Embed Size (px)
Citation preview
HSI2/VAL1 Silences AGL15 to Regulate the DevelopmentalTransition from Seed Maturation to Vegetative Growthin ArabidopsisOPEN
Naichong Chen,a,b Vijaykumar Veerappan,a,c Haggag Abdelmageed,a,d Miyoung Kang,a,b and Randy D. Allena,b,1
a Institute for Agricultural Biosciences, Oklahoma State University, Ardmore, Oklahoma 73401bDepartment of Biochemistry and Molecular Biology, Oklahoma State University, Stillwater, Oklahoma 74074cDepartment of Biology, Eastern Connecticut State University, Willimantic, Connecticut 06226dDepartment of Agricultural Botany, Faculty of Agriculture, Cairo University, Giza 12613, Egypt
ORCID IDs: 0000-0003-3982-0762 (N.C.); 0000-0002-1689-8332 (V.V.); 0000-0003-2111-2629 (R.D.A.)
Gene expression during seed development in Arabidopsis thaliana is controlled by transcription factors including LEAFYCOTYLEDON1 (LEC1) and LEC2, ABA INSENSITIVE3 (ABI3), FUSCA3 (FUS3), known as LAFL proteins, and AGAMOUS-LIKE15(AGL15). The transition from seed maturation to germination and seedling growth requires the transcriptional silencing ofthese seed maturation-specific factors leading to downregulation of structural genes including those that encode seedstorage proteins, oleosins, and dehydrins. During seed germination and vegetative growth, B3-domain protein HSI2/VAL1 isrequired for the transcriptional silencing of LAFL genes. Here, we report chromatin immunoprecipitation analysis indicatingthat HSI2/VAL1 binds to the upstream sequences of the AGL15 gene but not at LEC1, ABI3, FUS3, or LEC2 loci. Functionalanalysis indicates that the HSI2/VAL1 B3 domain interacts with two RY elements upstream of the AGL15 coding region and atleast one of them is required for HSI2/VAL1-dependent AGL15 repression. Expression analysis of the major seed maturationregulatory genes LEC1, ABI3, FUS3, and LEC2 in different genetic backgrounds demonstrates that HSI2/VAL1 is epistatic toAGL15 and represses the seed maturation regulatory program through downregulation of AGL15 by deposition of H3K27me3at this locus. This hypothesis is further supported by results that show that HSI2/VAL1 physically interacts with the PolycombRepressive Complex 2 component protein MSI1, which is also enriched at the AGL15 locus.
INTRODUCTION
Seed development in plants involves two major phases:morphogenesis and maturation. During morphogenesis, specifi-cation of various developmental domains of the embryo is followedby thedifferentiationof tissuesandorgansystems.Thesubsequentseed maturation phase is characterized by accumulation ofstorage carbohydrates, proteins, and oils followed by acquisitionofdesiccation toleranceanddormancy.Establishmentof theseedmaturation program requires at least four master regulators,collectively known as the LAFL group of transcription factors(reviewed in Jia et al., 2014). This group includes the CAAT-boxfamily protein LEAFY COTYLEDON1 (LEC1) (Lotan et al., 1998),and theB3-family proteins ABSCISICACID INSENSITIVE3 (ABI3),FUSCA3 (FUS3), and LEC2 (Luerßen et al., 1998; Stone et al.,2001; Finkelstein et al., 2008). These LAFL regulators formacomplex genetic network andact in concert to promote the seedmaturation program by activating the expression of downstreamgenes related to storage product accumulation and desiccationtolerance (Holdsworth et al., 2008; Suzuki and McCarty 2008; Jiaet al. 2013).
During embryogenesis, AGAMOUS-Like15 (AGL15), a MADSdomain transcriptional regulator, is preferentially expressed andprimarily accumulates during early stages of seed development(Heck et al., 1995; Rounsley et al., 1995). InBrassica napus, maize(Zea mays), and Arabidopsis thaliana, AGL15 transcription re-mains high during embryo morphogenesis until the seeds start todry (Perry et al., 1996). Constitutive expression of AGL15 pro-motes seed development and somatic embryogenesis in Arabi-dopsis and loss-of-function mutation in both AGL15 and theclosely related MADS domain gene AGL18 leads to decreasedsomatic embryo tissue production (Harding et al., 2003; Thakareet al., 2008). AGL15 has been found to physically bind to thechromatin of LEC2, ABI3, and FUS3 and upregulate the expres-sion of these genes during embryogenesis (Zheng et al., 2009). Inprevious reports, we showed that expression ofAGL15 is stronglyupregulated in both hsi2-2 knockout Arabidopsis plants and inhsi2-4plants that carryaC-to-Ysubstitution in theHSI2PHDmotif(Veerappan et al., 2012). This change in expression is associatedwith a dramatic loss of histone 3 lysine 27 trimethylation(H3K27me3) at the AGL15 locus (Veerappan et al., 2014).Under favorable conditions, seed storage reserves are remobi-
lized and degraded to provide energy for subsequent germinationand seedling growth. The seed maturation program is inhibited ingerminated seeds by two transcriptional repressors, HIGH-LEVELEXPRESSION OF SUGAR INDUCIBLE GENE2 (HSI2) and HSI2-LIKE1 (HSL1), to ensure the developmental transition fromseeds toseedlings (Tsukagoshi et al., 2005, 2007). HSI2 and HSL1 are alsonamed as VP1/ABI3-LIKE1 (VAL1) and VAL2, respectively (Suzuki
1 Address correspondence to [email protected] author responsible for distribution of materials integral to the findingspresented in this article in accordance with the policy described in theInstructions for Authors (www.plantcell.org) is: Randy D. Allen ([email protected]).OPENArticles can be viewed without a subscription.www.plantcell.org/cgi/doi/10.1105/tpc.17.00655
The Plant Cell, Vol. 30: 600–619, March 2018, www.plantcell.org ã 2018 ASPB.

etal., 2007).HSI2andHSL1sharehighsequencesimilarity and, likeLEC2, ABI3, and FUS3, belong to the B3 DNA binding domain-containing family of transcription factors. Upon germination, ex-pression of HSI2 and HSL1 is induced and reaches to peak at 5 dafter germination (Tsukagoshi et al., 2005, 2007). HSI2 and HSL1show partial functional redundancy. Although morphologicallynormal, hsi2 or hsl1 mutants show increased expression of seedmaturation genes during seedling growth. However, double hsi2hsl1 mutants show much higher expression of seed maturationgenes, leading to inviable seedlingswith embryonic characteristics(Suzuki etal., 2007;Tsukagoshiet al., 2007;Holdsworthetal., 2008,Suzuki and McCarty; 2008).
The B3 domain is a plant-specific DNA binding domain thatspecifically interacts with RY DNA motifs (CATGCA) in targetgenes (Suzuki et al. 1997; Reidt et al. 2000; Mönke et al. 2004;Braybrook et al. 2006; Jia et al. 2013; Qüesta et al., 2016; Yuanetal., 2016). Themajorityofgenesupregulatedbymore than4-foldin hsi2 hsl1 double mutant seedlings contain one or more RYmotifs in the upstream regionor thefirst intron (Suzuki et al., 2007).Recently, electrophoretic mobility shift and yeast-one hybridassays indicate that the HSI2-B3 domain binds to RY elements ofFLOWERING LOCUS C (FLC) and represses its expression(Qüesta et al., 2016; Yuan et al., 2016). In addition, HSI2 containsother conserved motifs, including a plant homeodomain (PHD),cysteine and tryptophan residue-containing domain (CW), and anethylene-responsive element binding factor-associated amphi-philic repression (EAR) domain. The PHD finger is a zinc fingerdomain that is reported to recognize histone 3 trimethylated atlysine 4 (H3K4me3) and di- or trimethylated lysine 27 (H3K27me2/3) (Chakravarty et al., 2009; Zeng et al., 2010; Yuan et al., 2016).Genetic disruptionof theHSI2-PHDdomain inArabidopsis resultsin decreased H3K27me2/3 and increased H3K4me3 enrichment
at certain seed-related genes, correlatingwith ectopic expressionof those genes in seedlings (Veerappan et al., 2012, 2014; Yuanet al., 2016). The CW domain also recognizes H3K4me2 andH3K4me3 in vitro (Hoppmann et al. 2011) and the HSL1-CWdomain interacts with HISTONE DEACETYLASE19 (HDA19)in vivo to downregulate a subset of seed maturation genes byalteration of histone acetylation at the target gene loci during seedgermination (Zhou et al., 2013). The EAR domain is a repressivemotif that recruits corepressors, such as SWI-independent 3 andTOPLESS that subsequently recruit a histone deacetylase com-plex (HDAC) to targetgene loci (Tsukagoshi et al., 2005;Ohtaetal.,2001; Kazan, 2006; Kagale and Rozwadowski, 2011), and dis-ruption of the EAR motif of HSI2 reduces its ability to repress theexpressionofa luciferase reporter genedrivenby thesweetpotatoSporamin A1 gene promoter in Arabidopsis protoplasts (Tsukagoshietal.,2005).However, the functionsof theseHSI2 functionaldomainsand their interactions in the repression of seed maturation genes inseedlings remain unclear.Polycomb group (PcG) proteins regulate cell fate and identity in
higher eukaryotes by chromatin-mediated gene repression. InArabidopsis, twogroupsofPcGs,PolycombRepressiveComplex1 (PRC1) and PRC2, exist in multiple variants due to gene familyexpansion and functional diversification (Derkacheva andHennig,2014). During vegetative development in Arabidopsis, PRC2consists ofCURLYLEAF (CLF) orSWINGER (SWN), EMBRYONICFLOWER2 (EMF2), FERTILIZATION INDEPENDENT ENDO-SPERM (FIE), and MULTICOPY SUPPRESSOR of IRA1 (MSI1).CLF and SWN have methyltransferase activity and specificallycatalyze trimethylation of lysine 27 of histone H3 (H3K27me3) attarget gene loci to establish a repressive chromatin state(Chanvivattana, et al., 2004; Yoshida, et al., 2001; Jiang, et al.,2008;Kinoshita, et al., 2001).MSI1 is aWD40 repeat protein that is
AGL15 Is a Regulatory Target of HSI2/VAL1 601

