Embed Size (px)
Citation preview

Vol. 61, No. 4INFECrION AND IMMUNITY, Apr. 1993, p. 1185-11930019-9567/93/041185-09$02.00/0Copyright X 1993, American Society for Microbiology
Enhancement of Oxidative Response and Damage Caused byHuman Neutrophils to Aspergillus fimigatus Hyphae
by Granulocyte Colony-Stimulating Factorand Gamma Interferon
EMMANUEL ROILIDES,lt KATRIN UHLIG,1l DAVID VENZON,2PHILIP A. PIZZO,1 AND THOMAS J. WALSH`*
Infectious Diseases Section, Pediatric Branch, 1 and Biostatistics andData Management Section,2 National Cancer Institute,
Bethesda, Maryland 20892
Received 2 October 1992/Accepted 8 January 1993
Invasive aspergillosis is a serious fungal infection caused by the proliferation and invasion of Aspergillushyphae in tissue. Neutrophils (PMNs) are the most important line of defense against AspergiUlus hyphae. Toinvestigate the role of granulocyte colony-stimulating factor (G-CSF) and gamma interferon (IFN-y) againstAspergilusfiumigatus, we studied the effects of the two cytokines on the oxidative burst and the capacity ofnormal human PMNs to damage hyphae of the organism. G-CSF enhanced PMN oxidative burst measured assuperoxide anion (02) production in response to N-formylmethionyl leucyl phenylalanine, serum opsonizedhyphae, and nonopsonized hyphae by 75, 37, and 24%, respectively, compared with control PMNs (P < 0.015).IFN-y also induced increases of 52, 71, and 961%, respectively, in response to the same stimuli (P < 0.006). Inaddition, the capacity of PMNs to damage hyphae as measured by the 3-4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MMT) colorimetric metabolic assay was significantly enhanced by G-CSF andIFN-y (P < 0.01 and < 0.05, respectively). The enhancement was achieved irrespective of serum opsonizationof the hyphae, suggesting upregulatory actions of the two cytokines on signal pathways specific for opsonizedand nonopsonized hyphae. The combination of the two cytokines exhibited an additive effect at the higherconcentrations compared with the effects of the cytokines alone (P < 0.05). Pretreatment of PMNs with proteinsynthesis inhibitors showed that IFN-y activates PMN function through transcriptional regulation, whereas theeffect of G-CSF does not require new proteins. These in vitro effects suggest modulatory roles for G-CSF andIFN-y in the host defense against AspergiUus hyphae irrespective of serum opsonization and a potential utilityof the cytokines as adjuncts for the prevention and possible treatment of invasive aspergillosis.
Invasive aspergillosis has emerged as an important causeof mortality in persistently neutropenic patients with canceror aplastic anemia and in patients with phagocytic defectssuch as chronic granulomatous disease (12, 47, 48). Anincreased susceptibility to Aspergillus infections has alsobeen described in patients with AIDS (14). Aspergillusfumigatus is the most common isolate from these patients.Although the alveolar macrophages have been shown to bethe first-line effector cells that inhibit germination of theinhaledAspegillus conidia, neutrophils (PMNs) are the cellsthat damage the escaping hyphae by secretion of microbici-dal oxidative and possibly nonoxidative metabolites and thusprevent the establishment of invasive disease (15, 43). Thestudy of PMN-hypha interactions has been largely facilitatedby the recent adaptation of the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) colorimetricmethod (25, 32, 38) for the measurement of hyphal damage.
Previous studies have shown that hyphae of other fungisuch as Candida albicans are killed by PMNs following anoxidative metabolic burst (16, 26). Furthermore, in the case
* Corresponding author.t Present address: Third Department of Pediatrics, University of
Thessaloniki, Hippokration Hospital, GR-54642 Thessaloniki,Greece.
t Present address: Department of Internal Medicine, GeorgetownUniversity Hospital, Washington, DC 20007.
of C. albicans, PMNs respond to both opsonized and nonop-sonized pseudohyphae with activation of the oxidative met-abolic burst and subsequent release of microbicidal metab-olites such as hydrogen peroxide (H202), hypochlorous acid,and chloramines. Hyphae of Aspergillus spp. are also sus-ceptible to these microbicidal substances (24) and may elicitsimilar responses from the attacking PMNs.
Granulocyte colony-stimulating factor (G-CSF) andgamma interferon (IFN--y) have been shown to upregulatecertain antimicrobial PMN functions such as oxidative met-abolic burst in response to soluble stimuli (9, 23), phagocy-tosis, and bactericidal activity of PMNs (19, 41, 44). Inaddition, our previous work has suggested that each of thesecytokines enhances the oxidative metabolic burst of PMNsin response to particulate stimuli, such as C. albicansblastoconidia and pseudohyphae (40).
Little is known, however, about whether the effects ofcytokines such as G-CSF and IFN-y on the oxidative burstand fungicidal activities of PMNs against hyphae of As-pergillus spp. are similar to their effects on other fungi.Diversity of results among different fungi and cytokines hasbeen previously reported by a number of investigators (17,20, 29, 41, 42) and may be critical in pathogenesis and hostdefenses against specific fungal infections. Thus, in thisstudy, we undertook to assess the effects of the two cyto-kines on the oxidative burst of normal human PMNs inresponse to hyphae ofA. fumigatus and compare them with
1185

1186 ROILIDES ET AL.
the soluble stimulus N-formylmethionyl leucyl phenylala-nine (FMLP). In addition, we examined the effects of thecytokines alone or in combination on the fungicidal activityof PMNs.
(This study was presented in part at the 31st InterscienceConference on Antimicrobial Agents and Chemotherapy,Chicago, Ill., 29 September to 2 October 1991.)
MATERIALS AND METHODS
Organism. A well-characterized clinical isolate of A. flu-migatus from a cancer patient with invasive aspergillosiswas used for the experiments described in this report. Thestrain was preserved in frozen potato dextrose agar slants at-70°C. Conidia were harvested from the slants by scrapingthe surface of the slants and suspending the product inphosphate-buffered saline. After filtration through three lay-ers of sterile gauze, the conidia were washed twice, sus-pended by vigorous agitation in phosphate-buffered saline,and kept in suspension at 4°C. On the day before theexperiment, a suspension of 5 x 104 or 1 x 105 conidia per mlin yeast nitrogen base supplemented with 2% glucose wasmade, and 1 ml-aliquots were plated in each well of 24-wellplates (Costar, Cambridge, Mass.). The inoculated plateswere incubated for approximately 15 h at 30°C for germina-tion of the conidia to hyphae. By that time, more than 95%of the conidia had germinated to hyphae approximately 150to 200 ,um long. At that point, the hyphae either were usedimmediately for experiments or were stored at 4°C for 1 to 2h. An additional suspension of 106 conidia per ml in yeastnitrogen base supplemented with 2% glucose was incubatedat 30°C in 50-ml Falcon polypropylene tubes (Becton Dick-inson, Lincoln Park, N.J.) for approximately 15 h to yieldhyphae for superoxide production assays. After the incuba-tion, the walls of the tubes were scraped thoroughly, and thehyphae were vortexed, washed twice with Hanks balancedsalt solution (HBSS), and used in superoxide experiments.
Cytokines and other reagents. Recombinant human G-CSFproduced in Escherichia coli was obtained from Amgen Inc.,Thousand Oaks, Calif., and had a specific activity of 2 x 108U/mg of protein as determined by granulocyte-macrophagecolony formation of nonadherent bone marrow cells insemisolid media. Recombinant human IFN--y was kindlysupplied by Genentech Inc., South San Francisco, Calif.,and had a specific activity of 4 x 107 U/mg. Both prepara-tions contained <0.1 ng of protein endotoxin per mg astested by the limulus amebocyte lysate assay. Cyclohexim-ide, actinomycin D, MIT, FMLP, cytochrome c, and de-oxycholate Na were all purchased from Sigma ChemicalCo., St. Louis, Mo. Isopropanol was purchased from BakerInc., Phillipsburg, N.J. Buffers and RPMI 1640 were pre-pared at the National Institutes of Health Media Depart-ment, National Institutes of Health, Bethesda, Md. HumanAB normal serum was purchased from GIBCO Laborato-ries, Grand Island, N.Y.
