Embed Size (px)
Citation preview

SUMMARY
The development of plant virology as a science is, al-most totally, a 20th century phenomenon. Early paperson plant viruses emphasize pathogenicity and ultra-fil-terability and report transmission by leafhopper andaphid vectors, and from soil or through seed. Successiveadvances were largely driven by access to new tech-niques. For example, biochemical approaches, and lo-cal-lesion and serological assays, led to purification andcrystallization of virus particles, and their identificationas nucleoproteins; electron microscopy and X-ray crys-tallography revealed their detailed structure. Biologicaltests revealed mite, nematode, chytrid and plasmodio-phorid vectors. Viroids were discovered. Proof of theinfectivity of viral RNA and DNA, followed by sequenc-ing of complete viral genomes, allowed their genes andcontrol elements to be defined. Although some viralgenomes were monopartite and others multipartite,each typically encoded proteins involved in replication,movement within the plant, particle structure and, com-monly, transmission by vectors. The processes involvedin vector transmission, viral replication, disease induc-tion and resistance, and the role of gene silencing, werethen studied intensively. Genetic variability was ana-lyzed and short-term evolution explored. A soundly-based virus taxonomy was devised and now includesover 70 plant virus genera containing some 800 species.Virus ecological and epidemiological research estab-lished patterns of virus spread and aided the develop-ment of control measures. These include schemes forproducing virus-tested stocks of vegetatively propagatedplants, quarantine regulations, application of vector-controlling chemicals to crops and use of virus-resistantcultivars that contain conventional or man-made resist-ance genes. These and other changes in agriculturalpractices have resulted in substantial increases in cropyields and food security. Since about 1950, results of re-search on viruses infecting the whole spectrum of host
Corresponding author: B.D. HarrisonFax: +44.1382562426E-mail: [email protected]
taxa have been integrated to constitute virology, withplant virology contributing many seminal discoveries.The biological phase of plant virology has progressed tomolecular biological and molecular genetic phases, andplant viruses have become tools in molecular biology,cell biology and biotechnology.
INTRODUCTION
To review the advances in plant virology made in the20th century is a formidable task because few seminaldiscoveries predate this period and subsequent progresswas increasingly rapid. Indeed, the study of viruses didnot become widely known as ‘virology’ before about1950 (Luria, 1953). However, the development of plantvirology is not simply a matter of dating major discover-ies. It also concerns the people involved, their rationalthought, serendipity and mistakes; and the enormousand continuing advance in techniques. Progress hassomewhat resembled the Dance of the Seven Veils, witha new technique enabling another veil to be removedand increasingly intimate details of the subject to be re-vealed. In the following account, to give some historicalperspective, I shall consider developments in each suc-cessive quarter century.
1900-1925 DIVERSE BEGINNINGS
During the first quarter of the 20th century, the mostimportant techniques available for studying plant virus-es were transmission by inoculation of sap and by graft-ing, and ultra-filtration to differentiate viruses from bac-terial pathogens (Table 1). Mosaic and leaf curl diseasesymptoms were reported in cultivated plants, includingtobacco, potato, cucurbits, legumes and sugar beet. In-ternal symptoms, including crystalline and amorphousinclusion bodies, were described in plants with tobaccomosaic disease (Ivanowski, 1903). Transmission byleafhoppers (Takami, 1901), by aphids (Doolittle, 1916;Jagger, 1916) and by beetles (Smith, 1924), and fromsoil (Behrens, 1899) and through seed (Reddick andStewart, 1919), was reported or confirmed. Viral
Journal of Plant Pathology (2009), 91 (3), 509-520 Edizioni ETS Pisa, 2009 509
INVITED REVIEW
A BRIEF OUTLINE OF THE DEVELOPMENT OF PLANT VIROLOGYIN THE 20TH CENTURY
B.D. Harrison
Scottish Crop Research Institute, Invergowrie, Dundee DD2 5DA, UK
001_JPPReview(Harrison)_509 23-11-2009 17:23 Pagina 509

510 Plant virology in the 20th century Journal of Plant Pathology (2009), 91 (3), 509-520
Table 1. Some important techniques and research aids devised or adopted in plant virus research.
Period Technique/research aid
1900-1925. Diverse beginnings
Transmission by sap and graft inoculation Transmission by insect vectors Ultrafiltration Use of test plants
1925-1950. The impact of biochemistry
Local lesion infectivity assay Serological assay Protein chemistry and protein isolation Centrifugation X-ray diffraction analysis Abrasives as aids in transmission by manual inoculation Cross-protection tests Vector culturing Screened glasshouses
Virus acquisition by vectors fed through a membrane or injected Early electron microscopy
1950-1975. The rise of molecular biology
Preparative and analytical ultracentrifugation Density-gradient centrifugation Electron microscopy of shadowcast specimens Negative staining in electron microscopy Ultrathin sectioning Gel-diffusion serological tests
Polyacrylamide gel electrophoresis of proteins and nucleic acids Amino-acid sequencing of proteins Nucleic acid isolation Radioactive labelling Chemical mutagenesis X-ray crystallography Protoplast systems Heat therapy Meristem-tip culture Application of vector-killing chemicals to crops
Introgression in crop plants of resistance genes from wild relatives
1975-2000. Viral genomes and molecular genetics
Reverse transcription Restriction enzymes Nucleic acid cloning and sequencing Polymerase chain reaction (PCR)
Computerised analysis of nucleotide and amino-acid sequencesSouthern and other blotting procedures
Site-directed mutagenesis Enzyme-linked immunosorbent assay (ELISA) Immunosorbent electron microscopy (ISEM) Monoclonal antibodies Rapid diagnostic procedures Production of transgenic plants Protein markers: jellyfish green fluorescent protein Confocal laser-scanning microscopy of living cells Gene silencing Mapping host genes and use of gene markers
001_JPPReview(Harrison)_509 23-11-2009 17:23 Pagina 510

Journal of Plant Pathology (2009), 91 (3), 509-520 B.D. Harrison 511
pathogens seemed unable to enter plants unaided but,once established, they spread through the whole plantand to its vegetative progeny, although usually notthrough true seed.
