Embed Size (px)
Citation preview

Jorrrnnl of Nrrirodirnristn Raven Press. New York 0 19R6 International Society for Neurochemistry
Human Skeletal Muscle Cells Synthesise a Neuronotrophic Factor Reactive with Spinal Neurons
Patrick Doherty, John G. Dickson, Thomas P. Flanigan, and Frank S. Walsh
lnstitrcte of Neurology, London, England
Abstract: Retrograde trophic influences originating in the skeletal musculature have been postulated to be involved in regulating survival and differentiation of embryonic motor neurons and reactive terminal sprouting of mature motor fibres. We have previously described the use of a quantitative immunoassay for neurofilament protein t o bioassay in vitro the cell-type-specific neuronotrophic ac- tivity of nerve growth factor (NGF) on sensory ganglion neurons. In the present study, the effect of media con- ditioned by adult human muscle cells (MCM) on the in vitro development of chicken spinal neurons has been studied using a similar approach. Significant increases in neurofilament protein levels in 7-day chicken embryonic spinal cord cultures were found with doses of MCM pro-
tein as low as 0.4 bg/ml, with a dose-response relation- ship yielding maximal and half-maximal effects at 4 and I pg/ml, respectively. Maximal increases in neurofilarnent protein levels were associated with an approximate two- fold increase in neuronal cell survival. MCM also induced increases in choline acetyltransferase activity in chick spinal cord cultures. In both the absence and presence of NGF, M C M did not increase neurofilament protein expression in primary cultures of sensory neurons. Key Words: Spinal neurons-Cell culture-Immunoassay- Immunofluorescence - Neurofilaments - Neuronotro- phic factor. Doherty P. et al. Human skeletal muscle cells synthesise a neuronotrophic factor reactive with spinal neurons. J . Neurochem. 46, 133-139 (1986).
The development of the vertebrate nervous system is accompanied by an overproduction of many cell types that is subsequently regulated by the phenomenon of naturally occurring neuronal cell death (Jacobson, 1978; Hamburger and Oppen- heim, 1982). This phenomenon, which has now been described for many neurons, including ct- motor neurons, has been attributed at least in part to a competition for a limiting supply of a target tissue-derived trophic factor (Purves, 1980; Brown et al., 1981; Hamburger and Oppenheim, 1982). Support for this model of retrograde trophic control has come from many studies that have demon- strated a clear relationship between the size of the postsynaptic field and the relative amount of neu- ronal cell death. For example, during development of the chick motor system, the natural cell death of motor neurons can be reduced by grafting on extra limb tissue or alternatively increased by extirpation of limb tissue (Hamburger, 1958; Hollyday and Hamburger, 1976).
The identification and characterisation of the ef- fects of nerve growth factor (NGF) on the devel- opment of the sympathetic and sensory nervous system, and in particular its ability to reduce both natural and experimentally induced cell death (Thoenen and Barde, 1980; Yanker and Shooter, 1982) further supports the concept that the avail- ability of a muscle-derived growth factor may be required for the survival and morphological devel- opment of embryonic motor neurons, as well as per- haps the maintenance of function of the adult motor system.
The successful application of in vitro cell culture techniques in the characterisation of the effects of NGF on neuronal cell survival and morphological differentiation has led to the adoption of similar bioassays in an attempt to identify a muscle-derived trophic activity (Barde et al., 1983).
We have previously demonstrated that an en- zyme-linked immunoadsorbent assay (ELISA) for neurofilament protein can be used to determine an
Received May 9, 1985; accepted July 4. 1985. Address correspondence and reprint requests 10 Dr. P. Do-
herty at Institute of Neurology, Department of Neurochemistry, Queen Square, London, WCIN 3BG. U.K.
Abbreviations used: ChAT, choline acetyltransferase: DMEM, Dulbecco's modified medium; DRG, dorsal root ganglia; ELISA,
enzyme-linked immunoadsorbent assay; FCS, foetal calf serum; HRP-SAM. horseradish peroxidase-conjugated sheep anti- mouse immunoglobulin; HS, horse serum; McAb, monoclonal antibody; MCM. muscle cell conditioned media; MNGF. motor neuron growth factor; NGF. nerve growth factor; OPD, o-phen- ylenediamine; PBS, phosphate-buffered saline.
I33

134 P. DOHERTY ET AL.
arbitrary index of neuronal cell survival and mor- phological differentiation in primary cultures initi- a ted from both human and chick nervous tissue (Doherty et al., 1984a,b). Considering the demon- stration of the target cell-specific nature of the ef- fect of NGF on neurofilament protein expression as a model for classical neuronotrophic activity (Do- herty et al., 1984b). we have utilised this ELISA assay to determine whether human skeletal muscle cell conditioned media (MCM) contains an activity directed against spinal as compared with sensory neuronal cultures. A preliminary account of this work has been reported (Doherty et al., 1985).
