Embed Size (px)
Citation preview

ARTICLE
Identification and Regulation of Tissue-Specificcis-Acting Elements Associated With theHuman AP-2� GeneJ. Zhang1,2 and T. Williams1,3*
Mice lacking transcription factor AP-2� exhibit defects in the formation of the head, body wall, heart, neural tube, eye,and limbs, reflecting important sites of AP-2� expression in the developing embryo. AP-2� is also expressed in thepostnatal mammary gland and has been linked to tumor progression and defects in growth regulation in the breast.We have used a transgenic mouse approach to identify tissue-specific cis-acting sequences associated withexpression of the human AP-2� gene. Our analysis indicates that multiple elements located throughout the genecontribute to expression in the trigeminal ganglia, spinal cord, mammary gland, and epidermis. A discrete cis-elementlocated within the fifth intron is required for expression in the face and limbs, and we have derived a permanent lineof AP-2�::lacZ transgenic mice to assess expression of this latter enhancer throughout morphogenesis. We alsointroduced this transgene into an AP-2�–null mouse background and detected subtle alterations of its expressionwithin the progress zone and apical ectodermal ridge of the forelimbs. Similar changes in lacZ expression were observedwithin the zeugopod, and these correlated with defects in radius condensation in AP-2�–knockout mice. Taken together,these findings indicate that cell:cell communication within the forelimb is altered in the absence of AP-2� and reveal novelregulatory potential for AP-2� in limb development. Developmental Dynamics 228:194–207, 2003.© 2003 Wiley-Liss, Inc.
Key words: lacZ reporter; transgenic mouse; AP-2�; transcription factor; limb; face; mammary gland; spinal cord; geneexpression; regulatory elements
Received 12 May 2003; Accepted 13 June 2003
INTRODUCTION
The human AP-2� gene is the arche-typal member of the AP-2 family of“basic-helix-span-helix” transcriptionfactors (Williams et al., 1988; Moseret al., 1995; Bosher et al., 1996; Ou-lad-Abdelghani et al., 1996; Hilger-Eversheim et al., 2000; Zhao et al.,2001; Cheng et al., 2002; Feng andWilliams, 2003). This gene family reg-ulates important aspects of verte-
brate development and is linkedwith tumor progression in both breastand skin cancer (Schorle et al., 1996;Zhang et al., 1996; Moser et al.,1997a; Nottoli et al., 1998; Turner etal., 1998; Gee et al., 1999; Bar-Eli,2001; Auman et al., 2002; Brewer etal., 2002; Werling and Schorle, 2002;Zhang et al., 2003). During mouseand chick embryogenesis, AP-2� ex-pression has first been reported in
premigratory neural crest cells, andis maintained in these cells as theymigrate throughout the trunk andcranial regions of the embryo(Mitchell et al., 1991; Shen et al.,1997). Several tissues derived fromthe neural crest continue to expressAP-2� during their growth and mor-phogenesis, including the frontona-sal process, branchial arches, andperipheral nervous system ganglia
1Department of Molecular, Cellular and Developmental Biology, Yale University, New Haven, Connecticut2College of Life Science, Hunan Normal University, Changsha, Hunan, People’s Republic of China3Departments of Craniofacial Biology and Cellular and Structural Biology, University of Colorado Health Sciences Center, Denver, ColoradoGrant sponsor: NIH; Grant numbers: DE12728; CA77833.*Correspondence to: T. Williams, Departments of Craniofacial Biology and Cellular and Structural Biology, BRB151, Campus Box C286,University of Colorado Health Sciences Center, 4200 East Ninth Avenue, Denver, CO 80262. E-mail: [email protected]
DOI 10.1002/dvdy.10365
DEVELOPMENTAL DYNAMICS 228:194–207, 2003
© 2003 Wiley-Liss, Inc.

(Mitchell et al., 1991; Shen et al.,1997). An additional prominent siteof AP-2� expression is in the develop-ing limb bud. AP-2� transcripts canbe detected soon after the limb budappears, and expression persists inthe mesenchyme of the progresszone during subsequent outgrowth.Significant AP-2� expression is alsoobserved in the developing epider-mis, kidney, cerebellum, spinal cord,and eye. AP-2� levels decline later inembryogenesis, although expressionin some tissues, such as the ductalepithelium of the mammary gland,persists into adulthood (Zhang et al.,2003). Indeed, altered regulation ofAP-2� expression within the maturemammary gland has been linked tothe progression of human breastcancer (Turner et al., 1998; Gee etal., 1999).
Gene targeting experiments haveshown that the AP-2� gene is re-quired for multiple developmentalprocesses in vertebrates. Mice lack-ing both copies of the AP-2� genedie perinatally and exhibit severemalformation of the head and trunk,including exencephaly and thora-coabdominoschisis (Schorle et al.,1996; Zhang et al., 1996). Furtheranalysis has shown that AP-2� is in-dependently required for at least sixmajor morphogenetic events duringembryogenesis: formation of theneural tube, face, eye, body-wall,cardiovascular system, and fore-limbs (Nottoli et al., 1998; West-Mayset al., 1999; Brewer et al., 2002). Theabsence of AP-2� can also affectcell fate determination and geneexpression in the skin and nervoussystem (Maytin et al., 1999; Krameret al., 2000).
The range of phenotypes ob-served in the absence of AP-2� dis-plays a significant overlap with thosecaused by teratogenic levels of reti-noic acid (Ross et al., 2000). More-over, the expression of AP-2� can bealtered by the application of reti-noic acid both in vivo and in vitro(Williams et al., 1988; Luscher et al.,1989; Shen et al., 1997), strongly sug-gesting that AP-2� is an importantcomponent of this morphogen’smechanism of action. Indeed, withrespect to craniofacial develop-ment, the AP-2� gene may providean important link between the envi-
ronmental and genetic causes ofhuman birth defects. Experiments inthe chick indicate that alterations inAP-2� expression in the developingface correlate with the teratogeniceffects of retinoic acid in this tissue(Shen et al., 1997). In addition, ge-netic mapping studies indicate thatthe human AP-2� gene (TFAP2A) islocated at chromosome 6p24, a lo-cus that is associated with craniofa-cial abnormalities, including orofa-cial clefting (Davies et al., 1995,1999; Topping et al., 2002). This latterfinding is significant, given that micewith a single functional copy of theAP-2� gene frequently display facialdysplasia characterized by a twistedsnout and dental malocclusion (Not-toli et al., 1998). Moreover, micelacking both copies of AP-2�, andchimeric mice containing a mixtureof wild-type and AP-2�–null cells,have severe facial clefts (Schorle etal., 1996; Zhang et al., 1996; Nottoli etal., 1998). The orofacial defectscommonly associated with theAP-2� gene are classified as cleft lipwith or without clefting of the pri-mary palate (CL/P) and are distinctfrom clefts of the secondary palate(Wilkie and Morriss-Kay, 2001). CL/Ppathologies result from defective fu-sion of the facial prominences dur-ing a critical period of embryogene-sis, i.e., �embryonic day (E)9.5–E12.5 in the mouse. The observa-tion that the AP-2� gene is highlyexpressed in the facial prominencesduring this time period suggests thatthis transcription factor directly reg-ulates growth and fusion of the facefrom within these tissues. Therefore, itis likely that identification and char-acterization of the cis-acting se-quences responsible for AP-2� ex-pression in the facial prominenceswould advance our understandingof the regulation of craniofacial de-velopment.
Data obtained in chick andmouse experimental systems alsodemonstrate that AP-2� may havean important role in patterning theforelimb. In the chick, removal of theapical ectodermal ridge (AER)causes a rapid loss of AP-2� expres-sion in the mesenchyme, and thisloss precedes the severe reductionin the size of the limb bud (Shen etal., 1997). Treatment with fibroblast
growth factor (FGF) 4 rescues limbbud outgrowth and leads to a re-covery of AP-2� expression in thelimb mesenchyme. Taken together,these data indicate that AP-2� inthe mesenchyme may act down-stream of regulatory signals emanat-ing from the AER in regulating out-growth and patterning of the limbbud. A patterning function for AP-2�in the limb is supported by the find-ing that mice in which there is a lossor alteration of AP-2� expression fre-quently exhibit forelimb defects. Themajority of AP-2�-null mice exhibitforelimb phocomelia, characterizedby loss of the radius (Schorle et al.,1996; Zhang et al., 1996). More strik-ing phenotypes are seen in chimericmice in which the juxtaposition ofwild-type and AP-2�–null cells canlead to forelimb duplications (Nottoliet al., 1998). These data indicatethat cross-talk between populationsof cells may be critical for the ex-pression and function of AP-2�. Re-cently, studies on the single Dro-sophila AP-2 gene (dAP-2) haveshown that it may also regulate legmorphology through both cell-au-tonomous and nonautonomousmechanisms (Kerber et al., 2001;Monge et al., 2001).
Given the importance of AP-2�during mammalian development,and its possible role in breast can-cer, we wish to understand the reg-ulatory mechanisms responsible forits specific expression pattern. Wepreviously have identified theAP-2� proximal promoter andmapped conserved initiator andoctamer sequences within this re-gion that are responsible for thebasal expression of this gene(Creaser et al., 1996). Here, by us-ing reporter gene analysis in trans-genic mice, we have extendedthese studies by identifying cis-reg-ulatory elements associated withthe human AP-2� gene that directexpression to tissues including thelimb, face, and mammary gland.Furthermore, we have used theseAP-2�::lacZ transgenes to analyzechanges in the distribution of cellsthat are capable of using theseAP-2� regulatory sequences in theAP-2�-null background. These stud-ies indicate that communicationbetween the progress zone and
AP-2� EXPRESSION DURING MORPHOGENESIS 195
apical ectodermal ridge is alteredin the absence of AP-2�, and re-veal that the AP-2� gene may besubject to autoregulation.
