Embed Size (px)
Citation preview

Steroids 70 (2005) 458–463
Identification and characterization of 1,25D3-membrane-associated rapidresponse, steroid (1,25D3-MARRS)-binding protein in rat IEC-6 cells
Benjamin Rohea, Susan E. Saffordb, Ilka Nemerec, Mary C. Farach-Carsona,∗a Department of Biological Sciences, University of Delaware, Newark, DE 19716, USA
b Department of Biology, Lincoln University, PA, USAc Department of Nutrition and Food Sciences, Utah State University, Logan, UT, USA
Available online 1 April 2005
Abstract
We report the presence of a mammalian equivalent of the avianMembrane-AssociatedRapidResponse,Steroid (1,25D3-MARRS)-bindingprotein specific for 1,25(OH)2D3 in a rat small intestinal cell line, IEC-6, that demonstrates rapid responses to the steroid hormone. Identificationof transcript and protein was achieved using RT-PCR with several specific primer sets, Western blot analysis with two separate antibodiesrecognizing distinct regions of the protein, ribozyme knockdown and immunohistochemistry. Promoter analysis of the 1000-bp upstream regiono ′ oc tels ,25DM Togethertp es.©
K
1
[n1daC1otmps
rtherity
rallls
a 5ranein
ealentnals to
sencer
0d
f the 1,25D3-MARRS gene in several species revealed the presence of a conserved smad-3 element in the 5proximal promoter region, but nlassical vitamin D response element (VDRE). Treatment of IEC-6 cells with transforming growth factor�1 (TGF�1) increased steady-staevels of 1,25D3-MARRS (mRNA and protein) approximately two-fold over a 24-h period. In contrast, treatment with 1,25(OH)2D3 failed toignificantly change 1,25D3-MARRS protein or mRNA levels. Localization studies showed rapid nuclear translocation of a pool of 13-ARRS protein after 1,25(OH)2D3 treatment, suggesting that the protein is subject to membrane-initiated signal pathway activation.
hese data point to complex interactions between the two important 1,25(OH)2D3 sensitive response systems in intestinal cells, 1,25D3-MARRSrotein and the well-studied nVDR, that together work to fine tune intestinal Ca2+ absorption in a variety of avian and mammalian speci2005 Elsevier Inc. All rights reserved.
eywords:Steroid; Vitamin D3; Intestine; Calcium absorption; 1,25D3-MARRS; Rapid response
. Introduction
The nuclear receptor for 1,25-dihydroxyvitamin D31,25(OH)2D3] plays a well-characterized role in the mainte-ance of systemic Ca2+ homeostasis. Mice null for the nuclear,25-vitamin D3 receptor (nVDR) display numerous skeletalefects secondary to reduced intestinal absorption of Ca2+
nd hypocalcemia that largely can be rescued by elevation ofa2+ in the diet[1]. Intestinal uptake of Ca2+ stimulated by,25(OH)2D3 also requires activation of a rapid response thatccurs more quickly than transcriptional regulation through
he nVDR[2]. We recently reported the identification of aembrane-linked, rapidly reacting steroid hormone-bindingrotein that is involved in 1,25(OH)2D3-stimulated ion ab-orption in the intact chick intestine[3]. The 1,25D3-MARRS
∗ Corresponding author. Tel.: +1 302 831 2277; fax: +1 302 831 2281.E-mail address:[email protected] (M.C. Farach-Carson).
