Embed Size (px)
Citation preview

Human pluripotent stem cells as a model oftrophoblast differentiation in both normaldevelopment and diseaseMariko Horiia,b,1, Yingchun Lia,b,1, Anna K. Wakelanda,b,1, Donald P. Pizzoa, Katharine K. Nelsona,b, Karen Sabatinib,c,Louise Chang Laurentb,c, Ying Liud,e,f, and Mana M. Parasta,b,2
aDepartment of Pathology, University of California, San Diego, La Jolla, CA 92093; bSanford Consortium for Regenerative Medicine, University of California,San Diego, La Jolla, CA 92093; cDepartment of Reproductive Medicine, University of California, San Diego, La Jolla, CA 92093; dDepartment ofNeurosurgery, Center for Stem Cell and Regenerative Medicine, University of Texas Health Sciences Center, Houston, TX 77030; eThe Senator Lloyd andB. A. Bentsen Center for Stroke Research, University of Texas Health Sciences Center, Houston, TX 77030; and fThe Brown Foundation Institute of MolecularMedicine for the Prevention of Human Diseases, University of Texas Health Sciences Center, Houston, TX 77030
Edited by R. Michael Roberts, University of Missouri–Columbia, Columbia, MO, and approved May 25, 2016 (received for review March 24, 2016)
Trophoblast is the primary epithelial cell type in the placenta, atransient organ required for proper fetal growth and develop-ment. Different trophoblast subtypes are responsible for gas/nutrientexchange (syncytiotrophoblasts, STBs) and invasion and maternalvascular remodeling (extravillous trophoblasts, EVTs). Studies ofearly human placental development are severely hampered by thelack of a representative trophoblast stem cell (TSC) model with thecapacity for self-renewal and the ability to differentiate into bothSTBs and EVTs. Primary cytotrophoblasts (CTBs) isolated fromearly-gestation (6–8 wk) human placentas are bipotential, a phe-notype that is lost with increasing gestational age. We have iden-tified a CDX2+/p63+ CTB subpopulation in the early postimplantationhuman placenta that is significantly reduced later in gestation. Wedescribe a reproducible protocol, using defined medium containingbone morphogenetic protein 4 by which human pluripotent stemcells (hPSCs) can be differentiated into CDX2+/p63+ CTB stem-likecells. These cells can be replated and further differentiated intoSTB- and EVT-like cells, based on marker expression, hormone secre-tion, and invasive ability. As in primary CTBs, differentiation ofhPSC-derived CTBs in low oxygen leads to reduced human chorionicgonadotropin secretion and STB-associated gene expression, in-stead promoting differentiation into HLA-G+ EVTs in an hypoxia-inducible, factor-dependent manner. To validate further the utilityof hPSC-derived CTBs, we demonstrated that differentiation of tri-somy 21 (T21) hPSCs recapitulates the delayed CTB maturation andblunted STB differentiation seen in T21 placentae. Collectively, ourdata suggest that hPSCs are a valuable model of human placentaldevelopment, enabling us to recapitulate processes that result inboth normal and diseased pregnancies.
human pluripotent stem cells | cytotrophoblast | extravillous trophoblast |syncytiotrophoblast | placenta
Trophoblast is the first lineage specified in the developingembryo. Following blastocoel formation, cells segregate into
the inner cell mass (ICM), which gives rise to the epiblast andembryo proper; the outer trophectoderm (TE) cells give rise toextraembryonic ectoderm, the epithelial portion of the placenta(1). Much of what we know about this first lineage specificationcomes from studies in mouse (2–4). In fact, the combination ofabilities to derive trophoblast stem cells (TSCs) from mouseblastocysts and to evaluate the compartment-specific contributionof specific genes during embryogenesis has resulted in the iden-tification of numerous transcription factors and signaling pathwaysinvolved in TE specification and placental development (2, 3, 5).Results from such studies have led to identification of Cdx2(caudal type homeobox 2), a member of the caudal-related ho-meobox transcription factor gene family, the expression of whichhas been shown to be required for maintenance of TE and TSCsin the mouse embryo (6, 7). Other transcription factors, including
Elf5 (Ets domain transcription factor) and Eomes (Eomeso-dermin), also have been shown to be required for maintenance ofthe TSC fate in the mouse (8, 9).Significantly less is known about TE specification and the TSC
niche in the human embryo (10, 11). Recent studies have pointed tomajor differences in gene expression between mouse and humanpreimplantation embryos (12, 13). One major distinction is thecoexpression of CDX2 and POU5F1 (POU domain, class 5, ho-meobox 1)/OCT4 (octamer-binding protein 4) in the early TE ofhuman embryos, whereas in the mouse embryo these two genes areexpressed exclusively in the TE and ICM, respectively, each recip-rocally regulating the expression of the other (6, 12, 13). Otherdifferences include the lack of EOMES and ELF5 expression inhuman TE (13). Based on these findings, the absence of a prolif-erating trophoblast compartment in the early human embryo (1, 10),and the inability to derive human TSCs from such preimplantationembryos (14), it has been proposed that the human TSC niche mayin fact reside in the early postimplantation placenta.In the absence of a human TSC model, researchers have turned
to human pluripotent stem cells (hPSCs). Since 2002, when Xuet al. (15) first published the finding that bone morphogeneticprotein 4 (BMP4) induces the expression of trophoblast-relatedgenes in hPSCs, multiple groups have used these cells as a modelfor studying trophoblast lineage specification (16–22). The ma-jority of these studies, including our own (21), have used BMP4
Significance
Human pluripotent stem cells (hPSCs) continue to be underap-preciated as a model for studying trophoblast differentiation. Inthis study, we provide a reproducible, two-step protocol by whichhPSCs can be differentiated into bipotential cytotrophoblast (CTB)stem-like cells and subsequently into functional, terminally dif-ferentiated trophoblasts. In addition, we provide evidence thatthe response of hPSC-derived CTBs to low oxygen is similar to thatof primary CTBs. Finally, using trisomy 21-affected hPSCs, weshow, for the first time to our knowledge, that hPSCs can model atrophoblast differentiation defect. We propose that hPSCs aresuperior to other currently available models for studying humantrophoblast differentiation.
Author contributions: M.M.P. designed research; M.H., Y. Li, A.K.W., D.P.P., K.K.N., andK.S. performed research; M.H., Y. Li, A.K.W., L.C.L., and Y. Liu contributed new reagents/analytic tools; M.H., Y. Li, A.K.W., K.S., L.C.L., and M.M.P. analyzed data; and M.H., Y. Li,A.K.W., and M.M.P. wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.1M.H., Y. Li, and A.K.W. contributed equally to this work.2To whom correspondence should be addressed. Email: [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1604747113/-/DCSupplemental.
E3882–E3891 | PNAS | Published online June 20, 2016 www.pnas.org/cgi/doi/10.1073/pnas.1604747113
Dow
nloa
ded
by g
uest
on
May
27,
202
0
in the presence of feeder-conditioned medium (FCM), resultingin the expression of some mesoderm markers and thereforegenerating doubt about the true identity of these hPSC-derivedcells (23). Nevertheless, follow-up studies using more definedculture conditions have confirmed the identity of these cells astrophoblasts (20).Most recently, Lee et al. (24) have proposed criteria for defining
trophoblasts based on expression of a set of markers, includingELF5. Although a laudable attempt at standardization, this studyfails to account for differences in gene expression across gestationalage and falls short of defining syncytiotrophoblasts (STBs) (24).To confirm the utility of hPSCs for modeling trophoblast dif-ferentiation, we instead asked whether these cells can recapitu-late functional phenotypes of primary trophoblasts during bothnormal development and disease. We previously have identifiedp63, a member of the p53 family of nuclear proteins, as a markerspecific to proliferative cytotrophoblasts (CTBs) in the humanplacenta (21, 25, 26). We now have identified a subpopulation ofCTBs in the early human placenta that are double-positive forp63 and CDX2; this CTB subpopulation is greatly reduced in thesecond trimester and is temporally associated with the loss ofbipotential differentiation of CTBs (27). In addition, we describea completely defined culture condition, containing BMP4, bywhich CDX2+/p63+ CTB stem-like cells can be efficiently andreproducibly derived from hPSCs. Furthermore, we show thathPSC-derived CTBs respond to low oxygen in a manner similarto primary CTBs. Finally, we provide the first, to our knowledge,
proof-of-concept data for the ability of hPSCs to model a tro-phoblast differentiation defect, using trisomy 21 (T21)-affectedhPSCs.