MATERIALS AND METHODS
Antibody reagents The neurofilament monoclonal antibody RT97 was ob-
tained as ascites fluids (Anderton et al., 1982). Horse- radish peroxidase-conjugated sheep anti-mouse immu- noglobulin (HRP-SAM) was prepared by the method of Nakane and Kawaoi (1974) using affinity-purified anti- bodies reactive with mouse immunoglobulin (Ig). Rho- damine-conjugated sheep anti-mouse Ig was prepared as previously described (Dickson et al., 1982) and was used at a 1:20 dilution. All antibody dilutions for ELISA and indirect immunofluorescence were into phosphate-buff- ered saline (PBS, containing per litre: NaCI, 8.5 g; KCI, 0.4 g; Na,HPO,, 0.71 g; glucose, 4.5 g; sucrose, 8.58 g; CaCl * 2H,O, 0.26 g; MgSO, * 7H,O, 0.20 g; adjusted to pH 7.2 with 5 M HCI) supplemented with 10% (vol/vol) heat-inactivated foetal calf serum (FCS).
Preparation of human skeletal muscle conditioned media
A muscle biopsy sample obtained from an 18-month- old male was dissociated and grown in primary culture on gelatin-coated tissue culture plates according to Yasin et al. (1977). Mononucleate cell proliferation was stimu- lated by the presence of 10% heat-inactivated FCS in Dulbecco's moditied Eagle's medium (DMEM). After 5 days, when cultures were approximately 70% confluent, media were changed to DMEM supplemented with 5% heat-inactivated horse serum (HS) and 0.5% FCS to fa- cilitate myoblast fusion into multinucleate myotubes. After a further 3 days the cell cultures were washed three times with serum-free medium and batches of standard MCM prepared. Fifteen-millilitre aliquots of DMEM were incubated with myotube cultures for four sequential 24-h periods and harvested. The MCM was stored at 4"C, pooled, and concentrated approximately 10 times by using an ultratidtration membrane with a molecular weight exclusion limit of 10,OOO. The concentrate was brought back to volume with fresh DMEM and reconcentrated twice. A final, approximately 7.5-fold concentrated stock solution of MCM contained 40 pg/ml protein. Unconcen- trated MCM was collected by incubating cultures with a single 15-1111 aliquot of DMEM for a continuous 6-day period. The protein concentration of the soluble fraction, collected following centrifugation at 9OOO g for 45 min was 25 pg/ml. Both batches of MCM were sterilized by pas- sage through a 0.2-pm filter, aliquoted, and stored in liquid nitrogen. Immediately prior to use, aliquots were thawed and heat-inactivated HS added to a final concen-
tration of 5-10% (vol/vol). Unless otherwise stated, all experiments were conducted with the concentrated MCM. MCM was added to culture wells immediately prior to cell seeding.
Cell culture of sensory and spinal neurons Dorsal root ganglia (DRG) obtained from 9-day chick
embryos and spinal cords from 7-day embryos were dis- sected free of meningeal tissue and dissociated by treat- ment with 0.05% (wtivol) trypsin at 37°C for 30 min. This was followed by trituration with a 19-gauge needle in cul- ture media that consisted of DMEM supplemented with 10% HS.
All primary cultures were initiated by seeding 100 FI of a dissociated cell suspension in DMEM supplemented with 7.5% HS into individual wells of a collagen- or col- lagen/polylysine-coated Linbro microtitre plate, each containing 100 p,l of culture media with twice the NGF andor MCM present. Seeding cell numbers were 0.5 X 106/ml for chick DRG, and 1 x 106/ml for chick spinal cord. Cytosine arabinoside (lo-' M) was added to all cul- tures after 2 days to suppress the growth of nonneuronal cells. Cultures were maintained at 37°C under a water- saturated atmosphere of 90% air, 10% CO,.
Indirect immunofluorescence and cell counts of neurofilament-positive cells
Indirect immunofluorescence staining was carried out as previously described (Dickson et al., 1982; Doherty et al., 19846) using monoclonal antibody RT97 to identify and localise neurofilaments in cultures. Cultures grown in 35-mm tissue culture dishes were processed identically as for neurofilament assay up to and including incubation with a 1:200 dilution of RT97 ascitic fluid. Cultures were then washed three times with PBS containing 10% FCS and incubated with rhodamine-conjugated sheep anti- mouse Ig for 60 min at room temperature. After further washing, coverslips were mounted on the cultures in 50% (vol/vol) glycerol in PBS and cells viewed on a Leitz Di- alux microscope. Counts of neurofilament-positive pro- cess bearing cells were taken from a series of represen- tative fields from these cultures.
