Embed Size (px)
Citation preview

Ubiquitylation: its role and medical significance
Zoltán LipinszkiInstitute of Biochemistry
October 25, 2017
„Practice-oriented, student-friendly modernization of the biomedical education for strengthening the international competitiveness of the rural Hungarian universities”TÁMOP-4.1.1.C-13/1/KONV-2014-0001

• The proteins of the body are in a dynamic state of synthesis and degradation!– It is thought that we degrade and resynthesize ~3-5% of
our cellular proteins daily.• Paradigm that cellular processes are controlled
mainly by only transcription and translation must be changed.
Cellular homeostasisCellular homeostasis

Why are proteins degraded?
• Quality control– Proteins become denatured/misfolded/damaged
• Elevated temperatures (37°C)– Proteins being synthesized are folded incorrectly
• Regulation of biological pathways– Cell cycle– Receptor mediated endocytosis– Synaptic remodeling

Schoenheimer: proteins are in a dynamic turnover

De Duve: protein degradation in lysosomes

Energy dependence of protein degradation tyrosine aminotransferase

TABLE 1.- Resolution of the ATP-dependent cell-free proteolytic system into complementing activities
Az ATP-függő proteolitikus aktivitás frakcionálása DEAE-cellulóz kromatográfiával


"for the discovery of ubiquitin-mediated protein degradation"
Nobel Prize in Chemistry, 2004
Aaron Ciechanover Avram Hershko Irwin Rose

Discovery timeline of the ubiquitin proteasome system

Cyclin levels fluctuate during cell cycle

Mitotic cyclin destruction box
COOHH2N
Cyclin A
Cyclin B1Cyclin B2
Arg-Thr-Val-Leu-Gly-Val-Ile-Gly-Asp
Arg-Thr-Ala-Leu-Gly-Asp-Ile-Gly-Asn
Arg-Ala-Ala-Leu-Gly-Glu-Ile-Gly-Asn

Ubiquitin
composed of 76 amino acids
found only in eukaryotes
highly conserved
synthesized as a polyprotein
forms a heat-stable compact globular structure
exist either in free form or as attachment to other proteins
serves as a tag that marks proteins for degradation

1-MQIFVKTLTGKTITLEVESSDTIDNVKSKIQDKEGIPPDQQRLIF-451-MQIFVKTLTGKTITLEVESSDTIDNVKAKIQDKEGIPPDQQRLIF-451-MQIFVKTLTGKTITLEVEPSDTIENVKAKIQDKEGIPPDQQRLIF-451-MQIFVKTLTGKTITLEVEPSDTIENVKAKIQDKEGIPPDQQRLIF-45
46-AGKQLEDGRTLSDYNIQKESTLHLVLRLRGG-7646-AGKQLEDGRTLADYNIQKESTLHLVLRLRGG-7646-AGKQLEDGRTLSDYNIQKESTLHLVLRLRGG-7646-AGKQLEDGRTLSDYNIQKESTLHLVLRLRGG-76
Fission yeast humanGreen pea fruitfly
The structure of ubiquitin is conserved from yeast to human

Ubiquitin is synthesized as a polyprotein
Ubiquitin Ubiquitin Ubiquitin UbiquitinRibosomal protein
Transcription/Translation
Ub C-terminal hydrolase