Separation of PMNs. PMNs were prepared from hepa-rinized (5 to 10 U/ml) venous blood by dextran sedimenta-tion followed by centrifugation over Ficoll and hypotoniclysis of erythrocytes as previously described (4, 39). Theywere then suspended in HBSS without Ca2" and Mg' . Theresulting cell preparations consisted of more than 95% viablePMNs as judged by trypan blue exclusion and counting on ahemacytometer.Priming of PMNs. Before the assays, PMNs (2 x 107/ml)
were incubated with various concentrations of G-CSF orIFN-y in HBSS without Ca2" and Mg2+ at 37°C for various
periods. After this pretreatment, PMNs were used for assayswithout any further washing or other treatment. For someselected experiments, PMNs were pretreated with the pro-tein synthesis inhibitors cycloheximide and actinomycin Dfor 15 min prior to and during the 90-min priming of the cellswith cytokines. The concentrations of the inhibitors usedwere 1 and 5 ,ug/ml, respectively.
Superoxide production by PMNs. Production of superoxideanion by PMNs in response to the synthetic tripeptideFMLP and to hyphae of A. fumigatus was assessed by thesuperoxide dismutase-inhibitable reduction of cytochrome cspectrophotometrically (39). PMNs (106) were mixed with 50,uM cytochrome c. As a stimulus, either 0.5 p,M FMLP orhyphae in an effector-to-target cell (EST) ratio of 1/1 wereadded to the PMNs in 1 ml of HBSS, and the mixture wasincubated on a shaker at 37°C for 15 min. In some experi-ments, the hyphae were preopsonized by incubation in 50%AB human serum at 37°C for 20 to 30 min. The serum wasderived from normal donors, and its complement activitywas not defined. Control tubes containing all of the constit-uents listed above plus superoxide dismutase (40 ,ug/ml) orhyphae alone were also included. After incubation, thereaction tubes were centrifuged and the supernatants wereremoved. The amount of 02 produced was measured in thesupernatants as the difference in A550 for the reaction mix-ture and the control. For this purpose, a Gilford 260 spec-trophotometer (Ciba-Corning Diagnostics Corp., Oberlin,Ohio) was used. Superoxide produced by 106 PMNs wasthen calculated by using the millimolar extinction coefficientfor reduced cytochrome c. For each individual donor, theamount of superoxide produced by control PMNs wassubtracted from the amount of superoxide produced bycytokine-pretreated PMNs, and the absolute difference wascalculated.PMN-induced damage of hyphae. The damage to hyphae
caused by PMNs was assessed by the MIT colorimetricassay previously described in detail for assessing viability oftumor cells (32) and fungi (25, 38). This assay takes advan-tage of the fact that metabolically active (growing) hyphaeconvert the yellow tetrazolium salt MTT to a blue formazanderivative with maximum absorbance at 560 to 570 nm,whereas damaged hyphae fail to do so (25, 38). Briefly, thesupernatant of the wells was aspirated with a 19-gaugeneedle connected to a vacuum aspirator, leaving the hyphaeattached at the bottoms of the wells. Particular care wastaken not to let the hyphae dry for more than 1 to 2 min. Thehyphae adhered firmly to the bottoms of the wells and werenot detached from the plates by repetitive washings. PMNsthat had previously been treated with buffer or variousconcentrations of G-CSF or IFN-y at a final E/T ratio of 5/1,10/1, or 20/1 and in a final volume of 1 ml ofHBSS containing20 mM HEPES (N-2-hydroxyethylpiperazine-N'-2-ethane-sulfonic acid) buffer were added to the wells. In thoseexperiments, in which assessment of the effect of cytokineson the killing of opsonized hyphae was the goal, 10% ABhuman serum (100 pl) was also added to each well. After theappropriate time of incubation at 37°C, supernatants werecompletely aspirated from the wells and PMNs were lysedby adding 300 ,ul of 0.5% sodium deoxycholate. The hyphaewere then washed with sterile distilled water three times, 1ml of RPMI 1640 with L-glutamine and without phenol redbut containing 0.5% mg of MIT per ml was added to eachwell, and the plates were further incubated at 37°C for threeadditional hours. The wells were then aspirated dry, 200 ,ulof isopropanol was added to each well, and the plates wereswirled gently until all of the blue precipitate had dissolved.
INFECT. IMMUN.
CYTOKINE AND NEUTROPHIL RESPONSES TO A. FUMIGATUS 1187
Volumes (150 ,J) of isopropanol were transferred to thewells of a 96-well Immulon I plate (Dynatech Laboratories,Chantilly, Va.), and the color was measured on a TitertekMultiscan microplate spectrophotometer (Flow Laborato-ries, McLean, Va.) at the wavelengths 570 and 690 nm. Awell containing only isopropanol was used as a blank.Control wells containing hyphae and buffer only but notPMNs were included in each experiment. Fungicidal activity(damage) was calculated by using the following formula: %damage of hyphae = [(OD of control wells - OD of testwells)/OD of control wells] x 100, where OD is opticaldensity and control wells are the wells containing hyphaeand buffer only. Each condition was tested in duplicate ortriplicate, and the results were averaged.
Statistics. Differences in superoxide production and hy-phal damage between baseline and individual concentrationswere assessed by unbalanced two-way analysis of variance,with PMN donor and concentration as factors. Least-squares regression analysis was employed to test for lineartrends as functions of log(10 + concentration) or log(1 +concentration). In every regression model, the baseline foreach donor was included as a parameter to be estimated. Thedistributions of residuals were consistent with normality. AllP values reported are two tailed and are derived from theanalysis of variance unless otherwise stated.
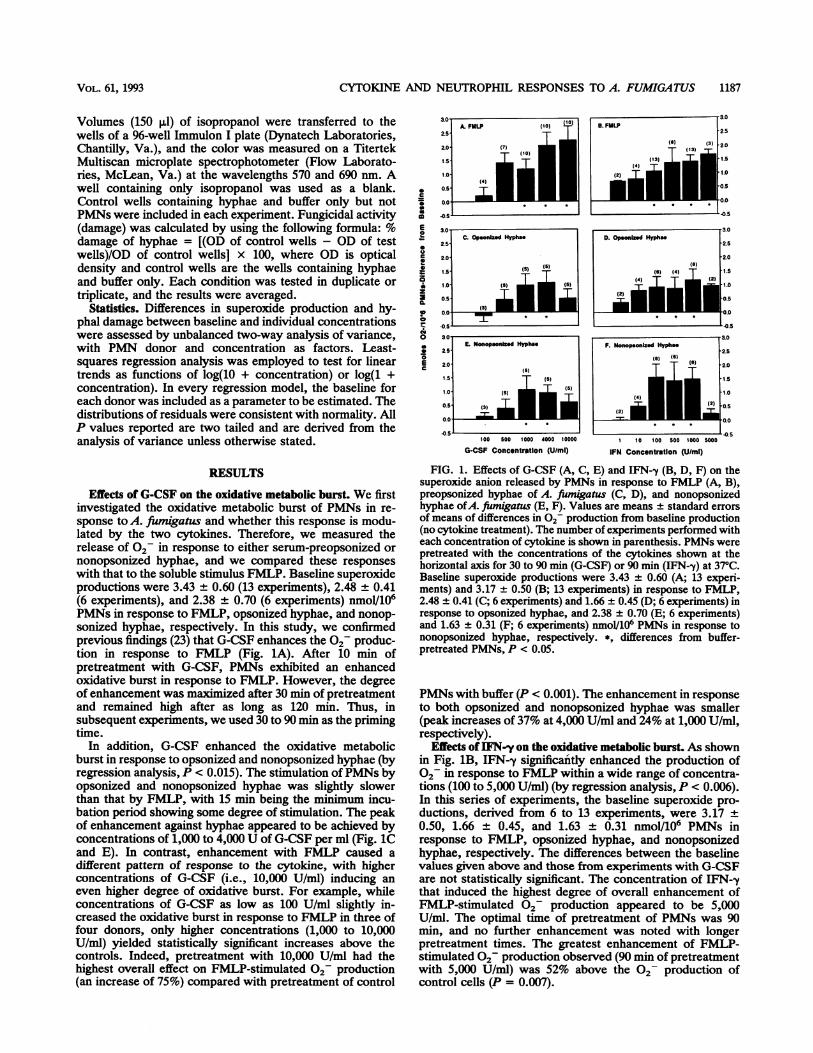
3.0 (10)A. FMILP (1o) E.FUIAP
2s -2.