Quantitation of neurofilament protein in neural cell cultures
The ELISA assay for neurofilament protein has been described in detail elsewhere (Doherty et al., 19840,h). Briefly, cell cultures were fixed and permeabilised with 4% paraformaldehyde and 0.2% Triton X-100 in PBS, re- spectively. Following incubation with PBS containing 10% FCS to block nonspecific protein binding sites, cul- tures were incubated for 60 rnin at 20°C with a 1:500 di- lution of RT97. Cultures were washed twice with PBS supplemented with FCS and incubated with a 1500 di- lution of HRP-SAM for a further 60 min. Cultures were then washed three times with PBS, twice with distilled H20, and finally incubated with 50 pI of 0.2% (wt/vol) O- phenylenediamine (OPD) and 0.02% (vol/vol) H,Oz in ci- trate buffer. After 30 min the conversion of OPD to its oxidised product was stopped by the addition of 50 FI of 4.5 M H2S0,. Aliquots of the reaction mixtures were then transferred to a fresh microtitre plate and product for- mation determined by optical density measurements at 492 nm using a Flow Titretek Multiscan apparatus. Ap- propriate control incubation mixtures were used as ref-
1. Neurochern.. Vol. 46, No. 1. 1986
SPINAL NEURON NEURONOTROPHIC FACTOR 135
erence samples. A linear relationship between oxidised OPD concentration and optical density, and a linear rate of production of oxidised OPD by the HRP-SAM with time were found under all experimental conditions.
Choline acetyltransferase assay Cultures prepared in 16-mm wells were rinsed twice
with PBS and lysed with 90 *I of lysis buffer consisting of 14.6 g of NaC1.6.8 g of KH,PO,. 0.15 g of EDTA, and 5 ml of Triton X-100 (pH 7.4) per litre. Each culture was scraped, transferred into an Eppendorf microfuge tube, and sonicated for 20 s , and centrifuged at 9,000 g for 2 min. Choline acetyltransferase (ChAT; EC 2.3.1.6.) was assayed in the supernatant by the method of Fonnum (1975).
RESULTS
MCM induces a n increase in neuronal survival and neurofilarnent protein levels in chick spinal cord cell cultures
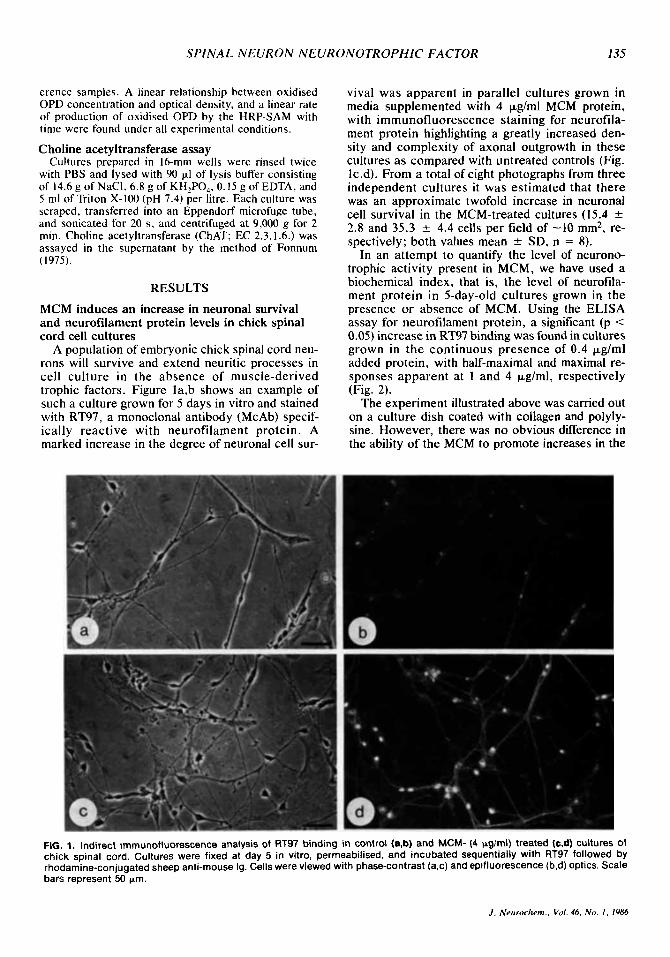
A population of embryonic chick spinal cord neu- rons will survive and extend neuritic processes in cell culture in the absence of muscle-derived trophic factors. Figure la,b shows an example of such a culture grown for 5 days in vitro and stained with RT97, a monoclonal antibody (McAb) specif- ically reactive with neurofilament protein. A marked increase in the degree of neuronal cell sur-
vival was apparent in parallel cultures grown in media supplemented with 4 pg/ml MCM protein, with immunofluorescence staining for neurofila- ment protein highlighting a greatly increased den- sity and complexity of axonal outgrowth in these cultures as compared with untreated controls (Fig. Ic,d). From a total of eight photographs from three independent cultures it was estimated that there was an approximate twofold increase in neuronal cell survival in the MCM-treated cultures (15.4 _t
2.8 and 35.3 & 4.4 cells per field of -10 mm2, re- spectively; both values mean & SD, n = 8).