RESULTSMultiple AP-2� cis-RegulatorySequences Are LocatedDownstream of theTranscriptional Start Sites
We conducted a transient trans-genic analysis in the mouse to iden-
tify the sequence elements responsi-ble for the expression of AP-2� in thedeveloping mammalian embryo.Previously, in vitro analysis of theAP-2� basal promoter indicated thatit does not contain a classic TATAbox but instead relies on an initiatorelement that works in associationwith a conserved octamer motif justupstream (Creaser et al., 1996).Given the atypical nature of theAP-2� initiation sites, we decided toidentify tissue-specific cis-regulatory
elements required for the in vivo ex-pression of AP-2� in the context of itsown promoter. For these purposes, abasic construct was made, C1 Xho �
gal, which contained approximately200 nucleotides of the human AP-2�
promoter upstream of the start sites.This construct possesses all the afore-mentioned sequence elements, thetranslational start site, and the first 13codons of AP-2� fused in frame tothe lacZ coding region (Fig. 1A). Byusing this basic plasmid, we then de-
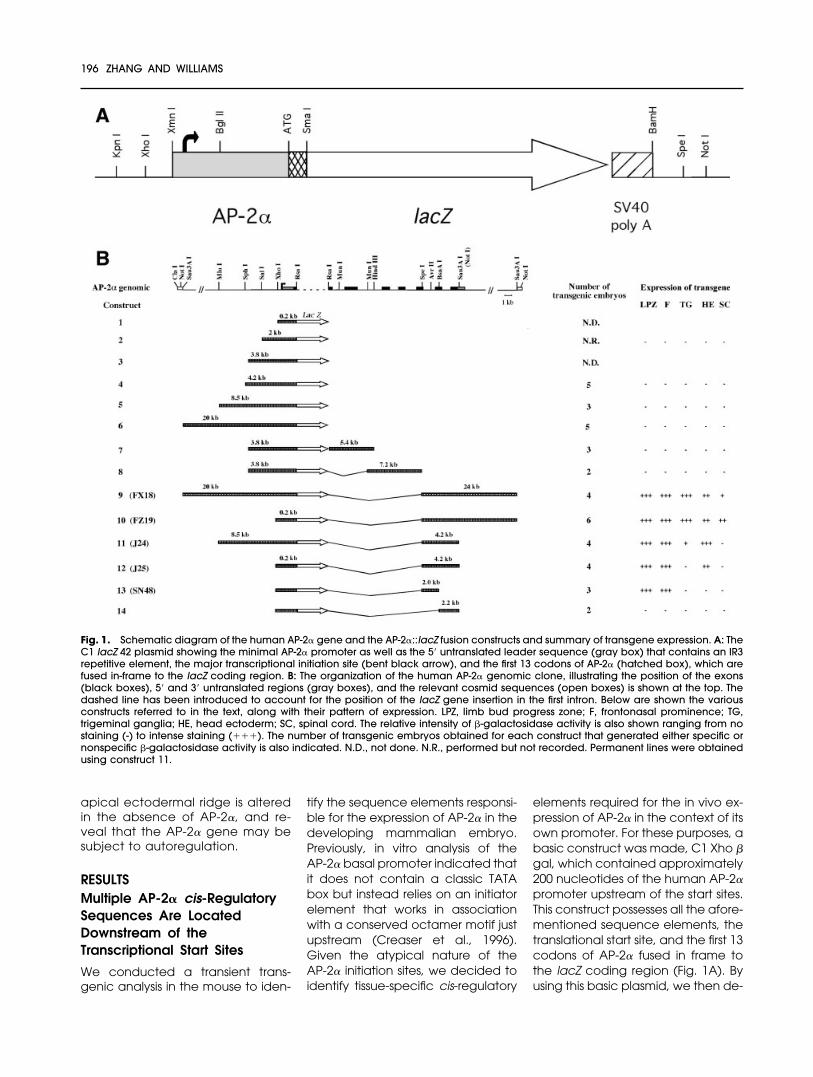
Fig. 1. Schematic diagram of the human AP-2� gene and the AP-2�::lacZ fusion constructs and summary of transgene expression. A: TheC1 lacZ 42 plasmid showing the minimal AP-2� promoter as well as the 5� untranslated leader sequence (gray box) that contains an IR3repetitive element, the major transcriptional initiation site (bent black arrow), and the first 13 codons of AP-2� (hatched box), which arefused in-frame to the lacZ coding region. B: The organization of the human AP-2� genomic clone, illustrating the position of the exons(black boxes), 5� and 3� untranslated regions (gray boxes), and the relevant cosmid sequences (open boxes) is shown at the top. Thedashed line has been introduced to account for the position of the lacZ gene insertion in the first intron. Below are shown the variousconstructs referred to in the text, along with their pattern of expression. LPZ, limb bud progress zone; F, frontonasal prominence; TG,trigeminal ganglia; HE, head ectoderm; SC, spinal cord. The relative intensity of �-galactosidase activity is also shown ranging from nostaining (-) to intense staining (���). The number of transgenic embryos obtained for each construct that generated either specific ornonspecific �-galactosidase activity is also indicated. N.D., not done. N.R., performed but not recorded. Permanent lines were obtainedusing construct 11.
196 ZHANG AND WILLIAMS
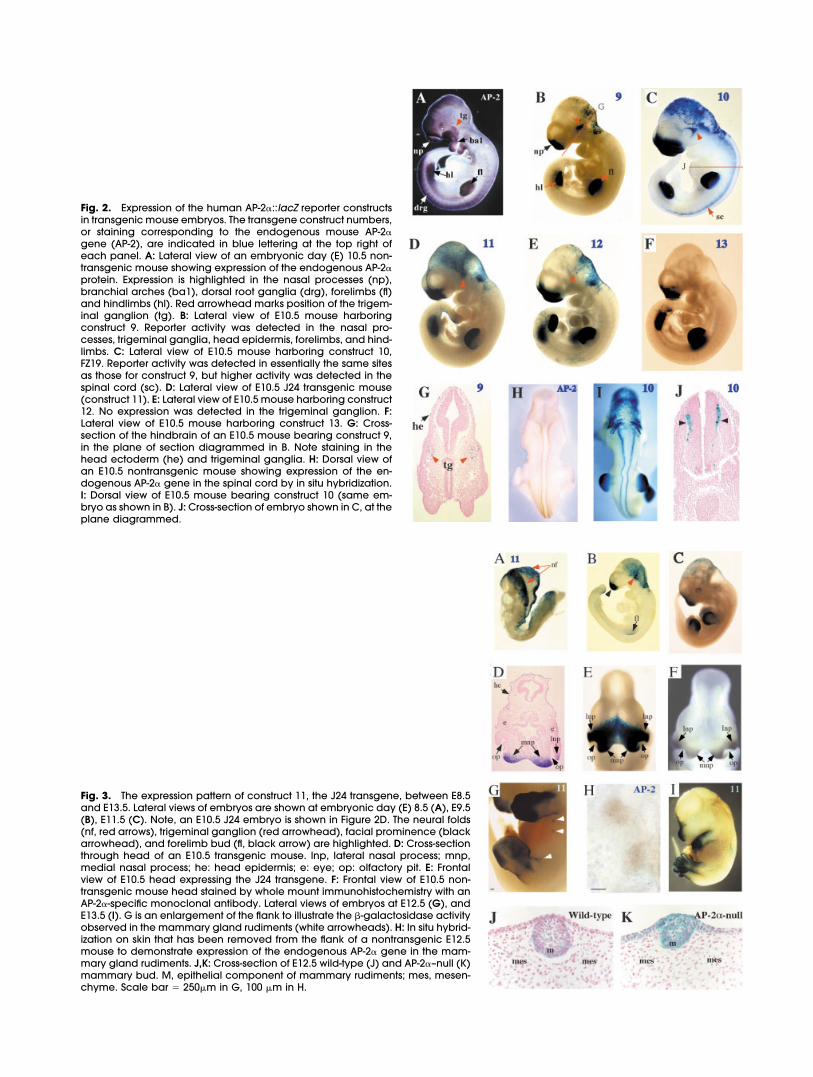
Fig. 2. Expression of the human AP-2�::lacZ reporter constructsin transgenic mouse embryos. The transgene construct numbers,or staining corresponding to the endogenous mouse AP-2�gene (AP-2), are indicated in blue lettering at the top right ofeach panel. A: Lateral view of an embryonic day (E) 10.5 non-transgenic mouse showing expression of the endogenous AP-2�protein. Expression is highlighted in the nasal processes (np),branchial arches (ba1), dorsal root ganglia (drg), forelimbs (fl)and hindlimbs (hl). Red arrowhead marks position of the trigem-inal ganglion (tg). B: Lateral view of E10.5 mouse harboringconstruct 9. Reporter activity was detected in the nasal pro-cesses, trigeminal ganglia, head epidermis, forelimbs, and hind-limbs. C: Lateral view of E10.5 mouse harboring construct 10,FZ19. Reporter activity was detected in essentially the same sitesas those for construct 9, but higher activity was detected in thespinal cord (sc). D: Lateral view of E10.5 J24 transgenic mouse(construct 11). E: Lateral view of E10.5 mouse harboring construct12. No expression was detected in the trigeminal ganglion. F:Lateral view of E10.5 mouse harboring construct 13. G: Cross-section of the hindbrain of an E10.5 mouse bearing construct 9,in the plane of section diagrammed in B. Note staining in thehead ectoderm (he) and trigeminal ganglia. H: Dorsal view ofan E10.5 nontransgenic mouse showing expression of the en-dogenous AP-2� gene in the spinal cord by in situ hybridization.I: Dorsal view of E10.5 mouse bearing construct 10 (same em-bryo as shown in B). J: Cross-section of embryo shown in C, at theplane diagrammed.
Fig. 3. The expression pattern of construct 11, the J24 transgene, between E8.5and E13.5. Lateral views of embryos are shown at embryonic day (E) 8.5 (A), E9.5(B), E11.5 (C). Note, an E10.5 J24 embryo is shown in Figure 2D. The neural folds(nf, red arrows), trigeminal ganglion (red arrowhead), facial prominence (blackarrowhead), and forelimb bud (fl, black arrow) are highlighted. D: Cross-sectionthrough head of an E10.5 transgenic mouse. lnp, lateral nasal process; mnp,medial nasal process; he: head epidermis; e: eye; op: olfactory pit. E: Frontalview of E10.5 head expressing the J24 transgene. F: Frontal view of E10.5 non-transgenic mouse head stained by whole mount immunohistochemistry with anAP-2�-specific monoclonal antibody. Lateral views of embryos at E12.5 (G), andE13.5 (I). G is an enlargement of the flank to illustrate the �-galactosidase activityobserved in the mammary gland rudiments (white arrowheads). H: In situ hybrid-ization on skin that has been removed from the flank of a nontransgenic E12.5mouse to demonstrate expression of the endogenous AP-2� gene in the mam-mary gland rudiments. J,K: Cross-section of E12.5 wild-type (J) and AP-2�–null (K)mammary bud. M, epithelial component of mammary rudiments; mes, mesen-chyme. Scale bar � 250�m in G, 100 �m in H.

