Embed Size (px)
Citation preview

How can biotechnology help with biofuel production?
Changing the plant:
greater yield (photosynthesis) domestication of partially domesticated plants (e.g. architecture, making plants more easily harvested)
Changing the biomass components for easier processing: ratio of cellulose to lignin; structure of lignin; structure of cellulose; incorporating cellulase; new triglycerides that yield better biodiesel.
Selecting new microorganisms (Synthetic genomics): efficiency of pentose fermentation; combining cellulose hydrolysis and fermentation; new fermentation end-products insoluble in water.
Producing better enzymes:
gene shuffling technologies

What regulatory issues should be kept in mind?
• Plant improvement will undoubtedly include genetic modification (engineering)
• Many people will object to industrial traits in GMO food crops (corn, soybeans) because accidental presence of these seeds in food cannot be avoided (no matter how hard we try).
• Biomass crops (switchgrass, miscanthus, poplar, eucalyptus etc) must be carefully considered before financial resources are invested. Poplar has excellent genomic resources, but produces pollen that travels far and has many wild relatives; switchgrass has few genomic resources but is a perennial that can be harvested before it flowers.
• Ensuring fail-safe pollen sterility will be crucial to securing regulatory approval and public acceptance.

How much solar radiation can be converted to biomass?
How can this be improved? How much?




C3 and C4 photosynthesis
C3 plants fix CO2 by the Calvin- Benson pathway in the mesophyll cells by Rubisco. The substrates are ribulose bisphosphate and CO2 gas dissolved in the chloroplast. The product is a 3-carbon compound (3-phosphoglycerate).
C4 plants fix CO2 first in the mesophyll cells by PEP carboxylase. The substrates are phosphoenolpyruvate and HCO3. The product, malate or another 4-carbon compound), is transported to the bundle sheath cells were CO2 is first released and then fixed again by the Calvin-Benson pathway. The equilibrium of CO2 in H2O favors HCO3, and PEP carboxylase cannot react with O2. At the low concentrations of CO2 in the mesophyll, C4 photosynthesis is more efficient than C3.
C4 plants have increased water andnitrogen use efficiencies.
C3
C4

The Calvin cycle for CO2 assimilation has 3 important parts
Rubisco

Rubisco can catalyze carboxylation and
oxygenation of ribulose bisphosphate (RuBP) Which reaction takes place
depends on the concentration of the two gasses at the site of the reaction in the chloroplast.
When photosynthesis evolved 3 billion years ago, the atmosphere was CO2 rich and O2 poor. Now it is the reverse.
The enzyme evolved to increase its discrimination (specificity) but this came at the expense of the turnover rate (only 3 reaction per second). Rubisco is the laziest enzyme in the world!
Carboxylation
Oxygenation
C4 plants concentrate CO2 at the site of Rubisco carboxylation, therebyminimizing the oxygenation reaction.

In C3 plants, photorespiration recycles the C in PO4-glycolate produced in the oxygenation reaction of Rubisco to glycerate with the release of 1/4 of
the C as CO2. The reactions involve 3 different organelles.
This is called photo-respiration becauseit only occurs in the light (when Rubisco is active), releasesCO2 (in the mito=chondria) and usesO2 (in the peroxi-somes)



Sedoheptulose-1,7-bisphosphatase (SBPase)
Faster regeneration of ribulose bisphosphate causes more rapid photosynthesis resulting in bigger plants


Creating a bypass for photorespiration in the chloroplast using five bacterial genes.
WT
D+E+F
D+E+F+G+T
D,E and F = 3 subunits of bacterial glycolate dehydrogenase; G+T = glycolateCarboligase and tartonic semialdehyde reductase

Enhanced biomass yield by genetic modification of growth processes
poplar
control control
Arabidopsis
Mendel
The transgene is a transcription factor from Mendel, a Bay Area biotech company. Will this hold up in the field?

Lignin and biofuel production
• Lignin has negative impacts on biofuel prodution– lignin impedes access of hydrolytic enzymes to wall
polysaccharides– lignin adsorbs hydrolytic enzymes– lignin interferes with pretreatment processes– lignin degradation products inhibit fermentation– lignin is not fermentable (but its energy can be utilized)
• Lignin is essential for plant viability; all the genes ar known
• Can reducing lignin content, composition, and tissue specificity improve biomass quality? Plants seem to tolerate substantial lignin modification.