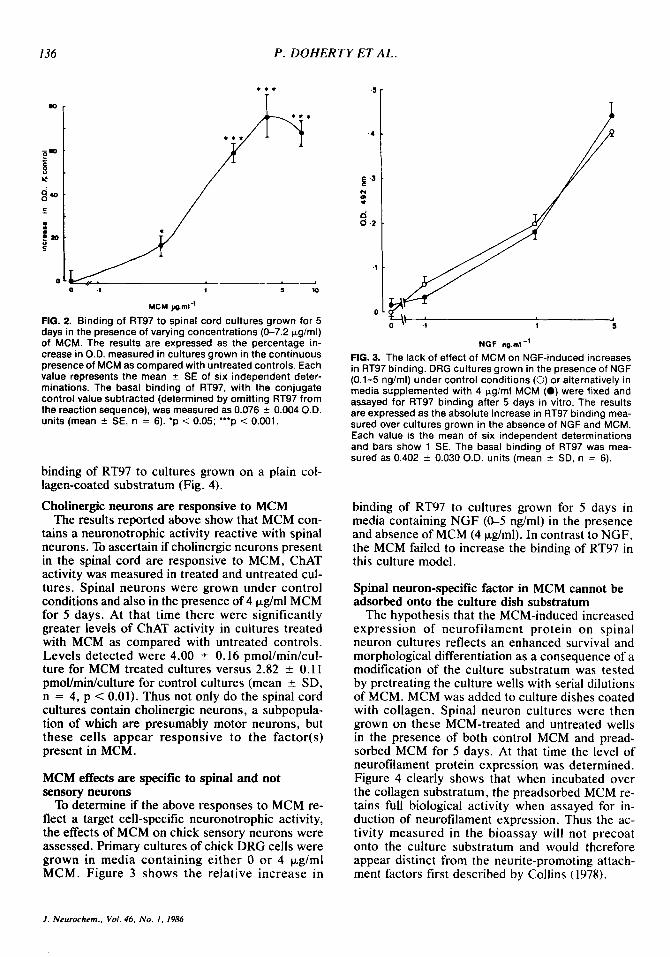
In an attempt to quantify the level of neurono- trophic activity present in MCM, we have used a biochemical index, that is, the level of neurofila- ment protein in 5-day-old cultures grown in the presence o r absence of MCM. Using the ELISA assay for neurofilament protein, a significant (p < 0.05) increase in RT97 binding was found in cultures grown in the continuous presence of 0.4 pg/ml added protein, with half-maximal and maximal re- sponses apparent at 1 and 4 pg/ml, respectively (Fig. 2).
The experiment illustrated above was carried out on a culture dish coated with collagen and polyly- sine. However, there was no obvious difference in the ability of the MCM to promote increases in the
FIG. 1. Indirect irnrnunofluorescence analysis of RT97 binding in control (a,b) and MCM- (4 pglrnl) treated (c,d) cultures of chick spinal cord. Cultures were fixed at day 5 in vitro, permeabilised, and incubated sequentially with RT97 followed by rhodamine-conjugated sheep anti-mouse lg. Cells were viewed with phase-contrast (as) and epifluorescence (b,d) optics. Scale bars represent 50 Frn.
1. Neururhem.. Vol . 46, No. I . 1986
136 P . DOHERTY ET AL
*.* T
0 .l 1 5 u )
MCM )rOrnt4
FIG. 2. Binding of RT97 to spinal cord cultures grown for 5 days in the presence of varying concentrations (C7.2 pg/ml) of MCM. The results are expressed as the percentage in- crease in O.D. measured in cultures grown in the continuous presence of MCM as compared with untreated controls. Each value represents the mean 2 SE of six independent deter- minations. The basal binding of RT97, with the conjugate control value subtracted (determined by omitting RT97 from the reaction sequence), was measured as 0.076 5 0.004 OD. units (mean ? SE, n = 6). ‘p < 0.05; ***p < 0.001.
binding of RT97 to cultures grown on a plain col- lagen-coated substratum (Fig. 4).
Cholinergic neurons are responsive to MCM The results reported above show that MCM con-
tains a neuronotrophic activity reactive with spinal neurons. To ascertain if cholinergic neurons present in the spinal cord are responsive to MCM, ChAT activity was measured in treated and untreated cul- tures. Spinal neurons were grown under control conditions and also in the presence of 4 pg/ml MCM for 5 days. At that time there were significantly greater levels of ChAT activity in cultures treated with MCM as compared with untreated controls. Levels detected were 4.00 * 0.16 pmol/min/cul- ture for MCM treated cultures versus 2.82 2 0.1 I pmol/min/culture for control cultures (mean 2 SD, n = 4, p < 0.01). Thus not only do the spinal cord cultures contain cholinergic neurons, a subpopula- tion of which are presumably motor neurons, but these cells appear responsive to the factor(s) present in MCM.
MCM effects are specific to spinal and not sensory neurons
To determine if the above responses to MCM re- flect a target cell-specific neuronotrophic activity, the effects of MCM on chick sensory neurons were assessed. Primary cultures of chick DRG cells were grown in media containing either 0 or 4 pg/ml MCM. Figure 3 shows the relative increase in
1 5
NCF ng.rnt-’
FIG. 3. The lack of effect of MCM on NGF-induced increases in RT97 binding. DRG cultures grown in the presence of NGF (0.1-5 ng/ml) under control conditions (0) or alternatively in media supplemented with 4 pg!ml MCM (0) were fixed and assayed for RT97 binding after 5 days in vitro. The results are expressed as the absolute increase in RT97 binding mea- sured over cultures grown in the absence of NGF and MCM. Each value is the mean of six independent determinations and bars show 1 SE. The basal binding of RT97 was mea- sured as 0.402 2 0.030 O.D. units (mean 2 SD, n = 6).
binding of RT97 to cultures grown for 5 days in media containing NGF (&5 ng/ml) in the presence and absence of MCM (4 pg/ml). In contrast to NGF, the MCM failed to increase the binding of RT97 in this culture model.