(Membrane-AssociatedRapidResponse,Steroid specific fo1,25(OH)2D3) protein bears no sequence similarity tonVDR, but the central domain shares regional similato a class of intracellular 1,25(OH)2D3-binding proteinsidentified in rickets-prone New World primates[4].1,25D3-MARRS protein was first purified from basolatemembranes (BLM) of chicken intestinal epithelial ceand the transcript encoding it was found to include ′untranslated region, a signal peptide, no transmembdomain and two thioredoxin motifs known to be involvedprotein interactions including activation of NF-�B (via relhomology domains) in the nucleus[5]. We now report thpresence and characterization of a mammalian equivto 1,25D3-MARRS protein present in a rat small intesticell line, IEC-6, that demonstrates rapid response1,25(OH)2D3 [6]. Promoter analysis of 1,25D3-MARRSgene in several mammalian species revealed the preof a conserved smad-3 element in the 5′ proximal promote
039-128X/$ – see front matter © 2005 Elsevier Inc. All rights reserved.oi:10.1016/j.steroids.2005.02.016

B. Rohe et al. / Steroids 70 (2005) 458–463 459
region, but no vitamin D response element (VDRE). Thisreport describes the results of our initial analysis of effects oftreatment of IEC-6 cells with transforming growth factor�1(TGF�1) and 1,25(OH)2D3 on 1,25D3-MARRS expressionand localization.
2. Experimental
2.1. Cell culture
Rat intestinal epithelial IEC-6 and IEC-18 cell lines werepurchased from ATCC (Manassas, VA) and maintained inDMEM with 5% (v/v) heat-inactivated fetal bovine serum(FBS). MG63 human osteosarcoma cells were purchasedfrom ATCC and maintained in F12 media with 10% FBS.MC3T3 E1 (subclone 14) mouse pre-osteoblastic cells werepurchased from ATCC and maintained in� MEM with 10%FBS and 1% HEPES. Cells were passaged at 80% confluencywith trypsin/EDTA. Cells were incubated at 37◦C with 5%CO2. All media were purchased from Gibco Invitrogen Cor-poration (Carslbad, CA). Cells were withdrawn from serum24 h prior to treatment. Treatments were performed in serum-free medium at indicated concentrations.
2.2. RT-PCR
aseQ meduQ d us-i n.S ibcoI al-t boveu db
2
awnf ents.P ice.C cmflR e for3 wasc t− y kita mso (w/v)B nft ak-i
transfer buffer with 10% methanol was accomplished at 20 Vfor 20 min. Blots were blocked overnight in 5% (w/v) nonfatdry milk in 0.01% (v/v) PBS-Tween 20 (PBST20) at 40◦C.Ab099 or ERp57 antibodies (Affinity Bioreagents, Golden,CO) were added to the block buffer at 1:2500 and incubatedfor 2 h with shaking. Blots were washed 3X in PBST20 for5 min with shaking. Goat anti-rabbit HRP-conjugated sec-ondary antibody (Pierce) was added at 1:200,000 in PBST20and incubated for 2 h with shaking. Blots then were washed3X in PBST20 for 5 min with shaking and incubated inSuperSignal® West Dura Extended Duration Substrate fromPierce. Blots then were exposed to film. Five micrograms ofBLM from chicken intestine was used as positive control forAB099. All steps were performed at room temperature unlessotherwise indicated.
2.4. Ribozyme production and transfection
1,25D3-MARRS mRNA was analyzed and ribozyme wasproduced and tested in vitro as previously described[3,7].IEC-6 cells were transfected using LipofectamineTM reagentand protocol from Invitrogen. Colonies were grown and se-lected in media containing 250�g/ml ZeocinTM (Invitrogen).
2.5. Gene and promoter analysis
n1 ivep ) wasoe ati-cg
2
-s ith-d d in2 henw 1%s d in0 for5 /PBS3 on-c ht at4 3w nti-b tiono erew k.T con-c thed -i . All
Cells were grown to 80% confluency and total RNA wxtracted by RNAeasy Mini-prep® kits and protocol fromiagen (Valencia, CA). Reverse transcription was perforsing 500-ng RNA and OmniscriptTM kit and protocol fromiagen. Polymerase chain reaction (PCR) was performe
ng Hot Star TaqTM polymerase kit and protocol from Qiageamples were electrophoresed on 1.2% (w/v) agarose (G
nvitrogen Corporation) in TAE at 90 V for 45–60 min. Reime reverse transcription (RT)-PCR was performed as asing QuantitectTM SYBR® Green kit and protocol suppliey Qiagen.