veloped a series of constructs inwhich up to 20 kb of upstream AP-2�sequences were attached to thelacZ gene (constructs 1–6, Fig. 1B).Members of this series were next in-jected into one-cell mouse embryos,and the transgenic founder micewere analyzed subsequently bywhole-mount �-galactosidase stain-ing at E10.5. This developmentaltime point was chosen for the initialanalysis because it enables AP-2�expression to be assayed in both thelimb bud and in neural crest deriva-tives, such as the face and cranialganglia. Despite the importance ofthe promoter proximal region forbasal level expression in vitro, we didnot detect any strong lacZ expres-sion in vivo from constructs contain-ing this element when it was placedsolely in association with upstreamsequences. Even the presence of 20kb of upstream AP-2� genomic se-quences (construct 6) failed to pro-duce any strong and consistent pat-tern of lacZ expression in transgenicmice at E10.5.
We next examined if cis-regulatorysequences might reside in the re-gions downstream of the AP-2� startsites. The human AP-2� gene consistsof seven exons and six introns thatspan 18 kb (Bauer et al., 1994). Alarge transgene was made contain-ing 20 kb of upstream sequences inassociation with a 24 kb fragment ofdownstream sequences. These lattersequences encompassed the fifthand sixth introns and the 20-kb re-gion immediately downstream ofthe stop codon (construct 9, Fig. 1B).At E10.5, this construct was able todirect strong expression of the trans-gene to the mesenchyme of thefrontonasal process and limb buds(Fig. 2B), which are major sites of en-dogenous AP-2� expression at thisstage of development (Fig. 2A andMitchell et al., 1991). Additionalstaining was observed in the trigem-inal ganglia, and in dorsolateral cellswithin the brain and spinal cord (Fig.2B,G, and data not shown), whichagain correspond to sites of endog-enous AP-2� expression. Transgeneexpression was also observed in filesof cells that emanated from the dor-sal surface of the head. In cross-sec-tion, these cells occurred within the
head ectoderm and were not com-ponents of the neural crest (Fig. 2G).
The 24-kb downstream fragmentwas next tested in the context of theminimal 0.2-kb human AP-2� pro-moter. The pattern of lacZ activitydirected by this shorter transgene,(Fig. 1B, construct 10; Fig. 2C,I) wassimilar to that seen with the afore-mentioned transgene that con-tained 20 kb of upstream sequence(Fig. 1B, construct 9; Fig. 2B). Thesefindings indicate that major cis-reg-ulatory elements responsible forAP-2� expression occur downstreamof the start site. However, one no-ticeable difference in the activity ofthe two transgenes was that con-struct 10 was more strongly ex-pressed within the spinal cord (Fig.2C,I,J) than construct 9 (Fig. 2B). Thislatter observation may indicate thatnegative regulatory sequences re-side upstream of the basal pro-moter, and their removal allows forhigher levels of spinal cord expres-sion from the shorter transgene.
Because regulatory sequencesappeared to be concentrateddownstream of the start site, wegenerated and tested additionaltransgenes containing sequencesfrom the more promoter proximal in-tron sequences within introns 1–4.However, a construct composed ofthe first intron and part of the secondintron attached to 3.8 kb of AP-2�upstream regulatory sequences (Fig.1B, construct 7), failed to produceany specific staining. Similarly, afragment containing the majority ofthe second intron and all of the thirdand fourth introns (Fig. 1B, construct8) did not produce any consistent�-galactosidase staining pattern.Therefore, we concentrated our fur-ther transgenic analysis on the moredownstream sequence elements,from intron 5 onward, that recapitu-lated multiple aspects of the endog-enous AP-2� expression pattern.
Regulatory ElementsResponsible for FacialMesenchyme and Limb BudExpression Reside Within theFifth Intron
Two constructs were made in whichthe 4.2-kb genomic region that
spans exons 5 through 7 was at-tached to either 8.5 kb or 0.2 kb ofupstream sequence (Fig. 1B, con-structs 11 and 12, respectively). Theshorter transgene, construct 12 (J25),directed �-galactosidase expressionto the limb, face, and head ecto-derm (Fig. 2E), indicating that majorregulatory sequences lie within thefifth and sixth introns. The addition of8.3 kb of further upstream se-quences in the plasmid J24 (con-struct 11) gave the same pattern ofexpression as construct 12 but alsorestored expression at a low level tothe trigeminal ganglia (Fig. 2D, ar-rowhead). This latter finding indi-cates that there are regulatory se-quences responsible for AP-2�expression upstream of the start site,although they may need to cooper-ate with downstream sequences toyield a high level of expression. Un-like construct 10, neither construct11 nor 12 produced expression in thespinal cord, suggesting that the ele-ments responsible for the expressionin spinal cord were located down-stream of the coding region. The4.2-kb fragment that directed ex-
Fig. 4. Analysis of embryonic day (E) 12.5spinal cord expression in transgenic micebearing construct 10 (FZ19). A,B: Dorsal andlateral views of an E12.5 transgenic mouse,respectively. C,D: Cross-sections of the E12.5transgenic mouse shown in B, in the planesof section diagrammed.Fig. 5. Comparison of J24 lacZ transgeneexpression between wild-type (WT) and AP-2�-null (Null) embryos. A,B: Lateral view ofwild-type (A) and AP-2�–null (B) J24 em-bryos at embryonic day (E) 10.5. C,D: Fron-tal view of a wild-type (C) and an AP-2�–null(D) embryonic head at E10.5. E,F: Lateralview of wild-type (E) and AP-2�–null (F) em-bryo at E12.5. G,H: Frontal view of a wild-type (G) and an AP-2�–null (H) embryonichead at E12.5.Fig. 7. Alterations in J24 lacZ transgeneexpression in the forelimb buds of AP-2�–null embryos correlate with agenesis of theradius. Forelimbs from wild-type (A–C) andAP-2�–null (D,E) embryos were analyzed forthe expression of the J24 lacZ transgene.A,D: Dorsal view of embryonic day (E) 12.5wild-type (A) and AP-2�-null (D) forelimbs.B,E: Cross-section of the forelimbs shown inA and D at the planes diagrammed. Thepositions of the condensing radius (r) andulna (u) are shown. The mutant forelimb in Elacked a radius (arrow). C: Cross-section ofE13.5 wild-type forelimb in the region of thedigits.
198 ZHANG AND WILLIAMS
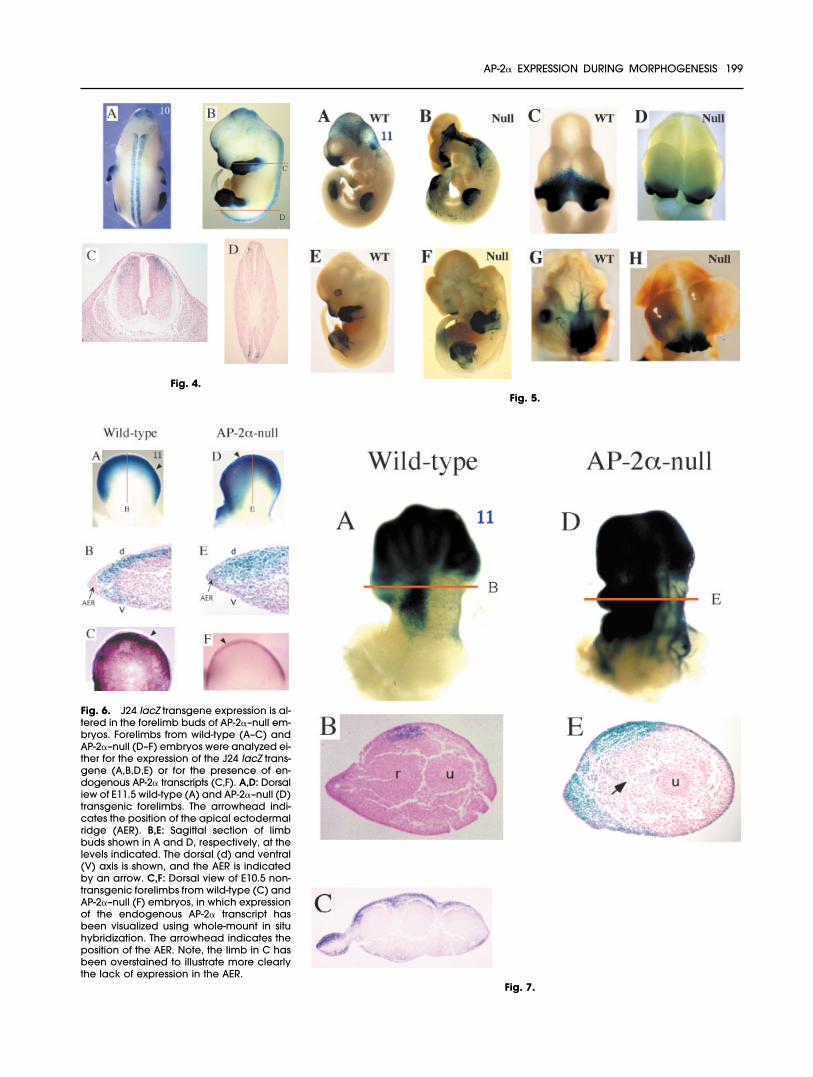
Fig. 4.Fig. 5.
Fig. 6. J24 lacZ transgene expression is al-tered in the forelimb buds of AP-2�–null em-bryos. Forelimbs from wild-type (A–C) andAP-2�–null (D–F) embryos were analyzed ei-ther for the expression of the J24 lacZ trans-gene (A,B,D,E) or for the presence of en-dogenous AP-2� transcripts (C,F). A,D: Dorsaliew of E11.5 wild-type (A) and AP-2�–null (D)transgenic forelimbs. The arrowhead indi-cates the position of the apical ectodermalridge (AER). B,E: Sagittal section of limbbuds shown in A and D, respectively, at thelevels indicated. The dorsal (d) and ventral(V) axis is shown, and the AER is indicatedby an arrow. C,F: Dorsal view of E10.5 non-transgenic forelimbs from wild-type (C) andAP-2�–null (F) embryos, in which expressionof the endogenous AP-2� transcript hasbeen visualized using whole-mount in situhybridization. The arrowhead indicates theposition of the AER. Note, the limb in C hasbeen overstained to illustrate more clearlythe lack of expression in the AER.
