Embed Size (px)
Citation preview

Mutation Research, 226 (1989) 69-73 69 Elsevier
MTRL 0204
Hyperthermia-induced modulation of killing and mutation by UV and N-methyl-N'-nitro-N-nitrosoguanidine in V79 cells
S u b r a t a B a n e r j e e , G a y a r a m B h a u m i k a n d S u k h e n d u B. B h a t t a c h a r j e e
Crystallography and Molecular Biology Division, Saha Institute o f Nuclear Physics, Calcutta - 700 064 (India)
(Accepted 18 January 1989)
Keywords: Hyperthermia; Cell killing; UV; MNNG
Summary
Hyperthermic exposures of V79 cells did not affect the killing by UV light, whereas it enhanced MNNG- induced killing. Such hyperthermic exposure increased the mutation induction (resistance to 6-thioguanine) by both UV and MNNG. The timing of heat exposure, before or after the treatments, had no effect on the result in cases of cytotoxicity and mutagenesis.
Even though a considerable amount of research is being done to elucidate the mechanism or pro- cesses governing cellular responses to elevated temperatures in the range of 40-46°C (Hahn, 1982), not much is known about the genetic effect of heat on mammalian cells, particularly when used in combination with various chemicals and radiation. Since initiation for carcinogenesis most probably occurs through somatic mutation (Ramel et al., 1984), the clinical potential of heat dictates that we understand the mutagenic response of cells in relation to hyperthermia. Distinct enzymatic systems are involved in the repair of damage due to ionizing, ultraviolet and alkylating agents (Fried-
Correspondence: Dr. S.B. Bhattacharjee, Professor and Head, Crystallography and Molecular Biology Division, Saha Institute of Nuclear Physics, Sector 1, Block 'AF', Bidhan Nagar, Calcutta - 700 064 (India).
berg, 1985) and comparison between the interac- tion of heat and these agents may provide some in- sight into the mechanisms of heat-induced effects on mammalian cells.
Synthesis of DNA, RNA and proteins is known to be affected by hyperthermia (McCormick and Penman, 1969; Mondovi et al., 1969). Hyperther- mia can cause sister-chromatid exchanges (Liv- ingston and Dethlefsen, 1979) and chromosomal aberrations in mammalian cells (Dewey at al., 1978). Temporary inhibition of DNA polymerase activities in mammalian cells has also been reported due to heat exposures (Spiro et al., 1984;
D i c k o m e y et al., 1987). These heat-induced changes in the genetic material and cellular capaci- ty of repair (Corry et al., 1977; Mills and Meyn, 1983) may then affect the extent and fidelity of DNA repair. In this communication, we have in- vestigated the influence of hyperthermia on muta- tion induction by ultraviolet light (UV) and an
0165-7992/89/$ 03.50 © 1989 Elsevier Science Publishers B.V. (Biomedical Division)
70
alkylating agent, N-methyl-N'-nitro-N-nitroso- guanidine (MNNG). Our results indicate that hyperthermia (45°C, 10 min) can potentiate the lethal effect of MNNG but not that of UV. However, it distinctly modulates the mutagenic effect of both agents.
Materials and methods
Chemicals MNNG, neomycin sulfate and 6-thioguanine
(TG) were purchased from Sigma Chemicals and penicillin and streptomycin sulfate from local pharmaceutical sources. A stock solution of MNNG was prepared immediately before use and diluted to the desired concentration. A stock solu- tion of TG was made in dimethyl sulfoxide (DMSO). Trypsin (1:250) was purchased from Dif- co laboratories.
Cell culture Details about the culture condition, media etc.
have already been published (Bhattacharjee and Pal, 1982; Bhattacharjee et al., 1982). In brief, CH-V79 (male, lung fibroblasts) cells were grown as monolayer in Corning glass Petri dishes or disposable plastic Petri dishes in modified Eagle's medium enriched with all non-essential amino acids and vitamins but without hypoxanthine and thymidine and supplemented with 10°70 dialyzed goat serum, penicillin, streptomycin and neo- mycin.
Heat shock and UV irradiation Heat treatment was carried out in a water bath
fitted with a circulating pump and precision- controlled thermostat which could regulate the temperature within +_0.01°C. Asynchronously growing monolayer cells in 55-mm Petri dishes were placed in a perforated rack and immersed in water preheated to the desired temperature (45.0°C). The temperature of the medium reached the desired value within 4 min; the zero time of treatment started immediately after 4 rain. To avoid pH shock, fresh medium was always used for heat treatment.