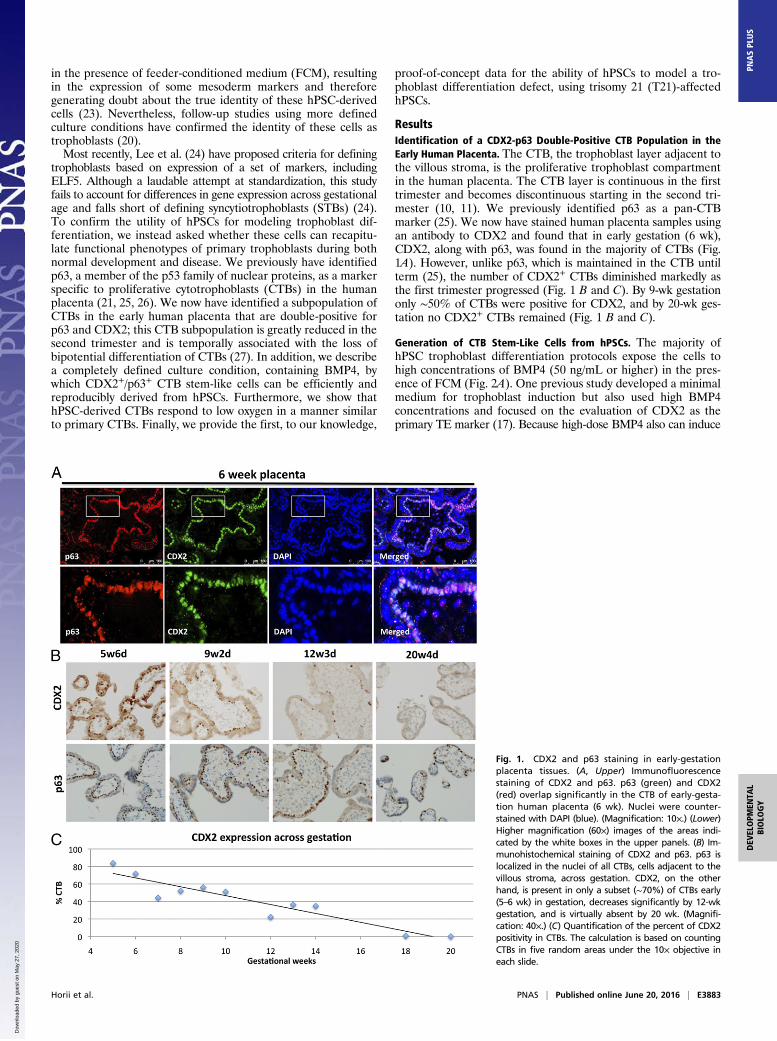
ResultsIdentification of a CDX2-p63 Double-Positive CTB Population in theEarly Human Placenta. The CTB, the trophoblast layer adjacent tothe villous stroma, is the proliferative trophoblast compartmentin the human placenta. The CTB layer is continuous in the firsttrimester and becomes discontinuous starting in the second tri-mester (10, 11). We previously identified p63 as a pan-CTBmarker (25). We now have stained human placenta samples usingan antibody to CDX2 and found that in early gestation (6 wk),CDX2, along with p63, was found in the majority of CTBs (Fig.1A). However, unlike p63, which is maintained in the CTB untilterm (25), the number of CDX2+ CTBs diminished markedly asthe first trimester progressed (Fig. 1 B and C). By 9-wk gestationonly ∼50% of CTBs were positive for CDX2, and by 20-wk ges-tation no CDX2+ CTBs remained (Fig. 1 B and C).
Generation of CTB Stem-Like Cells from hPSCs. The majority ofhPSC trophoblast differentiation protocols expose the cells tohigh concentrations of BMP4 (50 ng/mL or higher) in the pres-ence of FCM (Fig. 2A). One previous study developed a minimalmedium for trophoblast induction but also used high BMP4concentrations and focused on the evaluation of CDX2 as theprimary TE marker (17). Because high-dose BMP4 also can induce
Fig. 1. CDX2 and p63 staining in early-gestationplacenta tissues. (A, Upper) Immunofluorescencestaining of CDX2 and p63. p63 (green) and CDX2(red) overlap significantly in the CTB of early-gesta-tion human placenta (6 wk). Nuclei were counter-stained with DAPI (blue). (Magnification: 10×.) (Lower)Higher magnification (60×) images of the areas indi-cated by the white boxes in the upper panels. (B) Im-munohistochemical staining of CDX2 and p63. p63 islocalized in the nuclei of all CTBs, cells adjacent to thevillous stroma, across gestation. CDX2, on the otherhand, is present in only a subset (∼70%) of CTBs early(5–6 wk) in gestation, decreases significantly by 12-wkgestation, and is virtually absent by 20 wk. (Magnifi-cation: 40×.) (C) Quantification of the percent of CDX2positivity in CTBs. The calculation is based on countingCTBs in five random areas under the 10× objective ineach slide.
Horii et al. PNAS | Published online June 20, 2016 | E3883
DEV
ELOPM
ENTA
LBIOLO
GY
PNASPL
US
Dow
nloa
ded
by g
uest
on
May
27,
202
0

mesoderm (23), we asked whether, in that minimal medium (17),low-dose BMP4 (10 ng/mL) by itself is able to initiate trophoblastdifferentiation of hPSCs into CTBs, as evaluated by a com-prehensive panel of markers, including p63 and CDX2. We ini-tially used WA09/H9 ES cells (ESCs) but also tested this protocolon WA01/H1 and SIVF21 ESCs as well as on induced pluripotentstem cells (iPSCs) derived in the L.C.L. and Y. Liu laboratories,all of which were adapted to StemPro according to the manu-facturer’s instructions, with similar results in each case (Fig. S1).We refer to this method as the “two-step protocol” (Fig. 2B).In the first step, StemPro-adapted hPSCs were treated with a
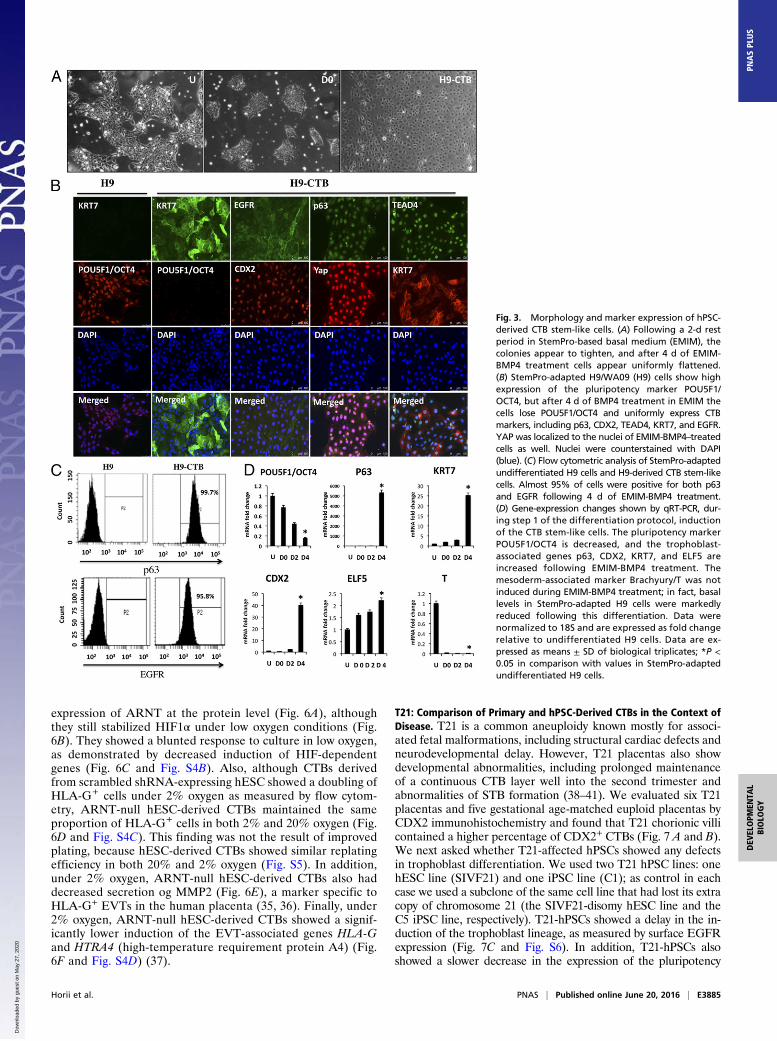
DMEM/F12-based minimal medium (EMIM) (17) for 2 d andthen were cultured in EMIM containing 10 ng/mL BMP4 for upto 4 d (Fig. 2B). After 2 d in EMIM alone, the colonies appearedto tighten and become better delineated; following BMP4treatment, cells uniformly adopted a more flattened appearancebut continued to grow to near confluence by day 4 (Fig. 3A). Wethen proceeded to evaluate markers of undifferentiated CTBsby immunostaining: Compared with undifferentiated hPSCs,BMP4-treated cells showed loss of the pluripotency markerPOU5F1/OCT4; conversely, the CTB-associated genes p63,CDX2, TEAD4 (TEA domain family member 4), KRT7 (keratin,type II cytoskeletal 7), and EGFR (EGF receptor) were all highlyexpressed in the hPSC-derived CTBs (Fig. 3B). In addition, YAP(Yes-associated protein) was localized to the nuclei of thesehPSC-derived CTBs (Fig. 3B). After 4 d of BMP4 treatment,more than 95% of the cells were positive for both p63 and EGFRas measured by flow cytometry (Fig. 3C). Next, we investigatedgene expression during CTB induction of hPSCs. Analysis byquantitative RT-PCR (qRT-PCR) showed that the expression ofPOU5F1/OCT4 decreased and the expression of p63, CDX2,and KRT7 increased significantly following BMP4 treatment(Fig. 3D). In addition, the mouse TSC-associated marker ELF5was induced twofold, whereas the mesoderm-associated markerT/BRACHYURY was reduced from the basal levels present inStemPro-adapted cells and was not induced by low-dose BMP4treatment (Fig. 3D). These data indicate that, with only 4 d of
low-dose BMP4 treatment in this defined medium, hPSCs uni-formly assume a CTB stem-like phenotype. Interestingly, as inisolated primary CTBs but unlike p63+ CTBs in vivo (26), hPSC-derived CTBs were not highly proliferative and could not bepropagated under these culture conditions (Fig. S2).