2.0-(7) ~~~ ~~~~~~~~~~~(a)(3) -2.0O(0) (13)
10O (2)1 1.0
c41's
0
011Ir-r.109z10a0.
p0zA0
E0c
0.0
-0.5
2.0-C. Opsonized Hyph..
2.5'
2.0'
1.5'
2.0's'.O55 T (5
0.5-
0.c
0.)EG NonoponcazdHyp (/e
2.5-
2.0'(5)
1.5 F (5)
1.0'
-0.5100 500 1000 4000 10000
G-CSF Concentration (U/mi)
D. Op,mkoi Hyph2I1 1 . 2 . 5~~~~~~~~~~.12.0
(8)(6) (4)(T 1.5
1.0(2)
0.5
0.0
3.0F. Nonopseoie dHyph 2.5
2.5
1.0(4)15
(2) o0.5(2)
0.0
-0.51 10 100 500 1000 5000
IFN Concentration (U/ml)
RESULTS
Effects of G-CSF on the oxidative metabolic burst. We firstinvestigated the oxidative metabolic burst of PMNs in re-sponse to A. fiumigatus and whether this response is modu-lated by the two cytokines. Therefore, we measured therelease of 02 in response to either serum-preopsonized ornonopsonized hyphae, and we compared these responseswith that to the soluble stimulus FMLP. Baseline superoxideproductions were 3.43 + 0.60 (13 experiments), 2.48 0.41(6 experiments), and 2.38 + 0.70 (6 experiments) nmol/106PMNs in response to FMLP, opsonized hyphae, and nonop-sonized hyphae, respectively. In this study, we confirmedprevious findings (23) that G-CSF enhances the 02 produc-tion in response to FMLP (Fig. 1A). After 10 min ofpretreatment with G-CSF, PMNs exhibited an enhancedoxidative burst in response to FMLP. However, the degreeof enhancement was maximized after 30 min of pretreatmentand remained high after as long as 120 min. Thus, insubsequent experiments, we used 30 to 90 min as the primingtime.
In addition, G-CSF enhanced the oxidative metabolicburst in response to opsonized and nonopsonized hyphae (byregression analysis, P < 0.015). The stimulation of PMNs byopsonized and nonopsonized hyphae was slightly slowerthan that by FMLP, with 15 min being the minimum incu-bation period showing some degree of stimulation. The peakof enhancement against hyphae appeared to be achieved byconcentrations of 1,000 to 4,000 U of G-CSF per ml (Fig. 1Cand E). In contrast, enhancement with FMLP caused adifferent pattern of response to the cytokine, with higherconcentrations of G-CSF (i.e., 10,000 U/ml) inducing aneven higher degree of oxidative burst. For example, whileconcentrations of G-CSF as low as 100 U/ml slightly in-creased the oxidative burst in response to FMLP in three offour donors, only higher concentrations (1,000 to 10,000U/ml) yielded statistically significant increases above thecontrols. Indeed, pretreatment with 10,000 U/ml had thehighest overall effect on FMLP-stimulated 02 production(an increase of 75%) compared with pretreatment of control
FIG. 1. Effects of G-CSF (A, C, E) and IFN--y (B, D, F) on thesuperoxide anion released by PMNs in response to FMLP (A, B),preopsonized hyphae of A. fiumigatus (C, D), and nonopsonizedhyphae ofA. fimigatus (E, F). Values are means ± standard errorsof means of differences in 2- production from baseline production(no cytokine treatment). The number of experiments performed witheach concentration of cytokine is shown in parenthesis. PMNs werepretreated with the concentrations of the cytokines shown at thehorizontal axis for 30 to 90 min (G-CSF) or 90 min (IFN--y) at 37°C.Baseline superoxide productions were 3.43 ± 0.60 (A; 13 experi-ments) and 3.17 ± 0.50 (B; 13 experiments) in response to FMLP,2.48 ± 0.41 (C; 6 experiments) and 1.66 ± 0.45 (D; 6 experiments) inresponse to opsonized hyphae, and 2.38 ± 0.70 (E; 6 experiments)and 1.63 ± 0.31 (F; 6 experiments) nmol/106 PMNs in response tononopsonized hyphae, respectively. *, differences from buffer-pretreated PMNs, P < 0.05.
PMNs with buffer (P < 0.001). The enhancement in responseto both opsonized and nonopsonized hyphae was smaller(peak increases of 37% at 4,000 U/ml and 24% at 1,000 U/ml,respectively).
Effects ofIFN-y on the oxidative metabolic burst. As shownin Fig. 1B, IFN-y significantly enhanced the production of02 in response to FMLP within a wide range of concentra-tions (100 to 5,000 U/ml) (by regression analysis, P < 0.006).In this series of experiments, the baseline superoxide pro-ductions, derived from 6 to 13 experiments, were 3.17 +0.50, 1.66 + 0.45, and 1.63 ± 0.31 nmol/106 PMNs inresponse to FMLP, opsonized hyphae, and nonopsonizedhyphae, respectively. The differences between the baselinevalues given above and those from experiments with G-CSFare not statistically significant. The concentration of IFN-ythat induced the highest degree of overall enhancement ofFMLP-stimulated 02- production appeared to be 5,000U/ml. The optimal time of pretreatment of PMNs was 90min, and no further enhancement was noted with longerpretreatment times. The greatest enhancement of FMLP-stimulated 02- production observed (90 min of pretreatmentwith 5,000 U/ml) was 52% above the 02- production ofcontrol cells (P = 0.007).
0.0
I.
-0.5
VOL. 61, 1993
1188 ROILIDES ET AL.
IFN--y also enhanced the oxidative metabolic burst inresponse to opsonized A. fumigatus hyphae in a patternsimilar to that of G-CSF (Fig. 1D). While the slope ofenhancement of 02 production in response to FMLP con-tinued to rise with concentrations higher than 1,000 U/ml,the curve Of 02 production in response to either serum-preopsonized or nonopsonized hyphae reached a peak in therange of concentrations from 100 to 1,000 U/ml. In contrastto G-CSF, however, the increase Of 02 production inresponse to hyphae was larger than that in response toFMLP. For example, a 71% peak increase over that withcontrol PMNs was achieved in response to opsonizedhyphae after pretreatment of the cells with 1,000 U of IFN--yper ml. The peak increase Of 02 production in response tononopsonized hyphae achieved at 100 U/ml was 96% abovethe control.
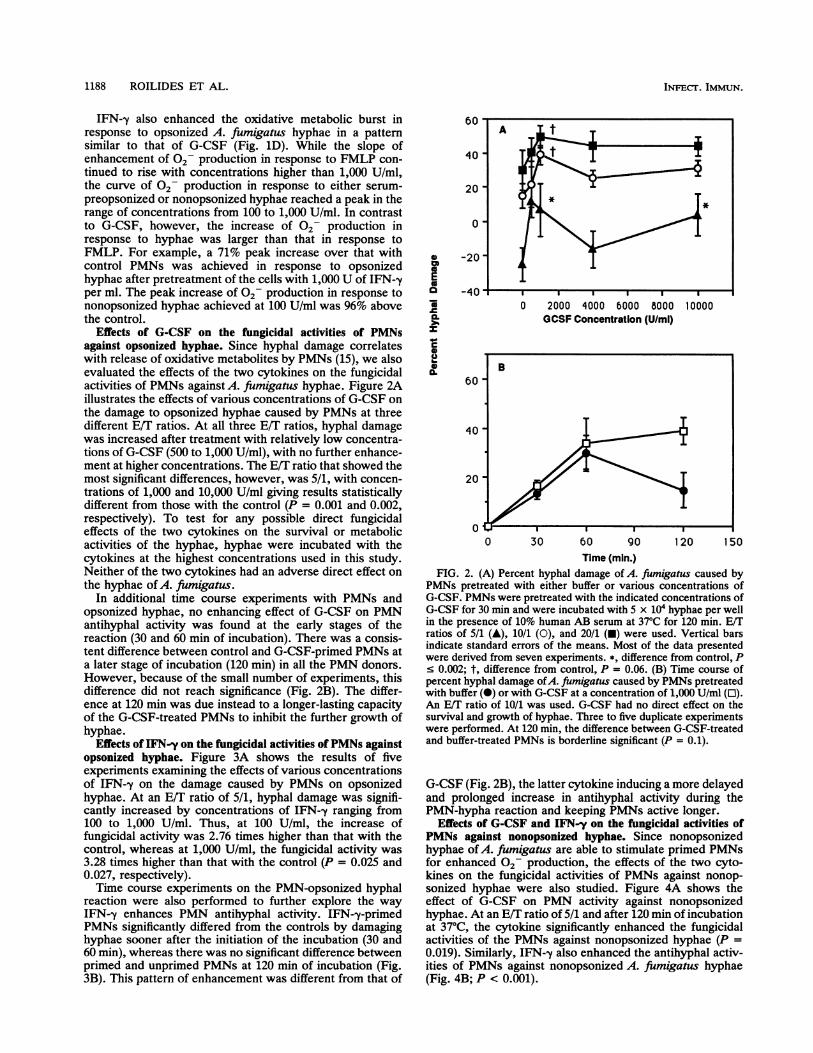
Effects of G-CSF on the fungicidal activities of PMNsagainst opsonized hyphae. Since hyphal damage correlateswith release of oxidative metabolites by PMNs (15), we alsoevaluated the effects of the two cytokines on the fungicidalactivities of PMNs against A. fumigatus hyphae. Figure 2Aillustrates the effects of various concentrations of G-CSF onthe damage to opsonized hyphae caused by PMNs at threedifferent E/T ratios. At all three E/T ratios, hyphal damagewas increased after treatment with relatively low concentra-tions of G-CSF (500 to 1,000 U/ml), with no further enhance-ment at higher concentrations. The E/T ratio that showed themost significant differences, however, was 5/1, with concen-trations of 1,000 and 10,000 U/ml giving results statisticallydifferent from those with the control (P = 0.001 and 0.002,respectively). To test for any possible direct fungicidaleffects of the two cytokines on the survival or metabolicactivities of the hyphae, hyphae were incubated with thecytokines at the highest concentrations used in this study.Neither of the two cytokines had an adverse direct effect onthe hyphae of A. fumigatus.