.3. Western blotting
Cells were grown to 80% confluency and serum withdror 24 h then treated as described in individual experimrotein extraction was performed using RIPA buffer onells were washed with PBS twice, then cells in T-75asks received 500�l and 6-well plates received 100�l coldIPA buffer. Flasks and plates then were incubated on ic0 min and cells were scraped to collect extract. Extractentrifuged at 13,000 rpm for 20 min at 4◦C and stored a20◦C. Protein was quantified using BCA Protein assand protocol from Pierce (Rockford, IL). Thirty micrograf total protein was loaded and electrophoresed on 10%IS/TRIS NupageTM gels with MOPS buffer from Invitroge
or 40 min at 200 V. Gels were equilibrated in 2X NupageTM
ransfer buffer with 10% (v/v) methanol for 15 min with shng. Semi-dry transfer onto nitrocellulose in 2X NupageTM
Gene locus location was obtained by OMIM (http://www.cbi.nlm.nih.gov/entrez/query.fcgi?db=OMIM). The first000 bp immediately 5′ of the translational start site (putatromoter region) for three species (human, rat, mousebtained using the Ensembl Genome Browser (http://www.nsembl.org/). The putative promoter region was systemally analyzed using Genomatix MatInspector (http://www.enomatix.de/products/MatInspector/index.html).
.6. Immunohistochemistry
IEC-6 cells were plated onto poly-l-lysine-coated tisue culture slides overnight in complete medium, then wrawn from serum for 24 h prior to study. Cells were fixe% (v/v) paraformaldehyde in PBS on ice for 15 min, tashed 3× with PBS for 5 min and washed once with 0.0odium azide in PBS for 10 min. Cells were permeabilize.2% (v/v) Triton X-100/PBS with 1% (v/v) goat serummin. Slides then were washed in 1% normal goat serumX for 5 min. Ab099 primary antibody was added at 1:50 centration in 1% goat serum/PBS and incubated overnig0◦C. Slides then were washed in 1% goat serum/PBS×ith shaking. FITC-conjugated anti-rabbit secondary aody (Sigma, St. Louis, MO) was added at a concentraf 1:25 in PBS and incubated in the dark for 1 h. Slides washed 3× with PBS for 5 min with shaking in the darOPRO 3 nuclear stain was added to the slides at aentration of 1:4000 in PBS and incubated for 15 min inark. Slides were washed 3× with PBS for 5 min with shak
ng in the dark and then imaged by confocal microscopy
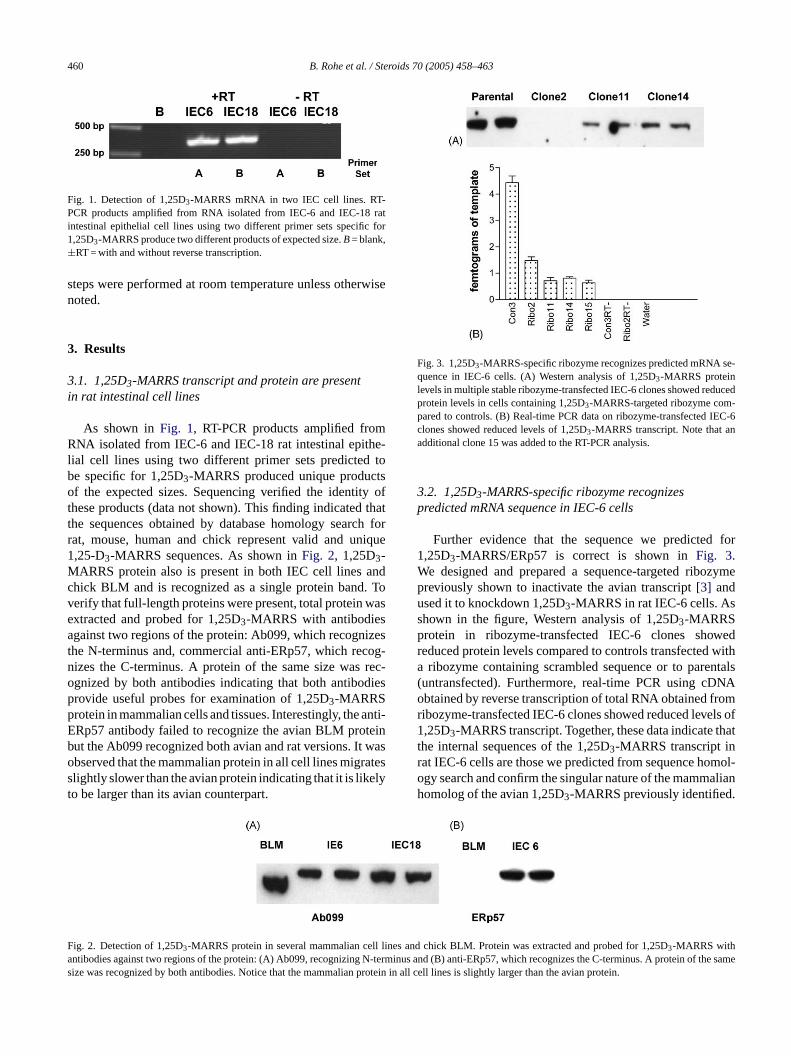
460 B. Rohe et al. / Steroids 70 (2005) 458–463
Fig. 1. Detection of 1,25D3-MARRS mRNA in two IEC cell lines. RT-PCR products amplified from RNA isolated from IEC-6 and IEC-18 ratintestinal epithelial cell lines using two different primer sets specific for1,25D3-MARRS produce two different products of expected size.B= blank,±RT = with and without reverse transcription.