an indispensable component ofPRC2at all developmental stages(Köhler et al., 2003; Schönrock et al., 2006). MSI1 is required forseed development and loss of function causes female sterility andseed abortion (Hennig et al., 2003; Köhler et al., 2003). TransgenicMSI1 cosuppression lines (msi1-cs), in which MSI1 expression isreduced to 5%of that in wild-type plants, lose repression of someEMF2-target genes and show strong defects in vegetative andreproductive development leading to sterility (Hennig et al., 2003;Köhler et al., 2003). Recently, Mehdi et al. (2016) reported thatMSI1 interactswithHDA19 to formahistonedeacetylasecomplexinvolved in the suppression of ABA receptor gene expression,which is thought to attenuate ABA signaling in Arabidopsis. Thus,MSI1 may have functions outside of the classical PRC2 context.
Here, we report that two RY elements located upstream ofAGL15 are direct targets of theB3domain ofHSI2 andat least oneof these elements is required for HSI2-dependent silencing of theAGL15 promoter. Furthermore, the PRC2 protein MSI1 interactswith HSI2 and is required for full transcriptional silencing of theAGL15 promoter. We propose a regulatory model in which theLAFL genes LEC1,ABI3, FUS3, and LEC2 are positively regulatedby AGL15. HSI2-dependent silencing of AGL15 expression inArabidopsis seedlings results in downregulation of these genesand repression of the seed maturation developmental pathway.
RESULTS
HSI2 Is Enriched at the AGL15 Promoter Region
Since expressionofAGL15 is strongly upregulated andH3K27me3enrichment is reduced in hsi2 mutants (Veerappan et al., 2012,2014), we hypothesize that the AGL15 locus is a direct regulatorytarget ofHSI2. Todetermine ifHSI2 interactswithAGL15 in vivo, anepitope-tagged HSI2-rescued Arabidopsis line was generated bythe introduction ofHSI2-HA coding sequence under control of thenative HSI2 promoter (HSI2pro:HSI2-HA) into hsi2-2 knockoutplants that contain the GSTF8pro:LUC (hsi2-2LUC) reporter gene(Veerappan et al., 2012, 2014). Expression of the luciferase re-porter gene and selected seed-specific major transcriptionalregulator genes in plants of the rescued linewas reduced from thehigh levels seen in hsi2-2LUC plants to levels similar to that in wild-typeplants, indicating thatHSI2-HA is functional (Figures1A to1C).Expression of HSI2-HA in the rescued line was confirmed byimmunoblot analysis using anti-HA (Figure 1D). Chromatin im-munoprecipitation (ChIP) assays were conducted on these plantsto investigate the enrichment of HSI2 on chromatin of seedmaturation regulatory genes including LEC1, ABI3, FUS3, LEC2,andAGL15. Different primer sets were used to query promoter (P)and the coding regions (C) of candidate target genes by ChIP-qPCR. Significant HSI2 enrichment was detected on the proximalpromoter regions (P1 and P2) and 5ʹ coding area (C1) of AGL15,with strongest enrichment at P1. Significant HSI2 enrichment wasnot detected at the distal upstream region (P3) or the far down-streamcoding regionC2 (Figure 2). SignificantHSI2 accumulationwas not observed in chromatin of the promoter or coding se-quences of LEC1, ABI3, FUS3, or LEC2 loci (Figure 2). Thus,among these master transcription factor genes, only AGL15appears to be enriched for HSI2 under the test conditions used.
RY Elements Are Required for HSI2-Dependent Repressionof AGL15
Sequence analysis indicated the presence of two canonical RYelements in the region upstream of the transcriptional start site ofAGL15. This region corresponds to P1, which showed significantHSI2enrichment inChIP-qPCRassays (Figure2).Toexaminetheroleof these RY elements in AGL15 repression by HSI2, a luciferasereporter gene controlled by theAGL15promoter (AGL15pro:LUC) ora mutated AGL15 promoter in which both RY elements were dis-rupted (AGL15mpro:LUC) was tested by transient expression inNicotiana benthamiana leaves by agroinfiltration (Figure 3A). Sub-stantial levelsof luminescence, indicating luciferaseactivity, andLUCmRNA were seen in leaves agroinfiltrated with either the AGL15pro:LUC or the AGL15mpro:LUC construct alone (Figure 3B). However,expression from theAGL15pro:LUC reporter construct was stronglyreduced when coinfiltrated with an HSI2-expressing effector con-struct, while expression from the AGL15mp:LUC construct wasunaffected by HSI2 coexpression (Figure 3B). Furthermore, lumi-nescence and luciferase expression assays of seedlings of stabletransgenic Arabidopsis lines that contained the AGL15pro:LUC orAGL15mpro:LUC constructs in a wild-type genetic backgroundshowed that loss of the RY elements in this promoter led to de-repression of theAGL15 promoter, resulting in increased expressionof the luciferase reporter gene (Figures 3C and 3D).To investigate whether both RY elements are required for HSI2-
dependent AGL15 transcriptional repression, luciferase reportergenes driven by mutated AGL15 promoters in which one orthe other RY element was disrupted (AGL15mpro1:LUC andAGL15mpro2:LUC) were developed (Figure 3A) and agroinfiltrated,with or without the HSI2-expressing effector construct, into N.benthamiana leaves for transient expression assays. High levels ofluciferase activity and LUC mRNA were detected when either theAGL15mpro1:LUC or the AGL15mpro2:LUC construct was tran-siently expressed inN. benthamiana leaves. However, reporter geneexpressionwassignificantly reducedwheneitherof theseconstructswas coexpressed with HSI2 (Figure 3E). These results demonstratethat, at least in agroinfiltrated N. benthamiana leaves, a single RYelement isnecessaryandsufficient forHSI2-dependent repressionoftranscription by the AGL15 promoter.
B3 Domain Is Required for HSI2 to Bind to RY Elements andRepress AGL15
The B3 domain of HSI2 recognizes and binds to RY motifs in theFLC silencing elements and is required to repress FLC expression(Qüesta et al., 2016;Yuanet al., 2016). Since, as shown inFigure 3,an RY element is required for the HSI2-dependent repression ofthe AGL15 promoter, the HSI2 B3 domain may also play a role inAGL15 repression. To test this hypothesis, an effector constructthat encodes a B3 domain-deleted HSI2 (HSI2-DB3) was co-infiltrated with the AGL15pro:LUC reporter construct into N.benthamiana leaves (Figure 4A). Expressionof theAGL15pro:LUCreporter gene was strongly repressed by coexpression with HSI2but, when coexpressed with HSI2-DB3, reporter gene expressionremained high (Figure 4B). Previously reported soluble structureanalysis indicated that two lysine residues, which correspond toK297 and K314 of HSI2, are located on the DNA binding surfaces
602 The Plant Cell
of ABI3 and RAV1 and these residues were predicted to makedirect contact with DNA (Yamasaki et al., 2004). Furthermore, itwas demonstrated that the DNA binding activity of maize VIV-IPOROUS1 (zmVP1), a member of the ABI3 family of B3 domainproteins, was lost almost completely when K519, which corre-spondswith K297 of HSI2, was replaced by arginine (Suzuki et al.,2014). Therefore, we predicted that these two conserved lysinesare required for the function of HSI2. To test this hypothesis, K297and K314 were replaced by arginine (K297R and K314R) by site-directed mutagenesis (Figure 4C). The HSI2pro:HSI2mB3 con-structwas transformed into hsi2-2LUCArabidopsis tomeasure theability of HSI2mB3 to repress the expression of the AGL15 genesin planta. Expression of HSI2mB3-GFP was confirmed in theHSI2mB3 transgenic line by the GFP expression (Figure 4D).Relative expression of AGL15 in HSI2mB3 seedlings was sig-nificantly higher than in WTLUC plants and similar to the levels inhsi2-2LUC (Figure 4D; Supplemental Figure 1). Taken together,these results demonstrate that the HSI2 B3 domain is required forHSI2-dependent repression of the AGL15 promoter.
To test whether the HSI2 B3 domain can bind directly to the RYelements in theAGL15 promoter in vitro, His-Trx-taggedHSI2-B3
fusionproteinwasexpressed inEscherichia coliandpurifiedusingDynabeads. For DNA pull-down experiments, this recombinantproteinwas incubated independentlywithDNAprobes containingtwo intact RY elements or probes in which the 5ʹ (RY1) or 3ʹ (RY2),or both RY elements were mutated (Figure 5A). Following purifi-cation of the protein-DNA complexes, the probe was quantifiedusing qPCR. The HSI2 B3 domain-containing fusion proteinbound strongly with DNA fragments containing tandem RY ele-ments, while little binding activity was detected when incubatedwith DNA fragments in which both RY elements were mutated(Figure 5A). Binding of probes that contain single intact RY ele-ments in the HSI2 B3 domain was reduced by more than 50%compared with the probe with two RY elements. To confirm thatsingle RY elements can bind the HSI2 B3 domain in vitro, elec-trophoretic mobility shift assays (EMSAs) were also performed.RY1- and RY2-containing DNA probes from the AGL15 proximalpromoter were labeled with biotin at the 3ʹ end. EMSA resultsshowed that HSI2-B3 was able to bind to both RY1 and RY2-containing probes (Figure 5B). Probes in which either RY1 or RY2sequences were mutated failed to bind HSI2-B3, and an HSI2-B3polypeptide with a disrupted B3 domain failed to bind intact RY1
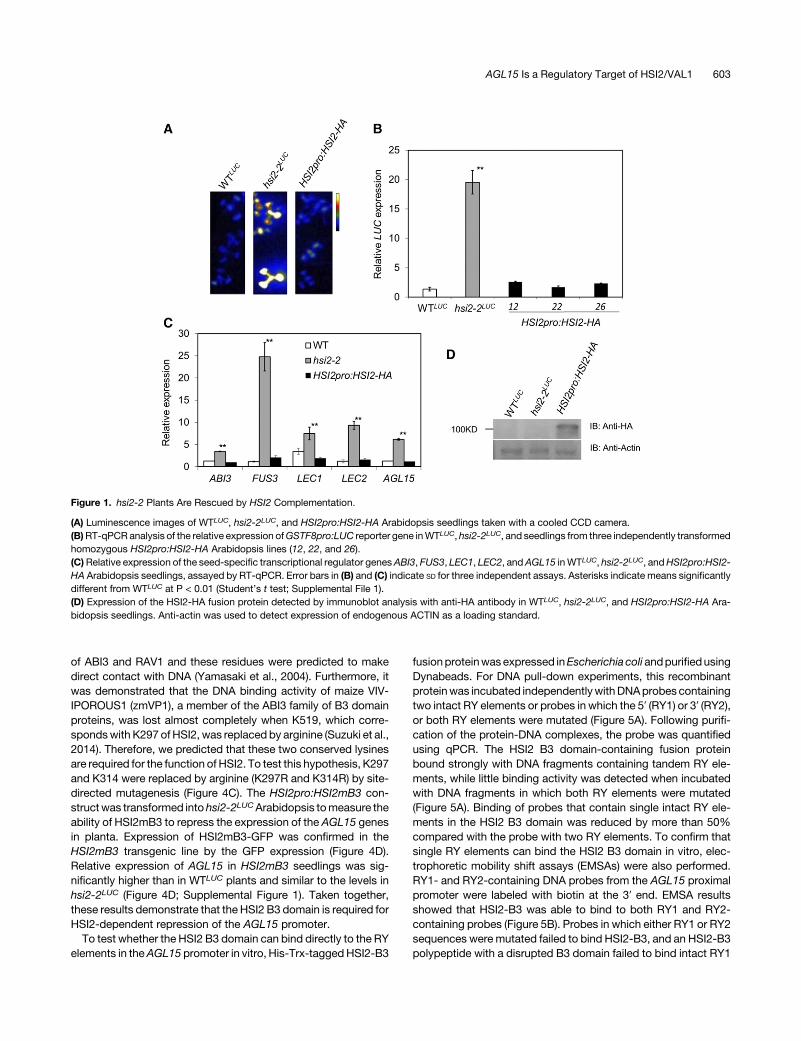
Figure 1. hsi2-2 Plants Are Rescued by HSI2 Complementation.
(A) Luminescence images of WTLUC, hsi2-2LUC, and HSI2pro:HSI2-HA Arabidopsis seedlings taken with a cooled CCD camera.(B)RT-qPCR analysis of the relative expression ofGSTF8pro:LUC reporter gene inWTLUC, hsi2-2LUC, and seedlings from three independently transformedhomozygous HSI2pro:HSI2-HA Arabidopsis lines (12, 22, and 26).(C)Relative expression of the seed-specific transcriptional regulator genesABI3, FUS3, LEC1, LEC2, andAGL15 inWTLUC, hsi2-2LUC, andHSI2pro:HSI2-HA Arabidopsis seedlings, assayed by RT-qPCR. Error bars in (B) and (C) indicate SD for three independent assays. Asterisks indicate means significantlydifferent from WTLUC at P < 0.01 (Student’s t test; Supplemental File 1).(D) Expression of the HSI2-HA fusion protein detected by immunoblot analysis with anti-HA antibody in WTLUC, hsi2-2LUC, and HSI2pro:HSI2-HA Ara-bidopsis seedlings. Anti-actin was used to detect expression of endogenous ACTIN as a loading standard.
AGL15 Is a Regulatory Target of HSI2/VAL1 603

and RY2 probes. These results indicate that the HSI2-B3 domaincan specifically binds to the both AGL15 RY1 and RY2 elements.
To investigate if disruption of the HSI2 B3 domain affects HSI2enrichment at theAGL15 locus in plant cells, we performed ChIP-qPCRassaysusinghsi2-2Arabidopsis leaf protoplasts to expressHSI2-GFP and HSI2mB3-GFP under control of the HSI2 nativepromoter, as described in Methods. Our analysis revealed that,comparedwithHSI2, accumulation of HSI2mB3at P1, P2, andC1areas decreased significantly (Figure 5C). However, comparedwith control protoplasts that express GFP only, enrichment ofHSI2mB3atP1andP2siteswassomewhatgreater. This suggests
that othermotifs, in addition to theB3domain, couldbe involved inthe recruitment of HSI2 to its target loci.
PHD Domain Is Required for HSI2 to RepressAGL15 Expression
HSI2-dependent transcriptional repression of GSTF8pro:LUC re-porter gene, along with other HSI2-repressed genes, includingAGL15, is compromised in hsi2-4 plants that carry a C-to-Y sub-stitutionatposition65 in thePHDdomain (Veerappanetal., 2012). Totest the hypothesis that the HSI2 PHD domain is required for HSI2
Figure 2. HSI2 Accumulation at the AGL15 Locus.
ChIP-qPCRanalysisofHSI2/VAL1enrichmentatLEC1,FUS3,ABI3,LEC2, andAGL15genes inwild-typeandHSI2-rescuedArabidopsisplants.Schematicrepresentations of genes and assayed genomic regions are indicated, with P indicating promoter regions andC indicating coding regions. The positions ofRYelementswithin upstreamsequencesof thesegenesare indicated. These regionswere interrogatedbyChIP-qPCRanalysis of 7-d-old transgenic plantsharboringHSI2pro:HSI2-HAusingHAantibody. TheamountofDNAafterChIPwasnormalizedusingACT2asan internal standard.Data representmeansofthreeqPCRreactions fromeachof three independentChIPassays.Errorbars indicate SD.Asterisks indicatemeanssignificantlydifferent fromthewild typeatP < 0.01 (Student’s t test; Supplemental File 1).
604 The Plant Cell