1925-1950 THE IMPACT OF BIOCHEMISTRY
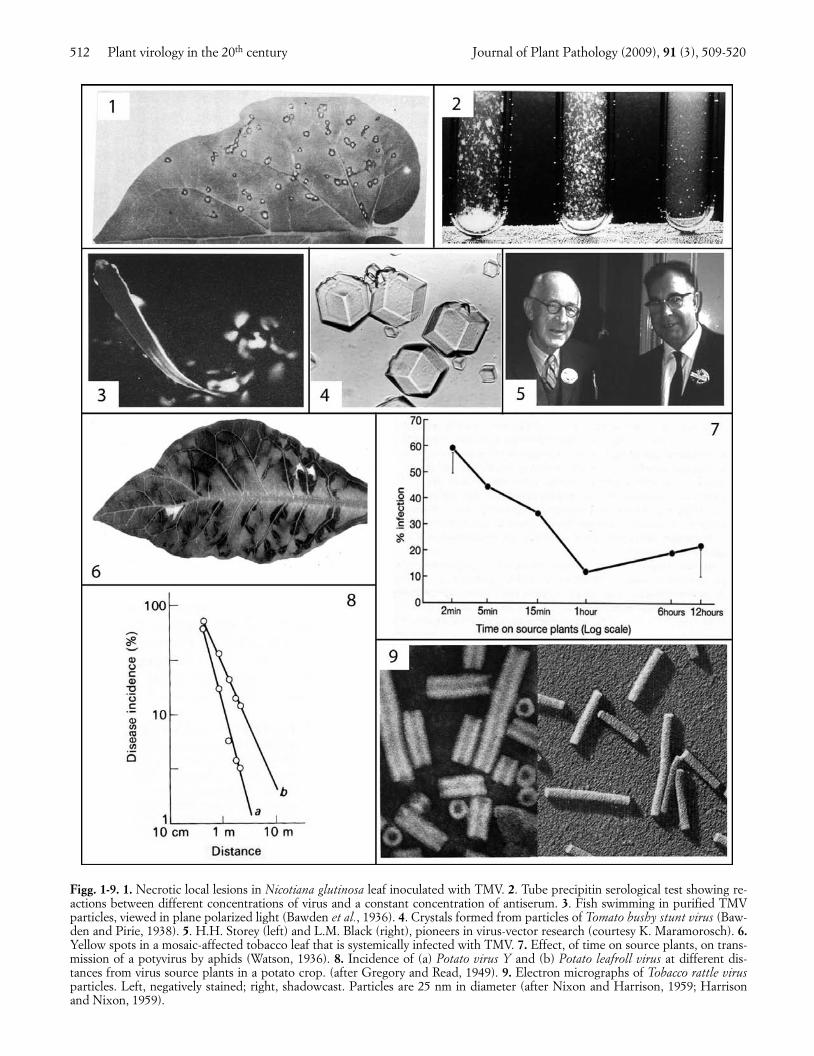
The second quarter century saw the impact of theemerging science of biochemistry, especially aspects in-volving proteins, and the development of much im-proved quantitative assays. For example, FrancisHolmes (1929) discovered the local lesion response tomanual inoculation (Fig. 1), and went on to show thatit could be the basis of a much improved infectivity as-say, and also of a highly effective form of resistancecontrolled by a single dominant gene (Holmes, 1934).Animal virologists did not have anything like this. Inaddition, serological assay methods (Fig. 2) were ap-plied to plant viruses, especially by Helen Purdy (Pur-dy, 1928). Work to isolate and purify viral entities, de-scribed initially by Kenneth Smith as like ‘looking for ablack cat in a dark cellar without being sure it wasthere’, now began in earnest. Success was achievedwhen Wendell Stanley (1935) applied methods thatwere being used by colleagues to purify enzymes. Hedescribed infectious preparations of ‘needle-like crys-tals’ of Tobacco mosaic virus (TMV), and concludedthey consisted entirely of protein. However, more care-ful analysis by Fred Bawden and Bill Pirie (Bawden etal., 1936) showed that such purified preparations con-sisted of 94% protein and 6% ribonucleic acid. Stan-ley’s needles were in fact paracrystals produced by sideto side aggregation of elongated particles, so explainingthe anisotropy of flow seen when preparations wereswirled in plane polarized light, a phenomenon report-ed earlier by Takahashi and Rawlins (1932). It can beseen when a fish swims through purified TMV (Fig. 3).In addition, J.D. Bernal and I. Fankuchen, Bawden andPirie’s collaborators, showed by X-ray diffractionanalysis that the viral entities had a repeating substruc-ture (Bawden et al., 1936). Soon afterwards, Bawdenand Pirie (1938) went on to purify the particles ofTomato bushy stunt virus (TBSV), and produced true 3-dimensional crystals (Fig. 4). The discovery that aminute pathogen could be crystallized cast a new lighton viruses in general, caused much debate about theirpossible origins and stimulated biophysicists to charac-terize the virus particles in more detail. In particular,development of the electron microscope (Kausche etal., 1939) was to revolutionize knowledge of the mor-phology of virus particles.
At the plant level, McKinney (1929) found that vari-ant forms of TMV could be cultured from yellow spotsthat appeared in some systemically infected leaves (Fig.6), and later Knight (1947) showed that the protein in
particles of such a variant had an altered amino-acidcomposition: thus viruses mutated, and the mutantscould be sub-cultured and distinguished chemically.McKinney (1929) and Thung (1931) discovered, inde-pendently, that plants infected with one strain of a viruswere resistant to subsequent infection by other strainsof the same virus (termed cross-protection), althoughthey remained susceptible to other viruses. Samuel(1934) and Bennett (1937) plotted the pattern of spreadof viruses through a plant, concluding that local cell-to-cell spread was through plasmodesmata whereas long-distance spread was through the phloem.
Eriophyid mites (Amos et al., 1927), whiteflies(Kirkpatrick, 1931), thrips (Pittman, 1927) and mealybugs (Box, 1945) were added to the list of vectors ofindividual viruses and a start made towards unravellingthe mechanisms of vector transmission. Haydon Storey(1932, 1933; Fig. 5), working in Africa, discovered thepersistent, circulative transmission of Maize streakvirus (MSV) by leafhoppers and showed that a non-vector race of the leafhopper, differing in a major sex-linked gene for transmission, could transmit when itsgut wall was punctured. Neither this virus nor Beetcurly top virus (BCTV), studied by Bennett and Wal-lace (1938), seemed to multiply in vector leafhoppers.However, Fukushi (1940) reported transmission ofRice dwarf virus (RDV) through leafhopper eggs formany successive generations, suggesting that it didmultiply in the leafhoppers, and later Lindsay Black(Fig. 5) and Myron Brakke (1952) proved that Woundtumor virus (WTV) replicated in its leafhopper vector.Meanwhile, Marion Watson (1936) found, remarkably,that aphids transmitted several viruses less frequentlythe longer they fed on virus source plants (Fig. 7), andthat few aphids retained these viruses for longer thanone hour. She called this ‘non-persistent’ transmission.Later she found a contrasting type of transmissionwhen studying Beet yellows virus (BYV): here transmis-sion frequency increased with increase in length, bothof the acquisition feed and of the inoculation feed, andthe virus persisted in the aphid for up to 3 days (Wat-son, 1940). This was the prototype of ‘semi-persistent’transmission.
Field studies in this period led to the beginnings ofplant virus epidemiology and its mathematical treat-ment, especially by Van der Plank (1946), and by Gre-gory (1948). Field spread of virus from plant to plantwas analyzed in terms of time and space (Fig. 8). The ef-fects of vector numbers and activity, of vector efficiencyand of virus-vector relations, began to be recognized.On the applied side, schemes were started, notably inthe Netherlands and the British Isles, to select andpropagate relatively virus-free planting stocks of pota-toes, a development later extended to many other vege-tatively propagated crops.