steps were performed at room temperature unless otherwisenoted.
3. Results
3.1. 1,25D3-MARRS transcript and protein are presentin rat intestinal cell lines
As shown inFig. 1, RT-PCR products amplified fromRNA isolated from IEC-6 and IEC-18 rat intestinal epithe-lial cell lines using two different primer sets predicted tobe specific for 1,25D3-MARRS produced unique productsof the expected sizes. Sequencing verified the identity ofthese products (data not shown). This finding indicated thatthe sequences obtained by database homology search forrat, mouse, human and chick represent valid and unique1,25-D3-MARRS sequences. As shown inFig. 2, 1,25D3-MARRS protein also is present in both IEC cell lines andchick BLM and is recognized as a single protein band. Toverify that full-length proteins were present, total protein wasextracted and probed for 1,25D3-MARRS with antibodiesagainst two regions of the protein: Ab099, which recognizesthe N-terminus and, commercial anti-ERp57, which recog-nizes the C-terminus. A protein of the same size was rec-ognized by both antibodies indicating that both antibodiespp anti-E teinb waso atess elyt
Fig. 3. 1,25D3-MARRS-specific ribozyme recognizes predicted mRNA se-quence in IEC-6 cells. (A) Western analysis of 1,25D3-MARRS proteinlevels in multiple stable ribozyme-transfected IEC-6 clones showed reducedprotein levels in cells containing 1,25D3-MARRS-targeted ribozyme com-pared to controls. (B) Real-time PCR data on ribozyme-transfected IEC-6clones showed reduced levels of 1,25D3-MARRS transcript. Note that anadditional clone 15 was added to the RT-PCR analysis.
3.2. 1,25D3-MARRS-specific ribozyme recognizespredicted mRNA sequence in IEC-6 cells
Further evidence that the sequence we predicted for1,25D3-MARRS/ERp57 is correct is shown inFig. 3.We designed and prepared a sequence-targeted ribozymepreviously shown to inactivate the avian transcript[3] andused it to knockdown 1,25D3-MARRS in rat IEC-6 cells. Asshown in the figure, Western analysis of 1,25D3-MARRSprotein in ribozyme-transfected IEC-6 clones showedreduced protein levels compared to controls transfected witha ribozyme containing scrambled sequence or to parentals(untransfected). Furthermore, real-time PCR using cDNAobtained by reverse transcription of total RNA obtained fromribozyme-transfected IEC-6 clones showed reduced levels of1,25D3-MARRS transcript. Together, these data indicate thatthe internal sequences of the 1,25D3-MARRS transcript inrat IEC-6 cells are those we predicted from sequence homol-ogy search and confirm the singular nature of the mammalianhomolog of the avian 1,25D3-MARRS previously identified.
