Embed Size (px)
Citation preview

Hindawi Publishing CorporationOxidative Medicine and Cellular LongevityVolume 2013 Article ID 678473 10 pageshttpdxdoiorg1011552013678473
Research ArticleLack of HXK2 Induces Localization ofActive Ras in Mitochondria and Triggers Apoptosis inthe Yeast Saccharomyces cerevisiae
Loredana Amigoni12 Enzo Martegani12 and Sonia Colombo12
1 Department of Biotechnology and Biosciences University of Milano-Bicocca Piazza Della Scienza 2 20126 Milan Italy2 SysBio Centre of Systems Biology Piazza Della Scienza 2 20126 Milan Italy
Correspondence should be addressed to Sonia Colombo soniacolombounimibit
Received 12 June 2013 Revised 18 July 2013 Accepted 24 July 2013
Academic Editor Paula Ludovico
Copyright copy 2013 Loredana Amigoni et al This is an open access article distributed under the Creative Commons AttributionLicense which permits unrestricted use distribution and reproduction in any medium provided the original work is properlycited
We recently showed that activated Ras proteins are localized to the plasmamembrane and in the nucleus in wild-type cells growingexponentially on glucose while in the hxk2Δ strain they accumulated mainly in mitochondria An aberrant accumulation ofactivated Ras in these organelles was previously reported and correlated to mitochondrial dysfunction accumulation of ROS andcell death Here we show that addition of acetic acid to wild-type cells results in a rapid recruitment of Ras-GTP from the nucleusand the plasma membrane to the mitochondria providing a further proof that Ras proteins might be involved in programmed celldeath Moreover we show that Hxk2 protects against apoptosis in S cerevisiae In particular cells lacking HXK2 and showing aconstitutive accumulation of activated Ras at the mitochondria are more sensitive to acetic-acid-induced programmed cell deathcompared to the wild type strain Indeed deletion of HXK2 causes an increase of apoptotic cells with several morphologicaland biochemical changes that are typical of apoptosis including DNA fragmentation externalization of phosphatidylserine andROS production Finally our results suggest that apoptosis induced by lack of Hxk2 may not require the activation of Yca1 themetacaspase homologue identified in yeast
1 Introduction
In Saccharomyces cerevisiae the highly homologous genesRAS1 and RAS2 encode small G-proteins that are activated bythe guanine nucleotide exchange factors (GEFs) Cdc25 andSdc25 [1 2] and inactivated by theGTPase-activating proteins(GAPs) Ira1 and Ira2 [3] GEFs and GAPs control the switchof the two small monomeric proteins between the activeGTP-bound and the inactive GDP-bound state The Ras pro-teins and theGPCR system [4ndash6] constitute twobranches thatmodulate the activity of adenylate cyclase (Cyr1) accordingto the glucose availability in the environment In turn Cyr1[7] activates cAMP-dependent protein kinase (PKA) throughcAMPThe amount of this secondmessenger is also regulatedat the level of degradation by the two phosphodiesterasesPde1 and Pde2 PKA plays a major role in the modulation ofmetabolism stress resistance cell growth proliferation mor-phogenesis and aging [8]
Recently our group expressed a probe consisting of aGFPfusion with a trimeric Ras Binding Domain of Raf1 (eGFP-RBD3) which binds Ras-GTP with a much higher affinitythan Ras-GDP to investigate the localization of active Ras inwild-type and in mutant strains in the cAMPPKA pathway[9] Our results showed that in W303-1A wild-type cells theprobe is localized essentially at the plasma membrane and inthe nucleus while in hxk2Δ cells the fluorescent signal accu-mulated in internal membranes and mitochondria [9] Thispeculiar localization of activated Ras2 was previously foundin S cerevisiae cells lackingWhi2p function a protein knownto influence cell cycle exit under conditions of nutritionalstress [10] The loss of Whi2p function led to accumulationof damaging ROS and cell death that displayed the hallmarksof apoptosis More recently it has been shown that also inmammalian cells translocation of activated K-RAS pro-tein to mitochondria caused mitochondrial dysfunction andincreased ROS generation [11]
2 Oxidative Medicine and Cellular Longevity
Apoptosis plays a crucial role in embryogenesis develop-ment tissue homeostasis and disease control inmulticellularorganisms In the last two decades the budding yeast S cere-visiae has become a useful model organism for studying thisprocess [12ndash15] The basic molecular machinery executingprogrammed cell death is phylogenetically conserved in yeastas well as animals Yeast orthologues of mammalian genesrelated to apoptosis coding for caspase (Yca1) the apoptosis-inducing factor (Aif1) the AIF-homologous mitochondrion-associated inducer of death (Ndi1) the serine protease OMI(Nma111) the endonuclease G (Nuc1) and the endo-exo-nuclease Tat-D (scTat-D) [12 15ndash20] have been characterizedThe apoptotic pathway in S cerevisiae can be activated by sev-eral mutations including cdc48-S565G [13] the inactivationof the UBP10 gene coding for a deubiquitinating enzyme [21]or by overexpression of the mammalian apoptotic cell deathregulator Bax [22] Moreover apoptotic cell death is alsoinduced by exogenous toxic agents such as hydrogen peroxide[23] formic acid [24] acetic acid [25] and others In par-ticular acetic-acid-induced apoptosis has been investigatedin detail and it has been shown that ROS accumulation andrelease of cytochrome c to the cytosol take place and thatH2O2is a trigger for acetic-acid-induced apoptosis [26ndash29]
In addition at least two death pathways can be activatedin yeast acetic-acid-induced apoptosis one is dependenton cyt c release which requires YCA1 and the other(s) isindependent of it [16 30] The yeast caspase Yca1 can protectyeast cells against multiple distinct forms of lethal insultssuch as exposure to metals (iron manganese cadmium) tolowdoses of valpronic acid and the previousmentioned aceticacid to toxins produced by virus killer toxins and others [31]On the other side in many instances Yca1 is not necessaryfor cell death For example external stimuli such as formicacid or copper or apoptosis derived from defective N-glyco-sylation in cells lackingOst2p the yeast homolog of themam-malian defender of apoptosis-1 are independent ofYCA1 [31]
In this work we provide data indicating that a correlationexists between programmed cell death and localization ofactive Ras proteins to mitochondria First of all we showthat addition of acetic acid to wild-type cells causes withinfive minutes a delocalization of the eGFP-RBD3 probe fromplasma membrane and nucleus to mitochondria Further-more we show that in hxk2Δ cells showing a constitutivelocalization of activeRas at themitochondria addition of ace-tic acid causes an increase of ROS accumulation mitochon-drial dysfunction and cell death compared with the wild-type strain It is known that hexokinase 2 functions as a glyco-lytic enzyme in the cytoplasm and as a regulator of gene tran-scription of several Mig1-regulated genes in the nucleus [3233] In this paper we provide data showing a new role for hex-okinases 2 as an antiapoptotic factor in this microorganism
2 Materials and Methods
21 Yeast Strains and Media Strains used in this studyW303-1A (MATa ade2-1 can1-100 his3-1115 leu2-3112 trp1-1 ura3-1) [34] YSH310 (MATa W303-1A with hxk2LEU2)[34] yca1Δ (MATa W303-1A with yca1URA3) (this study)
eGFP-RBD3 Rhodamine Merge
0 min
5 min
pH
35 m
in
40 mMacetic acid
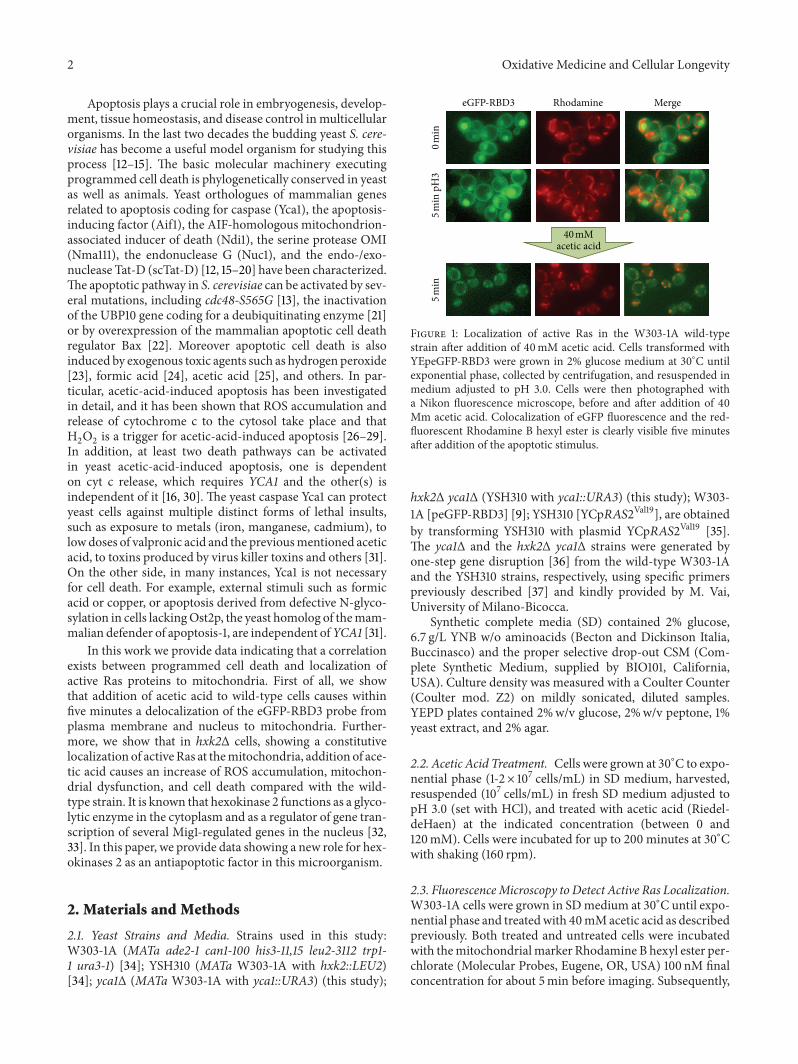
Figure 1 Localization of active Ras in the W303-1A wild-typestrain after addition of 40mM acetic acid Cells transformed withYEpeGFP-RBD3 were grown in 2 glucose medium at 30∘C untilexponential phase collected by centrifugation and resuspended inmedium adjusted to pH 30 Cells were then photographed witha Nikon fluorescence microscope before and after addition of 40Mm acetic acid Colocalization of eGFP fluorescence and the red-fluorescent Rhodamine B hexyl ester is clearly visible five minutesafter addition of the apoptotic stimulus
hxk2Δ yca1Δ (YSH310 with yca1URA3) (this study) W303-1A [peGFP-RBD3] [9] YSH310 [YCp1198771198601198782Val19] are obtainedby transforming YSH310 with plasmid YCp1198771198601198782Val19 [35]The yca1Δ and the hxk2Δ yca1Δ strains were generated byone-step gene disruption [36] from the wild-type W303-1Aand the YSH310 strains respectively using specific primerspreviously described [37] and kindly provided by M VaiUniversity of Milano-Bicocca
Synthetic complete media (SD) contained 2 glucose67 gL YNB wo aminoacids (Becton and Dickinson ItaliaBuccinasco) and the proper selective drop-out CSM (Com-plete Synthetic Medium supplied by BIO101 CaliforniaUSA) Culture density was measured with a Coulter Counter(Coulter mod Z2) on mildly sonicated diluted samplesYEPD plates contained 2wv glucose 2wv peptone 1yeast extract and 2 agar
22 Acetic Acid Treatment Cells were grown at 30∘C to expo-nential phase (1-2times 107 cellsmL) in SD medium harvestedresuspended (107 cellsmL) in fresh SD medium adjusted topH 30 (set with HCl) and treated with acetic acid (Riedel-deHaen) at the indicated concentration (between 0 and120mM) Cells were incubated for up to 200 minutes at 30∘Cwith shaking (160 rpm)
23 FluorescenceMicroscopy to Detect Active Ras LocalizationW303-1A cells were grown in SDmedium at 30∘C until expo-nential phase and treatedwith 40mMacetic acid as describedpreviously Both treated and untreated cells were incubatedwith themitochondrialmarker Rhodamine B hexyl ester per-chlorate (Molecular Probes Eugene OR USA) 100 nM finalconcentration for about 5min before imaging Subsequently
Oxidative Medicine and Cellular Longevity 3
SD pH 53
W303-1A
hxk2
W303-1A
hxk2
W303-1A
hxk2
W303-1A
hxk2
SD pH 30
NT
40 mM
80 mM
120 mM
(a)
0
20
40
60
80
100
120
140
0 50 100 150 2000
20
40
60
80
100
120
140
0 50 100 150 200Time (min)
W303-1A
Surv
ival
cells
()
Surv
ival
cells
()
Time (min)
hxk2
(b)
Figure 2 hxk2Δ cells exhibit inhibition of cell growth and hypersensitivity to acetic acid (a) W303-1A and hxk2Δ cells were harvested andresuspended (1-2times 107 cellsmL) in SD medium adjusted either at pH 53 or at pH 30 (set with HCl) in the absence (NT) or in the presenceof 40 80 or 120mM acetic acid Cells were incubated for 200min at 30∘C with shaking (160 rpm) After treatment cells were harvestedand resuspended at the same concentration (108 cellsmL) in milliQ water 5 microliter from a concentrated suspension and from 10-folddilutions of each culture was spotted onto YEPD plates and incubated at 30∘C for 3 days (b) Cell survival ofW303-1A and hxk2Δ strains Cellviability of W303-1A and hxk2Δ untreated cells (998771) or treated with 80mM (◼) and 120mM (e) acetic acid was analyzed at indicated times bymeasuring colony-forming units (cfu) after 2 days of growth at 30∘C Cell survival (100) corresponds to the cfu at time zero The means of4 independent experiments with standard deviations are reported
4 Oxidative Medicine and Cellular Longevity
40 120583L of cells suspension was seeded on concanavalin A(Sigma-Aldrich Milano Italy) coated cover glass for 10min(100 120583gmL) The cover glass was washed 4 times using theproper medium and put on top of a Thoma chamber Ima-ges were acquired with a Nikon Eclipse E600 microscopeequipped with a 60X 14 oil Plan-Apochromat objective anda standard FITC filter set for GFP fluorescence Images wererecorded digitally using a Leica DC 350F camera and pro-cessed using Adobe Photoshop (Adobe Systems Inc)
24 Acetic Acid Sensitivity This assay was performed essen-tially as described by Casatta et al [38] Exponential-phasecells were harvested and resuspended (107 cellsmL) in SDmedium adjusted either at pH 53 or at pH 30 (set with HCl)and containing 0 40 80 or 120mM acetic acid Cells wereincubated for 200min at 30∘C with shaking (160 rpm) Aftertreatment cells (10-fold serial dilutions) were spotted ontoYEPD plates and incubated at 30∘C for 3 days
25 Viability Assay Cells were grown in SDmedium at 30∘Cuntil exponential phase and treated with acetic acid asdescribed previously At different times (0 30 60 and 200minutes) during acetic acid treatment cell number was cal-culated and 400 cells were plated Viability was determined bymeasuring colony-forming units (cfu) after 2 days of growthon YEPD agar plates at 30∘C The percentage of viable cellsresulted in dividing the number of surviving colonies of thetreated sample by the number of surviving colonies of thesame culture before acid acetic addition
26 Dihydrorhodamine 123 (DHR123) Staining ROS (reac-tive oxygen species) were detected with DHR123 (SigmaAldrich) essentially as described by Madeo et al [23] Cellswere grown in SD medium at 30∘C until exponential phaseand treated with acetic acid as described previously DHR123was added directly to the culture medium at the final con-centration of 5120583gmL from a 25120583g120583L stock solution After2 hours of incubation cells were diluted to 106 cellsmL andanalyzed using a FACScan instrument (Becton Dickinson)at low flow rate with excitation and emission settings of 488and 525ndash550 nm (filter FL 1) A total of 20000 events wereacquired for each sample and data were processed usingWinMDI 29 software
27 410158406-Diamidino-2-phenylindole (DAPI) Staining Cellswere fixed with 37 formaldehyde for 30 minutes stainedwith 2120583gmL of DAPI for 10 minutes washed with distilledwater and resuspended in 50 glycerol solution Images wereacquired with a Nikon Eclipse E600 fluorescence microscopeusing a DAPI filter recorded digitally using a Leica DC 350Fcamera and processed using Adobe Photoshop (Adobe Sys-tems Inc)
28 Annexin V and Propidium Iodide (PI) Staining (FITC)conjugated recombinant Annexin V (Immuno Tools) wasused for the detection of phosphatidylserine exposed in themembrane of apoptotic cells Cells were harvested after 200minutes of acetic acid treatment as reported previously
0102030405060708090
100
Acetic acid
DH
R123
()
lowast
lowast
lowast
lowast
lowastlowast
0 mM 40 mM 80 mM 120 mM
Figure 3 ROS accumulation in W303-1A hxk2Δ and hxk2Δ cellsexpressing the activated Ras2Val19 allele after treatment with aceticacid W303-1A (black bars) hxk2Δ (gray bars) and hxk2Δ cellsexpressing the activated Ras2Val19 allele (white bars) exponentiallygrowing cells were treated with different concentrations (40ndash80ndash120mM) of acetic acid for 200 minutes at 30∘C ROS accumulationwas assayed using the dye dihydrorhodamine 123 (DHR123) by flowcytometry The means of 3 independent experiments with standarddeviations are reported Studentrsquos 119905-test lowast119875 lt 005 and lowastlowast119875 lt 001
washed with sorbitol buffer (1M sorbitol 01M NaH2PO4
pH 80) and the cell wall was digested with Zymolyase 20T(Seikagaku Biobusiness Corporation) for about 35 minutes at37∘C Cells were then washed two times with binding buffer(10mM HepesNaOH pH 74 140mM NaCl 25mM CaCl
2
12M sorbitol) Spheroplasts were resuspended in 35 120583L ofbinding buffer and incubated with 25 120583L of Annexin V and2 120583L of a PI (Fluka) working solution (50120583gmL) for 15minutes in the dark at room temperature After staining thesamples were resuspended in binding buffer and analyzedusing a FACScan instrument (Becton Dickinson) using FL1-H filter on 119909-axis and FL 2-H filter on 119910-axis A total of30000 events were acquired for each sample and data wereprocessed using WinMDI 29 software
29 331015840-Dihexyloxacarbocyanine Iodide (DiOC6) StainingThe mitochondrial morphology and membrane potentialwere assessed by staining with DiOC
6(Molecular Probes
Invitrogen) Cells were grown in SD medium at 30∘C untilexponential phase and treated with acetic acid as describedpreviously DiOC6 was added directly to the culture mediumat the final concentration of 175 nM for 15 minutes in the darkat room temperature After staining the cells were analyzedusing a FACScan instrument (Becton Dickinson) using FL1-H filter A total of 20000 events were acquired for eachsample and data were processed using WinMDI 29 soft-ware Images were also acquired with a Nikon Eclipse E600microscope equipped with a 60X 14 oil Plan-Apochromatobjective and a standard FITC filter set
3 Results and Discussion
31 Effect of Acetic Acid on the Localization of Active Ras inGlucose-Growing Cells In a recent study we showed that
Oxidative Medicine and Cellular Longevity 5
0
5
10
15
20
25
30
35
40A
nnex
in V
pos
(
)
Acetic acid
lowastlowast
lowastlowast
0 mM 40 mM 80 mM 120 mM
(a)
0
5
10
15
20
25
30
35
40
PI p
os (
)
Acetic acid
lowast
lowast
0 mM 40 mM 80 mM 120 mM
(b)
0
10
20
30
40
50
60
70
80
Acetic acid
Ann
exin
V+
PI p
os (
)
lowast
lowast
0 mM 40 mM 80 mM 120 mM
(c)
Figure 4 The occurrence of acetic-acid-induced cell death in hxk2Δ cells is characterized by markers of both apoptosis and necrosisAssessment of cell death by FITC-coupled annexinV and PI stainingW303-1A (black bars) and hxk2Δ (gray bars) exponentially growing cellswere treatedwith different concentrations (40ndash80ndash120mM) of acetic acid for 200minutes at 30∘C before being processed for determination ofphosphatidylserine externalization andmembrane integrity by flow cytometry 30000 events have been evaluatedThemeans of 3 independentexperiments with standard deviations are reported Studentrsquos 119905-test lowast119875 lt 005 and lowastlowast119875 lt 001
active Ras proteins accumulated mainly in the plasma mem-brane and in the nucleus in exponentially growing wild-typecells while they accumulated inmitochondria in cells deletedin the HXK2 gene indicating that this enzyme was involvedin the proper localization of these small GTPases [9] Therole played by active Ras in mitochondria is not knownalthough other reports have found Ras-GTP associated withthese organelles both inmammals and lower eukaryotes andhave linked it to cancer and apoptosis Leadsham et al [10]demonstrated that in S cerevisiae loss of Whi2p function aprotein known to influence cell cycle exit under conditions ofnutritional depletion determined an aberrant accumulationof activated Ras at the mitochondria In this mutant thefailure to shut down Ras signalling by addressing Ras to thevacuole would lead tomitochondrial dysfunction accumula-tion of damaging ROS and cell deathMore recently Hu et al[11] using a tetracycline inducible model demonstrated thatin mammalian cells association of K-rasG12V proteins withmitochondria induced mitochondrial dysfunction increasedROS accumulation and a metabolic switch from oxidativephosphorylation to glycolysis
To investigate whether a correlation exists between themitochondrial localization of active Ras and programmed
cell death we evaluated the effect of the addition of aceticacid a well-known apoptotic stimulus [25] to wild-typecells expressing the eGFP-RBD3 probe on the localizationof Ras-GTP proteins To this aim exponentially growingcells were collected by centrifugation resuspended in low pHmedium (pH 30) and pictures were taken at the fluorescencemicroscope before and after addition of 40mM acetic acidOur results showed that within 5 minutes this apoptoticstimulus caused the localization of active Ras proteins exclu-sively to the mitochondria reinforcing the hypothesis ofthe involvement of these proteins in programmed cell death(Figure 1) In wild-type cells resuspended in low pHmediumfor 5 minutes without acetic acid added active Ras proteinsaccumulated mainly in the plasma membrane and in thenucleus indicating that acidification of the medium did notinfluence the localization of these small G proteins (Figure 1)A mitochondrial localization of Ras-GTP was also observedwhen a low concentration of H
2O2was added to the medium
(data not shown) indicating that delocalization of active Rasto these organelles is a more general response to differentapoptotic stimuli and suggesting that mitochondrial localiza-tion of Ras2-GTP is actually important for the induction ofapoptosis in yeast
6 Oxidative Medicine and Cellular Longevity
W303-1A
256
0
Fluorescence
Even
ts
100
101
102
103
104
0 mM
40 mM
80 mM
120 mM
Fluorescence
Even
ts
256
010
010
110
210
310
4
0 mM
40 mM
80 mM
120 mM
hxk2
(a)
W303-1A
0 mM 80 mM
hxk2
(b)
Figure 5 Mitochondrial membrane potential and morphology in W303-1A and hxk2Δ strains upon acetic acid treatment W303-1A andhxk2Δ exponentially growing cells were treated with different concentrations (40ndash80ndash120mM) of acetic acid for 200 minutes at 30∘C DiOC
6
uptake was assessed using both flow cytometry (a) and fluorescence microscopy (b) Photomicrographs illustrate the alteration of the tubularmitochondrial network to clustered mitochondrial morphology particularly exacerbated in hxk2Δ cells after acetic acid treatment
32 Deletion of HXK2 Enhances Cell Death and IncreasesIntracellular ROS Levels The results presented previouslysuggest that localization of active Ras proteins to mitochon-dria might be involved in programmed cell death since theseproteins are localized to mitochondria following additionof acetic acid Since in glucose-growing hxk2Δ cells activeRas is constitutively located in mitochondria [9] we nextanalyzed the behavior of the hxk2Δmutant under conditions
that caused apoptosisWhile the growth of hxk2Δmutant wasalmost indistinguishable from the wild-type strain at pH 53it exhibited an increased sensitivity to acetic acid stress at lowpH (Figure 2(a)) suggesting that Hxk2 is required for normaltolerance to acetic acid treatment Cell survival of wild-typeand hxk2Δ cells after induction of apoptosis with acetic acidwas further tested in a plating assay After treatment withdifferent concentrations (80 and 120mM) of acetic acid for
Oxidative Medicine and Cellular Longevity 7
up to 200 minutes at 30∘C hxk2Δ cells showed a significantdose-dependent reduction in cell survival when comparedwith wild-type cells (Figure 2(b)) Taken together these datasuggest that Hxk2 is required for normal tolerance to aceticacid treatment
Since an aberrant accumulation of activated Ras in mito-chondria accompanied to accumulation of ROS has alreadybeen reported both in mammals and yeast [10 11] and sinceROS play a pivotal role in apoptotic cell death FACS analyseswere performed to evaluate the accumulation of ROS in wild-type and hxk2Δ cells treated with different concentrationsof acetic acid for 200min at 30∘C In particular DHR123was used to determine the accumulation of ROS in the cellssince this compound can easily permeate them and can bequantitatively oxidized to a green fluorescent product inthe presence of ROS The percentage of oxidized R123 wasabout the double for hxk2Δ cells compared to wild-type cells200min after treatment with either 40 or 80mM acetic acid(Figure 3) To substantiate the hypothesis that an aberrantaccumulation of activated Ras in mitochondria might leadtomitochondrial dysfunction and accumulation of damagingROS we expressed the dominant active RAS2Val19 allelewhich was reported to show a much higher level of Ras2-GTP [39] in hxk2Δ cells Our results showed that indeed theexpression of this allele in the hxk2Δ background caused afurther increase in the level of ROS both in growing cells andafter treatment with acetic acid (Figure 3)These data suggestthat conditions that presumably cause a highermitochondrialaccumulation of active Ras may contribute to accumulationof ROS and cell death
33 Loss of Hxk2 Causes an Increase of Both Apoptosis andNecrosis The results presented previously suggest that Hxk2might have an antiapoptotic activity To better characterizethe nature of cell death triggered by addition of acetic acid tohxk2Δ cells we quantified phenotypic changes indicative ofapoptosis While apoptotic DNA condensation was detectedby DAPI staining combined Annexin Vpropidium iodide(PI) staining was used to discriminate between early apop-totic (Annexin V+PIminus) late apoptoticsecondary necrotic(Annexin V+PI+) and necrotic (Annexin VminusPI+) deathsDAPI staining showed that nuclei of untreated cells wereround while the nuclear DNA was condensed in bothwild-type and hxk2Δ cells treated with 80mM acetic acidbeing the extent of this phenotype more pronounced for themutant strain compared to the wild-type strain (data notshown) Annexin Vpropidium iodide (PI) staining revealedthat hxk2Δ-facilitated cell death was accompanied by anincrease in both apoptotic and necroticmarkers (Figure 4) Inparticular untreated hxk2Δ cells showed a higher percentageof both early apoptotic and necrotic cells compared withthe wild-type strain which further increased upon treatmentwith acetic acid
34 Deletion of HXK2 Causes Hyperpolarization of Mitochon-dria Mitochondrial membrane potential (ΔΨ
119898) is a useful
indicator of mitochondrial function To measure ΔΨ119898 we
used the cationic lipophylic dye DiOC6 which accumulates
0102030405060708090
100110
DH
R 12
3 (
)
Acetic acid
lowast
lowast
lowast
0 mM 40 mM 80 mM 120 mM
Figure 6 Effect of YCA1 deletion on ROS accumulation aftertreatment with acetic acid W303-1A (black bars) yca1Δ (blackhatched bars) hxk2Δ (gray bars) and hxk2Δ-yca1Δ (white bars)exponentially growing cells were treated with different concen-trations (40ndash80ndash120mM) of acetic acid for 200 minutes at 30∘CROS accumulation was assayed by flow cytometry using the dyedihydrorhodamine 123 (DHR123) Studentrsquos 119905-test lowast119875 lt 005
in mitochondria in accordance with ΔΨ119898 Wild-type and
hxk2Δ cells were treatedwith different concentration of aceticacid (40ndash80ndash120mM) for 200min at 30∘C collected andstained with this dye FACS analysis showed that mitochon-drial membrane potential clearly increased in both wild-typeand hxk2Δ cells after treatment with acetic acid comparedwith untreated control cells with the hyperpolarizationof mitochondria being much stronger in the mutant cellscomparedwithwild-type cells (Figure 5(a)) In parallel mito-chondrial morphology was assessed by using DiOC
6 both
in wild-type and hxk2Δ cells At low concentration this dyespecifically stains the mitochondrial membranes in a mannerthat depends on membrane potential and can be observed byfluorescence microscopy Before treatment with acetic acidbothwild-type and hxk2Δ cells displayed a tubularmitochon-drial morphology indicating that these mitochondria werehealthy and possessed a membrane potential (Figure 5(b))By contrast the mitochondrial membranes present in bothwild-type and hxk2Δ cells appeared rounded and highlyfragmented after treatment with acetic acid (Figure 5(b))This conversion of mitochondrial morphology from tubu-lar to punctuate structures is likely to occur by excessivemitochondrial fission and has already been observed in yeastapoptosis induced by acetic acid treatment [40] ImportantlyDiOC
6staining was greatly increased and more pronounced
in hxk2Δ cells compared to wild-type cells after treatmentwith acetic acid confirming the increase of fluorescenceobserved in these mutant cells by FACS analysis Thesedata indicate that both in wild-type and hxk2Δ cells aceticacid treatment caused hyperpolarization of mitochondrialmembrane with consequent damage of these organelles andloss of functionality This effect was more pronounced inhxk2Δ cells compared to wild-type cells
35 Acetic-Acid-Induced Cell Death in the hxk2Δ Strain Is Yca1Independent At least two death pathways can be activated
8 Oxidative Medicine and Cellular Longevity
lowastlowast
lowastlowast
lowast
lowastlowast
0 mM 40 mM 80 mM 120 mMAcetic acid
0
5
10
15
20
25
30
35
40A
nnex
in V
pos
(
)
(a)
lowast
lowast
lowastlowast
lowastlowast
0 mM 40 mM 80 mM 120 mM0
10
20
30
40
50
60
70
80
PI p
os (
)
Acetic acid
(b)
lowastlowast
lowast
lowast
0 mM 40 mM 80 mM 120 mM0
10
20
30
40
50
60
70
80
Acetic acid
Ann
exin
V+
PI p
os (
)
(c)
Figure 7 Effect of YCA1 deletion on cell death after treatment with acetic acid W303-1A (black bars) yca1Δ (black hatched bars) hxk2Δ(gray bars) and hxk2Δ-yca1Δ (white bars) exponentially growing cells were treated with different concentrations (40ndash80ndash120mM) of aceticacid for 200 minutes at 30∘C Cell death was assessed by flow cytometry using FITC-coupled annexin V and PI co-staining to determinatethe externalization of phosphatidylserine and the membrane integrityThemeans of 3 independent experiments with standard deviations arereported Studentrsquos 119905-test lowast119875 lt 005 and lowastlowast119875 lt 001
in yeast acetic-acid-induced apoptosis one dependent oncytochromec release which requires YCA1 and the other(s)independent of it [16 30] Consequently we next investigatedwhether the yeast caspase Yca1 might be involved in acetic-acid-induced cell death in hxk2Δ cells In particular toassess whether Yca1 plays a role in acetic-acid-induced ROSaccumulation in hxk2Δ cells we treated hxk2Δ and hxk2Δyca1Δ cells with different concentration of acetic acid (40ndash80ndash120mM) for 200min at 30∘C and measured the ROSaccumulation Our results showed that deletion of YCA1 inhxk2Δ cells had no effect on acetic-acid-induced ROS accu-mulation (Figure 6) suggesting that the generation of ROSin hxk2Δ cells upon acetic acid stress is Yca1 independentHowever the level of intracellular ROS was consistentlyhigher in the hxk2Δ yca1Δ doublemutant growing on glucosemedium compared with the hxk2Δ mutant Similarly Du etal [24] showed that formic acid induced Yca1-independentapoptosis-like cell death and that the burst of ROS duringcell death occurred earlier and at a higher level in the yca1Δstrain than in the wild-type strain Moreover Khan et al [41]determined the level of oxidized proteins in yeast cells underH2O2stress and showed that lack of Yca1 abrogated apoptosis
but elevated intracellular oxidized proteins compared with