Differentiation Capacity of hPSC-Derived CTBs. To determine themultipotency of hPSC-derived CTBs, we tested their ability todifferentiate terminally into both STBs and extravillous tropho-blasts (EVTs). Because the cells reach confluency at day 4 aftertreatment with EMIM-BMP4, we attempted to replate the cellsat a lower density (∼2–4 × 104 cells/cm2). To replate hPSC-derived CTBs, we tested different cell-detachment methods,including trypsin, StemDS, and EDTA; we obtained the mostconsistent results with trypsin. We also tested replating on Geltrexboth in the minimal medium EMIM and in FCM and foundthat cells plated significantly better in the presence of FCM(Fig. S3A).After replating and culture in FCM supplemented with 10 ng/mL
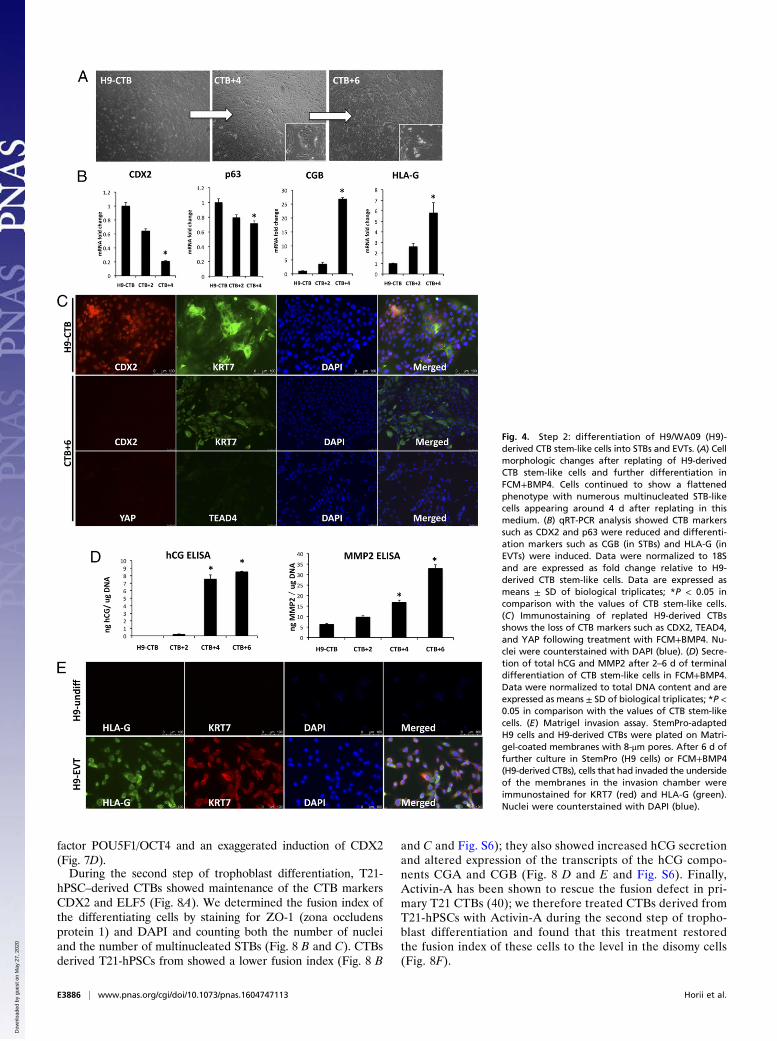
human BMP4 (FCM+BMP4), the cells continued to differentiate(Fig. 4). Morphologically, 4–6 d after FCM+BMP4 treatment, cellscontinued to show an epithelial (flattened) phenotype with nu-merous multinucleated cells (Fig. 4A). Based on qRT-PCR analysis,CTB markers such as CDX2 and p63 were reduced, and differen-tiation markers, including CGB (chorionic gonadotropin subunitbeta; an STB marker) and HLA-G (an EVT marker), were in-duced (Fig. 4B). Loss of CTB markers was further evident byimmunostaining, showing loss of expression of CDX2, TEAD4, andYAP following culture in FCM-BMP4 (Fig. 4C). Functional tro-phoblast differentiation was confirmed by the secretion of hCG(human CG, the pregnancy hormone and a marker of STBs) andMMP2 (matrix metalloproteinase-2, an EVT marker) (Fig. 4D). Inaddition, we confirmed the presence of invasive HLA-G+ EVTsusing a Matrigel invasion assay followed by immunostaining (Fig.4E). The presence of FCM appeared to be optimal for differen-tiation, because differentiation in minimal medium with BMP4resulted in significantly lower levels of differentiation markers, in-cluding hCG secretion and surface HLA-G+ cells (Fig. S3 B–D).Together, based on both marker expression and function, these dataconfirm the bipotential differentiation ability of hPSC-derived CTBsin FCM+BMP4.
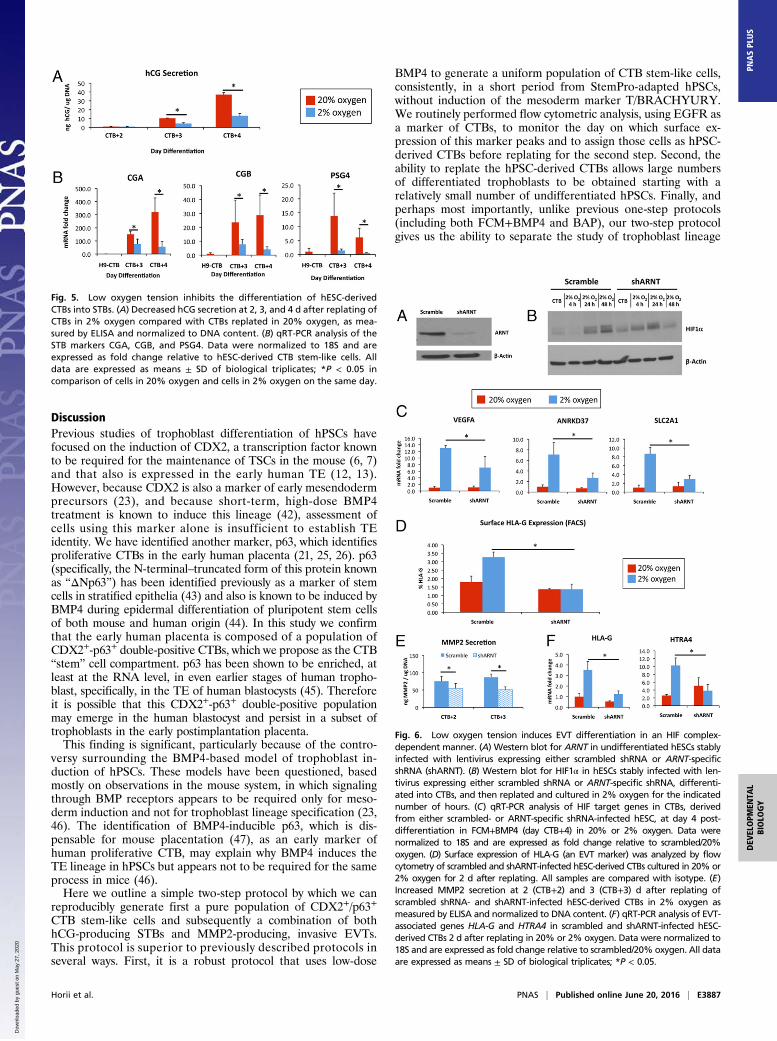
Behavior of hPSC-Derived CTBs in Low Oxygen. The effect of lowoxygen on the differentiation of STBs has been well established.Specifically, the production of hCG- and STB-associated tran-scripts is known to be inhibited by low oxygen tension (28, 29).We replated hPSC-derived CTBs in either 20% or 2% oxygenand continued to differentiate them in FCM+BMP4 (step 2).Under 2% oxygen, the cells produced significantly less hCG (Fig.5A and Fig. S4A); they also showed a significant decrease in theexpression of STB-associated transcripts, including CGA (chorionicgonadotropin subunit alpha), CGB, and PSG4 (pregnancy-specificβ-1-glycoprotein 4) (Fig. 5B).Culture under low oxygen tension is also known to promote
the initial differentiation of CTBs into HLA-G+ EVTs (30), al-though further differentiation into fully mature, invasive EVTs isinhibited (31); in addition, an intact hypoxia-inducible factor(HIF), a transcription factor complex mediating a significantportion of the cellular response to low oxygen (32), has beenshown to be required for the differentiation of rodent TSCs intotrophoblast giant cells, which are analogous to human EVTs(33, 34). To test the effect of different oxygen tensions on EVTdifferentiation, we generated hESCs stably expressing either ascrambled shRNA or shRNA specific to ARNT (aryl hydrocar-bon receptor nuclear translocator; also called “HIF1β”); which isrequired for the formation of an intact HIF complex (32). Weused two different hESC lines, H9/WA09 and SIVF-disomy hESCs(see below). ARNT-null hESCs showed significantly reduced
Fig. 2. Representations of trophoblast differentiation protocols. (A) Tradi-tional (one-step) trophoblast differentiation protocol using BMP4 in thepresence of FCM. This protocol was used by the majority of previous studiesfor trophoblast differentiation of hPSCs. (B) Our modified (two-step) tro-phoblast differentiation protocol. Following a 2-d “rest” in StemPro-basedminimal medium (EMIM), hPSCs are cultured in EMIM plus 10 ng/mL BMP4for up to 4 d to form uniformly flattened CTB stem-like colonies. These cellsthen are trypsinized and replated to be terminally differentiated into STBsand EVTs in FCM plus 10 ng/mL BMP4. H9, H9/WA09 cells; U, undifferenti-ated hPSCs in StemPro; D0, hPSCs after 2-d rest in EMIM; D1–D4, daysafter treatment with EMIM+BMP4; CTB+1 to CTB+6, days after replatingin FCM+BMP4.
