L. 'Ababat NLL hb s.
P. vuZgaris ' Po r r i l o t NLL - P. vuZgaxis 'Red kidneyt N LL VN, M s . M t l .
P. vuZgaris Dwarf
P. vuZgaris 'Pinto
- a, m, b s , Motl, Ma
NLL VN, Mos, Motl, Ma.
P. vuZgaris 'Munroe1 NLL - P. vuZga- ' Lavica ' NLL -
contd.

TABLE 3 . Contd,
Host Symptoms
CMV BY MY
P i s m s a t i v m L. - Vicia faba L. 'Early long pod' NLL
V i g n a unguicuZatu (L.) Walp. NLL 'Early Ramshorn'
V. unguicu~ata ' Barsat i mutant ' NLL
V . unquiculatu 'Puss Ebphasli ' N L L
SOLAN ACEAE
Capsicm a n m m L. - - Datura stramoniwn L . CL L
Lycope~sicon esculentwn Mill. ' Pusa Rubyt - Nicotiam e a c w n L . ' Xanthi-ncl Mos, M t l . - N. rust ica L. 'White Pathar ' Mos, M o t l . - Petunia hybrida V i l m . - -
CLL = Chlorot ic l o c a l l e s ions ; Ma = Malformation of leaves; Mos = Mosaic; Motl = Mottle; NLL = Necrot ic l o c a l l e s ions ; TN = Top necros is ; VN = Veinal necros is ; - = No symptoms.
111. I N VITRO PROPERTIES
A . Cucumber Mosaic Virus
1. Thermal i n a c t i v a t i o n m i n t (TIPI
The assay p l a n t s inocula ted with t h e s tandard e x t r a c t t r e a t e d
a t 4S0, So0, 5S0, 60°, 6s0, and 7 0 ' ~ produced l o c a l l e s i o n s whereas no
loca l l e s i o n s developed with the e x t r a c t heated a t 7S0c o r more (Table 4)
The TIP o f t h e v i r u s was t h u s between 70' and 75'~.

TABLE 4. Influence of hea t ing t h e standard e x t r a c t of cucumber mosaic v i r u s a t d i f f e r e n t temperatures f o r 10 minutes on i n f e c t i v i t y
Exposure temperature (OC) Number o f loca l l e s ions*
4 5
5 0
5 5
6 0
6 5
7 0
75
80
8 5
9 0
Control
*Average o f s i x r e p l i c a t i o n s wi th range o f l e s i o n s i n parentheses
2 . Dilution end po in t (DEP)
Assay p l a n t s inocu la ted with 0, lo-', and lo - )
d i l u t i o n s of t h e s tandard e x t r a c t produced l o c a l l e s i o n s . No l o c a l
l e s i o n s were produced when t h e p l a n t s were inocu la ted wi th t h e
e x t r a c t d i l u t e d t o 1 0 ' ~ o r above (Table 5 ) . Thus t h e DEP o f t h e
v i r u s war between and ld4.
3 . Longevity in vitm
The s tandard e x t r a c t s t o r e d a t mom temperature (22' - 24'~)
l o s t i t s i n f e c t i v i t y on t h e f i f t h day (Table 6) .
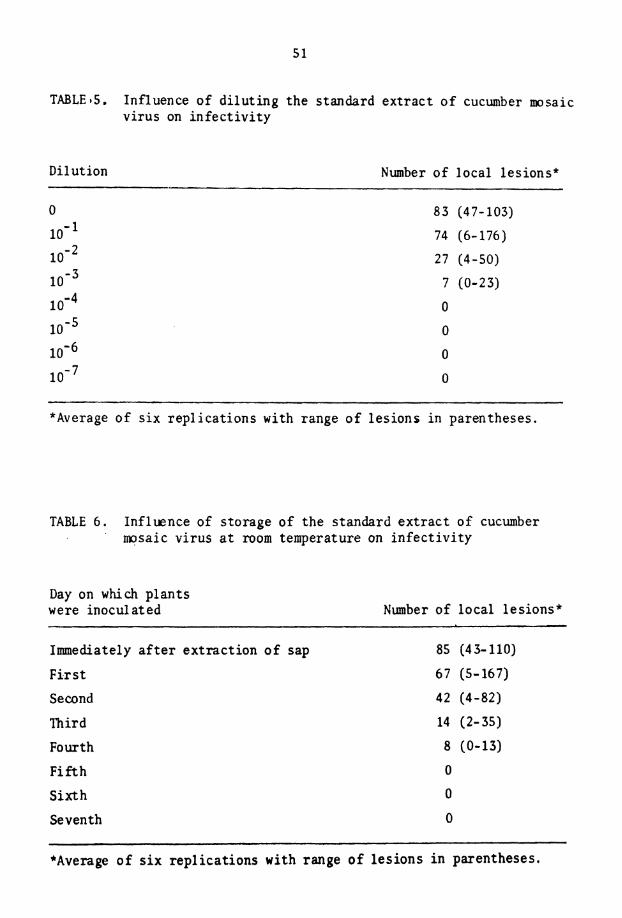
TABLE*5. Influence o f d i l u t i n g t h e standard e x t r a c t o f cucumber mosaic v i r u s on i n f e c t i v i t y
Di lut ion Number of l o c a l l e s ions*
0
*Average of s i x r e p l i c a t i o n s wi th range o f l e s i o n s i n parentheses .
TABLE 6 . Infllrence of s to rage o f t h e standard e x t r a c t o f cucumber mqsaic v i r u s a t room temperature on i n f e c t i v i t y
Day on which p l a n t s were inocula ted Number of l o c a l l e s i o n s *
Immediately a f t e r e x t r a c t i o n of sap
F i r s t
Second
Third
Fourth
F i f t h
S i x t h
Seventh
*Average o f six r e p l i c a t i o n s wi th range o f l e s i o n s i n parentheses.

B. Bean Yellow Mosaic Virus *
1. Thermal inac t iva t ion point (TIP)
The chickpea p l a n t s inocula ted with t h e standard ex t rac t
heated a t 45', 50' and 5 5 ' ~ f o r 10 minutes produced disease symptoms
(Table 7). However, no symptoms were no t i ced when the p l a n t s were
inocula ted with the e x t r a c t heated a t 60°c o r more. The TIP o f t h e
v i r u s was thus between 55' and 60'~.
2. Dilution end po in t [DEP)
P lan t s inocula ted with t h e s tandard e x t r a c t d i l u t e d to 10"
and produced d i sease symptoms but no t with d i l u t i o n s t h e r e a f t e r
[Table 8) . The DEP o f t h e v i r u s was thus between and
3 . Longevity i n v i t r o
The v i r u s sap when s t o r e d a t room temperature (22-24'~) l o s t
i t s i n f e c t i v i t y on t h e 2nd day (Table 9 ) .
TABLE 7. Influence o f hea t ing the s tandard e x t r a c t of bean yellow m s a i c v i r u s a t d i f f e r e n t temperatures f o r 10 minutes on i n f e c t i v i t y
Expo sure temperature (OC)
4 5
50
5 5
60
65
7 0
75
80
85
9 0
Control
No. o f p l a n t s No. o f p l a n t s inocu la ted i n f e c t e d
10 7
10 5
10 3
10 0
10 0
10 0
10 0
10 0
10 0
10 0
10 10
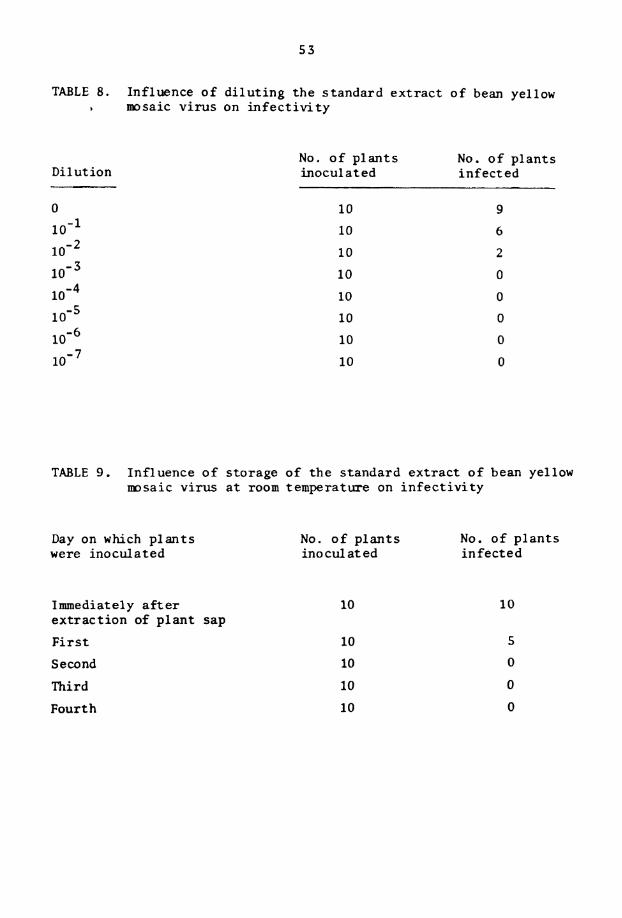
TABLE 8. Inf luence o f d i l u t i n g t h e s t a n d a r d e x t r a c t o f bean yellow , mosaic v i r u s on i n f e c t i v i t y
Di lu t ion No. o f p l a n t s No. o f p l a n t s inocu la ted i n f e c t e d
TABLE 9 . In f luence o f s to rage o f t h e s t andard e x t r a c t o f bean yellow m s a i c v i r u s a t room temperature on i n f e c t i v i t y
Day on which p l a n t s were inocu la ted
Immediately a f t e r e x t r a c t i o n of p l a n t sap
F i r s t
Second
Third
Four th
No. o f p l a n t s No. o f p l a n t s ino cul a t ed i n f e c t e d

IV. INSECT TRANSMISSION #
The r e s u l t s on t h e transmission o f t h e two v i ruses by t h e
two aphid species a r e given i n Table 10. Both the vi ruses could be
t ransmit ted by Mzjzus persicae and Aphis cmccivora in a non-pers is tent
manner though t h e l a t t e r was found t o be m r e e f f i c i e n t than the
former.
V. PURIFICATION
A . Cucumber Mosaic Virus
The p u r i f i c a t i o n procedure adopted i n t h e present s tud ies
r e s u l t e d in highly p u r i f i e d and i n f e c t i o u s v i rus . .When butanol was
used f o r i n i t i a l c l a r i f i c a t i o n , m s t o f t h e chloroplas t mater ia l
could be e l iminated and thus a c l e a r amber-coloured supernatant was
obta ined a f t e r low speed cen t r i fuga t ion . m e v i r u s could e a s i l y be
concentrated by PEG p r e c i p i t a t i o n and d i f f e r e n t i a l cen t r i fuga t ion .
The high speed cen t r i fuga t ion r e s u l t e d i n a g lassy p e l l e t and by which
time almost a l l t h e green colour could be e l iminated. Further pur i -
f i c a t i o n by densi ty gradient cen t r i fuga t ion r e s u l t e d i n a s ing le ,
th ick, l i g h t s c a t t e r i n g zone in dens i ty gradient tube (Fig. 7a). When
t h e gradient tube was subjected t o f r a c t i o n a t i o n i n an ISCO f r a c t i o n -
a t o r , a s i n g l e absorption peak could be obta ined corresponding t o t h e
v i r u s zone (Fig. 7b). 'Ihe r e s u l t s of t h e i n f e c t i v i t y assays o f
gradient f r a c t i o n s ind ica ted t h a t t h e i n f e c t i v i t y was confined t o the
W absorbing aomponent (Fig. 7b). A c l e a r g lassy p e l l e t was obtained
i n high speed , cen t r i fuga t ion of t h e d i l u t e d v i r u s zone and could be
e a s i l y suspended i n buf fe r .
?he homogeneity of t h e p u r i f i e d v i r u s could b e f u r t h e r
assessed spectrophotometrically. The absorption spectrum was found
t o be typ ica l of nucleoprote in (Fig . 7c) , with absorpt ion maximum a t