A single 15-W germicidal lamp with output pre- dominantly at 254 nm was used for UV exposures. The incident dose rate was 2.00 j /m2/s as deter- mined by chemical actinometry (Hatchard and Parker, 1956).
Determination o f cellular survival and mutant frequencies
Details of the techniques used in survival and mutational assay have been published (Ghosh and Bhattacharjee, 1988). Briefly, after different heat and UV or MNNG treatments (1 h) exponentially growing cells were washed with phosphate- buffered saline (PBS), trypsinized and counted in a hemocytometer, diluted appropriately to yield 50-250 colonies per dish, and seeded in triplicate in 55-mm Petri dishes for each dilution. The cells were then incubated at 37°C in a humidified 5070 CO2 atmosphere for colony formation. After 7-9 days, colonies were stained with 0.1 °7o methylene blue and visible colonies were counted. The cloning efficiency of control cells was about 65°7o.
For mutational studies, the replating technique has been used. The endpoint selected was resistance to the drug TG. After different mutagenic treat- ments exponential cells were allowed to grow in
0 i t~.
0.1
7-
0.01 i I l I 5 10 15 20
UV FLUENCE (Jlm 2 )
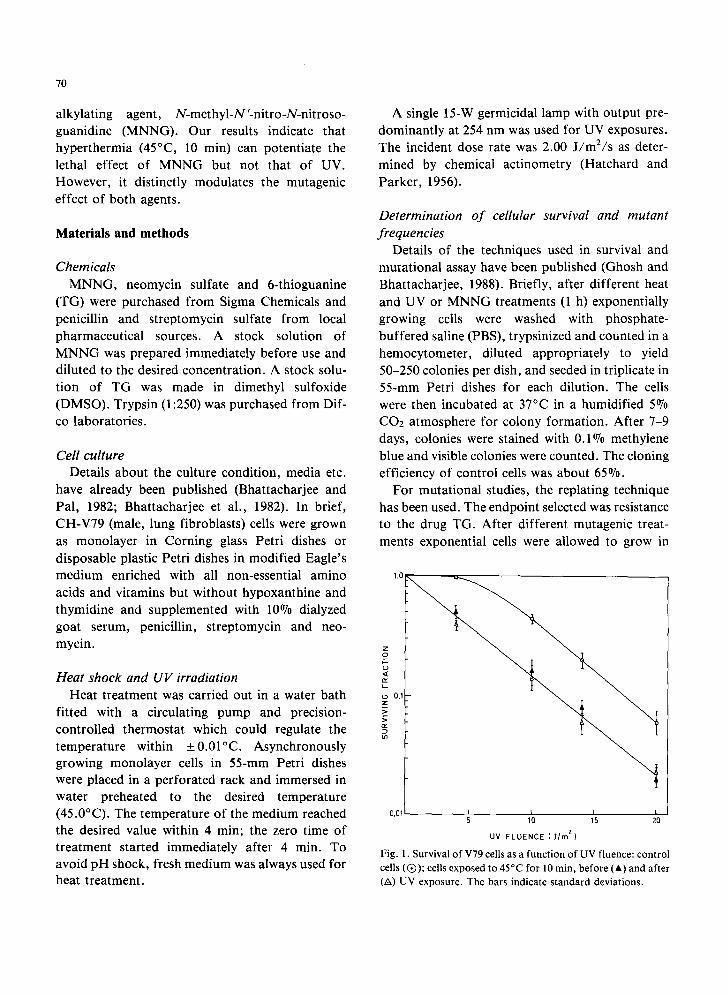
Fig. 1. Survival of V79 cells as a funct ion of UV fluence: control cells (Q) ; cells exposed to 45°C for lO rain, before (A) and after (A) UV exposure. The bars indicate standard deviations.
normal growth medium for 9-10 days for mutation expression with 3-4 subcultures during this period. Thereafter, the cells were diluted to 1 × 105 cells/100-mm Corning dish for growth in the presence of TG at a concentration of 3 ttg/ml in complete growth medium. For each dose, 5 plates were used. Resistant colonies were counted after 10-14 days; the cloning efficiency of the cells was also determined at the same time by plating 250 cells in triplicate in normal growth medium and counting the resultant colonies after 7-9 days. Ex- perimental data shown represent the average of 2-4 independent observations with the standard devia- tions.