Spinal neuron-specific factor in MCM cannot be adsorbed onto the culture dish substratum
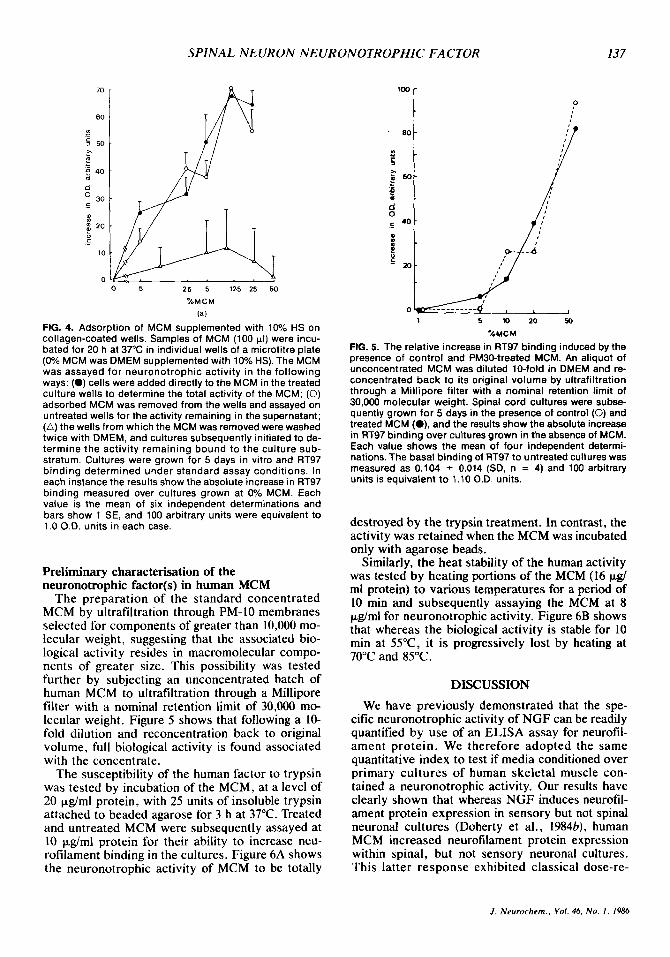
The hypothesis that the MCM-induced increased expression of neurofilament protein on spinal neuron cultures reflects an enhanced survival and morphological differentiation as a consequence of a modification of the culture substratum was tested by pretreating the culture wells with serial dilutions of MCM. MCM was added to culture dishes coated with collagen. Spinal neuron cultures were then grown on these MCM-treated and untreated wells in the presence of both control MCM and pread- sorbed MCM for 5 days. At that time the level of neurofilament protein expression was determined. Figure 4 clearly shows that when incubated over the collagen substratum, the preadsorbed MCM re- tains full biological activity when assayed for in- duction of neurofilament expression. Thus the ac- tivity measured in the bioassay will not precoat onto the culture substratum and would therefore appear distinct from the neurite-promoting attach- ment factors first described by Collins (1978).
1. Neurorhem.. Vol. 46, N o . I , 1986
SPlNAL NEURON NEURONOTROPHIC FACTOR 137
?
25 5 125 25 50 0
0 5
%MCM
(a)
FIG. 4. Adsorption of MCM supplemented with 10% HS on collagen-coated wells. Samples of MCM (100 PI) were incu- bated for 20 h at 37°C in individual wells of a microtitre plate (0% MCM was DMEM supplemented with 10% HS). The MCM was assayed for neuronotrophic activity in the following ways: (0) cells were added directly to the MCM in the treated culture wells to determine the total activity of the MCM; (0) adsorbed MCM was removed from the wells and assayed on untreated wells for the activity remaining in the supernatant; (A) the wells from which the MCM was removed were washed twice with DMEM, and cultures subsequently initiated to de- termine the activity remaining bound to the culture sub- stratum. Cultures were grown for 5 days in vitro and RT97 binding determined under standard assay conditions. In each instance the results show the absolute increase in RT97 binding measured over cultures grown at 0% MCM. Each value is the mean of six independent determinations and bars show 1 SE, and 100 arbitrary units were equivalent to 1 .O O.D. units in each case.
Preliminary characterisation of the neuronotrophic factor(s) in human MCM
The preparation of the standard concentrated MCM by ultrafiltration through PM-I0 membranes selected for components of greater than 10,000 mo- lecular weight, suggesting that the associated bio- logical activity resides in macromolecular compo- nents of greater size. This possibility was tested further by subjecting an unconcentrated batch of human MCM to ultrafiltration through a Millipore filter with a nominal retention limit of 30,000 mo- lecular weight. Figure 5 shows that following a 10- fold dilution and reconcentration back to original volume, full biological activity is found associated with the concentrate.