TABLE 10. Aphid transmission o f cucumber mosaic and bean yellow mosaic v i r u s e 2
Aphid species
No. o f p lants Percent No. of aphids b No. o f p lants infected transmission used per p lant inoculatedC CMV BY MV CMV BYMV
Mjzw persicae
Aphis craccivora.
a. Data from two t r i a l s .
b. 'he aphids were given one hour f a s t i n g period and one minute acquisi t ion feed period.
c. Inoculation feeding period was 1 2 hours. Ten-day-old p lants were used f o r inoculation.
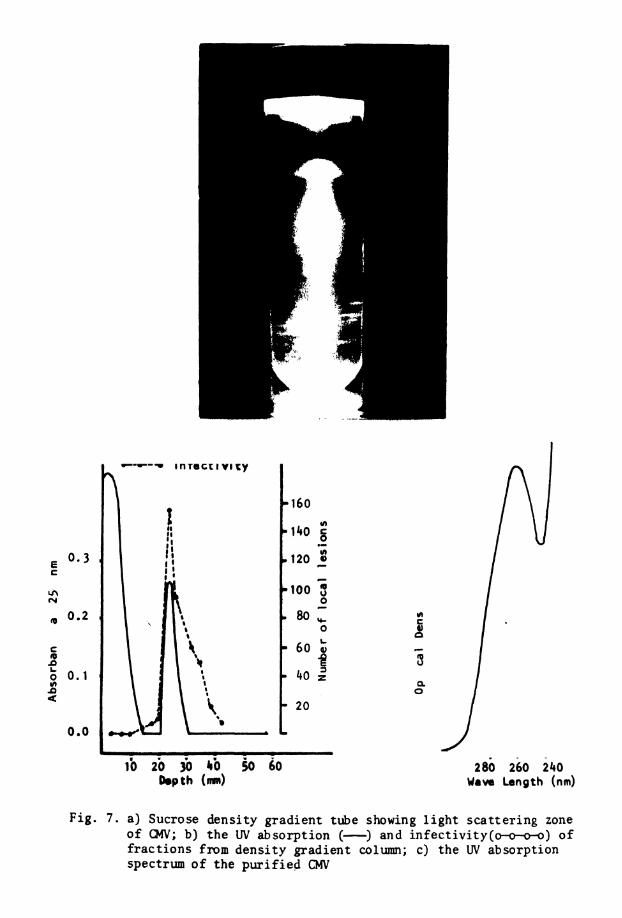
2d 260 240 W I W Length (nm)
Fig. 7 . a ) Sucrose dens i ty g rad ien t tube showing l i g h t s c a t t e r i n g zone of W; b) t h e W absorpt ion (-) and infectivity(-) of f r a c t i o n s from d e n s i t y g rad ien t column; c) t h e W absorpt ion spectrum of t h e p u r i f i e d CMV

260 nm and minirmun a t 240 nn. I t had A260/A280J A280/A260J and $
A max/A min r a t i o s o f 1.55, 0.64, and 1.27, r e spec t ive ly .
The v i r u s y i e l d ranged from 8 t o 10 mg/100 g of p lan t t i s s u e
when ca lcu la ted spectrophotometrically.
B. Bean Yellow Mosaic Virus
Since a s a t i s f a c t o r y loca l l e s i o n hos t could not be found,
the success o f t h e p u r i f i c a t i o n procedure adopted was assessed only
through t h e use o f t h e e l e c t r o n microscope. With t h e experience
gained from d i f f e r e n t p u r i f i c a t i o n procedures t r i e d , t h e following
conclusions could be drawn:
Carbon t e t r a c h l o r i d e was b e t t e r than chloroform o r butanol
f o r pre l iminary c l a r i f i c a t i o n .
r Addition o f carbon t e t r a c h l o r i d e i n t h e Waring blendor
during homgenizat ion was found t o be b e t t e r than adding
it a f t e r homogenization.
Use o f PEG alone was found t o be b e t t e r than PEG + NaC1.
Overnight incubation o f t h e PEG dissolved prepara t ion was
found t o be b e t t e r than incubat ion f o r 2 hours.
High speed cen t r i fuga l f o r c e s increased p a r t i c l e breakage.
Use of a sucrose pad was found use fu l i n e l imina t ing most
o f t h e green mate r i a l .
Out o f severa l methods used, t h e one adopted i n t h e present
i n v e s t i g a t i o n (descr ibed under Mater ia ls and Methods) was found t o be

s a t i g f a c t o r y . But i n densi ty gradient cen t r i fuga t ion , t h e v i r u s zone
was no t compact and c lea r . Rather, an inconspicuous, d i f fused zone
was obtained. However, t h i s zone, when p e l l e t e d consis ted o f high
concentrat ion o f v i r u s p a r t i c l e s ; though, the re was some degree of
breakage.
The o p t i c a l cha rac te r s and o t h e r physico-chemical p roper t i e s
could not be performed s i n c e t h e v i r u s p repara t ion , f r e e o f host
components could not be obtained.
VI . ELECTRON MICROSCOPY
A. CM! P u r i f i e d Prepara t ion
The negat ively s t a ined prepara t ion o f t h e p u r i f i e d CMV
contained a homogenous population o f spher ica l p a r t i c l e s which
measured about 30 nm i n diameter (Fig. 8 ) .
B . Bean Yellow Mosaic Virus
1. Leaf d ip prepara t ion
The negat ively s t a i n e d 1 eaf d ip prepara t ion revealed the
presence of f lexuous rods (Fig. 9 ) . From t h e l eng th d i s t r i b u t i o n
histogram (Fig. 10) i t i s c l e a r t h a t most o f t h e p a r t i c l e s measured
between 740 and 760 nm with a modal l e n g t h o f 750 nm. The width was
determined t o be about 15 nrn.
2 . Pur i f i ed prepara t ion
A negat ively s t a i n e d p repara t ion o f the p u r i f i e d BYMV
contained flexuous rods. Though the re was a breakage o f p a r t i c l e s
t o some ex ten t , aggregation was not observed (Fig . 11) .


m w o rd 4 h n O cdo .ri F 4 0 C, Q) 1- k k - rd a m 14 * a M.rl X c a -
* d
g % 2 5 2 :
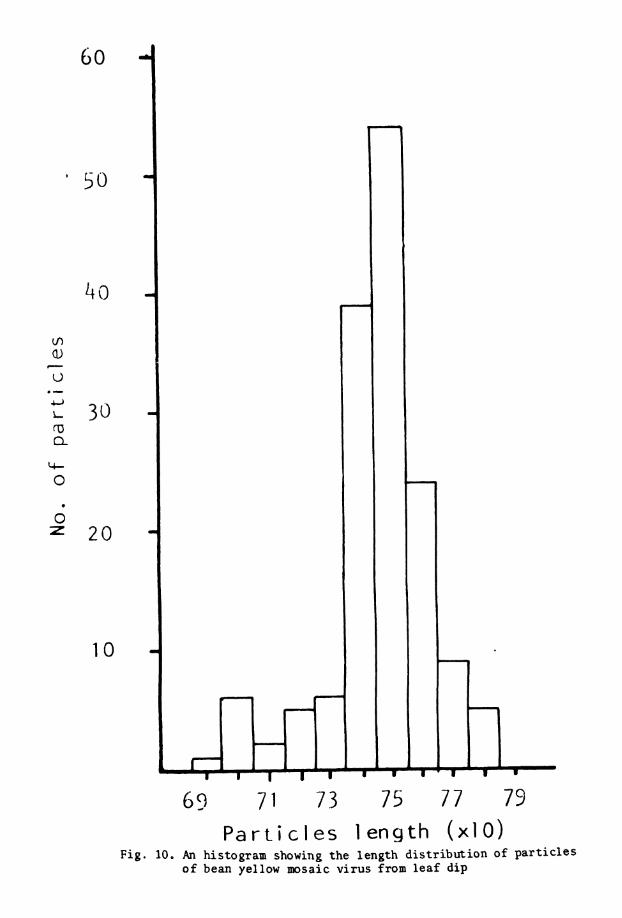
P a r t i c l e s l e n g t h ( ~ 1 0 ) F i g . 10. An histogram showing the length d i s t r i bu t i on o f p a r t i c l e s
of bean yellow mosaic v i r u s f r o m l e a f d ip


3. Ul t ra th in sec t ions o f BYMV-infected chickpea leaves
Electron microscopy of u l t r a t h i n sec t ions o f BYMV-infected , chickpea leaves revealed the presence o f pinwheel inc lus ions with
curved p l a t e s , and laminate aggregates (Fig. 1 2 ) .
JII. IDENTIFICATION OF VIRUSES BY SEROLOGY
A. Double-Diffusion i n Agar
Clear p r e c i p i t i n bands could be observed when crude sap from
v i rus - infected chickpea p l a n t s and p u r i f i e d prepara t ions of CMV
(Fig. 13a) and BYMV (Fig. 13b) were made t o r e a c t wi th t h e i r respect ive
a n t i s e r a . The p u r i f i e d CMV sometimes produced two p r e c i p i t i n bands
with i t s antiserum, a s t r a i g h t one nea r t h e antiserum well and a
curved one near t h e ant igen well (Fig. 13c) . No reac t ion e i t h e r with
the heal thy sap o r b u f f e r control was observed wi th a n t i s e r a o f the
two vi ruses .
B. Enzyme-Linked Innnunosorbent Assay (ELISA)
For i d e n t i f i c a t i o n o f CMV by ELISA, t h e anti-CMV obtained
from D r . H.E. Waterworth, USDA, Maryland, U.S.A. was used (CMV-S) .
A p o s i t i v e reac t ion (development o f yellow colour) could be observed
both with i n f e c t i o u s crude sap and p u r i f i e d v i r u s a t y-globulin
concentra t ion o f 1:100 and conjugate concentra t ions o f 1: 100 and
1 :SO0 (Fig. 14) . No r e a c t i o n was observed e i t h e r with the heal thy
sap o r buf fe r .
The ELISA could not be performed w i t h BYMV due t o non-
a v a i l a b i l i t y o f good antiserum.

Fig. 12. Electron pbtomicrographs of uitratfuii sections cf BW- infected chickpea l ea f showing: (a) pinwheels (PW) and (b) laminate aggregates (LA)
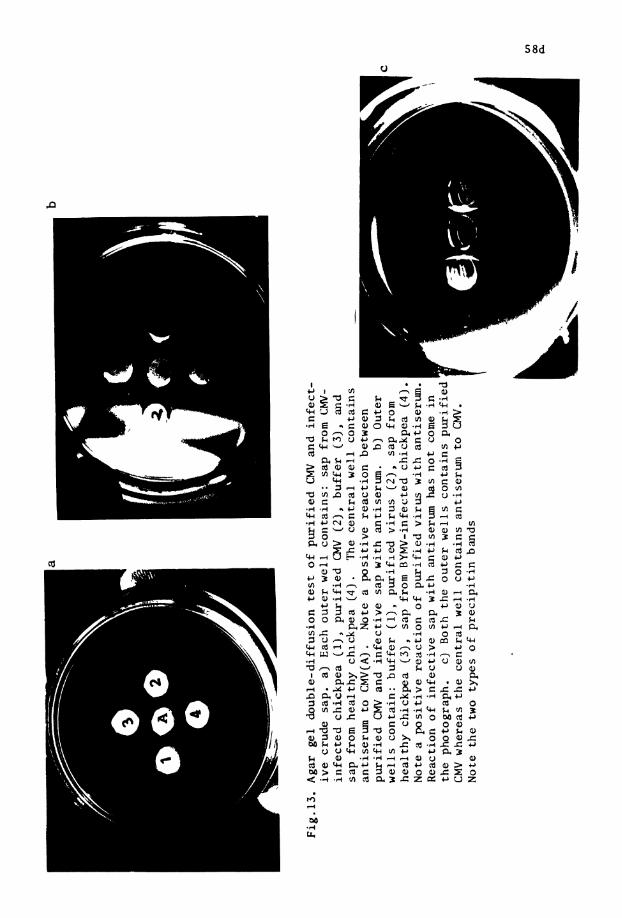
Fig. 13. Agar gel double-diffusion t e s t o f p u r i f i e d CMV and i n f e c t - i v e crude sap. a ) Each o u t e r wel l c o n t a i n s : sap from CMV- i n f e c t e d chickpea ( I ) , p u r i f i e d CMV (2), b u f f e r (3 ) ) and saD from hea l thy chickpea (41. The c e n t r a l wel l c o n t a i n s - L . <
ant iserum t o CMV(A). Note a p o s i t i v e r e a c t i o n between p u r i f i e d CMV and i n f e c t i v e sap with ant iserum. b) Outer w e l l s con ta in : b u f f e r ( I ) , p u r i f i e d v i r u s ( 2 ) , sap from heal thy chickpea ( 3 ) , s a p from BYW-infected chickpea ( 4 ) . Note a p o s i t i v e r e a c t i o n o f p u r i f i e d v i r u s with antiserum. Reaction o f i n f e c t i v e sap with antiserum has n o t come i n t h e photograph. c) Both t h e o u t e r w e l l s c o n t a i n s p u r i f i e d CMV whereas t h e c e n t r a l well c o n t a i n s ant iserum t o CMV. Note t h e two types o f p r e c i p i t i n bands
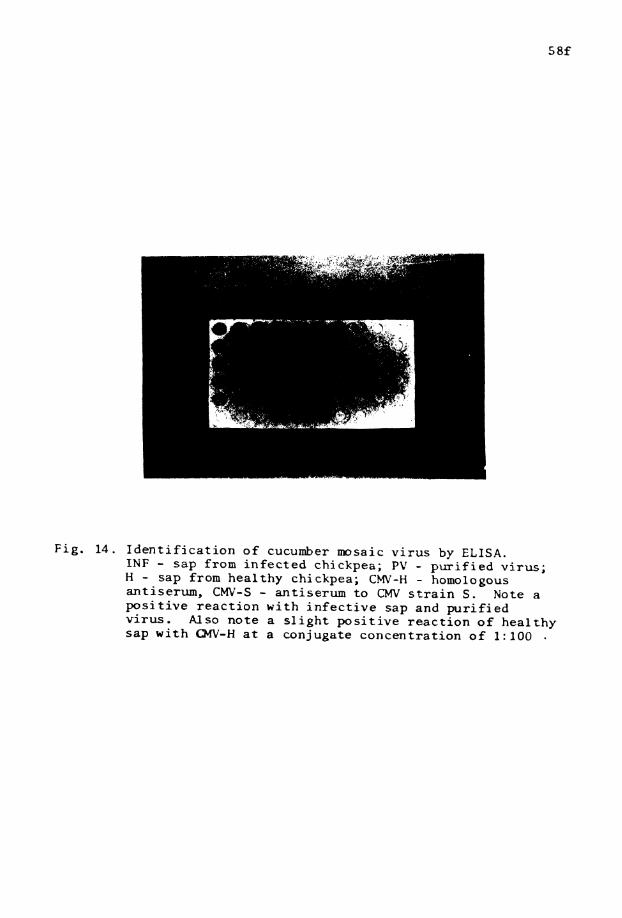
Fig . 1 4 . I d e n t i f i c a t i o n o f cucumber mosaic v i r u s by ELISA. INF - s a p from i n f e c t e d chickpea; PV - p u r i f i e d virus; H - sap from h e a l t h y chickpea; CMV-H - homologous ant iserum, CMV-S - an t i se rum t o CMV s t r a i n S . Note a p o s i t i v e r e a c t i o n w i t h i n f e c t i v e s a p and p u r i f i e d v i r u s . Also n o t e a s l i g h t p o s i t i v e r e a c t i o n o f h e a l t h y s a p with CMV-H a t a conjugate c o n c e n t r a t i o n o f 1:100 .