Figure 3. RY Elements Are Required for HSI2 Repression of AGL15 Expression.
(A) Schematic representation of reporter and effector constructs used to assay the function of RY elements in the AGL15 upstream sequence. In reporterconstructs, a luciferase gene is driven by the AGL15 promoter or an RY-mutated AGL15 promoter. Red bars represent mutation sites.(B) Luminescence images and relative expression of luciferase mRNA, as determined by RT-qPCR assays in N. benthamiana leaves coinfiltrated with thereporter and effector gene combinations indicated. RT-PCR analysis of HSI2 and NbActin gene expression in each infiltrated area.(C) Luminescence image of Arabidopsis seedlings that contain either AGL15pro:LUC or AGL15mpro:LUC reporter genes.(D) Relative expression of luciferase reporter gene, by RT-qPCR, corresponding to the luminescence image above in Arabidopsis seedlings.(E) Luminescence images and relative expression of luciferase reporter genes, assayed by RT-qPCR from N. benthamiana leaves coinfiltrated withcombinations of reporter and effector constructs, as indicated. HSI2 and NbActin gene expression was assayed by RT-PCR in each infiltrated area. Datarepresent means of three qPCR reactions from three independent assays per genotype. Error bars indicate SD. Asterisks indicate that luciferase activity ininfiltrations with the 35Spro:HSI2 effector is significantly different from those without the effector at P < 0.01 (Student’s t test; Supplemental File 1).
AGL15 Is a Regulatory Target of HSI2/VAL1 605

transcriptional repression of AGL15, expression of AGL15pro:LUCreporter genewhencoexpressedwith intactHSI2or amutant formofHSI2 in which the eight conserved amino acids of the PHD domainare substituted (C39S, C42S, C65S, C68S, H73L, C76S, C93S, andC96S) was analyzed by transient expression in N. benthamianaleaves (Figures 6A and 6B). As expected, both luciferase activity andmRNA expression were significantly reduced when AGL15pro:LUCwascoexpressedwith intactHSI2butcoexpressionwithHSI2mPHDhadnosignificant effect on reporter geneexpression (Figure6C). Thetranscriptional repressor activity of HSI2mPHD was also assayed instable transgenic Arabidopsis plants that express the HSI2pro:HSI2mPHD-HA construct in the hsi2-2LUC background. Expressionof HSI2mPHD-HA was confirmed in these plants by immunoblotanalysis using anti-HA (Figure 6D). Relative expression of the nativeAGL15 gene in HSI2mPHD seedlings was significantly upregulatedcompared with WTLUC, and the upregulation was similar to that in
hsi2-2LUCseedlings (Figure6E;SupplementalFigure1).Theseresultsconfirm that the HSI2 PHD domain is required for transcriptionalrepression of the AGL15 promoter.TheHSI2 PHDdomain has been reported to recognize andbind
to H3K27me3 in vitro (Yuan et al., 2016) and disruption of HSI2PHD domain leads to decreased H3K27me3 chromatin marks attheAGL15 locus (Veerappanetal., 2014), resulting inderepressionofAGL15 expression. One possible role for the HSI2 PHD domaincould be that it is involved in targetingHSI2 to specific loci. To testthis hypothesis, ChIP-qPCR assays were performed to measurethe enrichment of PHD-mutated HSI2 at the AGL15 locus inHSI2pro:HSI2mPHD-HA and HSI2pro:HSI2-HA transgenic Ara-bidopsis seedlings (Figure 6F). Compared with HSI2-HA, theaccumulation of HSI2mPHD-HA was significantly lower at theAGL15 locus, showing levels similar to wild-type seedlings. Thus,an intact PHD domain is required for HSI2 enrichment at the
Figure 4. B3 Domain Is Required for HSI2 Activity.
(A)Schematic representation of reporter and effector constructs used to assay the function of the HSI2 B3 domain. In the reporter construct, the luciferasegene is driven by the AGL15 promoter. In effector constructs, expression of intact HSI2 or HSI2-DB3 is controlled by the CaMV 35S promoter.(B)Luminescence imageproducedbyacooledCCDcameraand relative expressionof the luciferase reporter gene fromN.benthamiana leavescoinfiltratedwith combinations of reporters and effectors, as indicated, was assayed by RT-qPCR. HSI2 and NbActin gene expression in each infiltrated area wasassayed by RT-PCR.(C)Amino acid sequence alignment of the N-terminal B3 domains among Arabidopsis HSI2, HSL1, VAL3, ABI3, LEC2, FUS3, RAV1, RAV2, andmaize VP1.Red stars indicate substitution mutation sites within the HSI2 B3 domain.(D) Relative expression of endogenous AGL15 in WTLUC, hsi2-2LUC, and HSI2pro:HSI2mB3-GFP Arabidopsis lines was assayed by RT-qPCR andnormalized by EF1a.HSI2mB3-GFP and EF1a gene expression inWTLUC, hsi2-2LUC, andHSI2mB3-GFP Arabidopsis lines was assayed by RT-PCR. Datarepresent means of three qPCR reactions for each locus from three independent assays. Error bars indicate SD. Asterisks indicate significantly differentmeans at P < 0.01 (Student’s t test; Supplemental File 1).
606 The Plant Cell

AGL15 locus, indicating that it isnecessary forHSI2binding invivo(Figure 6F). Using pull-down assays, we confirmed that a fusionprotein that contained the HSI2 PHD domain could interact withH3K27me3peptidesas reportedbyYuanet al. (2016) andshowedthat it could also interact with H3K4me3 peptides (Figure 6G).These interactions were lost, for the most part, when a fusionprotein that contained amutated PHDdomain (C65Y) was used inthe pull-down assay (Figure 6G).
EAR Motif Is Required for Full HSI2 Activity
The HSI2 EAR motif was shown to be necessary for full HSI2-dependent repression of a luciferase reporter gene under controlof the sugar-responsive Sporamin promoter from sweet potato(Ipomoea batatas;Spomin) in Arabidopsis protoplasts (Tsukagoshi
et al., 2005). Therefore, we tested whether the EAR motif con-tributes to HSI2-dependent transcriptional repression of theAGL15 promoter. Coinfiltration of a construct encoding EARmotif-deletedHSI2 (HSI2-DEAR)with theAGL15pro:LUC reportergene construct into N. benthamiana leaves resulted in reducedluciferase expression compared with infiltration with the reportergene alone but repression was only about half that seen withcoexpression of intact HSI2 (Figures 7A and 7B). These resultsshow that expression of HSI2 that lacks the EAR motif leads topartial derepression of AGL15, indicating that this motif contrib-utes to, but is not essential for, the transcriptional repressor ac-tivity of HSI2 in this assay.Protein sequence comparison indicates that the EAR motif
includes a core of three conserved amino acids that are predicted
Figure 5. HSI2 B3 Domain Interacts with RY Elements in the AGL15 Promoter.
(A) DNA binding assay corresponding to the putative RY binding sites of the HSI2-B3 domain. DNA fragments from AGL15 proximal promoter areacontainingRYandmutatedRYelements bound toHis-Trx-B3 fusion protein orHis-Trx control were quantifiedbyqPCRafter elution. Data representmeansof three technical replicates from three independent assays. Boxes and red letters denote RY elements andmutated sites, respectively. Error bars indicateSD. Asterisks indicate means significantly different from control reactions (His-Trx) at P < 0.01 (Student’s t test; Supplemental File 1).(B)EMSAsof interactionbetweenRY1andRY2, andmutant,mRY1, andmRY2probeswithHis-HSI2B3andmutantHis-HSI2mB3proteins. Freeprobeandbound probe bands are indicated.(C)Enrichment of HSI2-GFP andHSI2mB3-GFP at theAGL15 locus in Arabidopsis protoplasts assayed byChIP-qPCR. Interrogated regions are indicatedin the gene diagram. Data represent means of three ChIP-qPCR assays from three independent assays for each genotype. Error bars indicate SD. Asterisksindicate means significantly different from control (GFP) at P < 0.01 (Student’s t test; Supplemental File 1).
AGL15 Is a Regulatory Target of HSI2/VAL1 607
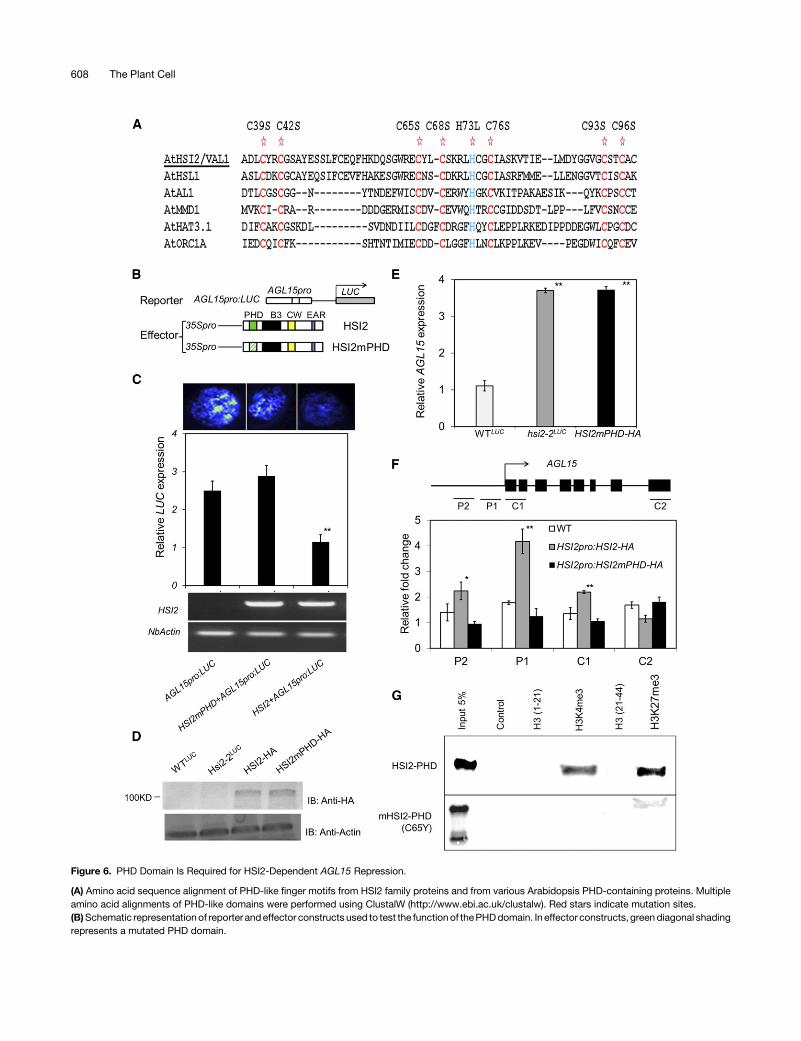
Figure 6. PHD Domain Is Required for HSI2-Dependent AGL15 Repression.
(A) Amino acid sequence alignment of PHD-like finger motifs from HSI2 family proteins and from various Arabidopsis PHD-containing proteins. Multipleamino acid alignments of PHD-like domains were performed using ClustalW (http://www.ebi.ac.uk/clustalw). Red stars indicate mutation sites.(B)Schematic representationof reporter andeffector constructs used to test the functionof thePHDdomain. In effector constructs, greendiagonal shadingrepresents a mutated PHD domain.
608 The Plant Cell