Fig. 7.
AP-2� EXPRESSION DURING MORPHOGENESIS 199

pression to the face, limbs, and ec-toderm was further divided into twopieces, corresponding to introns fiveor six, and tested in association withthe minimal promoter (Fig. 1B, con-structs 13 and 14, respectively). The1.95-kb fifth intron was able to directexpression to the face and limbs(construct 13; Fig. 2F), whereas thesixth intron did not possess any activ-ity in this context (construct 14). Thus,an enhancer element specific forthe mesenchyme of the limb andfrontonasal prominence resideswithin the fifth intron of the humanAP-2� gene.
Spatiotemporal Analysis of J24Transgene Expression Revealscis-Regulatory SequencesDirecting Mammary Gland andEpithelial-Specific GeneExpression
To perform a more comprehensiveanalysis of the spatiotemporal regu-lation of AP-2� transgene expressionin the limb, face, and trigeminalganglia, we generated stable trans-genic mouse lines with construct 11,J24 (Fig. 3). Two lines were obtained,which both gave identical patternsof lacZ expression, and one of theselines was analyzed in more detail.During embryogenesis, expression ofthis transgene was first detected be-tween E8.0 and E8.5, in the cranialneural folds (Fig. 3A). The onset oftransgene expression in the embryoand its location surrounding the neu-ral plate approximated to the ex-pression pattern of the endogenousAP-2� gene. Between E9.5 andE11.5, expression associated with theneural folds became concentratedin files of ectodermal cells that em-anated from the dorsal surface ofthe head (Fig. 3B–D).
By E9.5, �-galactosidase activitybecame apparent in the frontona-sal prominence (Fig. 3B) and per-sisted through E13.5, becomingmore restricted to the midline of theface over this time period. Histologicexamination at E10.5 indicated thattransgene expression was confinedto the mesenchyme of the medialand lateral nasal processes and wasnot present in the facial ectodermat this stage (Fig. 3D). The timing and
spatial distribution of �-galactosi-dase activity within the mesen-chyme of the medial and lateral na-sal processes during this periodrecapitulates that of the endoge-nous AP-2� protein (Fig. 3E,F). Thesefindings suggest we have identifiedan enhancer element in the humanAP-2� gene that can direct a verysimilar spatiotemporal pattern ofcraniofacial expression as the en-dogenous mouse gene. The J24transgene also contained se-quences that were capable of di-recting �-galactosidase activity tothe trigeminal ganglia, another siteof endogenous AP-2� expression.Transgene expression in these gan-glia was apparent from E9.5 to E10.5but was not observed at subsequentstages (Figs. 2D, 3B, and data notshown).
The J24 lacZ transgene also reca-pitulated the expression pattern ofthe endogenous AP-2� gene in thelimb bud progress zone. Expressioncould first be detected in a patch ofmesenchyme in the nascent fore-limb bud at E9.5 (Fig. 3B). By E10.5,expression was readily apparent in aregion corresponding to theprogress zone in both fore- and hind-limb buds (Fig. 2D). Expression wasthen maintained in the distal limbmesenchyme from E11.5–13.5, be-coming progressively confined tothe autopod during these stages(Fig. 3C,G,I). A second, more limited,expression domain could be ob-served at E12.5 and E13.5 as a dorsalstripe in the mesenchyme of the ze-ugopod (Fig. 3G,I). We will discussthe histologic analysis of transgeneexpression in wild-type limbs in thefinal part of the Results section, in acomparison to the data obtained inAP-2�–null limbs.
J24 transgene expression also be-came evident in two further tissuesby E12.5: the developing genital tu-bercle (data not shown) and the ep-ithelial compartment of the mam-mary gland rudiments (Fig. 3G,J).Mammary-specific transgene ex-pression was observed in all five pairsof mammary buds at this time point.In Figure 3G, arrowheads indicatethree visible buds that are discern-ible as blue dots on the flank,whereas the other two buds on thisside are obscured by the limbs. Be-
cause AP-2� expression was not pre-viously noted in the embryonicmammary glands, in situ hybridiza-tion was used to extend this analysis.In agreement with the transgenicdata, we determined that endoge-nous AP-2� transcripts were also ex-pressed in the mammary gland rudi-ments at E12.5 (Fig. 3H). Transgeneexpression was confined to the epi-thelial component of the mammarygland bud, and no reporter activitywas detected in the surroundingmesenchyme (Fig. 3J). Extensive ec-todermal expression of the endoge-nous AP-2� gene has also been re-ported in the developing hairfollicles of the epidermis (Byrne et al.,1994). The J24 transgene also di-rected widespread punctate �-ga-lactosidase staining correspondingto this aspect of AP-2� expression byE13.5 (Fig. 3I). This age was the lasttime point analyzed by whole-mount analysis since we and othershave noted that stain penetrationproblems occur in mice from E13 on-ward (Hogan et al., 1994). The AP-2�gene is not extensively expressedduring the postnatal period; there-fore, we have not performed a de-tailed analysis of J24 expression inthe adult at this juncture. Neverthe-less, we have noted that J24 trans-gene activity occurred within theductal epithelia of the mammarygland at postnatal time points (datanot shown) in common with the ex-pression of the endogenous AP-2�gene (Zhang et al., 2003). This latterfinding suggests that the enhancerwe have identified may be criticalfor regulating AP-2� function in nor-mal breast physiology and duringthe progression of breast cancer(Bosher et al., 1995; Turner et al.,1998; Gee et al., 1999; Zhang et al.,2003).
Analysis of SequencesRequired for Expression in theMammary Gland, GenitalTubercle, and Spinal Cord
Given the potential importance of thesequences required for mammary-specific AP-2� expression, further stud-ies were undertaken to determinetheir location. The embryonic mam-mary buds are not apparent at theE10.5 time point we had used previ-
200 ZHANG AND WILLIAMS

ously for transient analysis. Therefore,a new set of E12.5 transient transgenicembryos were generated and ana-lyzed by using either construct 5,which contains only the upstream se-quences present in J24, or construct12, which has the minimal promoterand all of the J24 downstream ele-ments. Unlike J24, neither of theseconstructs were capable of generat-ing expression in the E12.5 mammarygland rudiments (data not shown).Thus, expression in the embryonicmammary gland required coopera-tion between cis-regulatory elementslocated both upstream and down-stream of the transcriptional initiationsites, a situation analogous to that ob-served for expression within the tri-geminal ganglia. In contrast, the se-quences required for expression withinthe genital tubercle at E12.5 weremore localized. LacZ expression wasobtained in transgenic mice harbor-ing construct 12 but was not seen withconstruct 5 (data not shown). Thesefindings position the regulatory se-quences required for genital tubercleexpression downstream of the startsite within introns five and six.
Construct 10, which containedmore extensive downstream se-quence than J24, directed expres-sion to the face, limbs, trigeminalganglia, head ectoderm, and spinalcord at E10.5 (Fig. 2C). We also an-alyzed this transgene in a transientanalysis at E12.5 to determinewhether there were any changes inthe expression pattern (Fig. 4). �-ga-lactosidase activity remained robustin the limbs, face, and spinal cord(Fig. 4A,B). However, expression inthe trigeminal ganglia and head ec-toderm were no longer apparent atE12.5, and no new expression do-mains were observed. The contin-ued expression in the limb and faceand loss of expression in the headectoderm and trigeminal gangliawere consistent with the data ob-tained with the J24 transgene atE12.5. With respect to the spinalcord, construct 10 expression wasmore dorsal in aspect at E12.5 whencompared with E10.5 (Fig. 4C, com-pare with 2J). Moreover, there was achange in the dorsoventral positionof the lacZ-positive cells along therostral caudal axis of the spinal cord.Cells expressing the transgene were
restricted to more dorsolateral posi-tions toward the head (Fig. 4C), incontrast to their more mediolateralpositions toward the tail (Fig. 4D). To-gether, these findings indicate thatwe have identified the appropriateregulatory sequences necessary forspinal cord expression, because thespatiotemporal expression patternof this human lacZ transgene corre-sponds to that of the endogenousmouse AP-2� gene in presumptivespinal sensory interneurons (com-pare Fig. 2H and 2I; Mitchell et al.,1991). Further mapping studies sug-gest that the spinal cord regulatorysequences occur within the most 3�18 kb of the AP-2� sequences thatwe have tested (data not shown).
Expression of the Transgene inthe Forelimb Buds Is Expandedin AP-2�–Null Mice
We wished to determine whethercell populations that would normallyexpress AP-2� were specifically al-tered in the AP-2�–knockout mousebackground. Therefore, we crossedthe J24 transgene into the AP-2�–nullbackground so that the lacZ markercould be used to follow the distribu-tion of such cell populations. In gen-eral, we found that significant ex-pression of the lacZ transgene stilloccurred in the appropriate tissuesin the absence of AP-2�, althoughsuch tissues—including the face andmammary buds—were frequentlydisplaced due to the dysmorphol-ogy (Fig. 5). These results indicatedthat the loss of AP-2� gene expres-sion did not necessarily lead to areduced population size for the cellsin which it was usually expressed, asituation we have also noted with alacZ knock-in allele of AP-2� (Breweret al., 2002). However, there weretwo exceptions to this general trend.The first change was a decrease intransgene expression in the trigemi-nal ganglia, a result that was ex-pected because these structuresare greatly reduced in size in AP-2�–null mice (data not shown; Schorleet al., 1996; Zhang et al., 1996). Morestriking and unexpected were the al-terations of J24 transgene expressionin the forelimbs of AP-2�–null micewhen compared with their wild-typelittermates.