Ubiquitin is conjugated to proteins
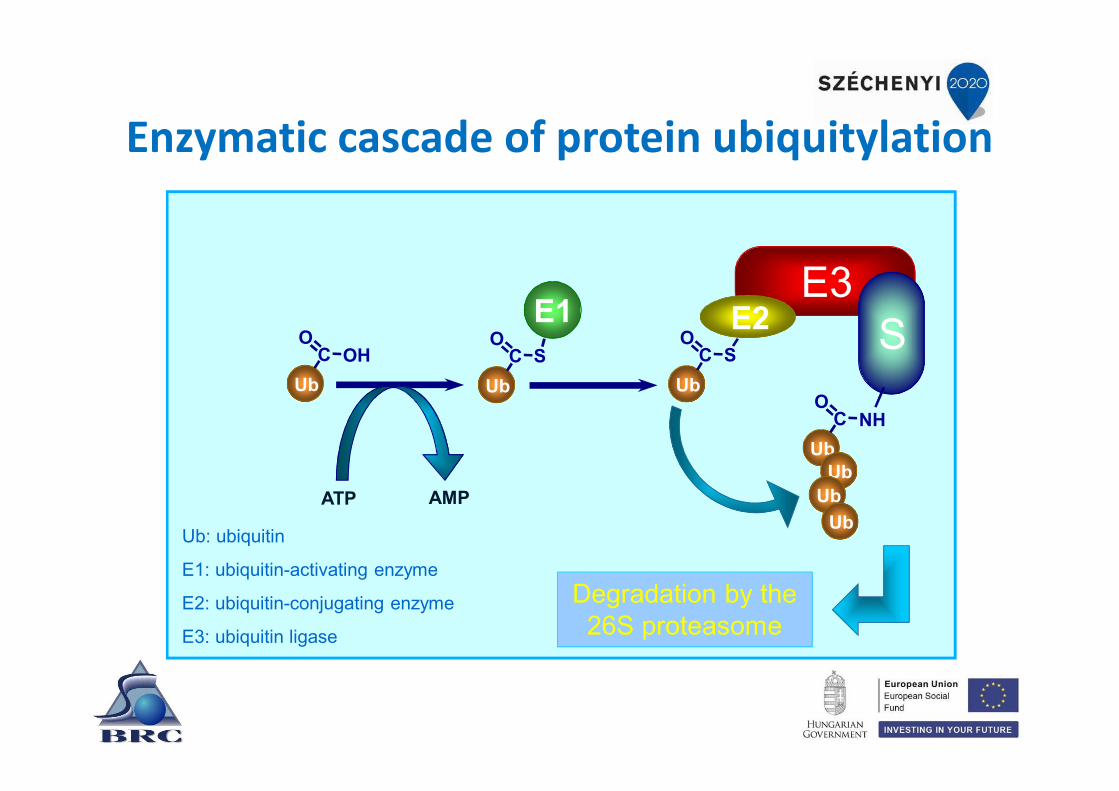
Enzymatic cascade of protein ubiquitylation
Ub: ubiquitin
E1: ubiquitin-activating enzyme
E2: ubiquitin-conjugating enzyme
E3: ubiquitin ligase
UbC OH
O
E3E2 SC S
O
UbC S
O
Ub
E1
ATP AMP
UbC NH
O
UbUb
Ub
Degradation by the 26S proteasome

The ubiquitin system is enormous
The genes of the UPS constitutes ~5% of the genome
• E1’s - 1-2 activating enzymes• E2’s - 10-20 conjugating enzymes• E3’s - 500-800 ubiquitin ligase - drives specificity• DUBs - 100 ubiquitin specific proteases

The hierarchical structure of the ubiquitin system

Subunit S. cerevisiae S. pombe Gerincesek Drosophila APC1 Apc1 Cut4 Apc1/Tsg24 shattered
APC2 Apc2/Rsi1 Apc2 Apc2 DmApc2
APC3 Cdc27 Nuc2 Apc3/Cdc27 mákos
APC4 Apc4 Cut20 Apc4 DmApc4
APC5 Apc5 Apc5 Apc5 ida
APC6 Cdc16 Cut9 Apc6/Cdc16 DmCdc16
APC7 --- --- Apc7 DmApc7
APC8 Cdc23 Cut23 Apc8/Cdc23 DmApc8
APC9 Apc9 --- --- ---
APC10 Apc10/Doc1 Apc10 Apc10 DmApc10
APC11 Apc11 Apc11 Apc11 lemming
APC12 Cdc26 Hcn1 Cdc26 Cdc26
APC13 Swm1 Apc13 Apc13 Apc13
The APC subunits are evolutionarily conserved

A speculative model of protein ubiquitylation by the APC

What about the protease?
• Previous studies demonstrated that the activity of the protease was ATP dependent (not just ubiquitination requires ATP)
• What is it composed of?• Where is it located?• How is it selective toward ubiquitinated
proteins?• Why does it need ATP?