tobecritical forEAR function (Tsukagoshi et al., 2005;Zhanget al.,2013). Site-directed mutagenesis was used to replace the highlyconserved leucineatposition730within theEARmotifwithalanine(L730A) (Figure 7C). The transcriptional repressor activity ofHSI2mEAR was assayed in planta by stable transformation of theHSI2pro:HSI2mEAR-HA construct into hsi2-2LUC Arabidopsis.Expression of HSI2mEAR-HA was confirmed in the HSI2mEARtransgenic line by immunoblot analysis using anti-HA (Figure 7D).Luminescence and LUC mRNA expression in HSI2mEAR seed-lings was much stronger than in WTLUC plants, which expressintact HSI2, butwas significantly weaker than hsi2-2LUC knockoutplants (Figure 7E). Relative expressionof the nativeAGL15gene inHSI2mEAR seedlings was also derepressed significantly com-pared withWTLUC but, like the reporter gene, failed to reach to theexpression level seen in hsi2-2LUC seedlings (Figure 7F;Supplemental Figure 1). These results confirm that the HSI2 EARmotif contributes to HSI2-mediated transcriptional repression ofthe native AGL15 gene.
CW Domain Does Not Contribute to HSI2-DependentRepression of AGL15
Previous research showed that the CW domain of HSL1, but notof HSI2, physically interacts with HDA19, leading to histonedeacetylation of chromatin at its target loci and thus plays a role intheir repression (Zhou et al., 2013). Therefore, we examinedwhether the CW domain of HSI2 contributes to the repression ofAGL15. Based on protein sequence alignment between HSI2 andother CW domain-containing proteins in Arabidopsis, two tryp-tophan and four cysteine residues were predicted to be critical forCW domain function (Figure 8A). These four conserved cysteinesat positions 547, 550, 568, and 580 were replaced by serine(C547S, C550S, C568S, and C580S) through site-directed mu-tagenesis (Figure 8A). The HSI2pro:HSI2mCW construct wastransformed into hsi2-2LUC Arabidopsis to measure the ability ofHSI2mCW to repress the expression of the GSTF8pro:LUC re-porter and AGL15 genes in planta. Expression of HSI2mCW-HAwas confirmed in the HSI2mCW transgenic line by immunoblotanalysis using anti-HA (Figure 8B). Luminescence from theGSTF8pro:LUC reporter and relative expression of both LUC
mRNA and the native AGL15 gene in HSI2mCW seedlings wassimilar to the expression levels seen in WTLUC, but was stronglyrepressedcomparedwithhsi2-2LUCseedlings (Figures8Cand8D;Supplemental Figure 1). Since the ability of HSI2 to represstranscription of either the GSTF8pro:LUC transgene or nativeAGL15 was not deleteriously affected by multiple substitutionmutationsatcriticalpositions in theCWdomain, it appears that theCW domain is not required for the HSI2-mediated transcriptionalrepression in this system.
HSI2 Represses Key Regulators of Seed Maturation throughDownregulation of AGL15
Based on the results reported here, we hypothesize that HSI2directly targets AGL15 but not LEC1, ABI3, FUS3, and LEC2,leading to repression of the seed maturation program. Based onthis hypothesis, we predict that HSI2 is epistatic to AGL15 andrepresses key regulators of seed maturation via the down-regulation of AGL15. To test this prediction, the relative expres-sionofLEC1,ABI3,FUS3, andLEC2wasexamined in seedlingsofArabidopsis hsi2-2 and agl15-4 T-DNA knockout lines and anAGL15-overexpressing line (Figure 9A). Changes in the expres-sion of AGL15 in these lines was confirmed by RT-qPCR analysis(Figure 9B). Since AGL15 expression is repressed in wild-typeseedlings, we predicted that agl15 loss-of-function mutationsshould have little effect on the expression of downstream targetgenesand thisproved tobe thecase,asexpressionofLEC1,ABI3,FUS3, and LEC2 in agl15-4 seedlings was not significantly re-duced relative to thewild type (Figure 9A). However, expression ofLEC1,ABI3,FUS3, andLEC2wassignificantly elevated inAGL15-overexpressing plants, consistent with AGL15 functioning toactivate the expression of these genes (Zheng et al., 2009), yet theincrease inLAFLgeneexpression in these lineswasnotequivalentto the levels seen inhsi2-2seedlings. Interestingly, relative tohsi2-2, hsi2-2 agl15-4 double knockout seedlings showed significantlyreduced expression of LEC1, ABI3, FUS3, and LEC2 (Figure 9A),yet the levels of expression remained significantly greater than inwild-type plants. We interpret these results to indicate that HSI2-dependent silencing of these LAFL seed maturation regulatorygenes in Arabidopsis seedlings is indirect and mediated, in large
Figure 6. (continued).
(C) Luminescence image and relative expression of luciferase reporter genes, measured by RT-qPCR, from N. benthamiana leaves coinfiltrated withcombinations of reporter and effector constructs, as indicated. HSI2 and NbActin gene expression in each infiltrated area was assayed by RT-PCR.(D) HSI2mPHD protein was detected by immunoblot assays using anti-HA antibody, anti-actin was used to detect actin as a loading standard.(E)Relative expression of nativeAGL15 inWTLUC,hsi2-2LUC, andHSI2pro:HSI2mPHD-HA transgenic Arabidopsis plants. All data representmeansof threeRT-qPCR reactions from three independent assays for each genotype or treatment. Error bars indicate SD. Asterisks indicate means significantly differentfrom control (WTLUC) at P < 0.01 (Student’s t test; Supplemental File 1).(F) Results of ChIP-qPCR analyses to compare the enrichment of HSI2-HA and HSI2mPHD-HA at the AGL15 locus in Arabidopsis plants. Interrogatedregionsare indicated in thegenediagram.Data representmeansof threeChIP-qPCRassays fromthreebiological replicates.Errorbars indicate SD.Asterisksindicate means significantly different from control plants (WT) at P < 0.01 (Student’s t test; Supplemental File 1).(G) In vitro histone peptide binding assay of GST-fused PHD domain of HSI2 with H3K4me3 and H3K27me3 polypeptides. Various biotinylated histonepeptidesbound toGST-fusionPHDdomainofHSI2wereprecipitatedusing streptavidin agarosebeadsandsubjected to immunoblot assayusinganti-GSTantibody. Input indicates 5% of GST-fusion proteins used for pull-down assay. Control indicates no histone peptide in reaction. H3 (1-21), nonmethylatedhistoneH3 amino acid residues 1 to 21, and used as control for H3K4me3 (histoneH3, 1–21, trimethylated at Lys-4); H3 (21-44), nonmethylated histoneH3amino acid residues 21 to 44, and used as control for H3K27me3 (histoneH3, 21–44 trimethylated at Lys-27). Amino acid changedbypointmutation (C65Y)is indicated based on the position within the full-length protein.
AGL15 Is a Regulatory Target of HSI2/VAL1 609

Figure 7. EAR Domain Is Important for Full HSI2 Activity.
(A)Schematic representationof reporter andeffectorconstructsused to test the functionof theEARdomain.Effectorconstructsencodeeither intactHSI2ora C-terminal deletion that lacks the EAR domain.(B) Luminescence image and relative expression analysis of the LUCmRNA, by RT-qPCR, from N. benthamiana leaves coinfiltrated with combinations ofreporter and effector constructs, as indicated. RT-PCR analysis of HSI2 and NbActin gene expression in infiltrated areas as above.(C)Amino acid sequence alignment of EARmotifs fromHSI2 family proteins and fromvariousArabidopsis EAR-containingproteins alignedusingClustalW.Red star indicates a single substitution mutation used to disrupt the EAR motif of HSI2 protein.(D)HSI2mEAR protein expression in transgenic plants was detected by immunoblot assays using anti-HA antibody. Anti-actin was used to detect actin asa loading standard.(E) Luminescence images to compare WTLUC, hsi2-2LUC, and HSI2pro:HSI2mEAR-HA Arabidopsis seedlings and relative expression analysis of the LUCreporter gene corresponding to the luminescence image. RT-qPCR assays were normalized by EF1a.
610 The Plant Cell
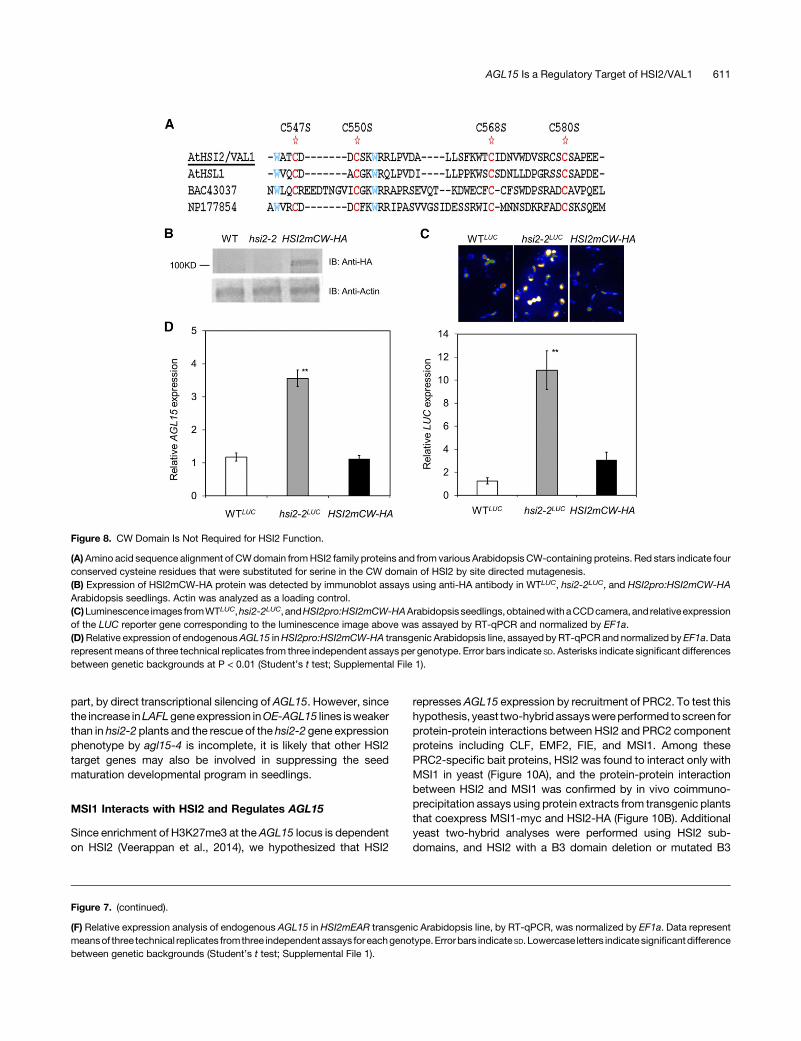
part, by direct transcriptional silencing of AGL15. However, sincethe increase inLAFLgeneexpression inOE-AGL15 lines isweakerthan in hsi2-2 plants and the rescue of the hsi2-2 gene expressionphenotype by agl15-4 is incomplete, it is likely that other HSI2target genes may also be involved in suppressing the seedmaturation developmental program in seedlings.
MSI1 Interacts with HSI2 and Regulates AGL15
Since enrichment of H3K27me3 at the AGL15 locus is dependenton HSI2 (Veerappan et al., 2014), we hypothesized that HSI2
represses AGL15 expression by recruitment of PRC2. To test thishypothesis, yeast two-hybrid assayswereperformed toscreen forprotein-protein interactions between HSI2 and PRC2 componentproteins including CLF, EMF2, FIE, and MSI1. Among thesePRC2-specific bait proteins, HSI2 was found to interact only withMSI1 in yeast (Figure 10A), and the protein-protein interactionbetween HSI2 and MSI1 was confirmed by in vivo coimmuno-precipitation assays using protein extracts from transgenic plantsthat coexpress MSI1-myc and HSI2-HA (Figure 10B). Additionalyeast two-hybrid analyses were performed using HSI2 sub-domains, and HSI2 with a B3 domain deletion or mutated B3
Figure 7. (continued).
(F) Relative expression analysis of endogenous AGL15 in HSI2mEAR transgenic Arabidopsis line, by RT-qPCR, was normalized by EF1a. Data representmeansof three technical replicates fromthree independentassays foreachgenotype.Errorbars indicate SD.Lowercase letters indicatesignificantdifferencebetween genetic backgrounds (Student’s t test; Supplemental File 1).
Figure 8. CW Domain Is Not Required for HSI2 Function.
(A)Amino acid sequence alignment of CWdomain fromHSI2 family proteins and from various Arabidopsis CW-containing proteins. Red stars indicate fourconserved cysteine residues that were substituted for serine in the CW domain of HSI2 by site directed mutagenesis.(B) Expression of HSI2mCW-HA protein was detected by immunoblot assays using anti-HA antibody in WTLUC, hsi2-2LUC, and HSI2pro:HSI2mCW-HAArabidopsis seedlings. Actin was analyzed as a loading control.(C)Luminescence images fromWTLUC,hsi2-2LUC, andHSI2pro:HSI2mCW-HAArabidopsis seedlings,obtainedwithaCCDcamera, and relativeexpressionof the LUC reporter gene corresponding to the luminescence image above was assayed by RT-qPCR and normalized by EF1a.(D)Relative expression of endogenousAGL15 inHSI2pro:HSI2mCW-HA transgenic Arabidopsis line, assayed by RT-qPCR and normalized byEF1a. Datarepresentmeans of three technical replicates from three independent assays per genotype. Error bars indicate SD. Asterisks indicate significant differencesbetween genetic backgrounds at P < 0.01 (Student’s t test; Supplemental File 1).
AGL15 Is a Regulatory Target of HSI2/VAL1 611