In additional time course experiments with PMNs andopsonized hyphae, no enhancing effect of G-CSF on PMNantihyphal activity was found at the early stages of thereaction (30 and 60 min of incubation). There was a consis-tent difference between control and G-CSF-primed PMNs ata later stage of incubation (120 min) in all the PMN donors.However, because of the small number of experiments, thisdifference did not reach significance (Fig. 2B). The differ-ence at 120 min was due instead to a longer-lasting capacityof the G-CSF-treated PMNs to inhibit the further growth ofhyphae.
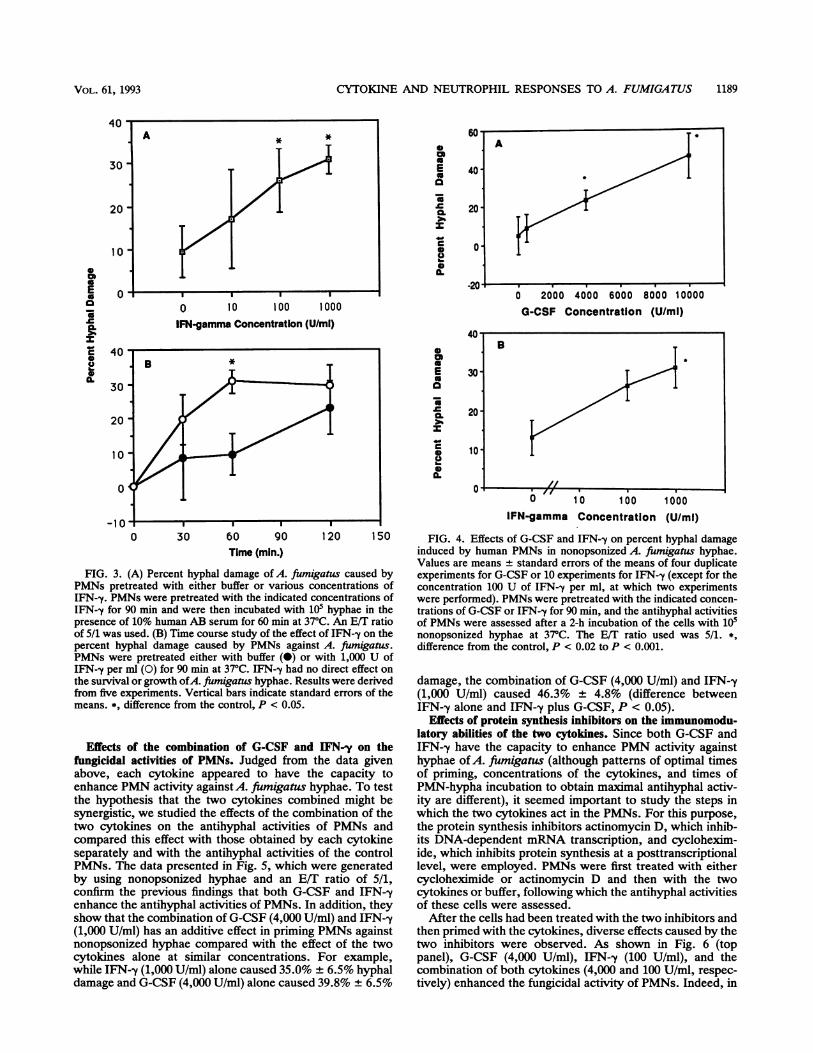
Effects of IFN-y on the fungicidal activities of PMNs againstopsonized hyphae. Figure 3A shows the results of fiveexperiments examining the effects of various concentrationsof IFN--y on the damage caused by PMNs on opsonizedhyphae. At an E/T ratio of 5/1, hyphal damage was signifi-cantly increased by concentrations of IFN--y ranging from100 to 1,000 U/ml. Thus, at 100 U/ml, the increase offungicidal activity was 2.76 times higher than that with thecontrol, whereas at 1,000 U/ml, the fungicidal activity was3.28 times higher than that with the control (P = 0.025 and0.027, respectively).Time course experiments on the PMN-opsonized hyphal
reaction were also performed to further explore the wayIFN--y enhances PMN antihyphal activity. IFN--y-primedPMNs significantly differed from the controls by damaginghyphae sooner after the initiation of the incubation (30 and60 min), whereas there was no significant difference betweenprimed and unprimed PMNs at 120 min of incubation (Fig.3B). This pattern of enhancement was different from that of
40 -
20 -
0
0
0
Ea
a.
CL
0
0.
-20
-40 -
40
20
0
0 2000 4000 6000 8000 10000GCSF Concentration (U/ml)
0 30 60 90Time (min.)
120 150
FIG. 2. (A) Percent hyphal damage of A. fiumigatus caused byPMNs pretreated with either buffer or various concentrations ofG-CSF. PMNs were pretreated with the indicated concentrations ofG-CSF for 30 min and were incubated with 5 x 104 hyphae per wellin the presence of 10% human AB serum at 37°C for 120 min. E/Tratios of 5/1 (A), 10/1 (0), and 20/1 (A) were used. Vertical barsindicate standard errors of the means. Most of the data presentedwere derived from seven experiments. *, difference from control, Pc 0.002; t, difference from control, P = 0.06. (B) Time course ofpercent hyphal damage of A. fumigatus caused by PMNs pretreatedwith buffer (0) or with G-CSF at a concentration of 1,000 U/ml (E).An E/T ratio of 10/1 was used. G-CSF had no direct effect on thesurvival and growth of hyphae. Three to five duplicate experimentswere performed. At 120 min, the difference between G-CSF-treatedand buffer-treated PMNs is borderline significant (P = 0.1).
G-CSF (Fig. 2B), the latter cytokine inducing a more delayedand prolonged increase in antihyphal activity during thePMN-hypha reaction and keeping PMNs active longer.
Effects of G-CSF and IFN-y on the fungicidal activities ofPMNs against nonopsonized hyphae. Since nonopsonizedhyphae of A. fumigatus are able to stimulate primed PMNsfor enhanced 02 production, the effects of the two cyto-kines on the fungicidal activities of PMNs against nonop-sonized hyphae were also studied. Figure 4A shows theeffect of G-CSF on PMN activity against nonopsonizedhyphae. At an EST ratio of 5/1 and after 120 min of incubationat 37°C, the cytokine significantly enhanced the fungicidalactivities of the PMNs against nonopsonized hyphae (P =
0.019). Similarly, IFN-y also enhanced the antihyphal activ-ities of PMNs against nonopsonized A. fumigatus hyphae(Fig. 4B; P < 0.001).
A
INFECT. IMMUN.