Lignin modification may decrease the need for pretreatment

Effect of lignin content on enzymatic recovery of sugars from Miscanthus
D Vrije et al (2002) Int J Hydrogen Energy 27,1381
Lignin
Switchgrasscomposition
cellulose
Hemi cellulose

Lignin biosynthetic pathway in woody angiosperms(e.g. aspen or poplar)

Lignin biotechnology: Antisense CAD and COMT genes in poplars
Red color shows differenttype of lignin has beenproduced.
Kraft pulping of tree trunks showed that the reduced-CAD lines had improved characteristics, allowing easier de-lignification, using smaller amounts of chemicals, while yielding more high-quality pulp. This work highlights the potential of engineering wood quality for more environmentally benign papermaking without interfering with tree growth or fitness. Nature Biotechnology 20, 607 - 612 (2002)

To determine relationships between lignin content/composition and chemical/enzymatic saccharification, we analyzed previously generated alfalfa lines expressing antisense constructs for downregulating lignin biosynthesis independently at six different steps: cinnamate 4-hydroxylase (C4H); hydroxycinnamoyl CoA: shikimate hydroxycinnamoyl
transferase (HCT); coumaroyl shikimate 3-hydroxylase (C3H); caffeoyl CoA 3-O-methyltransferase (CCoAOMT); ferulate 5-hydroxylase (F5H); or caffeic acid 3-O-
methyltransferase (COMT) Dixon lab. Nature Biotechnology 25, 759 - 761 (2007) w/o acid pretreatment
after acid pretreatment

Cleavable Lignin Concept
OHOH OH
OHO
O
OO
OHO
OO
OHO
O O
OH
OO
OH
OO O
OH
OH
OO O
OH
Simplified degradation
Modified synthesis
Normal precursor
Novel precursor
ester Rosemarinic acid = ester

Plants have 120 different ABC transporters in the PM, vacuolar membrane, peroxisomes and
mitochondria. What is their specificity? How are they targetted?
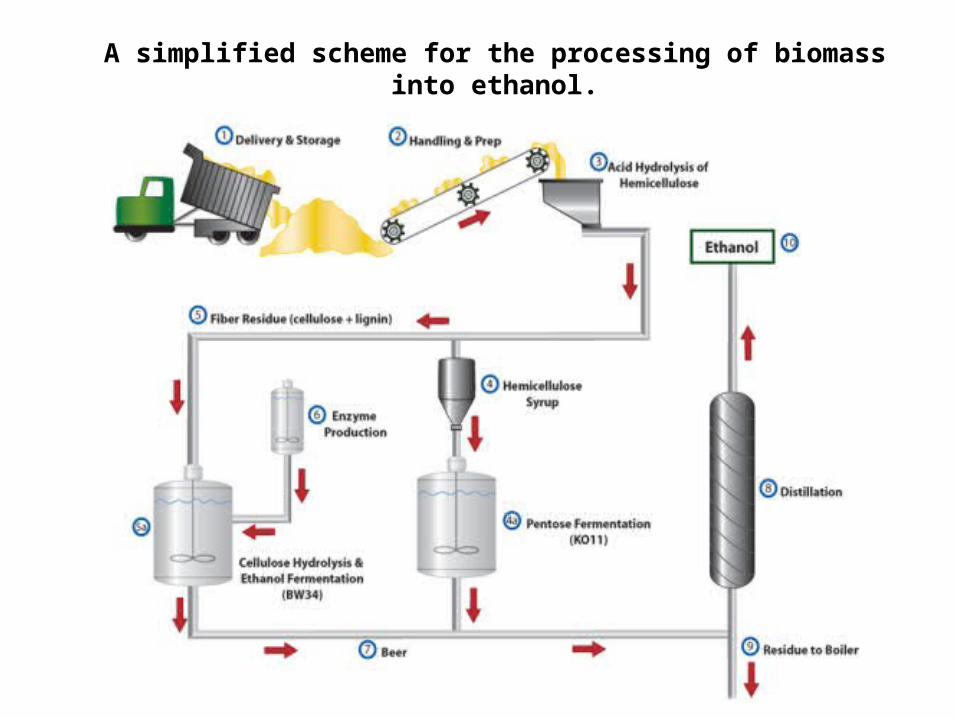
A simplified scheme for the processing of biomass into ethanol.