Results
Survival of V79 cells as a function of UV fluence
1.0
0.5
z o
ta .
tt~
0.1 t f 0.t75 0.25 0.50 1.0
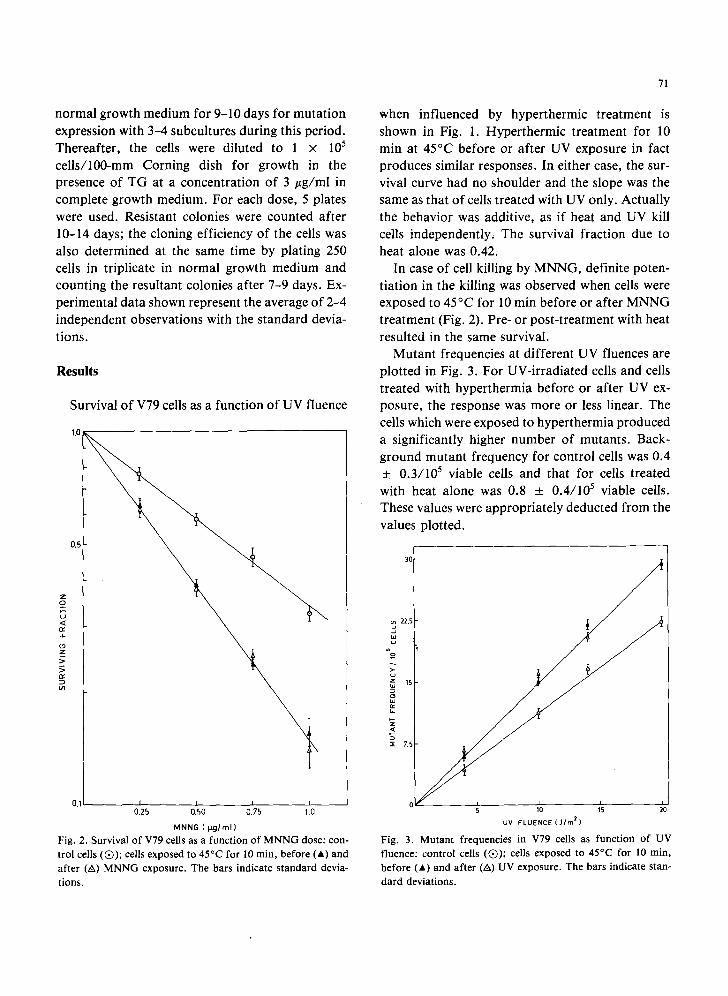
MNNG ( t ~ j / r n l } Fig. 2. Survival of V79 cells as a function o f M N N G dose: con- trol cells (Q) ; cells exposed to 45°C for l0 min, before (A) and after (A) MNNG exposure. The bars indicate s tandard devia- tions.
71
when influenced by hyperthermic treatment is shown in Fig. 1. Hyperthermic treatment for 10 min at 45°C before or after UV exposure in fact produces similar responses. In either case, the sur- vival curve had no shoulder and the slope was the same as that of cells treated with UV only. Actually the behavior was additive, as if heat and UV kill cells independently, The survival fraction due to heat alone was 0.42.
In case of cell killing by MNNG, definite poten- tiation in the killing was observed when cells were exposed to 45°C for l0 min before or after MNNG treatment (Fig. 2). Pre- or post-treatment with heat resulted in the same survival.
Mutant frequencies at different UV fluences are plotted in Fig. 3. For UV-irradiated cells and cells treated with hyperthermia before or after UV ex- posure, the response was more or less linear. The cells which were exposed to hyperthermia produced a significantly higher number of mutants. Back- ground mutant frequency for control cells was 0.4 _ 0.3/105 viable cells and that for cells treated with heat alone was 0.8 _+ 0.4/105 viable cells. These values were appropriately deducted from the values plotted.
~ 22: ~ ~
o >-
t a
z
5 =E 7.5
0 5 10 15 20
UV FLUENCE [J/rn 2)
Fig. 3. Mutan t frequencies in V79 cells as function of UV fluence: control cells (Q) ; cells exposed to 45°C for 10 min, before (A) and after (A) UV exposure. The bars indicate stan-
dard deviations.

72
9O
70 ..a
so
so
~ 3o
~ 2o
10
0 0.25 0.50 0.?5 10
MNNG (p,g/rnl)
Fig. 4. Mutant frequencies in V79 cells as function of MNNG dose: control cells (Q); cells exposed to 45°C for 10 min, before (A) and after (A) MNNG exposure. The bars indicate standard deviations.