Proteasomethe cellular chamber of doom
Composed of at least 64 subunits with a molecular mass of about 2.5 MDa
Barrel-shaped 20S catalytic core particle
Two 19S regulatory cap particles
Major substrates: polyubiquitylated proteins
Cleaves proteins in an ATP dependent manner
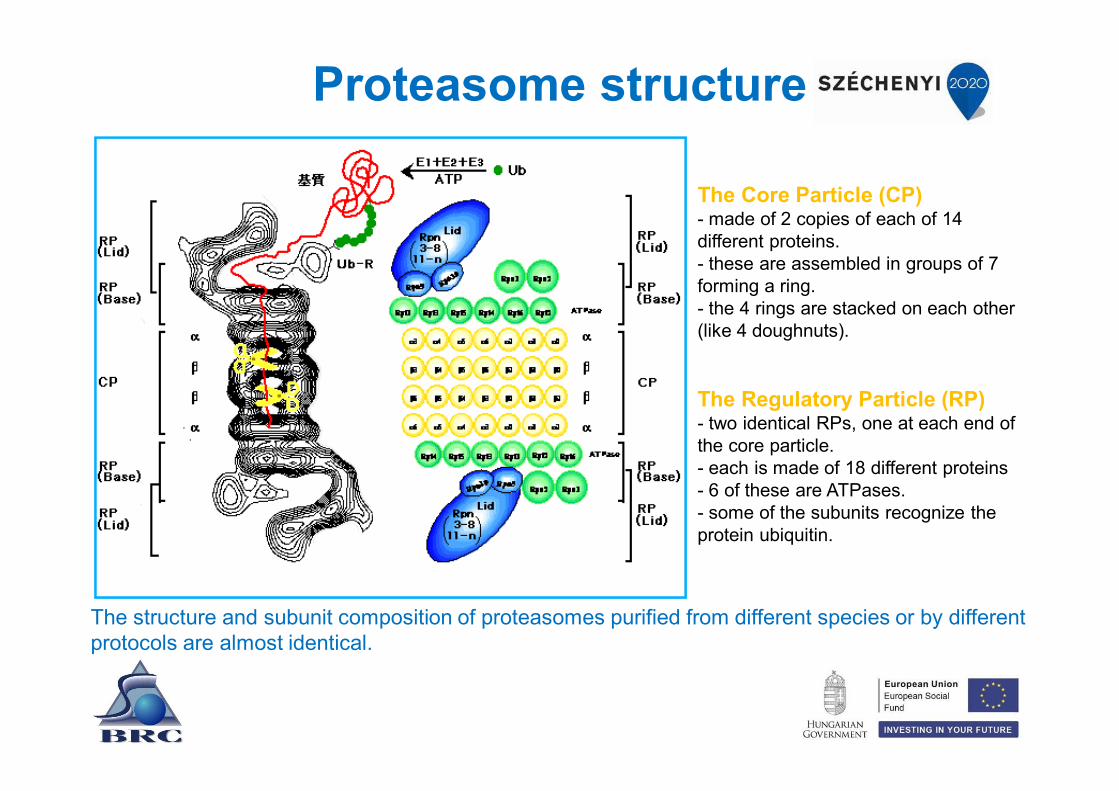
Proteasome structure
The structure and subunit composition of proteasomes purified from different species or by different protocols are almost identical.
The Core Particle (CP)- made of 2 copies of each of 14 different proteins. - these are assembled in groups of 7 forming a ring. - the 4 rings are stacked on each other (like 4 doughnuts).
The Regulatory Particle (RP)- two identical RPs, one at each end of the core particle. - each is made of 18 different proteins - 6 of these are ATPases. - some of the subunits recognize the protein ubiquitin.

Mechanism of protein degradation
- The complex binds to ubiquitin-recognizing site(s) on the regulatory particle.
- The protein is unfolded by the ATPases using the energy of ATP.
- The unfolded protein is translocated into the central cavity of the core particle.
- Several active sites on the inner surface of the two middle "doughnuts" break various specific peptide bonds of the chain.
- This produces a set of peptides averaging about 8 amino acids long. These leave the core particle
- The regulatory particle releases the ubiquitins for reuse.