F ll linesa -termi of ts rotein
rovide useful probes for examination of 1,25D3-MARRSrotein in mammalian cells and tissues. Interestingly, theRp57 antibody failed to recognize the avian BLM prout the Ab099 recognized both avian and rat versions. Itbserved that the mammalian protein in all cell lines migrlightly slower than the avian protein indicating that it is liko be larger than its avian counterpart.
ig. 2. Detection of 1,25D3-MARRS protein in several mammalian centibodies against two regions of the protein: (A) Ab099, recognizing Nize was recognized by both antibodies. Notice that the mammalian p
and chick BLM. Protein was extracted and probed for 1,25D3-MARRS withnus and (B) anti-ERp57, which recognizes the C-terminus. A proteinhe samein all cell lines is slightly larger than the avian protein.
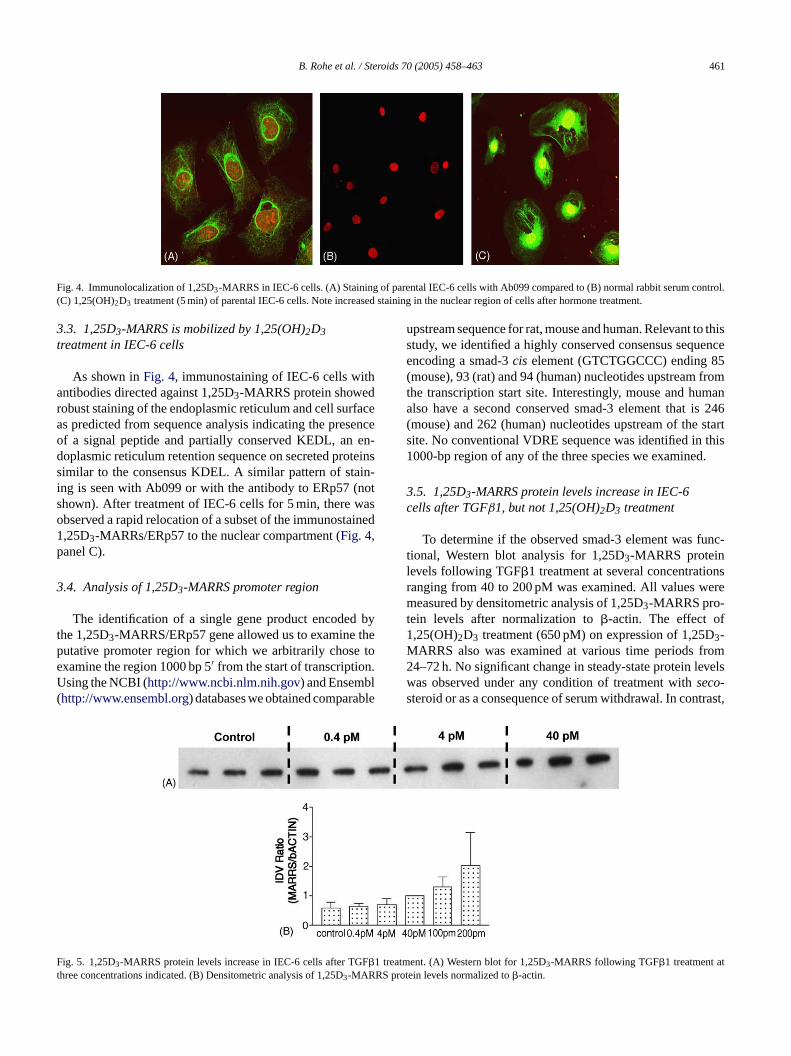
B. Rohe et al. / Steroids 70 (2005) 458–463 461
Fig. 4. Immunolocalization of 1,25D3-MARRS in IEC-6 cells. (A) Staining of parental IEC-6 cells with Ab099 compared to (B) normal rabbit serum control.(C) 1,25(OH)2D3 treatment (5 min) of parental IEC-6 cells. Note increased staining in the nuclear region of cells after hormone treatment.