001_JPPReview(Harrison)_509 23-11-2009 17:23 Pagina 511
512 Plant virology in the 20th century Journal of Plant Pathology (2009), 91 (3), 509-520
Figg. 1-9. 1. Necrotic local lesions in Nicotiana glutinosa leaf inoculated with TMV. 2. Tube precipitin serological test showing re-actions between different concentrations of virus and a constant concentration of antiserum. 3. Fish swimming in purified TMVparticles, viewed in plane polarized light (Bawden et al., 1936). 4. Crystals formed from particles of Tomato bushy stunt virus (Baw-den and Pirie, 1938). 5. H.H. Storey (left) and L.M. Black (right), pioneers in virus-vector research (courtesy K. Maramorosch). 6.Yellow spots in a mosaic-affected tobacco leaf that is systemically infected with TMV. 7. Effect, of time on source plants, on trans-mission of a potyvirus by aphids (Watson, 1936). 8. Incidence of (a) Potato virus Y and (b) Potato leafroll virus at different dis-tances from virus source plants in a potato crop. (after Gregory and Read, 1949). 9. Electron micrographs of Tobacco rattle virusparticles. Left, negatively stained; right, shadowcast. Particles are 25 nm in diameter (after Nixon and Harrison, 1959; Harrisonand Nixon, 1959).
001_JPPReview(Harrison)_509 23-11-2009 17:23 Pagina 512

Journal of Plant Pathology (2009), 91 (3), 509-520 B.D. Harrison 513
1950-1975 THE RISE OF MOLECULAR BIOLOGY
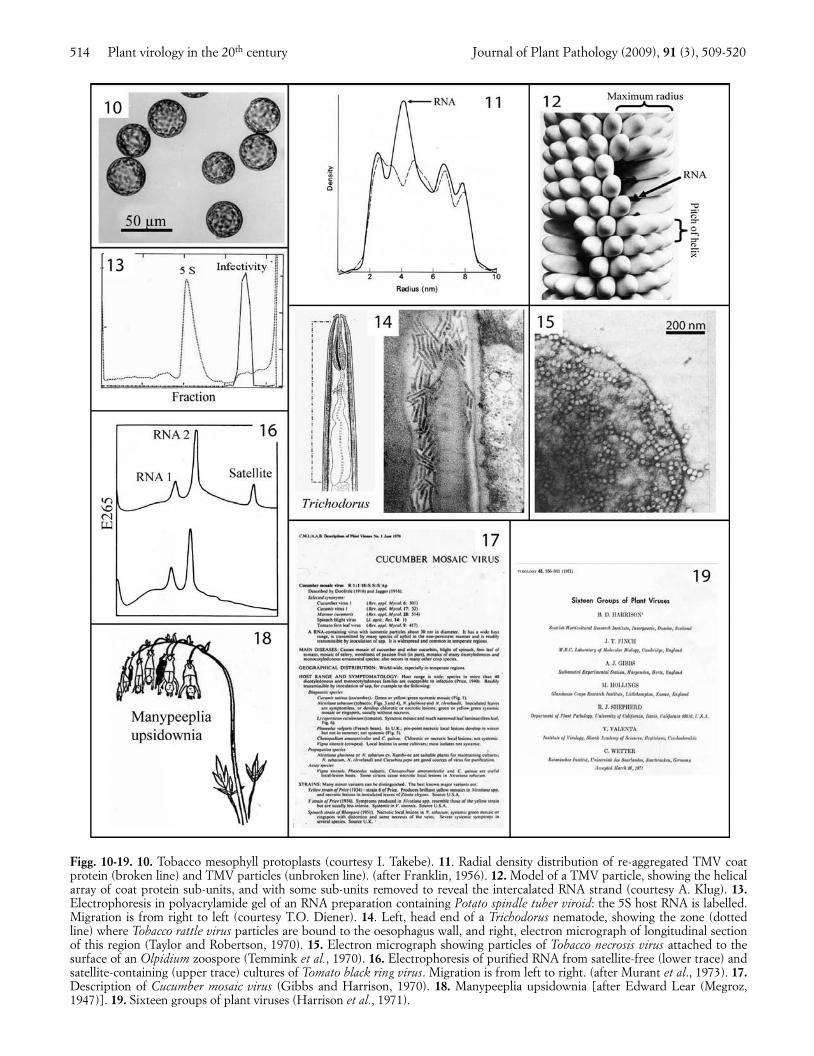
Plant virology entered what can be called its ‘GoldenAge’ in the second half of the 20th century. The years1950-1975 saw the rise of molecular biology and theadoption of many powerful new techniques (Table 1;Fig. 9). Protoplast systems (Fig. 10), pioneered byTakebe and Otsuki (1969), allowed single-step virusgrowth curves to be studied. X-ray diffraction workshowed that TMV particles are helical (Watson, 1954),with 16 and one third protein subunits in each turn ofthe helix (Franklin and Holmes, 1958). To find the posi-tion of the embedded RNA, the radial density distribu-tion was established for intact TMV particles by DonCaspar and for cylindrical aggregates of TMV proteinby Rosalind Franklin. The position of the RNA was in-dicated by comparing these plots. The plots were pub-lished in two contiguous letters to Nature (Caspar, 1956;Franklin, 1956; Fig. 11). A sign of those times is thatFranklin was so keen to get the work published that sheapparently wrote both papers! These advances led tothe well-known helical structure of TMV particles (Fig.12), the first for a virus.
Fraenkel-Conrat and Williams (1955) published afine paper that described the reassembly of TMV parti-cles from their isolated protein and RNA components.Moreover, the reassembled particles were infective al-though the protein and RNA preparations apparentlywere not, resulting in the authors’ claim to have createdinfectivity. However, like Bawden and Pirie 18 years ear-lier, Fraenkel-Conrat and Williams had failed to recog-nize the instability of the isolated RNA, and it was Gier-er and Schramm (1956) who proved that purified TMVRNA was infective. Gierer and Mundry (1958) followedthis up by producing TMV mutants through treatingthe RNA with nitrous acid to deaminate its nucleotidebases. Analysis of the resulting amino-acid replacementsin the coat protein of such TMV mutants, especially byTsugita (1962) and Wittmann (1962), helped to crackthe genetic code.
Work with other viruses began to reveal the varietyof genomes and genetic systems found among plantviruses. Although only one size of TMV RNA moleculewas needed for infection, two or three sizes were need-ed with several other viruses, and with Tobacco rattlevirus (TRV) two sizes were needed for a normal infec-tion but one size could give an abnormal infection (Lis-ter, 1966; Sanger, 1968). Bob Shepherd was the first toreport a DNA genome in a plant virus, Cauliflower mo-saic virus (CaMV) (Shepherd et al., 1968). Basil Kassa-nis gave the name ‘satellite’ to a virus he found in somecultures of Tobacco necrosis virus (TNV): the satellitevirus could only replicate in cells also infected by TNV(Kassanis and Nixon, 1961). Similarly, non-essential butbiologically active satellite RNA molecules were foundin cultures of various other viruses (Schneider, 1971;
Fig. 16). Ted Diener found that the potato spindle tuberagent was a free circular RNA molecule of only about100,000 daltons (Diener, 1971; Fig. 13), much smallerthan a viral genome, and named it a viroid, the first ofmany.