V I 11. PHYSICO-CHEMICAL PROPERTIES OF ?HE PURIFIED CMV
A. Sedimentation Coeff ic ien t (SZOW)
The homogeneity o f t h e p u r i f i e d CMV was ind ica t ed by t h e
presence o f a s ing le Schle i ren peak (Fig. 15) i n t h e ana ly t i ca l u l t r a -
cent r i fuge . The SZ0W o f t he p u r i f i e d CMV was ca l cu la t ed t o be 104s.
B. Buoyant Density
In t h e Cesium c h l o r i d e equi l ibr ium dens i ty gradient c e n t r i -
fugat ion , t h e p u r i f i e d CMV banded a t a d e n s i t y l e v e l o f 1.325 g/ml.
The dens i ty l e v e l s and UV absorpt ion o f d i f f e r e n t f r a c t i o n s o f t he
g rad ien t a r e given i n Fig. 16.
C. The PAGE o f Coat Pro te in
The PAGE o f SDS-disrupted coat p r o t e i n o f CMV revea led t h e
presence o f a s i n g l e polypeptide. The banding p a t t e r n in t h e ge l s and
the scanning p a t t e r n o f t h e W - c o a t p r o t e i n and marker p r o t e i n s a r e
given i n Fig. 17a and 18. Based on t h e l i n e a r r e l a t i o n s h i p between
migrat ion o f marker p r o t e i n s and t h e logar i thm o f t h e i r known molecular
weights (Fig. l a d ) , t h e molecular weight o f CMV-coat p ro t e in was
est imated t o be 25,000 da l tons .
D. The PAGE o f Nucleic Acids
The PAGE o f SDS-disrupted CMV r evea led t h e presence o f fou r
RNA spec i e s (Fig. 1%). The e l ec t rophores i s o f t h e d i s rup ted v i r u s
t h a t was f i r s t t r e a t e d wi th RNase d id n o t produce any bands.

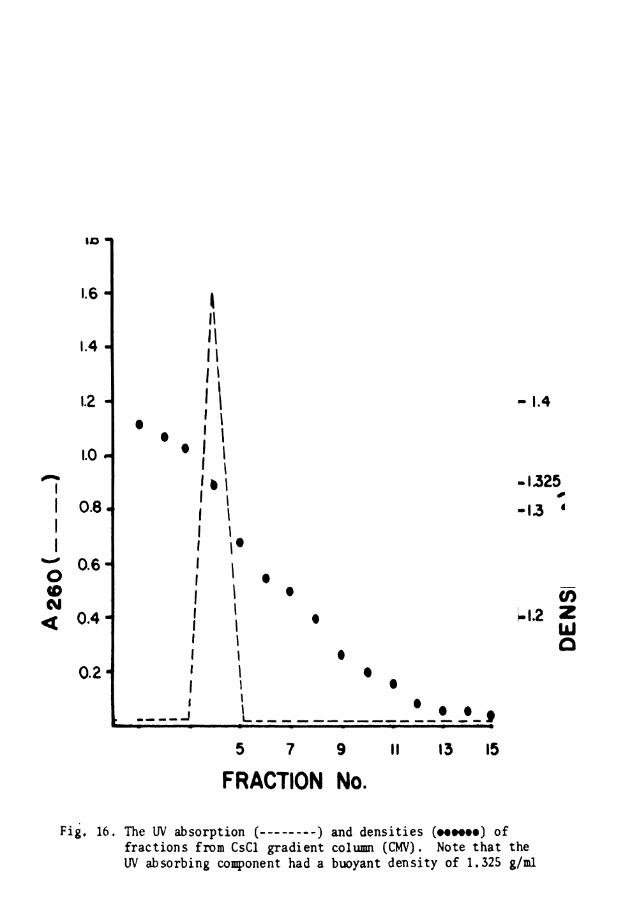
FRACTION No.
~ i g . 16, The UV absorpt ion ( - - - - - - - - ) and d e n s i t i e s (oeooee) of f r a c t i o n s from CsCl gradient column (CMV) . Note t h a t the W absorbing component had a buoyant dens i ty of 1.325 g/ml

Fig. 17. a ) The PAGE of (B) W - c o a t p r o t e i n , (C) marker p r o t e i n s and (A) coe l ec tmphores i s of B and C. Marker p r o t e i n s are: Phosphorylase B , ml . w t . 92,500(1); Bovine serum albumin, ml.wt. 66,200 ( 2 and 3 s ince t h i s m r k e r has s p l i t i n t o two); Ovalbumin, ml.wt. 45,000(4); Carbonic anhydrase, mol.wt. 31,000(5); Soybean t r y p s i n i n h i b i t o r y , ml . w t . 21.500(6) and Lysozyme, ml.wt. 14,400(7) . Note t h a t v i r u s p r o t e i n has not reso lved from carbonic anhydrase. Migration is from top t o bottom.
b) The PAGE o f CMV-RNAs, showing f o u r RNA s p e c i e s , Migration i s from t o p t o bottom.
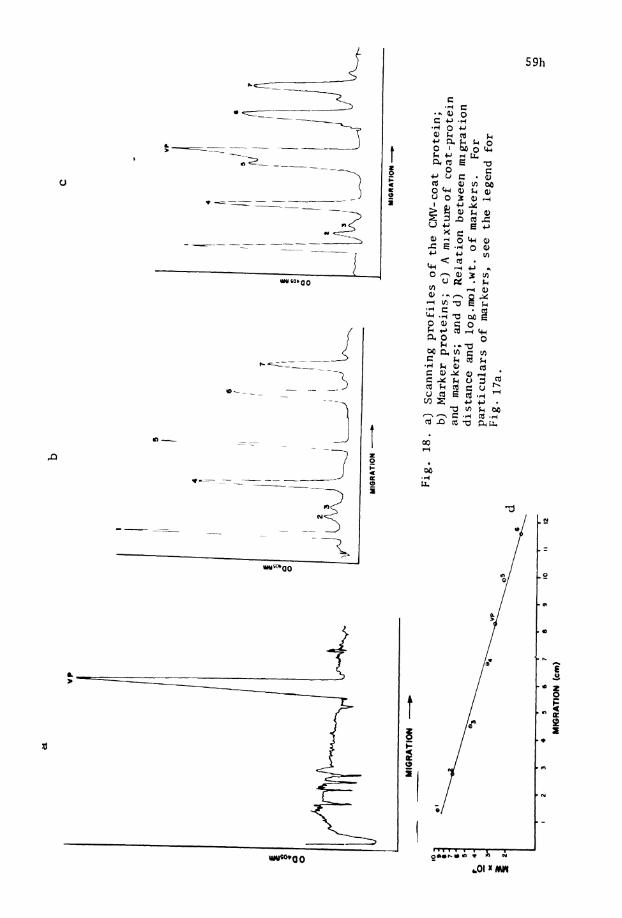

IX. PRODUCTION OF ANTISERUM TO CMV
The immunization schedule followed produced a reasonably
high t i t r e d antiserum t o CMV. In Ouchterlony double-diffusion t e s t
the t i t r e was determined t o be 1/1024 when t e s t e d wi th the p u r i f i e d
v i rus . No reac t ion could be no t i ced with t h e heal thy sap i n t h i s
t e s t . However, when t e s t e d by ELISA, a very n e g l i g i b l e p o s i t i v e
react ion was no t i ced a t a conjugate concentrat ion of 1:100; no reac t ion
could be no t i ced a t a conjugate concentra t ion o f 1:500 pig. 14) .
X. SEROLOGICAL RELATIONSHIP BETWEEN 'IHE PRESENT STRAIN OF CM, WITH OTHER KNOWN STRAINS
The i n t e n s i t i e s of the p r e c i p i t i n bands formed i n Ouchter-
lony double-diffusion t e s t when t h e p u r i f i e d CMV was made t o r e a c t
with a n t i s e r a t o d i f f e r e n t known s t r a i n s o f CMV a r e given i n Table 11.
The r e s u l t s i n d i c a t e t h a t the p resen t CMV s t r a i n i s r e l a t i v e l y more
c losely r e l a t e d t o OW-C pool, followed by CMV-IX and M-CMV.
TABLE 11. Serological a f f i n i t y of p resen t CMV s t r a i n with o the r known s t r a i n s o f CMV
- - -
S t r a i n Source o f antiserum Reaction
CMV-C pool
am- I X
CMV- S
U-CMV
X- CMV
Q- m' M-w T- CMV
Homologous
H.A. S c o t t (U.S.A.)
H.E. Watemorth (U.S.A.)
H. E . Waterworth (U.S.A.)
R. I .B . Francki (Aus t ra l i a )
R. I.B. Francki (Aus t ra l i a )
R.I.B. Francki (Aus t ra l i a )
R . I . B . Francki (Aust ra l ia)
R.I.B. Francki (Aust ra l ia)
ICRISAT

XI. SCREENING FOR DISEASE RESISTANCE
A . Cucumber Mosaic Virus ,
The data on the react ions o f chickpea l i n e s to CMV a r e
presented i n Appendix I . I t i s evident t h a t only two l i ne s , ICC-1781
and -8203, remained f r e e from CMV infec t ion , one l i n e ICC-9006 showed
10% infec t ion and the remaining 140 l i n e s showed in fec t ion ranging
from 33 t o 100%.
B . Bean Yellow Mosaic Virus
'Ihe data on the reac t ions o f t h e 106 chickpea l i n e s to BYMV
a r e presented in Appendix 11. The r e s u l t s show t h a t 9 l i ne s ; ICC-607, -6999,
-1468, -2162, -2342, -3440, -3598, -4045, and -11550, remained f ree
from BYMV i n f ec t ion , four l i n e s ; ICC-1416, -3916, -4093 and -4192,
showed 10% infec t ion , 12 l i ne s ; ICC-187, -1121, -1754, -3634, -3918,
-3919, -3921, -3969, -4188, -6989, -7000 and -8207, showed infec t ion
between 10 and 20%. A l l the remaining 81 l i n e s showed infec t ion
ranging from 22 t o 100%.