Figure 9. Expression of LEC1, ABI3, FUS3, and LEC2 in hsi2 agl15 Plants.
(A)Relative expressionofLEC1,ABI3,FUS3, andLEC2wasmeasured inwild-type, agl15-4,AGL15-overexpressor (OE-AGL15),hsi2-2, andhsi2-2 agl15-4Arabidopsis lines by RT-qPCR.(B)RelativeexpressionofAGL15 inwild-type,agl15-4,OE-AGL15,hsi2-2, andhsi2-2agl15-4Arabidopsis lines.Theagl15-4 line isnull andexpression in theOE-AGL15 is;2003higher than thewild type.DatawerenormalizedbyEF1aandrepresentmeansof three technical replicates fromthree independentRNAsamples for each genotype. Error bars indicate SD. Lowercase letters indicate significant differences (P < 0.01) between genetic backgrounds (Student’st test; Supplemental File 1).
612 The Plant Cell
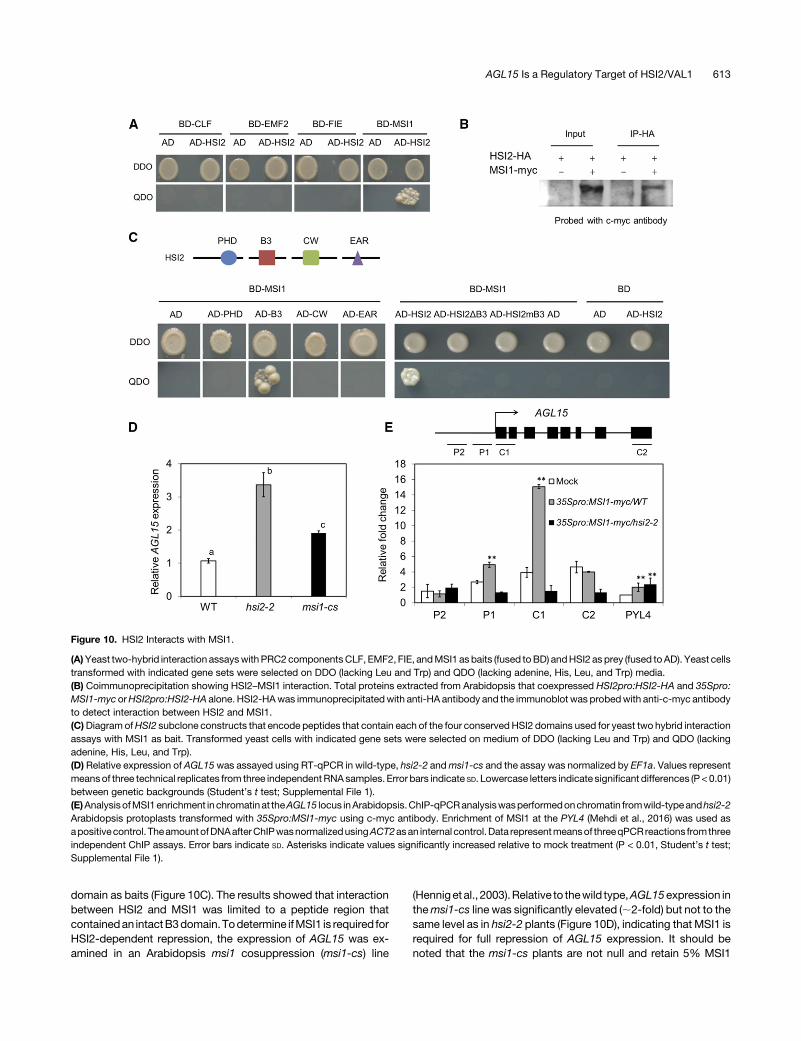
domain as baits (Figure 10C). The results showed that interactionbetween HSI2 and MSI1 was limited to a peptide region thatcontainedan intactB3domain.Todetermine ifMSI1 is required forHSI2-dependent repression, the expression of AGL15 was ex-amined in an Arabidopsis msi1 cosuppression (msi1-cs) line
(Henniget al., 2003).Relative to thewild type,AGL15expression inthemsi1-cs line was significantly elevated (;2-fold) but not to thesame level as in hsi2-2 plants (Figure 10D), indicating that MSI1 isrequired for full repression of AGL15 expression. It should benoted that the msi1-cs plants are not null and retain 5% MSI1
Figure 10. HSI2 Interacts with MSI1.
(A)Yeast two-hybrid interaction assayswithPRC2componentsCLF, EMF2, FIE, andMSI1 as baits (fused toBD) andHSI2 as prey (fused toAD). Yeast cellstransformed with indicated gene sets were selected on DDO (lacking Leu and Trp) and QDO (lacking adenine, His, Leu, and Trp) media.(B) Coimmunoprecipitation showing HSI2–MSI1 interaction. Total proteins extracted from Arabidopsis that coexpressed HSI2pro:HSI2-HA and 35Spro:MSI1-myc orHSI2pro:HSI2-HA alone. HSI2-HAwas immunoprecipitatedwith anti-HA antibody and the immunoblot was probedwith anti-c-myc antibodyto detect interaction between HSI2 and MSI1.(C)Diagram ofHSI2 subclone constructs that encode peptides that contain each of the four conservedHSI2 domains used for yeast two hybrid interactionassays with MSI1 as bait. Transformed yeast cells with indicated gene sets were selected on medium of DDO (lacking Leu and Trp) and QDO (lackingadenine, His, Leu, and Trp).(D) Relative expression of AGL15was assayed using RT-qPCR in wild-type, hsi2-2 andmsi1-cs and the assay was normalized by EF1a. Values representmeansof three technical replicates from three independentRNAsamples. Error bars indicate SD. Lowercase letters indicate significant differences (P<0.01)between genetic backgrounds (Student’s t test; Supplemental File 1).(E)AnalysisofMSI1enrichment inchromatinat theAGL15 locus inArabidopsis.ChIP-qPCRanalysiswasperformedonchromatin fromwild-typeandhsi2-2Arabidopsis protoplasts transformed with 35Spro:MSI1-myc using c-myc antibody. Enrichment of MSI1 at the PYL4 (Mehdi et al., 2016) was used asapositivecontrol. TheamountofDNAafterChIPwasnormalizedusingACT2asan internal control.Data representmeansof threeqPCRreactions fromthreeindependent ChIP assays. Error bars indicate SD. Asterisks indicate values significantly increased relative to mock treatment (P < 0.01, Student’s t test;Supplemental File 1).
AGL15 Is a Regulatory Target of HSI2/VAL1 613

expression relative towild-typeplants. To test if, likeHSI2,MSI1 isenriched at the AGL15 locus, protoplasts isolated from wild-typeand hsi2-2Arabidopsis plants were transformedwith aMSI1-mycexpressing construct (35Spro:MSI1-myc) and analyzed by ChIP-qPCR (Figure 10E). The results showed significantly higher MSI1signal at the proximal promoter and 5ʹ coding region of theAGL15locus in wild-type Arabidopsis protoplasts than in hsi2-2 proto-plasts, indicating that accumulation ofMSI1 at theAGL15 locus isHSI2 dependent. Since enrichment ofMS1 at the 5ʹ coding regionis stronger than at the proximal promoter, it is possible that HSI2may recruit MSI1 to form a PRC2 nucleation site at the proximalpromoter region of AGL15, which then expands into the 5ʹ codingregion. We interpret these results to indicate that HSI2-mediatedrepression ofAGL15 is dependent on thePRC2componentMSI1.
DISCUSSION
The major regulatory genes of seed maturation, LEC1, ABI3,FUS3, and LEC2, constitute a genetic pathway to activate theseed maturation program (Lotan et al. 1998; Luerßen et al., 1998;Stone et al., 2001; Finkelstein et al., 2008; Holdsworth et al., 2008;Suzuki and McCarty 2008). AGL15 is reported to act upstream ofthese key regulatory genes to promote their expression duringseed development (Zheng et al., 2009). The regulatory modelshown in Figure 11 summarizes our results and those presentedbyChhunetal. (2016) regarding the roleofHSI2 in the repressionofthe seed maturation developmental program in seedlings aftergermination. Accordingly, expression of several key transcrip-tional regulators of the seedmaturationgeneexpressionprogram,including LEC1, ABI3, FUS3, LEC2, and AGL15, is upregulated inhsi2-2 loss-of-function Arabidopsis seedlings (Figure 1C; Suzukiet al. 2007, Tsukagoshi et al. 2007; Holdsworth et al. 2008, SuzukiandMcCarty 2008; Veerappan et al., 2012). Increased expressionofABI3andLEC1wasaccompaniedby relativelyminordecreasesin H3K27me3 deposition at these loci and no change was seen atthe FUS3 and LEC2 loci (Veerappan et al., 2014). On the otherhand, H3K27me3 was strongly decreased at the AGL15 locus inboth hsi2 knockout and PHD domain mutants (hsi2-2 and hsi2-4,respectively; Veerappan et al., 2014). Results of ChIP analysesindicate that thatHSI2-HA is substantially enriched in theproximalpromoter region of the AGL15 locus, which contains two putativeB3 domain binding RY elements, but no detectable enrichment ofHSI2-HA was observed at the LEC1, ABI3, FUS3, or LEC2 loci inour assays (Figure 2). Based on these results, we predict that theexpression of AGL15 in Arabidopsis seedlings is directly re-pressed byHSI2, while downregulation of LEC1,ABI3, FUS3, andLEC2 expression is likely to be indirect and possibly a result of thedownregulation of AGL15. These findings conflict with those re-ported by Chhun et al. (2016), who found enrichment of HSI2 at 5ʹregions of LEC1, LEC2, ABI3, and FUS3 loci, leading them topropose these genes as direct targets of HSI2 transcriptionalrepression. However, the plants used for their ChIP experimentsexpressed HA-tagged HSI2 under control of the strongly con-stitutiveCaMV 35S promoter instead of the nativeHSI2 promoterand had transcript levels 30-fold higher than the wild type. Oneexplanation for theseconflicting results couldbe thatHSI2has lowaffinity for the RY sites upstream of these LAFL genes and binds
only when intracellular concentrations of HSI2 are quite high, as inthe HSI2-HA overexpressing plants used by Chhun et al. (2016).Whether interactionunder theseconditions is relevant towild-typeplants has yet to be elucidated. Thus, our results indicate that,when expressed under control of its native promoter, HSI2-HAaccumulates at theAGL15 locus but not at LAFL gene loci in 7-d-old Arabidopsis seedlings. It remains possible that HSI2 couldinteract with LAFL genes at other developmental stages or underdifferent physiological conditions.Analysis of the direct transcriptional silencing of the AGL15
promoter by transient coexpression of luciferase reporter genescontrolled by either the intact AGL15 promoter or AGL15 pro-moters in which the RY elements were disrupted indicate that atleastoneRYelement is required forHSI2-dependent repressionofthe AGL15 promoter (Figure 3). Protein-DNA binding assaysconfirmed that theHSI2-B3domain is able to bind to eachof theseRY elements individually and binding increased additively whentwo intactRYelementswerepresent,whilebindingwasabrogatedwith the disruption of both RY elements (Figure 5A). Furthermore,while the RY elements found in the AGL15 promoter include thecore consensus sequence 5ʹ-CATGC-3ʹ, they vary somewhatfromtheelements targetedbyLEC2,FUS3,andABI3, inwhichG ispreferred immediately upstreamof the core sequence (Baud et al.,2016). So, it is possible that HSI2 may have stronger affinity toa subset of RY elements that differ somewhat from those rec-ognizedmost strongly by other B3-domain proteins, but this is yetto be tested directly.The B3 domain deletion effector construct (HSI2-DB3) failed to
repress AGL15 promoter activity when coinfiltrated into N. ben-thamiana leaves with the AGL15pro:LUC reporter, and a HSI2 B3mutant with K279R K314R substitutions failed to rescue thetranscriptional repression of AGL15 in transgenic hsi2-2 plants(Figure 4). These data indicate that the B3 domain is requiredfor HSI2-mediated transcriptional repression in this system.Furthermore, the HSI2 PHD domain is also required for AGL15 re-pressionactivity. Thisoutcomewaspredictedby thederepressionof AGL15 expression and associated loss of H3K27me3 chro-matinmarks in thehsi2-4mutant line (Veerappanetal., 2012,2014)and was confirmed here by coexpression of the HSI2mPHD ef-fector construct with the AGL15pro:LUC reporter in N. ben-thamiana leaves. The PHD domain was reported to be a histonecode reader that recognizes H3K4me3 and H3K27me2/3, andweconfirmed the interaction between HSI2 and thesemethylated H3peptides in vitro (Figure6G;Li et al., 2006,Chakravarty et al., 2009;Zeng et al., 2010; Yuan et al., 2016). Therefore, it is possible thattargeting of HSI2 to the AGL15 promoter requires both DNAbinding activity of the B3 domain and interaction of the PHDdomainwithhistonemethylationmarksassociatedwithchromatinat theAGL15 locus.OurChIP results confirmed that amutant formof HSI2 with a disrupted PHD domain fails to accumulate at the 5ʹregion of the AGL15 locus in vivo (Figure 6). Through RNA se-quencing analysis, Schneider et al. (2016) identified two HSI2splice variants: a full-length form and a truncated form that lackscoding sequences for the PHD domain. Translation of the trun-cated HSI2 transcript would produce a protein variant, similar toHSL2 that contains intact B3, CW, and EAR motifs but lacks thePHD domain (Jia et al., 2014). These authors suggest that thisputative protein variant could still silence target genes by binding
614 The Plant Cell