Along with the rapidly increasing numbers and typesof viruses being described, new kinds of vectors werediscovered, including nematodes (Hewitt et al., 1958),chytrids (Teakle, 1960) and plasmodiophorids (Estesand Brakke, 1966; Canova, 1966). Viruses apparently donot multiply in these soil-inhabiting vectors but themechanisms of transmission are diverse. For example,Fig. 14 shows the site of attachment of TRV particlesalong the wall of the oesophagus of a Trichodorus nema-tode (Taylor and Robertson, 1970) and Fig. 15 showsTNV particles bound to the outer surface of an Olpidi-um zoospore (Temmink et al., 1970). The ecology andepidemiology of these soil-borne viruses proved to pres-ent interesting contrasts with those of viruses that haveaerial vectors (Harrison, 1977). Among viruses with in-sect vectors, persistent transmission of WTV involvedsystemic infection of the leafhoppers (Shikata andMaramorosch, 1965; Sinha, 1965), whereas non-persist-ent transmission of potyviruses required a virus-speci-fied ‘helper component’ protein (Govier and Kassanis,1974).
To provide a readily accessible source of the greatlyincreasing amount of information on plant viruses, theseries of Descriptions of Plant Viruses was started(Gibbs and Harrison, 1970; Fig. 17). Individual De-scriptions were written by an international array of au-thors, and tightly edited. These Descriptions, now pub-lished by the Association of Applied Biologists, mod-ernized and provided with genome sequences andcolour illustrations, are available on the Internet forover 400 viruses. The greatly increasing amount of dataavailable enabled a more soundly based virus taxonomy,with approved virus names, to be developed, amid con-siderable controversy. Some, mainly animal, virologists,favoured an approach to the taxonomy that was basedon a few selected properties, and divided the viruses ina top-down manner, from Orders down to Species, withlatinized binomials for the names of individual viruses.Other animal virologists, and most plant virologists, ofwhich I was one, favoured a bottom-up approach tobuilding taxa, which were based on all available viralproperties, and used common names, such as ‘tobaccomosaic virus’, for individual viruses. The rival viewswere presented forcefully and I recall the top-down andlatinized name approach being lampooned as upside-down thinking by Peter Wildy, at the Moscow Interna-tional Congress of Microbiology in 1968, with the aid ofan Edward Lear cartoon that depicted the mythicalplant Manypeeplia upsidownia (Fig. 18). Eventually thebottom-up approach was largely adopted and the first16 groups (now genera) of plant viruses were proposed
001_JPPReview(Harrison)_509 23-11-2009 17:23 Pagina 513
514 Plant virology in the 20th century Journal of Plant Pathology (2009), 91 (3), 509-520
Figg. 10-19. 10. Tobacco mesophyll protoplasts (courtesy I. Takebe). 11. Radial density distribution of re-aggregated TMV coatprotein (broken line) and TMV particles (unbroken line). (after Franklin, 1956). 12. Model of a TMV particle, showing the helicalarray of coat protein sub-units, and with some sub-units removed to reveal the intercalated RNA strand (courtesy A. Klug). 13.Electrophoresis in polyacrylamide gel of an RNA preparation containing Potato spindle tuber viroid: the 5S host RNA is labelled.Migration is from right to left (courtesy T.O. Diener). 14. Left, head end of a Trichodorus nematode, showing the zone (dottedline) where Tobacco rattle virus particles are bound to the oesophagus wall, and right, electron micrograph of longitudinal sectionof this region (Taylor and Robertson, 1970). 15. Electron micrograph showing particles of Tobacco necrosis virus attached to thesurface of an Olpidium zoospore (Temmink et al., 1970). 16. Electrophoresis of purified RNA from satellite-free (lower trace) andsatellite-containing (upper trace) cultures of Tomato black ring virus. Migration is from left to right. (after Murant et al., 1973). 17.Description of Cucumber mosaic virus (Gibbs and Harrison, 1970). 18. Manypeeplia upsidownia [after Edward Lear (Megroz,1947)]. 19. Sixteen groups of plant viruses (Harrison et al., 1971).
001_JPPReview(Harrison)_509 23-11-2009 17:23 Pagina 514

in 1971, with names such as Bromovirus, Potexvirus andTobamovirus (Harrison et al., 1971; Fig. 19). Now thereare about 70 recognized genera and more than 800 rec-ognized species of plant viruses.
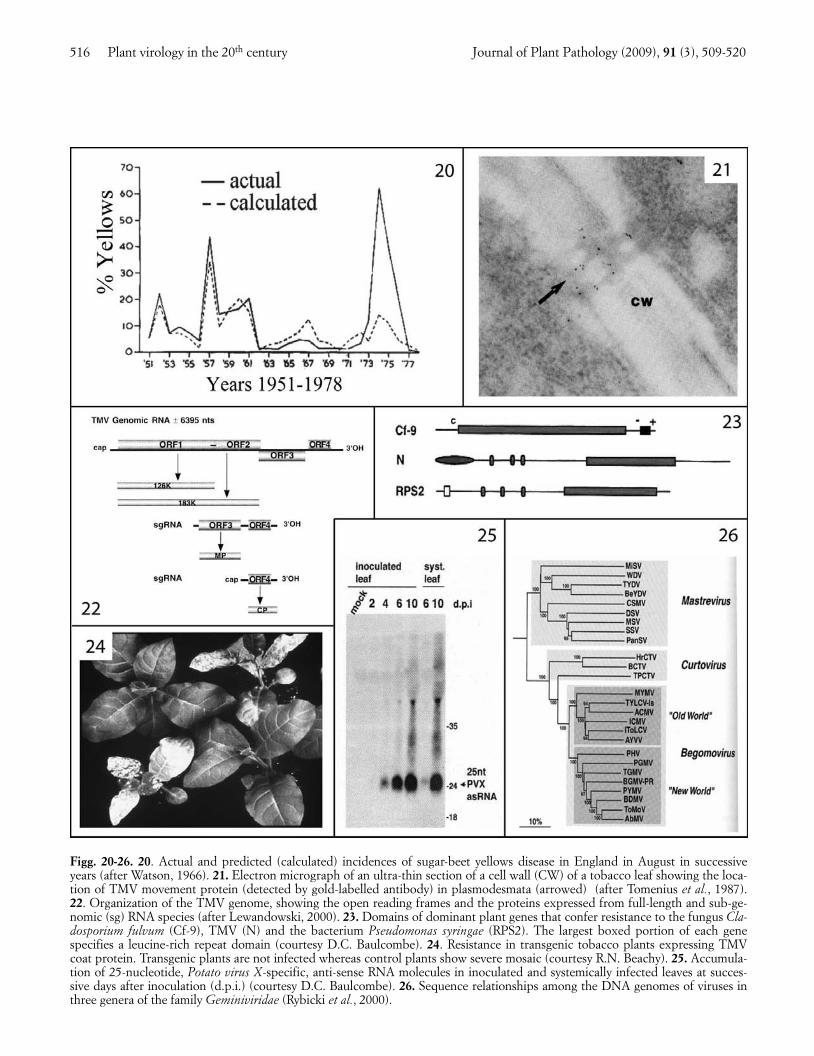
During this period good progress was being made incontrolling plant virus diseases. Systemic organophos-phate insecticides became widely used to control aerialvectors (e.g. Hull and Heathcote, 1967), and nemati-cides to kill vector nematodes in soil (Harrison et al.,1963). In exceptional instances, such as for beet yellow-ing viruses in sugar beet in Britain, a way was devisedfor predicting, from aphid and weather data beforesowing, what the likely disease incidence would be 3-4months later (Watson, 1966; Fig. 20). Thus models thatexplained the progression of epidemics were evolvinginto predictive modelling, which could be used to im-prove the timing and efficacy of pesticide applications.In addition, heat therapy (Kassanis, 1950) and apicalmeristem culture (Morel and Martin, 1952) were adopt-ed for eliminating virus infection from master clones ofvegetatively propagated species, and increasing num-bers of virus-resistant crop cultivars were bred. In duecourse, virus control based on combining pesticidalcontrol of vectors with other control methods, andknown as integrated virus control, was superceded bycontrol obtained by any combination of procedures de-signed to prevent economically important crop losses,known as integrated virus management.