wild-type cells Consequently both our results and data in theliterature [24 41] clearly suggest a linkage between ROS pro-duction and Yca1 activation during apoptosis in yeast Finallycytofluorometric quantification of phosphatidylserine exter-nalization andormembrane permeabilization (annexinVPIcostaining) further confirmed that Yca1p did not influencethe acetic-acid-induced cell death in hxk2Δ cells (Figure 7)These results strongly suggest that apoptosis induced by aceticacid in hxk2Δ cells is largely Yca1 independent
4 Conclusion
Several studies indicate that the Ras proteinmay be physicallyassociated with mitochondria both in yeast and mammals[10 11 42ndash44] This study provides results indicating thatassociation of active Ras to these organelles might be linkedto apoptosis First of all we show that addition of acetic acida well-known apoptotic stimulus in S cerevisiae to growingwild-type cells determined a delocalization of active Ras pro-teins from nuclei and plasma membrane to mitochondriaFurthermore we show that addition of acetic acid to hxk2Δcells showing a constitutive localization of active Ras inmito-chondria [9] enhanced ROS production and cell death
Oxidative Medicine and Cellular Longevity 9
compared with wild-type cells Moreover expression of thedominant active RAS2Val19 allele in the hxk2Δ backgroundcaused an even higher increase in the level of ROS both ingrowing cells and after treatment with acetic acid providinga further proof to the hypothesis that conditions that presum-ably cause a higher mitochondrial accumulation of activeRas may contribute to accumulation of ROS and cell deathWe also show that in hxk2Δ cells addition of acetic acidincreased cell death compared with the wild-type strain withthe typical markers of apoptosis highlighting a new rolefor Hexokinase 2 as an antiapoptotic factor in S cerevisiaecells However at this stage we cannot exclude a correlationbetween the well-known functions of Hxk2 in glucose repres-sion and signalling and the role of Ras in inducing apoptosis
Finally in this study we show that lack of Hxk2 inducesapoptosis via a mitochondria-mediated pathway withoutmetacaspase Yca1 involvement since deletion of YCA1 in thehxk2Δ background did not abrogate the acetic-acid-inducedaccumulation of ROS and did not decrease the percentage ofboth apoptotic and necrotic cells
Acknowledgments
The authors thank J Winderickx University of Leuven Bel-gium for the gift of strains and M Vai Universita Milano-Bicocca Milan for the gift of primers This work was sup-ported by FAR grant to Sonia Colombo and by founding ofProgram SysBioNet Italian Roadmap Research Infrastruc-ture 2012
References
[1] S JonesM-L Vignais and J R Broach ldquoTheCDC25 protein ofSaccharomyces cerevisiae promotes exchange of guanine nucle-otides bound to RasrdquoMolecular and Cellular Biology vol 11 no5 pp 2641ndash2646 1991
[2] S Rudoni S Colombo P Coccetti and E Martegani ldquoRole ofguanine nucleotides in the regulation of the RascAMPpathwayin Saccharomyces cerevisiaerdquo Biochimica et Biophysica Acta vol1538 no 2-3 pp 181ndash189 2001
[3] K TanakaMNakafuku T Satoh et al ldquoS cerevisiae genes IRA1and IRA2 encode proteins that may be functionally equivalentto mammalian ras GTPase activating proteinrdquo Cell vol 60 no5 pp 803ndash807 1990
[4] S Zaman S I Lippman X Zhao and J R Broach ldquoHow Sac-charomyces responds to nutrientsrdquo Annual Review of Geneticsvol 42 pp 27ndash81 2008
[5] JMThevelein and J H DeWinde ldquoNovel sensingmechanismsand targets for the cAMP-protein kinase A pathway in the yeastSaccharomyces cerevisiaerdquoMolecularMicrobiology vol 33 no 5pp 904ndash918 1999
[6] F Rolland JWinderickx and JMThevelein ldquoGlucose-sensingand -signallingmechanisms in yeastrdquo FEMS Yeast Research vol2 no 2 pp 183ndash201 2002
[7] G F Casperson N Walker and H R Bourne ldquoIsolation of thegene encoding adenylate cyclase in Saccharomyces cerevisiaerdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 82 no 15 pp 5060ndash5063 1985
[8] B Smets R Ghillebert P De Snijder et al ldquoLife in the midstof scarcity adaptations to nutrient availability in Saccharomycescerevisiaerdquo Current Genetics vol 56 no 1 pp 1ndash32 2010
[9] S Broggi E Martegani and S Colombo ldquoLive-cell imaging ofendogenous Ras-GTP shows predominant Ras activation at theplasma membrane and in the nucleus in Saccharomyces cere-visiaerdquoThe International Journal of Biochemistry amp Cell Biologyvol 45 no 2 pp 384ndash394 2013
[10] J E Leadsham K Miller K R Ayscough et al ldquoWhi2p linksnutritional sensing to actin-dependent Ras-cAMP-PKA regu-lation and apoptosis in yeastrdquo Journal of Cell Science vol 122no 5 pp 706ndash715 2009
[11] YHuW LuG Chen et al ldquoK-rasG12V transformation leads tomitochondrial dysfunction and a metabolic switch from oxida-tive phosphorylation to glycolysisrdquo Cell Research vol 22 no 2pp 399ndash412 2012
[12] S Buttner T Eisenberg D Carmona-Gutierrez et al ldquoEndonu-clease G regulates budding yeast life and deathrdquoMolecular Cellvol 25 no 2 pp 233ndash246 2007
[13] F Madeo E Frohlich and K-U Frohlich ldquoA yeast mutantshowing diagnostic markers of early and late apoptosisrdquo Journalof Cell Biology vol 139 no 3 pp 729ndash734 1997
[14] FMadeo E Herker SWissing H Jungwirth T Eisenberg andK-U Frohlich ldquoApoptosis in yeastrdquo Current Opinion in Micro-biology vol 7 no 6 pp 655ndash660 2004
[15] F Madeo E Herker C Maldener et al ldquoA caspase-related pro-tease regulates apoptosis in yeastrdquo Molecular Cell vol 9 no 4pp 911ndash917 2002
[16] N Guaragnella C Pereira M J Sousa et al ldquoYCA1 participatesin the acetic acid induced yeast programmed cell death also in amanner unrelated to its caspase-like activityrdquo FEBS Letters vol580 no 30 pp 6880ndash6884 2006
[17] W Li L Sun Q Liang J Wang W Mo and B Zhou ldquoYeastAMID homologue Ndi1p displays respiration-restricted apop-totic activity and is involved in chronological agingrdquoMolecularBiology of the Cell vol 17 no 4 pp 1802ndash1811 2006
[18] S Wissing P Ludovico E Herker et al ldquoAn AIF orthologueregulates apoptosis in yeastrdquo Journal of Cell Biology vol 166 no7 pp 969ndash974 2004
[19] B Fahrenkrog U Sauder and U Aebi ldquoThe S cerevisiae HtrA-like protein Nma111p is a nuclear serine protease that mediatesyeast apoptosisrdquo Journal of Cell Science vol 117 no 1 pp 115ndash126 2004
[20] J Qiu J-H Yoon and B Shen ldquoSearch for apoptotic nucleasesin yeast role of Tat-D nuclease in apoptotic DNA degradationrdquoThe Journal of Biological Chemistry vol 280 no 15 pp 15370ndash15379 2005
[21] I Orlandi M Bettiga L Alberghina and M Vai ldquoTranscrip-tional profiling of ubp10 null mutant reveals altered subtelo-meric gene expression and insurgence of oxidative stressresponserdquoThe Journal of Biological Chemistry vol 279 no 8 pp6414ndash6425 2004
[22] M Ligr F Madeo E Frohlich W Hilt K-U Frohlich and DH Wolf ldquoMammalian Bax triggers apoptotic changes in yeastrdquoFEBS Letters vol 438 no 1-2 pp 61ndash65 1998
[23] F Madeo E Frohlich M Ligr et al ldquoOxygen stress a regulatorof apoptosis in yeastrdquo Journal of Cell Biology vol 145 no 4 pp757ndash767 1999
[24] L Du Y Su D Sun et al ldquoFormic acid induces Yca1p-inde-pendent apoptosis-like cell death in the yeast Saccharomycescerevisiaerdquo FEMS Yeast Research vol 8 no 4 pp 531ndash539 2008
10 Oxidative Medicine and Cellular Longevity
[25] P LudovicoM J SousaM T Silva C Leao andMCorte-RealldquoSaccharomyces cerevisiae commits to a programmed cell deathprocess in response to acetic acidrdquoMicrobiology vol 147 no 9pp 2409ndash2415 2001
[26] P Ludovico F Rodrigues AAlmeidaMT Silva A Barrientosand M Corte-Real ldquoCytochrome c release and mitochondriainvolvement in programmed cell death induced by acetic acidin Saccharomyces cerevisiaerdquo Molecular Biology of the Cell vol13 no 8 pp 2598ndash2606 2002
[27] N Guaragnella L Antonacci S Passarella E Marra and SGiannattasio ldquoHydrogen peroxide and superoxide anion pro-duction during acetic acid-induced yeast programmed celldeathrdquo Folia Microbiologica vol 52 no 3 pp 237ndash240 2007
[28] S Giannattasio A Atlante L Antonacci et al ldquoCytochrome c isreleased from coupled mitochondria of yeast en route to aceticacid-induced programmed cell death and can work as an elec-tron donor and a ROS scavengerrdquo FEBS Letters vol 582 no 10pp 1519ndash1525 2008
[29] N Guaragnella L Antonacci S Giannattasio E Marra and SPassarella ldquoCatalase T and Cu Zn-superoxide dismutase in theacetic acid-induced programmed cell death in Saccharomycescerevisiaerdquo FEBS Letters vol 582 no 2 pp 210ndash214 2008
[30] N Guaragnella A Bobba S Passarella E Marra and S Gian-nattasio ldquoYeast acetic acid-induced programmed cell death canoccurwithout cytochrome c releasewhich requiresmetacaspaseYCA1rdquo FEBS Letters vol 584 no 1 pp 224ndash228 2010
[31] F Madeo D Carmona-Gutierrez J Ring S Buttner T Eisen-berg and G Kroemer ldquoCaspase-dependent and caspase-inde-pendent cell death pathways in yeastrdquo Biochemical and Biophys-ical Research Communications vol 382 no 2 pp 227ndash231 2009
[32] DAhuatzi A Riera R Pelaez PHerrero andFMoreno ldquoHxk2regulates the phosphorylation state of Mig1 and therefore itsnucleocytoplasmic distributionrdquo The Journal of BiologicalChemistry vol 282 no 7 pp 4485ndash4493 2007
[33] FMoreno D Ahuatzi A Riera C A Palomino and P HerreroldquoGlucose sensing through the Hxk2-dependent signalling path-wayrdquo Biochemical Society Transactions vol 33 no 1 pp 265ndash268 2005
[34] B JThomas and R Rothstein ldquoElevated recombination rates intranscriptionally active DNArdquo Cell vol 56 no 4 pp 619ndash6301989
[35] L C Robinson J B Gibbs M S Marshall I S Sigal and KTatchell ldquoCDC25 a component of the RAS-adenylate cyclasepathway in Saccharomyces cerevisiaerdquo Science vol 235 no 4793pp 1218ndash1221 1987
[36] R J Rothstein ldquoOne-step gene disruption in yeastrdquoMethods inEnzymology vol 101 pp 202ndash211 1983
[37] M Bettiga L Calzari I Orlandi L Alberghina and M VaildquoInvolvement of the yeast metacaspase Yca1 in ubp10Δ-pro-grammed cell deathrdquo FEMS Yeast Research vol 5 no 2 pp 141ndash147 2004
[38] N Casatta A Porro I Orlandi L Brambilla andM Vai ldquoLackof Sir2 increases acetate consumption and decreases extracellu-lar pro-aging factorsrdquo Biochimica et Biophysica Acta vol 1833no 3 pp 593ndash601 2013
[39] S Colombo D Ronchetti J M Thevelein J Winderickx andE Martegani ldquoActivation state of the Ras2 protein and glucose-induced signaling in Saccharomyces cerevisiaerdquo The Journal ofBiological Chemistry vol 279 no 45 pp 46715ndash46722 2004
[40] P Fabrizio and V D Longo ldquoChronological aging-inducedapoptosis in yeastrdquo Biochimica et Biophysica Acta vol 1783 no7 pp 1280ndash1285 2008
[41] M A S Khan P B Chock and E R Stadtman ldquoKnockout ofcaspase-like gene YCA1 abrogates apoptosis and elevates oxi-dized proteins in Saccharomyces cerevisiaerdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 102 no 48 pp 17326ndash17331 2005
[42] J CWolfman S M Planchon J Liao and AWolfman ldquoStruc-tural and functional consequences of c-N-Ras constitutivelyassociated with intact mitochondriardquo Biochimica et BiophysicaActa vol 1763 no 10 pp 1108ndash1124 2006
[43] T G Bivona S E Quatela B O Bodemann et al ldquoPKC regu-lates a farnesyl-electrostatic switch on K-Ras that promotes itsassociation with Bcl-XL on mitochondria and induces apopto-sisrdquoMolecular Cell vol 21 no 4 pp 481ndash493 2006
[44] A Rebollo D Perez-Sala and C Martınez-A ldquoBcl-2 differen-tially targets K- N- and H-Ras to mitochondria in IL-2 sup-plemented or deprived cells implications in prevention of apop-tosisrdquo Oncogene vol 18 no 35 pp 4930ndash4939 1999
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
2 Oxidative Medicine and Cellular Longevity
Apoptosis plays a crucial role in embryogenesis develop-ment tissue homeostasis and disease control inmulticellularorganisms In the last two decades the budding yeast S cere-visiae has become a useful model organism for studying thisprocess [12ndash15] The basic molecular machinery executingprogrammed cell death is phylogenetically conserved in yeastas well as animals Yeast orthologues of mammalian genesrelated to apoptosis coding for caspase (Yca1) the apoptosis-inducing factor (Aif1) the AIF-homologous mitochondrion-associated inducer of death (Ndi1) the serine protease OMI(Nma111) the endonuclease G (Nuc1) and the endo-exo-nuclease Tat-D (scTat-D) [12 15ndash20] have been characterizedThe apoptotic pathway in S cerevisiae can be activated by sev-eral mutations including cdc48-S565G [13] the inactivationof the UBP10 gene coding for a deubiquitinating enzyme [21]or by overexpression of the mammalian apoptotic cell deathregulator Bax [22] Moreover apoptotic cell death is alsoinduced by exogenous toxic agents such as hydrogen peroxide[23] formic acid [24] acetic acid [25] and others In par-ticular acetic-acid-induced apoptosis has been investigatedin detail and it has been shown that ROS accumulation andrelease of cytochrome c to the cytosol take place and thatH2O2is a trigger for acetic-acid-induced apoptosis [26ndash29]
In addition at least two death pathways can be activatedin yeast acetic-acid-induced apoptosis one is dependenton cyt c release which requires YCA1 and the other(s) isindependent of it [16 30] The yeast caspase Yca1 can protectyeast cells against multiple distinct forms of lethal insultssuch as exposure to metals (iron manganese cadmium) tolowdoses of valpronic acid and the previousmentioned aceticacid to toxins produced by virus killer toxins and others [31]On the other side in many instances Yca1 is not necessaryfor cell death For example external stimuli such as formicacid or copper or apoptosis derived from defective N-glyco-sylation in cells lackingOst2p the yeast homolog of themam-malian defender of apoptosis-1 are independent ofYCA1 [31]
In this work we provide data indicating that a correlationexists between programmed cell death and localization ofactive Ras proteins to mitochondria First of all we showthat addition of acetic acid to wild-type cells causes withinfive minutes a delocalization of the eGFP-RBD3 probe fromplasma membrane and nucleus to mitochondria Further-more we show that in hxk2Δ cells showing a constitutivelocalization of activeRas at themitochondria addition of ace-tic acid causes an increase of ROS accumulation mitochon-drial dysfunction and cell death compared with the wild-type strain It is known that hexokinase 2 functions as a glyco-lytic enzyme in the cytoplasm and as a regulator of gene tran-scription of several Mig1-regulated genes in the nucleus [3233] In this paper we provide data showing a new role for hex-okinases 2 as an antiapoptotic factor in this microorganism
2 Materials and Methods
21 Yeast Strains and Media Strains used in this studyW303-1A (MATa ade2-1 can1-100 his3-1115 leu2-3112 trp1-1 ura3-1) [34] YSH310 (MATa W303-1A with hxk2LEU2)[34] yca1Δ (MATa W303-1A with yca1URA3) (this study)
eGFP-RBD3 Rhodamine Merge
0 min
5 min
pH
35 m
in
40 mMacetic acid
Figure 1 Localization of active Ras in the W303-1A wild-typestrain after addition of 40mM acetic acid Cells transformed withYEpeGFP-RBD3 were grown in 2 glucose medium at 30∘C untilexponential phase collected by centrifugation and resuspended inmedium adjusted to pH 30 Cells were then photographed witha Nikon fluorescence microscope before and after addition of 40Mm acetic acid Colocalization of eGFP fluorescence and the red-fluorescent Rhodamine B hexyl ester is clearly visible five minutesafter addition of the apoptotic stimulus
hxk2Δ yca1Δ (YSH310 with yca1URA3) (this study) W303-1A [peGFP-RBD3] [9] YSH310 [YCp1198771198601198782Val19] are obtainedby transforming YSH310 with plasmid YCp1198771198601198782Val19 [35]The yca1Δ and the hxk2Δ yca1Δ strains were generated byone-step gene disruption [36] from the wild-type W303-1Aand the YSH310 strains respectively using specific primerspreviously described [37] and kindly provided by M VaiUniversity of Milano-Bicocca
Synthetic complete media (SD) contained 2 glucose67 gL YNB wo aminoacids (Becton and Dickinson ItaliaBuccinasco) and the proper selective drop-out CSM (Com-plete Synthetic Medium supplied by BIO101 CaliforniaUSA) Culture density was measured with a Coulter Counter(Coulter mod Z2) on mildly sonicated diluted samplesYEPD plates contained 2wv glucose 2wv peptone 1yeast extract and 2 agar
22 Acetic Acid Treatment Cells were grown at 30∘C to expo-nential phase (1-2times 107 cellsmL) in SD medium harvestedresuspended (107 cellsmL) in fresh SD medium adjusted topH 30 (set with HCl) and treated with acetic acid (Riedel-deHaen) at the indicated concentration (between 0 and120mM) Cells were incubated for up to 200 minutes at 30∘Cwith shaking (160 rpm)
23 FluorescenceMicroscopy to Detect Active Ras LocalizationW303-1A cells were grown in SDmedium at 30∘C until expo-nential phase and treatedwith 40mMacetic acid as describedpreviously Both treated and untreated cells were incubatedwith themitochondrialmarker Rhodamine B hexyl ester per-chlorate (Molecular Probes Eugene OR USA) 100 nM finalconcentration for about 5min before imaging Subsequently
Oxidative Medicine and Cellular Longevity 3
SD pH 53
W303-1A
hxk2
W303-1A
hxk2
W303-1A
hxk2
W303-1A
hxk2
SD pH 30
NT
40 mM
80 mM
120 mM
(a)
0
20
40
60
80
100
120
140
0 50 100 150 2000
20
40
60
80
100
120
140
0 50 100 150 200Time (min)
W303-1A
Surv
ival
cells
()
Surv
ival
cells
()
Time (min)
hxk2
(b)
Figure 2 hxk2Δ cells exhibit inhibition of cell growth and hypersensitivity to acetic acid (a) W303-1A and hxk2Δ cells were harvested andresuspended (1-2times 107 cellsmL) in SD medium adjusted either at pH 53 or at pH 30 (set with HCl) in the absence (NT) or in the presenceof 40 80 or 120mM acetic acid Cells were incubated for 200min at 30∘C with shaking (160 rpm) After treatment cells were harvestedand resuspended at the same concentration (108 cellsmL) in milliQ water 5 microliter from a concentrated suspension and from 10-folddilutions of each culture was spotted onto YEPD plates and incubated at 30∘C for 3 days (b) Cell survival ofW303-1A and hxk2Δ strains Cellviability of W303-1A and hxk2Δ untreated cells (998771) or treated with 80mM (◼) and 120mM (e) acetic acid was analyzed at indicated times bymeasuring colony-forming units (cfu) after 2 days of growth at 30∘C Cell survival (100) corresponds to the cfu at time zero The means of4 independent experiments with standard deviations are reported
4 Oxidative Medicine and Cellular Longevity
40 120583L of cells suspension was seeded on concanavalin A(Sigma-Aldrich Milano Italy) coated cover glass for 10min(100 120583gmL) The cover glass was washed 4 times using theproper medium and put on top of a Thoma chamber Ima-ges were acquired with a Nikon Eclipse E600 microscopeequipped with a 60X 14 oil Plan-Apochromat objective anda standard FITC filter set for GFP fluorescence Images wererecorded digitally using a Leica DC 350F camera and pro-cessed using Adobe Photoshop (Adobe Systems Inc)
24 Acetic Acid Sensitivity This assay was performed essen-tially as described by Casatta et al [38] Exponential-phasecells were harvested and resuspended (107 cellsmL) in SDmedium adjusted either at pH 53 or at pH 30 (set with HCl)and containing 0 40 80 or 120mM acetic acid Cells wereincubated for 200min at 30∘C with shaking (160 rpm) Aftertreatment cells (10-fold serial dilutions) were spotted ontoYEPD plates and incubated at 30∘C for 3 days
25 Viability Assay Cells were grown in SDmedium at 30∘Cuntil exponential phase and treated with acetic acid asdescribed previously At different times (0 30 60 and 200minutes) during acetic acid treatment cell number was cal-culated and 400 cells were plated Viability was determined bymeasuring colony-forming units (cfu) after 2 days of growthon YEPD agar plates at 30∘C The percentage of viable cellsresulted in dividing the number of surviving colonies of thetreated sample by the number of surviving colonies of thesame culture before acid acetic addition
26 Dihydrorhodamine 123 (DHR123) Staining ROS (reac-tive oxygen species) were detected with DHR123 (SigmaAldrich) essentially as described by Madeo et al [23] Cellswere grown in SD medium at 30∘C until exponential phaseand treated with acetic acid as described previously DHR123was added directly to the culture medium at the final con-centration of 5120583gmL from a 25120583g120583L stock solution After2 hours of incubation cells were diluted to 106 cellsmL andanalyzed using a FACScan instrument (Becton Dickinson)at low flow rate with excitation and emission settings of 488and 525ndash550 nm (filter FL 1) A total of 20000 events wereacquired for each sample and data were processed usingWinMDI 29 software
27 410158406-Diamidino-2-phenylindole (DAPI) Staining Cellswere fixed with 37 formaldehyde for 30 minutes stainedwith 2120583gmL of DAPI for 10 minutes washed with distilledwater and resuspended in 50 glycerol solution Images wereacquired with a Nikon Eclipse E600 fluorescence microscopeusing a DAPI filter recorded digitally using a Leica DC 350Fcamera and processed using Adobe Photoshop (Adobe Sys-tems Inc)
28 Annexin V and Propidium Iodide (PI) Staining (FITC)conjugated recombinant Annexin V (Immuno Tools) wasused for the detection of phosphatidylserine exposed in themembrane of apoptotic cells Cells were harvested after 200minutes of acetic acid treatment as reported previously
0102030405060708090
100
Acetic acid
DH
R123
()
lowast
lowast
lowast
lowast
lowastlowast
0 mM 40 mM 80 mM 120 mM
Figure 3 ROS accumulation in W303-1A hxk2Δ and hxk2Δ cellsexpressing the activated Ras2Val19 allele after treatment with aceticacid W303-1A (black bars) hxk2Δ (gray bars) and hxk2Δ cellsexpressing the activated Ras2Val19 allele (white bars) exponentiallygrowing cells were treated with different concentrations (40ndash80ndash120mM) of acetic acid for 200 minutes at 30∘C ROS accumulationwas assayed using the dye dihydrorhodamine 123 (DHR123) by flowcytometry The means of 3 independent experiments with standarddeviations are reported Studentrsquos 119905-test lowast119875 lt 005 and lowastlowast119875 lt 001
washed with sorbitol buffer (1M sorbitol 01M NaH2PO4
pH 80) and the cell wall was digested with Zymolyase 20T(Seikagaku Biobusiness Corporation) for about 35 minutes at37∘C Cells were then washed two times with binding buffer(10mM HepesNaOH pH 74 140mM NaCl 25mM CaCl
2
12M sorbitol) Spheroplasts were resuspended in 35 120583L ofbinding buffer and incubated with 25 120583L of Annexin V and2 120583L of a PI (Fluka) working solution (50120583gmL) for 15minutes in the dark at room temperature After staining thesamples were resuspended in binding buffer and analyzedusing a FACScan instrument (Becton Dickinson) using FL1-H filter on 119909-axis and FL 2-H filter on 119910-axis A total of30000 events were acquired for each sample and data wereprocessed using WinMDI 29 software
29 331015840-Dihexyloxacarbocyanine Iodide (DiOC6) StainingThe mitochondrial morphology and membrane potentialwere assessed by staining with DiOC
6(Molecular Probes
Invitrogen) Cells were grown in SD medium at 30∘C untilexponential phase and treated with acetic acid as describedpreviously DiOC6 was added directly to the culture mediumat the final concentration of 175 nM for 15 minutes in the darkat room temperature After staining the cells were analyzedusing a FACScan instrument (Becton Dickinson) using FL1-H filter A total of 20000 events were acquired for eachsample and data were processed using WinMDI 29 soft-ware Images were also acquired with a Nikon Eclipse E600microscope equipped with a 60X 14 oil Plan-Apochromatobjective and a standard FITC filter set
3 Results and Discussion
31 Effect of Acetic Acid on the Localization of Active Ras inGlucose-Growing Cells In a recent study we showed that
Oxidative Medicine and Cellular Longevity 5
0
5
10
15
20
25
30
35
40A
nnex
in V
pos
(
)
Acetic acid
lowastlowast
lowastlowast
0 mM 40 mM 80 mM 120 mM
(a)
0
5
10
15
20
25
30
35
40
PI p
os (
)
Acetic acid
lowast
lowast
0 mM 40 mM 80 mM 120 mM
(b)
0
10
20
30
40
50
60
70
80
Acetic acid
Ann
exin
V+
PI p
os (
)
lowast
lowast
0 mM 40 mM 80 mM 120 mM
(c)
Figure 4 The occurrence of acetic-acid-induced cell death in hxk2Δ cells is characterized by markers of both apoptosis and necrosisAssessment of cell death by FITC-coupled annexinV and PI stainingW303-1A (black bars) and hxk2Δ (gray bars) exponentially growing cellswere treatedwith different concentrations (40ndash80ndash120mM) of acetic acid for 200minutes at 30∘C before being processed for determination ofphosphatidylserine externalization andmembrane integrity by flow cytometry 30000 events have been evaluatedThemeans of 3 independentexperiments with standard deviations are reported Studentrsquos 119905-test lowast119875 lt 005 and lowastlowast119875 lt 001
active Ras proteins accumulated mainly in the plasma mem-brane and in the nucleus in exponentially growing wild-typecells while they accumulated inmitochondria in cells deletedin the HXK2 gene indicating that this enzyme was involvedin the proper localization of these small GTPases [9] Therole played by active Ras in mitochondria is not knownalthough other reports have found Ras-GTP associated withthese organelles both inmammals and lower eukaryotes andhave linked it to cancer and apoptosis Leadsham et al [10]demonstrated that in S cerevisiae loss of Whi2p function aprotein known to influence cell cycle exit under conditions ofnutritional depletion determined an aberrant accumulationof activated Ras at the mitochondria In this mutant thefailure to shut down Ras signalling by addressing Ras to thevacuole would lead tomitochondrial dysfunction accumula-tion of damaging ROS and cell deathMore recently Hu et al[11] using a tetracycline inducible model demonstrated thatin mammalian cells association of K-rasG12V proteins withmitochondria induced mitochondrial dysfunction increasedROS accumulation and a metabolic switch from oxidativephosphorylation to glycolysis
To investigate whether a correlation exists between themitochondrial localization of active Ras and programmed
cell death we evaluated the effect of the addition of aceticacid a well-known apoptotic stimulus [25] to wild-typecells expressing the eGFP-RBD3 probe on the localizationof Ras-GTP proteins To this aim exponentially growingcells were collected by centrifugation resuspended in low pHmedium (pH 30) and pictures were taken at the fluorescencemicroscope before and after addition of 40mM acetic acidOur results showed that within 5 minutes this apoptoticstimulus caused the localization of active Ras proteins exclu-sively to the mitochondria reinforcing the hypothesis ofthe involvement of these proteins in programmed cell death(Figure 1) In wild-type cells resuspended in low pHmediumfor 5 minutes without acetic acid added active Ras proteinsaccumulated mainly in the plasma membrane and in thenucleus indicating that acidification of the medium did notinfluence the localization of these small G proteins (Figure 1)A mitochondrial localization of Ras-GTP was also observedwhen a low concentration of H
2O2was added to the medium
(data not shown) indicating that delocalization of active Rasto these organelles is a more general response to differentapoptotic stimuli and suggesting that mitochondrial localiza-tion of Ras2-GTP is actually important for the induction ofapoptosis in yeast
6 Oxidative Medicine and Cellular Longevity
W303-1A
256
0
Fluorescence
Even
ts
100
101
102
103
104
0 mM
40 mM
80 mM
120 mM
Fluorescence
Even
ts
256
010
010
110
210
310
4
0 mM
40 mM
80 mM
120 mM
hxk2
(a)
W303-1A
0 mM 80 mM
hxk2
(b)
Figure 5 Mitochondrial membrane potential and morphology in W303-1A and hxk2Δ strains upon acetic acid treatment W303-1A andhxk2Δ exponentially growing cells were treated with different concentrations (40ndash80ndash120mM) of acetic acid for 200 minutes at 30∘C DiOC
6
uptake was assessed using both flow cytometry (a) and fluorescence microscopy (b) Photomicrographs illustrate the alteration of the tubularmitochondrial network to clustered mitochondrial morphology particularly exacerbated in hxk2Δ cells after acetic acid treatment
32 Deletion of HXK2 Enhances Cell Death and IncreasesIntracellular ROS Levels The results presented previouslysuggest that localization of active Ras proteins to mitochon-dria might be involved in programmed cell death since theseproteins are localized to mitochondria following additionof acetic acid Since in glucose-growing hxk2Δ cells activeRas is constitutively located in mitochondria [9] we nextanalyzed the behavior of the hxk2Δmutant under conditions
that caused apoptosisWhile the growth of hxk2Δmutant wasalmost indistinguishable from the wild-type strain at pH 53it exhibited an increased sensitivity to acetic acid stress at lowpH (Figure 2(a)) suggesting that Hxk2 is required for normaltolerance to acetic acid treatment Cell survival of wild-typeand hxk2Δ cells after induction of apoptosis with acetic acidwas further tested in a plating assay After treatment withdifferent concentrations (80 and 120mM) of acetic acid for
Oxidative Medicine and Cellular Longevity 7
up to 200 minutes at 30∘C hxk2Δ cells showed a significantdose-dependent reduction in cell survival when comparedwith wild-type cells (Figure 2(b)) Taken together these datasuggest that Hxk2 is required for normal tolerance to aceticacid treatment