E3884 | www.pnas.org/cgi/doi/10.1073/pnas.1604747113 Horii et al.
Dow
nloa
ded
by g
uest
on
May
27,
202
0
expression of ARNT at the protein level (Fig. 6A), althoughthey still stabilized HIF1α under low oxygen conditions (Fig.6B). They showed a blunted response to culture in low oxygen,as demonstrated by decreased induction of HIF-dependentgenes (Fig. 6C and Fig. S4B). Also, although CTBs derivedfrom scrambled shRNA-expressing hESC showed a doubling ofHLA-G+ cells under 2% oxygen as measured by flow cytom-etry, ARNT-null hESC-derived CTBs maintained the sameproportion of HLA-G+ cells in both 2% and 20% oxygen (Fig.6D and Fig. S4C). This finding was not the result of improvedplating, because hESC-derived CTBs showed similar replatingefficiency in both 20% and 2% oxygen (Fig. S5). In addition,under 2% oxygen, ARNT-null hESC-derived CTBs also haddecreased secretion og MMP2 (Fig. 6E), a marker specific toHLA-G+ EVTs in the human placenta (35, 36). Finally, under2% oxygen, ARNT-null hESC-derived CTBs showed a signif-icantly lower induction of the EVT-associated genes HLA-Gand HTRA4 (high-temperature requirement protein A4) (Fig.6F and Fig. S4D) (37).
T21: Comparison of Primary and hPSC-Derived CTBs in the Context ofDisease. T21 is a common aneuploidy known mostly for associ-ated fetal malformations, including structural cardiac defects andneurodevelopmental delay. However, T21 placentas also showdevelopmental abnormalities, including prolonged maintenanceof a continuous CTB layer well into the second trimester andabnormalities of STB formation (38–41). We evaluated six T21placentas and five gestational age-matched euploid placentas byCDX2 immunohistochemistry and found that T21 chorionic villicontained a higher percentage of CDX2+ CTBs (Fig. 7 A and B).We next asked whether T21-affected hPSCs showed any defectsin trophoblast differentiation. We used two T21 hPSC lines: onehESC line (SIVF21) and one iPSC line (C1); as control in eachcase we used a subclone of the same cell line that had lost its extracopy of chromosome 21 (the SIVF21-disomy hESC line and theC5 iPSC line, respectively). T21-hPSCs showed a delay in the in-duction of the trophoblast lineage, as measured by surface EGFRexpression (Fig. 7C and Fig. S6). In addition, T21-hPSCs alsoshowed a slower decrease in the expression of the pluripotency
Fig. 3. Morphology and marker expression of hPSC-derived CTB stem-like cells. (A) Following a 2-d restperiod in StemPro-based basal medium (EMIM), thecolonies appear to tighten, and after 4 d of EMIM-BMP4 treatment cells appear uniformly flattened.(B) StemPro-adapted H9/WA09 (H9) cells show highexpression of the pluripotency marker POU5F1/OCT4, but after 4 d of BMP4 treatment in EMIM thecells lose POU5F1/OCT4 and uniformly express CTBmarkers, including p63, CDX2, TEAD4, KRT7, and EGFR.YAP was localized to the nuclei of EMIM-BMP4–treatedcells as well. Nuclei were counterstained with DAPI(blue). (C) Flow cytometric analysis of StemPro-adaptedundifferentiated H9 cells and H9-derived CTB stem-likecells. Almost 95% of cells were positive for both p63and EGFR following 4 d of EMIM-BMP4 treatment.(D) Gene-expression changes shown by qRT-PCR, dur-ing step 1 of the differentiation protocol, inductionof the CTB stem-like cells. The pluripotency markerPOU5F1/OCT4 is decreased, and the trophoblast-associated genes p63, CDX2, KRT7, and ELF5 areincreased following EMIM-BMP4 treatment. Themesoderm-associated marker Brachyury/T was notinduced during EMIM-BMP4 treatment; in fact, basallevels in StemPro-adapted H9 cells were markedlyreduced following this differentiation. Data werenormalized to 18S and are expressed as fold changerelative to undifferentiated H9 cells. Data are ex-pressed as means ± SD of biological triplicates; *P <0.05 in comparison with values in StemPro-adaptedundifferentiated H9 cells.
Horii et al. PNAS | Published online June 20, 2016 | E3885
DEV
ELOPM
ENTA
LBIOLO
GY
PNASPL
US
Dow
nloa
ded
by g
uest
on
May
27,
202
0
factor POU5F1/OCT4 and an exaggerated induction of CDX2(Fig. 7D).During the second step of trophoblast differentiation, T21-
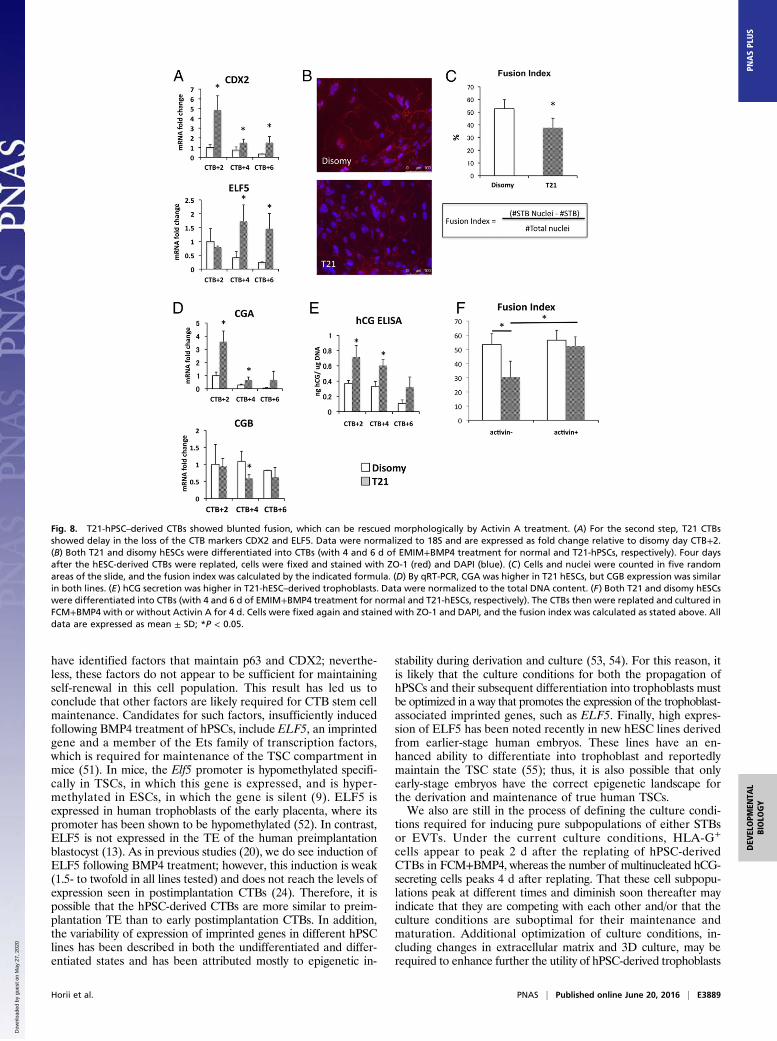
hPSC–derived CTBs showed maintenance of the CTB markersCDX2 and ELF5 (Fig. 8A). We determined the fusion index ofthe differentiating cells by staining for ZO-1 (zona occludensprotein 1) and DAPI and counting both the number of nucleiand the number of multinucleated STBs (Fig. 8 B and C). CTBsderived T21-hPSCs from showed a lower fusion index (Fig. 8 B
and C and Fig. S6); they also showed increased hCG secretionand altered expression of the transcripts of the hCG compo-nents CGA and CGB (Fig. 8 D and E and Fig. S6). Finally,Activin-A has been shown to rescue the fusion defect in pri-mary T21 CTBs (40); we therefore treated CTBs derived fromT21-hPSCs with Activin-A during the second step of tropho-blast differentiation and found that this treatment restoredthe fusion index of these cells to the level in the disomy cells(Fig. 8F).