Efficiency of biomass conversion
Dry stover 1 ton (1000 kg)
Cellulose content × 0.45
Cellulose conversion and recovery efficiency × 0.76 (too high?)
Ethanol stoichiometric yield × 0.51
Glucose fermentation efficiency × 0.75
Yield from glucose 131 kg ethanol = 151 L (40 gallons)
Dry stover 1 ton (1000 kg)
Hemicellulose content × 0.29
Hemicellulose conversion and recovery efficiency × 0.90
Ethanol stoichiometric yield × 0.51
Xylose fermentation efficiency × 0.50
Yield from xylose 66 kg ethanol = 76 L (20 gallons)

Getting the sugars out of ligno-cellulose: acid versus enzyme
• Acid hydrolysis (cheap)Pentoses are released more quickly than hexoses. Use 0.5 %
sulfuric, 180 C for 5 to 10 minutes. For cellulose hydrolysis: 1% sulfuric acid (dilute) for 12 seconds at
237 C in a continuous flow reactor. If you heat longer you get byproducts of acid breakdown like furfural, which inhibit fermentation later on. Process is rapid but yield is only 50 % of all the glucose.
Enzymatic hydrolysis (still expensive) Lignocellulose must be made into fine pellets if the enzymes are to
have access to the cellulose molecules. Multiple cellulolytic enzymes are needed. Hemicellulases are relatively inefficient.
.

Looking at Nature: Organisms has evolved 2 ways of degrading cellulose. In both cases the cellulase enzymes
have cellulose binding domains (CDBs)
• Aerobic degradation by white rot fungi (e.g. Trichoderma) and aerobic bacteria. This requires endoglucanases that cleave glycosidc bonds in the amorphous part of the molecule creating reducing and non-reducing ends. Exoglucanases (cellobiohydrolases) that release cellobiose (a disaccharide) and cellobiase to release glucose from cellobiose.
• Anaerobic degradation. Best studied is the cellulosome of Clostridium thermocellum. A multiprotein complex anchored on the surface of the bacterium by non-catalytic proteins. This bacterium can also ferment glucose to ethanol.

White rot fungi degrade biomass aerobically with
cellulases, hemicellulases and
ligninases.
Degradation requires endoglucanases that
Cleave glycosidic bonds in the amorphous
part of the molecule creating reducing and
non-reducing ends. Exoglucanases
(cellobiohydrolases) that release cellobiose
(a disaccharide) and cellobiase to release
glucose from cellobiose.


Cellulose binding domains have 3 aromatic amino acid residues (e.g. tyrosine or tryptophan) that form a planar
structure able to bind to a cellulose sheet.
Different CDBs target different sites on the cellulose surface. They (1) recognize specific sites, (2) bring the catalytic site of the enzyme in close proximity to the cellulose, and (3) pull the cellulose molecule out of the sheet so the glycosidc bond is accessible.
By making chimeric proteins it is possible to determine the specificity of different CBDs.

Cellulose binding modules/domains are primarily found on fungal cellobiohydrolases and are
essential for the breakdown of crystalline cullulose.
• Plant endoglucanases generally do not have CDBs A: plasma membrane anchored with large extracellular domain. B: secreted C: secreted but with a cellulose binding domain at the C-terminus.

This “crust” fungus does not make mushrooms but crust-
like fruiting bodies underneath wood pieces or logs
The 30-million base-pair genome of Phanerochaete chrysosporium strain RP78 has been sequenced using a whole genome shotgun approach. The P. chrysosporium genome reveals an impressive array of genes encoding
secreted oxidases, peroxidases and hydrolytic enzymes that cooperate in wood decay.

Lignin degradation is an enzyme-mediated oxidation, involving the initial transfer of single electrons to the intact lignin. Electrons are transferred to other parts of the molecule in uncontrolled chain reactions, leading to breakdown of the polymer.
* lignin peroxidase (previously known as ligninase). This is an iron-containing enzyme which accepts two electrons from hydrogen peroxide (H2O2), then passes them as single electrons to the lignin molecule. * manganese peroxidase, which acts in a similar way to lignin peroxidase but oxidizes manganese (from H2O2) as an intermediate in the transfer of electrons to lignin. * laccase, a phenol oxidase which directly oxidizes the lignin molecule * several hydrogen-peroxide-generating enzymes - for example glucose oxidase which generates H2O2 from glucose (a product of cellulose or hemicellulose breakdown).