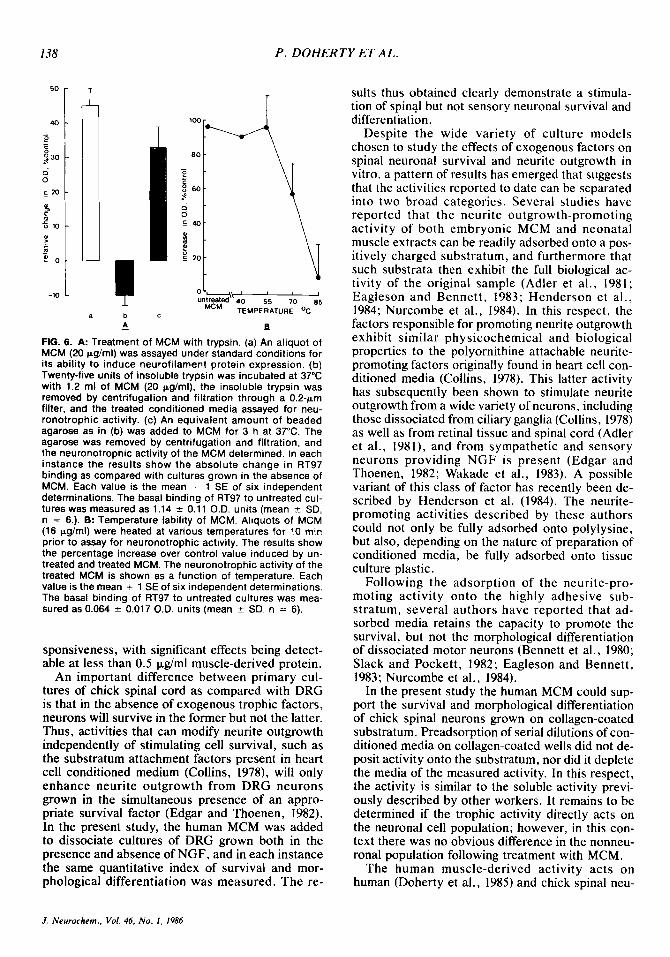
The susceptibility of the human factor to trypsin was tested by incubation of the MCM, at a level of 20 pg/ml protein, with 25 units of insoluble trypsin attached to beaded agarose for 3 h at 37°C. Treated and untreated MCM were subsequently assayed at 10 p g h l protein for their ability to increase neu- rofilament binding in the cultures. Figure 6A shows the neuronotrophic activity of MCM to be totally
FIG. 5. The relative increase in RT97 binding induced by the presence of control and PM3O-treated MCM. An aliquot of unconcentrated MCM was diluted 10-fold in DMEM and re- concentrated back to its original volume by ultrafiltration through a Millipore filter with a nominal retention limit of 30,000 molecular weight. Spinal cord cultures were subse- quently grown for 5 days in the presence of control (0) and treated MCM (O), and the results show the absolute increase in RT97 binding over cultures grown in the absence of MCM. Each value shows the mean of four independent determi- nations. The basal binding of RT97 to untreated cultures was measured as 0.104 2 0.014 (SD, n = 4) and 100 arbitrary units is equivalent t o 1.10 O.D. units.
destroyed by the trypsin treatment. In contrast, the activity was retained when the MCM was incubated only with agarose beads.
Similarly, the heat stability of the human activity was tested by heating portions of the MCM (16 pg/ ml protein) to various temperatures for a period of 10 min and subsequently assaying the MCM at 8 pg/ml for neuronotrophic activity. Figure 6B shows that whereas the biological activity is stable for 10 min at SYC, it is progressively lost by heating at 70°C and 85°C.
DISCUSSION
We have previously demonstrated that the spe- cific neuronotrophic activity of NGF can be readily quantified by use of an ELISA assay for neurofil- ament protein. We therefore adopted the same quantitative index to test if media conditioned over primary cultures of human skeletal muscle con- tained a neuronotrophic activity. Our results have clearly shown that whereas NGF induces neurofil- ament protein expression in sensory but not spinal neuronal cultures (Doherty et al., 1984b), human MCM increased neurofilament protein expression within spinal, but not sensory neuronal cultures. This latter response exhibited classical dose-re-
1. Neurorhern.. Vol. 46. No. I . 1986
P . DOHERTY ET AL.
sults thus obtained clearly demonstrate a stimula- tion of spinal but not sensory neuronal survival and differentiation.