Fig. 4. Step 2: differentiation of H9/WA09 (H9)-derived CTB stem-like cells into STBs and EVTs. (A) Cellmorphologic changes after replating of H9-derivedCTB stem-like cells and further differentiation inFCM+BMP4. Cells continued to show a flattenedphenotype with numerous multinucleated STB-likecells appearing around 4 d after replating in thismedium. (B) qRT-PCR analysis showed CTB markerssuch as CDX2 and p63 were reduced and differenti-ation markers such as CGB (in STBs) and HLA-G (inEVTs) were induced. Data were normalized to 18Sand are expressed as fold change relative to H9-derived CTB stem-like cells. Data are expressed asmeans ± SD of biological triplicates; *P < 0.05 incomparison with the values of CTB stem-like cells.(C) Immunostaining of replated H9-derived CTBsshows the loss of CTB markers such as CDX2, TEAD4,and YAP following treatment with FCM+BMP4. Nu-clei were counterstained with DAPI (blue). (D) Secre-tion of total hCG and MMP2 after 2–6 d of terminaldifferentiation of CTB stem-like cells in FCM+BMP4.Data were normalized to total DNA content and areexpressed as means ± SD of biological triplicates; *P <0.05 in comparison with the values of CTB stem-likecells. (E) Matrigel invasion assay. StemPro-adaptedH9 cells and H9-derived CTBs were plated on Matri-gel-coated membranes with 8-μm pores. After 6 d offurther culture in StemPro (H9 cells) or FCM+BMP4(H9-derived CTBs), cells that had invaded the undersideof the membranes in the invasion chamber wereimmunostained for KRT7 (red) and HLA-G (green).Nuclei were counterstained with DAPI (blue).
E3886 | www.pnas.org/cgi/doi/10.1073/pnas.1604747113 Horii et al.
Dow
nloa
ded
by g
uest
on
May
27,
202
0
DiscussionPrevious studies of trophoblast differentiation of hPSCs havefocused on the induction of CDX2, a transcription factor knownto be required for the maintenance of TSCs in the mouse (6, 7)and that also is expressed in the early human TE (12, 13).However, because CDX2 is also a marker of early mesendodermprecursors (23), and because short-term, high-dose BMP4treatment is known to induce this lineage (42), assessment ofcells using this marker alone is insufficient to establish TEidentity. We have identified another marker, p63, which identifiesproliferative CTBs in the early human placenta (21, 25, 26). p63(specifically, the N-terminal–truncated form of this protein knownas “ΔNp63”) has been identified previously as a marker of stemcells in stratified epithelia (43) and also is known to be induced byBMP4 during epidermal differentiation of pluripotent stem cellsof both mouse and human origin (44). In this study we confirmthat the early human placenta is composed of a population ofCDX2+-p63+ double-positive CTBs, which we propose as the CTB“stem” cell compartment. p63 has been shown to be enriched, atleast at the RNA level, in even earlier stages of human tropho-blast, specifically, in the TE of human blastocysts (45). Thereforeit is possible that this CDX2+-p63+ double-positive populationmay emerge in the human blastocyst and persist in a subset oftrophoblasts in the early postimplantation placenta.This finding is significant, particularly because of the contro-
versy surrounding the BMP4-based model of trophoblast in-duction of hPSCs. These models have been questioned, basedmostly on observations in the mouse system, in which signalingthrough BMP receptors appears to be required only for meso-derm induction and not for trophoblast lineage specification (23,46). The identification of BMP4-inducible p63, which is dis-pensable for mouse placentation (47), as an early marker ofhuman proliferative CTB, may explain why BMP4 induces theTE lineage in hPSCs but appears not to be required for the sameprocess in mice (46).Here we outline a simple two-step protocol by which we can
reproducibly generate first a pure population of CDX2+/p63+
CTB stem-like cells and subsequently a combination of bothhCG-producing STBs and MMP2-producing, invasive EVTs.This protocol is superior to previously described protocols inseveral ways. First, it is a robust protocol that uses low-dose
BMP4 to generate a uniform population of CTB stem-like cells,consistently, in a short period from StemPro-adapted hPSCs,without induction of the mesoderm marker T/BRACHYURY.We routinely performed flow cytometric analysis, using EGFR asa marker of CTBs, to monitor the day on which surface ex-pression of this marker peaks and to assign those cells as hPSC-derived CTBs before replating for the second step. Second, theability to replate the hPSC-derived CTBs allows large numbersof differentiated trophoblasts to be obtained starting with arelatively small number of undifferentiated hPSCs. Finally, andperhaps most importantly, unlike previous one-step protocols(including both FCM+BMP4 and BAP), our two-step protocolgives us the ability to separate the study of trophoblast lineage
Fig. 5. Low oxygen tension inhibits the differentiation of hESC-derivedCTBs into STBs. (A) Decreased hCG secretion at 2, 3, and 4 d after replating ofCTBs in 2% oxygen compared with CTBs replated in 20% oxygen, as mea-sured by ELISA and normalized to DNA content. (B) qRT-PCR analysis of theSTB markers CGA, CGB, and PSG4. Data were normalized to 18S and areexpressed as fold change relative to hESC-derived CTB stem-like cells. Alldata are expressed as means ± SD of biological triplicates; *P < 0.05 incomparison of cells in 20% oxygen and cells in 2% oxygen on the same day.
Fig. 6. Low oxygen tension induces EVT differentiation in an HIF complex-dependent manner. (A) Western blot for ARNT in undifferentiated hESCs stablyinfected with lentivirus expressing either scrambled shRNA or ARNT-specificshRNA (shARNT). (B) Western blot for HIF1α in hESCs stably infected with len-tivirus expressing either scrambled shRNA or ARNT-specific shRNA, differenti-ated into CTBs, and then replated and cultured in 2% oxygen for the indicatednumber of hours. (C) qRT-PCR analysis of HIF target genes in CTBs, derivedfrom either scrambled- or ARNT-specific shRNA-infected hESC, at day 4 post-differentiation in FCM+BMP4 (day CTB+4) in 20% or 2% oxygen. Data werenormalized to 18S and are expressed as fold change relative to scrambled/20%oxygen. (D) Surface expression of HLA-G (an EVT marker) was analyzed by flowcytometry of scrambled and shARNT-infected hESC-derived CTBs cultured in 20% or2% oxygen for 2 d after replating. All samples are compared with isotype. (E)Increased MMP2 secretion at 2 (CTB+2) and 3 (CTB+3) d after replating ofscrambled shRNA- and shARNT-infected hESC-derived CTBs in 2% oxygen asmeasured by ELISA and normalized to DNA content. (F) qRT-PCR analysis of EVT-associated genes HLA-G and HTRA4 in scrambled and shARNT-infected hESC-derived CTBs 2 d after replating in 20% or 2% oxygen. Data were normalized to18S and are expressed as fold change relative to scrambled/20% oxygen. All dataare expressed as means ± SD of biological triplicates; *P < 0.05.
Horii et al. PNAS | Published online June 20, 2016 | E3887
DEV
ELOPM
ENTA
LBIOLO
GY
PNASPL
US
Dow
nloa
ded
by g
uest
on
May
27,
202
0

specification (hPSC → CTB) from terminal trophoblast differ-entiation (CTB → STB and EVT). This ability is important,particularly because it allows the characterization of differentstages of trophoblast differentiation and will improve compara-tive studies with primary cells.One of the major reasons hPSCs have not been widely used for
studying early implantation events has been the paucity of dataregarding their ability to model human trophoblast differentiationduring both normal development and disease. One recent studyshowed blunted trophoblast differentiation of hESCs carrying anunbalanced chromosomal translocation t (11, 22) after BMP4treatment (48). Because this translocation is commonly associatedwith miscarriage, the authors argued that the hESCs carrying thischromosomal aberration are a good model for studying implan-tation failure (48). However, because the mechanism of implan-tation failure in these cases is not known, the model could not bevalidated further. We decided to evaluate two well-establishedphenomena in trophoblast differentiation: the response of CTBsto low oxygen tension and the differentiation defects of T21-affected CTBs.The culture of primary CTBs in low oxygen is known to inhibit
hCG production and reduce the expression of STB-associatedgenes (28, 29), a phenotype that was replicated with hPSC-derivedCTBs in our study. The effect of low oxygen on EVT differenti-ation is more complex. Low oxygen tension appears to induce thefirst step of EVT differentiation, seen as the expansion of theHLA-G+ cell column EVTs, in both first-trimester human pla-cental explants (31, 49) and isolated primary trophoblasts (30).However, differentiation into mature, invasive EVT appears to be
thwarted in low oxygen (29, 49, 50). Using our two-step protocol,we evaluated the first step of EVT differentiation, which involvesthe induction of HLA-G expression, and found that in low (2%)oxygen the surface HLA-G+ cells were expanded in a HIF complex-dependent manner. We could not evaluate the second stepof EVT differentiation (i.e., invasion) independently, because wecould not replate the HLA-G+ cells.We next turned to another well-studied model of a human
trophoblast differentiation defect: T21. As mentioned previously,it is known that T21 placentas maintain a continuous layer ofCTBs well into the second trimester (38). We evaluated theseplacentas further by immunohistochemistry and noted that CDX2is maintained longer in the CTBs of T21 placentas than in euploidplacentas. Interestingly, with BMP4 treatment, T21-hPSCs in-duced CDX2 at much higher levels and also maintained thismarker longer after the induction of terminal trophoblast differ-entiation (step 2). More importantly, and again similar to previousstudies using primary T21 CTBs (39–41), T21-hPSC–derivedCTBs showed a defect in the formation of multinucleated STBs,which was alleviated with Activin-A treatment. These data show,for the first time to our knowledge, a well-established trophoblastdifferentiation defect that can be recapitulated using hPSCs.There remain some limitations to the use of hPSCs for mod-
eling trophoblast differentiation. In particular, to date we havebeen unable to maintain the p63+/CDX2+ CTBs in an undif-ferentiated state. Several factors may be at play here, includingsuboptimal culture conditions and/or insufficient expression offactors promoting self-renewal. With respect to the former, weare in the process of performing small-molecule screens and
Fig. 7. Both T21 placenta and T21-hPSC–derived CTBs showed delay in differentiation to CTBs. (A) Six T21 and five normal (euploid) placentas, all between14- and 15-wk gestational age, were immunostained for CDX2. Representative micrographs show a continuous layer of CDX2+ CTBs in T21 villi and fewer suchcells in normal villi. (B) For each of the areas of the placenta shown in A, five random areas were selected. CDX2-positive and -negative CTBs were quantified.Data are expressed as means ± SD; *P < 0.05. (C) Flow cytometry showed maximum expression of EGFR (a surface CTB marker) on day 6 in T21-hPSCs and onday 4 in the disomy control hPSCs, showing the delay in CTB induction in the T2-hPSCs. All results are shown in comparison with the isotype control. (D) Basedon qRT-PCR, T21 cells showed a delay both in the reduction of the pluripotency marker POU5F1/OCT4 and in the induction of the CTB marker CDX2. CDX2levels also were higher and were maintained longer in T21-hPSCs. Data were normalized to 18S and are expressed as fold change relative to disomy day 0.Data are expressed as means ± SD of biological triplicates; *P < 0.05.