Induction of mutation by MNNG as influenced by hyperthermia is shown in Fig, 4. Here also, hyperthermic treatment resulted in a very signifi- cant increase in mutation induction at each dose level of MNNG studied. For example, at 0.5/zg/ml MNNG concentration, control ceils gave 21 x 10 -s mutant frequency, whereas heat-exposed cells gave 40 x 10 -5 mutant frequency at the same dose of treatment. Timing of heat treatment either before or after MNNG exposure had little effect on the yield.
Discussion
Our experiments with Chinese hamster cells demonstrate that cellular killing can be enhanced by hyperthermia for the alkylating agent MNNG, but not for 254-nm UV light. Dewey (1984) has reported similar observations for alkylating agents and Mitchell et al. (1985) for UV light. The exact nature of the biochemical process which causes the observed differential survival response between hea t -MNNG and heat-UV is not clear. It is reason- able to expect that this difference could be related to the sensitivity of different repair enzymes to hyperthermia. For instance, DNA polymerases ot
and/3 have been shown to be affected by hyperther- mic treatment (Spiro et al., 1984). Since, however, both these enzymes are believed to be involved in repair of MNNG- and UV-induced damage (Fried- berg, 1985), thermal inhibition of these poly- merases may not be the reason for the observed dif- ferential sensitivity of the 2 agents to heat. Other enzymes whose variations could influence the slope and shoulder of the survival curves are methyl transferase, ligases, exonuclease, endonuclease, topoisomerase, etc. It has been shown that excision repair-defective xeroderma pigmentosum (XP) variant cells exhibited a non-shouldered survival response without any detectable increase in UV sensitivity relative to that of normal cells, as was observed here after hyperthermic treatment (Maher and McCormick, 1976). Similarly, a decreae in the slope of the alkylating damage-induced survival curve was observed in ceils with a reduced level of methyl transferase (Day et al., 1980). Recently, Warters and Brizgys (1988) have suggested that topoisomerase II may have an important function in thermal toxicity. However, presently no definite data are available which indicate whether heat- induced inhibition or inactivation of these enzymes could be an important rate-limiting agent in hyper- thermic cellular sensitization. Another possibility to be considered is the effect on cellular mem- branes. Modification of the cellular membrane by heat (Lepock, 1982) could also be a cause of the enhancement of cytotoxicity for some agents.
An interesting observation is the modulation of UV- and MNNG-induced mutation by hyperther- mia. The failure to find a difference in the nature of response in the mutation curve because of hyper- thermia suggests that the toxic and mutagenic ef- fects do not have a one-to-one correlation. In this connection it is worth while to consider the obser- vation made by McCready and Cook (1984) and Harless and Hewitt (1987) that repair replication in mammalian cells is a nuclear matrix-associated process. Warters and Stone (1983) have shown that heat-induced alteration in the nuclear matrix could inhibit DNA replication. Heat may thus alter the DNA repair enzymes themselves or cause an altera- tion on the repair substrate, i.e., the nucleoprotein

complex in chromatin and the nuclear matrix, such that the accessibility of repair enzymes is impeded. A reduced accessibility will probably affect fidelity of DNA repair and may account for the observed increase in mutant yield.
Acknowledgements
We thank Mr. Ashis Dutta, Mr. Utpal Basu and Mr. Radhey Mohan Tewary for their excellent technical assistance.
Some of these data and ideas were presented at the 12th Conference of Environmental Mutagen Society of India at Hisar (India).
References
Bhattacharjee, S.B., and B. Pal (1982) Tetracycline induced mutation in Chinese hamster cells, Mutation Res., 101, 329-338.
Bhattacharjee, S.B., S. Chatterjee and B. Pal (1982) Survival and mutation of Chinese hamster cells after ultraviolet ir- radiation and caffeine treatment, Mutation Res., 106, 137-146.
Corry, P.M., S. Robinson and S. Getz (1977) Hyperthermic ef- fect on DNA repair mechanisms, Radiology, 123, 475-482.
Day, R.S., Ill, C.H.J. Ziolkowski, D.A. Scudiero, S.A. Meyer and M.R. Mattern (1980) Human tumor cell strains defective in the repair of alkylation damage, Carcinogenesis, 1, 21-32.
Dewey, W.C. (1984) Interaction of heat with radiation and chemotherapy, Cancer Res., 44 (Suppl.), 47145-47205.
Dewey, W.C., S.A. Sapareto and D.A. Betten (1978) Hyper- thermic radiosensitization of synchronous Chinese hamster cells: relationship between lethality and chromosomal aberra- tions, Radiation Res., 76, 48-59.