The active sites of the proteasome

Proteasomes degrade proteins in a highly processive fashion
"bite-chew" model
active sites with chymotrypsin, trypsin and caspase-like activities
active sites work in an organized manner
active sites regulate each other's activity
products are small oligopeptides with 3-20 residues

Since many substrate proteins and manyprocesses are involved in ubiquitylation, it is notsurprising that malfunctions of the ubiquitin-proteasome system have been implicated –directly or indirectly – in the etiology of manyinherited and acquired human diseases.
Ubiquitin system and disease

Ubiquitylation is reversible
ProteinProtein
DUBsDeubiquitylating enzymes
E1, E2, E3Ubiquitylating enzymes

Deubiquitylating enzymes (DUBs)Deubiquitylating enzymes (DUBs)
Cysteine-proteasesMetallo-
proteases

Catalytic functions of DUBs

Processes regulated by ubiquitin-mediated protein degradation
ApoptosisCell cycle
Development
Stress response
Protein degradation
Antigen presentation
InflammationCancer
Neuronal abnormalities
Metabolism
transcription
DNA repair
Signaltransduction

Human pathologies resulting from disorders in protein degradation
Mohammed AliPope John Paul II Michael J. Fox

Parkinson disease
synuclein synuclein
parkin
parkin
Dopamine producingcells die

von Hippel-Lindau syndrome
• Caused by mutation of the VHL tumor suppressor gene.
• This leads to the development of angio-blastoma in the CNS and the retina.
• Also leads to the development of cysts in kidneys and renal cell carcinoma.

The VHL protein regulates the degradation of HIF transcription factor
HIFVHL
oxygen
hypoxia inducible factor
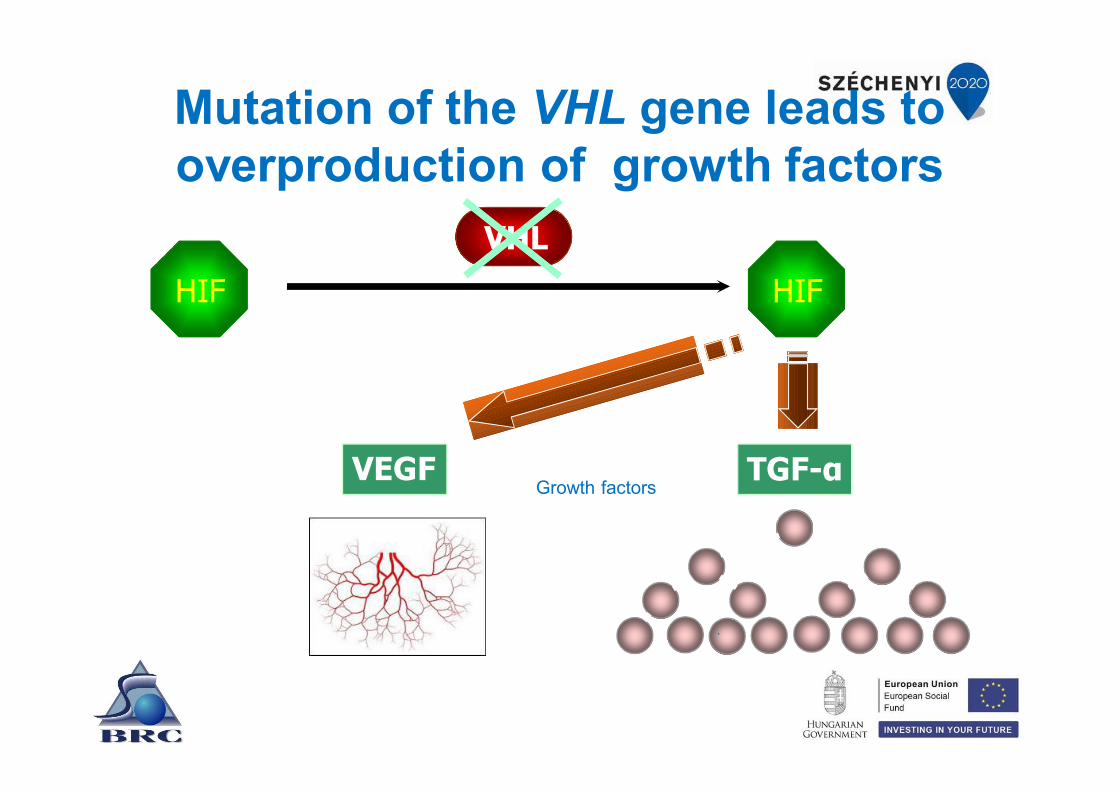
Mutation of the VHL gene leads to overproduction of growth factors
HIF HIF
VHL
VEGF TGF-αGrowth factors