Despite the wide variety of culture models chosen to study the effects of exogenous factors on spinal neuronal survival and neurite outgrowth in vitro, a pattern of results has emerged that suggests that the activities reported to date can be separated into two broad categories. Several studies have reported that the neurite outgrowth-promoting activity of both embryonic MCM and neonatal muscle extracts can be readily adsorbed onto a pos- itively charged substratum, and furthermore that such substrata then exhibit the full biological ac- t iv i ty of the original sample (Adler et al., 1981; Eagleson and Bennett, 1983; Henderson et al., 1984; Nurcombe et al., 1984). In this respect, the factors responsible for promoting neurite outgrowth 6
FIG. 6. A: Treatment of MCM with trypsin. (a) An aliquot of MCM (20 pg/ml) was assayed under standard conditions for its ability to induce neurofilament protein expression. (b) Twenty-five units of insoluble trypsin was incubated at 37°C with 1.2 ml of MCM (20 pg/ml), the insoluble trypsin was removed by centrifugation and filtration through a 0.2-km filter, and the treated conditioned media assayed for neu- ronotrophic activity. (c) An equivalent amount of beaded agarose as in (b) was added to MCM for 3 h at 37°C. The agarose was removed by centrifugation and filtration, and the neuronotrophic activity of the MCM determined. In each instance the results show the absolute change in RT97 binding as compared with cultures grown in the absence of MCM. Each value is the mean + 1 SE of six independent determinations. The basal binding of RT97 to untreated cul- tures was measured as 1.14 2 0.11 O.D. units (mean 5 SD, n = 6.). 8: Temperature lability of MCM. Aliquots of MCM (16 pglml) were heated at various temperatures for 10 min prior to assay for neuronotrophic activity. The results show the percentage increase over control value induced by un- treated and treated MCM. The neuronotrophic activity of the treated MCM is shown as a function of temperature. Each value is the mean + 1 SE of six independent determinations. The basal binding of RT97 to untreated cultures was mea- sured as 0.064 f 0.017 O.D. units (mean 2 SD, n = 6).
sponsiveness, with significant effects being detect- able at less than 0.5 pg/ml muscle-derived protein.
An important difference between primary cul- tures of chick spinal cord as compared with DRG is that in the absence of exogenous trophic factors, neurons will survive in the former but not the latter. Thus, activities that can modify neurite outgrowth independently of stimulating cell survival, such as the substratum attachment factors present in heart cell conditioned medium (Collins, 1978), will only enhance neurite outgrowth from DRG neurons grown in the simultaneous presence of an appro- priate survival factor (Edgar and Thoenen, 1982). In the present study, the human MCM was added to dissociate cultures of DRG grown both in the presence and absence of NGF, and in each instance the same quantitative index of survival and mor- phological differentiation was measured. The re-
exhibit similar physicochemical and bioiogical properties to the polyornithine attachable neurite- promoting factors originally found in heart cell con- ditioned media (Collins, 1978). This latter activity has subsequently been shown to stimulate neurite outgrowth from a wide variety of neurons, including those dissociated from ciliary ganglia (Collins, 1978) as well as from retinal tissue and spinal cord (Adler et al., 1981), and from sympathetic and sensory neurons providing N G F is present (Edgar and Thoenen, 1982; Wakade et al., 1983). A possible variant of this class of factor has recently been de- scribed by Henderson et al. (1984). The neurite- promoting activities described by these authors could not only be fully adsorbed onto polylysine, but also, depending on the nature of preparation of conditioned media, be fully adsorbed onto tissue culture plastic.
Following the adsorption of the neurite-pro- moting activity onto the highly adhesive sub- stratum, several authors have reported that ad- sorbed media retains the capacity to promote the survival, but not the morphological differentiation of dissociated motor neurons (Bennett et al., 1980; Slack and Pockett, 1982; Eagleson and Bennett, 1983; Nurcombe et al., 1984).
In the present study the human MCM could sup- port the survival and morphological differentiation of chick spinal neurons grown on collagen-coated substratum. Preadsorption of serial dilutions of con- ditioned media on collagen-coated wells did not de- posit activity onto the substratum, nor did it deplete the media of the measured activity. In this respect, the activity is similar to the soluble activity previ- ously described by other workers. It remains to be determined if the trophic activity directly acts on the neuronal cell population; however, in this con- text there was no obvious difference in the nonneu- ronal population following treatment with MCM.
The human muscle-derived activity acts on human (Doherty et al., 1985) and chick spinal neu-
.I. Neurochem., Vol. 46, No. I , 1986

SPINAL NEURON NEURONOTROPHIC FACTOR 139
rons, but not chick sensory neurons. The ELISA assay for neurofilament proteins provides a rapid and highly sensitive means to quantitate this ac- tivity arbitrarily. As such, the assay can serve as a convenient tool in the purification and further char- acterisation of the human factor(s), with a view to the identification of the cell types that can synthe- sise and respond to the activity.
REFERENCES Adler R., Manthorpe M., Skaper S. D., and Varon S. (1981)
Poly-ornithine attached neurite-promoting factors (PNPFs). Culture sources and responsive neurons. Brain Res. 206, 129-1 44.
Anderton B. H.. Breinburg D., Downes M. J., Green P. J., Tom- Iinson B. E.. Ulrich J. , Wood J. N., and Kahn J. (1982) Monoclonal antibodies show that neurofibrillary tangles and neurofilaments share antigenic determinants. Narure 298, 84-86.