directly to RY motifs through the B3 domain. However, resultsreportedhere indicate that this truncated formofHSI2 isunlikely tobe an effective repressor of AGL15.
Disruption of the HSI2 EAR motif reduced its ability to repressthe expression of a luciferase reporter gene controlled by thesweet potato Sporamin A1 promoter in Arabidopsis protoplasts(Tsukagoshi et al., 2005). Likewise, coexpressionof theHSI2-DEARconstruct with theAGL15pro:LUC reporter gene inN. benthamianaleaves or stable expression of an HSI2 construct with a pointmutation within the EAR domain in Arabidopsis plants resulted incompromised transcriptional repressor activity (Figure7).However,the activity remained substantially higher than inhsi2-2, B3, or PHDdomain mutants. Thus, while the EAR motif is necessary for fullactivity, it appears toplay a reduced role inHSI2-dependentAGL15transcriptional repression compared with the B3 or PHD domains.Furthermore, expressionof aHSI2mCWconstruct inwhich the fourconserved cysteines were replaced by serine was able to fullyrescueAGL15 repression inArabidopsishsi2-2seedlings (Figure8),indicating that the intact HSI2 CWdomain is not required for HSI2-mediated repression ofAGL15. While theCWdomain of HSL1wasreported to interactwith HDA19 in vivo to downregulate a subset ofseed maturation genes by alteration of histone acetylation at thetarget gene loci during seed germination, HDA19 binding with theHSI2 CW domain was not detected (Zhou et al. 2013).
Todetermine ifsilencingofAGL15 is theprimaryHSI2-dependentpathway for the repression of these seed maturation regulatory
genes, the expression of LAFL genes in plants with altered ex-pression of AGL15 was analyzed (Figure 9). Expression of LAFLgenes was increased in Arabidopsis plants that overexpressAGL15, though not as strongly as in hsi2-2. Expression of LAFLgenes was strongly decreased in hsi2-2 agl15-4 double mutantsrelative to hsi2-2, but remained somewhat higher than inwild-typeor agl15-4 plants. These results provide strong evidence thatHSI2-dependent silencing ofAGL15 is amajor control point in thedownregulation of LAFL gene expression in seedlings, thoughadditional mechanisms appear to be required for full transcrip-tional repression. These results donot support thehypothesis thatHSI2 directly silences LAFLgene expression since, if thiswere thecase, expressionof thesegeneswouldbeexpected to remainhighinhsi2-2 agl15-4doublemutants. Therefore,AGL15 is required forhigh-level expression of LAFL gene in hsi2 seedlings, but it seemslikely that another, as yet unidentified, HSI2 target gene (or genes)is necessary for full derepression of the seed maturation de-velopmental program in this mutant.Since the derepression of AGL15 in hsi2-2 plants is associated
with a lossofH3K27me3marks and thedepositionofH3K4me3atthe silenced gene loci (Veerappan et al., 2014), we predicted thatPRC2 is involved in the HSI2-dependent silencing of this gene.Using directed yeast two-hybrid and coimmunoprecipitationassays, we found that the essential PRC2 component MSI1(Köhler et al., 2003; Schönrock et al., 2006) physically interactswith the HSI2 B3 domain (Figure 10). Analysis of AGL15
Figure 11. Proposed Model for the Repression of Seed Maturation Gene Expression in Arabidopsis Seedlings.
According to thismodel, HSI2 dimers (Chhunet al., 2016)maydirectly target theAGL15promoter bybinding toRYelements andhistonemethylationmarksthrough itsB3andPHDdomains, respectively. HSI2 recruits PRC2by interactionwithMSI1 to repressAGL15 expression throughdeposition ofH3K27me3marks. Additional interactions with MED13 andHDA6, reported by Chhun et al. (2016), are also shown. Since AGL15 positively regulates the expression oftheLAFLgenes thatencode theseedmaturation transcription factorsLEC1,ABI3,FUS3,andLEC2,silencingofAGL15byHSI2 leads to reducedexpressionof these regulatory genes. In hsi2null plants, on the other hand, AGL15 expression remains high and LAFL gene expression is activated, leading to ectopicexpressionof the seedmaturation genetic pathway. Solid lines indicate directly upregulated targets ofAGL15 (Zheng et al., 2009).While our results indicatethat silencingofAGL15 is amajor component ofLAFLgene repression in seedlings, another potential target(s) ofHSI2could alsoplaya role in this regulatorymechanism.
AGL15 Is a Regulatory Target of HSI2/VAL1 615

expression inmsi1-csmutantseedlings, inwhichMSI1expressionis reduced by 95% (Hennig et al., 2003) indicated that MSI1 isrequired for full repression of AGL15 expression in planta. Theseresults suggest two, potentially overlapping, regulatory scenarios.First, HSI2 may be involved in establishing AGL15 silencing byinteracting with H3K4me3 marks associated with the activelytranscribed AGL15 gene to initiate the formation of a repressivechromatin state. Second, HSI2 could be involved in maintenanceof AGL15 silencing by interacting with existing H3K27me3 marksand recruiting PRC2 to add additional silencing-associatedmodifications, thereby reinforcing transcriptional repression. Inanimal systems, PRC2 activity is inhibited by active chromatinmarks such as H3K4me3 and H3K36me3, which block the in-teractionof theH3N-terminal tailwith theNurf55-Su(z)12complex(Schmitges et al., 2011). In Arabidopsis, this interaction can bemodulated by incorporation of the Su(z)12 ortholog VRN2 ratherthan EMF2, which allows PRC2 to autonomously add H3K27me3without overwriting active chromatin domains (Schmitges et al.,2011).
Comparative transcriptomic analysis of hsi2, hsl1, and hsi2 hsl1doublemutants indicated that the transcriptional silencingactivitiesof HSI2 and HSL1 are strongly synergistic (Suzuki et al., 2007).According to those data, expression ofAGL15 in hsi2 hsl1 is 4-foldhigher than in hsi2, indicating that HSL1 is also involved in thetranscriptional silencing of AGL15. A mechanistic explanation forthe functional cooperation between HSI2 and HSL1 in the tran-scriptional silencingofseedmaturationgenesduringgermination isyet tobefully formulated.Theseproteinssharesignificantsequencesimilarity and, while their functions overlap, they are not fully re-dundant (Suzuki et al., 2007; Tsukagoshi et al., 2007). Chhun et al.(2016) reported thatHSI2andHSL1can interactwitheachotherandare predicted to form both homodimers and heterodimers. BothHSI2 and HSL1 are also reported to interact with the TRAP250domain of MED13, a subunit of the MEDIATOR CDK8 repressivemodule.While theCWdomain of HSL1binds specifically toHDA19(Zhouetal.,2013),HSI2 is reported tospecificallybind tothehistonedeacetylaseHDA6 (Chhunetal., 2016).Thus,asshown inFigure11,it seems possible that HSI2 homodimers are targeted to the up-stream sequences of AGL15 by interaction with RY DNA elementsand modified histones, where it recruits PRC2, along with othercorepressors, includingMED-CDK8andHDA6, toformarepressivecomplex. It is also possible that complexes based on HSI2 HSL1heterodimers, that include HDA19, could also form. However, theimportance of the HSL1 B3 and PHD-like domains in targeting thisprotein to AGL15 or other regulated genes has yet to be directlyinvestigated.
METHODS
Plant Materials and Growth Conditions
Arabidopsis (Arabidopsis thaliana) Columbia (Col-0; CS60000) wild-typeand loss-of-function alleles hsi2-2 (SALK_088606) and agl15-4(SALK_076234) were obtained from Arabidopsis Biological ResourcesCenter. Cosuppression linemsi1-cs and 35Spro:AGL15were provided byLars Hennig (Uppsala BioCenter, Swedish University of Agricultural Sci-ences and Linnean Center for Plant Biology) and Donna E. Fernandez(University of Wisconsin, Madison), respectively. WTLUC and hsi2-2LUC,which harbors GSTF8pro:LUC HSI2-responsive reporter gene in the
wild-type and HSI2 T-DNA knockout allele hsi2-2 (SALK_088606) back-ground, respectively, were described previously (Veerappan et al., 2012).For all the experiments, plants were grown under continuous illumination(fluorescent lampsat;200mmolm22 s21) at 24°Con0.3%Phytagel platescontaining 0.53 Murashige and Skoog (MS) salt, 0.5 g/L MES, 13Gamborg vitamin mix, and 1% sucrose (pH adjusted to 5.7).
Plasmid Constructs and Plant Transformation
The HSI2 promoter, consisting of a 1500-bp fragment immediately up-stream of the HSI2 start codon, was amplified by PCR and cloned intopGEMT-Easy vector (Promega). After sequencing, theHSI2 promoter wascleaved by NotI restriction enzyme and inserted into pENTR/D TOPOvector containing theHSI2 cDNA. Then, an LR recombination reactionwasperformed to introduce HSI2pro:HSI2 into the binary vector pEarley-gate301 containing an HA epitope tag at the C terminus. To generate35Spro:MSI1 constructs, pDONOR207 vector containing full-lengthMSI1coding sequence as subcloned into destination vector pGWB521. Ara-bidopsis plants were transformed using the floral dip method (Clough andBent, 1998). The primers used for generating these constructs are listed inSupplemental Data Set 1.
HSI2-HA Complementation
The HSI2pro:HSI2-HA construct was introduced into Agrobacteriumtumefaciens and used to transform hsi2-2LUC Arabidopsis plant throughthe floral dip method (Clough and Bent, 1998). The resulting T1 generationseeds were plated and screened on MS medium containing 5 mg/mLglufosinate (BASTA).Genotypingof resistantplantswasperformedbyPCRusing specific primers. Expression of the HSI2-HA fusion protein wasdetected by immunoblot analysis. Equal amounts of total protein fromWTLUC, hsi2-2LUC, and HSI2pro:HSI2-HA Arabidopsis seedlings wasresolved by SDS-PAGE and electroblotted onto PVDF membranes(Bio-Rad). The processed membranes were probed with anti-HA antibodyto detect HSI2-HA fusion protein or with anti-actin to detect endogenousACTIN for use as a loading standard.
Luciferase Imaging
Imaging of luciferase was performed using Andor iKON-M DU934N-BVCCDcamera (Andor Technology). AndorSOLIS (I) imaging software (AndorTechnology) was used for image acquisition and processing of lumines-cence.Prior to luminescence imaging,Arabidopsis seedlingsgrownonMSplatesand infiltratedNicotianabenthamiana leaveswereuniformly sprayedwith 2mM D-luciferin (Gold Biotechnology) in 0.01%Triton X-100 solution.After spraying with luciferin, seedlings and leaf samples were incubatedunder dark for 5 min. Exposure time for luminescence imaging was 5 min,unless otherwise specified.
RT-qPCR
RT-qPCR was performed using a StepOne Plus system (Applied Bio-systems)with iTAqSYBRGreenSupermixwithROX (Bio-Rad). RNase-freeDNase (Qiagen)-treated total RNA was used for cDNA synthesis usingiScript cDNAsynthesis kit (Bio-Rad). For eachexperiment, cDNAsynthesisreactions were performed on three independent RNA samples preparedfrom approximately five seedlings, each. Three qPCR reactions wereperformed for each cDNA sample. EF1A (AT5G60390) andHYGROMYCINPHOSPHOTRANSFERASE (HPT ) were used as reference genes for Ara-bidopsis and N. benthamiana, respectively. The relative expression ofgeneswascalculated according to theABIPrism7700sequencedetectionsystem (User Bulletin #2; Livak and Schmittgen, 2001). Primer sequencesused for RT-qPCR are listed in Supplemental Data Set 1.
616 The Plant Cell