E3888 | www.pnas.org/cgi/doi/10.1073/pnas.1604747113 Horii et al.
Dow
nloa
ded
by g
uest
on
May
27,
202
0
have identified factors that maintain p63 and CDX2; neverthe-less, these factors do not appear to be sufficient for maintainingself-renewal in this cell population. This result has led us toconclude that other factors are likely required for CTB stem cellmaintenance. Candidates for such factors, insufficiently inducedfollowing BMP4 treatment of hPSCs, include ELF5, an imprintedgene and a member of the Ets family of transcription factors,which is required for maintenance of the TSC compartment inmice (51). In mice, the Elf5 promoter is hypomethylated specifi-cally in TSCs, in which this gene is expressed, and is hyper-methylated in ESCs, in which the gene is silent (9). ELF5 isexpressed in human trophoblasts of the early placenta, where itspromoter has been shown to be hypomethylated (52). In contrast,ELF5 is not expressed in the TE of the human preimplantationblastocyst (13). As in previous studies (20), we do see induction ofELF5 following BMP4 treatment; however, this induction is weak(1.5- to twofold in all lines tested) and does not reach the levels ofexpression seen in postimplantation CTBs (24). Therefore, it ispossible that the hPSC-derived CTBs are more similar to preim-plantation TE than to early postimplantation CTBs. In addition,the variability of expression of imprinted genes in different hPSClines has been described in both the undifferentiated and differ-entiated states and has been attributed mostly to epigenetic in-
stability during derivation and culture (53, 54). For this reason, itis likely that the culture conditions for both the propagation ofhPSCs and their subsequent differentiation into trophoblasts mustbe optimized in a way that promotes the expression of the trophoblast-associated imprinted genes, such as ELF5. Finally, high expres-sion of ELF5 has been noted recently in new hESC lines derivedfrom earlier-stage human embryos. These lines have an en-hanced ability to differentiate into trophoblast and reportedlymaintain the TSC state (55); thus, it is also possible that onlyearly-stage embryos have the correct epigenetic landscape forthe derivation and maintenance of true human TSCs.We also are still in the process of defining the culture condi-
tions required for inducing pure subpopulations of either STBsor EVTs. Under the current culture conditions, HLA-G+
cells appear to peak 2 d after the replating of hPSC-derivedCTBs in FCM+BMP4, whereas the number of multinucleated hCG-secreting cells peaks 4 d after replating. That these cell subpopu-lations peak at different times and diminish soon thereafter mayindicate that they are competing with each other and/or that theculture conditions are suboptimal for their maintenance andmaturation. Additional optimization of culture conditions, in-cluding changes in extracellular matrix and 3D culture, may berequired to enhance further the utility of hPSC-derived trophoblasts
Fig. 8. T21-hPSC–derived CTBs showed blunted fusion, which can be rescued morphologically by Activin A treatment. (A) For the second step, T21 CTBsshowed delay in the loss of the CTB markers CDX2 and ELF5. Data were normalized to 18S and are expressed as fold change relative to disomy day CTB+2.(B) Both T21 and disomy hESCs were differentiated into CTBs (with 4 and 6 d of EMIM+BMP4 treatment for normal and T21-hPSCs, respectively). Four daysafter the hESC-derived CTBs were replated, cells were fixed and stained with ZO-1 (red) and DAPI (blue). (C) Cells and nuclei were counted in five randomareas of the slide, and the fusion index was calculated by the indicated formula. (D) By qRT-PCR, CGA was higher in T21 hESCs, but CGB expression was similarin both lines. (E) hCG secretion was higher in T21-hESC–derived trophoblasts. Data were normalized to the total DNA content. (F) Both T21 and disomy hESCswere differentiated into CTBs (with 4 and 6 d of EMIM+BMP4 treatment for normal and T21-hESCs, respectively). The CTBs then were replated and cultured inFCM+BMP4 with or without Activin A for 4 d. Cells were fixed again and stained with ZO-1 and DAPI, and the fusion index was calculated as stated above. Alldata are expressed as mean ± SD; *P < 0.05.
Horii et al. PNAS | Published online June 20, 2016 | E3889
DEV
ELOPM
ENTA
LBIOLO
GY
PNASPL
US
Dow
nloa
ded
by g
uest
on
May
27,
202
0

as a model. Overall, however, given the proper expression of manytrophoblast-associated genes, in proper progression from CTBinduction to STB/EVT differentiation, the proper response of theCTB stem-like cells to low oxygen, and the ability to recapitulatean STB differentiation defect with T21-affected iPSCs, we main-tain that the BMP4-based protocol, particularly the two-stepprotocol described herein, provides a bona fide model for thestudy of early human trophoblast differentiation. Currently, theonly other human model that provides a bipotential trophoblastprogenitor cell population is primary CTBs isolated from first-trimester placental tissue, to which access is highly limited. Otherhuman trophoblast cell lines, whether derived from choriocarci-noma (trophoblastic tumor) tissue or isolated primary trophoblasts,represent previously committed trophoblast lineages (either villousor extravillous) and thus do not provide a platform for evaluatinglineage segregation and TSC fate decisions. Our two-step protocolpresents a system for studying both the initial specification of thehuman trophoblast lineage and subsequent differentiation intoboth villous and extravillous lineages. We therefore proposethat, except in primary cells, our two-step protocol is superiorto other currently available models for studying early humantrophoblast differentiation.
Materials and MethodshPSC Culture and Differentiation. The hPSC aspect of the research wasperformed under a protocol approved by the University of California, SanDiego Institutional Review Board and Embryonic Stem Cell Research Over-sight Committee. Before differentiation, ESCs (WA09/H9 and WA01/H1 linesfrom WiCell and SIVF21 trisomy and disomy lines) and iPSCs [C1 (T21) and C5(disomy) lines] (56) were cultured under feeder-free conditions in StemPro(Invitrogen) medium with 12 ng/mL recombinant basic FGF (bFGF) (Invitrogen)on plates coated with Geltrex (1:200; BD Biosciences). In the first step, cellswere plated relatively sparsely (8 × 103/cm2 for WA09/H9 and 105/cm2 for SIVF,C1, and C5 cell lines). Cells were passaged with StemDS (ScienCell ResearchLaboratories) or Accutase (Life Technology) every 3–5 d. Only passages 10–20post-StemPro adaptation were used for all experiments described here. For thefirst phase of TE differentiation, hPSCs were first treated with a StemPro(Thermo Fisher)-based basal medium [KnockOut DMEM (Thermo Fisher)/F12containing 1% insulin-transferrin-selenium A, 1× nonessential amino acid, 2%(wt/vol) BSA, 2 mM L-glutamine, 100 ng/mL heparin sulfate] designated as“EMIM” (17) for 2 d. The cells thenwere switched to EMIMmedium containing10 ng/mL human BMP4 (R&D Systems) for 2–4 d, at which point they weredesignated hPSC-derived CTBs. For terminal differentiation, the hPSC-derivedCTBs were passaged using 0.05% trypsin and were replated on Geltrex-coatedplates in FCM [KnockOut DMEM/F12 containing 20% (vol/vol) KnockOut serumreplacement (Knockout SR; Thermo Fisher), 1× GlutaMAX (Thermo Fisher), and1× nonessential amino acid, 0.1 mM 2-mercaptoethanol; conditioned on irra-diated mouse embryonic fibroblasts for 24 h], supplemented with 10 ng/mLhuman BMP4 for the indicated number of days. For Activin-A rescue experi-ments, in addition to FCM+BMP4, the cells were treated with 5 ng/mL ActivinA (Stemgent). For culture under low oxygen, cells were incubated in an Xvivosystem under 2% oxygen (BioSpherix). For all experiments in hypoxia, themedium was changed, and cells were lysed (for RNA or protein analysis) orfixed (for flow cytometry) in the Xvivo work chamber under 2% oxygen.