1975-2000 VIRAL GENOMES AND MOLECULAR GENETICS
During the last quarter of the 20th century greatprogress was made on several fronts, especially ingenome analysis and molecular genetics, facilitated byadditional powerful techniques (Table 1). Proceduressuch as enzyme-linked immunosorbent assay (ELISA),immunosorbent electron microscopy (ISEM) and thepolymerase chain reaction (PCR) revolutionized theroutine detection and assay of the better studied viruses.The use of combinations of the new techniques made itpossible to obtain much new information on the celland tissue biology of viruses and on viral mechanisms ofmany kinds. The wealth of approaches possible, togeth-er with the small size and accessibility of plant viralgenomes, encouraged scientists on the fringes of virolo-gy to use plant viruses in model systems, resulting in aperiod of increased funding and more rapid progressthan would have been made by virologists alone.
In 1980, the circular double-stranded DNA of CaMVbecame the first plant viral genome to be completely se-quenced (Frank et al., 1980). Next came the single-stranded linear RNA genome of TMV (Goelet et al.,1982). These sequences allowed the constituent genesand control elements, and the translation products, to
be defined. When infectious clones became available(Ahlquist and Janda, 1984), site-directed mutagenesiswas used to investigate gene functions (Ishikawa et al.,1986), and genomes were manipulated to turn virusesinto expression vectors (Brisson et al., 1984). Thegenomes of viruses in different genera turned out tocontain a variety of numbers and arrangements ofgenes, which typically, as in the TMV genome (Fig. 22),included genes encoding the coat and replication pro-teins. However, several lines of evidence (Deom et al.,1987; Meshi et al., 1987; Tomenius et al., 1987; Fig. 21)showed that one or more additional virus-coded pro-teins were required for cell-to-cell movement of virusthrough modified plasmodesmata (Wolf et al., 1989;Oparka et al., 1999), and that these so-called ‘move-ment proteins’ of different viruses work in a variety ofways. In addition, a massive amount of work was doneon mechanisms of virus replication: on the replicationof single- and double-stranded viral RNA and DNA, onthe translation of viral open reading frames and process-ing of the resulting products, on the roles of virus-cod-ed and host enzymes in these events, and on the com-partmentalization of events in cells. These and relatedadvances would warrant a separate review, and will notbe described in detail here. However, five other mainlines of progress seem to merit special mention.
First, the action of dominant plant genes conferringhypersensitive resistance was found to be triggered bydifferent viral proteins (Saito et al., 1987), each specificfor a cognate host gene, and defined modifications inthese viral proteins were implicated in resistance-break-ing. Intriguing evidence began to emerge on the mecha-nisms of action of plant genes conferring hypersensitivi-ty or extreme resistance (Kohm et al., 1993). Several ofthese plant genes were cloned (Whitham et al., 1994),sequenced and found to encode protein domains similarto those being discovered in comparable work on genescontrolling resistance to bacteria, fungi or nematodes.Fig. 23 compares some of these pathogen resistancegenes, all of which encode a leucine-rich repeat domainthat is involved in specificity of action. Several recessivevirus-resistance genes proved to encode translation initi-ation factors (Lellis et al., 2002).
Secondly, viral coat protein genes (Powell Abel et al.,1986) and other virus-derived sequences were shown toconfer virus resistance when transferred to plant nuclearDNA (Fig. 24), and the mechanisms of action of thesetransgenic resistances explored (Nelson et al., 1987).Wang et al. (2000) devised an improved design of trans-genic sequences for conferring strong resistance. Somecrop plants with transgenic virus resistance have beencommercialized successfully (Gonsalves, 1998).
Thirdly, gene silencing was discovered (Lindbo et al.,1993) and its roles in regulating virus replication, and incross-protection between virus strains revealed (Coveyet al., 1997; Ratcliff et al., 1997). Short double-stranded
Journal of Plant Pathology (2009), 91 (3), 509-520 B.D. Harrison 515
001_JPPReview(Harrison)_509 23-11-2009 17:23 Pagina 515
516 Plant virology in the 20th century Journal of Plant Pathology (2009), 91 (3), 509-520
Figg. 20-26. 20. Actual and predicted (calculated) incidences of sugar-beet yellows disease in England in August in successiveyears (after Watson, 1966). 21. Electron micrograph of an ultra-thin section of a cell wall (CW) of a tobacco leaf showing the loca-tion of TMV movement protein (detected by gold-labelled antibody) in plasmodesmata (arrowed) (after Tomenius et al., 1987).22. Organization of the TMV genome, showing the open reading frames and the proteins expressed from full-length and sub-ge-nomic (sg) RNA species (after Lewandowski, 2000). 23. Domains of dominant plant genes that confer resistance to the fungus Cla-dosporium fulvum (Cf-9), TMV (N) and the bacterium Pseudomonas syringae (RPS2). The largest boxed portion of each genespecifies a leucine-rich repeat domain (courtesy D.C. Baulcombe). 24. Resistance in transgenic tobacco plants expressing TMVcoat protein. Transgenic plants are not infected whereas control plants show severe mosaic (courtesy R.N. Beachy). 25. Accumula-tion of 25-nucleotide, Potato virus X-specific, anti-sense RNA molecules in inoculated and systemically infected leaves at succes-sive days after inoculation (d.p.i.) (courtesy D.C. Baulcombe). 26. Sequence relationships among the DNA genomes of viruses inthree genera of the family Geminiviridae (Rybicki et al., 2000).
001_JPPReview(Harrison)_509 23-11-2009 17:23 Pagina 516

Journal of Plant Pathology (2009), 91 (3), 509-520 B.D. Harrison 517
virus-derived RNA molecules play a critical role(Hamilton and Baulcombe, 1999; Fig. 25). Viralgenomes were found typically to encode a suppressor ofgene silencing (Pruss et al., 1997), which is also involvedin synergy between unrelated viruses and can act in avariety of ways. When viruses with their genomes modi-fied to contain host nucleic acid sequences were used toinfect plants, the cognate plant genes could be silenced.
Fourthly, the wealth of viral genome sequences avail-able enabled the frequency of mutation and pseudo-re-combination to be assessed, true interspecific recombi-nation to be proved (Zhou et al., 1997) and phylogenet-ic relationships to be inferred within and between viralspecies, genera and, in some instances, families. Fig. 26represents the relationships among the genomes ofviruses assigned to three genera within the family Gemi-niviridae.
Fifthly, excellent progress was made in understand-ing vector-transmission mechanisms, not only those in-volved in non-persistent transmission of potyviruses(Berger and Pirone, 1986; Atreya et al., 1990), and per-sistent circulative transmission of luteoviruses (Gildowand Rochow, 1980; van den Heuvel et al., 1994; Braultet al., 1995), both involving aphids, but also those thatuse vectors of many other kinds.