Schematic presentation of the cellulosome of Clostridium thermocellum: CipA protein is drawn in green, catalytical components in orange, cell-wall binding components in blue, the bacterial cell in brown and the cellulose microfibril in black. The S-layer is punctuated. The cohesin modules are
numbered 1 to 9. The binding of the enzymes to specific positions (cohesins) is hypothetical, as is the linear orientation of the scaffoldin. The
scaffoldins (CipA) bound to OlpB are only sketched partially. All cellulosome components are not drawn to scale.
QuickTime™ and aTIFF (Uncompressed) decompressor
are needed to see this picture.

Different approaches to enzyme production for cellulose digestion
1. Culture one or more microorganisms on chips of the material to be degraded, so that they produce the right mixture of enzymes.
2. Isolate cellulase genes (and other genes) and by directed evolution select the genes that make heat stable enzymes with high catalytic activity.
3. Select microorganisms and optimize their activities so that they digest and ferment lignocellulose under high temperature and anaerobic conditions.

Pichia stipitis, closely related to the yeast symbionts living in in the hindgut of passalid beetles that degrade white-
rotted wood, are able to efficiently convert pentose in ethanol.
QuickTime™ and aTIFF (Uncompressed) decompressor
are needed to see this picture.

The abundance of the transcripts in the basic
metabolic pathways of Pichia stipitis under different growth
conditions.GA = glucose aerobicXA = xylose aerobic
GOL = glucose oxygen limitedXAL = xylose oxygen limited

How can pentoses be fermented?
• The pentose phosphate pathway can convert 3 pentose phosphates into 2 fructose-6-phosphates and 1 glyceraldehyde-3-P (GAP) which can both be used in glycolysis. However, F6P can also be oxidized in the oxidative part of the cycle to generate ribulose-5-P and NADPH. To make things go better you have to inhibit this part of the cycle and also take away G-3-P. This requires two bits of genetic engineering: overexpression of GDP1 to reduce GAP and elimination of G6P dehydrogenase (zwf1).
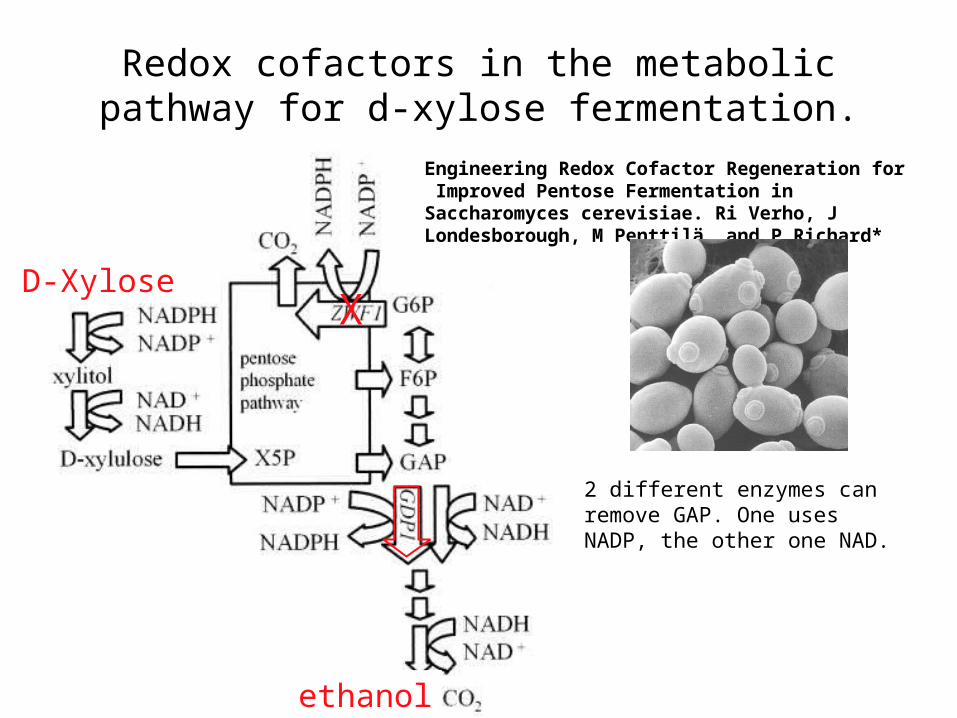
Redox cofactors in the metabolic pathway for d-xylose fermentation.
Engineering Redox Cofactor Regeneration for Improved Pentose Fermentation in Saccharomyces cerevisiae. Ri Verho, J Londesborough, M Penttilä, and P Richard*
X
ethanol
D-Xylose
2 different enzymes can remove GAP. One uses NADP, the other one NAD.