Whole-mount preparations ofwild-type limbs at E11.5 showed thatJ24 was expressed in distal limb tis-sue (Fig. 6A). Sagittal sectionsthrough the forelimb at this stageshowed that the �-galactosidaseactivity was present in the mesen-chyme and not in the overlying epi-dermis or in the apical ectodermalridge (Fig. 6B). Within the mesen-chyme of the progress zone, trans-gene activity was more pronouncedtoward the dorsum (Fig. 6B). Inmarked contrast, for the E11.5 AP-2�–null forelimb, additional lacZ re-porter activity was detected in theAER and the epidermis, as well as inits normal mesenchymal location(Fig. 6D,E). Furthermore, with respectto the progress zone, expression inthe AP-2�–null background now en-compassed the entire dorsoventralaxis of the limb bud, although as inthe wild-type it was still more con-centrated on the dorsal aspect (Fig.6E). To ascertain if these alterationswere a peculiarity of the transgeneor represented bona fide changes inAP-2� gene regulation, we exam-ined the expression of the nonfunc-tional targeted AP-2�–knockout al-lele. Although this allele is notcapable of producing a functionalprotein, it is still transcribed in the nullbackground. In situ hybridizationstudies confirmed that the endoge-nous AP-2� gene was also activatedin the AER of the mutant limbs (Fig.6F, arrowhead).
Whole-mount and histologic anal-ysis of J24 transgene expression wasalso performed at E12.5 and E13.5for wild-type forelimbs. At thesestages, the transgene was stillheavily expressed in the distal mes-enchyme of limb bud, although itwas less prevalent in the regions cor-responding to digit formation (Fig.7A; also Fig. 3G,I). In histologic sec-tions, expression was again mostpronounced in the dorsal region ofthe mesenchyme, adjacent to theregions of digit condensation (Fig.7C). Patches of expression couldalso be seen in the limb epidermisbeginning at E13.5 (Fig. 7C), in com-mon with the activation of epider-mal expression in other regions of theembryo at this stage (Fig. 3I).
At E12.5 and E13.5 a second do-main of expression occurred in the
AP-2� EXPRESSION DURING MORPHOGENESIS 201

zeugopod, specifically in a stripe ofmesenchyme that resides dorsal tothe position of radius condensation(Fig. 7A,B). When J24 was intro-duced into an AP-2�–null back-ground, alterations in transgene ex-pression were also apparent at E12.5in this region of the mouse forelimbbud. First, expression of the trans-gene was activated in the epidermisof the knockout mice (Fig. 7E) incommon with the E11.5 time point(Fig. 6E). Second, the discrete do-main of expression normally seen inthe mesenchyme dorsal to the ra-dius was often, but not always, re-placed by an expanded region ofreporter activity that encompassedthe lateral and ventral regions onthe posterior side of the limb bud(Fig. 7D,E). The partial penetrance ofthis altered expression pattern was in-triguing given that 65–70% of AP-2�–knockout animals normally lacked aradius (Zhang et al., 1996). Further in-vestigation revealed that there was asignificant correlation between thetwo phenomena: of 18 forelimbs, 12had the expanded domain of re-porter activity, and these also lackedareas of condensation correspondingto the position of the radius (Fig. 7E,and data not shown). The aberrantexpression of AP-2� specifically af-fects the patterning of the forelimband does not produce gross alter-ations of hindlimb morphology. In-triguingly, in contrast to the forelimb,the expression of the J24 lacZ trans-gene in the hindlimb was essentiallyidentical in both the wild-type andAP-2�–knockout backgrounds. In par-ticular, no expression was observed inthe apical ectodermal ridge of thehindlimb in either genetic back-ground, and there were no differ-ences in the mesenchymal domainsof expression (data not shown).
DISCUSSION
The AP-2� gene performs importantfunctions in several morphogeneticprocesses critical for mammaliandevelopment. It will be necessary todetermine how expression of thisgene is controlled before we canfully understand the mechanisms bywhich AP-2� participates in the reg-ulatory network that governs embry-ogenesis. In this report, by using a
series of AP-2�::lacZ transgenes, wehave demonstrated that the tissue-specific expression of AP-2� relies onmultiple regulatory elements. Fur-thermore, we have discovered thatthe pattern of gene expression gen-erated by these sequences can bealtered in the forelimb in the ab-sence of the AP-2� protein, indicatingthat this gene may have a hithertounsuspected negative autoregula-tory function.
We had originally determined thatthe basal promoter of the humanAP-2� gene encompassed the tran-scriptional initiation sites and the ad-jacent upstream region of �100 bp(Creaser et al., 1996). This sequencewas highly conserved between hu-man, mouse, and chicken and wasable to direct accurate transcrip-tional initiation in tissue culture cells.However, in the current study, wedemonstrate that this sequence byitself does not possess the requisitecis-regulatory elements to direct ap-propriate tissue-specific expressionin vivo. Moreover, the addition of upto 20 kb of genomic sequence 5� tothe start site failed to produce anyconsistent pattern of lacZ gene ex-pression. In contrast, the addition of24 kb of genomic DNA extendingdownstream from exon five pro-duced robust �-galactosidase activ-ity in multiple sites in the developingembryo. Specifically, the down-stream sequences contain cis-regu-latory elements directing expressionto the neural folds, frontonasalprominences, limb bud progresszone, trigeminal ganglia, centralnervous system, skin, mammarygland, and genital tubercle. Thesetissues correspond to important sitesof endogenous AP-2� expression,and the results indicate that wehave identified bona fide regulatorysequences responsible for many as-pects of AP-2� gene transcriptionwithin the developing embryo(Mitchell et al., 1991). Our analysisfurther illustrates that the individualcis-regulatory sequences are dis-persed over a large region of theAP-2� gene. Indeed, although weanalyzed �45 kb of human AP-2�genomic sequence the cis-actingelements responsible for AP-2� ex-pression in the developing eye, mi-grating neural crest and branchial
arches were not identified. This find-ing indicates that the cis-acting se-quences required for AP-2� expres-sion in the mesenchyme of the facialprominences are distinct from theregulatory elements needed in themigratory NCC that give rise to thefacial prominences. Similarly, distinctregulatory networks must be respon-sible for directing AP-2� expression inthe mesenchyme of the facial prom-inences as opposed to the branchialarches. An alternative possibility isthat our findings represent a funda-mental difference in regulation ofthe human and mouse AP-2� genesin their respective species. Thus, thehuman gene may have lost orgained some expression potentialwith respect to the mouse gene thatcould reflect differences in the utili-zation of AP-2� to sculpt body planin these distinct mammalian species.Alternatively, more subtle differ-ences in the cis-regulatory elementsbetween the human and mousegenes could alter the ability of thecognate transcription factors tobind to the human gene in the trans-genic mouse system with conse-quent effects on the expression pat-tern. Future experiments on both themouse and human AP-2� genes willbe necessary to determine whetherthere are species-specific functionaldifferences between the respectivecis-regulatory sequences and/orwhether additional sequence ele-ments reside even more distal to thepromoter and enhancer elementsanalyzed in this study.
Our analysis indicates that the in-teraction of multiple enhancer ele-ments is responsible for the appropri-ate tissue-specific expression of theAP-2� gene. The most defined en-hancer region resides within the fifthintron, and this sequence directsstrong lacZ expression to the fronto-nasal prominence and limb budprogress zone in association with thecore promoter. An examination ofthe sequence of the fifth intron en-hancer element did not reveal anyclearly defined cis-acting motifs. Inparticular, even though AP-2� ex-pression is responsive to retinoic acidapplication in the developing chickface (Shen et al., 1997), we wereunable to identify a conserved reti-noic acid responsive element based
202 ZHANG AND WILLIAMS

solely on the primary sequence.Therefore, a detailed molecular dis-section of this 1.95-kb intronic regionwill be required to pinpoint the cru-cial cis-acting motifs required forlimb and face-specific AP-2� expres-sion. Such an analysis will be espe-cially pertinent with respect tocraniofacial development becausethe AP-2� gene maps to a chromo-somal locus associated with humanorofacial clefting (Davies et al.,1995, 1999; Topping et al., 2002).Thus, it is possible that alterationswithin the AP-2� coding sequences,or within the associated cis-regula-tory elements, will contribute to hu-man craniofacial abnormalities. In-deed, the reason we chose to studythe human rather than mouse AP-2�cis-regulatory sequences was thepotential for understanding the ge-netic causes of human craniofacialdefects.
Sequence elements within the fifthintron can contribute to other as-pects of AP-2� expression in additionto those within the face and limb. Inconjunction with the sixth intron,these sequences produce expres-sion in the ectoderm of the head.Moreover, the region encompassingintrons five and six is also involved indirecting gene expression to the tri-geminal ganglia and mammarybuds. In these two tissues, the down-stream elements need to cooperatewith cis-regulatory sequences lo-cated in an 8.5-kb region upstreamof the basal promoter to producerobust tissue-specific expression. Atleast one additional downstream re-gion of the AP-2� gene is involved indriving expression within the trigemi-nal ganglia. This conclusion is basedon the finding that construct 10(FZ19), which consists mainly of the24-kb downstream sequence, is ex-pressed in the trigeminal gangliaeven though it lacks the 8.5-kb up-stream fragment discussed above.Based on the structure of construct10, the pertinent cis-regulatory ele-ments must be located 3� to theAP-2� coding region, specificallydownstream of exon seven. Thus,the expression of the endogenousAP-2� gene within the trigeminalganglia may result from the con-certed action of multiple, dispersed,cis-acting elements. Also note that
sequences located downstream ofthe AP-2� coding region are not onlyinvolved in producing tissue-specificgene expression within the trigemi-nal ganglia but also within the cen-tral nervous system. Further analyseswill be required to determine the ex-tent and importance of synergisticinteractions for AP-2� expression, es-pecially with respect to any regula-tory elements that exist in the moredistal sequences that were outsidethe scope of the current analysis. Fi-nally, given the preponderance ofenhancer sequences we have lo-cated downstream of the AP-2�transcriptional start site, it will be in-teresting to determine whether asimilar arrangement of regulatory se-quences occurs in the related mam-malian AP-2� and AP-2� genes(Moser et al., 1995; Bosher et al.,1996; Chazaud et al., 1996). Thegenomic organization of these threegenes is very similar, each consistingof seven exons and six introns (Baueret al., 1994), and they are also ex-pressed in overlapping patterns inthe neural crest, central nervous sys-tem, limb, and face (Mitchell et al.,1991; Chazaud et al., 1996; Moser etal., 1997b).