CYTOKINE AND NEUTROPHIL RESPONSES TO A. FUMIGATUS 1189
0
Ea7i
CL:0
0I.,
0 10 100 1000IFN-gamma Concentration (U/ml)
0S
aIs
CL
0
0a.
40 - I
20-
0
-20 I I
0 2000 4000 6000 8000 10000G-CSF Concentration (U/ml)
40-B
30X
20-
10-
04i0 10 100
IFN-gamma Concentration1000(U/ml)
0 30 60 90 120 150 FIG. 4. Effects of G-CSF and IFN-y on percent hyphal damage
Time (mln.) induced by human PMNs in nonopsonized A. fumigatus hyphae.Values are means + standard errors of the means of four duplicate
FIG. 3. (A) Percent hyphal damage of A. ftumigatus caused by experiments for G-CSF or 10 experiments for IFN-y (except for thePMNs pretreated with either buffer or various concentrations of concentration 100 U of IFN-y per ml, at which two experimentsIFN-y. PMNs were pretreated with the indicated concentrations of were performed). PMNs were pretreated with the indicated concen-
IFN-.y for 90 min and were then incubated with 105 hyphae in the trations of G-CSF or IFN--y for 90 min, and the antihyphal activitiespresence of 10% human AB serum for 60 min at 37°C. An E/T ratio of PMNs were assessed after a 2-h incubation of the cells with 105of 5/1 was used. (B) Time course study of the effect of IFN-y on the nonopsonized hyphae at 37C. The E/T ratio used was 5/1. *,
percent hyphal damage caused by PMNs against A. fumigatus. difference from the control, P < 0.02 to P < 0.001.PMNs were pretreated either with buffer (@) or with 1,000 U ofIFN--y per ml (0) for 90 min at 37°C. IFN--y had no direct effect on
the survival or growth ofA. fuimigatus hyphae. Results were derived damage, the combination of G-CSF (4,000 U/ml) and IFN-,yfrom five experiments. Vertical bars indicate standard errors of the (1,000 U/ml) caused 46.3% 4.8% (difference betweenmeans. *, difference from the control, P < 0.05. IFN-y alone and IFN-,y plus G-CSF, P < 0.05).
Effects of protein synthesis inhibitors on the immunomodu-latory abilities of the two cytokines. Since both G-CSF and
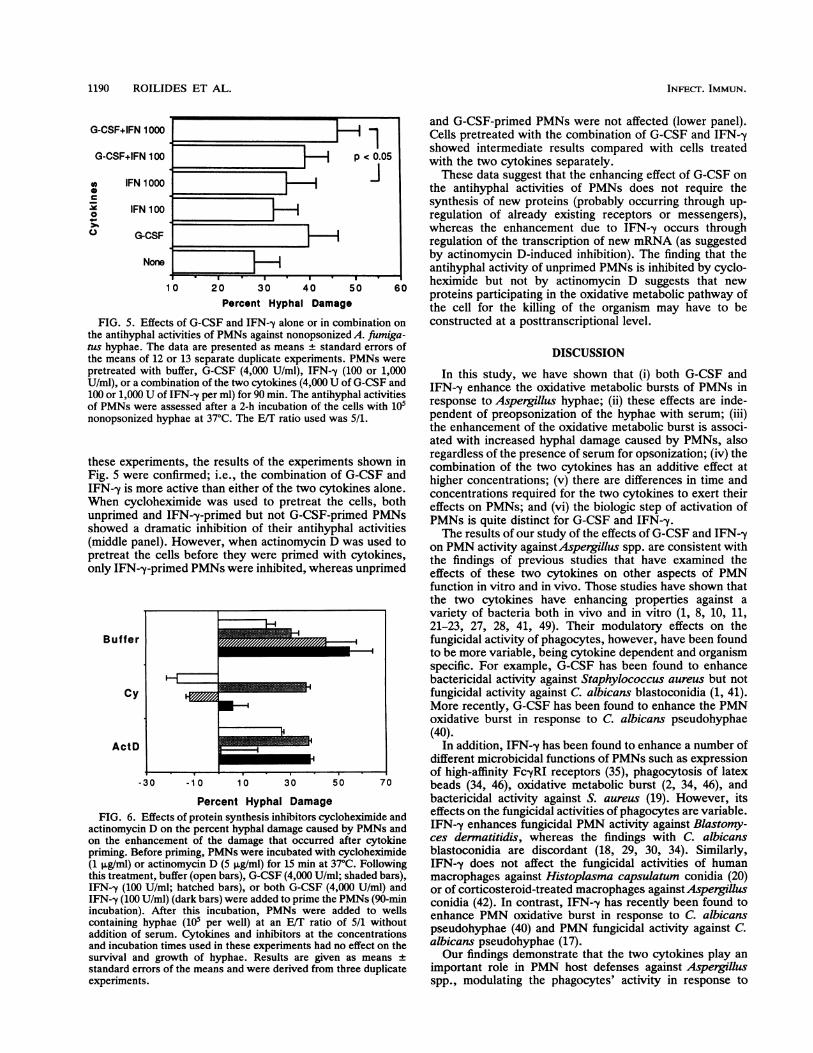
Effects of the combination of G-CSF and IFN-y on the IFN-,y have the capacity to enhance PMN activity againstfungicidal activities of PMNs. Judged from the data given hyphae of A. fumigatus (although patterns of optimal timesabove, each cytokine appeared to have the capacity to of priming, concentrations of the cytokines, and times ofenhance PMN activity against A. fiumigatus hyphae. To test PMN-hypha incubation to obtain maximal antihyphal activ-the hypothesis that the two cytokines combined might be ity are different), it seemed important to study the steps insynergistic, we studied the effects of the combination of the which the two cytokines act in the PMNs. For this purpose,
two cytokines on the antihyphal activities of PMNs and the protein synthesis inhibitors actinomycin D, which inhib-compared this effect with those obtained by each cytokine its DNA-dependent mRNA transcription, and cyclohexim-separately and with the antihyphal activities of the control ide, which inhibits protein synthesis at a posttranscriptionalPMNs. The data presented in Fig. 5, which were generated level, were employed. PMNs were first treated with eitherby using nonopsonized hyphae and an E/T ratio of 5/1, cycloheximide or actinomycin D and then with the twoconfirm the previous findings that both G-CSF and IFN-y cytokines or buffer, following which the antihyphal activitiesenhance the antihyphal activities of PMNs. In addition, they of these cells were assessed.show that the combination of G-CSF (4,000 U/ml) and IFN-y After the cells had been treated with the two inhibitors and(1,000 U/ml) has an additive effect in priming PMNs against then primed with the cytokines, diverse effects caused by thenonopsonized hyphae compared with the effect of the two two inhibitors were observed. As shown in Fig. 6 (topcytokines alone at similar concentrations. For example, panel), G-CSF (4,000 U/ml), IFN--y (100 U/ml), and thewhile IFN--y (1,000 U/ml) alone caused 35.0% + 6.5% hyphal combination of both cytokines (4,000 and 100 U/ml, respec-
damage and G-CSF (4,000 U/ml) alone caused 39.8% _ 6.5% tively) enhanced the fungicidal activity of PMNs. Indeed, in
40 '
30
20
10
A
0
00IaEaa
zi.C
0a.