Genetic engineering of yeast: knock out zwf and overexpress GDP1. This results
in better pentose fermentation.

How can pentoses be fermented?
• In certain cell types the pentose phosphate pathway is used to generate reducing power from glucose. This produces pentoses. To enhance fermentation of pentoses this part of the pathway has to be knocked out by eliminating G-6-P dehydrogenase.

This ABE (acetone, butanol, ethanol) fermentation process yields butanol, acetone, and ethanol in a ratio of 6:3:1, respectively. For each bushel of corn, then, the yield would be 1.3 gallons of butanol, 0.65 gallons of acetone, and 0.22 gallons of ethanol, each at a concentration of 1-2%. This ratio bodes well for butanol, but, with the use of yeast in the fermentation process rather than bacteria, ethanol has emerged as the most viable fuel. For the same bushel of corn, yeast fermentation yields 2.5 gallons of ethanol with a concentration of 10-15%. It was this great difference that likely established ethanol as the premier alcoholic fuel source. Butanol has the advantage that it is more like gasoline (energy density and air/fuel ratio needed) but it is more viscous. A joint venture of BP and DuPont is developing butanol as a transportation fuel.
Clostridium acetobutylicum

Part II The End

2H+ + Dred → H2 + Dox Photo-bioreactors with green algae or
cyanobacteria are used to test different strains for the ability to produce hydrogen gas.
Hydrogen gas is itself a renewable fuel, but it also finds many applications in industry including the manufacture of ammonia fertilizer, hydrogenated
fats and gasoline. Hydrogenases in green photosynthetic organisms are sensitive to oxygen,
and algae produce little or no hydrogen without genetic intervention and manipulation of the
culture.
Certain fermentative anaerobic bacteria, like Clostridium pasteurianum, produce hydrogen gas by transferring electrons from reduced ferredoxin
to protons when they catabolize sugars. This hydrogen does not leave the cells but is used as a
source of energy.

• d-Xylose is converted to d-xylulose through an NADPH-utilizing reductase and NAD+-utilizing dehydrogenase. d-Xylulose is then
phosphorylated to the pentose phosphate intermediate d-xylulose 5-phosphate (X5P). The products of the pentose phosphate pathway
are fructose-6-phosphate (F6P) and GAP. GAP is reduced through an NADP-GAPDH, encoded by GDP1, or by the endogenous NAD-
GAPDH, depending on cofactor availability. In the following reactions involving an NADH requiring alcohol dehydrogenase, equimolar
amounts of CO2 and ethanol are derived. A competing pathway for NADP+ is the oxidative part of the pentose phosphate pathway.
Glucose-6-phosphate (G6P) is derived from fructose-6-phosphate and can enter the oxidative part of the pentose phosphate pathway through G6PDH, which is encoded by the ZWF1 gene. G6P is
oxidized, thereby generating NADPH and CO2. The deletion of ZWF1 prevents this reaction.

Lignin biosynthetic pathway in woody angiosperms(e.g. aspen or poplar)
Secretion is followed by oxidativepolymerization catalyzed by peroxidase
<--- Phenylalanine

The crystalline regions of cellulose have intramolecular and intermolecular hydrogen bonds, allowing the linear glucan chains to form crystalline structures that exclude water and
enzymes.
Intramolecular:The H of the OH on C3 to the O that makes the glycosidicbonds.Intermolecular: The H of the OH on C6 to the O of the OH on C3. These are the bonds that make the very tight structure of cellulose microfibrils. Microfibrils have 30-40 chains each with 2000 to 10,000 glucose units.


Can we improve cellulose digestion?
cellulose
cellulaseCellulase B
New cellulase
cellulose
cellulase
New cellulose
glucan
cellulase
cellulose
cellulase

Pilot plant for the anaerobic fermentation of biomass into mixed alcohol fuel by a mixture of microorganisms.





