Since an aberrant accumulation of activated Ras in mito-chondria accompanied to accumulation of ROS has alreadybeen reported both in mammals and yeast [10 11] and sinceROS play a pivotal role in apoptotic cell death FACS analyseswere performed to evaluate the accumulation of ROS in wild-type and hxk2Δ cells treated with different concentrationsof acetic acid for 200min at 30∘C In particular DHR123was used to determine the accumulation of ROS in the cellssince this compound can easily permeate them and can bequantitatively oxidized to a green fluorescent product inthe presence of ROS The percentage of oxidized R123 wasabout the double for hxk2Δ cells compared to wild-type cells200min after treatment with either 40 or 80mM acetic acid(Figure 3) To substantiate the hypothesis that an aberrantaccumulation of activated Ras in mitochondria might leadtomitochondrial dysfunction and accumulation of damagingROS we expressed the dominant active RAS2Val19 allelewhich was reported to show a much higher level of Ras2-GTP [39] in hxk2Δ cells Our results showed that indeed theexpression of this allele in the hxk2Δ background caused afurther increase in the level of ROS both in growing cells andafter treatment with acetic acid (Figure 3)These data suggestthat conditions that presumably cause a highermitochondrialaccumulation of active Ras may contribute to accumulationof ROS and cell death
33 Loss of Hxk2 Causes an Increase of Both Apoptosis andNecrosis The results presented previously suggest that Hxk2might have an antiapoptotic activity To better characterizethe nature of cell death triggered by addition of acetic acid tohxk2Δ cells we quantified phenotypic changes indicative ofapoptosis While apoptotic DNA condensation was detectedby DAPI staining combined Annexin Vpropidium iodide(PI) staining was used to discriminate between early apop-totic (Annexin V+PIminus) late apoptoticsecondary necrotic(Annexin V+PI+) and necrotic (Annexin VminusPI+) deathsDAPI staining showed that nuclei of untreated cells wereround while the nuclear DNA was condensed in bothwild-type and hxk2Δ cells treated with 80mM acetic acidbeing the extent of this phenotype more pronounced for themutant strain compared to the wild-type strain (data notshown) Annexin Vpropidium iodide (PI) staining revealedthat hxk2Δ-facilitated cell death was accompanied by anincrease in both apoptotic and necroticmarkers (Figure 4) Inparticular untreated hxk2Δ cells showed a higher percentageof both early apoptotic and necrotic cells compared withthe wild-type strain which further increased upon treatmentwith acetic acid
34 Deletion of HXK2 Causes Hyperpolarization of Mitochon-dria Mitochondrial membrane potential (ΔΨ
119898) is a useful
indicator of mitochondrial function To measure ΔΨ119898 we
used the cationic lipophylic dye DiOC6 which accumulates
0102030405060708090
100110
DH
R 12
3 (
)
Acetic acid
lowast
lowast
lowast
0 mM 40 mM 80 mM 120 mM
Figure 6 Effect of YCA1 deletion on ROS accumulation aftertreatment with acetic acid W303-1A (black bars) yca1Δ (blackhatched bars) hxk2Δ (gray bars) and hxk2Δ-yca1Δ (white bars)exponentially growing cells were treated with different concen-trations (40ndash80ndash120mM) of acetic acid for 200 minutes at 30∘CROS accumulation was assayed by flow cytometry using the dyedihydrorhodamine 123 (DHR123) Studentrsquos 119905-test lowast119875 lt 005
in mitochondria in accordance with ΔΨ119898 Wild-type and
hxk2Δ cells were treatedwith different concentration of aceticacid (40ndash80ndash120mM) for 200min at 30∘C collected andstained with this dye FACS analysis showed that mitochon-drial membrane potential clearly increased in both wild-typeand hxk2Δ cells after treatment with acetic acid comparedwith untreated control cells with the hyperpolarizationof mitochondria being much stronger in the mutant cellscomparedwithwild-type cells (Figure 5(a)) In parallel mito-chondrial morphology was assessed by using DiOC
6 both
in wild-type and hxk2Δ cells At low concentration this dyespecifically stains the mitochondrial membranes in a mannerthat depends on membrane potential and can be observed byfluorescence microscopy Before treatment with acetic acidbothwild-type and hxk2Δ cells displayed a tubularmitochon-drial morphology indicating that these mitochondria werehealthy and possessed a membrane potential (Figure 5(b))By contrast the mitochondrial membranes present in bothwild-type and hxk2Δ cells appeared rounded and highlyfragmented after treatment with acetic acid (Figure 5(b))This conversion of mitochondrial morphology from tubu-lar to punctuate structures is likely to occur by excessivemitochondrial fission and has already been observed in yeastapoptosis induced by acetic acid treatment [40] ImportantlyDiOC
6staining was greatly increased and more pronounced
in hxk2Δ cells compared to wild-type cells after treatmentwith acetic acid confirming the increase of fluorescenceobserved in these mutant cells by FACS analysis Thesedata indicate that both in wild-type and hxk2Δ cells aceticacid treatment caused hyperpolarization of mitochondrialmembrane with consequent damage of these organelles andloss of functionality This effect was more pronounced inhxk2Δ cells compared to wild-type cells
35 Acetic-Acid-Induced Cell Death in the hxk2Δ Strain Is Yca1Independent At least two death pathways can be activated
8 Oxidative Medicine and Cellular Longevity
lowastlowast
lowastlowast
lowast
lowastlowast
0 mM 40 mM 80 mM 120 mMAcetic acid
0
5
10
15
20
25
30
35
40A
nnex
in V
pos
(
)
(a)
lowast
lowast
lowastlowast
lowastlowast
0 mM 40 mM 80 mM 120 mM0
10
20
30
40
50
60
70
80
PI p
os (
)
Acetic acid
(b)
lowastlowast
lowast
lowast
0 mM 40 mM 80 mM 120 mM0
10
20
30
40
50
60
70
80
Acetic acid
Ann
exin
V+
PI p
os (
)
(c)
Figure 7 Effect of YCA1 deletion on cell death after treatment with acetic acid W303-1A (black bars) yca1Δ (black hatched bars) hxk2Δ(gray bars) and hxk2Δ-yca1Δ (white bars) exponentially growing cells were treated with different concentrations (40ndash80ndash120mM) of aceticacid for 200 minutes at 30∘C Cell death was assessed by flow cytometry using FITC-coupled annexin V and PI co-staining to determinatethe externalization of phosphatidylserine and the membrane integrityThemeans of 3 independent experiments with standard deviations arereported Studentrsquos 119905-test lowast119875 lt 005 and lowastlowast119875 lt 001
in yeast acetic-acid-induced apoptosis one dependent oncytochromec release which requires YCA1 and the other(s)independent of it [16 30] Consequently we next investigatedwhether the yeast caspase Yca1 might be involved in acetic-acid-induced cell death in hxk2Δ cells In particular toassess whether Yca1 plays a role in acetic-acid-induced ROSaccumulation in hxk2Δ cells we treated hxk2Δ and hxk2Δyca1Δ cells with different concentration of acetic acid (40ndash80ndash120mM) for 200min at 30∘C and measured the ROSaccumulation Our results showed that deletion of YCA1 inhxk2Δ cells had no effect on acetic-acid-induced ROS accu-mulation (Figure 6) suggesting that the generation of ROSin hxk2Δ cells upon acetic acid stress is Yca1 independentHowever the level of intracellular ROS was consistentlyhigher in the hxk2Δ yca1Δ doublemutant growing on glucosemedium compared with the hxk2Δ mutant Similarly Du etal [24] showed that formic acid induced Yca1-independentapoptosis-like cell death and that the burst of ROS duringcell death occurred earlier and at a higher level in the yca1Δstrain than in the wild-type strain Moreover Khan et al [41]determined the level of oxidized proteins in yeast cells underH2O2stress and showed that lack of Yca1 abrogated apoptosis
but elevated intracellular oxidized proteins compared with
wild-type cells Consequently both our results and data in theliterature [24 41] clearly suggest a linkage between ROS pro-duction and Yca1 activation during apoptosis in yeast Finallycytofluorometric quantification of phosphatidylserine exter-nalization andormembrane permeabilization (annexinVPIcostaining) further confirmed that Yca1p did not influencethe acetic-acid-induced cell death in hxk2Δ cells (Figure 7)These results strongly suggest that apoptosis induced by aceticacid in hxk2Δ cells is largely Yca1 independent
4 Conclusion
Several studies indicate that the Ras proteinmay be physicallyassociated with mitochondria both in yeast and mammals[10 11 42ndash44] This study provides results indicating thatassociation of active Ras to these organelles might be linkedto apoptosis First of all we show that addition of acetic acida well-known apoptotic stimulus in S cerevisiae to growingwild-type cells determined a delocalization of active Ras pro-teins from nuclei and plasma membrane to mitochondriaFurthermore we show that addition of acetic acid to hxk2Δcells showing a constitutive localization of active Ras inmito-chondria [9] enhanced ROS production and cell death
Oxidative Medicine and Cellular Longevity 9
compared with wild-type cells Moreover expression of thedominant active RAS2Val19 allele in the hxk2Δ backgroundcaused an even higher increase in the level of ROS both ingrowing cells and after treatment with acetic acid providinga further proof to the hypothesis that conditions that presum-ably cause a higher mitochondrial accumulation of activeRas may contribute to accumulation of ROS and cell deathWe also show that in hxk2Δ cells addition of acetic acidincreased cell death compared with the wild-type strain withthe typical markers of apoptosis highlighting a new rolefor Hexokinase 2 as an antiapoptotic factor in S cerevisiaecells However at this stage we cannot exclude a correlationbetween the well-known functions of Hxk2 in glucose repres-sion and signalling and the role of Ras in inducing apoptosis
Finally in this study we show that lack of Hxk2 inducesapoptosis via a mitochondria-mediated pathway withoutmetacaspase Yca1 involvement since deletion of YCA1 in thehxk2Δ background did not abrogate the acetic-acid-inducedaccumulation of ROS and did not decrease the percentage ofboth apoptotic and necrotic cells
Acknowledgments
The authors thank J Winderickx University of Leuven Bel-gium for the gift of strains and M Vai Universita Milano-Bicocca Milan for the gift of primers This work was sup-ported by FAR grant to Sonia Colombo and by founding ofProgram SysBioNet Italian Roadmap Research Infrastruc-ture 2012
References
[1] S JonesM-L Vignais and J R Broach ldquoTheCDC25 protein ofSaccharomyces cerevisiae promotes exchange of guanine nucle-otides bound to RasrdquoMolecular and Cellular Biology vol 11 no5 pp 2641ndash2646 1991
[2] S Rudoni S Colombo P Coccetti and E Martegani ldquoRole ofguanine nucleotides in the regulation of the RascAMPpathwayin Saccharomyces cerevisiaerdquo Biochimica et Biophysica Acta vol1538 no 2-3 pp 181ndash189 2001
[3] K TanakaMNakafuku T Satoh et al ldquoS cerevisiae genes IRA1and IRA2 encode proteins that may be functionally equivalentto mammalian ras GTPase activating proteinrdquo Cell vol 60 no5 pp 803ndash807 1990
[4] S Zaman S I Lippman X Zhao and J R Broach ldquoHow Sac-charomyces responds to nutrientsrdquo Annual Review of Geneticsvol 42 pp 27ndash81 2008
[5] JMThevelein and J H DeWinde ldquoNovel sensingmechanismsand targets for the cAMP-protein kinase A pathway in the yeastSaccharomyces cerevisiaerdquoMolecularMicrobiology vol 33 no 5pp 904ndash918 1999
[6] F Rolland JWinderickx and JMThevelein ldquoGlucose-sensingand -signallingmechanisms in yeastrdquo FEMS Yeast Research vol2 no 2 pp 183ndash201 2002
[7] G F Casperson N Walker and H R Bourne ldquoIsolation of thegene encoding adenylate cyclase in Saccharomyces cerevisiaerdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 82 no 15 pp 5060ndash5063 1985
[8] B Smets R Ghillebert P De Snijder et al ldquoLife in the midstof scarcity adaptations to nutrient availability in Saccharomycescerevisiaerdquo Current Genetics vol 56 no 1 pp 1ndash32 2010
[9] S Broggi E Martegani and S Colombo ldquoLive-cell imaging ofendogenous Ras-GTP shows predominant Ras activation at theplasma membrane and in the nucleus in Saccharomyces cere-visiaerdquoThe International Journal of Biochemistry amp Cell Biologyvol 45 no 2 pp 384ndash394 2013
[10] J E Leadsham K Miller K R Ayscough et al ldquoWhi2p linksnutritional sensing to actin-dependent Ras-cAMP-PKA regu-lation and apoptosis in yeastrdquo Journal of Cell Science vol 122no 5 pp 706ndash715 2009
[11] YHuW LuG Chen et al ldquoK-rasG12V transformation leads tomitochondrial dysfunction and a metabolic switch from oxida-tive phosphorylation to glycolysisrdquo Cell Research vol 22 no 2pp 399ndash412 2012
[12] S Buttner T Eisenberg D Carmona-Gutierrez et al ldquoEndonu-clease G regulates budding yeast life and deathrdquoMolecular Cellvol 25 no 2 pp 233ndash246 2007
[13] F Madeo E Frohlich and K-U Frohlich ldquoA yeast mutantshowing diagnostic markers of early and late apoptosisrdquo Journalof Cell Biology vol 139 no 3 pp 729ndash734 1997
[14] FMadeo E Herker SWissing H Jungwirth T Eisenberg andK-U Frohlich ldquoApoptosis in yeastrdquo Current Opinion in Micro-biology vol 7 no 6 pp 655ndash660 2004
[15] F Madeo E Herker C Maldener et al ldquoA caspase-related pro-tease regulates apoptosis in yeastrdquo Molecular Cell vol 9 no 4pp 911ndash917 2002
[16] N Guaragnella C Pereira M J Sousa et al ldquoYCA1 participatesin the acetic acid induced yeast programmed cell death also in amanner unrelated to its caspase-like activityrdquo FEBS Letters vol580 no 30 pp 6880ndash6884 2006
[17] W Li L Sun Q Liang J Wang W Mo and B Zhou ldquoYeastAMID homologue Ndi1p displays respiration-restricted apop-totic activity and is involved in chronological agingrdquoMolecularBiology of the Cell vol 17 no 4 pp 1802ndash1811 2006
[18] S Wissing P Ludovico E Herker et al ldquoAn AIF orthologueregulates apoptosis in yeastrdquo Journal of Cell Biology vol 166 no7 pp 969ndash974 2004
[19] B Fahrenkrog U Sauder and U Aebi ldquoThe S cerevisiae HtrA-like protein Nma111p is a nuclear serine protease that mediatesyeast apoptosisrdquo Journal of Cell Science vol 117 no 1 pp 115ndash126 2004
[20] J Qiu J-H Yoon and B Shen ldquoSearch for apoptotic nucleasesin yeast role of Tat-D nuclease in apoptotic DNA degradationrdquoThe Journal of Biological Chemistry vol 280 no 15 pp 15370ndash15379 2005
[21] I Orlandi M Bettiga L Alberghina and M Vai ldquoTranscrip-tional profiling of ubp10 null mutant reveals altered subtelo-meric gene expression and insurgence of oxidative stressresponserdquoThe Journal of Biological Chemistry vol 279 no 8 pp6414ndash6425 2004
[22] M Ligr F Madeo E Frohlich W Hilt K-U Frohlich and DH Wolf ldquoMammalian Bax triggers apoptotic changes in yeastrdquoFEBS Letters vol 438 no 1-2 pp 61ndash65 1998
[23] F Madeo E Frohlich M Ligr et al ldquoOxygen stress a regulatorof apoptosis in yeastrdquo Journal of Cell Biology vol 145 no 4 pp757ndash767 1999
[24] L Du Y Su D Sun et al ldquoFormic acid induces Yca1p-inde-pendent apoptosis-like cell death in the yeast Saccharomycescerevisiaerdquo FEMS Yeast Research vol 8 no 4 pp 531ndash539 2008
10 Oxidative Medicine and Cellular Longevity
[25] P LudovicoM J SousaM T Silva C Leao andMCorte-RealldquoSaccharomyces cerevisiae commits to a programmed cell deathprocess in response to acetic acidrdquoMicrobiology vol 147 no 9pp 2409ndash2415 2001
[26] P Ludovico F Rodrigues AAlmeidaMT Silva A Barrientosand M Corte-Real ldquoCytochrome c release and mitochondriainvolvement in programmed cell death induced by acetic acidin Saccharomyces cerevisiaerdquo Molecular Biology of the Cell vol13 no 8 pp 2598ndash2606 2002
[27] N Guaragnella L Antonacci S Passarella E Marra and SGiannattasio ldquoHydrogen peroxide and superoxide anion pro-duction during acetic acid-induced yeast programmed celldeathrdquo Folia Microbiologica vol 52 no 3 pp 237ndash240 2007
[28] S Giannattasio A Atlante L Antonacci et al ldquoCytochrome c isreleased from coupled mitochondria of yeast en route to aceticacid-induced programmed cell death and can work as an elec-tron donor and a ROS scavengerrdquo FEBS Letters vol 582 no 10pp 1519ndash1525 2008
[29] N Guaragnella L Antonacci S Giannattasio E Marra and SPassarella ldquoCatalase T and Cu Zn-superoxide dismutase in theacetic acid-induced programmed cell death in Saccharomycescerevisiaerdquo FEBS Letters vol 582 no 2 pp 210ndash214 2008
[30] N Guaragnella A Bobba S Passarella E Marra and S Gian-nattasio ldquoYeast acetic acid-induced programmed cell death canoccurwithout cytochrome c releasewhich requiresmetacaspaseYCA1rdquo FEBS Letters vol 584 no 1 pp 224ndash228 2010
[31] F Madeo D Carmona-Gutierrez J Ring S Buttner T Eisen-berg and G Kroemer ldquoCaspase-dependent and caspase-inde-pendent cell death pathways in yeastrdquo Biochemical and Biophys-ical Research Communications vol 382 no 2 pp 227ndash231 2009
[32] DAhuatzi A Riera R Pelaez PHerrero andFMoreno ldquoHxk2regulates the phosphorylation state of Mig1 and therefore itsnucleocytoplasmic distributionrdquo The Journal of BiologicalChemistry vol 282 no 7 pp 4485ndash4493 2007
[33] FMoreno D Ahuatzi A Riera C A Palomino and P HerreroldquoGlucose sensing through the Hxk2-dependent signalling path-wayrdquo Biochemical Society Transactions vol 33 no 1 pp 265ndash268 2005
[34] B JThomas and R Rothstein ldquoElevated recombination rates intranscriptionally active DNArdquo Cell vol 56 no 4 pp 619ndash6301989
[35] L C Robinson J B Gibbs M S Marshall I S Sigal and KTatchell ldquoCDC25 a component of the RAS-adenylate cyclasepathway in Saccharomyces cerevisiaerdquo Science vol 235 no 4793pp 1218ndash1221 1987
[36] R J Rothstein ldquoOne-step gene disruption in yeastrdquoMethods inEnzymology vol 101 pp 202ndash211 1983
[37] M Bettiga L Calzari I Orlandi L Alberghina and M VaildquoInvolvement of the yeast metacaspase Yca1 in ubp10Δ-pro-grammed cell deathrdquo FEMS Yeast Research vol 5 no 2 pp 141ndash147 2004
[38] N Casatta A Porro I Orlandi L Brambilla andM Vai ldquoLackof Sir2 increases acetate consumption and decreases extracellu-lar pro-aging factorsrdquo Biochimica et Biophysica Acta vol 1833no 3 pp 593ndash601 2013
[39] S Colombo D Ronchetti J M Thevelein J Winderickx andE Martegani ldquoActivation state of the Ras2 protein and glucose-induced signaling in Saccharomyces cerevisiaerdquo The Journal ofBiological Chemistry vol 279 no 45 pp 46715ndash46722 2004
[40] P Fabrizio and V D Longo ldquoChronological aging-inducedapoptosis in yeastrdquo Biochimica et Biophysica Acta vol 1783 no7 pp 1280ndash1285 2008
[41] M A S Khan P B Chock and E R Stadtman ldquoKnockout ofcaspase-like gene YCA1 abrogates apoptosis and elevates oxi-dized proteins in Saccharomyces cerevisiaerdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 102 no 48 pp 17326ndash17331 2005
[42] J CWolfman S M Planchon J Liao and AWolfman ldquoStruc-tural and functional consequences of c-N-Ras constitutivelyassociated with intact mitochondriardquo Biochimica et BiophysicaActa vol 1763 no 10 pp 1108ndash1124 2006
[43] T G Bivona S E Quatela B O Bodemann et al ldquoPKC regu-lates a farnesyl-electrostatic switch on K-Ras that promotes itsassociation with Bcl-XL on mitochondria and induces apopto-sisrdquoMolecular Cell vol 21 no 4 pp 481ndash493 2006
[44] A Rebollo D Perez-Sala and C Martınez-A ldquoBcl-2 differen-tially targets K- N- and H-Ras to mitochondria in IL-2 sup-plemented or deprived cells implications in prevention of apop-tosisrdquo Oncogene vol 18 no 35 pp 4930ndash4939 1999
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Oxidative Medicine and Cellular Longevity 3
SD pH 53
W303-1A
hxk2
W303-1A
hxk2
W303-1A
hxk2
W303-1A
hxk2
SD pH 30
NT
40 mM
80 mM
120 mM
(a)
0
20
40
60
80
100
120
140
0 50 100 150 2000
20
40
60
80
100
120
140
0 50 100 150 200Time (min)
W303-1A
Surv
ival
cells
()
Surv
ival
cells
()
Time (min)
hxk2
(b)
Figure 2 hxk2Δ cells exhibit inhibition of cell growth and hypersensitivity to acetic acid (a) W303-1A and hxk2Δ cells were harvested andresuspended (1-2times 107 cellsmL) in SD medium adjusted either at pH 53 or at pH 30 (set with HCl) in the absence (NT) or in the presenceof 40 80 or 120mM acetic acid Cells were incubated for 200min at 30∘C with shaking (160 rpm) After treatment cells were harvestedand resuspended at the same concentration (108 cellsmL) in milliQ water 5 microliter from a concentrated suspension and from 10-folddilutions of each culture was spotted onto YEPD plates and incubated at 30∘C for 3 days (b) Cell survival ofW303-1A and hxk2Δ strains Cellviability of W303-1A and hxk2Δ untreated cells (998771) or treated with 80mM (◼) and 120mM (e) acetic acid was analyzed at indicated times bymeasuring colony-forming units (cfu) after 2 days of growth at 30∘C Cell survival (100) corresponds to the cfu at time zero The means of4 independent experiments with standard deviations are reported
4 Oxidative Medicine and Cellular Longevity
40 120583L of cells suspension was seeded on concanavalin A(Sigma-Aldrich Milano Italy) coated cover glass for 10min(100 120583gmL) The cover glass was washed 4 times using theproper medium and put on top of a Thoma chamber Ima-ges were acquired with a Nikon Eclipse E600 microscopeequipped with a 60X 14 oil Plan-Apochromat objective anda standard FITC filter set for GFP fluorescence Images wererecorded digitally using a Leica DC 350F camera and pro-cessed using Adobe Photoshop (Adobe Systems Inc)
24 Acetic Acid Sensitivity This assay was performed essen-tially as described by Casatta et al [38] Exponential-phasecells were harvested and resuspended (107 cellsmL) in SDmedium adjusted either at pH 53 or at pH 30 (set with HCl)and containing 0 40 80 or 120mM acetic acid Cells wereincubated for 200min at 30∘C with shaking (160 rpm) Aftertreatment cells (10-fold serial dilutions) were spotted ontoYEPD plates and incubated at 30∘C for 3 days
25 Viability Assay Cells were grown in SDmedium at 30∘Cuntil exponential phase and treated with acetic acid asdescribed previously At different times (0 30 60 and 200minutes) during acetic acid treatment cell number was cal-culated and 400 cells were plated Viability was determined bymeasuring colony-forming units (cfu) after 2 days of growthon YEPD agar plates at 30∘C The percentage of viable cellsresulted in dividing the number of surviving colonies of thetreated sample by the number of surviving colonies of thesame culture before acid acetic addition
26 Dihydrorhodamine 123 (DHR123) Staining ROS (reac-tive oxygen species) were detected with DHR123 (SigmaAldrich) essentially as described by Madeo et al [23] Cellswere grown in SD medium at 30∘C until exponential phaseand treated with acetic acid as described previously DHR123was added directly to the culture medium at the final con-centration of 5120583gmL from a 25120583g120583L stock solution After2 hours of incubation cells were diluted to 106 cellsmL andanalyzed using a FACScan instrument (Becton Dickinson)at low flow rate with excitation and emission settings of 488and 525ndash550 nm (filter FL 1) A total of 20000 events wereacquired for each sample and data were processed usingWinMDI 29 software
27 410158406-Diamidino-2-phenylindole (DAPI) Staining Cellswere fixed with 37 formaldehyde for 30 minutes stainedwith 2120583gmL of DAPI for 10 minutes washed with distilledwater and resuspended in 50 glycerol solution Images wereacquired with a Nikon Eclipse E600 fluorescence microscopeusing a DAPI filter recorded digitally using a Leica DC 350Fcamera and processed using Adobe Photoshop (Adobe Sys-tems Inc)
28 Annexin V and Propidium Iodide (PI) Staining (FITC)conjugated recombinant Annexin V (Immuno Tools) wasused for the detection of phosphatidylserine exposed in themembrane of apoptotic cells Cells were harvested after 200minutes of acetic acid treatment as reported previously
0102030405060708090
100
Acetic acid
DH
R123
()
lowast
lowast
lowast
lowast
lowastlowast
0 mM 40 mM 80 mM 120 mM
Figure 3 ROS accumulation in W303-1A hxk2Δ and hxk2Δ cellsexpressing the activated Ras2Val19 allele after treatment with aceticacid W303-1A (black bars) hxk2Δ (gray bars) and hxk2Δ cellsexpressing the activated Ras2Val19 allele (white bars) exponentiallygrowing cells were treated with different concentrations (40ndash80ndash120mM) of acetic acid for 200 minutes at 30∘C ROS accumulationwas assayed using the dye dihydrorhodamine 123 (DHR123) by flowcytometry The means of 3 independent experiments with standarddeviations are reported Studentrsquos 119905-test lowast119875 lt 005 and lowastlowast119875 lt 001
washed with sorbitol buffer (1M sorbitol 01M NaH2PO4
pH 80) and the cell wall was digested with Zymolyase 20T(Seikagaku Biobusiness Corporation) for about 35 minutes at37∘C Cells were then washed two times with binding buffer(10mM HepesNaOH pH 74 140mM NaCl 25mM CaCl
2
12M sorbitol) Spheroplasts were resuspended in 35 120583L ofbinding buffer and incubated with 25 120583L of Annexin V and2 120583L of a PI (Fluka) working solution (50120583gmL) for 15minutes in the dark at room temperature After staining thesamples were resuspended in binding buffer and analyzedusing a FACScan instrument (Becton Dickinson) using FL1-H filter on 119909-axis and FL 2-H filter on 119910-axis A total of30000 events were acquired for each sample and data wereprocessed using WinMDI 29 software
29 331015840-Dihexyloxacarbocyanine Iodide (DiOC6) StainingThe mitochondrial morphology and membrane potentialwere assessed by staining with DiOC
6(Molecular Probes
Invitrogen) Cells were grown in SD medium at 30∘C untilexponential phase and treated with acetic acid as describedpreviously DiOC6 was added directly to the culture mediumat the final concentration of 175 nM for 15 minutes in the darkat room temperature After staining the cells were analyzedusing a FACScan instrument (Becton Dickinson) using FL1-H filter A total of 20000 events were acquired for eachsample and data were processed using WinMDI 29 soft-ware Images were also acquired with a Nikon Eclipse E600microscope equipped with a 60X 14 oil Plan-Apochromatobjective and a standard FITC filter set
3 Results and Discussion
31 Effect of Acetic Acid on the Localization of Active Ras inGlucose-Growing Cells In a recent study we showed that
Oxidative Medicine and Cellular Longevity 5
0
5
10
15
20
25
30
35
40A
nnex
in V
pos
(
)
Acetic acid
lowastlowast
lowastlowast
0 mM 40 mM 80 mM 120 mM
(a)
0
5
10
15
20
25
30
35
40
PI p
os (
)
Acetic acid
lowast
lowast
0 mM 40 mM 80 mM 120 mM
(b)
0
10
20
30
40
50
60
70
80
Acetic acid
Ann
exin
V+
PI p
os (
)
lowast
lowast
0 mM 40 mM 80 mM 120 mM
(c)
Figure 4 The occurrence of acetic-acid-induced cell death in hxk2Δ cells is characterized by markers of both apoptosis and necrosisAssessment of cell death by FITC-coupled annexinV and PI stainingW303-1A (black bars) and hxk2Δ (gray bars) exponentially growing cellswere treatedwith different concentrations (40ndash80ndash120mM) of acetic acid for 200minutes at 30∘C before being processed for determination ofphosphatidylserine externalization andmembrane integrity by flow cytometry 30000 events have been evaluatedThemeans of 3 independentexperiments with standard deviations are reported Studentrsquos 119905-test lowast119875 lt 005 and lowastlowast119875 lt 001
active Ras proteins accumulated mainly in the plasma mem-brane and in the nucleus in exponentially growing wild-typecells while they accumulated inmitochondria in cells deletedin the HXK2 gene indicating that this enzyme was involvedin the proper localization of these small GTPases [9] Therole played by active Ras in mitochondria is not knownalthough other reports have found Ras-GTP associated withthese organelles both inmammals and lower eukaryotes andhave linked it to cancer and apoptosis Leadsham et al [10]demonstrated that in S cerevisiae loss of Whi2p function aprotein known to influence cell cycle exit under conditions ofnutritional depletion determined an aberrant accumulationof activated Ras at the mitochondria In this mutant thefailure to shut down Ras signalling by addressing Ras to thevacuole would lead tomitochondrial dysfunction accumula-tion of damaging ROS and cell deathMore recently Hu et al[11] using a tetracycline inducible model demonstrated thatin mammalian cells association of K-rasG12V proteins withmitochondria induced mitochondrial dysfunction increasedROS accumulation and a metabolic switch from oxidativephosphorylation to glycolysis
To investigate whether a correlation exists between themitochondrial localization of active Ras and programmed
cell death we evaluated the effect of the addition of aceticacid a well-known apoptotic stimulus [25] to wild-typecells expressing the eGFP-RBD3 probe on the localizationof Ras-GTP proteins To this aim exponentially growingcells were collected by centrifugation resuspended in low pHmedium (pH 30) and pictures were taken at the fluorescencemicroscope before and after addition of 40mM acetic acidOur results showed that within 5 minutes this apoptoticstimulus caused the localization of active Ras proteins exclu-sively to the mitochondria reinforcing the hypothesis ofthe involvement of these proteins in programmed cell death(Figure 1) In wild-type cells resuspended in low pHmediumfor 5 minutes without acetic acid added active Ras proteinsaccumulated mainly in the plasma membrane and in thenucleus indicating that acidification of the medium did notinfluence the localization of these small G proteins (Figure 1)A mitochondrial localization of Ras-GTP was also observedwhen a low concentration of H
2O2was added to the medium
(data not shown) indicating that delocalization of active Rasto these organelles is a more general response to differentapoptotic stimuli and suggesting that mitochondrial localiza-tion of Ras2-GTP is actually important for the induction ofapoptosis in yeast