SOME GENERAL CONCLUSIONS
The general conclusions I draw from this brief surveyof a large and complicated subject are as follows.A. Research progress has been dependent on, or greatly
facilitated by, the development of new techniques.B. Plant virus genomes, multiplication mechanisms, and
viral adaptations to plants and to survival in fieldconditions, represent series of variations on basicthemes.
C. Numerous discoveries in plant virology have led vi-rology in general: for example in virus particle com-position and structure, in the infectivity of viral nu-cleic acid, in single gene and transgenic resistancemechanisms, in mechanisms of vector transmissionthat do not involve replication in the vector, in the ef-fects and uses of gene silencing, and in the spatial as-pects of virus epidemiology.
D.Virus research has changed agricultural practice: forexample through schemes for propagating virus-freeplanting stocks, quarantine regulations, use of vector-controlling chemicals, breeding virus-resistant culti-vars and rapid diagnostic tests. Agricultural practicehas moved on from integrated virus control to inte-grated virus management. These changes have in-creased crop yields and food security, dramatically insome instances.
E. The focus of research has proceeded from viral struc-tures to functions, then to mechanisms and next to
manipulating such mechanisms. Plant viruses havebecome tools in molecular biology, cell biology andbiotechnology.
F. Plant virology has become a most ecumenical sci-ence, drawing on expertise from many disciplinesand sub-disciplines, and it continues to pose manyintellectual and practical challenges. These seem like-ly to increase in number and urgency as the worldpopulation grows, the climate changes, energysources become more expensive, food crops are usedfor non-food purposes or are replaced by non-foodcrops, and the need to safeguard the environment be-comes more compelling.
ACKNOWLEDGEMENTS
I am most grateful to Graham Cowan and to thosementioned in the figure legends for help with the illus-trations.
REFERENCES
Ahlquist P., Janda M., 1984. cDNA cloning and in vitro tran-scription of the complete brome mosaic virus genome.Molecular and Cellular Biology 4: 2876-2882.
Amos J., Hatton R.G., Knight R.C., Massee A.M., 1927. Ex-periments in the transmission of “Reversion” in black cur-rants. Annual Report of East Malling Research Station for1925: 126-150.
Atreya C.D., Raccah B., Pirone T.P., 1990. A point mutationin the coat protein abolishes aphid transmissibility of a po-tyvirus. Virology 178: 161-165.
Bawden F.C., Pirie N.W., 1938. Crystalline preparations oftomato bushy stunt virus. British Journal of ExperimentalPathology 19: 251-263.
Bawden F.C., Pirie N.W., Bernal J.D., Fankuchen I., 1936.Liquid crystalline substances from virus-infected plants.Nature 138: 1051-1052.
Behrens J., 1899. Weiter Beitrage zur Kenntnis derTabakpflanze. XIV. Die Mauche (Mauk) des Tabaks. Land-wirtschaflichen Versuchsstationen 52: 442-447.
Bennett C.W., 1937. Correlation between movement of thecurly top virus and translocation of food in tobacco andsugar beet. Journal of Agricultural Research 54: 479-502.
Bennett C.W., Wallace H.E., 1938. Relation of the curly topvirus to the vector, Eutettix tenellus. Journal of AgriculturalResearch 56: 31-51.
Berger P.H., Pirone T.P., 1986. The effect of helper compo-nent on the uptake and localization of potyviruses inMyzus persicae. Virology 153: 256-261.
Black L.M., Brakke M.K., 1952. Multiplication of wound-tu-mor virus in an insect vector. Phytopathology 42: 269-273.
Box H.E., 1945. Insect transmission of the ‘swollen-shoot’virus in West African cacao. Nature 155: 608-609.
Brault V., van den Heuvel J.F.J.M., Verbeek M., Ziegler-GraffV., Reutenauer A., Herrbach H., Garaud J-C., Guilley H.,
001_JPPReview(Harrison)_509 23-11-2009 17:23 Pagina 517

Richards K., Jonard G., 1995. Aphid transmission of beetwestern yellows luteovirus requires the minor capsid read-through protein P74. The EMBO Journal 14: 650-659.
Brisson N., Paszkowski J., Penswick J.R., Gronenborn B.,Potrykus I., Hohn T., 1984. Expression of a bacterial genein plants by using a viral vector. Nature 310: 511-514.
Canova A., 1966. Ricerche sulla malattie da virus delle Grami-nacee. III. Polymyxa graminis Led. vettore del virus delmosaico del Frumento. Phytopathologia Mediterranea 5:53-58.
Caspar D.L.D., 1956. Radial density distribution in the tobac-co mosaic virus particle. Nature 177: 928.
Covey S.N., Al-Kaff N.S., Langara A., Turner D.S., 1997.Plants combat infection by gene silencing. Nature 385:781-782.
Deom C.M., Oliver M.J., Beachy R.N., 1987. The 30-kilodal-ton gene product of tobacco mosaic virus potentiates virusmovement. Science 237: 389-394.
Diener T.O., 1971. Potato spindle tuber “virus”. IV. A repli-cating, low molecular weight RNA. Virology 45: 411-428.
Doolittle S.P., 1916. A new infectious mosaic disease of cu-cumber. Phytopathology 6: 145-147.
Estes A.P., Brakke M.K., 1966. Correlation of Polymyxagraminis with transmission of soil-borne wheat mosaicvirus. Virology 28: 772-774.
Fraenkel-Conrat H., Williams R.C., 1955. Reconstitution ofactive tobacco mosaic virus from its inactive protein andnucleic acid components. Proceedings of the National Acad-emy of Sciences USA 41: 690-698.
Frank A., Guilley H., Jonard G., Richards K., Hirth L., 1980.Nucleotide sequence of cauliflower mosaic virus DNA.Cell 21: 285-294.
Franklin R.E., 1956. Location of the ribonucleic acid in thetobacco mosaic virus particle. Nature 177: 928-930.
Franklin R.E., Holmes K.C., 1958. Tobacco mosaic virus: ap-plication of the method of isomorphous replacement to thedetermination of the helical parameters and radial densitydistribution. Acta Crystallographica 11: 213-220.
Fukushi T., 1940. Further studies on the dwarf disease of riceplant. Journal of the Faculty of Agriculture of Hokkaido Im-perial University 45: 83-154.
Gibbs A.J., Harrison B.D., 1970. Cucumber mosaic virus.CMI/AAB Descriptions of Plant Viruses No. 1
Gierer A., Mundry K.W., 1958. Production of mutants of to-bacco mosaic virus by chemical alteration of its ribonucleicacid in vitro. Nature 182: 1457-1458.
Gierer A., Schramm G., 1956. Infectivity of ribonucleic acidfrom tobacco mosaic virus. Nature 177: 702-703.
Gildow F.E., Rochow W.F., 1980. Role of accessory salivaryglands in aphid transmission of barley yellow dwarf virus.Virology 104: 97-108.
Goelet P., Lomonossoff G.P., Butler P.J.G., Akam M.E., GaitM.J., Karn J., 1982. Nucleotide sequence of tobacco mosa-ic virus RNA. Proceedings of the National Academy of Sci-ences USA 79: 5818-5822.
Gonsalves D., 1998. Control of papaya ringspot virus in pa-paya: a case study. Annual Review of Phytopathology 36:415-437.