In the course of mapping AP-2�cis-regulatory sequences, we alsogenerated permanent lines of AP-2�–null mice that carry the J24AP-2�::lacZ transgene. This ap-proach enabled us to follow the dis-tribution of cells that express thisAP-2�::lacZ marker in the face andlimbs of AP-2�-knockout mice. Theoutgrowth and patterning of thelimb bud relies on continued com-munication between the mesenchy-mal cells in the progress zone andthe apical ectodermal ridge (Tickle,2003). Signaling molecules of theFGF and hedgehog families andtheir cognate receptors are key par-ticipants in this process. However,the transcriptional mechanisms bywhich these signaling pathways areestablished and interpreted arelargely uncharacterized. Previousstudies in the chick have indicatedthat AP-2� expression in the progresszone responds rapidly to signals em-anating from the AER (Shen et al.,1997). The current data strongly sug-gest that the AP-2� gene can alsorespond to signaling information
passing in the opposite direction,i.e., from the progress zone to theAER. For such a cell nonautonomousmechanism, we would postulatethat when AP-2� protein is removedfrom the progress zone, a cell:cellsignaling cascade is disrupted sothat repression of the AP-2� gene nolonger occurs within the AER. This hy-pothesis is supported by studies inDrosophila, which indicate that thedAP-2 gene can act in a nonauto-nomous manner to regulate leg out-growth (Kerber et al., 2001; Mongeet al., 2001). Alternatively, a cell au-tonomous mechanism may be re-sponsible for the activation of thetransgene in the AER. In this in-stance, a very low level of AP-2� ex-pression would occur normally withinthe AER and this expression wouldrepress AP-2� promoter activity. Inthe absence of functional AP-2�protein, repression of the AP-2�gene would be relieved. Supportingthis latter hypothesis, AP-2 bindingsites have been identified within theAP-2� promoter, and it has beenshown previously that AP-2� canregulate its own expression in vitro(Bauer et al., 1994; Creaser et al.,1996). Furthermore, we did note thatthe level of J24 transgene activitywas consistently higher in the mam-mary glands of AP-2�–null mice, sug-gesting that AP-2� may normally au-toregulate its own expression in thistissue (compare Fig. 3J and K). In thefuture, detailed mapping of the se-quences responsible for AP-2� pro-moter activation within the limb willaid in distinguishing between thesecell-autonomous and non–cell-au-tonomous signaling mechanisms.From our analysis of the expression ofthe endogenous AP-2�–null allele inthe AP-2�–knockout background(Fig. 6F), we can deduce two furthermechanistic insights into this gene’stranscriptional control in the limb. Al-though the endogenous gene is ac-tivated in the AER of the knockoutforelimbs, there is a concurrent lossof expression in the progress zone.The absence of expression in theprogress zone may actually reflectthe nature of the AP-2�–null alleleused in this analysis (Zhang et al.,1996). The knockout allele we haveused lacks a large part of intron five,and this is the intron we have now
AP-2� EXPRESSION DURING MORPHOGENESIS 203

shown drives expression in theprogress zone. The finding that ex-pression of the knockout allele is ab-sent in the progress zone suggeststhat this enhancer is indeed criticalfor the normal expression of theAP-2� gene in this region. We arecurrently testing this idea by gener-ating a new allele of the AP-2� genethat is lacking this enhancer. Thesecond conclusion that can bemade from this analysis is that thesequences required for activation ofthe null allele in the AER are distinctfrom those present in intron 5, whichdrive expression in the mesen-chyme, because these latter se-quences have been deleted fromthe mouse genome.
The expression pattern of the lacZtransgene in the limb buds of wild-type and AP-2�–knockout mice alsoprovides an explanation for theagenesis of the radius observed inthe majority of AP-2�–null animals(Schorle et al., 1996; Zhang et al.,1996). Normally, the endogenousgene and the AP-2�::lacZ transgeneare expressed as a stripe in the dor-sal mesenchyme of the zeugopod.The timing and location of this ex-pression domain precedes conden-sation of the radius in the underlyingmesenchyme. In mice lacking AP-2�, this domain of expression fre-quently expands and eventually en-compasses more dorsolateral tissuewithin the forelimb. Significantly, formice in which this alteration in ex-pression occurs, condensation ofthe radius does not happen. Con-versely, AP-2�–null mice that main-tain a discrete stripe of expressionare still able to initiate radius con-densation. These findings support amodel in which the expression ofAP-2� in the dorsal mesenchymegenerates an important cell:cell sig-naling center for radius condensa-tion in the underlying tissue. In theabsence of AP-2�, this signaling cen-ter is either entirely lost, or possiblyrendered ineffective by being dis-persed over a wider area. An alter-native explanation for this phenom-enon is that agenesis of the radiusand the altered expression patternof AP-2� in the zeugopod are notlinked directly but that both are in-dicative of earlier defects within thedeveloping limb bud. In this regard,
it is apparent that in E12.5 AP-2�–nullmice the handplate is frequentlymisshapen (see Fig. 7, and J.Z. un-published observations). Experi-ments in murine, avian, and amphib-ian systems have demonstrated thatthe formation of anterior limb struc-tures can be affected by manipulat-ing the interaction between the AERand the progress zone, which mayact by lowering the amount of mes-enchyme in the developing limbbud (Alberch and Gale, 1983; Ohu-chi et al., 1997; Lettice et al., 1999).Thus, inappropriate proliferationand/or cell:cell signaling within theprogress zone and AER of AP-2�–nullmice could eventually result in theobserved abnormalities in the pat-tern of gene expression and boneformation in the zeugopod region.Defective early limb bud patterninghas also been proposed for theproblems with radius formation thatare seen in both RAR�� double-mu-tant mice and in mice lacking hoxa-11/hoxd-11 (Lohnes et al., 1994;Davis et al., 1995). These particularRAR and hox mutant combinationscause development defects in theforelimb to a greater extent than inthe hindlimb. Similarly, all the effectsof AP-2� so far documented areforelimb-specific: alterations werenot previously reported in the hind-limbs of either AP-2�-null mice or chi-meric animals (Schorle et al., 1996;Zhang et al., 1996; Nottoli et al.,1998). The finding that the activity ofthe AP-2� promoter is only alteredwithin the forelimb of the AP-2�–nullmouse when compared with thefore- and hindlimbs of wild-typemice may provide a mechanistic linkto explain this discrepancy. Finally,we note that the action of AP-2� inthe forelimb must be somewhat re-dundant, because radius formationstill occurs at a low frequency evenin the absence of AP-2�. Becausethe AP-2� gene is also expressed inthe developing limb bud, this re-lated gene may compensate for theloss of AP-2� with respect to limbpatterning (Chazaud et al., 1996).
In summary, we have identifieddistinct regulatory elements respon-sible for directing AP-2� expressionto important sites of AP-2� functionthroughout embryogenesis. Thedata obtained from our current
studies also indicate that AP-2� canregulate epithelial:mesenchymal in-teractions in the limb. It is at presentuncertain whether AP-2� acts bymeans of similar tissue interactionmechanisms to regulate other mor-phogenetic processes, such as de-velopment of the facial promi-nences, eye, and/or nervous system.Alternatively, in these systems AP-2�may act in a cell autonomous man-ner to control proliferation, differen-tiation, or cell survival (Schorle et al.,1996; Zhang et al., 1996; West-Mayset al., 1999; Kramer et al., 2000).Whether or not there is a unified cel-lular basis for the influence of AP-2�on developmental decisions, it re-mains possible that AP-2� still exertsits effects through a common set oftarget genes, such as cell adhesionmolecules, cell surface receptors,and cell-cycle regulated proteins(Mitchell et al., 1987; Leask et al.,1991; Fini et al., 1994; Zutter et al.,1994; Gaubatz et al., 1995; Bosher etal., 1996; Hennig et al., 1996; Zeng etal., 1997). Further analysis of the reg-ulatory network responsible for con-trolling AP-2� expression will comple-ment studies on these downstreamtargets and lead to greater insightinto many aspects of development.
EXPERIMENTAL PROCEDURES
Construction of Transgenes
The isolation of human cosmidclones spanning the AP-2� gene lo-cus was described previously(Creaser et al., 1996). Two of thesecosmid clones were used to makethe AP-2�::lacZ reporter constructs:the Cosh1 clone extends more 5� ofthe AP-2�–coding region, whereasCosh7 contains more 3� sequence.The construct C1 lacZ 42 was theparent plasmid for the generation ofthe AP-2�::lacZ fusions (Fig. 1A). Thesequence between nucleotides �10and �321 of the AP-2� gene wasamplified by polymerase chain re-action (PCR) using the forwardprimer TWY 4 (5�-GCC TAC TCG AGAAGC TTA TGC ATG TCG ACG GAAAAG TTT CTA CCA TTA GAG-3�) andreverse primer TWY 2 (5�-GCC TACCCG GGG TCC TCG TAC TTG ATATTA TCC-3�). The sequences corre-sponding to AP-2� are underlined inthese primers, and the flanking nu-
204 ZHANG AND WILLIAMS

cleotides contain recognition se-quences for the restriction enzymesXhoI, HindIII, NsiI, and SalI in TWY4,and SmaI in TWY 2. PCR was per-formed on Cosh1 that had been lin-earized with ClaI. PCR conditionswere 2 cycles at 94°C for 1 min 20sec, 52°C for 30 sec, 72°C for 3 min;29 cycles at 94°C for 40 sec, 72°C for3 min; and 1 cycle at 72°C for 12 min.Subsequently, the 360-bp PCR frag-ment was digested with XhoI andSmaI and cloned into the corre-sponding sites of the plasmid pLZFSV(Bradshaw et al., 1996). The integrityof the resulting C1 lacZ 42 plasmidwas confirmed by DNA sequencing.
A series of constructs containinggreater amounts of genomic DNAfused to lacZ were made by usingstandard cloning procedures andavailable restriction enzyme sites(see Fig. 1B). First, plasmids weremade that contained the indicatedamount of sequence upstream ofthe transcriptional start site: C1 Xho �gal, 190 bp; C1 Sal � gal, 2 kb; C1Acc � gal, 3.8 kb; C1 Sph � gal, 4.2kb; and C1 Mlu � gal, 8.5 kb (con-structs 1–5, Fig. 1B). The C1 FL � galplasmid (construct 6) contains themaximum amount of 5� flanking se-quence from the cosmid Cosh1. Thisplasmid was made by subcloning anapproximately 18-kb HindIII–SalIfragment of Cosh1 into the corre-sponding sites of C1 Sal � gal. Notethat the HindIII site is located withinthe pWE15 vector sequences, 60 bpfrom the junction with the humangenomic DNA. The actual upstreamlimit of the AP-2� genomic se-quence is an Sau3A I site.