6 Oxidative Medicine and Cellular Longevity
W303-1A
256
0
Fluorescence
Even
ts
100
101
102
103
104
0 mM
40 mM
80 mM
120 mM
Fluorescence
Even
ts
256
010
010
110
210
310
4
0 mM
40 mM
80 mM
120 mM
hxk2
(a)
W303-1A
0 mM 80 mM
hxk2
(b)
Figure 5 Mitochondrial membrane potential and morphology in W303-1A and hxk2Δ strains upon acetic acid treatment W303-1A andhxk2Δ exponentially growing cells were treated with different concentrations (40ndash80ndash120mM) of acetic acid for 200 minutes at 30∘C DiOC
6
uptake was assessed using both flow cytometry (a) and fluorescence microscopy (b) Photomicrographs illustrate the alteration of the tubularmitochondrial network to clustered mitochondrial morphology particularly exacerbated in hxk2Δ cells after acetic acid treatment
32 Deletion of HXK2 Enhances Cell Death and IncreasesIntracellular ROS Levels The results presented previouslysuggest that localization of active Ras proteins to mitochon-dria might be involved in programmed cell death since theseproteins are localized to mitochondria following additionof acetic acid Since in glucose-growing hxk2Δ cells activeRas is constitutively located in mitochondria [9] we nextanalyzed the behavior of the hxk2Δmutant under conditions
that caused apoptosisWhile the growth of hxk2Δmutant wasalmost indistinguishable from the wild-type strain at pH 53it exhibited an increased sensitivity to acetic acid stress at lowpH (Figure 2(a)) suggesting that Hxk2 is required for normaltolerance to acetic acid treatment Cell survival of wild-typeand hxk2Δ cells after induction of apoptosis with acetic acidwas further tested in a plating assay After treatment withdifferent concentrations (80 and 120mM) of acetic acid for
Oxidative Medicine and Cellular Longevity 7
up to 200 minutes at 30∘C hxk2Δ cells showed a significantdose-dependent reduction in cell survival when comparedwith wild-type cells (Figure 2(b)) Taken together these datasuggest that Hxk2 is required for normal tolerance to aceticacid treatment
Since an aberrant accumulation of activated Ras in mito-chondria accompanied to accumulation of ROS has alreadybeen reported both in mammals and yeast [10 11] and sinceROS play a pivotal role in apoptotic cell death FACS analyseswere performed to evaluate the accumulation of ROS in wild-type and hxk2Δ cells treated with different concentrationsof acetic acid for 200min at 30∘C In particular DHR123was used to determine the accumulation of ROS in the cellssince this compound can easily permeate them and can bequantitatively oxidized to a green fluorescent product inthe presence of ROS The percentage of oxidized R123 wasabout the double for hxk2Δ cells compared to wild-type cells200min after treatment with either 40 or 80mM acetic acid(Figure 3) To substantiate the hypothesis that an aberrantaccumulation of activated Ras in mitochondria might leadtomitochondrial dysfunction and accumulation of damagingROS we expressed the dominant active RAS2Val19 allelewhich was reported to show a much higher level of Ras2-GTP [39] in hxk2Δ cells Our results showed that indeed theexpression of this allele in the hxk2Δ background caused afurther increase in the level of ROS both in growing cells andafter treatment with acetic acid (Figure 3)These data suggestthat conditions that presumably cause a highermitochondrialaccumulation of active Ras may contribute to accumulationof ROS and cell death
33 Loss of Hxk2 Causes an Increase of Both Apoptosis andNecrosis The results presented previously suggest that Hxk2might have an antiapoptotic activity To better characterizethe nature of cell death triggered by addition of acetic acid tohxk2Δ cells we quantified phenotypic changes indicative ofapoptosis While apoptotic DNA condensation was detectedby DAPI staining combined Annexin Vpropidium iodide(PI) staining was used to discriminate between early apop-totic (Annexin V+PIminus) late apoptoticsecondary necrotic(Annexin V+PI+) and necrotic (Annexin VminusPI+) deathsDAPI staining showed that nuclei of untreated cells wereround while the nuclear DNA was condensed in bothwild-type and hxk2Δ cells treated with 80mM acetic acidbeing the extent of this phenotype more pronounced for themutant strain compared to the wild-type strain (data notshown) Annexin Vpropidium iodide (PI) staining revealedthat hxk2Δ-facilitated cell death was accompanied by anincrease in both apoptotic and necroticmarkers (Figure 4) Inparticular untreated hxk2Δ cells showed a higher percentageof both early apoptotic and necrotic cells compared withthe wild-type strain which further increased upon treatmentwith acetic acid
34 Deletion of HXK2 Causes Hyperpolarization of Mitochon-dria Mitochondrial membrane potential (ΔΨ
119898) is a useful
indicator of mitochondrial function To measure ΔΨ119898 we
used the cationic lipophylic dye DiOC6 which accumulates
0102030405060708090
100110
DH
R 12
3 (
)
Acetic acid
lowast
lowast
lowast
0 mM 40 mM 80 mM 120 mM
Figure 6 Effect of YCA1 deletion on ROS accumulation aftertreatment with acetic acid W303-1A (black bars) yca1Δ (blackhatched bars) hxk2Δ (gray bars) and hxk2Δ-yca1Δ (white bars)exponentially growing cells were treated with different concen-trations (40ndash80ndash120mM) of acetic acid for 200 minutes at 30∘CROS accumulation was assayed by flow cytometry using the dyedihydrorhodamine 123 (DHR123) Studentrsquos 119905-test lowast119875 lt 005
in mitochondria in accordance with ΔΨ119898 Wild-type and
hxk2Δ cells were treatedwith different concentration of aceticacid (40ndash80ndash120mM) for 200min at 30∘C collected andstained with this dye FACS analysis showed that mitochon-drial membrane potential clearly increased in both wild-typeand hxk2Δ cells after treatment with acetic acid comparedwith untreated control cells with the hyperpolarizationof mitochondria being much stronger in the mutant cellscomparedwithwild-type cells (Figure 5(a)) In parallel mito-chondrial morphology was assessed by using DiOC
6 both
in wild-type and hxk2Δ cells At low concentration this dyespecifically stains the mitochondrial membranes in a mannerthat depends on membrane potential and can be observed byfluorescence microscopy Before treatment with acetic acidbothwild-type and hxk2Δ cells displayed a tubularmitochon-drial morphology indicating that these mitochondria werehealthy and possessed a membrane potential (Figure 5(b))By contrast the mitochondrial membranes present in bothwild-type and hxk2Δ cells appeared rounded and highlyfragmented after treatment with acetic acid (Figure 5(b))This conversion of mitochondrial morphology from tubu-lar to punctuate structures is likely to occur by excessivemitochondrial fission and has already been observed in yeastapoptosis induced by acetic acid treatment [40] ImportantlyDiOC
6staining was greatly increased and more pronounced
in hxk2Δ cells compared to wild-type cells after treatmentwith acetic acid confirming the increase of fluorescenceobserved in these mutant cells by FACS analysis Thesedata indicate that both in wild-type and hxk2Δ cells aceticacid treatment caused hyperpolarization of mitochondrialmembrane with consequent damage of these organelles andloss of functionality This effect was more pronounced inhxk2Δ cells compared to wild-type cells
35 Acetic-Acid-Induced Cell Death in the hxk2Δ Strain Is Yca1Independent At least two death pathways can be activated
8 Oxidative Medicine and Cellular Longevity
lowastlowast
lowastlowast
lowast
lowastlowast
0 mM 40 mM 80 mM 120 mMAcetic acid
0
5
10
15
20
25
30
35
40A
nnex
in V
pos
(
)
(a)
lowast
lowast
lowastlowast
lowastlowast
0 mM 40 mM 80 mM 120 mM0
10
20
30
40
50
60
70
80
PI p
os (
)
Acetic acid
(b)
lowastlowast
lowast
lowast
0 mM 40 mM 80 mM 120 mM0
10
20
30
40
50
60
70
80
Acetic acid
Ann
exin
V+
PI p
os (
)
(c)
Figure 7 Effect of YCA1 deletion on cell death after treatment with acetic acid W303-1A (black bars) yca1Δ (black hatched bars) hxk2Δ(gray bars) and hxk2Δ-yca1Δ (white bars) exponentially growing cells were treated with different concentrations (40ndash80ndash120mM) of aceticacid for 200 minutes at 30∘C Cell death was assessed by flow cytometry using FITC-coupled annexin V and PI co-staining to determinatethe externalization of phosphatidylserine and the membrane integrityThemeans of 3 independent experiments with standard deviations arereported Studentrsquos 119905-test lowast119875 lt 005 and lowastlowast119875 lt 001
in yeast acetic-acid-induced apoptosis one dependent oncytochromec release which requires YCA1 and the other(s)independent of it [16 30] Consequently we next investigatedwhether the yeast caspase Yca1 might be involved in acetic-acid-induced cell death in hxk2Δ cells In particular toassess whether Yca1 plays a role in acetic-acid-induced ROSaccumulation in hxk2Δ cells we treated hxk2Δ and hxk2Δyca1Δ cells with different concentration of acetic acid (40ndash80ndash120mM) for 200min at 30∘C and measured the ROSaccumulation Our results showed that deletion of YCA1 inhxk2Δ cells had no effect on acetic-acid-induced ROS accu-mulation (Figure 6) suggesting that the generation of ROSin hxk2Δ cells upon acetic acid stress is Yca1 independentHowever the level of intracellular ROS was consistentlyhigher in the hxk2Δ yca1Δ doublemutant growing on glucosemedium compared with the hxk2Δ mutant Similarly Du etal [24] showed that formic acid induced Yca1-independentapoptosis-like cell death and that the burst of ROS duringcell death occurred earlier and at a higher level in the yca1Δstrain than in the wild-type strain Moreover Khan et al [41]determined the level of oxidized proteins in yeast cells underH2O2stress and showed that lack of Yca1 abrogated apoptosis
but elevated intracellular oxidized proteins compared with
wild-type cells Consequently both our results and data in theliterature [24 41] clearly suggest a linkage between ROS pro-duction and Yca1 activation during apoptosis in yeast Finallycytofluorometric quantification of phosphatidylserine exter-nalization andormembrane permeabilization (annexinVPIcostaining) further confirmed that Yca1p did not influencethe acetic-acid-induced cell death in hxk2Δ cells (Figure 7)These results strongly suggest that apoptosis induced by aceticacid in hxk2Δ cells is largely Yca1 independent
4 Conclusion
Several studies indicate that the Ras proteinmay be physicallyassociated with mitochondria both in yeast and mammals[10 11 42ndash44] This study provides results indicating thatassociation of active Ras to these organelles might be linkedto apoptosis First of all we show that addition of acetic acida well-known apoptotic stimulus in S cerevisiae to growingwild-type cells determined a delocalization of active Ras pro-teins from nuclei and plasma membrane to mitochondriaFurthermore we show that addition of acetic acid to hxk2Δcells showing a constitutive localization of active Ras inmito-chondria [9] enhanced ROS production and cell death
Oxidative Medicine and Cellular Longevity 9
compared with wild-type cells Moreover expression of thedominant active RAS2Val19 allele in the hxk2Δ backgroundcaused an even higher increase in the level of ROS both ingrowing cells and after treatment with acetic acid providinga further proof to the hypothesis that conditions that presum-ably cause a higher mitochondrial accumulation of activeRas may contribute to accumulation of ROS and cell deathWe also show that in hxk2Δ cells addition of acetic acidincreased cell death compared with the wild-type strain withthe typical markers of apoptosis highlighting a new rolefor Hexokinase 2 as an antiapoptotic factor in S cerevisiaecells However at this stage we cannot exclude a correlationbetween the well-known functions of Hxk2 in glucose repres-sion and signalling and the role of Ras in inducing apoptosis
Finally in this study we show that lack of Hxk2 inducesapoptosis via a mitochondria-mediated pathway withoutmetacaspase Yca1 involvement since deletion of YCA1 in thehxk2Δ background did not abrogate the acetic-acid-inducedaccumulation of ROS and did not decrease the percentage ofboth apoptotic and necrotic cells
Acknowledgments
The authors thank J Winderickx University of Leuven Bel-gium for the gift of strains and M Vai Universita Milano-Bicocca Milan for the gift of primers This work was sup-ported by FAR grant to Sonia Colombo and by founding ofProgram SysBioNet Italian Roadmap Research Infrastruc-ture 2012
References
[1] S JonesM-L Vignais and J R Broach ldquoTheCDC25 protein ofSaccharomyces cerevisiae promotes exchange of guanine nucle-otides bound to RasrdquoMolecular and Cellular Biology vol 11 no5 pp 2641ndash2646 1991
[2] S Rudoni S Colombo P Coccetti and E Martegani ldquoRole ofguanine nucleotides in the regulation of the RascAMPpathwayin Saccharomyces cerevisiaerdquo Biochimica et Biophysica Acta vol1538 no 2-3 pp 181ndash189 2001
[3] K TanakaMNakafuku T Satoh et al ldquoS cerevisiae genes IRA1and IRA2 encode proteins that may be functionally equivalentto mammalian ras GTPase activating proteinrdquo Cell vol 60 no5 pp 803ndash807 1990
[4] S Zaman S I Lippman X Zhao and J R Broach ldquoHow Sac-charomyces responds to nutrientsrdquo Annual Review of Geneticsvol 42 pp 27ndash81 2008
[5] JMThevelein and J H DeWinde ldquoNovel sensingmechanismsand targets for the cAMP-protein kinase A pathway in the yeastSaccharomyces cerevisiaerdquoMolecularMicrobiology vol 33 no 5pp 904ndash918 1999
[6] F Rolland JWinderickx and JMThevelein ldquoGlucose-sensingand -signallingmechanisms in yeastrdquo FEMS Yeast Research vol2 no 2 pp 183ndash201 2002
[7] G F Casperson N Walker and H R Bourne ldquoIsolation of thegene encoding adenylate cyclase in Saccharomyces cerevisiaerdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 82 no 15 pp 5060ndash5063 1985
[8] B Smets R Ghillebert P De Snijder et al ldquoLife in the midstof scarcity adaptations to nutrient availability in Saccharomycescerevisiaerdquo Current Genetics vol 56 no 1 pp 1ndash32 2010
[9] S Broggi E Martegani and S Colombo ldquoLive-cell imaging ofendogenous Ras-GTP shows predominant Ras activation at theplasma membrane and in the nucleus in Saccharomyces cere-visiaerdquoThe International Journal of Biochemistry amp Cell Biologyvol 45 no 2 pp 384ndash394 2013
[10] J E Leadsham K Miller K R Ayscough et al ldquoWhi2p linksnutritional sensing to actin-dependent Ras-cAMP-PKA regu-lation and apoptosis in yeastrdquo Journal of Cell Science vol 122no 5 pp 706ndash715 2009
[11] YHuW LuG Chen et al ldquoK-rasG12V transformation leads tomitochondrial dysfunction and a metabolic switch from oxida-tive phosphorylation to glycolysisrdquo Cell Research vol 22 no 2pp 399ndash412 2012
[12] S Buttner T Eisenberg D Carmona-Gutierrez et al ldquoEndonu-clease G regulates budding yeast life and deathrdquoMolecular Cellvol 25 no 2 pp 233ndash246 2007
[13] F Madeo E Frohlich and K-U Frohlich ldquoA yeast mutantshowing diagnostic markers of early and late apoptosisrdquo Journalof Cell Biology vol 139 no 3 pp 729ndash734 1997
[14] FMadeo E Herker SWissing H Jungwirth T Eisenberg andK-U Frohlich ldquoApoptosis in yeastrdquo Current Opinion in Micro-biology vol 7 no 6 pp 655ndash660 2004
[15] F Madeo E Herker C Maldener et al ldquoA caspase-related pro-tease regulates apoptosis in yeastrdquo Molecular Cell vol 9 no 4pp 911ndash917 2002
[16] N Guaragnella C Pereira M J Sousa et al ldquoYCA1 participatesin the acetic acid induced yeast programmed cell death also in amanner unrelated to its caspase-like activityrdquo FEBS Letters vol580 no 30 pp 6880ndash6884 2006
[17] W Li L Sun Q Liang J Wang W Mo and B Zhou ldquoYeastAMID homologue Ndi1p displays respiration-restricted apop-totic activity and is involved in chronological agingrdquoMolecularBiology of the Cell vol 17 no 4 pp 1802ndash1811 2006
[18] S Wissing P Ludovico E Herker et al ldquoAn AIF orthologueregulates apoptosis in yeastrdquo Journal of Cell Biology vol 166 no7 pp 969ndash974 2004
[19] B Fahrenkrog U Sauder and U Aebi ldquoThe S cerevisiae HtrA-like protein Nma111p is a nuclear serine protease that mediatesyeast apoptosisrdquo Journal of Cell Science vol 117 no 1 pp 115ndash126 2004
[20] J Qiu J-H Yoon and B Shen ldquoSearch for apoptotic nucleasesin yeast role of Tat-D nuclease in apoptotic DNA degradationrdquoThe Journal of Biological Chemistry vol 280 no 15 pp 15370ndash15379 2005
[21] I Orlandi M Bettiga L Alberghina and M Vai ldquoTranscrip-tional profiling of ubp10 null mutant reveals altered subtelo-meric gene expression and insurgence of oxidative stressresponserdquoThe Journal of Biological Chemistry vol 279 no 8 pp6414ndash6425 2004
[22] M Ligr F Madeo E Frohlich W Hilt K-U Frohlich and DH Wolf ldquoMammalian Bax triggers apoptotic changes in yeastrdquoFEBS Letters vol 438 no 1-2 pp 61ndash65 1998
[23] F Madeo E Frohlich M Ligr et al ldquoOxygen stress a regulatorof apoptosis in yeastrdquo Journal of Cell Biology vol 145 no 4 pp757ndash767 1999
[24] L Du Y Su D Sun et al ldquoFormic acid induces Yca1p-inde-pendent apoptosis-like cell death in the yeast Saccharomycescerevisiaerdquo FEMS Yeast Research vol 8 no 4 pp 531ndash539 2008
10 Oxidative Medicine and Cellular Longevity
[25] P LudovicoM J SousaM T Silva C Leao andMCorte-RealldquoSaccharomyces cerevisiae commits to a programmed cell deathprocess in response to acetic acidrdquoMicrobiology vol 147 no 9pp 2409ndash2415 2001
[26] P Ludovico F Rodrigues AAlmeidaMT Silva A Barrientosand M Corte-Real ldquoCytochrome c release and mitochondriainvolvement in programmed cell death induced by acetic acidin Saccharomyces cerevisiaerdquo Molecular Biology of the Cell vol13 no 8 pp 2598ndash2606 2002
[27] N Guaragnella L Antonacci S Passarella E Marra and SGiannattasio ldquoHydrogen peroxide and superoxide anion pro-duction during acetic acid-induced yeast programmed celldeathrdquo Folia Microbiologica vol 52 no 3 pp 237ndash240 2007
[28] S Giannattasio A Atlante L Antonacci et al ldquoCytochrome c isreleased from coupled mitochondria of yeast en route to aceticacid-induced programmed cell death and can work as an elec-tron donor and a ROS scavengerrdquo FEBS Letters vol 582 no 10pp 1519ndash1525 2008
[29] N Guaragnella L Antonacci S Giannattasio E Marra and SPassarella ldquoCatalase T and Cu Zn-superoxide dismutase in theacetic acid-induced programmed cell death in Saccharomycescerevisiaerdquo FEBS Letters vol 582 no 2 pp 210ndash214 2008
[30] N Guaragnella A Bobba S Passarella E Marra and S Gian-nattasio ldquoYeast acetic acid-induced programmed cell death canoccurwithout cytochrome c releasewhich requiresmetacaspaseYCA1rdquo FEBS Letters vol 584 no 1 pp 224ndash228 2010
[31] F Madeo D Carmona-Gutierrez J Ring S Buttner T Eisen-berg and G Kroemer ldquoCaspase-dependent and caspase-inde-pendent cell death pathways in yeastrdquo Biochemical and Biophys-ical Research Communications vol 382 no 2 pp 227ndash231 2009
[32] DAhuatzi A Riera R Pelaez PHerrero andFMoreno ldquoHxk2regulates the phosphorylation state of Mig1 and therefore itsnucleocytoplasmic distributionrdquo The Journal of BiologicalChemistry vol 282 no 7 pp 4485ndash4493 2007
[33] FMoreno D Ahuatzi A Riera C A Palomino and P HerreroldquoGlucose sensing through the Hxk2-dependent signalling path-wayrdquo Biochemical Society Transactions vol 33 no 1 pp 265ndash268 2005
[34] B JThomas and R Rothstein ldquoElevated recombination rates intranscriptionally active DNArdquo Cell vol 56 no 4 pp 619ndash6301989
[35] L C Robinson J B Gibbs M S Marshall I S Sigal and KTatchell ldquoCDC25 a component of the RAS-adenylate cyclasepathway in Saccharomyces cerevisiaerdquo Science vol 235 no 4793pp 1218ndash1221 1987
[36] R J Rothstein ldquoOne-step gene disruption in yeastrdquoMethods inEnzymology vol 101 pp 202ndash211 1983
[37] M Bettiga L Calzari I Orlandi L Alberghina and M VaildquoInvolvement of the yeast metacaspase Yca1 in ubp10Δ-pro-grammed cell deathrdquo FEMS Yeast Research vol 5 no 2 pp 141ndash147 2004
[38] N Casatta A Porro I Orlandi L Brambilla andM Vai ldquoLackof Sir2 increases acetate consumption and decreases extracellu-lar pro-aging factorsrdquo Biochimica et Biophysica Acta vol 1833no 3 pp 593ndash601 2013
[39] S Colombo D Ronchetti J M Thevelein J Winderickx andE Martegani ldquoActivation state of the Ras2 protein and glucose-induced signaling in Saccharomyces cerevisiaerdquo The Journal ofBiological Chemistry vol 279 no 45 pp 46715ndash46722 2004
[40] P Fabrizio and V D Longo ldquoChronological aging-inducedapoptosis in yeastrdquo Biochimica et Biophysica Acta vol 1783 no7 pp 1280ndash1285 2008
[41] M A S Khan P B Chock and E R Stadtman ldquoKnockout ofcaspase-like gene YCA1 abrogates apoptosis and elevates oxi-dized proteins in Saccharomyces cerevisiaerdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 102 no 48 pp 17326ndash17331 2005
[42] J CWolfman S M Planchon J Liao and AWolfman ldquoStruc-tural and functional consequences of c-N-Ras constitutivelyassociated with intact mitochondriardquo Biochimica et BiophysicaActa vol 1763 no 10 pp 1108ndash1124 2006
[43] T G Bivona S E Quatela B O Bodemann et al ldquoPKC regu-lates a farnesyl-electrostatic switch on K-Ras that promotes itsassociation with Bcl-XL on mitochondria and induces apopto-sisrdquoMolecular Cell vol 21 no 4 pp 481ndash493 2006
[44] A Rebollo D Perez-Sala and C Martınez-A ldquoBcl-2 differen-tially targets K- N- and H-Ras to mitochondria in IL-2 sup-plemented or deprived cells implications in prevention of apop-tosisrdquo Oncogene vol 18 no 35 pp 4930ndash4939 1999
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
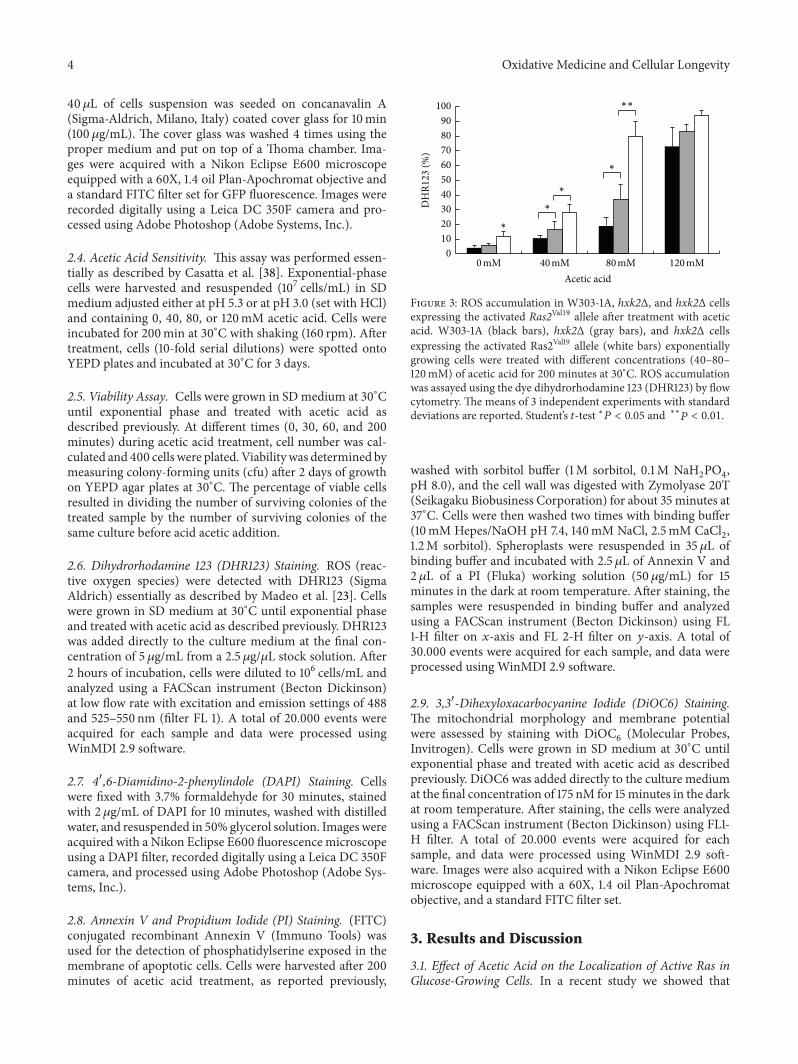
4 Oxidative Medicine and Cellular Longevity
40 120583L of cells suspension was seeded on concanavalin A(Sigma-Aldrich Milano Italy) coated cover glass for 10min(100 120583gmL) The cover glass was washed 4 times using theproper medium and put on top of a Thoma chamber Ima-ges were acquired with a Nikon Eclipse E600 microscopeequipped with a 60X 14 oil Plan-Apochromat objective anda standard FITC filter set for GFP fluorescence Images wererecorded digitally using a Leica DC 350F camera and pro-cessed using Adobe Photoshop (Adobe Systems Inc)
24 Acetic Acid Sensitivity This assay was performed essen-tially as described by Casatta et al [38] Exponential-phasecells were harvested and resuspended (107 cellsmL) in SDmedium adjusted either at pH 53 or at pH 30 (set with HCl)and containing 0 40 80 or 120mM acetic acid Cells wereincubated for 200min at 30∘C with shaking (160 rpm) Aftertreatment cells (10-fold serial dilutions) were spotted ontoYEPD plates and incubated at 30∘C for 3 days
25 Viability Assay Cells were grown in SDmedium at 30∘Cuntil exponential phase and treated with acetic acid asdescribed previously At different times (0 30 60 and 200minutes) during acetic acid treatment cell number was cal-culated and 400 cells were plated Viability was determined bymeasuring colony-forming units (cfu) after 2 days of growthon YEPD agar plates at 30∘C The percentage of viable cellsresulted in dividing the number of surviving colonies of thetreated sample by the number of surviving colonies of thesame culture before acid acetic addition
26 Dihydrorhodamine 123 (DHR123) Staining ROS (reac-tive oxygen species) were detected with DHR123 (SigmaAldrich) essentially as described by Madeo et al [23] Cellswere grown in SD medium at 30∘C until exponential phaseand treated with acetic acid as described previously DHR123was added directly to the culture medium at the final con-centration of 5120583gmL from a 25120583g120583L stock solution After2 hours of incubation cells were diluted to 106 cellsmL andanalyzed using a FACScan instrument (Becton Dickinson)at low flow rate with excitation and emission settings of 488and 525ndash550 nm (filter FL 1) A total of 20000 events wereacquired for each sample and data were processed usingWinMDI 29 software
27 410158406-Diamidino-2-phenylindole (DAPI) Staining Cellswere fixed with 37 formaldehyde for 30 minutes stainedwith 2120583gmL of DAPI for 10 minutes washed with distilledwater and resuspended in 50 glycerol solution Images wereacquired with a Nikon Eclipse E600 fluorescence microscopeusing a DAPI filter recorded digitally using a Leica DC 350Fcamera and processed using Adobe Photoshop (Adobe Sys-tems Inc)
28 Annexin V and Propidium Iodide (PI) Staining (FITC)conjugated recombinant Annexin V (Immuno Tools) wasused for the detection of phosphatidylserine exposed in themembrane of apoptotic cells Cells were harvested after 200minutes of acetic acid treatment as reported previously
0102030405060708090
100
Acetic acid
DH
R123
()
lowast
lowast
lowast
lowast
lowastlowast
0 mM 40 mM 80 mM 120 mM
Figure 3 ROS accumulation in W303-1A hxk2Δ and hxk2Δ cellsexpressing the activated Ras2Val19 allele after treatment with aceticacid W303-1A (black bars) hxk2Δ (gray bars) and hxk2Δ cellsexpressing the activated Ras2Val19 allele (white bars) exponentiallygrowing cells were treated with different concentrations (40ndash80ndash120mM) of acetic acid for 200 minutes at 30∘C ROS accumulationwas assayed using the dye dihydrorhodamine 123 (DHR123) by flowcytometry The means of 3 independent experiments with standarddeviations are reported Studentrsquos 119905-test lowast119875 lt 005 and lowastlowast119875 lt 001
washed with sorbitol buffer (1M sorbitol 01M NaH2PO4
pH 80) and the cell wall was digested with Zymolyase 20T(Seikagaku Biobusiness Corporation) for about 35 minutes at37∘C Cells were then washed two times with binding buffer(10mM HepesNaOH pH 74 140mM NaCl 25mM CaCl
2
12M sorbitol) Spheroplasts were resuspended in 35 120583L ofbinding buffer and incubated with 25 120583L of Annexin V and2 120583L of a PI (Fluka) working solution (50120583gmL) for 15minutes in the dark at room temperature After staining thesamples were resuspended in binding buffer and analyzedusing a FACScan instrument (Becton Dickinson) using FL1-H filter on 119909-axis and FL 2-H filter on 119910-axis A total of30000 events were acquired for each sample and data wereprocessed using WinMDI 29 software
29 331015840-Dihexyloxacarbocyanine Iodide (DiOC6) StainingThe mitochondrial morphology and membrane potentialwere assessed by staining with DiOC
6(Molecular Probes
Invitrogen) Cells were grown in SD medium at 30∘C untilexponential phase and treated with acetic acid as describedpreviously DiOC6 was added directly to the culture mediumat the final concentration of 175 nM for 15 minutes in the darkat room temperature After staining the cells were analyzedusing a FACScan instrument (Becton Dickinson) using FL1-H filter A total of 20000 events were acquired for eachsample and data were processed using WinMDI 29 soft-ware Images were also acquired with a Nikon Eclipse E600microscope equipped with a 60X 14 oil Plan-Apochromatobjective and a standard FITC filter set
3 Results and Discussion
31 Effect of Acetic Acid on the Localization of Active Ras inGlucose-Growing Cells In a recent study we showed that
Oxidative Medicine and Cellular Longevity 5
0
5
10
15
20
25
30
35
40A
nnex
in V
pos
(
)
Acetic acid
lowastlowast
lowastlowast
0 mM 40 mM 80 mM 120 mM
(a)
0
5
10
15
20
25
30
35
40
PI p
os (
)
Acetic acid
lowast
lowast
0 mM 40 mM 80 mM 120 mM
(b)
0
10
20
30
40
50
60
70
80
Acetic acid
Ann
exin
V+
PI p
os (
)
lowast
lowast
0 mM 40 mM 80 mM 120 mM
(c)
Figure 4 The occurrence of acetic-acid-induced cell death in hxk2Δ cells is characterized by markers of both apoptosis and necrosisAssessment of cell death by FITC-coupled annexinV and PI stainingW303-1A (black bars) and hxk2Δ (gray bars) exponentially growing cellswere treatedwith different concentrations (40ndash80ndash120mM) of acetic acid for 200minutes at 30∘C before being processed for determination ofphosphatidylserine externalization andmembrane integrity by flow cytometry 30000 events have been evaluatedThemeans of 3 independentexperiments with standard deviations are reported Studentrsquos 119905-test lowast119875 lt 005 and lowastlowast119875 lt 001
active Ras proteins accumulated mainly in the plasma mem-brane and in the nucleus in exponentially growing wild-typecells while they accumulated inmitochondria in cells deletedin the HXK2 gene indicating that this enzyme was involvedin the proper localization of these small GTPases [9] Therole played by active Ras in mitochondria is not knownalthough other reports have found Ras-GTP associated withthese organelles both inmammals and lower eukaryotes andhave linked it to cancer and apoptosis Leadsham et al [10]demonstrated that in S cerevisiae loss of Whi2p function aprotein known to influence cell cycle exit under conditions ofnutritional depletion determined an aberrant accumulationof activated Ras at the mitochondria In this mutant thefailure to shut down Ras signalling by addressing Ras to thevacuole would lead tomitochondrial dysfunction accumula-tion of damaging ROS and cell deathMore recently Hu et al[11] using a tetracycline inducible model demonstrated thatin mammalian cells association of K-rasG12V proteins withmitochondria induced mitochondrial dysfunction increasedROS accumulation and a metabolic switch from oxidativephosphorylation to glycolysis
To investigate whether a correlation exists between themitochondrial localization of active Ras and programmed