Thank you for your attention!
This work is supported by the European Union, co-financed by the European Social Fund, within the framework of " Practice-
oriented, student-friendly modernization of the biomedical education for strengthening the international
competitiveness of the rural Hungarian universities " TÁMOP-4.1.1.C-13/1/KONV-2014-0001 project.

Diverse biological roles of protein ubiquitination
Gabriella EndreInstitute of Genetics
October 25, 2017
„Practice-oriented, student-friendly modernization of the biomedical education for strengthening the international competitiveness of the rural Hungarian universities”TÁMOP-4.1.1.C-13/1/KONV-2014-0001

Ubiquitin with purple Lysine Residues
Ubiquitin
Ubiquitous in nature (hence the name)
Small polypeptide of 76 AA
Highly conserved in evolution: 3 AA differences between yeast and human homologues
Can mono or poly-ubiquinate
Lys-48 linked poly-ubiquitin chains common - proteasome
1975 - molecule

Ubiquitin processing and conjugation
early 1980s – pathway
and Irwin Rose University of California, Irvine, USA
Nobel Prize in Chemistry 2004
Aaron Ciechanover, Avram HershkoTechnion – Israel Institute of Technology, Haifa, Israel

Ubiquitin conjugation to substrates
ATP+
E1
E1
Activating
E2
Conjugating Ligating
E2
E2RING E3
E2HECT E3
substrate
substrate
12
substrate
ActivatingE1
ConjugatingE2
LigatingE3
Ub: general term here for monoubiquitin any or polyubiquitin chain

UPSUbiquitin - Proteasome System
UMSUbiquitin - Modification System
Degradation of proteins in proteosome
Alteredlongevity localizationand/or activity
of proteins

Ubiquitin labelling is not always fatal for the protein!
several non-proteolytic functions associated with the addition of
- a single ubiquitin molecule (mono-ubiquitination)
- or specific cases of polyubiquitination
affecting the substrate’s
cellular sub-location,
function
or its degradation through lysosomes

Ub modifications with their functional roles

Fates of ubiquitinated proteins
A: Monoubiquitinated integral membrane proteins are internalizedB: Polyubiquitinated integral membrane proteins can be degraded either by vacuoles or, via ERAD, the proteasomeC: Polyubiquitinated soluble proteins are proteasomally degraded (Lys-48) or play roles in DNA repair (Lys-63) D: Monoubiquitination and multiubiquitination of soluble proteins can lead to activation or inhibition of a protein’s activity
Guerra & Callis 2012 Plant Physiol160: 56–64

Example for the versatility of ubiquitination
I.
Regulation of Transcription Factors by ubiquitination

Regulating Transcription Factors (TFs) by ubiquitination
Three strategiesby controlling the
of the transcription factors
localization
activity
abundance

Regulating TFs by ubiquitinationlocalization
TF can be kept
outside the
nucleus by
interactions with
an inhibitor that
can be destroyed
by the UPS upon
a signal (like TF
NFκB and its
inhibitor IκB)
Another Ub-
family member
SUMO (S) can
directly
conjugate to
activators and
sequester them
into nuclear
bodies
I