Standard cloning procedureswere also used to generate a seriesof plasmids in which sequences lo-cated downstream of the AP-2�transcriptional initiation sites wereplaced 3� to the lacZ gene. In theplasmid pBS/ACC/M12 (construct 7),a 5.4-kb fragment of AP-2� genomicsequences, from the RsaI site in thefirst exon to the HindIII site in the sec-ond intron, was inserted into the 3�polylinker of the plasmid C1 Acc �gal. The plasmid pBS/ACC/MS (con-struct 8) was made in a similar man-ner, using a 7.5-kb fragment ofCosh1 that extends from an MunI sitein the second intron to the SpeI site inthe fifth exon. The plasmid FX18
(construct 9) is a derivative of C1 FL� gal (construct 6) that contains anadditional 24 kb of downstream se-quence extending from the SpeI sitein the fifth exon until the pWE15 vec-tor boundary of Cosh7. The plasmidFZ19 (construct 10) contains thesame downstream fragment placedinto C1 Xho � gal. The plasmids J24and J25 (constructs 11 and 12) weregenerated by cloning a down-stream 4.2-kb SpeI–NotI fragment ofCosh I into the corresponding sites ofC1 Mlu � gal (construct 5) and C1Xho � gal (construct 1), respectively.The downstream genomic fragmentspans from the SpeI site in the fifthexon to a Sau3A I site in the 3� un-translated region of the seventhexon, and then on to the adjacentNotI site in the pWE15 vector. Con-structs 13 and 14 are derivatives ofJ25 (construct 12). The plasmid SN48(construct 13) contains a 1.95-kb in-sert of AP-2� extending from the SpeIsite in the fifth exon to the BsaA I sitein the sixth exon. Plasmid SN25 (con-struct 14), contains a 2.2-kb frag-ment which spans from the BsaA Isite in the sixth exon to the Sau3A Isite in the 3� untranslated region ofthe seventh exon.
Generation of Transgenic Mice
Before microinjection, vector and in-sert sequences were separated bytreatment with appropriate restrictionenzymes, followed by sucrose densitycentrifugation. The fragments corre-sponding to the AP-2�::lacZ insertfragments were dialyzed extensivelyagainst 10 mM Tris � Cl; 1 mM EDTA, pH7.5. Transgenic mice were generatedby microinjecting DNA into pronucleiof fertilized oocytes of inbred FVBmice (Taconic). The embryos survivingthe microinjection were transferredinto oviducts of pseudopregnantCD-1 fosters (Charles River). The con-centration of DNA used in the micro-injection was adjusted to 1–5 �g/ml,based on the length of the con-structs. The founder embryos wereeither analyzed for the expression ofthe transgene at the embryonictime points indicated in the text, orwere allowed to develop to term.Transgenic mouse lines derived inthe latter instance were used to an-alyze the expression of the lacZ
marker in subsequent generations ofFVB embryos. Only transgenes thatgave a reproducible pattern of lacZexpression were considered to har-bor specific AP-2� cis-regulatory ele-ments. The derivation of AP-2�–knockout mice on a Black Swissbackground has been describedpreviously (Zhang et al., 1996). Theanalysis of transgene activity in theAP-2�–null background was per-formed on a mixed FVB/Black Swissbackground.
Isolation of Genomic DNA,Southern Blot, and PCRAnalysis
Identification of transgenic micewas performed by Southern blot orPCR analysis. Genomic DNA was iso-lated from either the placentas ofembryos or tails of 3- to 4-week-oldmice, as described (Laird et al.,1991). A total of 12 �g of genomicDNA was digested with the restric-tion enzyme BamHI, electropho-resed on a 0.8% agarose gel, andtransferred to nitrocellulose (Schlei-cher & Schuell) or Hybond N filters(Amersham Biosciences). The filterwas hybridized with a 3-kb BamHIfragment of the bacterial lacZ gene,which was either radioactively la-beled with 32P by random priming orlabeled nonradioactively with the“Genius Nonradioactive NucleicAcid Labeling and Detection Kit”(Roche Applied Science). Detectionwas then performed by autoradiog-raphy or light-emission, respectively.PCR was performed using the for-ward primer LacZ-F1 (5�-TAC AACGTC GTG ACT GGG AA-3�) and re-verse primer LacZ-R1 (5�-CCA GATAAC TGC CGT CAC TC-3�), whichare equivalent to nucleotides 45-65and 606-586 of the plasmidpMC1871 (Amersham Biosciences).The PCR conditions were 1 cycle at94°C for 1 min 20 sec; 32 cycles at94°C for 45 sec, 60°C for 45 sec, 72°Cfor 2 min; and 1 cycle at 72°C for 10min. A fragment of 562 bp was am-plified from mice containing thetransgene.
Detection of �-GalactosidaseActivity
Embryos at specific stages of devel-opment were obtained by timed
AP-2� EXPRESSION DURING MORPHOGENESIS 205

matings. The day of the vaginal plugwas considered 0.5 days post co-itum (E0.5). The embryos were dis-sected free from the maternal tissuesinto cold PBS, fixed for 30 min in0.25% glutaraldehyde, rinsed withPBS, and stained for �-galactosidaseactivity as described previously(Bieberich et al., 1990). Stained em-bryos were post-fixed in 4% parafor-maldehyde at 4°C overnight andembedded in paraffin for section-ing. Six- to 8-�m sections were cutand mounted on poly-L-lysine–coated slides. The sections werecounterstained with nuclear fast red.
In Situ Hybridization andImmunohistochemistry
Whole-mount in situ hybridizationwas performed essentially as de-scribed (Wilkinson, 1992) by using theplasmid TRIP1, which contains themouse AP-2� cDNA sequences cor-responding to amino acids 97 to 437inserted between the HindIII andEcoRI sites of pBluescript II KS- (Strat-agene). The plasmid was digestedwith HinddIII, and then transcribedwith T7 RNA polymerase in the pres-ence of digoxigenin-UTP, by usingconditions described by the manu-facturer (Roche Applied Science).Whole-mount immunohistochemistrywas performed as described (Mark etal., 1993) by using the 3B5 AP-2�-spe-cific monoclonal antibody (Zhang etal., 1996).
ACKNOWLEDGMENTSWe thank Frank Ruddle, CouduvalliShashikant, Suzanne Bradshaw, andCarol Arthur for assistance with trans-genic mouse production and analy-sis. We thank Ed Laufer for criticaldiscussion concerning limb morphol-ogy. We are also grateful to the Wil-liams’ laboratory, in particularStephanie Brewer, Amy Donner, andDanielle Nelson for critical reading ofthe manuscript, Stephanie Donald-son and Heidi Auman for generatingspecific strains of mice, and Mei Hanfor technical help. J.Z. was the recip-ient of a breast cancer research fel-lowship from the Department of De-fense (DAMD17-96-6094). T.W. wasfunded by the National Institutes ofHealth. The sequence of the fifth in-
tron has been deposited with Gen-bank, bankit no. 397758.
REFERENCES
Alberch P, Gale EA. 1983. Size-depen-dence during the development of theamphibian foot: colchicine induceddigital loss or reduction. J Embryol ExpMorphol 76:177–197.
Auman HJ, Nottoli T, Lakiza O, Winger Q,Donaldson S, Williams T. 2002. Transcrip-tion factor AP-2� is essential in the ex-tra-embryonic lineages for earlypostimplantation development. Devel-opment 129:2733–2747.
Bar-Eli M. 2001. Gene regulation in mela-noma progression by the AP-2 tran-scription factor. Pigment Cell Res 14:78–85.
Bauer R, Imhof A, Pscherer A, Kopp H,Moser M, Seegers S, Kerscher M, TainskyMA, Hofstaedter F, Buettner R. 1994.The genomic structure of the humanAP-2 transcription factor. Nucleic AcidsRes 22:1413–1420.
Bieberich CJ, Utset MF, Awgulewitsch A,Ruddle FH. 1990. Evidence for positiveand negative regulation of the Hox-3.1gene. Proc Natl Acad Sci U S A 87:8462–8466.
Bosher JM, Williams T, Hurst HC. 1995. Thedevelopmentally regulated transcrip-tion factor AP-2 is involved in c-erbB-2overexpression in human mammarycarcinoma. Proc Natl Acad Sci U S A92:744–747.
Bosher JM, Totty NF, Hsuan JJ, Williams T,Hurst HC. 1996. A family of AP-2 proteinsregulates c-erbB-2 expression in mam-mary carcinoma. Oncogene 13:1701–1707.
Bradshaw MS, Shashikant CS, Belting HG,Bollekens JA, Ruddle FH. 1996. A long-range regulatory element of Hoxc8identified by using the pClasper vector.Proc Natl Acad Sci U S A 93:2426–2430.
Brewer S, Jiang X, Donaldson S, Williams T,Sucov HM. 2002. Requirement forAP-2� in cardiac outflow tract morpho-genesis. Mech Dev 110:139–149.
Byrne C, Tainsky M, Fuchs E. 1994. Pro-gramming gene expression in develop-ing epidermis. Development 120:2369–2383.
Chazaud C, Oulad-Abdelghani M, Bouil-let P, Decimo D, Chambon P, Dolle P.1996. AP-2.2, a novel gene related toAP-2, is expressed in the forebrain,limbs and face during mouse embryo-genesis. Mech Dev 54:83–94.
Cheng C, Ying K, Xu M, Zhao W, Zhou Z,Huang Y, Wang W, Xu J, Zeng L, Xie Y,Mao Y. 2002. Cloning and character-ization of a novel human transcriptionfactor AP-2 beta like gene (TFAP2BL1).Int J Biochem Cell Biol 34:78–86.
Creaser P, D’Argenio DA, Williams T. 1996.Comparative and functional analysesof the AP-2 promoter indicates thatconserved octamer and initiator ele-ments are critical for activity. NucleicAcids Res 13:2597–2605.
Davies AF, Stephens RJ, Olavesen MG,Heather L, Dixon MJ, Magee A, Flinter F,Ragoussi J. 1995. Evidence of a locusfor orofacial clefting on human chro-mosome 6p24 and STS content map ofthe region. Hum Mol Genet 4:121–128.
Davies AF, Mirza G, Flinter F, Ragoussis J.1999. An interstitial deletion of 6p24-p25 proximal to the FKHL7 locus andincluding AP-2� that affects anterioreye chamber development. J MedGenet 36:708–710.