cell death we evaluated the effect of the addition of aceticacid a well-known apoptotic stimulus [25] to wild-typecells expressing the eGFP-RBD3 probe on the localizationof Ras-GTP proteins To this aim exponentially growingcells were collected by centrifugation resuspended in low pHmedium (pH 30) and pictures were taken at the fluorescencemicroscope before and after addition of 40mM acetic acidOur results showed that within 5 minutes this apoptoticstimulus caused the localization of active Ras proteins exclu-sively to the mitochondria reinforcing the hypothesis ofthe involvement of these proteins in programmed cell death(Figure 1) In wild-type cells resuspended in low pHmediumfor 5 minutes without acetic acid added active Ras proteinsaccumulated mainly in the plasma membrane and in thenucleus indicating that acidification of the medium did notinfluence the localization of these small G proteins (Figure 1)A mitochondrial localization of Ras-GTP was also observedwhen a low concentration of H
2O2was added to the medium
(data not shown) indicating that delocalization of active Rasto these organelles is a more general response to differentapoptotic stimuli and suggesting that mitochondrial localiza-tion of Ras2-GTP is actually important for the induction ofapoptosis in yeast
6 Oxidative Medicine and Cellular Longevity
W303-1A
256
0
Fluorescence
Even
ts
100
101
102
103
104
0 mM
40 mM
80 mM
120 mM
Fluorescence
Even
ts
256
010
010
110
210
310
4
0 mM
40 mM
80 mM
120 mM
hxk2
(a)
W303-1A
0 mM 80 mM
hxk2
(b)
Figure 5 Mitochondrial membrane potential and morphology in W303-1A and hxk2Δ strains upon acetic acid treatment W303-1A andhxk2Δ exponentially growing cells were treated with different concentrations (40ndash80ndash120mM) of acetic acid for 200 minutes at 30∘C DiOC
6
uptake was assessed using both flow cytometry (a) and fluorescence microscopy (b) Photomicrographs illustrate the alteration of the tubularmitochondrial network to clustered mitochondrial morphology particularly exacerbated in hxk2Δ cells after acetic acid treatment
32 Deletion of HXK2 Enhances Cell Death and IncreasesIntracellular ROS Levels The results presented previouslysuggest that localization of active Ras proteins to mitochon-dria might be involved in programmed cell death since theseproteins are localized to mitochondria following additionof acetic acid Since in glucose-growing hxk2Δ cells activeRas is constitutively located in mitochondria [9] we nextanalyzed the behavior of the hxk2Δmutant under conditions
that caused apoptosisWhile the growth of hxk2Δmutant wasalmost indistinguishable from the wild-type strain at pH 53it exhibited an increased sensitivity to acetic acid stress at lowpH (Figure 2(a)) suggesting that Hxk2 is required for normaltolerance to acetic acid treatment Cell survival of wild-typeand hxk2Δ cells after induction of apoptosis with acetic acidwas further tested in a plating assay After treatment withdifferent concentrations (80 and 120mM) of acetic acid for
Oxidative Medicine and Cellular Longevity 7
up to 200 minutes at 30∘C hxk2Δ cells showed a significantdose-dependent reduction in cell survival when comparedwith wild-type cells (Figure 2(b)) Taken together these datasuggest that Hxk2 is required for normal tolerance to aceticacid treatment
Since an aberrant accumulation of activated Ras in mito-chondria accompanied to accumulation of ROS has alreadybeen reported both in mammals and yeast [10 11] and sinceROS play a pivotal role in apoptotic cell death FACS analyseswere performed to evaluate the accumulation of ROS in wild-type and hxk2Δ cells treated with different concentrationsof acetic acid for 200min at 30∘C In particular DHR123was used to determine the accumulation of ROS in the cellssince this compound can easily permeate them and can bequantitatively oxidized to a green fluorescent product inthe presence of ROS The percentage of oxidized R123 wasabout the double for hxk2Δ cells compared to wild-type cells200min after treatment with either 40 or 80mM acetic acid(Figure 3) To substantiate the hypothesis that an aberrantaccumulation of activated Ras in mitochondria might leadtomitochondrial dysfunction and accumulation of damagingROS we expressed the dominant active RAS2Val19 allelewhich was reported to show a much higher level of Ras2-GTP [39] in hxk2Δ cells Our results showed that indeed theexpression of this allele in the hxk2Δ background caused afurther increase in the level of ROS both in growing cells andafter treatment with acetic acid (Figure 3)These data suggestthat conditions that presumably cause a highermitochondrialaccumulation of active Ras may contribute to accumulationof ROS and cell death
33 Loss of Hxk2 Causes an Increase of Both Apoptosis andNecrosis The results presented previously suggest that Hxk2might have an antiapoptotic activity To better characterizethe nature of cell death triggered by addition of acetic acid tohxk2Δ cells we quantified phenotypic changes indicative ofapoptosis While apoptotic DNA condensation was detectedby DAPI staining combined Annexin Vpropidium iodide(PI) staining was used to discriminate between early apop-totic (Annexin V+PIminus) late apoptoticsecondary necrotic(Annexin V+PI+) and necrotic (Annexin VminusPI+) deathsDAPI staining showed that nuclei of untreated cells wereround while the nuclear DNA was condensed in bothwild-type and hxk2Δ cells treated with 80mM acetic acidbeing the extent of this phenotype more pronounced for themutant strain compared to the wild-type strain (data notshown) Annexin Vpropidium iodide (PI) staining revealedthat hxk2Δ-facilitated cell death was accompanied by anincrease in both apoptotic and necroticmarkers (Figure 4) Inparticular untreated hxk2Δ cells showed a higher percentageof both early apoptotic and necrotic cells compared withthe wild-type strain which further increased upon treatmentwith acetic acid
34 Deletion of HXK2 Causes Hyperpolarization of Mitochon-dria Mitochondrial membrane potential (ΔΨ
119898) is a useful
indicator of mitochondrial function To measure ΔΨ119898 we
used the cationic lipophylic dye DiOC6 which accumulates
0102030405060708090
100110
DH
R 12
3 (
)
Acetic acid
lowast
lowast
lowast
0 mM 40 mM 80 mM 120 mM
Figure 6 Effect of YCA1 deletion on ROS accumulation aftertreatment with acetic acid W303-1A (black bars) yca1Δ (blackhatched bars) hxk2Δ (gray bars) and hxk2Δ-yca1Δ (white bars)exponentially growing cells were treated with different concen-trations (40ndash80ndash120mM) of acetic acid for 200 minutes at 30∘CROS accumulation was assayed by flow cytometry using the dyedihydrorhodamine 123 (DHR123) Studentrsquos 119905-test lowast119875 lt 005
in mitochondria in accordance with ΔΨ119898 Wild-type and
hxk2Δ cells were treatedwith different concentration of aceticacid (40ndash80ndash120mM) for 200min at 30∘C collected andstained with this dye FACS analysis showed that mitochon-drial membrane potential clearly increased in both wild-typeand hxk2Δ cells after treatment with acetic acid comparedwith untreated control cells with the hyperpolarizationof mitochondria being much stronger in the mutant cellscomparedwithwild-type cells (Figure 5(a)) In parallel mito-chondrial morphology was assessed by using DiOC
6 both
in wild-type and hxk2Δ cells At low concentration this dyespecifically stains the mitochondrial membranes in a mannerthat depends on membrane potential and can be observed byfluorescence microscopy Before treatment with acetic acidbothwild-type and hxk2Δ cells displayed a tubularmitochon-drial morphology indicating that these mitochondria werehealthy and possessed a membrane potential (Figure 5(b))By contrast the mitochondrial membranes present in bothwild-type and hxk2Δ cells appeared rounded and highlyfragmented after treatment with acetic acid (Figure 5(b))This conversion of mitochondrial morphology from tubu-lar to punctuate structures is likely to occur by excessivemitochondrial fission and has already been observed in yeastapoptosis induced by acetic acid treatment [40] ImportantlyDiOC
6staining was greatly increased and more pronounced
in hxk2Δ cells compared to wild-type cells after treatmentwith acetic acid confirming the increase of fluorescenceobserved in these mutant cells by FACS analysis Thesedata indicate that both in wild-type and hxk2Δ cells aceticacid treatment caused hyperpolarization of mitochondrialmembrane with consequent damage of these organelles andloss of functionality This effect was more pronounced inhxk2Δ cells compared to wild-type cells
35 Acetic-Acid-Induced Cell Death in the hxk2Δ Strain Is Yca1Independent At least two death pathways can be activated
8 Oxidative Medicine and Cellular Longevity
lowastlowast
lowastlowast
lowast
lowastlowast
0 mM 40 mM 80 mM 120 mMAcetic acid
0
5
10
15
20
25
30
35
40A
nnex
in V
pos
(
)
(a)
lowast
lowast
lowastlowast
lowastlowast
0 mM 40 mM 80 mM 120 mM0
10
20
30
40
50
60
70
80
PI p
os (
)
Acetic acid
(b)
lowastlowast
lowast
lowast
0 mM 40 mM 80 mM 120 mM0
10
20
30
40
50
60
70
80
Acetic acid
Ann
exin
V+
PI p
os (
)
(c)
Figure 7 Effect of YCA1 deletion on cell death after treatment with acetic acid W303-1A (black bars) yca1Δ (black hatched bars) hxk2Δ(gray bars) and hxk2Δ-yca1Δ (white bars) exponentially growing cells were treated with different concentrations (40ndash80ndash120mM) of aceticacid for 200 minutes at 30∘C Cell death was assessed by flow cytometry using FITC-coupled annexin V and PI co-staining to determinatethe externalization of phosphatidylserine and the membrane integrityThemeans of 3 independent experiments with standard deviations arereported Studentrsquos 119905-test lowast119875 lt 005 and lowastlowast119875 lt 001
in yeast acetic-acid-induced apoptosis one dependent oncytochromec release which requires YCA1 and the other(s)independent of it [16 30] Consequently we next investigatedwhether the yeast caspase Yca1 might be involved in acetic-acid-induced cell death in hxk2Δ cells In particular toassess whether Yca1 plays a role in acetic-acid-induced ROSaccumulation in hxk2Δ cells we treated hxk2Δ and hxk2Δyca1Δ cells with different concentration of acetic acid (40ndash80ndash120mM) for 200min at 30∘C and measured the ROSaccumulation Our results showed that deletion of YCA1 inhxk2Δ cells had no effect on acetic-acid-induced ROS accu-mulation (Figure 6) suggesting that the generation of ROSin hxk2Δ cells upon acetic acid stress is Yca1 independentHowever the level of intracellular ROS was consistentlyhigher in the hxk2Δ yca1Δ doublemutant growing on glucosemedium compared with the hxk2Δ mutant Similarly Du etal [24] showed that formic acid induced Yca1-independentapoptosis-like cell death and that the burst of ROS duringcell death occurred earlier and at a higher level in the yca1Δstrain than in the wild-type strain Moreover Khan et al [41]determined the level of oxidized proteins in yeast cells underH2O2stress and showed that lack of Yca1 abrogated apoptosis
but elevated intracellular oxidized proteins compared with
wild-type cells Consequently both our results and data in theliterature [24 41] clearly suggest a linkage between ROS pro-duction and Yca1 activation during apoptosis in yeast Finallycytofluorometric quantification of phosphatidylserine exter-nalization andormembrane permeabilization (annexinVPIcostaining) further confirmed that Yca1p did not influencethe acetic-acid-induced cell death in hxk2Δ cells (Figure 7)These results strongly suggest that apoptosis induced by aceticacid in hxk2Δ cells is largely Yca1 independent
4 Conclusion
Several studies indicate that the Ras proteinmay be physicallyassociated with mitochondria both in yeast and mammals[10 11 42ndash44] This study provides results indicating thatassociation of active Ras to these organelles might be linkedto apoptosis First of all we show that addition of acetic acida well-known apoptotic stimulus in S cerevisiae to growingwild-type cells determined a delocalization of active Ras pro-teins from nuclei and plasma membrane to mitochondriaFurthermore we show that addition of acetic acid to hxk2Δcells showing a constitutive localization of active Ras inmito-chondria [9] enhanced ROS production and cell death
Oxidative Medicine and Cellular Longevity 9
compared with wild-type cells Moreover expression of thedominant active RAS2Val19 allele in the hxk2Δ backgroundcaused an even higher increase in the level of ROS both ingrowing cells and after treatment with acetic acid providinga further proof to the hypothesis that conditions that presum-ably cause a higher mitochondrial accumulation of activeRas may contribute to accumulation of ROS and cell deathWe also show that in hxk2Δ cells addition of acetic acidincreased cell death compared with the wild-type strain withthe typical markers of apoptosis highlighting a new rolefor Hexokinase 2 as an antiapoptotic factor in S cerevisiaecells However at this stage we cannot exclude a correlationbetween the well-known functions of Hxk2 in glucose repres-sion and signalling and the role of Ras in inducing apoptosis
Finally in this study we show that lack of Hxk2 inducesapoptosis via a mitochondria-mediated pathway withoutmetacaspase Yca1 involvement since deletion of YCA1 in thehxk2Δ background did not abrogate the acetic-acid-inducedaccumulation of ROS and did not decrease the percentage ofboth apoptotic and necrotic cells
Acknowledgments
The authors thank J Winderickx University of Leuven Bel-gium for the gift of strains and M Vai Universita Milano-Bicocca Milan for the gift of primers This work was sup-ported by FAR grant to Sonia Colombo and by founding ofProgram SysBioNet Italian Roadmap Research Infrastruc-ture 2012
References
[1] S JonesM-L Vignais and J R Broach ldquoTheCDC25 protein ofSaccharomyces cerevisiae promotes exchange of guanine nucle-otides bound to RasrdquoMolecular and Cellular Biology vol 11 no5 pp 2641ndash2646 1991
[2] S Rudoni S Colombo P Coccetti and E Martegani ldquoRole ofguanine nucleotides in the regulation of the RascAMPpathwayin Saccharomyces cerevisiaerdquo Biochimica et Biophysica Acta vol1538 no 2-3 pp 181ndash189 2001
[3] K TanakaMNakafuku T Satoh et al ldquoS cerevisiae genes IRA1and IRA2 encode proteins that may be functionally equivalentto mammalian ras GTPase activating proteinrdquo Cell vol 60 no5 pp 803ndash807 1990
[4] S Zaman S I Lippman X Zhao and J R Broach ldquoHow Sac-charomyces responds to nutrientsrdquo Annual Review of Geneticsvol 42 pp 27ndash81 2008
[5] JMThevelein and J H DeWinde ldquoNovel sensingmechanismsand targets for the cAMP-protein kinase A pathway in the yeastSaccharomyces cerevisiaerdquoMolecularMicrobiology vol 33 no 5pp 904ndash918 1999
[6] F Rolland JWinderickx and JMThevelein ldquoGlucose-sensingand -signallingmechanisms in yeastrdquo FEMS Yeast Research vol2 no 2 pp 183ndash201 2002
[7] G F Casperson N Walker and H R Bourne ldquoIsolation of thegene encoding adenylate cyclase in Saccharomyces cerevisiaerdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 82 no 15 pp 5060ndash5063 1985
[8] B Smets R Ghillebert P De Snijder et al ldquoLife in the midstof scarcity adaptations to nutrient availability in Saccharomycescerevisiaerdquo Current Genetics vol 56 no 1 pp 1ndash32 2010
[9] S Broggi E Martegani and S Colombo ldquoLive-cell imaging ofendogenous Ras-GTP shows predominant Ras activation at theplasma membrane and in the nucleus in Saccharomyces cere-visiaerdquoThe International Journal of Biochemistry amp Cell Biologyvol 45 no 2 pp 384ndash394 2013
[10] J E Leadsham K Miller K R Ayscough et al ldquoWhi2p linksnutritional sensing to actin-dependent Ras-cAMP-PKA regu-lation and apoptosis in yeastrdquo Journal of Cell Science vol 122no 5 pp 706ndash715 2009
[11] YHuW LuG Chen et al ldquoK-rasG12V transformation leads tomitochondrial dysfunction and a metabolic switch from oxida-tive phosphorylation to glycolysisrdquo Cell Research vol 22 no 2pp 399ndash412 2012
[12] S Buttner T Eisenberg D Carmona-Gutierrez et al ldquoEndonu-clease G regulates budding yeast life and deathrdquoMolecular Cellvol 25 no 2 pp 233ndash246 2007
[13] F Madeo E Frohlich and K-U Frohlich ldquoA yeast mutantshowing diagnostic markers of early and late apoptosisrdquo Journalof Cell Biology vol 139 no 3 pp 729ndash734 1997
[14] FMadeo E Herker SWissing H Jungwirth T Eisenberg andK-U Frohlich ldquoApoptosis in yeastrdquo Current Opinion in Micro-biology vol 7 no 6 pp 655ndash660 2004
[15] F Madeo E Herker C Maldener et al ldquoA caspase-related pro-tease regulates apoptosis in yeastrdquo Molecular Cell vol 9 no 4pp 911ndash917 2002
[16] N Guaragnella C Pereira M J Sousa et al ldquoYCA1 participatesin the acetic acid induced yeast programmed cell death also in amanner unrelated to its caspase-like activityrdquo FEBS Letters vol580 no 30 pp 6880ndash6884 2006
[17] W Li L Sun Q Liang J Wang W Mo and B Zhou ldquoYeastAMID homologue Ndi1p displays respiration-restricted apop-totic activity and is involved in chronological agingrdquoMolecularBiology of the Cell vol 17 no 4 pp 1802ndash1811 2006
[18] S Wissing P Ludovico E Herker et al ldquoAn AIF orthologueregulates apoptosis in yeastrdquo Journal of Cell Biology vol 166 no7 pp 969ndash974 2004
[19] B Fahrenkrog U Sauder and U Aebi ldquoThe S cerevisiae HtrA-like protein Nma111p is a nuclear serine protease that mediatesyeast apoptosisrdquo Journal of Cell Science vol 117 no 1 pp 115ndash126 2004
[20] J Qiu J-H Yoon and B Shen ldquoSearch for apoptotic nucleasesin yeast role of Tat-D nuclease in apoptotic DNA degradationrdquoThe Journal of Biological Chemistry vol 280 no 15 pp 15370ndash15379 2005
[21] I Orlandi M Bettiga L Alberghina and M Vai ldquoTranscrip-tional profiling of ubp10 null mutant reveals altered subtelo-meric gene expression and insurgence of oxidative stressresponserdquoThe Journal of Biological Chemistry vol 279 no 8 pp6414ndash6425 2004
[22] M Ligr F Madeo E Frohlich W Hilt K-U Frohlich and DH Wolf ldquoMammalian Bax triggers apoptotic changes in yeastrdquoFEBS Letters vol 438 no 1-2 pp 61ndash65 1998
[23] F Madeo E Frohlich M Ligr et al ldquoOxygen stress a regulatorof apoptosis in yeastrdquo Journal of Cell Biology vol 145 no 4 pp757ndash767 1999
[24] L Du Y Su D Sun et al ldquoFormic acid induces Yca1p-inde-pendent apoptosis-like cell death in the yeast Saccharomycescerevisiaerdquo FEMS Yeast Research vol 8 no 4 pp 531ndash539 2008
10 Oxidative Medicine and Cellular Longevity
[25] P LudovicoM J SousaM T Silva C Leao andMCorte-RealldquoSaccharomyces cerevisiae commits to a programmed cell deathprocess in response to acetic acidrdquoMicrobiology vol 147 no 9pp 2409ndash2415 2001
[26] P Ludovico F Rodrigues AAlmeidaMT Silva A Barrientosand M Corte-Real ldquoCytochrome c release and mitochondriainvolvement in programmed cell death induced by acetic acidin Saccharomyces cerevisiaerdquo Molecular Biology of the Cell vol13 no 8 pp 2598ndash2606 2002
[27] N Guaragnella L Antonacci S Passarella E Marra and SGiannattasio ldquoHydrogen peroxide and superoxide anion pro-duction during acetic acid-induced yeast programmed celldeathrdquo Folia Microbiologica vol 52 no 3 pp 237ndash240 2007
[28] S Giannattasio A Atlante L Antonacci et al ldquoCytochrome c isreleased from coupled mitochondria of yeast en route to aceticacid-induced programmed cell death and can work as an elec-tron donor and a ROS scavengerrdquo FEBS Letters vol 582 no 10pp 1519ndash1525 2008
[29] N Guaragnella L Antonacci S Giannattasio E Marra and SPassarella ldquoCatalase T and Cu Zn-superoxide dismutase in theacetic acid-induced programmed cell death in Saccharomycescerevisiaerdquo FEBS Letters vol 582 no 2 pp 210ndash214 2008
[30] N Guaragnella A Bobba S Passarella E Marra and S Gian-nattasio ldquoYeast acetic acid-induced programmed cell death canoccurwithout cytochrome c releasewhich requiresmetacaspaseYCA1rdquo FEBS Letters vol 584 no 1 pp 224ndash228 2010
[31] F Madeo D Carmona-Gutierrez J Ring S Buttner T Eisen-berg and G Kroemer ldquoCaspase-dependent and caspase-inde-pendent cell death pathways in yeastrdquo Biochemical and Biophys-ical Research Communications vol 382 no 2 pp 227ndash231 2009
[32] DAhuatzi A Riera R Pelaez PHerrero andFMoreno ldquoHxk2regulates the phosphorylation state of Mig1 and therefore itsnucleocytoplasmic distributionrdquo The Journal of BiologicalChemistry vol 282 no 7 pp 4485ndash4493 2007
[33] FMoreno D Ahuatzi A Riera C A Palomino and P HerreroldquoGlucose sensing through the Hxk2-dependent signalling path-wayrdquo Biochemical Society Transactions vol 33 no 1 pp 265ndash268 2005
[34] B JThomas and R Rothstein ldquoElevated recombination rates intranscriptionally active DNArdquo Cell vol 56 no 4 pp 619ndash6301989
[35] L C Robinson J B Gibbs M S Marshall I S Sigal and KTatchell ldquoCDC25 a component of the RAS-adenylate cyclasepathway in Saccharomyces cerevisiaerdquo Science vol 235 no 4793pp 1218ndash1221 1987
[36] R J Rothstein ldquoOne-step gene disruption in yeastrdquoMethods inEnzymology vol 101 pp 202ndash211 1983
[37] M Bettiga L Calzari I Orlandi L Alberghina and M VaildquoInvolvement of the yeast metacaspase Yca1 in ubp10Δ-pro-grammed cell deathrdquo FEMS Yeast Research vol 5 no 2 pp 141ndash147 2004
[38] N Casatta A Porro I Orlandi L Brambilla andM Vai ldquoLackof Sir2 increases acetate consumption and decreases extracellu-lar pro-aging factorsrdquo Biochimica et Biophysica Acta vol 1833no 3 pp 593ndash601 2013
[39] S Colombo D Ronchetti J M Thevelein J Winderickx andE Martegani ldquoActivation state of the Ras2 protein and glucose-induced signaling in Saccharomyces cerevisiaerdquo The Journal ofBiological Chemistry vol 279 no 45 pp 46715ndash46722 2004
[40] P Fabrizio and V D Longo ldquoChronological aging-inducedapoptosis in yeastrdquo Biochimica et Biophysica Acta vol 1783 no7 pp 1280ndash1285 2008
[41] M A S Khan P B Chock and E R Stadtman ldquoKnockout ofcaspase-like gene YCA1 abrogates apoptosis and elevates oxi-dized proteins in Saccharomyces cerevisiaerdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 102 no 48 pp 17326ndash17331 2005
[42] J CWolfman S M Planchon J Liao and AWolfman ldquoStruc-tural and functional consequences of c-N-Ras constitutivelyassociated with intact mitochondriardquo Biochimica et BiophysicaActa vol 1763 no 10 pp 1108ndash1124 2006
[43] T G Bivona S E Quatela B O Bodemann et al ldquoPKC regu-lates a farnesyl-electrostatic switch on K-Ras that promotes itsassociation with Bcl-XL on mitochondria and induces apopto-sisrdquoMolecular Cell vol 21 no 4 pp 481ndash493 2006
[44] A Rebollo D Perez-Sala and C Martınez-A ldquoBcl-2 differen-tially targets K- N- and H-Ras to mitochondria in IL-2 sup-plemented or deprived cells implications in prevention of apop-tosisrdquo Oncogene vol 18 no 35 pp 4930ndash4939 1999
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
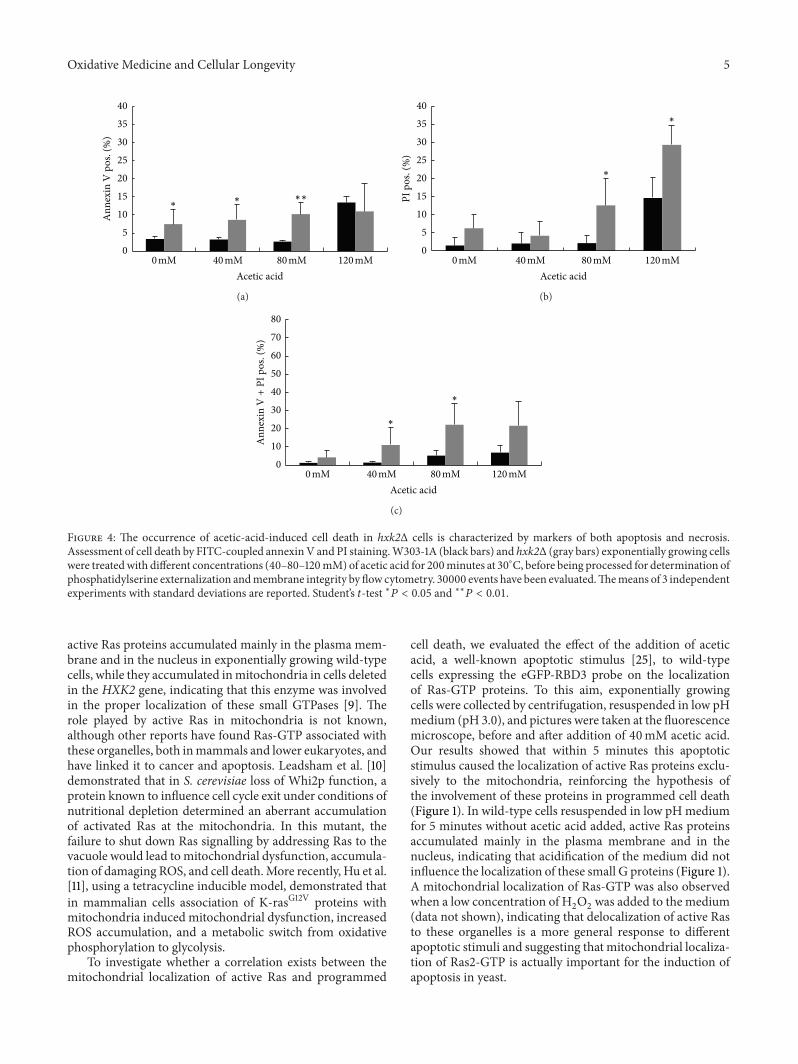
Oxidative Medicine and Cellular Longevity 5
0
5
10
15
20
25
30
35
40A
nnex
in V
pos
(
)
Acetic acid
lowastlowast
lowastlowast
0 mM 40 mM 80 mM 120 mM
(a)
0
5
10
15
20
25
30
35
40
PI p
os (
)
Acetic acid
lowast
lowast
0 mM 40 mM 80 mM 120 mM
(b)
0
10
20
30
40
50
60
70
80
Acetic acid
Ann
exin
V+
PI p
os (
)
lowast
lowast
0 mM 40 mM 80 mM 120 mM
(c)
Figure 4 The occurrence of acetic-acid-induced cell death in hxk2Δ cells is characterized by markers of both apoptosis and necrosisAssessment of cell death by FITC-coupled annexinV and PI stainingW303-1A (black bars) and hxk2Δ (gray bars) exponentially growing cellswere treatedwith different concentrations (40ndash80ndash120mM) of acetic acid for 200minutes at 30∘C before being processed for determination ofphosphatidylserine externalization andmembrane integrity by flow cytometry 30000 events have been evaluatedThemeans of 3 independentexperiments with standard deviations are reported Studentrsquos 119905-test lowast119875 lt 005 and lowastlowast119875 lt 001
active Ras proteins accumulated mainly in the plasma mem-brane and in the nucleus in exponentially growing wild-typecells while they accumulated inmitochondria in cells deletedin the HXK2 gene indicating that this enzyme was involvedin the proper localization of these small GTPases [9] Therole played by active Ras in mitochondria is not knownalthough other reports have found Ras-GTP associated withthese organelles both inmammals and lower eukaryotes andhave linked it to cancer and apoptosis Leadsham et al [10]demonstrated that in S cerevisiae loss of Whi2p function aprotein known to influence cell cycle exit under conditions ofnutritional depletion determined an aberrant accumulationof activated Ras at the mitochondria In this mutant thefailure to shut down Ras signalling by addressing Ras to thevacuole would lead tomitochondrial dysfunction accumula-tion of damaging ROS and cell deathMore recently Hu et al[11] using a tetracycline inducible model demonstrated thatin mammalian cells association of K-rasG12V proteins withmitochondria induced mitochondrial dysfunction increasedROS accumulation and a metabolic switch from oxidativephosphorylation to glycolysis
To investigate whether a correlation exists between themitochondrial localization of active Ras and programmed
cell death we evaluated the effect of the addition of aceticacid a well-known apoptotic stimulus [25] to wild-typecells expressing the eGFP-RBD3 probe on the localizationof Ras-GTP proteins To this aim exponentially growingcells were collected by centrifugation resuspended in low pHmedium (pH 30) and pictures were taken at the fluorescencemicroscope before and after addition of 40mM acetic acidOur results showed that within 5 minutes this apoptoticstimulus caused the localization of active Ras proteins exclu-sively to the mitochondria reinforcing the hypothesis ofthe involvement of these proteins in programmed cell death(Figure 1) In wild-type cells resuspended in low pHmediumfor 5 minutes without acetic acid added active Ras proteinsaccumulated mainly in the plasma membrane and in thenucleus indicating that acidification of the medium did notinfluence the localization of these small G proteins (Figure 1)A mitochondrial localization of Ras-GTP was also observedwhen a low concentration of H
2O2was added to the medium
(data not shown) indicating that delocalization of active Rasto these organelles is a more general response to differentapoptotic stimuli and suggesting that mitochondrial localiza-tion of Ras2-GTP is actually important for the induction ofapoptosis in yeast
6 Oxidative Medicine and Cellular Longevity
W303-1A
256
0
Fluorescence
Even
ts
100
101
102
103
104
0 mM
40 mM
80 mM
120 mM
Fluorescence
Even
ts
256
010
010
110
210
310
4
0 mM
40 mM
80 mM
120 mM
hxk2
(a)
W303-1A
0 mM 80 mM
hxk2
(b)
Figure 5 Mitochondrial membrane potential and morphology in W303-1A and hxk2Δ strains upon acetic acid treatment W303-1A andhxk2Δ exponentially growing cells were treated with different concentrations (40ndash80ndash120mM) of acetic acid for 200 minutes at 30∘C DiOC
6
uptake was assessed using both flow cytometry (a) and fluorescence microscopy (b) Photomicrographs illustrate the alteration of the tubularmitochondrial network to clustered mitochondrial morphology particularly exacerbated in hxk2Δ cells after acetic acid treatment
32 Deletion of HXK2 Enhances Cell Death and IncreasesIntracellular ROS Levels The results presented previouslysuggest that localization of active Ras proteins to mitochon-dria might be involved in programmed cell death since theseproteins are localized to mitochondria following additionof acetic acid Since in glucose-growing hxk2Δ cells activeRas is constitutively located in mitochondria [9] we nextanalyzed the behavior of the hxk2Δmutant under conditions
that caused apoptosisWhile the growth of hxk2Δmutant wasalmost indistinguishable from the wild-type strain at pH 53it exhibited an increased sensitivity to acetic acid stress at lowpH (Figure 2(a)) suggesting that Hxk2 is required for normaltolerance to acetic acid treatment Cell survival of wild-typeand hxk2Δ cells after induction of apoptosis with acetic acidwas further tested in a plating assay After treatment withdifferent concentrations (80 and 120mM) of acetic acid for
Oxidative Medicine and Cellular Longevity 7
up to 200 minutes at 30∘C hxk2Δ cells showed a significantdose-dependent reduction in cell survival when comparedwith wild-type cells (Figure 2(b)) Taken together these datasuggest that Hxk2 is required for normal tolerance to aceticacid treatment
Since an aberrant accumulation of activated Ras in mito-chondria accompanied to accumulation of ROS has alreadybeen reported both in mammals and yeast [10 11] and sinceROS play a pivotal role in apoptotic cell death FACS analyseswere performed to evaluate the accumulation of ROS in wild-type and hxk2Δ cells treated with different concentrationsof acetic acid for 200min at 30∘C In particular DHR123was used to determine the accumulation of ROS in the cellssince this compound can easily permeate them and can bequantitatively oxidized to a green fluorescent product inthe presence of ROS The percentage of oxidized R123 wasabout the double for hxk2Δ cells compared to wild-type cells200min after treatment with either 40 or 80mM acetic acid(Figure 3) To substantiate the hypothesis that an aberrantaccumulation of activated Ras in mitochondria might leadtomitochondrial dysfunction and accumulation of damagingROS we expressed the dominant active RAS2Val19 allelewhich was reported to show a much higher level of Ras2-GTP [39] in hxk2Δ cells Our results showed that indeed theexpression of this allele in the hxk2Δ background caused afurther increase in the level of ROS both in growing cells andafter treatment with acetic acid (Figure 3)These data suggestthat conditions that presumably cause a highermitochondrialaccumulation of active Ras may contribute to accumulationof ROS and cell death