Govier D.A., Kassanis B., 1974. A virus-induced componentof plant sap needed when plants acquire potato virus Yfrom purified preparations. Virology 61: 420-426.
Gregory P.H., 1948. The multiple-infection transformation.Annals of Applied Biology 35: 412-417.
Gregory P.H., Read D.R., 1949. The spatial distribution of in-sect-borne plant-virus diseases. Annals of Applied Biology36: 475-482.
Hamilton A.J., Baulcombe D.C., 1999. A species of small anti-sense RNA in posttranscriptional gene silencing in plants.Science 286: 950-952.
Harrison B.D., 1977. Ecology and control of viruses with soil-inhabiting vectors. Annual Review of Phytopathology 15:331-360.
Harrison B.D., Finch J.T., Gibbs A.J., Hollings M., ShepherdR.J., Valenta V., Wetter C., 1971. Sixteen groups of plantviruses. Virology 45: 356-363.
Harrison B.D., Nixon H.L., 1959. Separation and propertiesof particles of tobacco rattle virus with different lengths.Journal of General Microbiology 21: 569-581.
Harrison B.D., Peachey J.E., Winslow R.D., 1963. The use ofnematicides to control the spread of arabis mosaic virus byXiphinema diversicaudatum (Micol.). Annals of Applied Bi-ology 52: 243-255.
Hewitt W.B., Raski D.J., Goheen A.C., 1958. Nematode vec-tor of soil-borne fanleaf virus of grapevines. Phytopatholo-gy 48: 586-595.
Holmes F.O., 1929. Local lesions in tobacco mosaic. BotanicalGazette 87: 39-55.
Holmes F.O., 1934. Inheritance of ability to localize tobaccomosaic virus. Phytopathology 24: 984-1002.
Hull R., Heathcote G.D., 1967. Experiments on the time ofapplication of insecticides to decrease the spread of yel-lowing viruses of sugar beet. Annals of Applied Biology 60:469-478.
Ishikawa M., Meshi T., Motoyoshi F., Takamatsu N., OkadaY., 1986. In vitro mutagenesis of the putative replicasegenes of tobacco mosaic virus. Nucleic Acids Research 14:8291-8305.
Ivanovski D., 1903. Ueber die Mosaikkrankheit der Tabak-spflanze. Zeitschrift fur Pflanzenkrankheiten 13: 1-41.
Jagger I.C., 1916. Experiments with the cucumber mosaic dis-ease. Phytopathology 6: 148-151.
Kassanis B., 1950. Heat inactivation of leaf-roll virus in potatotubers. Annals of Applied Biology 37: 339-341.
Kassanis B., Nixon H.L., 1961. Activation of one tobacconecrosis virus by another. Journal of General Microbiology25: 459-471.
Kausche G.A., Pfankuch E., Ruska H., 1939. Die Sichtbarma-chung vom pflanzlichem Viren im Ubermikroskop. Natur-wissenschaften 27: 292-299.
Kirkpatrick T.W., 1931. Further studies on leaf-curl of cottonin the Sudan. Bulletin of Entomological Research 22: 323-363.
Kohm B.A., Goulden M.G., Gilbert J.E., Kavanagh T.A.,Baulcombe D.C., 1993. A potato virus X resistance genemediates an induced, nonspecific resistance in protoplasts.Plant Cell 5: 913-920.
518 Plant virology in the 20th century Journal of Plant Pathology (2009), 91 (3), 509-520
001_JPPReview(Harrison)_509 23-11-2009 17:23 Pagina 518

Knight C.A., 1947. The nature of some of the chemical differ-ences among strains of tobacco mosaic virus. Journal of Bi-ological Chemistry 171: 297-308.
Lellis A.D., Kasschau K.D., Whitham S.A., Carrington J.C.,2002. Loss-of-susceptibility mutants of Arabidopsisthaliana reveal an essential role for eIF(iso)4E during po-tyvirus infection. Current Biology 12: 1046-1051.
Lewandowski D.J., 2000. Genus Tobamovirus. In: van Regen-mortel M.H.V., Fauquet C.M., Bishop D.H.L., CarstensE.B., Estes M.K., Lemon S.M., Mayo M.A., McGeoch D.J.,Pringle C.R., Wickner R.B. (eds). Virus Taxonomy: SeventhReport of the International Committee on Taxonomy ofViruses, pp. 889-894. Academic Press, San Diego, CA, USA.
Lindbo J.A., Silva-Rosales L., Proebsting W.M., DoughertyW.G., 1993. Induction of a highly specific antiviral state intransgenic plants: implications for regulation of gene ex-pression and virus resistance. Plant Cell 5: 1749-1759.
Lister R.M., 1966. Possible relationships of virus-specificproducts of tobacco rattle virus infections. Virology 28:350-353.
Luria S.E., 1953. General Virology. John Wiley and Sons,New York, NY, USA.
McKinney H.H., 1929. Mosaic diseases in the Canary Islands,West Africa and Gibraltar. Journal of Agricultural Research39: 557-578.
Megroz R.L. (ed.), 1947. The Lear Omnibus, p.70. Hutchin-son, London, UK.
Meshi T., Watanabe Y., Saito T., Sugimoto A., Maeda T., Oka-da Y., 1987. Function of the 30kd protein of tobacco mo-saic virus: involvement in cell-to-cell movement and dis-pensability for replication. The EMBO Journal 6: 2557-2563.
Morel G., Martin C., 1952. Guerison de dahlias atteints d’unemaladie a virus. Compte Rendu Hebdomadaire des Séancesde l’Academie des Sciences Paris 235: 1324-1325.
Murant A.F., Mayo M.A., Harrison B.D., Goold R.A., 1973.Evidence for two functional RNA species and a ‘satellite’RNA in Tomato black ring virus. Journal of General Virolo-gy 19: 275-278.
Nelson R.S., Powell Abel P., Beachy R.N., 1987. Lesions andvirus accumulation in inoculated transgenic plants express-ing the coat protein gene of tobacco mosaic virus. Virology158: 126-132.
Nixon H.L., Harrison B.D., 1959. Electron microscopic evi-dence on the structure of the particles of tobacco rattlevirus. Journal of General Microbiology 21: 582-590.
Oparka K.J., Roberts A. G., Boevink P., Santa Cruz S.,Roberts I., Pradel K. S., Imlau A., Kotlizky G., Sauer N.,Epel B., 1999. Simple, but not branched, plasmodesmataallow the nonspecific trafficking of proteins in developingtobacco leaves. Cell 97: 743-754.
Pittman H.A., 1927. Spotted wilt in tomatoes. Journal of theCouncil for Scientific and Industrial Research (Australia) 1:74-77.
Powell Abel P., Nelson R.S., De B., Hoffman N., Rogers S.G.,Fraley R.T., Beachy R.N., 1986. Delay of disease develop-ment in transgenic plants that express the tobacco mosaicvirus coat protein gene. Science 232: 738-743.
Pruss G., Ge X., Shi X.M., Carrington J.C., Bowman VanceV., 1997. Plant viral synergism: the potyviral genome en-codes a broad-range pathogenicity enhancer that transacti-vates replication of heterologous viruses. Plant Cell 9: 859-868.
Purdy H.A., 1928. Immunologic reactions with tobacco mosa-ic virus. Proceedings of the Society for Experimental Biologyand Medicine 25: 702-703.