Can regulate the association of activators with co-activator proteins
Regulating TFs by ubiquitination
either directly:
by blocking the
association of an
activator with its
essential cofactor
or indirectly:
by facilitating
the exchange
of cofactors
with
an activator
activity

abundance
I - constitutive turnovermaintaining an activator in a constitutively unstable form -prompt transcriptional response when appropriate signal comes
II - trx-coupled destructionactivators are destroyed during the act of transcriptional activation as a way of limiting uncontrolled activation by any one DNA bound TF
Regulating TFs by ubiquitination

Numerous effects of ubiquitinationin one TF pathway: NF-kB
identified E3 ligases
identified DUBs
Wertz and Dixit, 2010

Mechanisms for Modulating Substrate Recognition by E3s
Pickart, 2004 Cell 116: 181–190,
posttranslational modificationsand other mechanisms known to regulate the recognition of cognate substrates by different E3s.

Wertz and Dixit, 2010
Example of the numerous effects of ubiquitination in one TF pathway
Lys-48-linked ubiquitin chains – redLys-63-linked ubiquitin chains - green

Example for the versatility of ubiquitination
II.
UbK63 Modification of Endocytic Cargoes

Examples of trafficking steps that involve UbK63 chains
Several channels, transporters and receptors undergo modification by UbK63 chains These endocytic cargoes are recognized for sorting to invaginated regions of the plasma membrane by a number of UBD-containing proteins.
Erpapazoglou et al., 2014 Cells

Example for the versatility of ubiquitination
III.
K63-Linked Ubiquitination in Selective Autophagy

Erpapazoglou et al., 2014 Cells
UbK63 chains positively regulate the autophagic clearance of aggresomes, mitochondria andintracellular bacteria by interfering with various steps of the process.
Involvement of UbK63 chains in selective autophagy

HOW the versatility of ubiquitinationis achieved?
Modulating Substrate Recognition by E3s

Mechanisms for Modulating Substrate Recognition by E3s
Pickart & Eddins 2004 BBA 1695: 55–72
Budding yeast
1 – E1
11 – E2
42 – E3U-box subclass: 2
>>E2/E3combinations
Plant Arabidopsis
2– E1
34– E2
>1300 – E3U-box subclass: 64
>>>>E2/E3combinations

Variety of domain compositions and organization for plant U-box proteins
Yee & Goring 2009 J. Exp. Botany

Many examples for the versatility of ubiquitination
in plants
hormone signalling
tailoring morphogenesis
responses to environmental challenges
self recognition (pollination)
battling pathogens
…

Roles of specific E3s in hormone signalling and photoperiod measurement
Vierstra, 2009 Nature Reviews

Control of self-incompatibility in flowers by the ubiquitin–26S proteasome system
Vierstra, 2009 Nature Reviews

Scheuring et al., 2009 BMC Plant Biol.
Sorting of plasma membrane and Golgi proteins into the vacuolar
degradation pathway
Model illustrating ubiquitin-mediated vacuolar transport of membrane proteins

Scheuring et al., 2009 BMC Plant Biol.
Ubiquitin causes internalization of a non-secretory reporter at the PM
Expression of non-secretory reporters to analyze the endocytic pathway.

Scheuring et al., 2009 BMC Plant Biol.
Ubiquitin causes a plasma membrane protein to traffic to the vacuole
Ubiquitin-dependent transport of an integral PM protein to the vacuole.