40
30
20
10
-1
VOL. 61, 1993
1190 ROILIDES ET AL.
G-CSF+IFN 1 00(
G-CSF+IFN 1 OC
0
0
0
IFN 1 OOC
IFN 1 OC
G-CSF
None
FIG. 5. Effec
and G-CSF-primed PMNs were not affected (lower panel).DL1+I~Cells pretreated with the combination of G-CSF and IFN-y. showed intermediate results compared with cells treated
P < 0.05 with the two cytokines separately.II|_ These data suggest that the enhancing effect of G-CSF on
the antihyphal activities of PMNs does not require thesynthesis of new proteins (probably occurring through up-
I__________lregulation of already existing receptors or messengers),whereas the enhancement due to IFN--y occurs through
1111hZ-I regulation of the transcription of new mRNA (as suggestedby actinomycin D-induced inhibition). The finding that the
______ r________._____._____.____ antihyphal activity of unprimed PMNs is inhibited by cyclo-2~ *1o 3 o 4 o s o 6 heximide but not by actinomycin D suggests that new1 0 20 30 40 50 60 proteins participating in the oxidative metabolic pathway ofPercent Hyphal Damage the cell for the killing of the organism may have to be
cts of G-CSF and IFN-y alone or in combination on constructed at a posttranscriptional level.the antihyphal activities of PMNs against nonopsonized A. fumiga-tus hyphae. The data are presented as means ± standard errors ofthe means of 12 or 13 separate duplicate experiments. PMNs were
pretreatedU/ml), or a100 or 1,00of PMNs v
nonopsoni
these expFig. 5 weIFN--y isWhen cyunprimedshowed a(middle ppretreat tonly IFN
Buffer
Cy
ActC
FIG. 6.actinomycon the en]priming. B(1 pLg/ml) (this treatmIFN-y (10IFN-y (10(incubationcontainingaddition oand incubasurvival astandard e
experimen
DISCUSSIONwith bufter, G-CSF (4,00( U/ml), I-N--y (1W0 or 1,WU In this study, we have shown that (i) both G-CSF andcombination of the two cytokines (4,000 U of G-CSF and IFN-y enhance the oxidative metabolic bursts of PMNs in
)O U of IFN-,y per ml) for 90 mm. The antihyphal activitieswere assessed after a 2-h incubation of the cells with 105 response to Aspergillus hyphae; (ii) these effects are inde-zed hyphae at 37°C. The E/T ratio used was 5/1. pendent of preopsonization of the hyphae with serum; (iii)
the enhancement of the oxidative metabolic burst is associ-ated with increased hyphal damage caused by PMNs, also
einttshown in regardless of the presence of serum for opsonization; (iv) theIer firmed, theresults of the expermnS combination of the two cytokines has an additive effect atre confirmed ia.e. the combination of G-CSF and higher concentrations; (v) there are differences in time andmclorexactie than useithe ofetreathetwo celtes,alon concentrations required for the two cytokines to exert theirclohexImide wasr usedbutnopretrt theSF cells,dbot effects on PMNs; and (vi) the biologic step of activation ofand IFN-yo-primed but not G-CSF-primed PMNs PMNs is quite distinct for G-CSF and IFN-^y.i dramatic inhibition of their antihyphal activities The results of our study of the effects of G-CSF andIFN-yy)anel). However, when actinomycin D was used to ohM acti againstAdyeofithe e spp are cns wIththe cells before they were primed with cytokines,
o PM atvit agis Aseglu sp''ecnssetwthecprimelldefr thywere sprimed the findings of previous studies that have examined the
--primedPMswrihbtd,weffects of these two cytokines on other aspects of PMNfunction in vitro and in vivo. Those studies have shown thatthe two cytokines have enhancing properties against avariety of bacteria both in vivo and in vitro (1, 8, 10, 11,21-23, 27, 28, 41, 49). Their modulatory effects on the
r M//////////////////////////////// fungicidal activity of phagocytes, however, have been foundto be more variable, being cytokine dependent and organismspecific. For example, G-CSF has been found to enhancebactericidal activity against Staphylococcus aureus but notfungicidal activity against C. albicans blastoconidia (1, 41).More recently, G-CSF has been found to enhance the PMNoxidative burst in response to C. albicans pseudohyphae(40).
In addition, IFN--y has been found to enhance a number ofdifferent microbicidal functions of PMNs such as expression
10. 0. ,of high-affinity Fc-yRI receptors (35), phagocytosis of latex30 -10 10 30 50 70 beads (34, 46), oxidative metabolic burst (2, 34, 46), and
Percent Hyphal Damage bactericidal activity against S. aureus (19). However, itsEffects of protein synthesis inhibitors cycloheximide and effects on the fungicidal activities of phagocytes are variable.in D on the percent hyphal damage caused by PMNs and IFN--y enhances fungicidal PMN activity against Blastomy-hancement of the damage that occurred after cytokine ces dermatitidis, whereas the findings with C. albicanslefore priming, PMNs were incubated with cycloheximide blastoconidia are discordant (18, 29, 30, 34). Similarly,Dr actinomycin D (5 pLg/ml) for 15 min at 37°C. Following IFN--y does not affect the fungicidal activities of humanient, buffer (open bars), G-CSF (4,000 U/ml; shaded bars), macrophages against Histoplasma capsulatum conidia (20)0 U/ml; hatched bars), or both G-CSF (4,000 U/ml) and or of corticosteroid-treated macrophages againstAspergillusD U/ml) (dark bars) were added to prime the PMNs (90-min conidia (42). In contrast, IFN--y has recently been found toL). After this incubation, PMNs were added to wells * * '
hyha(10 pe wel ata ai o / ihu enhance PMN oxidative burst In response to C. albicansrhyphae (105 per well) at an E/T ratio of 511 withoutf serum. Cytokines and inhibitors at the concentrations pseudohyphae (40) and PMN fungicidal activity against C.ation times used in these experiments had no effect on the albicans pseudohyphae (17).nd growth of hyphae. Results are given as means ± Our findings demonstrate that the two cytokines play anrrors of the means and were derived from three duplicate important role in PMN host defenses against AspergillusIts. spp., modulating the phagocytes' activity in response to
INFECT. IMMUN.

CYTOKINE AND NEUTROPHIL RESPONSES TO A. FUMIGATUS 1191
challenge with the organism. Using two standard assays, the02 release assay and the MIT assay of hyphal viability, wewere able to show that the two cytokines enhance both theoxidative burst of PMNs in response to Aspergillus hyphaeand the fungicidal activity of the same cells against hyphae.Despite the interexperimental variation of the baseline PMNactivity, we were able to demonstrate significant increases offungicidal activity within the same set of experiments andPMN donors due to cytokine pretreatment of PMNs.Our results with IFN--y are paralleled by a recently re-
ported ex vivo study of patients with chronic granulomatousdisease (known to have defective PMN function) receivingIFN-y who showed an increase in antihyphal activity (37). Inthis study, we correlated enhancement of fungicidal activitywith an increase of oxidative burst in response to the sameorganism. Moreover, we demonstrated that G-CSF pos-sesses a similar property that enhances fungicidal activityand that the two cytokines are synergistic. Potential effectsof the two cytokines on nonoxidative fungicidal mecha-nisms, however, have not been investigated here and areworth studying. Nonoxidative mechanisms do contribute tothe fungicidal activities of PMNs. Up to this time, however,to our knowledge, there have been no studies investigatingthe effects of cytokines on specific components of thenonoxidative microbicidal armamentarium of the PMN.Our data suggest that PMN activity against Aspergillus
hyphae (both oxidative metabolic burst and hyphal damage)can be enhanced by lower concentrations of G-CSF (500 to1,000 U/ml) than are required for other PMN activities suchas oxidative metabolic burst in response to FMLP andbactericidal activity (the peak occurring around 4,000 U/ml)(41) and PMN activity against C. albicans blastoconidia(significant enhancement not obtained by concentrations upto 10,000 U/ml). The basis for these organism-specific differ-ences remains to be elucidated.Opsonized A. fumigatus hyphae stimulate the phagocytic
cells by a mechanism different from that of the solublestimulus FMLP. Thus, while FMLP stimulates the cellmembrane by interaction with specific receptors, opsonizedhyphae stimulate the cell by attachment and interaction ofopsonins with Fc and C' receptors. Our findings suggest thatserum-opsonized A. fumigatus hyphae stimulate the oxida-tive burst of normal PMNs but with a somewhat lowerproduction of 02- than occurs after stimulation with FMLP.The finding of enhanced PMN stimulation against opsonizedhyphae by the two cytokines is consistent with previousreports that G-CSF and IFN--y modulate Fc-yRIII receptorson the PMN membrane and that IFN--y additionally in-creases the expression of Fc-yRI receptors (6, 7). The in-creased expression of the receptors may lead to a moreefficient interaction of the cell with opsonized organisms andconsequently to enhanced oxidative metabolic burst in re-sponse to these organisms.Of note, the two cytokines were able to enhance the
oxidative burst and hyphal damage even in response tononopsonized particulate stimuli. To our knowledge, this isa novel finding; that is, cytokines can enhance PMN stimu-lation in response to organisms through non-opsonin-specificreceptors and intracellular pathways. Such receptors havebeen found in the case of C. albicans pseudohyphae (man-nose receptors) (13). Similar receptors may be involved inthe case of Aspergillus spp., and G-CSF and IFN--y mayupregulate their numbers and/or affinities in a manner anal-ogous to the upregulation of FMLP receptors. However, toour knowledge, PMN receptors for cell wall surface compo-nents of Aspergillus hyphae have not yet been described.