33 Loss of Hxk2 Causes an Increase of Both Apoptosis andNecrosis The results presented previously suggest that Hxk2might have an antiapoptotic activity To better characterizethe nature of cell death triggered by addition of acetic acid tohxk2Δ cells we quantified phenotypic changes indicative ofapoptosis While apoptotic DNA condensation was detectedby DAPI staining combined Annexin Vpropidium iodide(PI) staining was used to discriminate between early apop-totic (Annexin V+PIminus) late apoptoticsecondary necrotic(Annexin V+PI+) and necrotic (Annexin VminusPI+) deathsDAPI staining showed that nuclei of untreated cells wereround while the nuclear DNA was condensed in bothwild-type and hxk2Δ cells treated with 80mM acetic acidbeing the extent of this phenotype more pronounced for themutant strain compared to the wild-type strain (data notshown) Annexin Vpropidium iodide (PI) staining revealedthat hxk2Δ-facilitated cell death was accompanied by anincrease in both apoptotic and necroticmarkers (Figure 4) Inparticular untreated hxk2Δ cells showed a higher percentageof both early apoptotic and necrotic cells compared withthe wild-type strain which further increased upon treatmentwith acetic acid
34 Deletion of HXK2 Causes Hyperpolarization of Mitochon-dria Mitochondrial membrane potential (ΔΨ
119898) is a useful
indicator of mitochondrial function To measure ΔΨ119898 we
used the cationic lipophylic dye DiOC6 which accumulates
0102030405060708090
100110
DH
R 12
3 (
)
Acetic acid
lowast
lowast
lowast
0 mM 40 mM 80 mM 120 mM
Figure 6 Effect of YCA1 deletion on ROS accumulation aftertreatment with acetic acid W303-1A (black bars) yca1Δ (blackhatched bars) hxk2Δ (gray bars) and hxk2Δ-yca1Δ (white bars)exponentially growing cells were treated with different concen-trations (40ndash80ndash120mM) of acetic acid for 200 minutes at 30∘CROS accumulation was assayed by flow cytometry using the dyedihydrorhodamine 123 (DHR123) Studentrsquos 119905-test lowast119875 lt 005
in mitochondria in accordance with ΔΨ119898 Wild-type and
hxk2Δ cells were treatedwith different concentration of aceticacid (40ndash80ndash120mM) for 200min at 30∘C collected andstained with this dye FACS analysis showed that mitochon-drial membrane potential clearly increased in both wild-typeand hxk2Δ cells after treatment with acetic acid comparedwith untreated control cells with the hyperpolarizationof mitochondria being much stronger in the mutant cellscomparedwithwild-type cells (Figure 5(a)) In parallel mito-chondrial morphology was assessed by using DiOC
6 both
in wild-type and hxk2Δ cells At low concentration this dyespecifically stains the mitochondrial membranes in a mannerthat depends on membrane potential and can be observed byfluorescence microscopy Before treatment with acetic acidbothwild-type and hxk2Δ cells displayed a tubularmitochon-drial morphology indicating that these mitochondria werehealthy and possessed a membrane potential (Figure 5(b))By contrast the mitochondrial membranes present in bothwild-type and hxk2Δ cells appeared rounded and highlyfragmented after treatment with acetic acid (Figure 5(b))This conversion of mitochondrial morphology from tubu-lar to punctuate structures is likely to occur by excessivemitochondrial fission and has already been observed in yeastapoptosis induced by acetic acid treatment [40] ImportantlyDiOC
6staining was greatly increased and more pronounced
in hxk2Δ cells compared to wild-type cells after treatmentwith acetic acid confirming the increase of fluorescenceobserved in these mutant cells by FACS analysis Thesedata indicate that both in wild-type and hxk2Δ cells aceticacid treatment caused hyperpolarization of mitochondrialmembrane with consequent damage of these organelles andloss of functionality This effect was more pronounced inhxk2Δ cells compared to wild-type cells
35 Acetic-Acid-Induced Cell Death in the hxk2Δ Strain Is Yca1Independent At least two death pathways can be activated
8 Oxidative Medicine and Cellular Longevity
lowastlowast
lowastlowast
lowast
lowastlowast
0 mM 40 mM 80 mM 120 mMAcetic acid
0
5
10
15
20
25
30
35
40A
nnex
in V
pos
(
)
(a)
lowast
lowast
lowastlowast
lowastlowast
0 mM 40 mM 80 mM 120 mM0
10
20
30
40
50
60
70
80
PI p
os (
)
Acetic acid
(b)
lowastlowast
lowast
lowast
0 mM 40 mM 80 mM 120 mM0
10
20
30
40
50
60
70
80
Acetic acid
Ann
exin
V+
PI p
os (
)
(c)
Figure 7 Effect of YCA1 deletion on cell death after treatment with acetic acid W303-1A (black bars) yca1Δ (black hatched bars) hxk2Δ(gray bars) and hxk2Δ-yca1Δ (white bars) exponentially growing cells were treated with different concentrations (40ndash80ndash120mM) of aceticacid for 200 minutes at 30∘C Cell death was assessed by flow cytometry using FITC-coupled annexin V and PI co-staining to determinatethe externalization of phosphatidylserine and the membrane integrityThemeans of 3 independent experiments with standard deviations arereported Studentrsquos 119905-test lowast119875 lt 005 and lowastlowast119875 lt 001
in yeast acetic-acid-induced apoptosis one dependent oncytochromec release which requires YCA1 and the other(s)independent of it [16 30] Consequently we next investigatedwhether the yeast caspase Yca1 might be involved in acetic-acid-induced cell death in hxk2Δ cells In particular toassess whether Yca1 plays a role in acetic-acid-induced ROSaccumulation in hxk2Δ cells we treated hxk2Δ and hxk2Δyca1Δ cells with different concentration of acetic acid (40ndash80ndash120mM) for 200min at 30∘C and measured the ROSaccumulation Our results showed that deletion of YCA1 inhxk2Δ cells had no effect on acetic-acid-induced ROS accu-mulation (Figure 6) suggesting that the generation of ROSin hxk2Δ cells upon acetic acid stress is Yca1 independentHowever the level of intracellular ROS was consistentlyhigher in the hxk2Δ yca1Δ doublemutant growing on glucosemedium compared with the hxk2Δ mutant Similarly Du etal [24] showed that formic acid induced Yca1-independentapoptosis-like cell death and that the burst of ROS duringcell death occurred earlier and at a higher level in the yca1Δstrain than in the wild-type strain Moreover Khan et al [41]determined the level of oxidized proteins in yeast cells underH2O2stress and showed that lack of Yca1 abrogated apoptosis
but elevated intracellular oxidized proteins compared with
wild-type cells Consequently both our results and data in theliterature [24 41] clearly suggest a linkage between ROS pro-duction and Yca1 activation during apoptosis in yeast Finallycytofluorometric quantification of phosphatidylserine exter-nalization andormembrane permeabilization (annexinVPIcostaining) further confirmed that Yca1p did not influencethe acetic-acid-induced cell death in hxk2Δ cells (Figure 7)These results strongly suggest that apoptosis induced by aceticacid in hxk2Δ cells is largely Yca1 independent
4 Conclusion
Several studies indicate that the Ras proteinmay be physicallyassociated with mitochondria both in yeast and mammals[10 11 42ndash44] This study provides results indicating thatassociation of active Ras to these organelles might be linkedto apoptosis First of all we show that addition of acetic acida well-known apoptotic stimulus in S cerevisiae to growingwild-type cells determined a delocalization of active Ras pro-teins from nuclei and plasma membrane to mitochondriaFurthermore we show that addition of acetic acid to hxk2Δcells showing a constitutive localization of active Ras inmito-chondria [9] enhanced ROS production and cell death
Oxidative Medicine and Cellular Longevity 9
compared with wild-type cells Moreover expression of thedominant active RAS2Val19 allele in the hxk2Δ backgroundcaused an even higher increase in the level of ROS both ingrowing cells and after treatment with acetic acid providinga further proof to the hypothesis that conditions that presum-ably cause a higher mitochondrial accumulation of activeRas may contribute to accumulation of ROS and cell deathWe also show that in hxk2Δ cells addition of acetic acidincreased cell death compared with the wild-type strain withthe typical markers of apoptosis highlighting a new rolefor Hexokinase 2 as an antiapoptotic factor in S cerevisiaecells However at this stage we cannot exclude a correlationbetween the well-known functions of Hxk2 in glucose repres-sion and signalling and the role of Ras in inducing apoptosis
Finally in this study we show that lack of Hxk2 inducesapoptosis via a mitochondria-mediated pathway withoutmetacaspase Yca1 involvement since deletion of YCA1 in thehxk2Δ background did not abrogate the acetic-acid-inducedaccumulation of ROS and did not decrease the percentage ofboth apoptotic and necrotic cells
Acknowledgments
The authors thank J Winderickx University of Leuven Bel-gium for the gift of strains and M Vai Universita Milano-Bicocca Milan for the gift of primers This work was sup-ported by FAR grant to Sonia Colombo and by founding ofProgram SysBioNet Italian Roadmap Research Infrastruc-ture 2012
References
[1] S JonesM-L Vignais and J R Broach ldquoTheCDC25 protein ofSaccharomyces cerevisiae promotes exchange of guanine nucle-otides bound to RasrdquoMolecular and Cellular Biology vol 11 no5 pp 2641ndash2646 1991
[2] S Rudoni S Colombo P Coccetti and E Martegani ldquoRole ofguanine nucleotides in the regulation of the RascAMPpathwayin Saccharomyces cerevisiaerdquo Biochimica et Biophysica Acta vol1538 no 2-3 pp 181ndash189 2001
[3] K TanakaMNakafuku T Satoh et al ldquoS cerevisiae genes IRA1and IRA2 encode proteins that may be functionally equivalentto mammalian ras GTPase activating proteinrdquo Cell vol 60 no5 pp 803ndash807 1990
[4] S Zaman S I Lippman X Zhao and J R Broach ldquoHow Sac-charomyces responds to nutrientsrdquo Annual Review of Geneticsvol 42 pp 27ndash81 2008
[5] JMThevelein and J H DeWinde ldquoNovel sensingmechanismsand targets for the cAMP-protein kinase A pathway in the yeastSaccharomyces cerevisiaerdquoMolecularMicrobiology vol 33 no 5pp 904ndash918 1999
[6] F Rolland JWinderickx and JMThevelein ldquoGlucose-sensingand -signallingmechanisms in yeastrdquo FEMS Yeast Research vol2 no 2 pp 183ndash201 2002
[7] G F Casperson N Walker and H R Bourne ldquoIsolation of thegene encoding adenylate cyclase in Saccharomyces cerevisiaerdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 82 no 15 pp 5060ndash5063 1985
[8] B Smets R Ghillebert P De Snijder et al ldquoLife in the midstof scarcity adaptations to nutrient availability in Saccharomycescerevisiaerdquo Current Genetics vol 56 no 1 pp 1ndash32 2010
[9] S Broggi E Martegani and S Colombo ldquoLive-cell imaging ofendogenous Ras-GTP shows predominant Ras activation at theplasma membrane and in the nucleus in Saccharomyces cere-visiaerdquoThe International Journal of Biochemistry amp Cell Biologyvol 45 no 2 pp 384ndash394 2013
[10] J E Leadsham K Miller K R Ayscough et al ldquoWhi2p linksnutritional sensing to actin-dependent Ras-cAMP-PKA regu-lation and apoptosis in yeastrdquo Journal of Cell Science vol 122no 5 pp 706ndash715 2009
[11] YHuW LuG Chen et al ldquoK-rasG12V transformation leads tomitochondrial dysfunction and a metabolic switch from oxida-tive phosphorylation to glycolysisrdquo Cell Research vol 22 no 2pp 399ndash412 2012
[12] S Buttner T Eisenberg D Carmona-Gutierrez et al ldquoEndonu-clease G regulates budding yeast life and deathrdquoMolecular Cellvol 25 no 2 pp 233ndash246 2007
[13] F Madeo E Frohlich and K-U Frohlich ldquoA yeast mutantshowing diagnostic markers of early and late apoptosisrdquo Journalof Cell Biology vol 139 no 3 pp 729ndash734 1997
[14] FMadeo E Herker SWissing H Jungwirth T Eisenberg andK-U Frohlich ldquoApoptosis in yeastrdquo Current Opinion in Micro-biology vol 7 no 6 pp 655ndash660 2004
[15] F Madeo E Herker C Maldener et al ldquoA caspase-related pro-tease regulates apoptosis in yeastrdquo Molecular Cell vol 9 no 4pp 911ndash917 2002
[16] N Guaragnella C Pereira M J Sousa et al ldquoYCA1 participatesin the acetic acid induced yeast programmed cell death also in amanner unrelated to its caspase-like activityrdquo FEBS Letters vol580 no 30 pp 6880ndash6884 2006
[17] W Li L Sun Q Liang J Wang W Mo and B Zhou ldquoYeastAMID homologue Ndi1p displays respiration-restricted apop-totic activity and is involved in chronological agingrdquoMolecularBiology of the Cell vol 17 no 4 pp 1802ndash1811 2006
[18] S Wissing P Ludovico E Herker et al ldquoAn AIF orthologueregulates apoptosis in yeastrdquo Journal of Cell Biology vol 166 no7 pp 969ndash974 2004
[19] B Fahrenkrog U Sauder and U Aebi ldquoThe S cerevisiae HtrA-like protein Nma111p is a nuclear serine protease that mediatesyeast apoptosisrdquo Journal of Cell Science vol 117 no 1 pp 115ndash126 2004
[20] J Qiu J-H Yoon and B Shen ldquoSearch for apoptotic nucleasesin yeast role of Tat-D nuclease in apoptotic DNA degradationrdquoThe Journal of Biological Chemistry vol 280 no 15 pp 15370ndash15379 2005
[21] I Orlandi M Bettiga L Alberghina and M Vai ldquoTranscrip-tional profiling of ubp10 null mutant reveals altered subtelo-meric gene expression and insurgence of oxidative stressresponserdquoThe Journal of Biological Chemistry vol 279 no 8 pp6414ndash6425 2004
[22] M Ligr F Madeo E Frohlich W Hilt K-U Frohlich and DH Wolf ldquoMammalian Bax triggers apoptotic changes in yeastrdquoFEBS Letters vol 438 no 1-2 pp 61ndash65 1998
[23] F Madeo E Frohlich M Ligr et al ldquoOxygen stress a regulatorof apoptosis in yeastrdquo Journal of Cell Biology vol 145 no 4 pp757ndash767 1999
[24] L Du Y Su D Sun et al ldquoFormic acid induces Yca1p-inde-pendent apoptosis-like cell death in the yeast Saccharomycescerevisiaerdquo FEMS Yeast Research vol 8 no 4 pp 531ndash539 2008
10 Oxidative Medicine and Cellular Longevity
[25] P LudovicoM J SousaM T Silva C Leao andMCorte-RealldquoSaccharomyces cerevisiae commits to a programmed cell deathprocess in response to acetic acidrdquoMicrobiology vol 147 no 9pp 2409ndash2415 2001
[26] P Ludovico F Rodrigues AAlmeidaMT Silva A Barrientosand M Corte-Real ldquoCytochrome c release and mitochondriainvolvement in programmed cell death induced by acetic acidin Saccharomyces cerevisiaerdquo Molecular Biology of the Cell vol13 no 8 pp 2598ndash2606 2002
[27] N Guaragnella L Antonacci S Passarella E Marra and SGiannattasio ldquoHydrogen peroxide and superoxide anion pro-duction during acetic acid-induced yeast programmed celldeathrdquo Folia Microbiologica vol 52 no 3 pp 237ndash240 2007
[28] S Giannattasio A Atlante L Antonacci et al ldquoCytochrome c isreleased from coupled mitochondria of yeast en route to aceticacid-induced programmed cell death and can work as an elec-tron donor and a ROS scavengerrdquo FEBS Letters vol 582 no 10pp 1519ndash1525 2008
[29] N Guaragnella L Antonacci S Giannattasio E Marra and SPassarella ldquoCatalase T and Cu Zn-superoxide dismutase in theacetic acid-induced programmed cell death in Saccharomycescerevisiaerdquo FEBS Letters vol 582 no 2 pp 210ndash214 2008
[30] N Guaragnella A Bobba S Passarella E Marra and S Gian-nattasio ldquoYeast acetic acid-induced programmed cell death canoccurwithout cytochrome c releasewhich requiresmetacaspaseYCA1rdquo FEBS Letters vol 584 no 1 pp 224ndash228 2010
[31] F Madeo D Carmona-Gutierrez J Ring S Buttner T Eisen-berg and G Kroemer ldquoCaspase-dependent and caspase-inde-pendent cell death pathways in yeastrdquo Biochemical and Biophys-ical Research Communications vol 382 no 2 pp 227ndash231 2009
[32] DAhuatzi A Riera R Pelaez PHerrero andFMoreno ldquoHxk2regulates the phosphorylation state of Mig1 and therefore itsnucleocytoplasmic distributionrdquo The Journal of BiologicalChemistry vol 282 no 7 pp 4485ndash4493 2007
[33] FMoreno D Ahuatzi A Riera C A Palomino and P HerreroldquoGlucose sensing through the Hxk2-dependent signalling path-wayrdquo Biochemical Society Transactions vol 33 no 1 pp 265ndash268 2005
[34] B JThomas and R Rothstein ldquoElevated recombination rates intranscriptionally active DNArdquo Cell vol 56 no 4 pp 619ndash6301989
[35] L C Robinson J B Gibbs M S Marshall I S Sigal and KTatchell ldquoCDC25 a component of the RAS-adenylate cyclasepathway in Saccharomyces cerevisiaerdquo Science vol 235 no 4793pp 1218ndash1221 1987
[36] R J Rothstein ldquoOne-step gene disruption in yeastrdquoMethods inEnzymology vol 101 pp 202ndash211 1983
[37] M Bettiga L Calzari I Orlandi L Alberghina and M VaildquoInvolvement of the yeast metacaspase Yca1 in ubp10Δ-pro-grammed cell deathrdquo FEMS Yeast Research vol 5 no 2 pp 141ndash147 2004
[38] N Casatta A Porro I Orlandi L Brambilla andM Vai ldquoLackof Sir2 increases acetate consumption and decreases extracellu-lar pro-aging factorsrdquo Biochimica et Biophysica Acta vol 1833no 3 pp 593ndash601 2013
[39] S Colombo D Ronchetti J M Thevelein J Winderickx andE Martegani ldquoActivation state of the Ras2 protein and glucose-induced signaling in Saccharomyces cerevisiaerdquo The Journal ofBiological Chemistry vol 279 no 45 pp 46715ndash46722 2004
[40] P Fabrizio and V D Longo ldquoChronological aging-inducedapoptosis in yeastrdquo Biochimica et Biophysica Acta vol 1783 no7 pp 1280ndash1285 2008
[41] M A S Khan P B Chock and E R Stadtman ldquoKnockout ofcaspase-like gene YCA1 abrogates apoptosis and elevates oxi-dized proteins in Saccharomyces cerevisiaerdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 102 no 48 pp 17326ndash17331 2005
[42] J CWolfman S M Planchon J Liao and AWolfman ldquoStruc-tural and functional consequences of c-N-Ras constitutivelyassociated with intact mitochondriardquo Biochimica et BiophysicaActa vol 1763 no 10 pp 1108ndash1124 2006
[43] T G Bivona S E Quatela B O Bodemann et al ldquoPKC regu-lates a farnesyl-electrostatic switch on K-Ras that promotes itsassociation with Bcl-XL on mitochondria and induces apopto-sisrdquoMolecular Cell vol 21 no 4 pp 481ndash493 2006
[44] A Rebollo D Perez-Sala and C Martınez-A ldquoBcl-2 differen-tially targets K- N- and H-Ras to mitochondria in IL-2 sup-plemented or deprived cells implications in prevention of apop-tosisrdquo Oncogene vol 18 no 35 pp 4930ndash4939 1999
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

6 Oxidative Medicine and Cellular Longevity
W303-1A
256
0
Fluorescence
Even
ts
100
101
102
103
104
0 mM
40 mM
80 mM
120 mM
Fluorescence
Even
ts
256
010
010
110
210
310
4
0 mM
40 mM
80 mM
120 mM
hxk2
(a)
W303-1A
0 mM 80 mM
hxk2
(b)
Figure 5 Mitochondrial membrane potential and morphology in W303-1A and hxk2Δ strains upon acetic acid treatment W303-1A andhxk2Δ exponentially growing cells were treated with different concentrations (40ndash80ndash120mM) of acetic acid for 200 minutes at 30∘C DiOC
6
uptake was assessed using both flow cytometry (a) and fluorescence microscopy (b) Photomicrographs illustrate the alteration of the tubularmitochondrial network to clustered mitochondrial morphology particularly exacerbated in hxk2Δ cells after acetic acid treatment
32 Deletion of HXK2 Enhances Cell Death and IncreasesIntracellular ROS Levels The results presented previouslysuggest that localization of active Ras proteins to mitochon-dria might be involved in programmed cell death since theseproteins are localized to mitochondria following additionof acetic acid Since in glucose-growing hxk2Δ cells activeRas is constitutively located in mitochondria [9] we nextanalyzed the behavior of the hxk2Δmutant under conditions
that caused apoptosisWhile the growth of hxk2Δmutant wasalmost indistinguishable from the wild-type strain at pH 53it exhibited an increased sensitivity to acetic acid stress at lowpH (Figure 2(a)) suggesting that Hxk2 is required for normaltolerance to acetic acid treatment Cell survival of wild-typeand hxk2Δ cells after induction of apoptosis with acetic acidwas further tested in a plating assay After treatment withdifferent concentrations (80 and 120mM) of acetic acid for
Oxidative Medicine and Cellular Longevity 7
up to 200 minutes at 30∘C hxk2Δ cells showed a significantdose-dependent reduction in cell survival when comparedwith wild-type cells (Figure 2(b)) Taken together these datasuggest that Hxk2 is required for normal tolerance to aceticacid treatment
Since an aberrant accumulation of activated Ras in mito-chondria accompanied to accumulation of ROS has alreadybeen reported both in mammals and yeast [10 11] and sinceROS play a pivotal role in apoptotic cell death FACS analyseswere performed to evaluate the accumulation of ROS in wild-type and hxk2Δ cells treated with different concentrationsof acetic acid for 200min at 30∘C In particular DHR123was used to determine the accumulation of ROS in the cellssince this compound can easily permeate them and can bequantitatively oxidized to a green fluorescent product inthe presence of ROS The percentage of oxidized R123 wasabout the double for hxk2Δ cells compared to wild-type cells200min after treatment with either 40 or 80mM acetic acid(Figure 3) To substantiate the hypothesis that an aberrantaccumulation of activated Ras in mitochondria might leadtomitochondrial dysfunction and accumulation of damagingROS we expressed the dominant active RAS2Val19 allelewhich was reported to show a much higher level of Ras2-GTP [39] in hxk2Δ cells Our results showed that indeed theexpression of this allele in the hxk2Δ background caused afurther increase in the level of ROS both in growing cells andafter treatment with acetic acid (Figure 3)These data suggestthat conditions that presumably cause a highermitochondrialaccumulation of active Ras may contribute to accumulationof ROS and cell death
33 Loss of Hxk2 Causes an Increase of Both Apoptosis andNecrosis The results presented previously suggest that Hxk2might have an antiapoptotic activity To better characterizethe nature of cell death triggered by addition of acetic acid tohxk2Δ cells we quantified phenotypic changes indicative ofapoptosis While apoptotic DNA condensation was detectedby DAPI staining combined Annexin Vpropidium iodide(PI) staining was used to discriminate between early apop-totic (Annexin V+PIminus) late apoptoticsecondary necrotic(Annexin V+PI+) and necrotic (Annexin VminusPI+) deathsDAPI staining showed that nuclei of untreated cells wereround while the nuclear DNA was condensed in bothwild-type and hxk2Δ cells treated with 80mM acetic acidbeing the extent of this phenotype more pronounced for themutant strain compared to the wild-type strain (data notshown) Annexin Vpropidium iodide (PI) staining revealedthat hxk2Δ-facilitated cell death was accompanied by anincrease in both apoptotic and necroticmarkers (Figure 4) Inparticular untreated hxk2Δ cells showed a higher percentageof both early apoptotic and necrotic cells compared withthe wild-type strain which further increased upon treatmentwith acetic acid
34 Deletion of HXK2 Causes Hyperpolarization of Mitochon-dria Mitochondrial membrane potential (ΔΨ
119898) is a useful
indicator of mitochondrial function To measure ΔΨ119898 we
used the cationic lipophylic dye DiOC6 which accumulates
0102030405060708090
100110
DH
R 12
3 (
)
Acetic acid
lowast
lowast
lowast
0 mM 40 mM 80 mM 120 mM
Figure 6 Effect of YCA1 deletion on ROS accumulation aftertreatment with acetic acid W303-1A (black bars) yca1Δ (blackhatched bars) hxk2Δ (gray bars) and hxk2Δ-yca1Δ (white bars)exponentially growing cells were treated with different concen-trations (40ndash80ndash120mM) of acetic acid for 200 minutes at 30∘CROS accumulation was assayed by flow cytometry using the dyedihydrorhodamine 123 (DHR123) Studentrsquos 119905-test lowast119875 lt 005
in mitochondria in accordance with ΔΨ119898 Wild-type and
hxk2Δ cells were treatedwith different concentration of aceticacid (40ndash80ndash120mM) for 200min at 30∘C collected andstained with this dye FACS analysis showed that mitochon-drial membrane potential clearly increased in both wild-typeand hxk2Δ cells after treatment with acetic acid comparedwith untreated control cells with the hyperpolarizationof mitochondria being much stronger in the mutant cellscomparedwithwild-type cells (Figure 5(a)) In parallel mito-chondrial morphology was assessed by using DiOC
6 both
in wild-type and hxk2Δ cells At low concentration this dyespecifically stains the mitochondrial membranes in a mannerthat depends on membrane potential and can be observed byfluorescence microscopy Before treatment with acetic acidbothwild-type and hxk2Δ cells displayed a tubularmitochon-drial morphology indicating that these mitochondria werehealthy and possessed a membrane potential (Figure 5(b))By contrast the mitochondrial membranes present in bothwild-type and hxk2Δ cells appeared rounded and highlyfragmented after treatment with acetic acid (Figure 5(b))This conversion of mitochondrial morphology from tubu-lar to punctuate structures is likely to occur by excessivemitochondrial fission and has already been observed in yeastapoptosis induced by acetic acid treatment [40] ImportantlyDiOC
6staining was greatly increased and more pronounced
in hxk2Δ cells compared to wild-type cells after treatmentwith acetic acid confirming the increase of fluorescenceobserved in these mutant cells by FACS analysis Thesedata indicate that both in wild-type and hxk2Δ cells aceticacid treatment caused hyperpolarization of mitochondrialmembrane with consequent damage of these organelles andloss of functionality This effect was more pronounced inhxk2Δ cells compared to wild-type cells
35 Acetic-Acid-Induced Cell Death in the hxk2Δ Strain Is Yca1Independent At least two death pathways can be activated
8 Oxidative Medicine and Cellular Longevity
lowastlowast
lowastlowast
lowast
lowastlowast
0 mM 40 mM 80 mM 120 mMAcetic acid
0
5
10
15
20
25
30
35
40A
nnex
in V
pos
(
)
(a)
lowast
lowast
lowastlowast
lowastlowast
0 mM 40 mM 80 mM 120 mM0
10
20
30
40
50
60
70
80
PI p
os (
)
Acetic acid
(b)
lowastlowast
lowast
lowast
0 mM 40 mM 80 mM 120 mM0
10
20
30
40
50
60
70
80
Acetic acid
Ann
exin
V+
PI p
os (
)
(c)
Figure 7 Effect of YCA1 deletion on cell death after treatment with acetic acid W303-1A (black bars) yca1Δ (black hatched bars) hxk2Δ(gray bars) and hxk2Δ-yca1Δ (white bars) exponentially growing cells were treated with different concentrations (40ndash80ndash120mM) of aceticacid for 200 minutes at 30∘C Cell death was assessed by flow cytometry using FITC-coupled annexin V and PI co-staining to determinatethe externalization of phosphatidylserine and the membrane integrityThemeans of 3 independent experiments with standard deviations arereported Studentrsquos 119905-test lowast119875 lt 005 and lowastlowast119875 lt 001
in yeast acetic-acid-induced apoptosis one dependent oncytochromec release which requires YCA1 and the other(s)independent of it [16 30] Consequently we next investigatedwhether the yeast caspase Yca1 might be involved in acetic-acid-induced cell death in hxk2Δ cells In particular toassess whether Yca1 plays a role in acetic-acid-induced ROSaccumulation in hxk2Δ cells we treated hxk2Δ and hxk2Δyca1Δ cells with different concentration of acetic acid (40ndash80ndash120mM) for 200min at 30∘C and measured the ROSaccumulation Our results showed that deletion of YCA1 inhxk2Δ cells had no effect on acetic-acid-induced ROS accu-mulation (Figure 6) suggesting that the generation of ROSin hxk2Δ cells upon acetic acid stress is Yca1 independentHowever the level of intracellular ROS was consistentlyhigher in the hxk2Δ yca1Δ doublemutant growing on glucosemedium compared with the hxk2Δ mutant Similarly Du etal [24] showed that formic acid induced Yca1-independentapoptosis-like cell death and that the burst of ROS duringcell death occurred earlier and at a higher level in the yca1Δstrain than in the wild-type strain Moreover Khan et al [41]determined the level of oxidized proteins in yeast cells underH2O2stress and showed that lack of Yca1 abrogated apoptosis
but elevated intracellular oxidized proteins compared with
wild-type cells Consequently both our results and data in theliterature [24 41] clearly suggest a linkage between ROS pro-duction and Yca1 activation during apoptosis in yeast Finallycytofluorometric quantification of phosphatidylserine exter-nalization andormembrane permeabilization (annexinVPIcostaining) further confirmed that Yca1p did not influencethe acetic-acid-induced cell death in hxk2Δ cells (Figure 7)These results strongly suggest that apoptosis induced by aceticacid in hxk2Δ cells is largely Yca1 independent
4 Conclusion
Several studies indicate that the Ras proteinmay be physicallyassociated with mitochondria both in yeast and mammals[10 11 42ndash44] This study provides results indicating thatassociation of active Ras to these organelles might be linkedto apoptosis First of all we show that addition of acetic acida well-known apoptotic stimulus in S cerevisiae to growingwild-type cells determined a delocalization of active Ras pro-teins from nuclei and plasma membrane to mitochondriaFurthermore we show that addition of acetic acid to hxk2Δcells showing a constitutive localization of active Ras inmito-chondria [9] enhanced ROS production and cell death
Oxidative Medicine and Cellular Longevity 9
compared with wild-type cells Moreover expression of thedominant active RAS2Val19 allele in the hxk2Δ backgroundcaused an even higher increase in the level of ROS both ingrowing cells and after treatment with acetic acid providinga further proof to the hypothesis that conditions that presum-ably cause a higher mitochondrial accumulation of activeRas may contribute to accumulation of ROS and cell deathWe also show that in hxk2Δ cells addition of acetic acidincreased cell death compared with the wild-type strain withthe typical markers of apoptosis highlighting a new rolefor Hexokinase 2 as an antiapoptotic factor in S cerevisiaecells However at this stage we cannot exclude a correlationbetween the well-known functions of Hxk2 in glucose repres-sion and signalling and the role of Ras in inducing apoptosis