ChIP Analysis
In transgenic Arabidopsis plants, ChIP assays were performed essentiallyas described (Yamaguchi et al., 2014; Veerappan et al., 2014) with minormodifications. Chromatin was extracted from 7-d-old seedlings grown inMS medium supplemented with 1% sucrose. The chromatin in theseseedlings was cross-linked with 1% formaldehyde. The resulting chro-matinwas sheared to fragmentswith 500 bp (200–1000 bp) average lengthby sonication and used for immunoprecipitation with commerciallyavailable anti-HA (Abcam; ab9110). After reversing the cross-links, im-munoprecipitated DNA was analyzed by qPCR using primers for specificregionsof theLAFLgenesLEC1,ABI3, andFUS3, alongwithAGL15. Threeindependent experiments, each using 500 mg of seedlings (25–30 in-dividual plants), were performed. Three technical replications for eachqPCRassaywere performed, andACT2was used as an internal control fornormalization. In Arabidopsis leaf protoplasts, the ChIP assays wereperformed as described previously (Lee et al., 2007; Du et al., 2009; Xionget al., 2013; Zhang et al., 2014). HSI2pro:HSI2-GFP and HSI2pro:HSI2mB3-GFPDNAwere transformed into hsi2-2Arabidopsis protoplastsfrom 14-d-old leaves, and 35Spro:MSI1-cmyc DNA was transformed intowild-type and hsi2-2 Arabidopsis protoplasts using the polyethyleneglycol-mediated transformation method. Protoplasts were incubated atroom temperature for 12 h under dark conditions. Protoplast chromatinwas cross-linked by 1% formaldehyde in W5 medium for 20 min andquenched with Gly (0.2 M) for 5 min. The protoplasts were then lysed, andtheDNAwas shearedon icewith sonication. The immunoprecipitationwasperformed with anti-GFP (ab290) and anti-myc (MA1-980). After reversingthe cross-links, the purified DNA was analyzed by qPCR using primers forspecific regions of AGL15. Each experiment was repeated at least twiceusing protoplasts from approximately five leaves. Three technical repli-cations for each qPCR assay were performed, and ACT2 was used as aninternal control for normalization. Primers of target genes used for qPCR inChIP analysis are listed in Supplemental Data Set 1.
In Vitro Histone Peptide Binding Assay
Sequencesencoding theHSI2PHDdomain (111aminoacids) andmutatedPHD domain (C65Y) were amplified from the HSI2 full-length cDNA andcloned into pENTR-D-TOPO vector by Gateway cloning system (In-vitrogen). ThepENTR-PHDandpENTR-mPHDplasmidswere recombinedinto thedestination vector pDEST15 (Invitrogen) toproduce fusionproteinswith an N-terminal GST tag. The plasmids were transformed into BL21-Rosetta cells.Escherichia coliwere grown in LBmediumcontaining 50mg/mL carbenicillin to OD600 = 0.6. Expression of recombinant protein wasinduced for 6 h at 28°C with 0.1 mM IPTG. Protein was extracted usingCelLytic B cell lysis buffer (Sigma-Aldrich). Recombinant fusion proteinswere purified by GST binding kit (Novagen).
Peptide binding assays were performed as previously described withminor modification (Lee et al., 2009). Briefly, 1 mg of GST fusion PHD fingerdomains was incubated with 1 mg of various biotinylated histone peptides,H3 (12-403 and 12-404), H3K4me3 (12-564), and H3K27me3 (12-565;Millipore) in bindingbuffer (30mMHEPES, pH7.5, 300mMNaCl, and0.1%[v/v] Nonidet P-40) overnight at 4°C, followed by addition of streptavidinagarosebeads (16-126;Millipore). After1h incubation,beadswerewashedwith binding buffer five times and eluted with 23 SDS sample loadingbuffer. The samples were separated by SDS-PAGE on 10% gels andsubjected to immunoblot analysis using anti-GST.
DNA Binding Assay
AnHSI2cDNAfragmentencodingapeptide(252aminoacids) thatcontainstheB3domainwas cloned into pENTR-D-TOPOvector and then recombined intotheE.coliexpressionvectorpET59-DEST.ExpressionofHis-Trx-B3 inRosettacells was induced with 0.4 mM isopropyl-1-thio-D-galactopyranoside at 37°C
for 3 h. The fusion protein was purified using Dynabeads His-tag isolation andpull-down kit (#10103D; Thermo Fisher) according to the manufacturer’sprotocol and quantified by the Bio-Rad protein assay reagent. DNA probesderived from the AGL15 promoter sequence, with RY and mutated RY ele-ments, were incubated with purified His-Trx or with the His-Trx-B3 fusionprotein. The DNA binding assays were performed as previously described(Wangetal., 2017). Inbrief, 0.3mgHis-B3proteinwasboundtoHis-tagspecificmagnetic beads (Dynabeads; Thermo Fisher) and incubated with 0.25 mMdouble-strandedDNAprobes for15minat roomtemperature.Dynabead-DNAcomplexeswere selectedwith amagnet and, afterwash andelution steps, theamountofprotein-boundprobewasdeterminedusingqPCR.Primersusedarelisted in Supplemental Data Set 1.
EMSA
EMSAwasperformed using double stranded biotinylatedDNAprobes andthe Lightshift Chemiluminescent EMSA kit (Thermo Scientific). Biotin 3ʹend-labeled DNA oligomers were prepared using a biotin end-labeling kit(Thermo Scientific), and double-stranded DNA probes were generated byannealing sense and antisense oligomers. Each 20-mL binding reactioncontained 50 fmol of biotin-labeled double-stranded DNAs, 5 mg of re-combinant His-HSI2B3 protein, and 50 ng/mL poly (dIdC) in binding buffer(10 mM Tris, 30 mM KCl, 0.1 mM EDTA, 1 mM DTT, 0.05% Nonidet P-40,and 6.5%glycerol, pH 7.9). Binding reactions were incubated for 20min atroom temperature resolved by electrophoresis of reaction samples on 5%polyacrylamide gels with TBE buffer. Detection of biotin-labeled DNA wasperformed by chemiluminescence according to the supplier instructions.
Yeast Two-Hybrid Assay
Yeast two-hybrid assays were performed using the Matchmaker GAL4-based two-hybrid system 3 (Clontech) according to the manufacturer’s in-structions. Sequences that encode full-length HSI2were subcloned into thepGADT7 vector, whereas the full-length CLF, EMF2, FIE, and MSI1 codingsequences were subcloned into the pGBKT7 vector. All constructs weretransformed intoyeaststrainAH109bythe lithiumacetatemethod,andyeastcells were grown on a minimal medium/-Leu/-Trp according to the manu-facturer’s instructions (Clontech). Transformed colonies were plated ontoa minimal medium/-Leu/-Trp/-His/-Ade to test for possible interactions.
In Vivo Coimmunoprecipitation
The MSI1 coding sequence was cloned into the binary vector pGWB521-myc. The construct was introduced into Agrobacterium strain GV3101 andtransformed into Arabidopsis hsi2-2 plants that had been complementedwith HSI2pro:HSI2-HA. Total protein was extracted from 9-d-old seedlingsusing lysis buffer (50 mM Tris-HCl, pH7.5, 150 mMNaCl, 1 mM EDTA, 10%glycerol, 2 mM NaVO4, 25 mM b-glycerophosphate, 10 mM NaF, 0.1%Tween20,1mMPMSF,andprotease inhibitorcocktail)and incubated30minat 4°Cwith gentle agitation. After centrifugation at 12,000g for 15min at 4°C,supernatantwas incubatedwithproteinAagarose/salmonspermDNAfor1hfor preclearing. The precleared protein solution was incubated with 50 mLHA-antibody conjugated agarose (Thermo Fisher #26181) overnight at 4°C.The supernatant was removed after centrifugation at 2000g for 2min at 4°C,and the agarose beads were washed three times with lysis buffer. Theproteins were eluted with 23 SDS sample loading buffer and analyzed byimmunoblotting using c-myc tag antibody (Thermo Fisher MA1-980).
Accession Numbers
Sequence data from this article can be found in the Arabidopsis GenomeInitiativeorGenBank/EMBLdatabasesunder theaccessionnumbers listedin Supplemental Table 1.
AGL15 Is a Regulatory Target of HSI2/VAL1 617

Supplemental Data
Supplemental Figure 1. Relative expression of AGL15 in multipleindependent transgenic lines.
Supplemental Table 1. Accession numbers or gene identifiers ofsequences used for multiple sequence alignment.
Supplemental Data Set 1. Sequences of primers used and genesanalyzed in this study.
Supplemental File 1. Statistical analyses.
ACKNOWLEDGMENTS
We thank Million Tadege for providing Gateway-compatible Y2H vectors,LarsHennig for providingmsi1-cs seeds,DonnaE.Fernandez for providingthe35Spro:AGL15seeds, and theArabidopsisBiological ResourceCenterat Ohio State University for providing the TDNA insertionmutants. We alsothank Fei Zhang, Mohamed Fokar, Angelika Reichert, Hui Wang, TezeraWatira, and Tyson Kerr for stimulating discussions and Million Tadege forcritical reading of the manuscript and stimulating discussions. This workwas supported by the Oklahoma Agricultural Experiment Station andOklahoma Center for the Advancement of Science and Technology (GrantPS16-009 to R.D.A.).
AUTHOR CONTRIBUTIONS
N.C. designed and performed the research, analyzed data, and wrote thearticle. V.V., H.A., andM.K. performed research andedited thearticle. R.D.A.designed the research, analyzed data, and edited the article.
Received August 21, 2017; revised January 30, 2018; accepted February20, 2018; published February 23, 2018.
REFERENCES
Baud, S., et al. (2016). Deciphering the molecular mechanisms un-derpinning the transcriptional control of gene expression by mastertranscriptional regulators in Arabidopsis seed. Plant Physiol. 171:1099–1112.
Braybrook, S.A., Stone, S.L., Park, S., Bui, A.Q., Le, B.H., Fischer,R.L., Goldberg, R.B., and Harada, J.J. (2006). Genes directlyregulated by LEAFY COTYLEDON2 provide insight into the controlof embryo maturation and somatic embryogenesis. Proc. Natl.Acad. Sci. USA 103: 3468–3473.
Chakravarty, S., Zeng, L., and Zhou, M.M. (2009). Structure and site-specific recognition of histone H3 by the PHD finger of human au-toimmune regulator. Structure 17: 670–679.
Chanvivattana, Y., Bishopp, A., Schubert, D., Stock, C., Moon, Y.-H.,Sung, Z.R., and Goodrich, J. (2004). Interaction of Polycomb-groupproteins controlling flowering in Arabidopsis. Development 131: 5263–5276.
Chhun, T., Chong, S.Y., Park, B.S., Wong, E.C., Yin, J.L., Kim, M., andChua, N.H. (2016). HSI2 repressor recruits MED13 and HDA6 to down-regulate seed maturation gene expression eirectly during Arabidopsisearly seedling growth. Plant Cell Physiol. 57: 1689–1706.
Clough, S.J., and Bent, A.F. (1998). Floral dip: a simplified method forAgrobacterium-mediated transformation of Arabidopsis thaliana.Plant J. 16: 735–743.
Derkacheva, M., and Hennig, L. (2014). Variations on a theme: Pol-ycomb group proteins in plants. J. Exp. Bot. 65: 2769–2784.
Du, L., Ali, G.S., Simons, K.A., Hou, J., Yang, T., Reddy, A.S., andPoovaiah, B.W. (2009). Ca2+/calmodulin regulates salicylic-acid-mediated plant immunity. Nature 457: 1154–1158.
Finkelstein, R., Reeves, W., Ariizumi, T., and Steber, C. (2008).Molecular aspects of seed dormancy. Annu. Rev. Plant Biol. 59:387–415.
Harding, E.W., Tang, W., Nichols, K.W., Fernandez, D.E., andPerry, S.E. (2003). Expression and maintenance of embryogenicpotential is enhanced through constitutive expression of AGAMOUS-Like 15. Plant Physiol. 133: 653–663.
Heck, G.R., Perry, S.E., Nichols, K.W., and Fernandez, D.E. (1995).AGL15, a MADS domain protein expressed in developing embryos.Plant Cell 7: 1271–1282.
Hennig, L., Taranto, P., Walser, M., Schönrock, N., and Gruissem,W. (2003). Arabidopsis MSI1 is required for epigenetic maintenanceof reproductive development. Development 130: 2555–2565.
Holdsworth, M.J., Bentsink, L., and Soppe, W.J.J. (2008). Molecularnetworks regulating Arabidopsis seed maturation, after-ripening,dormancy and germination. New Phytol. 179: 33–54.
Hoppmann, V., Thorstensen, T., Kristiansen, P.E., Veiseth, S.V.,Rahman, M.A., Finne, K., Aalen, R.B., and Aasland, R. (2011). TheCW domain, a new histone recognition module in chromatin pro-teins. EMBO J. 30: 1939–1952.
Jia, H., McCarty, D.R., and Suzuki, M. (2013). Distinct roles of LAFLnetwork genes in promoting the embryonic seedling fate in theabsence of VAL repression. Plant Physiol. 163: 1293–1305.
Jia, H., Suzuki, M., and McCarty, D.R. (2014). Regulation of the seedto seedling developmental phase transition by the LAFL and VALtranscription factor networks. Wiley Interdiscip. Rev. Dev. Biol. 3:135–145.
Jiang, D., Wang, Y., Wang, Y., and He, Y. (2008). Repression ofFLOWERING LOCUS C and FLOWERING LOCUS T by the Arabi-dopsis Polycomb repressive complex 2 components. PLoS One 3:e3404.
Kagale, S., and Rozwadowski, K. (2011). EAR motif-mediated tran-scriptional repression in plants: an underlying mechanism for epi-genetic regulation of gene expression. Epigenetics 6: 141–146.
Kazan, K. (2006). Negative regulation of defence and stress genes byEAR-motif-containing repressors. Trends Plant Sci. 11: 109–112.
Kinoshita, T., Harada, J.J., Goldberg, R.B., and Fischer, R.L. (2001).Polycomb repression of flowering during early plant development.Proc. Natl. Acad. Sci. USA 98: 14156–14161.
Köhler, C., Hennig, L., Spillane, C., Pien, S., Gruissem, W., andGrossniklaus, U. (2003). The Polycomb-group protein MEDEAregulates seed development by controlling expression of theMADS-box gene PHERES1. Genes Dev. 17: 1540–1553.
Lee, J.H., Yoo, S.J., Park, S.H., Hwang, I., Lee, J.S., and Ahn, J.H.(2007). Role of SVP in the control of flowering time by ambienttemperature in Arabidopsis. Genes Dev. 21: 397–402.
Lee, W.Y., Lee, D., Chung, W.I., and Kwon, C.S. (2009). ArabidopsisING and Alfin1-like protein families localize to the nucleus and bindto H3K4me3/2 via plant homeodomain fingers. Plant J. 58: 511–524.
Li, H., Ilin, S., Wang, W., Duncan, E.M., Wysocka, J., Allis, C.D., andPatel, D.J. (2006). Molecular basis for site-specific read-out of histoneH3K4me3 by the BPTF PHD finger of NURF. Nature 442: 91–95.
Livak, K.J., and Schmittgen, T.D. (2001). Analysis of relative geneexpression data using real-time quantitative PCR and the 2(-DeltaDelta C(T)) Method. Methods 25: 402–408.
Lotan, T., Ohto, M., Yee, K.M., West, M.A., Lo, R., Kwong, R.W.,Yamagishi, K., Fischer, R.L., Goldberg, R.B., and Harada, J.J.
618 The Plant Cell