D I S C U S S I O N A N D C O N C L U S I O N
Two v i ruses , v i z . , cucumber mosaic v i r u s (CMV) and bean
yellow mosaic v i r u s ( B W ) were found t o i n f e c t chickpeas under na tu ra l
condit ions. Their i d e n t i t y was confirmed by studying t h e i r character-
i s t i c s .
A d e t a i l e d study of t h e symptomatology by CMV and BYMV i n
chickpea ind ica ted t h a t both the v i ruses caused a s i m i l a r i n i t i a l
twis t ing of t h e terminal bud though the former caused a severe
twis t ing which terminated i n nec ros i s i n c o n t r a s t with a s l i g h t twist-
ing without nec ros i s caused by t h e l a t t e r . The CMV, a f t e r terminal
nec ros i s , caused two types of symptoms. In c e r t a i n v a r i e t i e s , i t
caused w i l t i n g and death of t h e p l a n t s whereas, i n o t h e r s p r o l i f e r a t i o n
and bushiness o f branches bearing small green l eaves . The l a t t e r
were commonly seen i n mechanically inocula ted chickpea and resembled
t h e symptoms produced by CMV under f i e l d condi t ions . The i n f e c t i o n
by BYMV a f t e r i n i t i a l twis t ing was followed by p r o l i f e r a t i o n o f both
the terminal and a x i l l a r y branches which a r e long and t h i n wi th
narrow leaves . The leaves below t h e p r o l i f e r a t e d branches showed
yellowing o r i n t e r v e i n a l ch lo ros i s o r mosaic depending upon t h e
genotypes used.
'Ihe present s t r a i n of CMV appears t o have a wide host range
as i t could i n f e c t 20 p lan t species belonging t o f i v e fami l i e s ;
A i zoaceae , Chenopodiaceae, Cucurbitaceae, Legminosae, and Sol anaceae,
Though i t produced l o c a l l e s ions on severa l p l a n t s t e s t e d , C. maranti- color was found t o be very good assay hos t and was, the re fo re , used
throughout t h e s t u d i e s . E a r l i e r , (=MV was repor ted t o produce systemic
mosaic on cucumber and f e r n leaf symptoms on tomato (Francki e t al . ,
1979). On t h e contrary , t h e p resen t s t r a i n of could n o t i n f e c t
tomato and produced l a r g e c h l o r o t i c patches on cucmber i n s t e a d o f a
systemic mosaic. Waterworth and Povish (1975) a l s o got s i m i l a r r e s u l t s

with CMV-IX. However, the cu l t iva r s of cucumber and tomato used i n the
present )s tudies were d i f fe ren t from those of Waterworth and Povish.
Though tobacco and pumpkin were reported t o be good hosts f o r v i rus
pur i f i ca t ion , i t was found t h a t chickpea supported b e t t e r v i rus multi-
p l i ca t ion , and therefore was used f o r v i rus pur i f i ca t ion i n the present
s tudies .
Though BYMV i s known to i n f e c t several non-legumes (Bos, 1970a),
the present s t r a i n appears to have a very r e s t r i c t e d host range as it
could not i n f e c t any p lan t species outs ide t h e Leguminosae. I t a l s o
produced veinal necros is of inoculated leaves and top necrosis o f some
french bean cu l t iva r s . The severe yellow mosaic s t r a i n of BYMV (Rex
and Zaumeyer, 1953) which a l so produced veinal necros is and top necros is
of french beans could a l s o not i n f e c t p l a n t s outs ide Leguminosae.
Based on the r e s u l t s of i n vitro proper t ies , the present
s t r a i n of CMV appears qu i t e s table . On the contrary, BYMV appears t o
be very unstable. However, it i s evident from t h e l i t e r a t u r e t h a t
in v i t r o proper t ies g rea t ly depend on the v i r u s source and t e s t p lan t
(Bos, 1970a).
Transmission of both the v i ruses by aphids d id not exceed
45%. I t i s well documented t h a t CMV t ransmission effici,ency va r ies
with p lant species (Simons, 1955; Simons, 1957). Similar ly , with CMV
and BYMV, Kaiser and Danesh (1971b) found t h a t transmission by
d i f f e r e n t aphid species was usually l e s s than 20% when chickpea was
used both as source and t e s t p lan t , whereas it was invar iably higher
when sweet clover and cucumber were used for CMV and BYMV, respect ively .
However, i n the present s tud ies t h e e f f e c t of d i f f e r e n t host species
on the transmission e f f i c i ency of aphids was no t studied.
The pur i f i ca t ion procedure adopted f o r CMV appeared e f f i c i e n t
a s judged by the W absorption i n ISCO f r a c t i o n a t o r and spectrophoto-

meter, i n f e c t i v i t y , and e lec t ron microscopy. A s ing le , sharp peak
obta ined i n the ISCO scanning p a t t e r n was an i n d i c a t i o n of i n t a c t
v i rus , without aggregation or degradation. In i n f e c t i v i t y t e s t s , the
f r a c t i o n s p resen t only i n the W absorbing component were found
i n f e c t i v e . Thus, the re was a c l o s e c o r r e l a t i o n between W absorbing
component and i n f e c t i v i t y which revea l s t h a t t h e zone i n t h e sucrose
gradient cons i s t ed of i n f e c t i v e v i r u s . The W absorbing p a t t e r n o f
f i n a l p u r i f i e d v i r u s i n spectrophotometer was dlso typ ica l o f nucleo-
p ro te in . Considerably higher y i e l d s (80-100 mg/kg o f t i s s u e ) were
obta ined by the p u r i f i c a t i o n procedure adopted. Yields o f 100-500
mg/kg t i s s u e have been repor ted i n the l i t e r a t u r e (Waterworth and
Povish, 1975; Francki e t aZ., 1979), though t h e y i e l d depended on
v i r u s s t r a i n and host p l a n t (Francki e t aZ., 1979).
The p a r t i c l e diameter (30 nm) obta ined f o r t h e present CMV
s t r a i n was i n agreement wi th o t h e r r e p o r t s (Francki e t aZ., 1966 and
Debmt e t at., 1974).
An e f f i c i e n t p u r i f i c a t i o n procedure could no t be evolved f o r
BYMV which belongs t o the pota to v i r u s Y (potyvirus) group. Viruses
of the potyvirus group a r e d i f f i c u l t t o p u r i f y because o f t h e i r
tendency t o aggregate, both end t o end and s i d e by s i d e (Shepherd and
Pound, 1960). Nevertheless, t h e r e have been severa l r e p o r t s on t h e
p u r i f i c a t i o n o f var ious v i ruses of t h i s group. The general conclusions
from these r e p o r t s a r e t h a t only small a m u n t s o f homogenous v i r u s . prepara t ions were obta ined wi th r a t h e r complicated procedures and t h a t
t h e methods success fu l ly used wi th a p a r t i c u l a r v i r u s could not
genera l ly be app l i ed f o r p u r i f i c a t i o n o f o t h e r e longate v i ruses . In
the present s t u d i e s , var ious methods used f o r BYMV p u r i f i c a t i o n d id
not y i e l d a high p u r i t y v i r u s . However, v i r u s p repara t ion o f s a t i s -
f a c t o r y p u r i t y could be obta ined by t h e procedure adopted. Urea t o
t h e concentra t ion ranging from 0.2 t o 1M has been used i n p u r i f i c a t i o n
o f d i f f e r e n t potyviruses t o avoid aggregation of t h e p a r t i c l e s

(Damirdagh and Shepherd, 1970; Uyeda e t aZ., 1975). In t h e present
s t u d i e s , 'the aggregat ion o f BYMV p a r t i c l e s could be avoided by using
0.2M urea i n a suspending medium. However, t h e breakage o f t h e v i r u s
p a r t i c l e s could no t be avoided completely d e s p i t e t h e use o f consi-
derably low c e n t r i f u g a l fo rces . The d i f f u s e d l i g h t s c a t t e r i n g zone
obta ined a f t e r a d e n s i t y g rad ien t c e n t r i f u g a t i o n might b e due t o t h e
breakage o f v i r u s p a r t i c l e s . Withdrawal o f t h e e n t i r e d i f fused zone
might have r e s u l t e d i n t h e contamination o f t h e v i r u s wi th p lan t
c o n s t i t u e n t s . With t h e r e s u l t , t h e v i r u s f r e e of host mater ia l could
no t be obta ined. For t h i s reason, t h e physico-chemical p r o p e r t i e s
c a r r i e d out f o r CMV could no t be performed f o r t h i s v i r u s . The mdal
length obta ined f o r this v i r u s was i n agreement wi th t h a t r epor ted f o r
o t h e r s t r a i n s of BYMV (Uyeda e t aZ., 1975; Kaiser and Danesh, 1971a).
Pinwheel inc lus ions observed i n u l t r a t h i n s e c t i o n s o f BYMV-infected
chickpea leaves were t y p i c a l o f po tyv i ruses (Mar te i l i and Russo, 1977)
t o which BYMV belongs.
In Ouchterlony double-di f fus ion t e s t , CMV sometimes produced
two p r e c i p i t i n bands. S c o t t (1968) and Devergne and Cardin (1970)
a l s o observed formation o f two p r e c i p i t i n l i n e s , a curved one n e a r
t h e ant igen we l l , caused by more o r l e s s i n t a c t v i r u s , and a s t r a i g h t
one near the antiserum wel l caused by degraded v i r u s . Sodium dodecyl
s u l f a t e - t r e a t e d BYMV, however, produced on ly a s i n g l e p r e c i p i t i n band.
The reason f o r a s l i g h t l y h igher sedimenta t ion value (104s)
obta ined f o r the p resen t (SiV s t r a i n than t h e repor ted values o f 98.6
(Francki e t aZ., 1966) and 101 (Tomlinson e t aZ., 1959) i s no t c l e a r .
The buoyant d e n s i t y of 1.325 g/ml obta ined f o r t h e p resen t
CMV s t r a i n was i n c l o s e agreement w i t h t h e va lues o f 1.361 and 1.359
f o r CMV-S and O W - D s t r a i n s , r e s p e c t i v e l y (Lot and Kaper, 1976).
E s s e n t i a l l y no W absorbing mate r i a l was p r e s e n t except i n t h e v i r u s
band, i n d i c a t i n g t h e high degree o f b m y a n t homgene i ty o f t h e
p u r i f i e d product .

The m l e c u l a r weight o f the CMV coat p ro te in obtained i n the , present s tud ies (25,000) was i n c l o s e agreement with the values o f
24,500 (Habil i and Francki, 1974) and 25,200 (van Regenmrtel e t aZ.,
1972).
The PAGE o f SDS-disrupted CMV revealed t h e presence of four
RNA species . The RNAs o f CMV have been va r iab ly described a s compris-
ing 4-6 d i f f e r e n t s i zed molecules. The presence o f f i t h and s i x t h
RNA components was claimed (Kaper and West, 1972; Peden and Symns,
1973) but was disputed when these components could not be cons i s t en t ly
ext racted from d i f f e r e n t s t r a i n s of CMV (Lot e t aZ., 1974) . However,
more recent work has demonstrated t h a t a t l e a s t one o f t h e s e components
(RNA 5) i s cons i s t en t ly present i n l a r g e proportions i n CMV-W (Wood and
b u t t s , 1975), and a l s o i n s t r a i n s D and R (Lot and Kaper, 1976).
Kaper and Waterworth (1977) discovered t h a t RNA 5 i s t h e causal agent
f o r a l e t h a l necros is d isease o f tomato p l a n t s . Though the presence o f
f i f t h and s i x t h WAS was doubtful , the presence o f t h e four l a r g e s t
components has never been i n doubt (Kaper and Tousignant, 1977). The
present s t r a i n o f Q.IV a l s o contained 4 RNAs. With CMV-bmmelina s t r a i n ,
Kaper and Tousignant (1977) demonstrated t h a t more than one passage i n
tobacco was required t o b u i l d up e l e c t r o p h o r e t i c a l l y de tec tab le
q u a n t i t i e s of RNA 5 . Thus, they a t t r i b u t e d t h a t c e r t a i n s t r a i n s of
CMV may contain undetectable q u a n t i t i e s o f RNA 5 whose syn thes i s can
be generated upon propagation i n c e r t a i n h o s t s . However, t h e e f f e c t
of propagation o f CMV i n d i f f e r e n t host p l a n t s on generation o f RNA 5
was not s tud ied i n t h e present s t u d i e s .
A good t i t r e d (1/1024) antiserum t o CMV produced i n t h e
p resen t s t u d i e s ind ica ted t h a t the i m u n i z a t i o n schedule followed was
good. The immunogenicity o f CMV was repor ted t o be poor (Francki
e t al., 1979), bu t could be enhanced by f i x a t i o n wi th formaldehyde
(Francki and Hab i l i , 1972) o r glutaraldehyde (Rao, 1982) . Using
t h e f i x e d v i r u s a s an ant igen, Rao (1982) produced antiserum to s i x