Finally in this study we show that lack of Hxk2 inducesapoptosis via a mitochondria-mediated pathway withoutmetacaspase Yca1 involvement since deletion of YCA1 in thehxk2Δ background did not abrogate the acetic-acid-inducedaccumulation of ROS and did not decrease the percentage ofboth apoptotic and necrotic cells
Acknowledgments
The authors thank J Winderickx University of Leuven Bel-gium for the gift of strains and M Vai Universita Milano-Bicocca Milan for the gift of primers This work was sup-ported by FAR grant to Sonia Colombo and by founding ofProgram SysBioNet Italian Roadmap Research Infrastruc-ture 2012
References
[1] S JonesM-L Vignais and J R Broach ldquoTheCDC25 protein ofSaccharomyces cerevisiae promotes exchange of guanine nucle-otides bound to RasrdquoMolecular and Cellular Biology vol 11 no5 pp 2641ndash2646 1991
[2] S Rudoni S Colombo P Coccetti and E Martegani ldquoRole ofguanine nucleotides in the regulation of the RascAMPpathwayin Saccharomyces cerevisiaerdquo Biochimica et Biophysica Acta vol1538 no 2-3 pp 181ndash189 2001
[3] K TanakaMNakafuku T Satoh et al ldquoS cerevisiae genes IRA1and IRA2 encode proteins that may be functionally equivalentto mammalian ras GTPase activating proteinrdquo Cell vol 60 no5 pp 803ndash807 1990
[4] S Zaman S I Lippman X Zhao and J R Broach ldquoHow Sac-charomyces responds to nutrientsrdquo Annual Review of Geneticsvol 42 pp 27ndash81 2008
[5] JMThevelein and J H DeWinde ldquoNovel sensingmechanismsand targets for the cAMP-protein kinase A pathway in the yeastSaccharomyces cerevisiaerdquoMolecularMicrobiology vol 33 no 5pp 904ndash918 1999
[6] F Rolland JWinderickx and JMThevelein ldquoGlucose-sensingand -signallingmechanisms in yeastrdquo FEMS Yeast Research vol2 no 2 pp 183ndash201 2002
[7] G F Casperson N Walker and H R Bourne ldquoIsolation of thegene encoding adenylate cyclase in Saccharomyces cerevisiaerdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 82 no 15 pp 5060ndash5063 1985
[8] B Smets R Ghillebert P De Snijder et al ldquoLife in the midstof scarcity adaptations to nutrient availability in Saccharomycescerevisiaerdquo Current Genetics vol 56 no 1 pp 1ndash32 2010
[9] S Broggi E Martegani and S Colombo ldquoLive-cell imaging ofendogenous Ras-GTP shows predominant Ras activation at theplasma membrane and in the nucleus in Saccharomyces cere-visiaerdquoThe International Journal of Biochemistry amp Cell Biologyvol 45 no 2 pp 384ndash394 2013
[10] J E Leadsham K Miller K R Ayscough et al ldquoWhi2p linksnutritional sensing to actin-dependent Ras-cAMP-PKA regu-lation and apoptosis in yeastrdquo Journal of Cell Science vol 122no 5 pp 706ndash715 2009
[11] YHuW LuG Chen et al ldquoK-rasG12V transformation leads tomitochondrial dysfunction and a metabolic switch from oxida-tive phosphorylation to glycolysisrdquo Cell Research vol 22 no 2pp 399ndash412 2012
[12] S Buttner T Eisenberg D Carmona-Gutierrez et al ldquoEndonu-clease G regulates budding yeast life and deathrdquoMolecular Cellvol 25 no 2 pp 233ndash246 2007
[13] F Madeo E Frohlich and K-U Frohlich ldquoA yeast mutantshowing diagnostic markers of early and late apoptosisrdquo Journalof Cell Biology vol 139 no 3 pp 729ndash734 1997
[14] FMadeo E Herker SWissing H Jungwirth T Eisenberg andK-U Frohlich ldquoApoptosis in yeastrdquo Current Opinion in Micro-biology vol 7 no 6 pp 655ndash660 2004
[15] F Madeo E Herker C Maldener et al ldquoA caspase-related pro-tease regulates apoptosis in yeastrdquo Molecular Cell vol 9 no 4pp 911ndash917 2002
[16] N Guaragnella C Pereira M J Sousa et al ldquoYCA1 participatesin the acetic acid induced yeast programmed cell death also in amanner unrelated to its caspase-like activityrdquo FEBS Letters vol580 no 30 pp 6880ndash6884 2006
[17] W Li L Sun Q Liang J Wang W Mo and B Zhou ldquoYeastAMID homologue Ndi1p displays respiration-restricted apop-totic activity and is involved in chronological agingrdquoMolecularBiology of the Cell vol 17 no 4 pp 1802ndash1811 2006
[18] S Wissing P Ludovico E Herker et al ldquoAn AIF orthologueregulates apoptosis in yeastrdquo Journal of Cell Biology vol 166 no7 pp 969ndash974 2004
[19] B Fahrenkrog U Sauder and U Aebi ldquoThe S cerevisiae HtrA-like protein Nma111p is a nuclear serine protease that mediatesyeast apoptosisrdquo Journal of Cell Science vol 117 no 1 pp 115ndash126 2004
[20] J Qiu J-H Yoon and B Shen ldquoSearch for apoptotic nucleasesin yeast role of Tat-D nuclease in apoptotic DNA degradationrdquoThe Journal of Biological Chemistry vol 280 no 15 pp 15370ndash15379 2005
[21] I Orlandi M Bettiga L Alberghina and M Vai ldquoTranscrip-tional profiling of ubp10 null mutant reveals altered subtelo-meric gene expression and insurgence of oxidative stressresponserdquoThe Journal of Biological Chemistry vol 279 no 8 pp6414ndash6425 2004
[22] M Ligr F Madeo E Frohlich W Hilt K-U Frohlich and DH Wolf ldquoMammalian Bax triggers apoptotic changes in yeastrdquoFEBS Letters vol 438 no 1-2 pp 61ndash65 1998
[23] F Madeo E Frohlich M Ligr et al ldquoOxygen stress a regulatorof apoptosis in yeastrdquo Journal of Cell Biology vol 145 no 4 pp757ndash767 1999
[24] L Du Y Su D Sun et al ldquoFormic acid induces Yca1p-inde-pendent apoptosis-like cell death in the yeast Saccharomycescerevisiaerdquo FEMS Yeast Research vol 8 no 4 pp 531ndash539 2008
10 Oxidative Medicine and Cellular Longevity
[25] P LudovicoM J SousaM T Silva C Leao andMCorte-RealldquoSaccharomyces cerevisiae commits to a programmed cell deathprocess in response to acetic acidrdquoMicrobiology vol 147 no 9pp 2409ndash2415 2001
[26] P Ludovico F Rodrigues AAlmeidaMT Silva A Barrientosand M Corte-Real ldquoCytochrome c release and mitochondriainvolvement in programmed cell death induced by acetic acidin Saccharomyces cerevisiaerdquo Molecular Biology of the Cell vol13 no 8 pp 2598ndash2606 2002
[27] N Guaragnella L Antonacci S Passarella E Marra and SGiannattasio ldquoHydrogen peroxide and superoxide anion pro-duction during acetic acid-induced yeast programmed celldeathrdquo Folia Microbiologica vol 52 no 3 pp 237ndash240 2007
[28] S Giannattasio A Atlante L Antonacci et al ldquoCytochrome c isreleased from coupled mitochondria of yeast en route to aceticacid-induced programmed cell death and can work as an elec-tron donor and a ROS scavengerrdquo FEBS Letters vol 582 no 10pp 1519ndash1525 2008
[29] N Guaragnella L Antonacci S Giannattasio E Marra and SPassarella ldquoCatalase T and Cu Zn-superoxide dismutase in theacetic acid-induced programmed cell death in Saccharomycescerevisiaerdquo FEBS Letters vol 582 no 2 pp 210ndash214 2008
[30] N Guaragnella A Bobba S Passarella E Marra and S Gian-nattasio ldquoYeast acetic acid-induced programmed cell death canoccurwithout cytochrome c releasewhich requiresmetacaspaseYCA1rdquo FEBS Letters vol 584 no 1 pp 224ndash228 2010
[31] F Madeo D Carmona-Gutierrez J Ring S Buttner T Eisen-berg and G Kroemer ldquoCaspase-dependent and caspase-inde-pendent cell death pathways in yeastrdquo Biochemical and Biophys-ical Research Communications vol 382 no 2 pp 227ndash231 2009
[32] DAhuatzi A Riera R Pelaez PHerrero andFMoreno ldquoHxk2regulates the phosphorylation state of Mig1 and therefore itsnucleocytoplasmic distributionrdquo The Journal of BiologicalChemistry vol 282 no 7 pp 4485ndash4493 2007
[33] FMoreno D Ahuatzi A Riera C A Palomino and P HerreroldquoGlucose sensing through the Hxk2-dependent signalling path-wayrdquo Biochemical Society Transactions vol 33 no 1 pp 265ndash268 2005
[34] B JThomas and R Rothstein ldquoElevated recombination rates intranscriptionally active DNArdquo Cell vol 56 no 4 pp 619ndash6301989
[35] L C Robinson J B Gibbs M S Marshall I S Sigal and KTatchell ldquoCDC25 a component of the RAS-adenylate cyclasepathway in Saccharomyces cerevisiaerdquo Science vol 235 no 4793pp 1218ndash1221 1987
[36] R J Rothstein ldquoOne-step gene disruption in yeastrdquoMethods inEnzymology vol 101 pp 202ndash211 1983
[37] M Bettiga L Calzari I Orlandi L Alberghina and M VaildquoInvolvement of the yeast metacaspase Yca1 in ubp10Δ-pro-grammed cell deathrdquo FEMS Yeast Research vol 5 no 2 pp 141ndash147 2004
[38] N Casatta A Porro I Orlandi L Brambilla andM Vai ldquoLackof Sir2 increases acetate consumption and decreases extracellu-lar pro-aging factorsrdquo Biochimica et Biophysica Acta vol 1833no 3 pp 593ndash601 2013
[39] S Colombo D Ronchetti J M Thevelein J Winderickx andE Martegani ldquoActivation state of the Ras2 protein and glucose-induced signaling in Saccharomyces cerevisiaerdquo The Journal ofBiological Chemistry vol 279 no 45 pp 46715ndash46722 2004
[40] P Fabrizio and V D Longo ldquoChronological aging-inducedapoptosis in yeastrdquo Biochimica et Biophysica Acta vol 1783 no7 pp 1280ndash1285 2008
[41] M A S Khan P B Chock and E R Stadtman ldquoKnockout ofcaspase-like gene YCA1 abrogates apoptosis and elevates oxi-dized proteins in Saccharomyces cerevisiaerdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 102 no 48 pp 17326ndash17331 2005
[42] J CWolfman S M Planchon J Liao and AWolfman ldquoStruc-tural and functional consequences of c-N-Ras constitutivelyassociated with intact mitochondriardquo Biochimica et BiophysicaActa vol 1763 no 10 pp 1108ndash1124 2006
[43] T G Bivona S E Quatela B O Bodemann et al ldquoPKC regu-lates a farnesyl-electrostatic switch on K-Ras that promotes itsassociation with Bcl-XL on mitochondria and induces apopto-sisrdquoMolecular Cell vol 21 no 4 pp 481ndash493 2006
[44] A Rebollo D Perez-Sala and C Martınez-A ldquoBcl-2 differen-tially targets K- N- and H-Ras to mitochondria in IL-2 sup-plemented or deprived cells implications in prevention of apop-tosisrdquo Oncogene vol 18 no 35 pp 4930ndash4939 1999
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Oxidative Medicine and Cellular Longevity 7
up to 200 minutes at 30∘C hxk2Δ cells showed a significantdose-dependent reduction in cell survival when comparedwith wild-type cells (Figure 2(b)) Taken together these datasuggest that Hxk2 is required for normal tolerance to aceticacid treatment
Since an aberrant accumulation of activated Ras in mito-chondria accompanied to accumulation of ROS has alreadybeen reported both in mammals and yeast [10 11] and sinceROS play a pivotal role in apoptotic cell death FACS analyseswere performed to evaluate the accumulation of ROS in wild-type and hxk2Δ cells treated with different concentrationsof acetic acid for 200min at 30∘C In particular DHR123was used to determine the accumulation of ROS in the cellssince this compound can easily permeate them and can bequantitatively oxidized to a green fluorescent product inthe presence of ROS The percentage of oxidized R123 wasabout the double for hxk2Δ cells compared to wild-type cells200min after treatment with either 40 or 80mM acetic acid(Figure 3) To substantiate the hypothesis that an aberrantaccumulation of activated Ras in mitochondria might leadtomitochondrial dysfunction and accumulation of damagingROS we expressed the dominant active RAS2Val19 allelewhich was reported to show a much higher level of Ras2-GTP [39] in hxk2Δ cells Our results showed that indeed theexpression of this allele in the hxk2Δ background caused afurther increase in the level of ROS both in growing cells andafter treatment with acetic acid (Figure 3)These data suggestthat conditions that presumably cause a highermitochondrialaccumulation of active Ras may contribute to accumulationof ROS and cell death
33 Loss of Hxk2 Causes an Increase of Both Apoptosis andNecrosis The results presented previously suggest that Hxk2might have an antiapoptotic activity To better characterizethe nature of cell death triggered by addition of acetic acid tohxk2Δ cells we quantified phenotypic changes indicative ofapoptosis While apoptotic DNA condensation was detectedby DAPI staining combined Annexin Vpropidium iodide(PI) staining was used to discriminate between early apop-totic (Annexin V+PIminus) late apoptoticsecondary necrotic(Annexin V+PI+) and necrotic (Annexin VminusPI+) deathsDAPI staining showed that nuclei of untreated cells wereround while the nuclear DNA was condensed in bothwild-type and hxk2Δ cells treated with 80mM acetic acidbeing the extent of this phenotype more pronounced for themutant strain compared to the wild-type strain (data notshown) Annexin Vpropidium iodide (PI) staining revealedthat hxk2Δ-facilitated cell death was accompanied by anincrease in both apoptotic and necroticmarkers (Figure 4) Inparticular untreated hxk2Δ cells showed a higher percentageof both early apoptotic and necrotic cells compared withthe wild-type strain which further increased upon treatmentwith acetic acid
34 Deletion of HXK2 Causes Hyperpolarization of Mitochon-dria Mitochondrial membrane potential (ΔΨ
119898) is a useful
indicator of mitochondrial function To measure ΔΨ119898 we
used the cationic lipophylic dye DiOC6 which accumulates
0102030405060708090
100110
DH
R 12
3 (
)
Acetic acid
lowast
lowast
lowast
0 mM 40 mM 80 mM 120 mM
Figure 6 Effect of YCA1 deletion on ROS accumulation aftertreatment with acetic acid W303-1A (black bars) yca1Δ (blackhatched bars) hxk2Δ (gray bars) and hxk2Δ-yca1Δ (white bars)exponentially growing cells were treated with different concen-trations (40ndash80ndash120mM) of acetic acid for 200 minutes at 30∘CROS accumulation was assayed by flow cytometry using the dyedihydrorhodamine 123 (DHR123) Studentrsquos 119905-test lowast119875 lt 005
in mitochondria in accordance with ΔΨ119898 Wild-type and
hxk2Δ cells were treatedwith different concentration of aceticacid (40ndash80ndash120mM) for 200min at 30∘C collected andstained with this dye FACS analysis showed that mitochon-drial membrane potential clearly increased in both wild-typeand hxk2Δ cells after treatment with acetic acid comparedwith untreated control cells with the hyperpolarizationof mitochondria being much stronger in the mutant cellscomparedwithwild-type cells (Figure 5(a)) In parallel mito-chondrial morphology was assessed by using DiOC
6 both
in wild-type and hxk2Δ cells At low concentration this dyespecifically stains the mitochondrial membranes in a mannerthat depends on membrane potential and can be observed byfluorescence microscopy Before treatment with acetic acidbothwild-type and hxk2Δ cells displayed a tubularmitochon-drial morphology indicating that these mitochondria werehealthy and possessed a membrane potential (Figure 5(b))By contrast the mitochondrial membranes present in bothwild-type and hxk2Δ cells appeared rounded and highlyfragmented after treatment with acetic acid (Figure 5(b))This conversion of mitochondrial morphology from tubu-lar to punctuate structures is likely to occur by excessivemitochondrial fission and has already been observed in yeastapoptosis induced by acetic acid treatment [40] ImportantlyDiOC
6staining was greatly increased and more pronounced
in hxk2Δ cells compared to wild-type cells after treatmentwith acetic acid confirming the increase of fluorescenceobserved in these mutant cells by FACS analysis Thesedata indicate that both in wild-type and hxk2Δ cells aceticacid treatment caused hyperpolarization of mitochondrialmembrane with consequent damage of these organelles andloss of functionality This effect was more pronounced inhxk2Δ cells compared to wild-type cells
35 Acetic-Acid-Induced Cell Death in the hxk2Δ Strain Is Yca1Independent At least two death pathways can be activated
8 Oxidative Medicine and Cellular Longevity
lowastlowast
lowastlowast
lowast
lowastlowast
0 mM 40 mM 80 mM 120 mMAcetic acid
0
5
10
15
20
25
30
35
40A
nnex
in V
pos
(
)
(a)
lowast
lowast
lowastlowast
lowastlowast
0 mM 40 mM 80 mM 120 mM0
10
20
30
40
50
60
70
80
PI p
os (
)
Acetic acid
(b)
lowastlowast
lowast
lowast
0 mM 40 mM 80 mM 120 mM0
10
20
30
40
50
60
70
80
Acetic acid
Ann
exin
V+
PI p
os (
)
(c)
Figure 7 Effect of YCA1 deletion on cell death after treatment with acetic acid W303-1A (black bars) yca1Δ (black hatched bars) hxk2Δ(gray bars) and hxk2Δ-yca1Δ (white bars) exponentially growing cells were treated with different concentrations (40ndash80ndash120mM) of aceticacid for 200 minutes at 30∘C Cell death was assessed by flow cytometry using FITC-coupled annexin V and PI co-staining to determinatethe externalization of phosphatidylserine and the membrane integrityThemeans of 3 independent experiments with standard deviations arereported Studentrsquos 119905-test lowast119875 lt 005 and lowastlowast119875 lt 001
in yeast acetic-acid-induced apoptosis one dependent oncytochromec release which requires YCA1 and the other(s)independent of it [16 30] Consequently we next investigatedwhether the yeast caspase Yca1 might be involved in acetic-acid-induced cell death in hxk2Δ cells In particular toassess whether Yca1 plays a role in acetic-acid-induced ROSaccumulation in hxk2Δ cells we treated hxk2Δ and hxk2Δyca1Δ cells with different concentration of acetic acid (40ndash80ndash120mM) for 200min at 30∘C and measured the ROSaccumulation Our results showed that deletion of YCA1 inhxk2Δ cells had no effect on acetic-acid-induced ROS accu-mulation (Figure 6) suggesting that the generation of ROSin hxk2Δ cells upon acetic acid stress is Yca1 independentHowever the level of intracellular ROS was consistentlyhigher in the hxk2Δ yca1Δ doublemutant growing on glucosemedium compared with the hxk2Δ mutant Similarly Du etal [24] showed that formic acid induced Yca1-independentapoptosis-like cell death and that the burst of ROS duringcell death occurred earlier and at a higher level in the yca1Δstrain than in the wild-type strain Moreover Khan et al [41]determined the level of oxidized proteins in yeast cells underH2O2stress and showed that lack of Yca1 abrogated apoptosis
but elevated intracellular oxidized proteins compared with
wild-type cells Consequently both our results and data in theliterature [24 41] clearly suggest a linkage between ROS pro-duction and Yca1 activation during apoptosis in yeast Finallycytofluorometric quantification of phosphatidylserine exter-nalization andormembrane permeabilization (annexinVPIcostaining) further confirmed that Yca1p did not influencethe acetic-acid-induced cell death in hxk2Δ cells (Figure 7)These results strongly suggest that apoptosis induced by aceticacid in hxk2Δ cells is largely Yca1 independent
4 Conclusion
Several studies indicate that the Ras proteinmay be physicallyassociated with mitochondria both in yeast and mammals[10 11 42ndash44] This study provides results indicating thatassociation of active Ras to these organelles might be linkedto apoptosis First of all we show that addition of acetic acida well-known apoptotic stimulus in S cerevisiae to growingwild-type cells determined a delocalization of active Ras pro-teins from nuclei and plasma membrane to mitochondriaFurthermore we show that addition of acetic acid to hxk2Δcells showing a constitutive localization of active Ras inmito-chondria [9] enhanced ROS production and cell death
Oxidative Medicine and Cellular Longevity 9
compared with wild-type cells Moreover expression of thedominant active RAS2Val19 allele in the hxk2Δ backgroundcaused an even higher increase in the level of ROS both ingrowing cells and after treatment with acetic acid providinga further proof to the hypothesis that conditions that presum-ably cause a higher mitochondrial accumulation of activeRas may contribute to accumulation of ROS and cell deathWe also show that in hxk2Δ cells addition of acetic acidincreased cell death compared with the wild-type strain withthe typical markers of apoptosis highlighting a new rolefor Hexokinase 2 as an antiapoptotic factor in S cerevisiaecells However at this stage we cannot exclude a correlationbetween the well-known functions of Hxk2 in glucose repres-sion and signalling and the role of Ras in inducing apoptosis
Finally in this study we show that lack of Hxk2 inducesapoptosis via a mitochondria-mediated pathway withoutmetacaspase Yca1 involvement since deletion of YCA1 in thehxk2Δ background did not abrogate the acetic-acid-inducedaccumulation of ROS and did not decrease the percentage ofboth apoptotic and necrotic cells
Acknowledgments
The authors thank J Winderickx University of Leuven Bel-gium for the gift of strains and M Vai Universita Milano-Bicocca Milan for the gift of primers This work was sup-ported by FAR grant to Sonia Colombo and by founding ofProgram SysBioNet Italian Roadmap Research Infrastruc-ture 2012
References
[1] S JonesM-L Vignais and J R Broach ldquoTheCDC25 protein ofSaccharomyces cerevisiae promotes exchange of guanine nucle-otides bound to RasrdquoMolecular and Cellular Biology vol 11 no5 pp 2641ndash2646 1991
[2] S Rudoni S Colombo P Coccetti and E Martegani ldquoRole ofguanine nucleotides in the regulation of the RascAMPpathwayin Saccharomyces cerevisiaerdquo Biochimica et Biophysica Acta vol1538 no 2-3 pp 181ndash189 2001
[3] K TanakaMNakafuku T Satoh et al ldquoS cerevisiae genes IRA1and IRA2 encode proteins that may be functionally equivalentto mammalian ras GTPase activating proteinrdquo Cell vol 60 no5 pp 803ndash807 1990
[4] S Zaman S I Lippman X Zhao and J R Broach ldquoHow Sac-charomyces responds to nutrientsrdquo Annual Review of Geneticsvol 42 pp 27ndash81 2008
[5] JMThevelein and J H DeWinde ldquoNovel sensingmechanismsand targets for the cAMP-protein kinase A pathway in the yeastSaccharomyces cerevisiaerdquoMolecularMicrobiology vol 33 no 5pp 904ndash918 1999
[6] F Rolland JWinderickx and JMThevelein ldquoGlucose-sensingand -signallingmechanisms in yeastrdquo FEMS Yeast Research vol2 no 2 pp 183ndash201 2002
[7] G F Casperson N Walker and H R Bourne ldquoIsolation of thegene encoding adenylate cyclase in Saccharomyces cerevisiaerdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 82 no 15 pp 5060ndash5063 1985
[8] B Smets R Ghillebert P De Snijder et al ldquoLife in the midstof scarcity adaptations to nutrient availability in Saccharomycescerevisiaerdquo Current Genetics vol 56 no 1 pp 1ndash32 2010
[9] S Broggi E Martegani and S Colombo ldquoLive-cell imaging ofendogenous Ras-GTP shows predominant Ras activation at theplasma membrane and in the nucleus in Saccharomyces cere-visiaerdquoThe International Journal of Biochemistry amp Cell Biologyvol 45 no 2 pp 384ndash394 2013
[10] J E Leadsham K Miller K R Ayscough et al ldquoWhi2p linksnutritional sensing to actin-dependent Ras-cAMP-PKA regu-lation and apoptosis in yeastrdquo Journal of Cell Science vol 122no 5 pp 706ndash715 2009
[11] YHuW LuG Chen et al ldquoK-rasG12V transformation leads tomitochondrial dysfunction and a metabolic switch from oxida-tive phosphorylation to glycolysisrdquo Cell Research vol 22 no 2pp 399ndash412 2012
[12] S Buttner T Eisenberg D Carmona-Gutierrez et al ldquoEndonu-clease G regulates budding yeast life and deathrdquoMolecular Cellvol 25 no 2 pp 233ndash246 2007
[13] F Madeo E Frohlich and K-U Frohlich ldquoA yeast mutantshowing diagnostic markers of early and late apoptosisrdquo Journalof Cell Biology vol 139 no 3 pp 729ndash734 1997
[14] FMadeo E Herker SWissing H Jungwirth T Eisenberg andK-U Frohlich ldquoApoptosis in yeastrdquo Current Opinion in Micro-biology vol 7 no 6 pp 655ndash660 2004
[15] F Madeo E Herker C Maldener et al ldquoA caspase-related pro-tease regulates apoptosis in yeastrdquo Molecular Cell vol 9 no 4pp 911ndash917 2002
[16] N Guaragnella C Pereira M J Sousa et al ldquoYCA1 participatesin the acetic acid induced yeast programmed cell death also in amanner unrelated to its caspase-like activityrdquo FEBS Letters vol580 no 30 pp 6880ndash6884 2006
[17] W Li L Sun Q Liang J Wang W Mo and B Zhou ldquoYeastAMID homologue Ndi1p displays respiration-restricted apop-totic activity and is involved in chronological agingrdquoMolecularBiology of the Cell vol 17 no 4 pp 1802ndash1811 2006
[18] S Wissing P Ludovico E Herker et al ldquoAn AIF orthologueregulates apoptosis in yeastrdquo Journal of Cell Biology vol 166 no7 pp 969ndash974 2004
[19] B Fahrenkrog U Sauder and U Aebi ldquoThe S cerevisiae HtrA-like protein Nma111p is a nuclear serine protease that mediatesyeast apoptosisrdquo Journal of Cell Science vol 117 no 1 pp 115ndash126 2004
[20] J Qiu J-H Yoon and B Shen ldquoSearch for apoptotic nucleasesin yeast role of Tat-D nuclease in apoptotic DNA degradationrdquoThe Journal of Biological Chemistry vol 280 no 15 pp 15370ndash15379 2005
[21] I Orlandi M Bettiga L Alberghina and M Vai ldquoTranscrip-tional profiling of ubp10 null mutant reveals altered subtelo-meric gene expression and insurgence of oxidative stressresponserdquoThe Journal of Biological Chemistry vol 279 no 8 pp6414ndash6425 2004
[22] M Ligr F Madeo E Frohlich W Hilt K-U Frohlich and DH Wolf ldquoMammalian Bax triggers apoptotic changes in yeastrdquoFEBS Letters vol 438 no 1-2 pp 61ndash65 1998
[23] F Madeo E Frohlich M Ligr et al ldquoOxygen stress a regulatorof apoptosis in yeastrdquo Journal of Cell Biology vol 145 no 4 pp757ndash767 1999
[24] L Du Y Su D Sun et al ldquoFormic acid induces Yca1p-inde-pendent apoptosis-like cell death in the yeast Saccharomycescerevisiaerdquo FEMS Yeast Research vol 8 no 4 pp 531ndash539 2008
10 Oxidative Medicine and Cellular Longevity
[25] P LudovicoM J SousaM T Silva C Leao andMCorte-RealldquoSaccharomyces cerevisiae commits to a programmed cell deathprocess in response to acetic acidrdquoMicrobiology vol 147 no 9pp 2409ndash2415 2001
[26] P Ludovico F Rodrigues AAlmeidaMT Silva A Barrientosand M Corte-Real ldquoCytochrome c release and mitochondriainvolvement in programmed cell death induced by acetic acidin Saccharomyces cerevisiaerdquo Molecular Biology of the Cell vol13 no 8 pp 2598ndash2606 2002
[27] N Guaragnella L Antonacci S Passarella E Marra and SGiannattasio ldquoHydrogen peroxide and superoxide anion pro-duction during acetic acid-induced yeast programmed celldeathrdquo Folia Microbiologica vol 52 no 3 pp 237ndash240 2007
[28] S Giannattasio A Atlante L Antonacci et al ldquoCytochrome c isreleased from coupled mitochondria of yeast en route to aceticacid-induced programmed cell death and can work as an elec-tron donor and a ROS scavengerrdquo FEBS Letters vol 582 no 10pp 1519ndash1525 2008
[29] N Guaragnella L Antonacci S Giannattasio E Marra and SPassarella ldquoCatalase T and Cu Zn-superoxide dismutase in theacetic acid-induced programmed cell death in Saccharomycescerevisiaerdquo FEBS Letters vol 582 no 2 pp 210ndash214 2008
[30] N Guaragnella A Bobba S Passarella E Marra and S Gian-nattasio ldquoYeast acetic acid-induced programmed cell death canoccurwithout cytochrome c releasewhich requiresmetacaspaseYCA1rdquo FEBS Letters vol 584 no 1 pp 224ndash228 2010
[31] F Madeo D Carmona-Gutierrez J Ring S Buttner T Eisen-berg and G Kroemer ldquoCaspase-dependent and caspase-inde-pendent cell death pathways in yeastrdquo Biochemical and Biophys-ical Research Communications vol 382 no 2 pp 227ndash231 2009
[32] DAhuatzi A Riera R Pelaez PHerrero andFMoreno ldquoHxk2regulates the phosphorylation state of Mig1 and therefore itsnucleocytoplasmic distributionrdquo The Journal of BiologicalChemistry vol 282 no 7 pp 4485ndash4493 2007
[33] FMoreno D Ahuatzi A Riera C A Palomino and P HerreroldquoGlucose sensing through the Hxk2-dependent signalling path-wayrdquo Biochemical Society Transactions vol 33 no 1 pp 265ndash268 2005
[34] B JThomas and R Rothstein ldquoElevated recombination rates intranscriptionally active DNArdquo Cell vol 56 no 4 pp 619ndash6301989
[35] L C Robinson J B Gibbs M S Marshall I S Sigal and KTatchell ldquoCDC25 a component of the RAS-adenylate cyclasepathway in Saccharomyces cerevisiaerdquo Science vol 235 no 4793pp 1218ndash1221 1987
[36] R J Rothstein ldquoOne-step gene disruption in yeastrdquoMethods inEnzymology vol 101 pp 202ndash211 1983
[37] M Bettiga L Calzari I Orlandi L Alberghina and M VaildquoInvolvement of the yeast metacaspase Yca1 in ubp10Δ-pro-grammed cell deathrdquo FEMS Yeast Research vol 5 no 2 pp 141ndash147 2004
[38] N Casatta A Porro I Orlandi L Brambilla andM Vai ldquoLackof Sir2 increases acetate consumption and decreases extracellu-lar pro-aging factorsrdquo Biochimica et Biophysica Acta vol 1833no 3 pp 593ndash601 2013
[39] S Colombo D Ronchetti J M Thevelein J Winderickx andE Martegani ldquoActivation state of the Ras2 protein and glucose-induced signaling in Saccharomyces cerevisiaerdquo The Journal ofBiological Chemistry vol 279 no 45 pp 46715ndash46722 2004
[40] P Fabrizio and V D Longo ldquoChronological aging-inducedapoptosis in yeastrdquo Biochimica et Biophysica Acta vol 1783 no7 pp 1280ndash1285 2008
[41] M A S Khan P B Chock and E R Stadtman ldquoKnockout ofcaspase-like gene YCA1 abrogates apoptosis and elevates oxi-dized proteins in Saccharomyces cerevisiaerdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 102 no 48 pp 17326ndash17331 2005
[42] J CWolfman S M Planchon J Liao and AWolfman ldquoStruc-tural and functional consequences of c-N-Ras constitutivelyassociated with intact mitochondriardquo Biochimica et BiophysicaActa vol 1763 no 10 pp 1108ndash1124 2006
[43] T G Bivona S E Quatela B O Bodemann et al ldquoPKC regu-lates a farnesyl-electrostatic switch on K-Ras that promotes itsassociation with Bcl-XL on mitochondria and induces apopto-sisrdquoMolecular Cell vol 21 no 4 pp 481ndash493 2006
[44] A Rebollo D Perez-Sala and C Martınez-A ldquoBcl-2 differen-tially targets K- N- and H-Ras to mitochondria in IL-2 sup-plemented or deprived cells implications in prevention of apop-tosisrdquo Oncogene vol 18 no 35 pp 4930ndash4939 1999