Another possibility is that the two cytokines modulate theearly responses that are elicited after initial stimulation ofthe cellular membrane receptors, although it has recentlybeen reported that only minor enhancement by IFN-y occursat the early steps of the oxidative burst in response to C.albicans hyphae (17). Nevertheless, this independence fromserum is biologically intriguing and may be clinically usefulin the activation of PMNs againstAspergillus hyphae at siteswith limited supply of opsonins.The finding of differential effects of protein synthesis
inhibitors on the augmentation of antihyphal activity ofPMNs by the two cytokines helps in understanding theintracellular pathways involved in priming the PMN byG-CSF and IFN--y. Very little is known about the signaltransduction of the two cytokines. G-CSF appeared not torequire synthesis of new proteins (probably acting throughupregulation of already existing receptors and protein kinaseC), IFN-y appeared to prime the microbicidal mechanisms ofPMNs through regulation of the transcription of new mRNA.In agreement with our results, the effect of IFN--y on thefungicidal activity of murine macrophages has recently beenfound to be inhibited by cycloheximide (5).The maximal enhancement in response to G-CSF is
achieved by concentrations ranging from 500 to 4,000 U/ml.These concentrations are well within the range of levels inserum achieved in patients receiving the recombinant cyto-kine intravenously or subcutaneously and correlate withPMN response (3, 31, 45). In contrast, although the concen-trations of IFN--y that were shown in this study to have aneffect (100 to 1,000 U/ml) are achievable clinically, theredoes not appear to be a correlation between levels of IFN-yin serum and degree of in vivo phagocytic activation (33).The findings of this study may have clinical implications
and suggest that PMNs can be primed by G-CSF and IFN--yto possess enhanced activity against Aspergillus spp., aproperty that may prevent invasive aspergillosis in immuno-compromised patients. Whether the results presented in thisreport correlate with in vivo activation is not known atpresent. Nevertheless, since PMNs are the main effectorcells against hyphal forms of important opportunistic fungisuch as Aspergillus spp., these in vitro effects may beassociated with increased in vivo stimulation of PMNs,increased subsequent killing of the hyphae, and clearingfrom the infected tissues. In support of our data, adminis-tration of G-CSF was recently reported to prolong survivalin normal and immunocompromised mice infected with A.fumigatus (36). Thus, they may be particularly important inamplifying host defenses against invasive aspergillosis inimmunocompromised hosts. These cytokines alone or com-bined with antifungal agents may also have an effect on thetreatment of established invasive disease. In the future,G-CSF and IFN--y may offer an adjunct to antifungal chemo-therapy in prevention and treatment of fungal infections inimmunocompromised patients, and appropriate studies ap-pear worthy of investigation.
ACKNOWLEDGMENT
We thank Alison Freifeld for assistance in obtaining IFN--y for theexperiments described in this study.
REFERENCES1. Arenberg, D., E. Navarro, E. Roilides, V. Thomas, J. Peter,
J. W. Lee, P. S. Francis, P. A. Pizzo, and T. J. Walsh. 1990.Effects of granulocyte colony-stimulating factor in experimentalhepatosplenic candidiasis in granulocytopenic rabbits, abstr.F90, p. 423. Abstr. 90th Annu. Meet. Am. Soc. Microbiol. 1990.
VOL. 61, 1993

1192 ROILIDES ET AL.
2. Bielefeldt Ohmann, H., and L. A. Babiuk. 1986. Alteration ofsome leukocyte functions following in vivo and in vitro expo-sure to recombinant bovine alpha- and gamma-interferon. J.Interferon Res. 6:123-136.
3. Bonilla, M. A., A. P. Gillio, M. Rugeiro, N. A. Kernan, J. A.Brochstein, M. Abboud, L. Fumagalli, M. Vincent, J. L. Gabri-love, K. Welte, L. M. Souza, and R. J. O'Reilly. 1989. Effects ofrecombinant human granulocyte colony-stimulating factor onneutropenia in patients with congenital agranulocytosis. N.Engl. J. Med. 320:1574-1580.
4. Boyum, A. 1968. Isolation of mononuclear cells and granulo-cytes from human blood: isolation of mononuclear cells by onecentrifugation and of granulocytes by combining centrifugationand sedimentation at 1 g. Scand. J. Clin. Lab. Invest. 21(Suppl.97):77-89.
5. Brummer, E., L. H. Hanson, and D. A. Stevens. 1991. Kineticsand requirements for activation of macrophages for fungicidalactivity: effect of protein synthesis inhibitors and immunosup-pressants on activation and fungicidal mechanism. Cell. Immu-nol. 132:236-245.
6. Buckle, A. M., and N. Hogg. 1989. The effect of IFN--y andcolony-stimulating factor on the expression of neutrophil cellmembrane receptors. J. Immunol. 143:2295-2301.
7. Buckle, A. M., Y. Jayaram, and N. Hogg. 1990. Colony-stimulating factors and interferon-gamma differentially affectcell surface molecules shared by monocytes and neutrophils.Clin. Exp. Immunol. 81:339-345.
8. Cairo, M. S., D. Mauss, S. Kommareddy, K. Norris, C. Ven, andH. Modanlou. 1990. Prophylactic or simultaneous administra-tion of recombinant human granulocyte colony-stimulating fac-tor in the treatment of group B streptococcal sepsis in neonatalrats. Pediatr. Res. 27:612-616.
9. Cassatella, M. A., R. Cappelli, V. Della Bianca, M. Grzeskowiak,S. Dusi, and G. Berton. 1988. Interferon-gamma activates hu-man neutrophil oxygen metabolism and exocytosis. Immunol-ogy 63:499-506.
10. Cohen, A. M., D. K. Hines, E. S. Korach, and B. J. Ratzkin.1988. In vivo activation of neutrophil function in hamsters byrecombinant human granulocyte colony-stimulating factor. In-fect. Immun. 56:2861-2865.
11. Cohen, A. M., K. M. Zsebo, H. Inoue, D. K. Hines, T. C. Boone,V. R. Chazin, L. Tsai, T. Ritch, L. M. Souza, E. S. Korach, andB. J. Ratzkin. 1987. In vivo stimulation of granulopoiesis byrecombinant human granulocyte colony-stimulating factor.Proc. Natl. Acad. Sci. USA 84:2484-2488.
12. Cohen, M. S., R. E. Isturiz, H. L. Malech, R. K. Root, C. M.Wilfert, L. Gutman, and R. H. Buckley. 1981. Fungal infectionin chronic granulomatous disease. The importance of the phago-cyte in defense against fungi. Am. J. Med. 71:59-66.
13. Danley, D. L., and A. E. Hilger. 1981. Stimulation of oxidativemetabolism in murine polymorphonuclear leukocytes by unop-sonized fungal cells: evidence for a mannose-specific mecha-nism. J. Immunol. 127:551-556.
14. Denning, D. W., S. E. Follansbee, M. Scolaro, S. Norris, H.Edelstein, and D. A. Stevens. 1991. Pulmonary aspergillosis inthe acquired immunodeficiency syndrome. N. Engl. J. Med.324:654-662.
15. Diamond, R. D., and R. Krzesicki. 1978. Mechanisms of attach-ment of neutrophils to C. albicans pseudohyphae in the absenceof serum, and of subsequent damage to pseudohyphae bymicrobicidal processes of neutrophils in vitro. J. Clin. Invest.61:360-369.
16. Diamond, R. D., R. Krzesicki, and W. Jao. 1978. Damage topseudohyphal form of C. albicans by neutrophils in the absenceof serum in vitro. J. Clin. Invest. 61:349-359.
17. Diamond, R. D., C. A. Lyman, and D. R. Wysong. 1991.Disparate effects of interferon-gamma and tumor necrosis fac-tor-alpha on early neutrophil respiratory burst and fungicidalresponses to Candida albicans hyphae in vitro. J. Clin. Invest.87:711-720.
18. Djeu, J. Y., D. K. Blanchard, D. Halkias, and H. Friedman.1986. Growth inhibition of Candida albicans by human polymor-phonuclear neutrophils: activation by interferon-gamma and
tumor necrosis factor. J. Immunol. 137:2980-2984.19. Edwards, S. W., J. E. Say, and V. Hughes. 1988. Gamma
interferon enhances the killing of Staphylococcus aureus byhuman neutrophils. J. Gen. Microbiol. 134:37-42.