s t r a i n s o f CMV with a t i t r e ranging from 1/128 t o 1/512. Tomlinson
e t aZ. (1958) produced antiserum t o unfixed CMV which had a t i t r e of
1/1024. However, they used p a r t i a l l y p u r i f i e d v i r u s and thus the
r e s u l t a n t antiserum might have contained an t ibod ies t o p l a n t prote ins
a l s o which might have contr ibuted t o the high t i t r e of antiserum. In
t h e present s t u d i e s , no react ion wi th t h e hea l thy sap was detected i n
agar double-diffusion t e s t , and hence, the reac t ion was a t t r i b u t e d
t o the v i r u s only. I n ELISA, a s l i g h t r e a c t i o n could be not iced with
t h e heal thy sap a l so . However, when the reactions t o the i n f e c t i v e and
heal thy sap were compared, the colour development i n case o f heal thy
sap was very neg l ig ib le . Even t h i s r e a c t i o n however was not observed
when conjugate concentrat ion was increased from 1: 100 t o 1:500.
The na tu ra l occurrence o f these two v i ruses was found t o be
l e s s than 1%. Considering the cropping p a t t e r n in ' Ind ia which includes
severa l cucurbitaceous and leguminous p l a n t s , which no t only harbour
these v i ruses b u t a l s o a i d i n t h e vector population buildup, the re
e x i s t s a p o t e n t i a l p o s s i b i l i t y f o r these v i ruses t o become widespread
i n the f u t u r e . The n a t u r a l occurrence o f CMV has a l ready been repor ted
by Dhingra e t aZ. (1979). Though t h e n a t u r a l occurrence o f BYMV was
no t repor ted , the survey r e s u l t s ind ica ted t h a t i t was p resen t i n
severa l p laces i n India such a s Fai zabad and Pantnagar (Ut t a r Pradesh) , Junagarh (Guj a r a t ) , Parbhani (Maharashtra) , and Tapparwaripora (Jammu
and Kashmir) (Dr. S.P.S. Beniwal, personal communication) . Therefore,
keeping t h e t h r e a t posed by t h e s e v i r u s e s i n view, the e l i t e chickpea
germplasm access ions were screened f o r r e s i s t a n c e t o these two v i ruses ,
and a number o f access ions were found f r e e from i n f e c t i o n . These
access ions a f t e r confirming t h e i r r e s i s t a n c e , could be used in chickpea
breeding program t o incorporate r e s i s t a n c e i n t o commercially acceptable
chickpea cul t i v a r s t o thwart any p o t e n t i a l danger caused by t h e s e
v i ruses t o chickpea c u l t i v a t i o n i n f u t u r e .

S U M M A R Y
Wo mechanically t r ansmiss ib le v i r u s e s , v iz . , cucumber mosaic
v i r u s (CMV) and bean yellow mosaic virus (BYMV) were found t o
n a t u r a l l y i n f e c t chickpeas. Both t h e v i r u s e s were i d e n t i f i e d by
studying t h e i r hos t range, physical p roper t i e s , i n s e c t transmission,
e l ec t ron microscopy and serology. E l i t e chickpea germplasm accessions
t h a t were e a r l i e r found t o be r e s i s t a n t t o w i l t , r o o t r o t , Ascochyta
b l i g h t and promising t o s t u n t (pea l e a f r o l l v i rus ) were screened
f o r r e s i s t a n c e t o these v i r u s e s . The r e s u l t s a r e summarized below:
1. I n i t i a l 1 y, both the v i ruses produced t w i s t i n g o f terminal
bud i n chickpea. Subsequently, CMV e i t h e r produced w i l t i n g and death
of p l a n t s o r p r o l i f e r a t i o n and bushiness of branches bearing very
s m l l green leaves and very few small pods, whereis BYMV produced
pro1 i f e r a t i o n of branches with narrow leaves , followed by production
o f d i s t o r t e d f lowers t h a t were converted i n t o very small pods having
s h r i v e l l e d and black seeds.
2 . CMV appeared t o have a wide hos t range i n f e c t i n g t h e p l a n t s
o f Aizoceae, Chenopodiaceae, Cucurbitaceae, Leguminosae and Solanaceae,
whereas BYMV could n o t i n f e c t hos t s o u t s i d e Leguminosae.
3 . The CMV had thermal i n a c t i v a t i o n p o i n t between 70' and 7S0c,
the d i l u t i o n end po in t between and and longevi ty i n v i h o
between 4 and 5 days. The corresponding f i g u r e s f o r BYMV a r e between
55' and 60°c, lo-* and and 1 and 2 days.
4. Both v i ruses could be t r ansmi t t ed by two aphid species , v i z . ,
Aphis cmccivora and Myaus persicae i n a non-pers is tent manner.
5 . The p u r i f i c a t i o n procedure developed f o r CMV yie lded 8-10
mg virus/100 g o f i n f e c t e d t i s s u e . A s a t i s f a c t o r y p u r i f i c a t i o n
procedure was a l s o developed f o r BYMV.

6. m e CMV measured about 30 nm i n diameter, whereas the length
and width of BYMV were found t o be 750 and 15 nm, respec t ive ly . Ultra-
t h i n s e c t i o n s of BYMV-infected chickpea l eaves revealed the presence
of pinwheel inc lus ions and laminate aggregates.
7. The sedimentation c o e f f i c i e n t o f p u r i f i e d CMV was determined
a s 104S, and the buoyant dens i ty a s 1.325 g/cm5.
8. The (3.IV coat p r o t e i n i s made up o f s i n g l e polypeptide wi th
a m l e c u l a r weight o f 25,000 da l tons .
The W - R N A contains four RNA spec ies .
10. An antiserum t o W produced i n a r a b b i t had a t i t r e o f
1/1024 a s determined i n agar double-dif fus ion t e s t ;
11. The p resen t chickpea s t r a i n o f CMV was found to be c lose ly
r e l a t e d t o CMV-C pool, followed by CMV-lx and M-CMV.
1 2 . Out o f 143 chickpea germplasm access ions t e s t e d f o r r e s i s -
tance t o CMV, two (ICC-1781 and -8203) remained f r e e from in fec t ion ,
one (ICC-9006) showed 10% i n f e c t i o n , whereas a l l o t h e r s showed in fec -
t i o n ranging from 33 t o 100%. For BYMV, 106 access ions were t e s t e d
and nine remained free from i n f e c t i o n , four showed 10% i n f e c t i o n ,
1 2 showed i n f e c t i o n between 10 t o 20%, whereas t h e remaining 81 showed
i n f e c t i o n ranging from 22 t o 100%.

LITERATURE CITED
ABU-SAMAH, N . , and J.W. RANDLES. 1981. A comparison of t h e nuc leo t ide
sequence homlogies o f t h r e e i s o l a t e s of bean yellow mosaic
v i ru s and t h e i r r e l a t i o n s h i p t o o t h e r potyvi ruses . Virology
110: 436-444.
ADESNIK, M. 1970. Polyacrylamide gel e l ec t rophores i s o f v i r a l RNA.
I n 'Methods i n Virology' , Vol. 5, ed. K. Maramorosch and
H. Koprowski, pp. 125-177. New York: Academic Press .
530 pp.
ALDEZ, W.C. 1959. Fac tors a f f e c t i n g t ransmiss ion o f bean yellow
mosaic v i r u s . J. Econ. Ent. 52: 260-262.
ANDERSON, C.W. 1955a. Vigna and Cro ta l a r i a v i ru ses i n F lo r ida . 11.
Notat ions concerning cowpea mosaic v i r u s (Mzrmr wigme).
Plant Dis. Reptr. 39: 349-353.
ANDERSON, C. W. 1955b. Vigna and Cro ta l a r i a v i ru ses i n F lor ida . 111.
Notat ions concerning i d e n t i f i c a t i o n d i f f i c u l t i e s , i nd i ca to r
p l an t s , pos s ib l e vec to r r e l a t i o n s h i p s and v i r u s maintenance.
P lan t Dis. Re~tr . 39: 354-357.
BALL, E.M. 1974, Semlog ica l t e s t s f o r t h e i d e n t i f i c a t i o n o f p l a n t
v i ru ses . Am.Phytopatho1. Soc., S t . Paul , 31 p.
BEEMSTER, A . B . R . and J.P.H. van de r WANT. 1951. Sero logica l i n v e s t i -
ga t ions on t h e haseolus v i r u s 1 and 2 . Antonie van Leeuwenhoek
J. Microbial. Se ro l . 17: 285-296.
BERCKS, R. 1960. Semlogische untersuchungen zur Differenziernung on
I so l a t en des Phaseolus v i r u s 2 and i h r e r Vemandtschaft mit
Phaseolus v i r u s 1. Phytopath. 2 . 39: 120-128.

BERCKS, R . 1961. Serological r e l a t i o n s h i p between potato v i rus Y , $
beet mosaic v i r u s and bean yellow mosaic v i r u s . Phytopath.
Z . 40: 357-365.
BETTO, E . , G . BELLI, G . G . CONTI, and G. GIUSSANI. 1964. On the
pur i f i ca t ion and e lect ron microscope demonstration of cucumber
mosaic v i rus . Phytopath. Z. 51: 85-89.
BHARGAVA, K.S. 1951. Some proper t i e s o f four s t r a i n s of cucumber
mosaic v i r u s . Ann. Appl. Biol. 38: 377-388.
BOS, L. 1970a. Bean yellow mosaic v i r u s . CMI/AAB Description o f
Plant Viruses No. 40.
BOS, L. 1970 b. The i d e n t i f i c a t i o n o f t h r e e new Ciruses i s o l a t e d
from Wistaria and Piswn i n t h e Netherlands and t h e problem
o f va r ia t ion i n the pota to v i r u s Y group. Neth. J. P1.
Path. 76: 8-46.
BOS, L . , D. J . HAGEDORN, and L . QUANTZ. 1960. Suggested procedures
f o r in te rna t iona l i d e n t i f i c a t i o n of legume v i ruses .
Tijdschr. P1 . Ziekt. 66: 328-343.
BRANDES, J . 1964. I d e n t i f i c a t i o n o f e longate p l a n t pathogenic v i ruses
on mrphological c r i t e r i a wi th assembly o f fundamental data
fo r t h e c l a s s i f i c a t i o n of these v i r u s e s . Mitt. b i o l . BdAnst.
Berl. 110: 130 pp.
BRANDES, J . , and L. QUANTZ. 1955. Electron microscopic s tud ies on
kidney bean v i ruses . Natumissenschaften 42 : 588.
BRIERLEY, P . , and F . F . SMIW. 1962. Three cowpea m s a i c v i ruses from
gladiolus . Plant Dis. Reptr. 46: 335-337.