Davis AP, Witte DP, Hsieh-Li HM, Potter SS,Capecchi MR. 1995. Absence of radiusand ulna in mice lacking hoxa-11 andhoxd-11. Nature 375:791–795.
Feng W, Williams T. 2003. Cloning andcharacterization of the mouse AP-2�gene: a novel family member ex-pressed in the developing olfactorybulb. Mol Cell Neurosci (in press).
Fini ME, Bartlett JD, Matsubara M, Rine-hart WB, Mody MK, Girard MT, RainvilleM. 1994. The rabbit gene for 92-kDamatrix metalloproteinase. Role of AP1and AP2 in cell type-specific transcrip-tion. J Biol Chem 269:28620–28628.
Gaubatz S, Imhof A, Dosch R, Werner O,Mitchell P, Buettner R, Eilers M. 1995.Transcriptional activation by Myc is un-der negative control by the transcrip-tion factor AP-2. EMBO J 14:1508–1519.
Gee JM, Robertson JF, Ellis IO, NicholsonRI, Hurst HC. 1999. Immunohistochemi-cal analysis reveals a tumour suppres-sor-like role for the transcription factorAP-2 in invasive breast cancer. J Pathol189:514–520.
Hennig G, Lowrick O, Birchmeier W, Beh-rens J. 1996. Mechanisms identified inthe transcriptional control of epithelialgene expression. J Biol Chem 271:595–602.
Hilger-Eversheim K, Moser M, Schorle H,Buettner R. 2000. Regulatory roles ofAP-2 transcription factors in vertebratedevelopment, apoptosis and cell-cy-cle control. Gene 260:1–12.
Hogan B, Beddington R, Costantini F,Lacy E. 1994. Manipulating the mouseembryo. Cold Spring Harbor: ColdSpring Harbor Laboratory Press.
Kerber B, Monge I, Mueller M, Mitchell PJ,Cohen SM. 2001. The AP-2 transcriptionfactor is required for joint formationand cell survival in Drosophila leg de-velopment. Development 128:1231–1238.
Kramer PR, Krishnamurthy R, Mitchell PJ,Wray S. 2000. Transcription factor Acti-vator Protein-2 is required for contin-ued luteinizing hormone-releasing hor-mone expression in the forebrain ofdeveloping mice. Endocrinology 141:1823–1838.
Laird PW, Zijderveld A, Linders K, RudnickiMA, Jaenisch R, Berns A. 1991. Simpli-fied mammalian DNA isolation proce-dure. Nucleic Acids Res 19:4293.
Leask A, Byrne C, Fuchs E. 1991. Transcrip-tion factor AP2 and its role in epider-mal-specific gene expression. Proc NatlAcad Sci U S A 88:7948–7952.
206 ZHANG AND WILLIAMS

Lettice L, Hecksher-Sorensen J, Hill RE.1999. The dominant hemimelia muta-tion uncouples epithelial-mesenchy-mal interactions and disrupts anteriormesenchyme formation in mouse hind-limbs. Development 126:4729–4736.
Lohnes D, Mark M, Mendelsohn C, DolleP, Dierich A, Gorry P, Gansmuller A,Chambon P. 1994. Function of the reti-noic acid receptors (RARs) during de-velopment. I. Craniofacial and skeletalabnormalities in RAR double mutants.Development 120:2723–2748.
Luscher B, Mitchell PJ, Williams T, Tjian R.1989. Regulation of transcription factorAP-2 by the morphogen retinoic acidand by second messengers. GenesDev 3:1507–1517.
Mark M, Lufkin T, Vonesch JL, Ruberte E,Olivo JC, Dolle P, Gorry P, LumsdenAPC. 1993. Two rhombomeres are al-tered in Hoxa-1 mutant mice. Develop-ment 119:319–338.
Maytin EV, Lin JC, Krishnamurthy R, Batch-varova N, Ron D, Mitchell PJ, HabenerJF. 1999. Keratin 10 gene expressionduring differentiation of mouse epider-mis requires transcription factors C/EBPand AP-2. Dev Biol 216:164–181.
Mitchell PJ, Wang C, Tjian R. 1987. Positiveand negative regulation of transcrip-tion in vitro: enhancer-binding proteinAP-2 is inhibited by SV40 T antigen. Cell50:847–861.
Mitchell PJ, Timmons PM, Hebert JM,Rigby PWJ, Tjian R. 1991. Transcriptionfactor AP-2 is expressed in neural crestcell lineages during mouse embryo-genesis. Genes Dev 5:105–119.
Monge I, Krishnamurthy R, Sims D, Hirth F,Spengler M, Kammermaier L, ReichertH, Mitchell PJ. 2001. Drosophila tran-scription factor AP-2 in proboscis, legand brain central complex develop-ment. Development 128:1239–1252.
Moser M, Imhof A, Pscherer A, Bauer R,Amselgruber W, Sinowatz F, HofstadterF, Schule R, Buettner R. 1995. Cloningand characterization of a second AP-2transcription factor: AP-2�. Develop-ment 121:2779–2788.
Moser M, Pscherer A, Roth C, Becker J,Mucher G, Zerres K, Dixkens C, Weis J,Guay-Woodford L, Buettner R, Fassler R.1997a. Enhanced apoptotic cell death
of renal epithelial cells in mice lackingtranscription factor AP-2�. Genes Dev11:1938–1948.
Moser M, Ruschoff J, Buettner R. 1997b.Comparative analysis of AP-2� andAP-2� gene expression during mouseembryogenesis. Dev Dyn 208:115–124.
Nottoli T, Hagopian-Donaldson S, ZhangJ, Perkins A, Williams T. 1998. AP-2-nullcells disrupt morphogenesis of the eye,face, and limbs in chimeric mice. ProcNatl Acad Sci U S A 95:13714–13719.
Ohuchi H, Nakagawa T, Yamamoto A,Araga A, Ohata T, Ishimaru Y, YoshiokaH, Kuwana T, Nohno T, Yamasaki M,Itoh N, Noji S. 1997. The mesenchymalfactor, FGF10, initiates and maintainsthe outgrowth of the chick limb budthrough interaction with FGF8, an api-cal ectodermal factor. Development124:2235–2244.
Oulad-Abdelghani M, Bouillet P, ChazaudC, Dolle P, Chambon P. 1996. AP-2.2: anovel AP-2-related transcription factorinduced by retinoic acid during differen-tiation of P19 embryonal carcinomacells. Exp Cell Res 225:338–347.
Ross SA, McCaffery PJ, Drager UC, DeLuca LM. 2000. Retinoids in embryonaldevelopment. Physiol Rev 80:1021–1054.
Schorle H, Meier P, Buchert M, Jaenisch R,Mitchell PJ. 1996. Transcription factorAP-2 essential for cranial closure andcraniofacial development. Nature 381:235–238.
Shen H, Wilkie T, Ashique AM, Narvey M,Zerucha T, Savino E, Williams T, Rich-man JM. 1997. Chicken transcriptionfactor AP-2: cloning, expression and itsrole in outgrowth of facial prominencesand limb buds. Dev Biol 188:248–266.
Tickle C. 2003. Patterning systems-fromone end of the limb to the other. DevCell 4:449–458.
Topping A, Harris P, Moss AL. 2002. The 6pdeletion syndrome: a new orofacialclefting syndrome and its implicationsfor antenatal screening. Br J Plast Surg55:68–72.
Turner BC, Zhang J, Gumbs AA, MaherMG, Kaplan L, Carter D, Glazer PM,Hurst HC, Haffty BG, Williams T. 1998.Expression of AP-2 transcription factorsin human breast cancer correlates with
the regulation of multiple growth factorsignalling pathways. Cancer Res 58:5466–5472.
Werling U, Schorle H. 2002. Transcriptionfactor gene AP-2� essential for earlymurine development. Mol Cell Biol 22:3149–3156.
West-Mays JA, Zhang J, Nottoli T, Hago-pian-Donaldson S, Libby D, Strissel KJ,Williams T. 1999. AP-2� transcriptionfactor is required for early morphogen-esis of the lens vesicle. Dev Biol 206:46–62.
Wilkie AO, Morriss-Kay GM. 2001. Genet-ics of craniofacial development andmalformation. Nat Rev Genet 2:458–468.
Wilkinson DG, editor. 1992. In situ hybrid-ization: a practical approach. Oxford:Oxford University Press.
Williams T, Admon A, Luscher B, Tjian R.1988. Cloning and expression of AP-2, acell-type-specific transcription factorthat activates inducible enhancer ele-ments. Genes Dev 2:1557–1569.
Zeng Y-X, Somasundaram K, El-Deiry WF.1997. AP2 inhibits cancer cell growthand activates p21WAF1/CIP1 expression.Nat Genet 15:78–82.
Zhang J, Hagopian-Donaldson S, Ser-bedzija G, Elsemore J, Plehn-DujowichD, McMahon AP, Flavell RA, Williams T.1996. Neural tube, skeletal and bodywall defects in mice lacking transcrip-tion factor AP-2. Nature 381:238–241.
Zhao F, Satoda M, Licht JD, Hayashizaki Y,Gelb BD. 2001. Cloning and character-ization of a novel mouse AP-2 transcrip-tion factor, Ap-2�, with unique DNAbinding and transactivation properties.J Biol Chem 276:40755–40760.
Zhang J, Brewer S, Huang J, Williams T.2003. Overexpression of transcriptionfactor AP-2� suppresses mammarygland growth and morphogenesis. DevBiol 256:127–145.
Zutter MM, Santoro SA, Painter AS, TsungYL, Gafford A. 1994. The human �2 in-tegrin gene promoter. Identification ofpositive and negative regulatory ele-ments important for cell-type and de-velopmentally restricted gene expres-sion. J Biol Chem 269:463–469.
AP-2� EXPRESSION DURING MORPHOGENESIS 207




![Basic and depression specific emotion identification in ...webdocs.cs.ualberta.ca/~zaiane/postscript/CICLING19-1.pdf · label emotion mining from text [2,11,17]. With the increasing](https://img.pdfslide.us/doc/110x75/5f4a6fb08857b1069945e82f/basic-and-depression-speciic-emotion-identiication-in-zaianepostscriptcicling19-1pdf.jpg)
