Generation of ARNT-Knockdown hESC Lines. pLKO.1-based scrambled shRNAwas obtained from Addgene. A scrambled shRNA and a set of five MissionshRNA Lentiviral constructs targeting the human ARNT gene were purchasedand packaged for transduction according to the manufacturer’s instructions(Sigm-Aldrich a). HEK293FT cells were transfected with the shRNA constructsDelta 8.2 and VSV-G using Lipofectamine 2000 (Sigma-Aldrich) according tothe manufacturer’s instructions. Lentiviral supernatants were concentratedwith PEG-it virus precipitation solution (System Biosciences). The concen-trated viral particles then were incubated with target cells in the presence of10 mg/mL Polybrene (Sigma-Aldrich). Packaging and infection efficiencywere tested using a GFP-expressing lentivirus (in the same pCMV-lentiviralconstruct used for ARNT). Of the five ARNT-specific shRNA constructs, threeworked best, based on protein knockdown in HEK293T cells; therefore,subsequent experiments were performed with this subset of three shRNAspooled together. hESCs were stably infected; after 3 d of infection, cellswere treated with 5 μg/mL puromycin (Sigma-Aldrich) to select cells stablytransduced over at least three passages. Infection efficiency was
determined by GFP expression. Knockdown efficiency was determined byWestern blot.
Human Placental Tissue Collection and Immunohistochemistry. Human pla-cental tissueswere collected under a protocol approved by theHumanResearchProtections Program Committee of the University of California, San DiegoInstitutional ReviewBoard; all patients gave informed consent for the collectionand use of these tissues. Serial sections of formalin-fixed, paraffin-embeddedplacental tissues were stained with mouse anti-p40 (ΔNp63-specific antibody,clone BC28; Biocare Medical) and rabbit anti-CDX2 (clone EPR2764Y; Abcam)antibodies, using a Ventana Discovery Ultra automated immunostainer withstandard antigen retrieval and reagents per the manufacturer’s protocol.
Flow Cytometric Analysis. For flow cytometry, cells were fixed and permeabilizedwith either 4% (wt/vol) paraformaldehyde for 10–15 min or with 100% ice-cold methanol for 30 min and then were washed three times in PBS sup-plemented with 10% (vol/vol) FBS. Cells then were incubated at roomtemperature for 1 h in flow cytometry (FC) buffer (0.5% BSA and 1% FBS inPBS) with an allophycocyanin (APC)-conjugated mouse anti-human EGFRantibody (clone AY13; BioLegend), phycoerythrin (PE)-conjugated mouseanti–HLA-G (MEM-G/9; EXBIO), or FITC-conjugated mouse anti-human p63antibody (clone 4A4; Santa Cruz). APC- or PE-conjugated mouse IgG (cloneMPOC-21; BioLegend), or FITC-conjugated mouse IgG (Santa Cruz) was usedas isotype IgG control. Cells were washed three times with FC buffer, andanalysis was carried out using a BD FACS-Canto Flow cytometer.
Immunostaining of Cells. For immunofluorescence staining, cells grown onGeltrex-coated coverslips were fixed with 4% (wt/vol) paraformaldehyde inPBS at room temperature for 10 min. Cells then were permeabilized with0.3% Triton X-100 for 10 min and incubated with primary antibodies,including mouse anti-p63 (4A4 clone; Sigma-Aldrich), rabbit anti-Ki67(ab15580; Abcam), mouse anti-KRT7 (clone OV-TL 12/30; Invitrogen), mouseanti-YAP (sc-15407; Santa Cruz), rabbit anti-EGFR (sc-03; Santa Cruz), rabbitanti-TEAD4 (HPA056896; Abcam), rabbit anti-CDX2 (clone EPR2764Y;Abcam), rabbit anti-OCT4 (ab19857; Abcam), rabbit anti–ZO-1 (ab59720;Abcam), or mouse anti–HLA-G (clone 4H84; Abcam) and were visualized byAlexa Fluor 488- or 595-conjugated goat secondary antibodies (Invitrogen).For nuclear staining, cells were incubated with DAPI (Invitrogen) for 5 min.
RNA Isolation and Quantitative Real-Time PCR. Total RNA was extracted usingeither the mirVana RNA Isolation Kit (Ambion) or the NucleoSpin RNA II Kit(Macherey-Nagel) according to the manufacturer’s protocol. The purity andconcentration of the samples were assessed with a NanoDrop 2000/2000cSpectrophotometer (Thermo Fisher Scientific). cDNA was prepared from500 ng RNA using iScript (Bio-Rad) in a 20-μL reaction and was diluted 1:5with nuclease-free water. qRT-PCR was performed using 4 μL of the dilutedcDNA, along with 500 nM of each primer and POWER SYBR Green PCR mastermix (Applied Biosystems) in a total reaction volume of 20 μL. qRT-PCR wasperformed using a System 7300 instrument (Applied Biosystems) or Step OnePlus (Applied Biosystems) and a one-step program: 95 °C for 10 min; 95 °C for30 s; 60 °C for 1 min, for 40 cycles. Unless otherwise stated, each experimentwas performed in triplicate, and all results were normalized against 18S rRNA.Relative mRNA expression levels, compared with 18S rRNA, were determined bythe comparative cycle threshold (ΔΔCT) method. All primer pairs (Table S1) werechecked for specificity using BLAST analysis and were checked by both agarosegel electrophoresis and thermal dissociation curves to ensure amplification of asingle product with the appropriate size and melting temperature.
Cell Invasion Assay. Matrigel-coated Transwell membranes (8.0-μm pore in-serts in 24-well BioCoat chambers; BD Biosciences) were used to perform invitro invasion assays. hPSCs and hPSC-derived CTBs were collected withStemDS or 0.05% trypsin, respectively. Cells were washed in cold 1× PBS andthen were resuspended in StemPro medium (hPSCs) or FCM plus 10 ng/mLBMP4 (hPSC-derived CTBs). Cells (1 × 104 per well) were placed in the upperchamber, and StemPro plus bFGF (for hPSCs) or FCM+BMP4 for (hPSC-derivedCTBs) was placed in the lower chamber. After 4 d, cells on the upper surfacewere removed by scrubbing with a cotton swab; filters then were fixed with4% paraformaldehyde and stained with mouse anti-KRT7 and rabbit anti-HLA-Gantibodies (as detailed above). Cells were visualized by goat anti-rabbit AlexaFluor 488- and goat anti-mouse Alexa Fluor 595-conjugated secondary anti-bodies. For nuclear staining, cells were incubated with DAPI for 5 min.
hCG Hormone and MMP2 Secretion Assays. Cell-culture supernatants werecollected and stored at −80 °C until use. Levels of total hCG were quantified
E3890 | www.pnas.org/cgi/doi/10.1073/pnas.1604747113 Horii et al.
Dow
nloa
ded
by g
uest
on
May
27,
202
0

using the hCG ELISA Kit (HC251F; Calbiotech Inc.), and levels of total MMP2were quantified using the human MMP2 ELISA Kit (ab100606; Abcam)according to the manufacturers’ protocols. The results were normalized tototal DNA content, quantified by DNeasy (Qiagen).
Statistical Analysis.Unless otherwise stated, the data presented are themean±SD of biological triplicates; the results shown are representative of three tofive independent experiments, each performed using a different preparationof cells. Student’s t test was performed, and P values below 0.05 were taken toindicate a statistically significant difference between the populations sampled.
ACKNOWLEDGMENTS. Uli Schmidt (Stem Cell Laboratory) derived the originalSIVF21 ESC line from which the subsequent SIVF21-disomy ESC line was derivedthrough subculture. This work was supported by California Institute for Regen-erative Medicine (CIRM) New Faculty Award RN2-00931, CIRM Physician-ScientistAward RN3-06396, and NIH/National Institute of Child Health and DevelopmentGrant R01-HD071100. M.H. was supported through CIRM Research and TrainingGrant TG2-01154 to the University of California, San Diego (UCSD). A.K.W. wassupported by UCSD Respiratory Biology Training Grant T32-HL098062. Y. Liu wassupported by CIRM Grant RT1-01107-1, the Memorial Hermann Foundation-Staman Ogilvie Fund, the Bentsen Stroke Center Fund, and Mission Connect–Texas Institute for Rehabilitation and Research Foundation.
1. Roberts RM, Fisher SJ (2011) Trophoblast stem cells. Biol Reprod 84(3):412–421.2. Senner CE, Hemberger M (2010) Regulation of early trophoblast differentiation - lessons
from the mouse. Placenta 31(11):944–950.3. Gasperowicz M, Natale DR (2011) Establishing three blastocyst lineages–Then what?
Biol Reprod 84(4):621–630.4. Pfeffer PL, Pearton DJ (2012) Trophoblast development. Reproduction 143(3):231–246.5. Tanaka S, Kunath T, Hadjantonakis AK, Nagy A, Rossant J (1998) Promotion of tro-
phoblast stem cell proliferation by FGF4. Science 282(5396):2072–2075.6. Niwa H, et al. (2005) Interaction between Oct3/4 and Cdx2 determines trophectoderm
differentiation. Cell 123(5):917–929.7. Strumpf D, et al. (2005) Cdx2 is required for correct cell fate specification and
differentiation of trophectoderm in the mouse blastocyst. Development 132(9):2093–2102.
8. Russ AP, et al. (2000) Eomesodermin is required for mouse trophoblast developmentand mesoderm formation. Nature 404(6773):95–99.
9. Ng RK, et al. (2008) Epigenetic restriction of embryonic cell lineage fate by methyl-ation of Elf5. Nat Cell Biol 10(11):1280–1290.
10. James JL, Carter AM, Chamley LW (2012) Human placentation from nidation to5 weeks of gestation. Part I: What do we know about formative placental develop-ment following implantation? Placenta 33(5):327–334.