BRIERLEY, P . , and R.V. TRAVIS. 1958. A v i r u l e n t s t r a i n of cucumber
'mosaic v i r u s from e a s t e r l i l y . Plant Dis. Reptr. 42: 1034-
1036.
CORBETT, M.K. 1958. A v i r u s d i s e a s e o f l u p i n s caused by bean yellow
mosaic v i rus . Phytopathology 48: 86-91.
COUSIN, R. 1969. Pea mosaic v i r u s . Comparative study o f s t r a i n s o f
pea m s a i c v i r u s and bean yellow mosaic v i r u s . Nature of
the r e s i s t a n c e t o c e r t a i n pea v a r i e t i e s t o these s t r a i n s .
Annls. Phytopath. 1: 201-212.
DAMIRDAM, I .S., and R.J. SHEPHERD. 1970. P u r i f i c a t i o n o f t h e tobacco
e tch and o t h e r v i r u s e s o f po ta to Y group. Phytopathology
60: 132-142.
DEBROT, E . A . , J.M. COSTA, and R . D . DE. UZCATEGUI. 1974. Natural
i n f e c t i o n o f Sa l i va splendens with cucumber mosaic v i r u s .
Phytopath. Z. 80: 193-198.
DEVERGNE, J.C. and L. CARDIN. 1970. Comparative se ro log ica l s tudy o f
some i s o l a t e s o f cucumber w s a i c v i r u s . Serological r e l a t i o n -
s h i p between t h e v i r u s and i t s so lub le ant igen. Annls.
Phytopath. 2: 639-661.
DEVERGNE, J.C. and L. CARDIN. 1975. Se ro log ica l r e l a t i o n s between
cucumis v i ruses (CMV, TAV, PSV). Annls. Phytopath. 7 : 255-
276.
MIINGRA, K.L. , V.V. CHENULU, md A. VERMA. 1979. A l e a f reduct ion
d i sease of Cicer arietinwn i n India , caused by a cucuw,
v i r u s . Curr. Sci. 48: 486-488,

WOLITTLE, S.P. 1916. A new in fec t ious mosaic d isease o f cucumber. $
Phytopathology 6: 145- 147.
WOLITTLE, S.P., and F.R. JONES. 1925. The mosaic d i sease i n t h e
garden pea and o t h e r legumes. Phytopathology 15: 763-771.
DOOLITTLE, S.P., and R.E. WEB. 1960. A s t r a i n of cucumis v i r u s 1
i n f e c t i o u s t o b l i g h t - r e s i s t a n t spinach. Phytopathology 50:
7-9.
DOUINE, L . , J. B. QUIOT, G . MARCHOUX and P. ARQ-IANGE. 1979. Revised
l i s t o f p lan t species suscep t ib le t o cucumber mosaic v i r u s .
Annls. Phytopath. 11: 439-475.
EL-ATTAR, S. , F.N. EL-DIN, and S,A. GHABRIAL. 1971. A s t r a i n o f
bean yellow mosaic v i r u s n a t u r a l l y occuring on broad bean
i n the Arab Republic of Egypt. Agric. Res. Rev. 49: 285-290.
ERWIN. D.C. 1958. E'usariwn l a t e r i t iwn f . c i c e r i , i n c i t a n t of Fusarium
w i l t o f Cicer arietintun. Phytopathology 48: 498-501.
ERWIN, D . C . , and W.C. SNYDER. 1958. Yellowing o f Garbanzo Beans.
Ca l i f . Agr. 12: 6, 16.
FAO. 1980. Production year book 34: 125.
F O R W I , B . , H.L, SANGER and F. CROSSMANN. 1966. Viruses on o i l
pumpkin. Nachrbl. d t . P f l a schu tzd iens t . , S t u t t g . 18: 113-116.
FRANCKI, R. I , B. 1972. P u r i f i c a t i o n o f v i r u s e s . In 'P r inc ip les and
Techniques i n Plant Virology1, ed. C.I. Kado and H.O. Agrawal,
pp. 295-335. New York: Van Nostrand - Reinhold. 688 pp.

FRANCKI, R.I.B., andN. HABILI. 1972. S t a b i l i z a t i o n of capsid
s t r u c t u r e and enhancement of immunogenicity o f cucumber
mosaic v i r u s ( Q s t r a i n ) by formaldehyde. Virology 48: 309-
315.
FRANCKI, R . I . B . , D.W. MOSSOP, and T. HATTA. 1979. Cucumber m s a i c
v i rus . CMI/AAB Description of Plant Viruses No. 213
(No. 1 rev i sed) .
FRANCKI, R. I. B . , J .W. RANDLES, 'I. C . CHAMBERS, and S.B. WILSON. 1966.
Some proper t i e s o f p u r i f i e d cucumber mosaic v i r u s (Q s t r a i n ) .
Virology 2 8 : 729-741.
FRANDSEN, N.O. 1952. Virusresistenzzuchtung b e i Phaseolus vulgaris L .
I . Phytopathologische Unterscuchungen. Zei t . Schr. Pflanzen-
zucht. 31: 381-420.
GIBBS, A.J., and B . D . HARRISON. 1970. Cucumber mosaic v i r u s . CMI/
AAB Description o f Plant Viruses No, 1.
GOEL, R . K , , and J.P. VERMA. 1973. Mosaic d isease o f r i d g e gourd
(Luf fa acutanguZa Roxb .) i n Haryana. J. Res. Haryana Agrl.
Univ. 3: 135-144.
COODING, G.V. Jr., and N . W . BING. 1970. Serological i d e n t i f i c a t i o n of
po ta to v i r u s Y and tobacco e t c h v i r u s using immunodiffusion
p l a t e s conta ining sodium dodecyl s u l f a t e . Phytopathology
60: 1293.
GOUDRIET, D.L. 1962. Eff ic iency o f var ious i n s e c t s a s v e c t o r s o f
cucumber mosaic and watermelon mosaic v i m s e s i n cantaloups . J. Ewn. Ent. 55: 519-520.

GOULD, A . R . , P. PALUKAITIS, R.H. SYMONS, and D.W. MOSSOP. 1978. I
Characterization of a s a t e l l i t e RNA associa ted with cucumber
m s a i c v i rus . Virology 84 : 443-455.
GRANETT, A.L . , and R. PROWIDENTI. 1975. P a r t i a l pur i f i ca t ion and
serological re la t ionship of th ree s t r a i n s of bean yellow
mosaic v i rus . Ann, Appl. Biol. 81: 413-415.
GROGAN, R . G . , and J.C. WALKER. 1948. I n t e r r e l a t i o n o f bean v i r u s 1
and bean v i rus 2 a s shown by cross protect ion t e s t s .
Phytopathology 38: 489-493.
HABILI, N . , and R. I. B. FRANCKI. 1974. Comparative s tudies on tomato
aspermy and cucumber mosaic v i ruses . I . Physical and
chemical proper t ies . Virology 57 : 392-401.
HABILI, N . , and R.I.B. FRANCKI. 1975. Comparative s tudies on tomato
aspermy and cucumber mosaic viruses. IV. Immunogenic and
serological proper t ies . Virology 64: 421-429.
HAMILTON, R.I. 1964. Serodiagnosis of bar ley s t r i p e mosaic f a c i l i t a t e d
by detergent. Phytopathology 54: 1290- 1291.
HANADA, K. , and H. TOCHIHARA. 1980. Genetic ana lys i s of cucumber m s a i c ,
peanut s t u n t and chrysanthemum mild ~ l lo t t l e v i ruses . Ann.
Phytopath. Soc. Japan 46: 159-168.
HARRISON, B.D. 1959, Cucumber mosaic v i r u s i n raspberry. Plant
Pathology 7: 109-111.
HEINZE, K . 1959. New vectors of pea enation mosaic v i rus and some
o the r v i ruses . Phytopath. Z . 35: 103-104.

H I L L , J . H . , and H. I . BENNER. 1976. Proper t ies of potyvirus RNAs:
Turnip mosaic, tobacco etch, and maize dwarf mosaic v i ruses .
Virology 75 : 4 19-432.
HORVAW, J., and J. SZIRM4I. 1973. Invest igat ions of a v i rus disease
of wild cucumber. Acta Phytopathologica Academiae Scientiarum
Hungaricae 8: 329-346.
HUTTINGA, H. 1973. Proper t ies of v i ruses o f potyvirus group. I .
A simple method t o pur i fy bean yellow mosaic vi rus , pea N mosaic v i r u s , l e t t u c e mosaice v i r u s and potato vi rus Y .
Neth. J. P1. Path. 79: 125-129.
HUTTINGA, H. 1975. Proper t ies o f v i ruses o f potyvirus group. 111.
A comparison o f buoyant dens i ty , S value; p a r t i c l e
wrphology and molecular weight of t h e coat protein sub-unit
o f 10 v i ruses and v i rus i s o l a t e s . Neth. J. P1. Path. 81:
58-63.
ICRISAT (Internat ional Crops Research I n s t i t u t e f o r the Semi-Arid
Tropics) . 1980. Annual Report 1978179. Patancheru, A. P.,
India: ICRISAT.
JAGGER, I.C. 1916. Experiments with t h e cucumber mosaic disease.
Phytopathology 6: 148-151.
JONES, A.T. 1976. An i s o l a t e o f cucumber m s s i c v i r u s from Rubus
phoenicolasius Maxim. Plant Pathology 25 : 137- 140.
KAISER, W .J., and D. DANESH. 1971a. Et io logy o f virus-induced w i l t
o f Cicer arietinwn, Phytopathology 61: 453-457.
KAISER, W .J., and D. DANESH. 1971b. Biology of four v i ruses a f f e c t i n g
Cicer arietinwn i n I ran . Phytopathology 61: 372-375.

KAISER, W . J . , D. DANESH, M. OKHOVAT, and H. M)SSAHEBI. 1968. Diseases
o f pulse crops (ed ib le legumes) i n I r an . Plant Dis. Reptr.
52: 687-691.
KAISER, W.J., G.M. MOSSAHEBI, and M. OKHOVAT. 1971. Al ternate hosts
of v i ruses a f f e c t i n g food legumes i n Iran. Iranian J . P1.
Path. 7: 25-29.
KANIKSKA, M. 1977. Cucumber mosaic v i r u s on gladiolus . Refe ra t imy
Zhurnal. 8: 1411.
KAPER, J.M. 1975. The chemical b a s i s o f v i r u s s t ruc tu re : Dissociat ion
and Reassembly (Fron t i e r s o f biology Vol . 39), Amsterdam:
North-Holland, 485 pp .
KAPER, J . M . , and G. G . RE. 1974. Redetermination of t h e RNA content
and the l i m i t i n g RNA s i z e of t h r e e s t r a i n s of cucumber
mosaic v i rus . Virology 60: 308-311.
KAPER, J.M., T.O. DIENER, and H.A. SCOTT. 1965. Some physical and
chemical p roper t i e s of cucumber m s a i c v i r u s ( s t r a i n Y) and
o f i t s i s o l a t e d r ibonuc le ic ac id . Virology 27: 54-72.
KAPER, J.M., and M.E. TOUSIGNANT. 1977. Cucumber mosaic v i r u s
a ssoc ia ted RNA 5. I . Role o f hos t p l a n t and helper s t r a i n
i n determining amount of assoc ia ted RNA 5 with v i r ions .
Virology 80: 186-195.
KAPER, J.M., and H.E. WATERWORTH. 1977. Cucumber mosaic v i r u s
a ssoc ia ted RNA 5: Causal agent f o r tomato nec ros i s .
Science 196: 429-431.

KAPER, J . M . , and C. K. WEST. 1972. Polyacrylamide gel separat ion and
molecular weight determination of the components of cucumber
xmsaic v i r u s WAS. Prep. Biochem. 2: 251-263.
KARL, E . 1971. New vec to rs f o r some non-pers is tent v i ruses . Arch.
Pf laschutz . 7: 337-342.
KENNEDY, J.S., M.F. DAY, and V.F. EASTOP. 1962. "A Conspectus of
Aphids a s Vectors of Plant viruses", Commonwealth I n s t .
Entomol., London, 114 pp.
KOMURO, Y. 1958. S tud ies on cucumber m s a i c v i r u s . 111. Host-range.
Ann. Phytopath. Soc. Japan 23: 235-239.
KOMURO, Y. 1973. Viruses i n Vegetables. Tokyo: ' Seibundo Shinkosha,
300 pp.
KUME, H., S . TANAKA, and D. MURAYAMA. 1970. A s t r a i n of bean yellow
xmsaic v i r u s i s o l a t e d from m s a i c diseased red clover p l a n t s .
Mem. Fac. Agric. Hokkaido Univ. 7: 434-448.
LAEMMLI, U.K. 1970. Cleavage o f s t r u c t u r a l p ro te ins during t h e
assembly of the head o f bacteriophage T4. Nature (London)
227: 680-685.
LEE', T.C., R.I.B. FRANCKI, and T. HATTA. 1979. A se r ious disease of
g lad io lus i n Aus t ra l i a caused by tomato spo t ted w i l t v i r u s .
Plant D i s . Reptr . 63: 345-348.
LISA, V . , G. BOCCARDO, G . D'AGOSTINO, G. DELLAVALLE, and M. dtAQUILIO.
1981. Character izat ion o f a potyvirus t h a t causes zucchini
yellow m s a i c . Phytopathology 71: 667-672.