(1998). Arabidopsis LEAFY COTYLEDON1 is sufficient to induceembryo development in vegetative cells. Cell 93: 1195–1205.
Luerßen, H., Kirik, V., Herrmann, P., and Miséra, S. (1998). FUSCA3encodes a protein with a conserved VP1/ABI3-like B3 domain whichis of functional importance for the regulation of seed maturation inArabidopsis thaliana. Plant J. 15: 755–764.
Mehdi, S., Derkacheva, M., Ramström, M., Kralemann, L.,Bergquist, J., and Hennig, L. (2016). The WD40 domain proteinMSI1 functions in a histone deacetylase complex to fine-tune ab-scisic acid signaling. Plant Cell 28: 42–54.
Mönke, G., Altschmied, L., Tewes, A., Reidt, W., Mock, H.P.,Bäumlein, H., and Conrad, U. (2004). Seed-specific transcriptionfactors ABI3 and FUS3: molecular interaction with DNA. Planta 219:158–166.
Ohta, M., Matsui, K., Hiratsu, K., Shinshi, H., and Ohme-Takagi, M.(2001). Repression domains of class II ERF transcriptional re-pressors share an essential motif for active repression. Plant Cell13: 1959–1968.
Perry, S.E., Nichols, K.W., and Fernandez, D.E. (1996). The MADSdomain protein AGL15 localizes to the nucleus during early stagesof seed development. Plant Cell 8: 1977–1989.
Qüesta, J.I., Song, J., Geraldo, N., An, H., and Dean, C. (2016).Arabidopsis transcriptional repressor VAL1 triggers Polycomb si-lencing at FLC during vernalization. Science 353: 485–488.
Reidt, W., Wohlfarth, T., Ellerström, M., Czihal, A., Tewes, A.,Ezcurra, I., Rask, L., and Bäumlein, H. (2000). Gene regulationduring late embryogenesis: the RY motif of maturation-specificgene promoters is a direct target of the FUS3 gene product. Plant J.21: 401–408.
Rounsley, S.D., Ditta, G.S., and Yanofsky, M.F. (1995). Diverse rolesfor MADS box genes in Arabidopsis development. Plant Cell 7:1259–1269.
Schmitges, F.W., et al. (2011). Histone methylation by PRC2 is in-hibited by active chromatin marks. Mol. Cell 42: 330–341.
Schneider, A., Aghamirzaie, D., Elmarakeby, H., Poudel, A.N., Koo,A.J., Heath, L.S., Grene, R., and Collakova, E. (2016). Potentialtargets of VIVIPAROUS1/ABI3-LIKE1 (VAL1) repression in de-veloping Arabidopsis thaliana embryos. Plant J. 85: 305–319.
Schönrock, N., Exner, V., Probst, A., Gruissem, W., and Hennig, L.(2006). Functional genomic analysis of CAF-1 mutants in Arabi-dopsis thaliana. J. Biol. Chem. 281: 9560–9568.
Stone, S.L., Kwong, L.W., Yee, K.M., Pelletier, J., Lepiniec, L.,Fischer, R.L., Goldberg, R.B., and Harada, J.J. (2001). LEAFYCOTYLEDON2 encodes a B3 domain transcription factor that in-duces embryo development. Proc. Natl. Acad. Sci. USA 98: 11806–11811.
Suzuki, M., Kao, C.Y., and McCarty, D.R. (1997). The conserved B3domain of VIVIPAROUS1 has a cooperative DNA binding activity.Plant Cell 9: 799–807.
Suzuki, M., Wang, H.H., and McCarty, D.R. (2007). Repression of theLEAFY COTYLEDON 1/B3 regulatory network in plant embryo de-velopment by VP1/ABSCISIC ACID INSENSITIVE 3-LIKE B3 genes.Plant Physiol. 143: 902–911.
Suzuki, M., and McCarty, D.R. (2008). Functional symmetry of the B3network controlling seed development. Curr. Opin. Plant Biol. 11:548–553.
Suzuki, M., Wu, S., Li, Q., and McCarty, D.R. (2014). Distinct func-tions of COAR and B3 domains of maize VP1 in induction of ectopicgene expression and plant developmental phenotypes in Arabi-dopsis. Plant Mol. Biol. 85: 179–191.
Thakare, D., Tang, W., Hill, K., and Perry, S.E. (2008). The MADS-domain transcriptional regulator AGAMOUS-LIKE15 promotes so-matic embryo development in Arabidopsis and soybean. PlantPhysiol. 146: 1663–1672.
Tsukagoshi, H., Morikami, A., and Nakamura, K. (2007). Two B3domain transcriptional repressors prevent sugar-inducible expres-sion of seed maturation genes in Arabidopsis seedlings. Proc. Natl.Acad. Sci. USA 104: 2543–2547.
Tsukagoshi, H., Saijo, T., Shibata, D., Morikami, A., and Nakamura,K. (2005). Analysis of a sugar response mutant of Arabidopsisidentified a novel B3 domain protein that functions as an activetranscriptional repressor. Plant Physiol. 138: 675–685.
Veerappan, V., Chen, N., Reichert, A.I., and Allen, R.D. (2014). HSI2/VAL1 PHD-like domain promotes H3K27 trimethylation to repressthe expression of seed maturation genes and complex transgenesin Arabidopsis seedlings. BMC Plant Biol. 14: 293.
Veerappan, V., Wang, J., Kang, M., Lee, J., Tang, Y., Jha, A.K., Shi,H., Palanivelu, R., and Allen, R.D. (2012). A novel HSI2 mutation inArabidopsis affects the PHD-like domain and leads to derepressionof seed-specific gene expression. Planta 236: 1–17.
Wang, H., Niu, L., Fu, C., Meng, Y., Sang, D., Yin, P., Wu, J., Tang,Y., Lu, T., Wang, Z.Y., Tadege, M., and Lin, H. (2017). Over-expression of the WOX gene STENOFOLIA improves biomass yieldand sugar release in transgenic grasses and display altered cyto-kinin homeostasis. PLoS Genet. 13: e1006649.
Xiong, Y., McCormack, M., Li, L., Hall, Q., Xiang, C., and Sheen, J.(2013). Glucose-TOR signalling reprograms the transcriptome andactivates meristems. Nature 496: 181–186.
Yamaguchi, N., Winter, C.M., Wu, M.F., Kwon, C.S., William, D.A.,and Wagner, D. (2014). Protocols: Chromatin immunoprecipitationfrom Arabidopsis tissues. The Arabidopsis Book 12: e0170, doi/10.1199/tab.0170.
Yamasaki, K., et al. (2004). Solution structure of the B3 DNA bindingdomain of the Arabidopsis cold-responsive transcription factorRAV1. Plant Cell 16: 3448–3459.
Yoshida, N., Yanai, Y., Chen, L., Kato, Y., Hiratsuka, J., Miwa, T.,Sung, Z.R., and Takahashi, S. (2001). EMBRYONIC FLOWER2,a novel polycomb group protein homolog, mediates shoot de-velopment and flowering in Arabidopsis. Plant Cell 13: 2471–2481.
Yuan, W., Luo, X., Li, Z., Yang, W., Wang, Y., Liu, R., Du, J., and He,Y. (2016). A cis cold memory element and a trans epigenome readermediate Polycomb silencing of FLC by vernalization in Arabidopsis.Nat. Genet. 48: 1527–1534.
Zeng, L., Zhang, Q., Li, S., Plotnikov, A.N., Walsh, M.J., and Zhou,M.M. (2010). Mechanism and regulation of acetylated histonebinding by the tandem PHD finger of DPF3b. Nature 466: 258–262.
Zhang, F., Wang, Y., Li, G., Tang, Y., Kramer, E.M., and Tadege, M.(2014). STENOFOLIA recruits TOPLESS to repress ASYMMETRICLEAVES2 at the leaf margin and promote leaf blade outgrowth inMedicago truncatula. Plant Cell 26: 650–664.
Zhang, H., Zhang, J., Quan, R., Pan, X., Wan, L., and Huang, R.(2013). EAR motif mutation of rice OsERF3 alters the regulation ofethylene biosynthesis and drought tolerance. Planta 237: 1443–1451.
Zheng, Y., Ren, N., Wang, H., Stromberg, A.J., and Perry, S.E.(2009). Global identification of targets of the Arabidopsis MADSdomain protein AGAMOUS-Like15. Plant Cell 21: 2563–2577.
Zhou, Y., et al. (2013). HISTONE DEACETYLASE19 interacts withHSL1 and participates in the repression of seed maturation genes inArabidopsis seedlings. Plant Cell 25: 134–148.
AGL15 Is a Regulatory Target of HSI2/VAL1 619

DOI 10.1105/tpc.17.00655; originally published online February 23, 2018; 2018;30;600-619Plant Cell
Naichong Chen, Vijaykumar Veerappan, Haggag Abdelmageed, Miyoung Kang and Randy D. AllenVegetative Growth in Arabidopsis
to Regulate the Developmental Transition from Seed Maturation toAGL15HSI2/VAL1 Silences
This information is current as of April 30, 2020
Supplemental Data /content/suppl/2018/02/23/tpc.17.00655.DC1.html
References /content/30/3/600.full.html#ref-list-1
This article cites 58 articles, 28 of which can be accessed free at:
Permissions https://www.copyright.com/ccc/openurl.do?sid=pd_hw1532298X&issn=1532298X&WT.mc_id=pd_hw1532298X
eTOCs http://www.plantcell.org/cgi/alerts/ctmain
Sign up for eTOCs at:
CiteTrack Alerts http://www.plantcell.org/cgi/alerts/ctmain
Sign up for CiteTrack Alerts at:
Subscription Information http://www.aspb.org/publications/subscriptions.cfm
is available at:Plant Physiology and The Plant CellSubscription Information for
ADVANCING THE SCIENCE OF PLANT BIOLOGY © American Society of Plant Biologists




![The B3-Domain Transcription Factor VAL1 Regulates the ...The B3-Domain Transcription Factor VAL1 Regulates the Floral Transition by RepressingFLOWERING LOCUS T1[OPEN] Yanjun Jing,a,2](https://img.pdfslide.us/doc/110x75/5ff5108b9228e622882d84c3/the-b3-domain-transcription-factor-val1-regulates-the-the-b3-domain-transcription.jpg)