Dikomey, E., W. Becket and K. Wielckens (1987) Reduction of DNA polymerase ~ activity of CHO cells by single and com- bined heat treatments, Int. J. Radiat. Biol., 52, 775-785.
Friedberg, E.C. (1985) DNA Repair, Freeman, New York. Ghosh, R., and S.B. Bhattacharjee (1988) Influence of nalidixic
acid on killing and mutation of V-79 cells exposed to different damaging agents, Mutation Res., 202, 71-75.
Hahn, G.M. (1982) Hyperthermia and Cancer, Plenum Press, New York.
73
Harless, J., and R.R. Hewitt (1987) Intranuclear localization of UV induced DNA repair in human VA 13 cells, Mutation Res., 183, 177-184.
Hatchard, C.G., and C.A. Parker (1956) A new sensitive chemical actinometer, II. Potassium ferrioxalate as a stan- dard chemical actinometer, Proc. R. Soc. A (London), 235, 518-536.
Kampinga, H.H., J.B.M. Torritsma and A.W.T. Konings (1985) Heat induced alterations in DNA polymerase activity of HeLa cells and of isolated nuclei. Relation to cell survival, Int. J. Radiat. Biol., 47, 29-40.
Lepock, J.R. (1982) Involvement of membranes in cellular responses to hyperthermia, Radiat. Res., 92, 433-438.
Livingston, G.K., and L.A. Dethlefsen (1979) Effects of hyper- thermia and X-irradiation on sister chromatid exchange (SCE) frequency in Chinese hamster ovary (CHO) cells, Radiat. Res., 77, 512-520.
Maher, V.M., and J.J. McCormick (1976) Effect of DNA repair on the cytotoxicity and mutagenicity of UV irradiation and of chemical carcinogens in normal and xeroderma pigmentosum cells, in: J.M. Yuhas, R.W. Tennant and J.D. Regan (Eds.), Biology of Radiation Carcinogenesis, Raven Press, New York, p. 129.
McCormick, W., and S. Penman (1969) Regulation of protein synthesis in HeLa cells: translation of elevated temperatures, J. Mol. Biol., 39, 315-333.
McCready, S.J., and P.R. Cook (1984) Lesions induced in DNA by ultraviolet light are repaired at the nuclear cage, J. Cell. Sci., 46, 365-386.
Mills, M.D., and R.E. Meyn (1983) Hyperthermic potentiation of unrejoined DNA strand breaks, Radiat. Res., 95,327-338.
Mitchell, R.E.J., B.P. Smith, N. Wheatly, A. Chan, S. Child and M.C. Paterson (1985) Sensitivity of hyperthermia-treated human cells to killing by ultraviolet or 3" radiation, Radiat. Res., 104, 234-241.
Mondovi, B., A. Finazzi Agro, G. Totillo, R. Stron, G. Morrica and A. Rossi Fanell (1969) The biochemical mechanism of selective heat sensitivity of cancer cells. II. Studies on nucleic acid and protein synthesis, Eur. J. Cancer, 5, 137-146.
Warters, R.L., and O.L. Stone (1983) Effects of hyperthermia on DNA replication in HeLa cells, Radiat. Res., 93, 71-84.
Warters, R.L., and L.M. Brizgys (1988) Effect of topo- isomerase II inhibitors on hyperthermic cytotoxicity, Cancer Res., 48, 3932-3928.
Communicated by F.H. Sobels
![farvardMenSupportNDE40 ath IFC BindG radStudenntlReceivees ...tech.mit.edu/V79/PDF/V79-N22.pdf · Bill Hecht, Ira Jaffe, Dan i Paul Robertson, Phil Robir ... Jire is p)] esently engaged](https://img.pdfslide.us/doc/110x75/5bc84da009d3f267298d3214/farvardmensupportnde40-ath-ifc-bindg-radstudenntlreceivees-techmiteduv79pdfv79-n22pdf.jpg)












![SUMMARY - NICNAS Web viewRCC (2008e) In vitro Chromosome Aberration Test in Chinese Hamster V79 Cells with [Notified polymer] (Study No. 1163702, July, 2008). Rossdorf, Germany,](https://img.pdfslide.us/doc/110x75/5a706c1f7f8b9ac0538bfbde/summary-nicnas-nbspdoc-fileweb-viewrcc-2008e.jpg)