11. Li Y, Parast MM (2014) BMP4 regulation of human trophoblast development. Int J DevBiol 58(2-4):239–246.
12. Niakan KK, Eggan K (2013) Analysis of human embryos from zygote to blastocystreveals distinct gene expression patterns relative to the mouse. Dev Biol 375(1):54–64.
13. Blakeley P, et al. (2015) Defining the three cell lineages of the human blastocyst bysingle-cell RNA-seq. Development 142(20):3613.
14. Kunath T, et al. (2014) Developmental differences in the expression of FGF receptorsbetween human and mouse embryos. Placenta 35(12):1079–1088.
15. Xu RH, et al. (2002) BMP4 initiates human embryonic stem cell differentiation totrophoblast. Nat Biotechnol 20(12):1261–1264.
16. Wu Z, et al. (2008) Combinatorial signals of activin/nodal and bone morphogenicprotein regulate the early lineage segregation of human embryonic stem cells. J BiolChem 283(36):24991–25002.
17. Erb TM, et al. (2011) Paracrine and epigenetic control of trophectoderm differenti-ation from human embryonic stem cells: The role of bone morphogenic protein 4 andhistone deacetylases. Stem Cells Dev 20(9):1601–1614.
18. Aghajanova L, et al. (2012) Comparative transcriptome analysis of human trophectodermand embryonic stem cell-derived trophoblasts reveal key participants in early implanta-tion. Biol Reprod 86(1):1–21.
19. Sudheer S, Bhushan R, Fauler B, Lehrach H, Adjaye J (2012) FGF inhibition directs BMP4-mediated differentiation of human embryonic stem cells to syncytiotrophoblast. StemCells Dev 21(16):2987–3000.
20. Amita M, et al. (2013) Complete and unidirectional conversion of human embryonicstem cells to trophoblast by BMP4. Proc Natl Acad Sci USA 110(13):E1212–E1221.
21. Li Y, et al. (2013) BMP4-directed trophoblast differentiation of human embryonicstem cells is mediated through a ΔNp63+ cytotrophoblast stem cell state.Development 140(19):3965–3976.
22. Warmflash A, Sorre B, Etoc F, Siggia ED, Brivanlou AH (2014) A method to recapitu-late early embryonic spatial patterning in human embryonic stem cells. Nat Methods11(8):847–854.
23. Bernardo AS, et al. (2011) BRACHYURY and CDX2 mediate BMP-induced differenti-ation of human and mouse pluripotent stem cells into embryonic and extraembryoniclineages. Cell Stem Cell 9(2):144–155.
24. Lee CQ, et al. (2016) What is trophoblast? A combination of criteria define humanfirst-trimester trophoblast. Stem Cell Rep 6(2):257–272.
25. Lee Y, et al. (2007) A unifying concept of trophoblastic differentiation and malig-nancy defined by biomarker expression. Hum Pathol 38(7):1003–1013.
26. Li Y, Moretto-Zita M, Leon-Garcia S, Parast MM (2014) p63 inhibits extravillous tro-phoblast migration and maintains cells in a cytotrophoblast stem cell-like state. Am JPathol 184(12):3332–3343.
27. McMaster MT, et al. (1995) Human placental HLA-G expression is restricted to dif-ferentiated cytotrophoblasts. J Immunol 154(8):3771–3778.
28. Alsat E, et al. (1996) Hypoxia impairs cell fusion and differentiation process in humancytotrophoblast, in vitro. J Cell Physiol 168(2):346–353.
29. Nelson DM, Johnson RD, Smith SD, Anteby EY, Sadovsky Y (1999) Hypoxia limits differ-entiation and up-regulates expression and activity of prostaglandin H synthase 2 in cul-tured trophoblast from term human placenta. Am J Obstet Gynecol 180(4):896–902.
30. Robins JC, Heizer A, Hardiman A, Hubert M, Handwerger S (2007) Oxygen tensiondirects the differentiation pathway of human cytotrophoblast cells. Placenta28(11-12):1141–1146.
31. Genbacev O, Zhou Y, Ludlow JW, Fisher SJ (1997) Regulation of human placentaldevelopment by oxygen tension. Science 277(5332):1669–1672.
32. Dunwoodie SL (2009) The role of hypoxia in development of the Mammalian embryo.Dev Cell 17(6):755–773.
33. Maltepe E, et al. (2005) Hypoxia-inducible factor-dependent histone deacetylase ac-tivity determines stem cell fate in the placenta. Development 132(15):3393–3403.
34. Chakraborty D, Rumi MA, Konno T, Soares MJ (2011) Natural killer cells directhemochorial placentation by regulating hypoxia-inducible factor dependent tro-phoblast lineage decisions. Proc Natl Acad Sci USA 108(39):16295–16300.
35. Bjørn SF, Hastrup N, Larsen JF, Lund LR, Pyke C (2000) Messenger RNA for membrane-type 2 matrix metalloproteinase, MT2-MMP, is expressed in human placenta of firsttrimester. Placenta 21(2-3):170–176.
36. Isaka K, et al. (2003) Expression and activity of matrix metalloproteinase 2 and 9 inhuman trophoblasts. Placenta 24(1):53–64.
37. Wang LJ, Cheong ML, Lee YS, Lee MT, Chen H (2012) High-temperature requirementprotein A4 (HtrA4) suppresses the fusogenic activity of syncytin-1 and promotestrophoblast invasion. Mol Cell Biol 32(18):3707–3717.
38. Roberts L, Sebire NJ, Fowler D, Nicolaides KH (2000) Histomorphological features ofchorionic villi at 10-14 weeks of gestation in trisomic and chromosomally normalpregnancies. Placenta 21(7):678–683.
39. Frendo JL, et al. (2001) Overexpression of copper zinc superoxide dismutase impairshuman trophoblast cell fusion and differentiation. Endocrinology 142(8):3638–3648.
40. Gerbaud P, et al. (2011) Mesenchymal activin-A overcomes defective human trisomy21 trophoblast fusion. Endocrinology 152(12):5017–5028.
41. Pidoux G, et al. (2012) Review: Human trophoblast fusion and differentiation: Lessonsfrom trisomy 21 placenta. Placenta 33(Suppl):S81–S86.
42. Zhang P, et al. (2008) Short-term BMP-4 treatment initiates mesoderm induction inhuman embryonic stem cells. Blood 111(4):1933–1941.
43. Senoo M, Pinto F, Crum CP, McKeon F (2007) p63 Is essential for the proliferativepotential of stem cells in stratified epithelia. Cell 129(3):523–536.
44. Aberdam D (2008) Epidermal stem cell fate: What can we learn from embryonic stemcells? Cell Tissue Res 331(1):103–107.
45. Bai Q, et al. (2012) Dissecting the first transcriptional divergence during human em-bryonic development. Stem Cell Rev 8(1):150–162.
46. Di-Gregorio A, et al. (2007) BMP signalling inhibits premature neural differentiationin the mouse embryo. Development 134(18):3359–3369.
47. Mills AA, et al. (1999) p63 is a p53 homologue required for limb and epidermalmorphogenesis. Nature 398(6729):708–713.
48. Shpiz A, et al. (2015) Human embryonic stem cells carrying an unbalanced trans-location demonstrate impaired differentiation into trophoblasts: An in vitro model ofhuman implantation failure. Mol Hum Reprod 21(3):271–280.
49. Caniggia I, et al. (2000) Hypoxia-inducible factor-1 mediates the biological effects ofoxygen on human trophoblast differentiation through TGFbeta(3). J Clin Invest105(5):577–587.
50. Genbacev O, Joslin R, Damsky CH, Polliotti BM, Fisher SJ (1996) Hypoxia alters earlygestation human cytotrophoblast differentiation/invasion in vitro and models theplacental defects that occur in preeclampsia. J Clin Invest 97(2):540–550.
51. Donnison M, et al. (2005) Loss of the extraembryonic ectoderm in Elf5 mutants leadsto defects in embryonic patterning. Development 132(10):2299–2308.
52. Hemberger M, Udayashankar R, Tesar P, Moore H, Burton GJ (2010) ELF5-enforcedtranscriptional networks define an epigenetically regulated trophoblast stem cellcompartment in the human placenta. Hum Mol Genet 19(12):2456–2467.
53. Nazor KL, et al. (2012) Recurrent variations in DNA methylation in human pluripotentstem cells and their differentiated derivatives. Cell Stem Cell 10(5):620–634.
54. Thiagarajan RD, Morey R, Laurent LC (2014) The epigenome in pluripotency anddifferentiation. Epigenomics 6(1):121–137.
55. Zdravkovic T, et al. (2015) Human stem cells from single blastomeres reveal pathwaysof embryonic or trophoblast fate specification. Development 142(23):4010–4025.
56. Chen C, et al. (2014) Role of astroglia in Down’s syndrome revealed by patient-derivedhuman-induced pluripotent stem cells. Nat Commun 5:4430.
Horii et al. PNAS | Published online June 20, 2016 | E3891
DEV
ELOPM
ENTA
LBIOLO
GY
PNASPL
US
Dow
nloa
ded
by g
uest
on
May
27,
202
0



![10000005505-Maintenance of Human Pluripotent Stem Cells …€¦ · The maintenance and expansion of human pluripotent stem cells (human embryonic stem [ES] cells and human induced](https://img.pdfslide.us/doc/110x75/6033bf7fdddc672302645fcf/10000005505-maintenance-of-human-pluripotent-stem-cells-the-maintenance-and-expansion.jpg)

