20. Fleischmann, J., B. Wu-Hsieh, and D. H. Howard. 1990. Theintracellular fate of Histoplasma capsulatum in human macro-phages is unaffected by recombinant human interferon--y. J.Infect. Dis. 161:143-145.
21. Gillio, A. P., M. A. Bonilla, G. K. Potter, J. L. Gabrilove, 0.Reilly, L. M. Souza, and K. Welte. 1987. Effects of recombinanthuman granulocyte colony-stimulating factor on hematopoieticreconstitution after autologous bone marrow transplantation inprimates. Transplant. Proc. 19:153-156.
22. Iguchi, K., S. Inoue, and A. Kumar. 1991. Effect of recombinanthuman granulocyte colony-stimulating factor administration innormal and experimentally infected newborn rats. Exp. Hema-tol. 19:352-358.
23. Kitagawa, S., A. You, L. M. Souza, M. Saito, Y. Miura, and F.Takaku. 1987. Recombinant human granulocyte colony-stimu-lating factor enhances superoxide release in human granulo-cytes stimulated by chemotactic peptide. Biochem. Biophys.Res. Commun. 144:1143-1146.
24. Levitz, S., and R. D. Diamond. 1984. Killing of Aspergillusfumigatus spores and Candida albicans yeast phase by theiron-hydrogen peroxide-iodide cytotoxic system: comparisonwith the myeloperoxidase-hydrogen peroxide-halide system.Infect. Immun. 43:1100-1102.
25. Levitz, S. M., and R. D. Diamond. 1985. A rapid colorimetricassay of fungal viability with the tetrazolium salt M1T. J. Infect.Dis. 152:938-945.
26. Lyman, C. A., E. R. Simons, D. A. Melnick, and R. D. Diamond.1987. Unopsonized Candida albicans hyphae stimulate a neu-trophil respiratory burst and a cytosolic calcium flux withoutmembrane depolarization. J. Infect. Dis. 156:770-776.
27. Matsumoto, M., S. Matsubara, T. Matsuno, M. Tamura, K.Hattori, H. Nomura, M. Ono, and T. Yokota. 1987. Protectiveeffect of human granulocyte colony-stimulating factor on micro-bial infection in neutropenic mice. Infect. Immun. 55:2715-2720.
28. Mizushima, Y., T. Morikage, T. Kuwahara, and S. Yano. 1990.Effects of granulocyte colony-stimulating factor on hematopoi-etic injury induced by anticancer drugs in mice. J. Biol. Re-sponse Modif. 9:576-583.
29. Morrison, C. J., E. Brummer, R. A. Isenberg, and D. A. Stevens.1987. Activation of murine polymorphonuclear neutrophils forfungicidal activity by recombinant gamma interferon. J. Leuko-cyte Biol. 41:434 440.
30. Morrison, C. J., E. Brummer, and D. A. Stevens. 1989. In vivoactivation of peripheral blood polymorphonuclear neutrophilsby gamma interferon results in enhanced fungal killing. Infect.Immun. 57:2953-2958.
31. Morstyn, G., G. J. Lieschke, W. Sheridan, J. Layton, J. Cebon,and R. M. Fox. 1989. Clinical experience with recombinanthuman granulocyte colony-stimulating factor. Semin. Hematol.26(Suppl. 2):9-13.
32. Mosmann, T. 1983. Rapid colorimetric assay for cellular growthand survival: application to proliferation and cytotoxicity as-says. J. Immunol. Methods 65:55-63.
33. Murray, H. W. 1988. Interferon--y, the activated macrophage,and host defense against microbial challenge. Ann. Intern. Med.108:595-608.
34. Perussia, B., M. Kobayashi, M. E. Rossi, I. Anegon, and G.Trinchieri. 1987. Immune interferon enhances properties ofhuman granulocytes: role of Fc receptors and effect of lympho-toxin, tumor necrosis factor, and granulocyte-macrophage col-ony-stimulating factor. J. Immunol. 138:765-774.
35. Petroni, K. C., L. Shen, and P. M. Guyre. 1988. Modulation ofhuman polymorphonuclear leukocyte IgG Fc receptors and Fcreceptor-mediated functions by IFN-gamma and glucocorti-coids. J. Immunol. 140:3467-3472.
36. Polak-Wyss, A. 1991. Protective effect of human granulocytecolony-stimulating factor (hG-CSF) on Cryptococcus and As-pergillus infections in normal and immunosuppressed mice.
INFECT. IMMUN.

CYTOKINE AND NEUTROPHIL RESPONSES TO A. FUMIGATUS 1193
Mycoses 34:205-215.37. Rex, J. H., J. E. Bennett, J. I. Gallin, H. L. Malech, E. S.
Decarlo, and D. A. Melnick. 1991. In vivo interferon-y therapyaugments the in vitro ability of chronic granulomatous diseaseneutrophils to damage Aspergillus hyphae. J. Infect. Dis. 163:849-852.
38. Rex, J. H., J. E. Bennett, J. I. Gallin, H. L. Malech, and D. A.Melnick. 1990. Normal and deficient neutrophils can cooperateto damage Aspergillus fumigatus hyphae. J. Infect. Dis. 162:523-528.
39. Roilides, E., S. Mertins, J. Eddy, T. J. Walsh, P. A. Pizzo, andM. Rubin. 1990. Impairment of neutrophil chemotactic andbactericidal function in HIV-infected children and partial rever-sal after in vitro exposure to granulocyte-macrophage colony-stimulating factor. J. Pediatr. 117:531-540.
40. Roilides, E., K. Uhlig, D. Venzon, P. A. Pizzo, and T. J. Walsh.1992. Neutrophil oxidative burst in response to blastoconidiaand pseudohyphae of Candida albicans: augmentation by gran-ulocyte colony-stimulating factor and interferon--y. J. Infect.Dis. 166:668-673.
41. Roilides, E., T. J. Walsh, P. A. Pizzo, and M. Rubin. 1991.Granulocyte-colony stimulating factor enhances the phagocyticand bactericidal activity of normal and defective human neutro-phils. J. Infect. Dis. 163:579-583.
42. Schaffner, A. 1985. Therapeutic concentrations of glucocorti-coids suppress the antimicrobial activity of human macrophageswithout impairing their responsiveness to gamma interferon. J.Clin. Invest. 76:1755-1764.
43. Schaffner, A., H. Douglas, and A. Braude. 1982. Selectiveprotection against conidia by mononuclear and against myceliaby polymorphonuclear phagocytes in resistance to Aspergillus:observations on these two lines of defense in vivo and in vitrowith human and mouse phagocytes. J. Clin. Invest. 69:617-631.
44. Shalaby, M. R., B. B. Aggarwal, E. Rinderknecht, L. P. Svender-sky, B. S. Finkle, and M. A. Palladino. 1985. Activation ofhuman polymorphonuclear neutrophil functions by interferon-gamma and tumor necrosis factor. J. Immunol. 135:2069-2073.
45. Sheridan, W., G. Morstyn, M. Wolf, A. Dodds, J. Lusk, D.Maher, J. E. Layton, M. Green, L. M. Souza, and R. Fox. 1989.Granulocyte colony-stimulating factor and neutrophil recoveryafter high-dose chemotherapy and autologous bone marrowtransplantation. Lancet ii:891-895.
46. Steinbeck, M. J., J. A. Roth, and M. L. Kaeberle. 1986.Activation of bovine neutrophils by recombinant interferon-gamma. Cell. Immunol. 98:137-144.
47. Walsh, T. J. 1990. Invasive aspergillosis in patients with neo-plastic diseases. Semin. Respir. Infect. 5:111-122.
48. Weinberger, M., I. Elattar, D. Marshall, S. M. Steinberg, R. L.Redner, N. S. Young, and P. A. Pizzo. 1992. Patterns of infectionin patients with aplastic anemia and the emergence ofAspergil-lus as a major cause of death. Medicine (Baltimore) 71:24-43.
49. Welte, K., M. A. Bonilla, A. P. Gillio, T. C. Boone, G. K. Potter,J. L. Gabrilove, M. A. S. Moore, R. J. O'Reily, and L. M. Souza.1987. Recombinant human granulocyte colony-stimulating fac-tor. Effects on hematopoiesis in normal and cyclophosphamide-treated primates. J. Exp. Med. 165:941-948.
VOL. 61, 1993





