LISTER, R.M. 1978. Application o f t h e enzyme-linked immunsorbent I
assay f o r d e t e c t i n g v i r u s e s i n soybean seed and p lan t s .
Phytopathology 68: 1393-1400.
LOT, H . , and J.M. KAPER. 1976. Physical and chemical d i f f e r e n t i a t i o n
of t h r e e s t r a i n s o f cucumber mosaic v i r u s and peanut s tun t
v i r u s . Virology 74: 209-22.
LOT, H . , G. MRClBUX, J. MARROU, J.M. KAPER, C.K. WEST, L . VAN VLQTEN-
DOTING, and R. HULL. 1974. Evidence f o r th ree functional
RNA species i n severa l s t r a i n s of cucumber m s a i c v i rus .
J. Gen. Viml. 22: 81-93.
LOT, H . , J. MARROU, J.B. QUIOT, and G. ESVAN. 1972. Contribution t o
t h e study o f cucumber mosaic v i r u s . I I . 'Rapid method of
v i r u s p u r i f i c a t i o n . Annls. Phytopath. 4: 25-28 .
McCLANAHAN, R. J . , and G . E . GWER. 1964. The r o l e o f i n s e c t s i n the
spread of cucumber mosaic v i r u s . Canad. J. P 1 . Sci . 44: 1-6.
McWHORTER, F . P . , and L . BOYLE. 1946. X-disease and o t h e r forms of
yellow bean mosaic i n western Oregon. Circ . Inform. 379,
Oregon a g r i c . Exp. S t a . Corva l l i s .
MALI, V.R. , and N.T. WANJANE. 1980. Dis to r t ion w s a i c - a new
v i r u s d i sease of chickpea in India . Curr. Sci . 49: 599-600.
MANGLITZ, G . R . , and K.W. KREITLOW. 1960. Vectors of a l f a l f a and
bean yellow mosaic v i r u s i n Ladino White clover. J. Econ.
Ent. 53: 113-115.
MARMOUX, G., L. WUINE, H, LOT, AND C. ESVAN. 1973. I d e n t i f i c a t i o n
e t es t imat ion dus poids moleculare des a c i d e s r ibonucleques,

due v i r u s de l a mosaique du concombre (VMC Souche D) . Comptre
Tendus hebdomadaire des seances de 1' Academie des Sciences,
P a r i s . 277: 1409-1412.
MARTELLI. G.P., and M. RUSSO. 1977. Plant v i r u s inclus ion bodies.
Adv. V i r . Res. 21: 175-266.
MATSUNAMI, M., T. SUETSUGU, and S. M3lDSHIMA. 1972. Detection of
cucumber mosaic v i r u s from pota to i n Japan. Res. Bul l .
P 1 . Prot. Ser., Japan 10: 17-22.
MAT?HEWS, R.E.F. 1979. C l a s s i f i c a t i o n and nomnclature o f v i ruses .
Third repor t o f the In te rna t iona l Committeeon Taxonomy of
Viruses. In tervi rology 12: 132-296.
MEDEIROS, A . G . 1963. Note on the occurrence of banana mosaic i n
Pernambuco s t a t e . Bolm. Tech. I n s t . Pesq. Agron. Pernambuco
4 : 13.
M3GHAL, S.M., and R . I . B . FRANCKI. 1976. Towards t h e system of the
i d e n t i f i c a t i o n and c l a s s i f i c a t i o n of potyviruses. I . Serology
and amino ac id composition o f s i x d i s t i n c t v i ruses . Virology
73: 350-362.
MOGHAL, S.M., and R.I .B . FRANCKI. 1981. Towards a system f o r the
i d e n t i f i c a t i o n and c l a s s i f i c a t i o n o f potyviruses. 11. Virus
p a r t i c l e length, symptomatology and cytopathology o f six
d i s t i n c t v i ruses . Virology 112: 210-216.
MORRIS, K. B.A.M., K.S. MIWE, and H.F. NEILSW. 1978. A s t r a i n o f
cucumber mosaic virus from Daphne. Plant Dis. Reptr. 62:
1008- 1012.

MOSSOP. D.W., and R.I.B. FRANCKI. 1977. Associa t ion o f RNA 3 with , aphid t ransmiss ion o f cucumber mosaic v i r u s . Virology 81:
177-181.
I(lOSSOP. D . W . , R. I. B. FRANCKI, and C. J . GRIVELL. 1976. Comparative
s t u d i e s on tomato aspermy and cucumber m s a i c v i r u s e s .
V. P u r i f i c a t i o n and p r o p e r t i e s o f a cucumber mosaic v i r u s
inducing severe ch lo ros i s . Virology 74: 544-546.
NEERGAARD, P. 1977. Seed Pathology, Vol. 1. London: k c m i l l a n Press
Ltd. 839 pp.
NENE, Y . L . 1980. Aworld l i s t o f pigeonpea (Ca;janus cajan ( L . ) Mi l l sp . )
and chickpea (Cicer arietinwn L. ) pathogens. ICRISAT Pulse
Pathology Progress Report no. 8, ~ a t a n c h e r u , A.P., India .
NENE, Y.L., M.P. HAWARE, and M.V. REDDY. 1978a. Diagnosis o f some
w i l t - l i k e d i so rde r s of chickpea (Cioer arietinwn L.) .
Information Bul l . No. 3, ICRISAT, Patancheru, A.P., India.
NENE, Y . L . , A. MENGISTU, J . B . SINCLAIR, and D. J. ROYSE. 1978b. An
annota ted b ib l iog raphy o f chickpea d i seases 1915-1976.
I n t e r n a t i o n a l Crops Research I n s t i t u t e f o r t he Semi-Arid
Tropics, Hyderabad, Ind ia , Information Bull. No. 1: 1-43.
NENE, Y.L., and M.V. REDDY. 1976. Pre l iminary information on chickpea
s t u n t . Tmp. Grain Legume Bu l l . 5 : 31-32.
PEDEN, K.W .C . , and R.H. SYMJNS. 1973. Cucumber mosaic v i r u s c o n t a i n s
a f u n c t i o n a l l y d iv ided genome. Virology 53: 487-492.
PIERCE, 1933, Viroses o f t he bean. Phytopathology 24: 87-115.

PRICE, W.C. 1934. I s o l a t i o n and study of some yellow s t r a i n s of 8
cucumber mosaic. Phytopathology 24: 742-761.
PRICE, W.C. 1940. Comparative hos t range of s i x p lan t v i ruses .
Am. J . Bot. 2 7 : 530-541.
FURCIFULL, D. E . , and D. L. BATCKELOR. 1977. Immunodiffusion t e s t s
with sodium dodecyl s u l f a t e (SDS) - t r e a t e d p lan t v i ruses
and p lan t v i r a l inc lus ions . Tech. Bul l . No. 788. Agric.
Expt. Sta . I n s t . Food and Agrl. Sciences, Univ. Flor ida ,
Gainesvi l le , 39 pp.
RAGOZZINO, A. 1973. A mosaic o f chard (Beta vulgaris L. , v a r . Ciela)
i n Compania. Annali d e l l a Facolta d i Science Agaric d e l l
Univers i ta d i Napoli i n P o r t i c i , 7: 57-6i .
RANDLES, J . W . , C. DAVIES, A. J. GIBBS, and T. HATTA. 1980. Amino ac id
composition o f capsid p r o t e i n a s a taxonomic c r i t e r i o n f o r
c l a s s i f y i n g t h e a typ ica l S s t r a i n o f bean yellow mosaic v i r u s .
Aust. J. Biol . Sc i . 33: 245-254.
RAO, A.L.N. 1982. Genetic s t u d i e s on cucumber mosaic and tomato
aspermy v i ruses . Ph. D. (1982) t h e s i s . Waite Agricul tura l
Research I n s t i t u t e , Univers i ty of Adelaide, South Aus t ra l i a .
REDDY, D.V.R . , R.M. LISTER, and R. RAJESWARI. 1981. Detection of
peanut mot t l e v i r u s i n peanuts employing enzyme-linked
immunosorbent assay ( i n p repara t ion) .
REX, T.H., and W.J. ZAUMEYER. 1953. A s t r a i n of yellow bean mosaic
v i r u s producing l o c a l l e s i o n s on tobacco. Phytopathology
43: 11-15.

SASTRY, K.C . , K.S.M. SASTRY, and S . J . SINGH. 1974. Studies on a v i r u s
causing mosaic d i s e a s e on Zinnia eZegans. Indian J. Mycol.
Pl . Path. 3: 165-168.
SCHMELZER, K . 1957. Contribution t o t h e knowledge of t h e transmis-
s i b i l i t y o f v i r u s e s by Cuscuta species . Phytopath. Z . 28:
1-56.
SCHMIDT, H . E . , and E . KARL. 1970. A contr ibut ion t o the a n a l y s i s of
v i r u s d i seases on hemp (Cannabis sat ivus) with regard t o the
hemp aphid (Phorodon cannabis) a s v i r u s vector . Zentbl . Bakt Paras i tkde . 125: 16-22.
SCHROEDER, W.T., and R . PROWIDENTI. ,1966. Further evidence t h a t
common pea m s a i c v i r u s (PV2) is a s t r a i n of bean yellow
mosaic v i r u s (BV2). P l a n t Dis. Reptr. 50: 337-340.
SCOTT, H.A. 1963. P u r i f i c a t i o n o f cucumber mosaic v i r u s . Virology
20: 103-106.
SCOTT, H.A. 1968. Se ro log ica l behaviour of cucumber mosaic v i r u s
( S t r a i n Y) and t h e v i r u s p r o t e i n . Virology 34: 79-90.
SHAGRUN, M . 1973. Bean yellow mosaic v i r u s on broad bean p l a n t s i n
Libya. I . I d e n t i f i c a t i o n of t h e causal agent. Libyan J.
Agric. 11: 33-38.
SHEPHERD, R . J . , and G.S. POUND. 1960. P u r i f i c a t i o n of t u r n i p mosaic
v i r u s . Phytopathology 50: 797-803.
SHOHARA, K . , and T. OSAKI. 1975. P r e c i p i t a t i o n and p u r i f i c a t i o n of
cucumber mosaic v i r u s by polyethylene glycol (PEG) and
reverse concentra t ion PEG grad ien t c e n t r i f u g a t i o n . Ann.
Phytopath. Soc. Japan 40: 265-267.

SIMONS, J.N. 1955. Some plant-vector-vi rus re la t ionsh ips of southern I
cucumber mosaic v i r u s , Phytopathology 45: 217-219.
SIMONS, J.N. 1957. Three s t r a i n s o f cucumber mosaic v i rus a f fec t ing
b e l l pepper i n t h e everglades area o f South Florida.
Phytopathology 47: 145-150.
SMITH, K.M. 1972. Text book of p lan t v i r u s diseases . Longman, London.
684 pp.
SWENSON, K . G . 1954. Aphid transmission of a bean yellow mosaic vi rus .
J. Econ. Ent. 47: 1121-1123.
SWENSON, K.G. 1957. Transmission o f bean yellow mosaic v i r u s by
aphids. J . Econ. Ent. 50: 727-731.
SWENSON, K . G . , and T.G. MARSH. 1967. Aphid transmission o f a cucumber
mosaic v i r u s from cherry. J. Econ. Ent. 60: 261-262.
SWENSON, K . G . , ar?d R.L. NELSON. 1959. Relation of aphids t o the
spread of cucumber mosaic v i r u s i n gladiolus . J . Econ. E n t .
52: 421-425.
SZALAY-MARZSO, L., and F. SOLYMOSSY. 1961. Inves t iga t ions on the
aphid vectors of t h e cucumber mosaic v i r u s on capsicum i n
Hungary. Acta Agron. Budapest 11: 329-343.
TAKANAMI, Y . , and K . TOMARU. 1969. Effect of EDTA i n cucumber mosaic
v i r u s and i t s app l ica t ion i n p u r i f i c a t i o n . Virology 37:
393-395.
TALENS, L.T. 1979. Cowpea v i ruses i n t h e Phi l ippines . 11. I s o l a t i o n
and i d e n t i f i c a t i o n o f a cowpea s t r a i n o f cucumber mosaic
v i rus . Phi l ippine Phytopathology 15: 51-57.

TAPIO, E. 1964. Virus d i seases on leguminous p lan t s . Maatalous j a , Koetoim. 18: 17-18.
TAYLOR, R . H . , and P . R . SMITH. 1968. The r e l a t i o n s h i p between bean
yellow mosaic v i r u s and pea mosaic v i rus . Aust. J. b i o l .
Sci . 21: 429-437.
THOTTAPPILLY, G . , K .F . HARRIS, and J . E . BATH. 1976. I d e n t i f i c a t i o n
of a top r o l l i n g s t r a i n of bean yellow mosaic v i r u s from
Michigan broad bean. Phytopath. 2 . 85: 183-187
TOMLINSON, J .A . , and D.G.A. WALKEY. 1967. Ef fec t s o f u l t r a son ic
treatments on t u r n i p mosaic v i r u s and pota to v i r u s X.
Virology 32: 267-268.
TOMLINSON, J .A., R. J. SHEPHERD, and J. C. WALKER. 1958. P u r i f i c a t i o n
and serology of cucumber mosaic v i r u s . Nature 182: 1616.
MMLINSON, J . A . , R . J . SHEPHERD, and J. C . WALKER. 1959. P u r i f i c a t i o n ,
p r o p e r t i e s and serology of cucumber mosaic v i r u s . Phyto-
pathology 49: 293-299.
TOPCHIISKA, M. , and I . TOPCHIISKI. 1977. I s o l a t i o n o f cucumber m s a i c
v i r u s from almond (Pncnus cmrygdaZus Stokes) i n Bulgaria.
Gradinarska b z a r s k a Nauka 13: 44-54.
TOSSO, A . B . D . , and C.V.D. VIDAL. 1976. Presence of bean yellow mosaic
v i r u s i n Chile. Agr icul tura Technica 36: 19-25.
TREMAINE, J.H. 1968. Some p r o p e r t i e s of a cucumber mosaic v i r u s
s t r a i n i s o l a t e d from Prmnus sp. Phytopathology 58: 797-801.
TU, J.C. 1980. Occurrence and i d e n t i f i c a t i o n o f a f lexuous rod v i r u s
from a m s a i c complex of white beans i n Southern Ontario.
Phytopath. 2. 99: 163-174.