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

8 Oxidative Medicine and Cellular Longevity
lowastlowast
lowastlowast
lowast
lowastlowast
0 mM 40 mM 80 mM 120 mMAcetic acid
0
5
10
15
20
25
30
35
40A
nnex
in V
pos
(
)
(a)
lowast
lowast
lowastlowast
lowastlowast
0 mM 40 mM 80 mM 120 mM0
10
20
30
40
50
60
70
80
PI p
os (
)
Acetic acid
(b)
lowastlowast
lowast
lowast
0 mM 40 mM 80 mM 120 mM0
10
20
30
40
50
60
70
80
Acetic acid
Ann
exin
V+
PI p
os (
)
(c)
Figure 7 Effect of YCA1 deletion on cell death after treatment with acetic acid W303-1A (black bars) yca1Δ (black hatched bars) hxk2Δ(gray bars) and hxk2Δ-yca1Δ (white bars) exponentially growing cells were treated with different concentrations (40ndash80ndash120mM) of aceticacid for 200 minutes at 30∘C Cell death was assessed by flow cytometry using FITC-coupled annexin V and PI co-staining to determinatethe externalization of phosphatidylserine and the membrane integrityThemeans of 3 independent experiments with standard deviations arereported Studentrsquos 119905-test lowast119875 lt 005 and lowastlowast119875 lt 001
in yeast acetic-acid-induced apoptosis one dependent oncytochromec release which requires YCA1 and the other(s)independent of it [16 30] Consequently we next investigatedwhether the yeast caspase Yca1 might be involved in acetic-acid-induced cell death in hxk2Δ cells In particular toassess whether Yca1 plays a role in acetic-acid-induced ROSaccumulation in hxk2Δ cells we treated hxk2Δ and hxk2Δyca1Δ cells with different concentration of acetic acid (40ndash80ndash120mM) for 200min at 30∘C and measured the ROSaccumulation Our results showed that deletion of YCA1 inhxk2Δ cells had no effect on acetic-acid-induced ROS accu-mulation (Figure 6) suggesting that the generation of ROSin hxk2Δ cells upon acetic acid stress is Yca1 independentHowever the level of intracellular ROS was consistentlyhigher in the hxk2Δ yca1Δ doublemutant growing on glucosemedium compared with the hxk2Δ mutant Similarly Du etal [24] showed that formic acid induced Yca1-independentapoptosis-like cell death and that the burst of ROS duringcell death occurred earlier and at a higher level in the yca1Δstrain than in the wild-type strain Moreover Khan et al [41]determined the level of oxidized proteins in yeast cells underH2O2stress and showed that lack of Yca1 abrogated apoptosis
but elevated intracellular oxidized proteins compared with
wild-type cells Consequently both our results and data in theliterature [24 41] clearly suggest a linkage between ROS pro-duction and Yca1 activation during apoptosis in yeast Finallycytofluorometric quantification of phosphatidylserine exter-nalization andormembrane permeabilization (annexinVPIcostaining) further confirmed that Yca1p did not influencethe acetic-acid-induced cell death in hxk2Δ cells (Figure 7)These results strongly suggest that apoptosis induced by aceticacid in hxk2Δ cells is largely Yca1 independent
4 Conclusion
Several studies indicate that the Ras proteinmay be physicallyassociated with mitochondria both in yeast and mammals[10 11 42ndash44] This study provides results indicating thatassociation of active Ras to these organelles might be linkedto apoptosis First of all we show that addition of acetic acida well-known apoptotic stimulus in S cerevisiae to growingwild-type cells determined a delocalization of active Ras pro-teins from nuclei and plasma membrane to mitochondriaFurthermore we show that addition of acetic acid to hxk2Δcells showing a constitutive localization of active Ras inmito-chondria [9] enhanced ROS production and cell death
Oxidative Medicine and Cellular Longevity 9
compared with wild-type cells Moreover expression of thedominant active RAS2Val19 allele in the hxk2Δ backgroundcaused an even higher increase in the level of ROS both ingrowing cells and after treatment with acetic acid providinga further proof to the hypothesis that conditions that presum-ably cause a higher mitochondrial accumulation of activeRas may contribute to accumulation of ROS and cell deathWe also show that in hxk2Δ cells addition of acetic acidincreased cell death compared with the wild-type strain withthe typical markers of apoptosis highlighting a new rolefor Hexokinase 2 as an antiapoptotic factor in S cerevisiaecells However at this stage we cannot exclude a correlationbetween the well-known functions of Hxk2 in glucose repres-sion and signalling and the role of Ras in inducing apoptosis
Finally in this study we show that lack of Hxk2 inducesapoptosis via a mitochondria-mediated pathway withoutmetacaspase Yca1 involvement since deletion of YCA1 in thehxk2Δ background did not abrogate the acetic-acid-inducedaccumulation of ROS and did not decrease the percentage ofboth apoptotic and necrotic cells
Acknowledgments
The authors thank J Winderickx University of Leuven Bel-gium for the gift of strains and M Vai Universita Milano-Bicocca Milan for the gift of primers This work was sup-ported by FAR grant to Sonia Colombo and by founding ofProgram SysBioNet Italian Roadmap Research Infrastruc-ture 2012
References
[1] S JonesM-L Vignais and J R Broach ldquoTheCDC25 protein ofSaccharomyces cerevisiae promotes exchange of guanine nucle-otides bound to RasrdquoMolecular and Cellular Biology vol 11 no5 pp 2641ndash2646 1991
[2] S Rudoni S Colombo P Coccetti and E Martegani ldquoRole ofguanine nucleotides in the regulation of the RascAMPpathwayin Saccharomyces cerevisiaerdquo Biochimica et Biophysica Acta vol1538 no 2-3 pp 181ndash189 2001
[3] K TanakaMNakafuku T Satoh et al ldquoS cerevisiae genes IRA1and IRA2 encode proteins that may be functionally equivalentto mammalian ras GTPase activating proteinrdquo Cell vol 60 no5 pp 803ndash807 1990
[4] S Zaman S I Lippman X Zhao and J R Broach ldquoHow Sac-charomyces responds to nutrientsrdquo Annual Review of Geneticsvol 42 pp 27ndash81 2008
[5] JMThevelein and J H DeWinde ldquoNovel sensingmechanismsand targets for the cAMP-protein kinase A pathway in the yeastSaccharomyces cerevisiaerdquoMolecularMicrobiology vol 33 no 5pp 904ndash918 1999
[6] F Rolland JWinderickx and JMThevelein ldquoGlucose-sensingand -signallingmechanisms in yeastrdquo FEMS Yeast Research vol2 no 2 pp 183ndash201 2002
[7] G F Casperson N Walker and H R Bourne ldquoIsolation of thegene encoding adenylate cyclase in Saccharomyces cerevisiaerdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 82 no 15 pp 5060ndash5063 1985
[8] B Smets R Ghillebert P De Snijder et al ldquoLife in the midstof scarcity adaptations to nutrient availability in Saccharomycescerevisiaerdquo Current Genetics vol 56 no 1 pp 1ndash32 2010
[9] S Broggi E Martegani and S Colombo ldquoLive-cell imaging ofendogenous Ras-GTP shows predominant Ras activation at theplasma membrane and in the nucleus in Saccharomyces cere-visiaerdquoThe International Journal of Biochemistry amp Cell Biologyvol 45 no 2 pp 384ndash394 2013
[10] J E Leadsham K Miller K R Ayscough et al ldquoWhi2p linksnutritional sensing to actin-dependent Ras-cAMP-PKA regu-lation and apoptosis in yeastrdquo Journal of Cell Science vol 122no 5 pp 706ndash715 2009
[11] YHuW LuG Chen et al ldquoK-rasG12V transformation leads tomitochondrial dysfunction and a metabolic switch from oxida-tive phosphorylation to glycolysisrdquo Cell Research vol 22 no 2pp 399ndash412 2012
[12] S Buttner T Eisenberg D Carmona-Gutierrez et al ldquoEndonu-clease G regulates budding yeast life and deathrdquoMolecular Cellvol 25 no 2 pp 233ndash246 2007
[13] F Madeo E Frohlich and K-U Frohlich ldquoA yeast mutantshowing diagnostic markers of early and late apoptosisrdquo Journalof Cell Biology vol 139 no 3 pp 729ndash734 1997
[14] FMadeo E Herker SWissing H Jungwirth T Eisenberg andK-U Frohlich ldquoApoptosis in yeastrdquo Current Opinion in Micro-biology vol 7 no 6 pp 655ndash660 2004
[15] F Madeo E Herker C Maldener et al ldquoA caspase-related pro-tease regulates apoptosis in yeastrdquo Molecular Cell vol 9 no 4pp 911ndash917 2002
[16] N Guaragnella C Pereira M J Sousa et al ldquoYCA1 participatesin the acetic acid induced yeast programmed cell death also in amanner unrelated to its caspase-like activityrdquo FEBS Letters vol580 no 30 pp 6880ndash6884 2006
[17] W Li L Sun Q Liang J Wang W Mo and B Zhou ldquoYeastAMID homologue Ndi1p displays respiration-restricted apop-totic activity and is involved in chronological agingrdquoMolecularBiology of the Cell vol 17 no 4 pp 1802ndash1811 2006
[18] S Wissing P Ludovico E Herker et al ldquoAn AIF orthologueregulates apoptosis in yeastrdquo Journal of Cell Biology vol 166 no7 pp 969ndash974 2004
[19] B Fahrenkrog U Sauder and U Aebi ldquoThe S cerevisiae HtrA-like protein Nma111p is a nuclear serine protease that mediatesyeast apoptosisrdquo Journal of Cell Science vol 117 no 1 pp 115ndash126 2004
[20] J Qiu J-H Yoon and B Shen ldquoSearch for apoptotic nucleasesin yeast role of Tat-D nuclease in apoptotic DNA degradationrdquoThe Journal of Biological Chemistry vol 280 no 15 pp 15370ndash15379 2005
[21] I Orlandi M Bettiga L Alberghina and M Vai ldquoTranscrip-tional profiling of ubp10 null mutant reveals altered subtelo-meric gene expression and insurgence of oxidative stressresponserdquoThe Journal of Biological Chemistry vol 279 no 8 pp6414ndash6425 2004
[22] M Ligr F Madeo E Frohlich W Hilt K-U Frohlich and DH Wolf ldquoMammalian Bax triggers apoptotic changes in yeastrdquoFEBS Letters vol 438 no 1-2 pp 61ndash65 1998
[23] F Madeo E Frohlich M Ligr et al ldquoOxygen stress a regulatorof apoptosis in yeastrdquo Journal of Cell Biology vol 145 no 4 pp757ndash767 1999
[24] L Du Y Su D Sun et al ldquoFormic acid induces Yca1p-inde-pendent apoptosis-like cell death in the yeast Saccharomycescerevisiaerdquo FEMS Yeast Research vol 8 no 4 pp 531ndash539 2008
10 Oxidative Medicine and Cellular Longevity
[25] P LudovicoM J SousaM T Silva C Leao andMCorte-RealldquoSaccharomyces cerevisiae commits to a programmed cell deathprocess in response to acetic acidrdquoMicrobiology vol 147 no 9pp 2409ndash2415 2001
[26] P Ludovico F Rodrigues AAlmeidaMT Silva A Barrientosand M Corte-Real ldquoCytochrome c release and mitochondriainvolvement in programmed cell death induced by acetic acidin Saccharomyces cerevisiaerdquo Molecular Biology of the Cell vol13 no 8 pp 2598ndash2606 2002
[27] N Guaragnella L Antonacci S Passarella E Marra and SGiannattasio ldquoHydrogen peroxide and superoxide anion pro-duction during acetic acid-induced yeast programmed celldeathrdquo Folia Microbiologica vol 52 no 3 pp 237ndash240 2007
[28] S Giannattasio A Atlante L Antonacci et al ldquoCytochrome c isreleased from coupled mitochondria of yeast en route to aceticacid-induced programmed cell death and can work as an elec-tron donor and a ROS scavengerrdquo FEBS Letters vol 582 no 10pp 1519ndash1525 2008
[29] N Guaragnella L Antonacci S Giannattasio E Marra and SPassarella ldquoCatalase T and Cu Zn-superoxide dismutase in theacetic acid-induced programmed cell death in Saccharomycescerevisiaerdquo FEBS Letters vol 582 no 2 pp 210ndash214 2008
[30] N Guaragnella A Bobba S Passarella E Marra and S Gian-nattasio ldquoYeast acetic acid-induced programmed cell death canoccurwithout cytochrome c releasewhich requiresmetacaspaseYCA1rdquo FEBS Letters vol 584 no 1 pp 224ndash228 2010
[31] F Madeo D Carmona-Gutierrez J Ring S Buttner T Eisen-berg and G Kroemer ldquoCaspase-dependent and caspase-inde-pendent cell death pathways in yeastrdquo Biochemical and Biophys-ical Research Communications vol 382 no 2 pp 227ndash231 2009
[32] DAhuatzi A Riera R Pelaez PHerrero andFMoreno ldquoHxk2regulates the phosphorylation state of Mig1 and therefore itsnucleocytoplasmic distributionrdquo The Journal of BiologicalChemistry vol 282 no 7 pp 4485ndash4493 2007
[33] FMoreno D Ahuatzi A Riera C A Palomino and P HerreroldquoGlucose sensing through the Hxk2-dependent signalling path-wayrdquo Biochemical Society Transactions vol 33 no 1 pp 265ndash268 2005
[34] B JThomas and R Rothstein ldquoElevated recombination rates intranscriptionally active DNArdquo Cell vol 56 no 4 pp 619ndash6301989
[35] L C Robinson J B Gibbs M S Marshall I S Sigal and KTatchell ldquoCDC25 a component of the RAS-adenylate cyclasepathway in Saccharomyces cerevisiaerdquo Science vol 235 no 4793pp 1218ndash1221 1987
[36] R J Rothstein ldquoOne-step gene disruption in yeastrdquoMethods inEnzymology vol 101 pp 202ndash211 1983
[37] M Bettiga L Calzari I Orlandi L Alberghina and M VaildquoInvolvement of the yeast metacaspase Yca1 in ubp10Δ-pro-grammed cell deathrdquo FEMS Yeast Research vol 5 no 2 pp 141ndash147 2004
[38] N Casatta A Porro I Orlandi L Brambilla andM Vai ldquoLackof Sir2 increases acetate consumption and decreases extracellu-lar pro-aging factorsrdquo Biochimica et Biophysica Acta vol 1833no 3 pp 593ndash601 2013
[39] S Colombo D Ronchetti J M Thevelein J Winderickx andE Martegani ldquoActivation state of the Ras2 protein and glucose-induced signaling in Saccharomyces cerevisiaerdquo The Journal ofBiological Chemistry vol 279 no 45 pp 46715ndash46722 2004
[40] P Fabrizio and V D Longo ldquoChronological aging-inducedapoptosis in yeastrdquo Biochimica et Biophysica Acta vol 1783 no7 pp 1280ndash1285 2008
[41] M A S Khan P B Chock and E R Stadtman ldquoKnockout ofcaspase-like gene YCA1 abrogates apoptosis and elevates oxi-dized proteins in Saccharomyces cerevisiaerdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 102 no 48 pp 17326ndash17331 2005
[42] J CWolfman S M Planchon J Liao and AWolfman ldquoStruc-tural and functional consequences of c-N-Ras constitutivelyassociated with intact mitochondriardquo Biochimica et BiophysicaActa vol 1763 no 10 pp 1108ndash1124 2006
[43] T G Bivona S E Quatela B O Bodemann et al ldquoPKC regu-lates a farnesyl-electrostatic switch on K-Ras that promotes itsassociation with Bcl-XL on mitochondria and induces apopto-sisrdquoMolecular Cell vol 21 no 4 pp 481ndash493 2006
[44] A Rebollo D Perez-Sala and C Martınez-A ldquoBcl-2 differen-tially targets K- N- and H-Ras to mitochondria in IL-2 sup-plemented or deprived cells implications in prevention of apop-tosisrdquo Oncogene vol 18 no 35 pp 4930ndash4939 1999
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Oxidative Medicine and Cellular Longevity 9
compared with wild-type cells Moreover expression of thedominant active RAS2Val19 allele in the hxk2Δ backgroundcaused an even higher increase in the level of ROS both ingrowing cells and after treatment with acetic acid providinga further proof to the hypothesis that conditions that presum-ably cause a higher mitochondrial accumulation of activeRas may contribute to accumulation of ROS and cell deathWe also show that in hxk2Δ cells addition of acetic acidincreased cell death compared with the wild-type strain withthe typical markers of apoptosis highlighting a new rolefor Hexokinase 2 as an antiapoptotic factor in S cerevisiaecells However at this stage we cannot exclude a correlationbetween the well-known functions of Hxk2 in glucose repres-sion and signalling and the role of Ras in inducing apoptosis
Finally in this study we show that lack of Hxk2 inducesapoptosis via a mitochondria-mediated pathway withoutmetacaspase Yca1 involvement since deletion of YCA1 in thehxk2Δ background did not abrogate the acetic-acid-inducedaccumulation of ROS and did not decrease the percentage ofboth apoptotic and necrotic cells
Acknowledgments
The authors thank J Winderickx University of Leuven Bel-gium for the gift of strains and M Vai Universita Milano-Bicocca Milan for the gift of primers This work was sup-ported by FAR grant to Sonia Colombo and by founding ofProgram SysBioNet Italian Roadmap Research Infrastruc-ture 2012
References
[1] S JonesM-L Vignais and J R Broach ldquoTheCDC25 protein ofSaccharomyces cerevisiae promotes exchange of guanine nucle-otides bound to RasrdquoMolecular and Cellular Biology vol 11 no5 pp 2641ndash2646 1991
[2] S Rudoni S Colombo P Coccetti and E Martegani ldquoRole ofguanine nucleotides in the regulation of the RascAMPpathwayin Saccharomyces cerevisiaerdquo Biochimica et Biophysica Acta vol1538 no 2-3 pp 181ndash189 2001
[3] K TanakaMNakafuku T Satoh et al ldquoS cerevisiae genes IRA1and IRA2 encode proteins that may be functionally equivalentto mammalian ras GTPase activating proteinrdquo Cell vol 60 no5 pp 803ndash807 1990
[4] S Zaman S I Lippman X Zhao and J R Broach ldquoHow Sac-charomyces responds to nutrientsrdquo Annual Review of Geneticsvol 42 pp 27ndash81 2008
[5] JMThevelein and J H DeWinde ldquoNovel sensingmechanismsand targets for the cAMP-protein kinase A pathway in the yeastSaccharomyces cerevisiaerdquoMolecularMicrobiology vol 33 no 5pp 904ndash918 1999
[6] F Rolland JWinderickx and JMThevelein ldquoGlucose-sensingand -signallingmechanisms in yeastrdquo FEMS Yeast Research vol2 no 2 pp 183ndash201 2002
[7] G F Casperson N Walker and H R Bourne ldquoIsolation of thegene encoding adenylate cyclase in Saccharomyces cerevisiaerdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 82 no 15 pp 5060ndash5063 1985
[8] B Smets R Ghillebert P De Snijder et al ldquoLife in the midstof scarcity adaptations to nutrient availability in Saccharomycescerevisiaerdquo Current Genetics vol 56 no 1 pp 1ndash32 2010
[9] S Broggi E Martegani and S Colombo ldquoLive-cell imaging ofendogenous Ras-GTP shows predominant Ras activation at theplasma membrane and in the nucleus in Saccharomyces cere-visiaerdquoThe International Journal of Biochemistry amp Cell Biologyvol 45 no 2 pp 384ndash394 2013
[10] J E Leadsham K Miller K R Ayscough et al ldquoWhi2p linksnutritional sensing to actin-dependent Ras-cAMP-PKA regu-lation and apoptosis in yeastrdquo Journal of Cell Science vol 122no 5 pp 706ndash715 2009
[11] YHuW LuG Chen et al ldquoK-rasG12V transformation leads tomitochondrial dysfunction and a metabolic switch from oxida-tive phosphorylation to glycolysisrdquo Cell Research vol 22 no 2pp 399ndash412 2012
[12] S Buttner T Eisenberg D Carmona-Gutierrez et al ldquoEndonu-clease G regulates budding yeast life and deathrdquoMolecular Cellvol 25 no 2 pp 233ndash246 2007
[13] F Madeo E Frohlich and K-U Frohlich ldquoA yeast mutantshowing diagnostic markers of early and late apoptosisrdquo Journalof Cell Biology vol 139 no 3 pp 729ndash734 1997
[14] FMadeo E Herker SWissing H Jungwirth T Eisenberg andK-U Frohlich ldquoApoptosis in yeastrdquo Current Opinion in Micro-biology vol 7 no 6 pp 655ndash660 2004
[15] F Madeo E Herker C Maldener et al ldquoA caspase-related pro-tease regulates apoptosis in yeastrdquo Molecular Cell vol 9 no 4pp 911ndash917 2002
[16] N Guaragnella C Pereira M J Sousa et al ldquoYCA1 participatesin the acetic acid induced yeast programmed cell death also in amanner unrelated to its caspase-like activityrdquo FEBS Letters vol580 no 30 pp 6880ndash6884 2006
[17] W Li L Sun Q Liang J Wang W Mo and B Zhou ldquoYeastAMID homologue Ndi1p displays respiration-restricted apop-totic activity and is involved in chronological agingrdquoMolecularBiology of the Cell vol 17 no 4 pp 1802ndash1811 2006
[18] S Wissing P Ludovico E Herker et al ldquoAn AIF orthologueregulates apoptosis in yeastrdquo Journal of Cell Biology vol 166 no7 pp 969ndash974 2004
[19] B Fahrenkrog U Sauder and U Aebi ldquoThe S cerevisiae HtrA-like protein Nma111p is a nuclear serine protease that mediatesyeast apoptosisrdquo Journal of Cell Science vol 117 no 1 pp 115ndash126 2004
[20] J Qiu J-H Yoon and B Shen ldquoSearch for apoptotic nucleasesin yeast role of Tat-D nuclease in apoptotic DNA degradationrdquoThe Journal of Biological Chemistry vol 280 no 15 pp 15370ndash15379 2005
[21] I Orlandi M Bettiga L Alberghina and M Vai ldquoTranscrip-tional profiling of ubp10 null mutant reveals altered subtelo-meric gene expression and insurgence of oxidative stressresponserdquoThe Journal of Biological Chemistry vol 279 no 8 pp6414ndash6425 2004
[22] M Ligr F Madeo E Frohlich W Hilt K-U Frohlich and DH Wolf ldquoMammalian Bax triggers apoptotic changes in yeastrdquoFEBS Letters vol 438 no 1-2 pp 61ndash65 1998
[23] F Madeo E Frohlich M Ligr et al ldquoOxygen stress a regulatorof apoptosis in yeastrdquo Journal of Cell Biology vol 145 no 4 pp757ndash767 1999
[24] L Du Y Su D Sun et al ldquoFormic acid induces Yca1p-inde-pendent apoptosis-like cell death in the yeast Saccharomycescerevisiaerdquo FEMS Yeast Research vol 8 no 4 pp 531ndash539 2008
10 Oxidative Medicine and Cellular Longevity
[25] P LudovicoM J SousaM T Silva C Leao andMCorte-RealldquoSaccharomyces cerevisiae commits to a programmed cell deathprocess in response to acetic acidrdquoMicrobiology vol 147 no 9pp 2409ndash2415 2001
[26] P Ludovico F Rodrigues AAlmeidaMT Silva A Barrientosand M Corte-Real ldquoCytochrome c release and mitochondriainvolvement in programmed cell death induced by acetic acidin Saccharomyces cerevisiaerdquo Molecular Biology of the Cell vol13 no 8 pp 2598ndash2606 2002
[27] N Guaragnella L Antonacci S Passarella E Marra and SGiannattasio ldquoHydrogen peroxide and superoxide anion pro-duction during acetic acid-induced yeast programmed celldeathrdquo Folia Microbiologica vol 52 no 3 pp 237ndash240 2007
[28] S Giannattasio A Atlante L Antonacci et al ldquoCytochrome c isreleased from coupled mitochondria of yeast en route to aceticacid-induced programmed cell death and can work as an elec-tron donor and a ROS scavengerrdquo FEBS Letters vol 582 no 10pp 1519ndash1525 2008
[29] N Guaragnella L Antonacci S Giannattasio E Marra and SPassarella ldquoCatalase T and Cu Zn-superoxide dismutase in theacetic acid-induced programmed cell death in Saccharomycescerevisiaerdquo FEBS Letters vol 582 no 2 pp 210ndash214 2008
[30] N Guaragnella A Bobba S Passarella E Marra and S Gian-nattasio ldquoYeast acetic acid-induced programmed cell death canoccurwithout cytochrome c releasewhich requiresmetacaspaseYCA1rdquo FEBS Letters vol 584 no 1 pp 224ndash228 2010
[31] F Madeo D Carmona-Gutierrez J Ring S Buttner T Eisen-berg and G Kroemer ldquoCaspase-dependent and caspase-inde-pendent cell death pathways in yeastrdquo Biochemical and Biophys-ical Research Communications vol 382 no 2 pp 227ndash231 2009
[32] DAhuatzi A Riera R Pelaez PHerrero andFMoreno ldquoHxk2regulates the phosphorylation state of Mig1 and therefore itsnucleocytoplasmic distributionrdquo The Journal of BiologicalChemistry vol 282 no 7 pp 4485ndash4493 2007
[33] FMoreno D Ahuatzi A Riera C A Palomino and P HerreroldquoGlucose sensing through the Hxk2-dependent signalling path-wayrdquo Biochemical Society Transactions vol 33 no 1 pp 265ndash268 2005
[34] B JThomas and R Rothstein ldquoElevated recombination rates intranscriptionally active DNArdquo Cell vol 56 no 4 pp 619ndash6301989
[35] L C Robinson J B Gibbs M S Marshall I S Sigal and KTatchell ldquoCDC25 a component of the RAS-adenylate cyclasepathway in Saccharomyces cerevisiaerdquo Science vol 235 no 4793pp 1218ndash1221 1987
[36] R J Rothstein ldquoOne-step gene disruption in yeastrdquoMethods inEnzymology vol 101 pp 202ndash211 1983
[37] M Bettiga L Calzari I Orlandi L Alberghina and M VaildquoInvolvement of the yeast metacaspase Yca1 in ubp10Δ-pro-grammed cell deathrdquo FEMS Yeast Research vol 5 no 2 pp 141ndash147 2004
[38] N Casatta A Porro I Orlandi L Brambilla andM Vai ldquoLackof Sir2 increases acetate consumption and decreases extracellu-lar pro-aging factorsrdquo Biochimica et Biophysica Acta vol 1833no 3 pp 593ndash601 2013
[39] S Colombo D Ronchetti J M Thevelein J Winderickx andE Martegani ldquoActivation state of the Ras2 protein and glucose-induced signaling in Saccharomyces cerevisiaerdquo The Journal ofBiological Chemistry vol 279 no 45 pp 46715ndash46722 2004
[40] P Fabrizio and V D Longo ldquoChronological aging-inducedapoptosis in yeastrdquo Biochimica et Biophysica Acta vol 1783 no7 pp 1280ndash1285 2008
[41] M A S Khan P B Chock and E R Stadtman ldquoKnockout ofcaspase-like gene YCA1 abrogates apoptosis and elevates oxi-dized proteins in Saccharomyces cerevisiaerdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 102 no 48 pp 17326ndash17331 2005
[42] J CWolfman S M Planchon J Liao and AWolfman ldquoStruc-tural and functional consequences of c-N-Ras constitutivelyassociated with intact mitochondriardquo Biochimica et BiophysicaActa vol 1763 no 10 pp 1108ndash1124 2006
[43] T G Bivona S E Quatela B O Bodemann et al ldquoPKC regu-lates a farnesyl-electrostatic switch on K-Ras that promotes itsassociation with Bcl-XL on mitochondria and induces apopto-sisrdquoMolecular Cell vol 21 no 4 pp 481ndash493 2006
[44] A Rebollo D Perez-Sala and C Martınez-A ldquoBcl-2 differen-tially targets K- N- and H-Ras to mitochondria in IL-2 sup-plemented or deprived cells implications in prevention of apop-tosisrdquo Oncogene vol 18 no 35 pp 4930ndash4939 1999
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

10 Oxidative Medicine and Cellular Longevity
[25] P LudovicoM J SousaM T Silva C Leao andMCorte-RealldquoSaccharomyces cerevisiae commits to a programmed cell deathprocess in response to acetic acidrdquoMicrobiology vol 147 no 9pp 2409ndash2415 2001
[26] P Ludovico F Rodrigues AAlmeidaMT Silva A Barrientosand M Corte-Real ldquoCytochrome c release and mitochondriainvolvement in programmed cell death induced by acetic acidin Saccharomyces cerevisiaerdquo Molecular Biology of the Cell vol13 no 8 pp 2598ndash2606 2002
[27] N Guaragnella L Antonacci S Passarella E Marra and SGiannattasio ldquoHydrogen peroxide and superoxide anion pro-duction during acetic acid-induced yeast programmed celldeathrdquo Folia Microbiologica vol 52 no 3 pp 237ndash240 2007
[28] S Giannattasio A Atlante L Antonacci et al ldquoCytochrome c isreleased from coupled mitochondria of yeast en route to aceticacid-induced programmed cell death and can work as an elec-tron donor and a ROS scavengerrdquo FEBS Letters vol 582 no 10pp 1519ndash1525 2008
[29] N Guaragnella L Antonacci S Giannattasio E Marra and SPassarella ldquoCatalase T and Cu Zn-superoxide dismutase in theacetic acid-induced programmed cell death in Saccharomycescerevisiaerdquo FEBS Letters vol 582 no 2 pp 210ndash214 2008
[30] N Guaragnella A Bobba S Passarella E Marra and S Gian-nattasio ldquoYeast acetic acid-induced programmed cell death canoccurwithout cytochrome c releasewhich requiresmetacaspaseYCA1rdquo FEBS Letters vol 584 no 1 pp 224ndash228 2010
[31] F Madeo D Carmona-Gutierrez J Ring S Buttner T Eisen-berg and G Kroemer ldquoCaspase-dependent and caspase-inde-pendent cell death pathways in yeastrdquo Biochemical and Biophys-ical Research Communications vol 382 no 2 pp 227ndash231 2009
[32] DAhuatzi A Riera R Pelaez PHerrero andFMoreno ldquoHxk2regulates the phosphorylation state of Mig1 and therefore itsnucleocytoplasmic distributionrdquo The Journal of BiologicalChemistry vol 282 no 7 pp 4485ndash4493 2007
[33] FMoreno D Ahuatzi A Riera C A Palomino and P HerreroldquoGlucose sensing through the Hxk2-dependent signalling path-wayrdquo Biochemical Society Transactions vol 33 no 1 pp 265ndash268 2005
[34] B JThomas and R Rothstein ldquoElevated recombination rates intranscriptionally active DNArdquo Cell vol 56 no 4 pp 619ndash6301989
[35] L C Robinson J B Gibbs M S Marshall I S Sigal and KTatchell ldquoCDC25 a component of the RAS-adenylate cyclasepathway in Saccharomyces cerevisiaerdquo Science vol 235 no 4793pp 1218ndash1221 1987
[36] R J Rothstein ldquoOne-step gene disruption in yeastrdquoMethods inEnzymology vol 101 pp 202ndash211 1983
[37] M Bettiga L Calzari I Orlandi L Alberghina and M VaildquoInvolvement of the yeast metacaspase Yca1 in ubp10Δ-pro-grammed cell deathrdquo FEMS Yeast Research vol 5 no 2 pp 141ndash147 2004
[38] N Casatta A Porro I Orlandi L Brambilla andM Vai ldquoLackof Sir2 increases acetate consumption and decreases extracellu-lar pro-aging factorsrdquo Biochimica et Biophysica Acta vol 1833no 3 pp 593ndash601 2013
[39] S Colombo D Ronchetti J M Thevelein J Winderickx andE Martegani ldquoActivation state of the Ras2 protein and glucose-induced signaling in Saccharomyces cerevisiaerdquo The Journal ofBiological Chemistry vol 279 no 45 pp 46715ndash46722 2004
[40] P Fabrizio and V D Longo ldquoChronological aging-inducedapoptosis in yeastrdquo Biochimica et Biophysica Acta vol 1783 no7 pp 1280ndash1285 2008
[41] M A S Khan P B Chock and E R Stadtman ldquoKnockout ofcaspase-like gene YCA1 abrogates apoptosis and elevates oxi-dized proteins in Saccharomyces cerevisiaerdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 102 no 48 pp 17326ndash17331 2005
[42] J CWolfman S M Planchon J Liao and AWolfman ldquoStruc-tural and functional consequences of c-N-Ras constitutivelyassociated with intact mitochondriardquo Biochimica et BiophysicaActa vol 1763 no 10 pp 1108ndash1124 2006
[43] T G Bivona S E Quatela B O Bodemann et al ldquoPKC regu-lates a farnesyl-electrostatic switch on K-Ras that promotes itsassociation with Bcl-XL on mitochondria and induces apopto-sisrdquoMolecular Cell vol 21 no 4 pp 481ndash493 2006
[44] A Rebollo D Perez-Sala and C Martınez-A ldquoBcl-2 differen-tially targets K- N- and H-Ras to mitochondria in IL-2 sup-plemented or deprived cells implications in prevention of apop-tosisrdquo Oncogene vol 18 no 35 pp 4930ndash4939 1999
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom












![Review Article - Hindawi Publishing Corporationdownloads.hindawi.com/journals/omcl/2019/9874159.pdf[33]. In another study, Roberts et al. reported a 25% reduced risk of MCI or dementia](https://img.pdfslide.us/doc/110x75/5f12eba9b32ade0284289ead/review-article-hindawi-publishing-33-in-another-study-roberts-et-al-reported.jpg)
