Scheuring et al., 2009 BMC Plant Biol.
Ubiquitin directs Golgi-localized proteins to the vacuole
Ubiquitin-dependent transport from the Golgi to the vacuole.
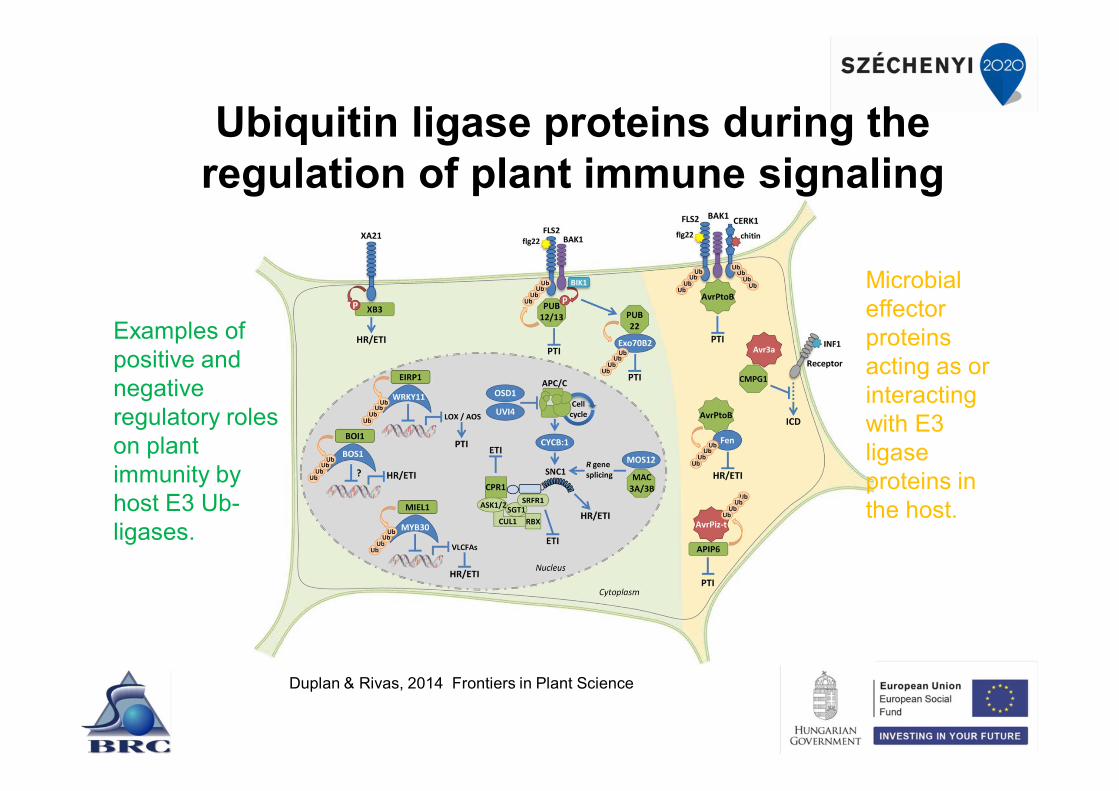
Ubiquitin ligase proteins during the regulation of plant immune signaling
Duplan & Rivas, 2014 Frontiers in Plant Science
Microbial effector proteins acting as or interacting with E3 ligase proteins in the host.
Examples of positive and negative regulatory roles on plant immunity by host E3 Ub-ligases.

Microbial effector proteins acting as ubiquitin E3 ligases???
YES!
Pathogens have “learned” how to (try to) fool the host

Some pathogen effectors interferingwith the plant ubiquitin system
Marino et al., 2012 Plant Physiol.
U-box and F-box effector proteins interfering directly or indirectly with the host UPS are color coded according to the pathogenicorganism and respectively representedby U and F symbols.

Control of Agrobacterium infection by VirF
Vierstra, 2009 Nature Reviews
During the pathogenesis cycle, a single-stranded DNA (T-strand) is synthesized from the Ti plasmid, coated withvirulence protein-E2 (VirE2) and transported into the plant host along with the VirF protein through the type-3secretion system (T3SS)

Microbial effector proteins acting as ubiquitin E3 ligases???
YES!
Pathogens have “learned” how to (try to) fool the host
Not only with plants, butGeneral feature

Manipulation of host ubiquitin pathwaysby Salmonella
Perrett et al., 2011 Frontiers in Microbiol.

Manipulation of host ubiquitin pathwaysby Yersinia
Perrett et al., 2011 Frontiers in Microbiol.

UBIQUITINATION
A VERSATILE POSTTRANSLATIONAL MODIFICATION

Thank you for your attention!
This work is supported by the European Union, co-financed by the European Social Fund, within the framework of " Practice-
oriented, student-friendly modernization of the biomedical education for strengthening the international
competitiveness of the rural Hungarian universities " TÁMOP-4.1.1.C-13/1/KONV-2014-0001 project.





