WEDA, I . , M. KOJIMA, and D. MURAYAMA. 1975. P u r i f i c a t i o n and >
serology of bean yellow m s a i c v i r u s . h. Phytopath. Soc.
Japan 41: 192-203.
WEMOTO, J . K . , R. PROWIDENTI, and W . T. SBIROEDER. 1972. Sero logica l
r e l a t i o n s h i p and de t ec t ion o f bean common and bean yellow
m s a i c v i r u s e s i n a g a r g e l . Ann. Appl . Biol . 71 : 235-242.
VAN REGENMORTEL, M . H . V . , D.A. HENDRY, and T. BALTZ. 1972. A re -
examination o f t he molecular s i z e of cucumber mosaic v i r u s
and i t s coa t p r o t e i n . Virology 49: 647-653.
VOVK, A.M. 1956. Cucumber mosaic v i r u s . Methods o f spread and
measures f o r c o n t r o l . Trud. I n s t . Genet. 23: 296-310.
WATERWORTH, H . E . , and W.R. POVISH. 1975. A v i r u s r e l a t e d t o cucumber
m s a i c v i r u s i s o l a t e d from imported Ixora p l a n t . Phytopath-
010gy 65: 728-729.
WILLISON, R.S., and M. WEINTRAUB. 1957. P r o p e r t i e s o f a s t r a i n of
cucumber mosaic v i r u s i s o l a t e d from prunus hos t s . Canad.
J. bt. 35: 763-771.
WOOD, K.R. , and R.H.A. COUTTS. 1975. P r e l i n a r y s t u d i e s on the RNA
components of t h r e e s t r a i n s o f cucumber m s a i c v i r u s .
Physiol . P l a n t . Pa tho l . 7: 139-145.
YILMAZ, M.A. 1979. Determination of molecular weight o f p r o t e i n s
f o r bean yellow m s a i c v i r u s by polyacrylamide ge l e l e c t r o -
phores i s . Journal o f Turkish Phytopathology 8: 97-100
ZAUMEYER, W .J., and H.H. FISHER. 1953. A new n e c r o t i c - l e s i o n producing
s t r a i n o f yellow bean msaic. Phytopathology 43: 45-49.

ZAUMEYER, W.J., and R.W. GOTH. 1963. Red c lover n e c r o t i c virus, t h e
c'ause of a s treak of peas. P l a n t Dis. Reptr. 47 : 10-14.
A P P E N D I X
I . Reaction of chickpea l i n e s t o cucumber mosaic v i r u s
S1. Number Number Percent No. ICC No. i n o c u l a t e d i n f e c t e d i n f e c t i o n
9 9 0
contd
cont d .
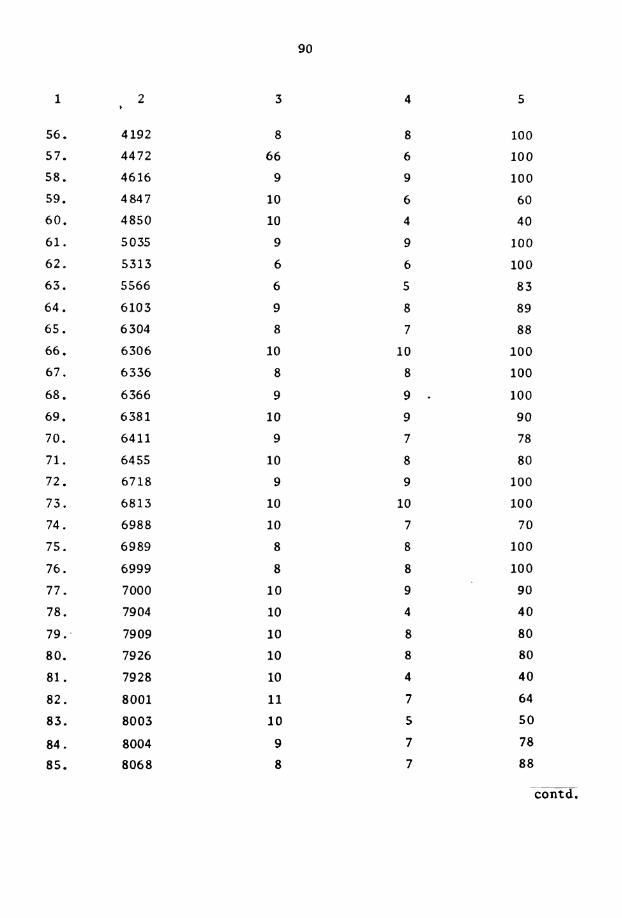
contd.
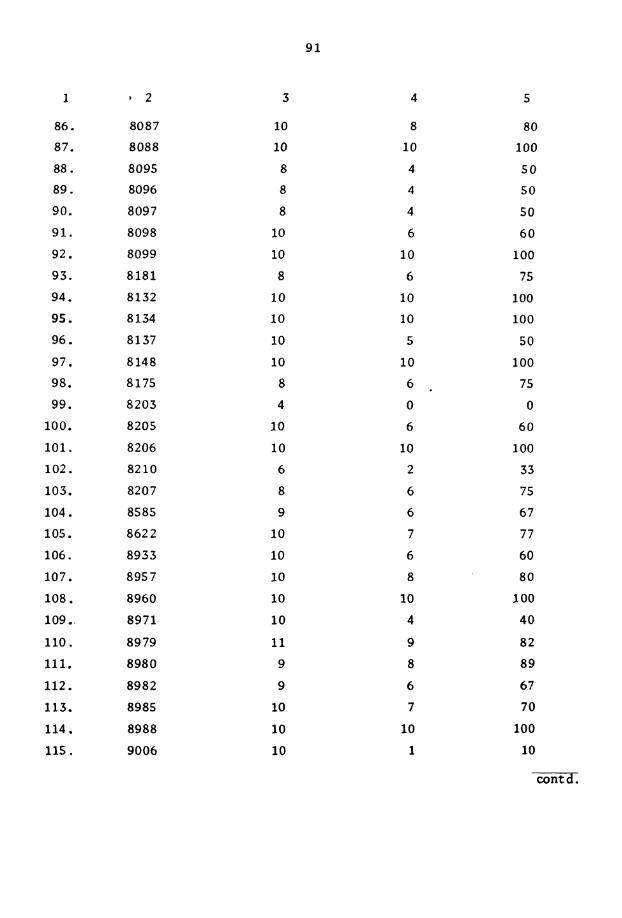
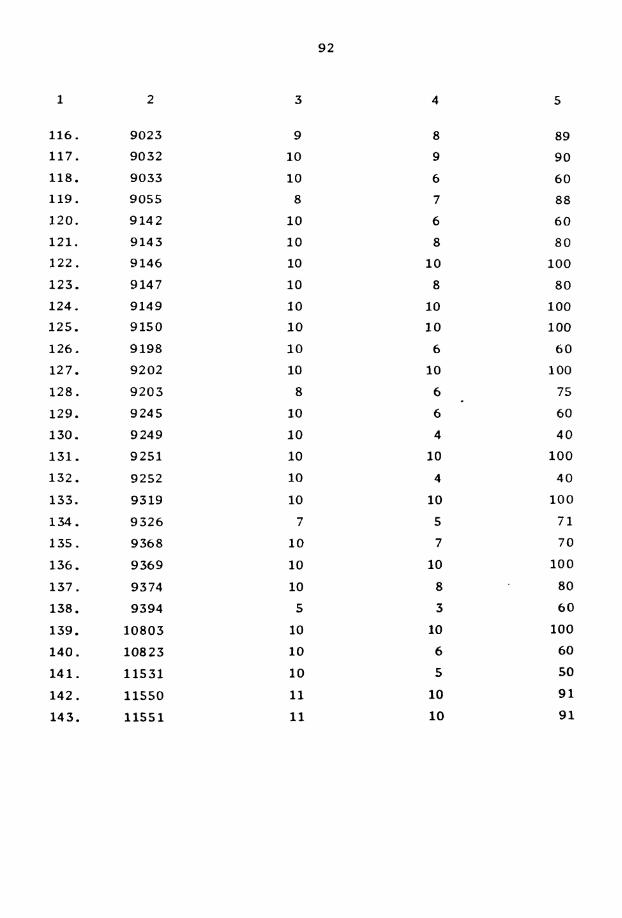
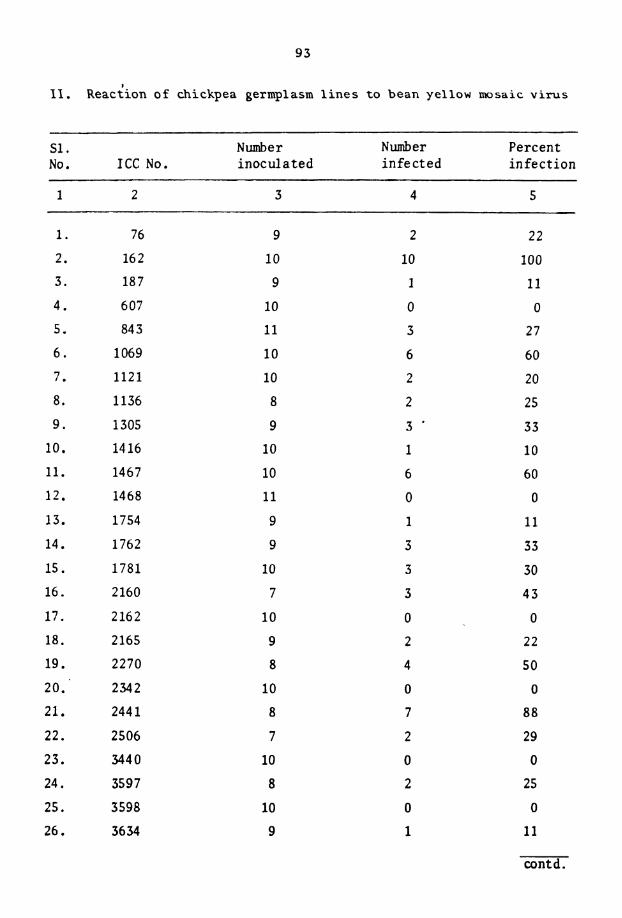
1
II. Reaction o f chickpea germplasm l i n e s t o bean yellow mosaic virus
S l . Number Number Percent No. ICC No. inocu la ted i n f e c t e d i n f e c t i o n
contd.
contd.
contd.

I , T. Venkata Chalam, was born on 1 August 1949 t o
Smt. Kesamma and S r i Narasaiah, i n Cwnbum, Prakasam D i s t r i c t , Andhra
Pradesh, Ind i a . I marr ied Miss S a v i t h r i o f Gadwal on 1 May 1977 and
we have one son*. I ob ta ined my B.Sc. degree in Agr i cu l tu re from
Andhra Pradesh A g r i c u l t u r a l Un ive r s i t y , Hyderabad i n 1970 and was
awarded my M.Sc. degree i n Agr i cu l tu re (P l an t Pathology) by G . B. Pant
Univers i ty o f Agr i cu l tu re and Te,:hnology, Pantnagar i n 1973.
I am on r e g u l a r s e r v i c e (Ass i s t an t P ro fe s so r ) o f Andhra
Pradesh A g r i c u l t u r a l Univers i ty . I j o ined t h e Ph. D . program (on s tudy
leave) i n 1979 and worked i n p l a n t v i ro logy under t h e guidance of
D r . Y .La Nene of ICRISAT and D r . J. Subbayya of APAU. During t h e course
o f t h i s i n v e s t i g a t i o n , I r ece ived ICRISAT fe l l owsh ip .