Barde Y.-A,, Edgar D., and Thoenen H. (1983) New neuro- trophic factors. Annu. Rev. Physiol. 45, 601412.
Bennett M. R., Lai K., and Nurcombe V. (1980) Identification of embryonic motor neurons in virro; their survival is de- pendent on skeletal muscle. Bruin Res. 190, 537-542.
Brown M. C., Holland R. I., and Hopkins W. G. (1981) Motor nerve sprouting. Annu. Rev. Neurosci. 4, 17-42.
Collins F. (1978) Induction of neurite outgrowth by a conditioned medium factor bound to the culture substratum. Proc. Nut/. Acad. Sci. USA 75, 5210-5213.
Dickson J . G., Flanigan T. P., and Walsh F. S. (1982) Cell surface antigens of human foetal brain and dorsal root ganglion cells in tissue culture, in Human Motor Neuron Diseases (Row- land L. P., ed), pp. 435-451. Raven Press, New York.
Doherty P., Dickson J . G., Flanigan T. P., and Walsh F. S. (19844 Quantitative evaluation of neurite outgrowth in cul- tures of human foetal brain and dorsal root ganglion cells using an enzyme-linked immunoadsorbent assay for human neurofilament protein. J. Neurochem. 42, 11 16-1 122.
Doherty P., Dickson J. G., Flanigan T. P., and Walsh F. S. (19846) The effect of nerve growth factor and its antibodies on neurofilament protein expression in primary cultures of sensory and spinal neurons. Neurosci. Lcrr. 52, 56-60.
Doherty P., Dickson J. G., Flanigan T. P., and Walsh F. S. (1985) Human muscle cell conditioned media stimulates neurofil- ament protein expression in primary cultures of human and chick spinal cord. J . Physiol. 360, 43P.
Eagleson K. L. and Bennett M. R. (1983) Survival of purified motor neurons in virro: effects of skeletal muscle-condi- tioned media. Neurosci. Lerr. 38, 187-192.
Edgar D. and Thoenen H. (1982) Modulation of NGF-induced survival of chick sympathetic neurons by contact with a conditioned medium factor bound to the culture substrate. Drv. Brain. Res. 5 , 89-92.
Fonnum F. (1975) A rapid radiochemical method for the deter- mination of choline acetyltransferase. J . Neurochem. 24, 407-409.
Hamburger V. (1958) Regression versus peripheral control of dif- ferentiation in motor hypoplasia. Am. J. Anat. 102,364-410.
Hamburger V. and Oppenheim R. W. (1982) Naturally occurring neuronal cell death in vertebrates. Neurosci. Comment 1 , 39-55.
Henderson C. E., Huchet M., and Changeux J. P. (1981) Neurite outgrowth from embryonic chicken spinal neurons is pro- moted by media conditioned by muscle cells. Proc. Narl. Acad. Sci. USA 78, 2625-2629.
Henderson C. E., Huchet M., and Changeux J. P. (1984) Neu- rite-promoting activities for embryonic spinal neurons and their developmental changes in the chick. Dev. B id . 104, 336-347.
Hollyday M. and Hamburger V. (1976) Reduction of the naturally occurring motor neuron loss by enlargement of the pe- riphery. J. Comp. Neurol. 170, 311-320.
Jacobson M. (1978) Developmental Neurobiology. Plenum Press, New York.
Nakane P. K. and Kawaoi A. (1974) Peroxidase-labelled anti- body, a new method of conjugation. J . Historhem. Cyto- chem. 22, 1084.
Nurcombe V., Hill M. A., Eagleson K. L., and Bennett M. R. (1984) Motor neuron survival and neuritic extension from spinal cord explants induced by factors released from de- nervated muscle. Brain Res. 291, 19-28.
Purves D. (1980) Neuronal competition. Nature 287, 585-586. Rogers S. L., Letourneau P. C., Palm S. L.. McCarthy J., and
Furcht L. (1983) Neurite extension by peripheral and central nervous system neurons in response to substratum-bound fibronectin and laminin. Dev. B i d . 98, 212-220.
Slack J. R. and Pockett S. (1982) Motor neurotrophic factor in denervated adult skeletal muscle. Brain Res. 247, 131140.
Thoenen H. and Barde Y.-A. (1980) Physiology of nerve growth factor. Physiol. Rev. 60, 1284-1335.
Wakade A. R., Edgar D., and Thoenan H. (1983) Both nerve growth factor and high K concentrations support the sur- vival of chick embryo sympathetic neurons. Exp. Cell Res. 144, 377-384.
Yanker B. A. and Shooter E. M. (1982) The biology and mech- anism of action of nerve growth factor. Annu. Rev. Biochem. 51, 845-868.
Yasin R., Van Beers G., Nurse K., Al-Ani S., Landon D. N., and Thompson E. J. (1977) A quantitative technique for growing human adult skeletal muscle in culture starting from mononucleated cells. J. Neuro/. Sci. 32, 347-360.
J . Neurochern., Vol. 46. No. I . 1986





