3.3. 1,25D3-MARRS is mobilized by 1,25(OH)2D3treatment in IEC-6 cells
As shown inFig. 4, immunostaining of IEC-6 cells withantibodies directed against 1,25D3-MARRS protein showedrobust staining of the endoplasmic reticulum and cell surfaceas predicted from sequence analysis indicating the presenceof a signal peptide and partially conserved KEDL, an en-doplasmic reticulum retention sequence on secreted proteinssimilar to the consensus KDEL. A similar pattern of stain-ing is seen with Ab099 or with the antibody to ERp57 (notshown). After treatment of IEC-6 cells for 5 min, there wasobserved a rapid relocation of a subset of the immunostained1,25D3-MARRs/ERp57 to the nuclear compartment (Fig. 4,panel C).
3.4. Analysis of 1,25D3-MARRS promoter region
The identification of a single gene product encoded bythe 1,25D3-MARRS/ERp57 gene allowed us to examine theputative promoter region for which we arbitrarily chose toexamine the region 1000 bp 5′ from the start of transcription.Using the NCBI (http://www.ncbi.nlm.nih.gov) and Ensembl(http://www.ensembl.org) databases we obtained comparable
upstream sequence for rat, mouse and human. Relevant to thisstudy, we identified a highly conserved consensus sequenceencoding a smad-3cis element (GTCTGGCCC) ending 85(mouse), 93 (rat) and 94 (human) nucleotides upstream fromthe transcription start site. Interestingly, mouse and humanalso have a second conserved smad-3 element that is 246(mouse) and 262 (human) nucleotides upstream of the startsite. No conventional VDRE sequence was identified in this1000-bp region of any of the three species we examined.
3.5. 1,25D3-MARRS protein levels increase in IEC-6cells after TGFβ1, but not 1,25(OH)2D3 treatment
To determine if the observed smad-3 element was func-tional, Western blot analysis for 1,25D3-MARRS proteinlevels following TGF�1 treatment at several concentrationsranging from 40 to 200 pM was examined. All values weremeasured by densitometric analysis of 1,25D3-MARRS pro-tein levels after normalization to�-actin. The effect of1,25(OH)2D3 treatment (650 pM) on expression of 1,25D3-MARRS also was examined at various time periods from24–72 h. No significant change in steady-state protein levelswas observed under any condition of treatment withseco-steroid or as a consequence of serum withdrawal. In contrast,
F Ftreatm tt S pro
ig. 5. 1,25D3-MARRS protein levels increase in IEC-6 cells after TG�1hree concentrations indicated. (B) Densitometric analysis of 1,25D3-MARR
ent. (A) Western blot for 1,25D3-MARRS following TGF�1 treatment atein levels normalized to�-actin.

462 B. Rohe et al. / Steroids 70 (2005) 458–463
Fig. 6. 1,25D3-MARRS transcript levels increase in IEC-6 cells after TGF�1treatment. Three concentrations of TGF�1 were added to IEC-6 cells asdescribed Section2. Controls without reverse transcription (−RT) are alsoshown. Approximately a two-fold increase in steady-state mRNA levels wasobserved after treatment with TGF�1. Control = vehicle treated.
as shown inFig. 5, steady-state protein levels increased two-fold at the lower concentrations (40 pM) and three-fold at thehigher concentration (200 pM) of TGF�1 that we tested.
3.6. 1,25D3-MARRS transcript levels increase in IEC-6cells after TGFβ1 treatment
To determine if the increased protein levels were corre-lated with increases in steady-state levels of mRNA encod-ing 1,25D3-MARRS, TGF�1 was added to IEC-6 cells asdescribed in Section2. Three concentrations were tested tocoincide with the protein studies, and RNA was collectedfor real-time RT-PCR. Controls without reverse transcriptionalso were performed. As shown inFig. 6, approximately atwo-fold increase in steady-state mRNA levels was observedafter treatment with TGF�1 at doses of 100–200 pM. No con-sistent change was observed at the lower dose of 40 pM.