Ratcliff F., Harrison B.D., Baulcombe D.C., 1997. A similari-ty between viral defense and gene silencing in plants. Sci-ence 276: 1558-1560.
Reddick E., Stewart V.B., 1919. Transmission of the virus ofbean mosaic in seed and observations on the thermal-deathpoint of seed and virus. Phytopathology 9: 445-450.
Rybicki E.P., Briddon R.W., Brown J.K., Fauquet C.M.,Maxwell D.P., Harrison B.D., Markham P.G., Bisaro D.M.,Robinson D., Stanley J., 2000. Family Geminiviridae. In:van Regenmortel M.H.V., Fauquet C.M., Bishop D.H.L.,Carstens E.B., Estes M.K., Lemon S.M., Mayo M.A., Mc-Geoch D.J., Pringle C.R., Wickner R.B. (eds.). Virus Tax-onomy: Seventh Report of the International Committee onTaxonomy of Viruses, pp. 285-297. Academic Press, SanDiego, CA, USA.
Saito T., Meshi T., Takamatsu N., Okada Y., 1987. Coat pro-tein gene sequence of tobacco mosaic virus encodes a hostresponse determinant. Proceedings of the National Acade-my of Sciences USA 84: 6074-6077.
Samuel G., 1934. The movement of tobacco mosaic viruswithin the plant. Annals of Applied Biology 21: 90-111.
Sanger H.L., 1968. Characteristics of tobacco rattle virus. IEvidence that its two particles are functionally defectiveand mutually complementing. Molecular and General Ge-netics 101: 346-367.
Schneider I.R., 1971. Characteristics of a satellite-like virus oftobacco ringspot virus. Virology 45: 108-122.
Shepherd R.J., Wakeman R.J., Romanko R.R., 1968. DNA incauliflower mosaic virus. Virology 36: 150-152.
Shikata E., Maramorosch K., 1965. Electron microscopic evi-dence for the systemic invasion of an insect host by a plantpathogenic virus. Virology 27: 461-475.
Sinha R.C., 1965. Sequential infection and distribution ofwound-tumor virus in the internal organs of a vector afteringestion of virus. Virology 26: 673-686.
Smith C.E., 1924. Transmission of cowpea mosaic by the beanleaf beetle. Science 60: 268.
Stanley W.M., 1935. Isolation of a crystalline protein possess-ing the properties of tobacco mosaic virus. Science 81: 644-645.
Storey H.H., 1932. The inheritance by an insect vector of theability to transmit a plant virus. Proceedings of the RoyalSociety of London B 112: 46-60.
Storey H.H., 1933. Investigations of the mechanism of thetransmission of plant viruses by insect vectors. I. Proceed-ings of the Royal Society of London B 113: 463-485.
Takahashi W.N., Rawlins T.E., 1932. Method for determiningshape of colloidal particles: application in study of tobaccomosaic virus. Proceedings of the Society for ExperimentalBiology and Medicine 30: 155-157.
Journal of Plant Pathology (2009), 91 (3), 509-520 B.D. Harrison 519
001_JPPReview(Harrison)_509 23-11-2009 17:23 Pagina 519

Takami N., 1901. On dwarf disease of rice plant and“tsumaguro-yokabai”. Journal of the Japanese AgriculturalSociety 241: 22-30.
Takebe I., Otsuki Y., 1969. Infection of tobacco mesophyllprotoplasts by tobacco mosaic virus. Proceedings of the Na-tional Academy of Sciences USA 64: 843-848.
Taylor C.E., Robertson W.M., 1970. Location of tobacco rat-tle virus in the nematode vector Trichodorus pachydermusSeinhorst. Journal of General Virology 6: 179-182.
Teakle D.S., 1960. Association of Olpidium brassicae and to-bacco necrosis virus. Nature 188: 431-432.
Temmink J.H.M., Campbell R.N., Smith P.R., 1970. Specifici-ty and site of in vitro acquisition of tobacco necrosis virusby zoospores of Olpidium brassicae. Journal of General Vi-rology 9: 201-213.
Thung T.H., 1931. Smetstof en plantencel bij enkelevirusziekten van de tabaksplant. Handelingen van het 6deNederlandsch-Indisch Natuurwetenschappelijk Congres,Bandoeng 1931, 450-453.
Tomenius K., Clapham D., Meshi T., 1987. Localization byimmunogold cytochemistry of the virus-coded 30K proteinin plasmodesmata of leaves infected with tobacco mosaicvirus. Virology 160: 363-371.
Tsugita A., 1962. The proteins of mutants of TMV: composi-tion and structure of chemically evoked mutants of TMV-RNA. Journal of Molecular Biology 5: 284-292.
Van den Heuvel J.F.J.M., Verbeek M., van der Wilk F., 1994.Endosymbiotic bacteria associated with circulative trans-mission of potato leafroll virus by Myzus persicae. Journalof General Virology 75: 2559-2565.
Van der Plank J.E., 1946. A method for estimating the num-ber of random groups of adjacent diseased plants in a ho-mogeneous field. Transactions of the Royal Society of SouthAfrica 31: 269-278.
Wang M-B., Abbott D.C., Waterhouse P.M., 2000. A singlecopy of a virus-derived transgene encoding hairpin RNAgives immunity to barley yellow dwarf virus. MolecularPlant Pathology 1: 347-356.
Watson J.D., 1954. The structure of Tobacco mosaic virus. I.X-ray evidence of a helical arrangement of sub-unitsaround the longitudinal axis. Biochimica et Biophysica Acta13: 10-19.
Watson M.A., 1936. Factors affecting the amount of infectionobtained by aphis transmission of the virus Hy. III. Philo-sophical Transactions of the Royal Society of London B 226:457-489.
Watson M.A., 1940. Studies on the transmission of sugar-beetyellows virus by the aphis, Myzus persicae (Sulz.). Proceed-ings of the Royal Society of London B 128: 535-552.
Watson M.A., 1966. The relation of annual incidence of beetyellowing viruses in sugar beet to variations in weather.Plant Pathology 15: 145-149.
Whitham S., Dinesh-Kumar S.P., Choi D., Hehl R., Corr C.,Baker B., 1994. The product of tobacco mosaic virus re-sistance gene N: similarity to Toll and the interleukin-1 re-ceptor. Cell 78: 1101-1115.
Wittmann H.G., 1962. Proteinundersuchungen an Mutantendes Tabakmosaikvirus als Beitrag zum Problem desgenetischen Codes. Zeitschrift für Vererbungslehre 93:491-530.
Wolf S., Deom C.M., Beachy R.N., Lucas W.J., 1989. Move-ment protein of tobacco mosaic virus modifies plasmodes-matal size exclusion limit. Science 246: 377-379.
Zhou X., Liu Y. L., Calvert L., Munoz C., Otim-Nape G.W.,Robinson D.J., Harrison B.D., 1997. Evidence that DNA-A of a geminivirus associated with severe cassava mosaicdisease in Uganda has arisen by interspecific recombina-tion. Journal of General Virology 78: 2101-2111.
520 Plant virology in the 20th century Journal of Plant Pathology (2009), 91 (3), 509-520
001_JPPReview(Harrison)_509 23-11-2009 17:23 Pagina 520