4. Discussion
The function of the nuclear receptor for 1,25(OH)2D3,nVDR, as a transcriptional regulator is reasonably wellunderstood. However, the responses of target cells to1,25(OH)2D3 are too complex to be explained purely bythe classical actions of the nVDR. It has been suggestedt dings onses[ cificc idap nalci al,r 46)[ gnalp ilar-
ity to the nVDR[3,5]. In this paper, we report the furthercharacterization of 1,25D3-MARRS in two 1,25(OH)2D3responsive rat intestinal cell lines[6] using RT-PCR,Western blot analysis, and immunolocalization. We alsoreport a limited analysis of the promoter region and potentialregulation of abundance by 1,25(OH)2D3 and TGF�1.
The rat protein is slightly larger than the avian proteinwhen assessed by SDS-PAGE, which may indicate additionalpost-translational modifications are present. Bioinformaticanalysis revealed numerous potential sites for modificationincluding those by several protein kinases, as well as forlipid addition [5]. Studies to examine this are presentlyunderway. The size differences cannot be explained by theprimary structure, since both the chicken and rat versionsof 1,25D3-MARRS consist of 505 amino acids from startmethionine to C-terminus. The chicken protein terminates ina KEDL sequence, but the rat protein terminates in a QEDLsequence, interestingly as do the homologues in mice andhuman which have 504 amino acids. This divergence at theC-terminus may explain the failure of the ERp57 antibodyto recognize the chicken protein as shown inFig. 2. There isan overall 79.6% sequence identity at the amino acid levelbetween chicken and rat.
Treatment with vitamin D hormone triggers rapid nucleartranslocation but does not elevate transcript or protein lev-els. In untreated cells, a majority of the protein resides int n oft enr ulumr heKM lized[w rp fac-t itst
o-s t. Ex-a oft rveds nsis-ti 5DM teinl e thatw n as-s ffectso
rmeda 5DM ellswI DMt f in-
hat a novel membrane-associated, rapidly responteroid hormone receptor must mediate acute resp8] Alternatively, it has been proposed that context-speonformational flexibility of the nVDR is linked to rapctions of the hormone[9]. A novel 1,25(OH)2D3-bindingrotein was recently identified in the BLM of chick intestiells and termed 1,25D3-MARRS [3,5]. 1,25D3-MARRS
s identical to a previously identified multifunctionedox-sensitive protein named ERp57 (OMIM #60203,10]. 1,25D3-MARRS sequence analysis revealed a sieptide, no transmembrane domain and no obvious sim
he endoplasmic reticulum, consistent with the functiohe K(Q)EDL. This variant of the classical KDEL has beeported to function as an alternative endoplasmic reticetention sequence[11], but it may not be as efficient as tDEL, thus explaining transit of a portion of the 1,25D3-ARRS protein to the cell surface where it has been loca
12]. Nuclear translocation after hormone treatment[12,thisork] could permit 1,25D3-MARRS to interact with otheroteins present in the nucleus including transcription
ors. It is intriguing to speculate that this might occur viawo rel interacting domains[3,5].
The gene encoding 1,25D3-MARRS resides at chromome 15p15 in human, and at chromosome 3q35 in ramination of the 1000-bp region immediately upstream
he transcriptional start site indicated that a highly consemad-3 consensus element is present, but no VDRE. Coent with this, we found that TGF�1, but not 1,25(OH)2D3,ncreased steady-state transcript and protein levels of 1,23-
ARRS. TGF�1 increased steady-state mRNA and proevels 2–3-fold in a dose-dependent manner. The increase observed was slightly higher at the 200 pM dose wheessed at the protein level, but we have not examined ef TGF�1 on protein stability.
When taken together, the analyses that we have perfot the gene, transcript and protein levels indicate that 1,23-ARRS is present in a variety of mammalian and avian chere it can participate in rapid responses to 1,25(OH)2D3.
n this context, we speculate that upregulation of 1,253-ARRS by physiological concentrations of TGF�1 acting
hrough a smad-3 element might increase sensitivity o

B. Rohe et al. / Steroids 70 (2005) 458–463 463
testinal cells to 1,25(OH)2D3 and contribute to increasedcalcium absorption. Of interest, smad-3-deficient mice wereshown recently to have higher proliferation rates in intesti-nal mucosa than their wild-type counterparts[13], consistentwith a role that we might envision for 1,25D3-MARRS inmaintaining differentiated functions of intestinal epithelialcells. Experiments to further assess the function of the smad-3 element in IEC-6 cells are underway.
Acknowledgments
This work was supported in part by a grant from theNIDCR DE12641 (MCF-C), in part by USDA/CSREES2004-35206-14134 (IN) and in part by NIAMS AR48554(SES).
References
[1] Amling M, Priemel M, Holzmann T, Chapin K, Rueger JM, BaronR, et al. Rescue of the skeletal phenotype of vitamin D receptor-ablated mice in the setting of normal mineral ion homeostasis: for-mal histomorphometric and biomechanical analyses. Endocrinology1999;140(11):4982–7.
[2] Nemere I, Norman AW. Transcaltachia, vesicular calcium trans-port, and microtubule-associated calbindin-D28K: emerging views
on.
anks
a 1,25(OH)2D3 membrane binding protein (1,25D3-MARRS) andphosphate uptake in intestinal cells. Proc Natl Acad Sci USA2004;101(19):7392–7.
[4] Adams JS, Chen H, Chun RF, Nguyen L, Wu S, Ren SY, et al. Novelregulators of vitamin D action and metabolism: lessons learned atthe Los Angeles zoo. J Cell Biochem 2003;88(2):308–14.
[5] Nemere I, Safford SE, Rohe B, DeSouza MM, Farach-Carson MC.Identification and characterization of 1,25D3-membrane-associatedrapid response, steroid (1,25D3-MARRS)-binding protein. J SteroidBiochem Mol Biol 2004;89–90:281–5.
[6] Armbrecht HJ, Boltz MA, Hodam TL, Kumar VB. Differential re-sponsiveness of intestinal epithelial cells to 1,25-dihydroxyvitaminD3—role of protein kinase C. J Endocrinol 2001;169(1):145–51.
[7] Liu R, Rohe B, Carson DD, Farach-Carson MC. A rapid and sim-ple nonradioactive method for in vitro testing of ribozyme activity.Antisense Nucl Acid Drug Dev 2002;12(4):283–8.
[8] Marcinkowska E. A run for a membrane vitamin D receptor. BiolSignals Recept 2001;10(6):341–9.
[9] Norman AW, Mizwicki MT, Norman DP. Steroid-hormone rapid ac-tions, membrane receptors and a conformational ensemble model.Nat Rev Drug Discov 2004;3(1):27–41.
[10] Frickel EM, Frei P, Bouvier M, Stafford WF, Helenius A, Glock-shuber R, et al. ERp57 is a multifunctional thiol-disulfide oxidore-ductase. J Biol Chem 2004;279(18):18277–87.
[11] Andres DA, Rhodes JD, Meisel RL, Dixon JE. Characterizationof the carboxyl-terminal sequences responsible for protein reten-tion in the endoplasmic reticulum. J Biol Chem 1991;266(22):14277–82.
[12] Nemere I, Ray R, McManus W. Immunochemical studies on theputative plasmalemmal receptor for 1,25(OH)2D3. I. Chick intestine.Am J Physiol Endocrinol Metab 2000;278(6):1104–14.
[ ki Y.urine(2):
of 1,25-dihydroxyvitamin D3-mediated intestinal calcium absorptiMiner Electrolyte Metab 1990;16(2–3):109–14.
[3] Nemere I, Farach-Carson MC, Rohe B, Sterling TM, NormAW, Boyan BD, et al. Ribozyme knockdown functionally lin
13] Tokumasa A, Katsuno T, Tanaga TS, Yokote K, Saito Y, SuzuReduction of Smad3 accelerates re-epithelialization in a mmodel of colitis. Biochem Biophys Res Commun 2004;317377–83.





























