Embed Size (px)
Citation preview

HOST LARVAL MORTALITY IN AN EXPERIMENTAL HOST-PARASITE POPULATION
T . BURNETT Research Institute, Canada Department qf Agriculture, Bellenrille, Ontario
Received April 8, 1964
Abstract T w o populations of T r i a k ~ r d e s ~itpnrrrrimrrnt (\Vwta..) and its chalcid para-
$te Encnrsin jnrmoso Gahiun wcrc reared oil tomato pl:t~tts in the . eenhor lse a t I 2-76 "F lor 2h weeks. .4Ithough the ahunclanrr o l bnth q)ecies Aurtu;~ted with peaks or increasing xrnplilude, the popula~ior~ that wn.i ir~i t ially l a r ~ c r rei~iainetl so thro~rghaut the period of sampling hecatrse rhr pnrasi te inflicted ~ i n ~ i l a r ratrs of mmtality En both c;lses. 'The fluctuations nf rhe twn separate poptilatioris were syncbronixcd throuahout the 1)criad of propa~;atinn. H n ~ t rnnrlalil y , which scsrlTtctl either from nlrnnst immediate killinr: ni host scales foIlnwing irttadc hy atlull parasites or from tleatll of host larv:lr fallowinq par;l+itimtion and tlevelop- ment of parasite proqenv, was tlrt~rmined by parasitr density, hnst s i~e , and po- 4 ldy by a uuml~er of other factors su<:h ns thcaxc structtlrc of hnsz larval popul:i- tintla, age of adult par;lhite~, and sucr.tllt-ncc oi leaves 011 which tltc hosi larvae develaprl. The interaction CII host a ~ ~ r l parasitc prodl~cetl r>*cles in the age strtlctilrps of host anrl l,,>rasiie poptllatir~n.; ~li;rt, in turn, influencerl the inter- aciinn of the tx.o species. l'hr ~ l r a t h of hnst Inrvae fol!o%~ill~ attack by adult parasites was a lorn1 o l host p r o t ~ c ~ i o l ~ . a9 i t ensurerl thc r:tpid rleclinc in ihe ahu~iclnncc of t h r parasite popi~li~tion nr~d rrras, t h ~ r c l u r ~ , the prirnilry [actor in the mairltennnce of the host-parnitr syster~l.
As the chaIcid parasite Rncnrsifi jormosa Gahan (falnily Et~lophidae, order Mymenoptera) is, r~ncler rertain circumstnnccs. an important agent in the control ol the greenhouse whitefly. 7i-tale?1rod~.r m ~ ~ o r a r i o r ~ ~ m (Westw.) (Family AIyerodidae, ordw Hornoptera), it might bc employed to better advan taga i f the various processes in thc in terartion between host and parasite populatinns were undcrstond. I n addition, propagation of the cornpletel?t freely-interacting populations with nverlapping generations should, together with a number nl other host-parasite svstems. csti~l~lish q~~nIitatively many of t h e processes thal: occur hehvccn twn trapl~ic levels of a natural community and therecore provide reliable postulates lor the interpretation of some natural events. For both practical and scientific reasons, therefore, a series of T. vnfiornriorztm - E. fbrmosn populations were propagated in the greenhouse and details of the interaction hetween t he species recordetl (Burnett 1%0a, b).
When T. vaporariorzrm and E. jorpnosa were reared in the greenhouse on tamato plants, at 72-76 O F , lor 8 months, t h e numbers of adults of both species fluctuated with peaks of increasing amplitude (Burnett 196Oaj. Although the fluctuations ol populations were in phase, thcre was much variation in the magnitude of peak numbers in thosf: pairs of populations which were con- nected by Iasgc openine in the glass partition separating them and hetween which, therelore, adults of host and parasite couId disperse readily. Because a number of population curves were ohserver1 in the preliminary experiments, the causes of variations in t h e fluctuations of adtdt: host and parasite popuIa- tions were analyzed before the details of the interaction in a given population were investigated. The average densities of host and parasite populations
Canadian Journal of Zoology. Volume 42 (1964)
Can
. J. Z
ool.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
MIC
HIG
AN
ST
AT
E U
NIV
ER
SIT
Y o
n 12
/05/
14Fo
r pe
rson
al u
se o
nly.

746 CANADIAN JOURNAL OF ZOOLOGY. VOL. 42, 1964
were related to their initial densities; the higher the initial densities of host and parasite, the higher the subsequent densities. The magnitudes of the fluctuations were determined by the periods of infestation of the two species, i.e., by the degree of synchronization between host and parasite populations a t the beginning of the experiment. Within a given population, however, the fluctuations in the numbers of adult hosts, and of adult parasites, were similar in magnitude and phase a t each sampling point (Burnett 1960b). These obser- vations suggested that in adjacent and contemporaneous populations the parasite destroyed a constant portion of immature hosts in populations of different sizes, that the age structures of host and parasite populations were important factors in the interaction, and that dispersal and dispersion of the two species had little effect on the incidence of host mortality.
A distinction between the importance of the ability of E. formosa adults to find hosts and alternative factors in determining the fluctuations in abundance of host and parasite may be made by propagating two well-synchronized host- parasite populations of initially different densities. The significance of differ- ence between initial densities should be apparent in the subsequent numbers of adult hosts and parasites. Variation in host mortality may be compared both between and within populations and mortality may be related to varia- tion in age structure of host and parasite populations and to variation in numbers of adult parasites.
This paper deals with the types and extent of mortality among immature stages of T. vaporariorum when populations of T. vaporariorum and its para- site E. formosa are reared in the greenhouse a t 72-76 O F .
Notes on the Biology of Host and Parasite Species Although the biology of E. formosa was reported in some detail by Speyer
(1927) and the parasite has been reared continuously since 1929 a t the Research Institute, Belleville, some aspects of its relation to its host needed further investigation for an interpretation of parasite-induced mortality of immature hosts. Accordingly, a number of supplementary experiments were conducted to determine the range of certain processes in the host-parasite interaction.
( a ) Parasitism of Hosts i n Relation to Parasite Density All four stages of host larvae are attacked by the parasite adults and either
are almost immediately killed and turn brown or are parasitized and eventually die and turn black as the single parasite larva matures inside each scale. Whether a host is successfully parasitized or killed is influenced by the density of adult parasites and by the size of the host attacked; hosts tend to be killed a t high parasite densities but to be parasitized a t lower densities; smaller hosts tend to be killed but larger ones to be parasitized (Burnett 1962). The increase in percentage of hosts parasitized as parasite density was decreased was observed only over a narrow range of parasite density, from one to four host larvae per adult parasite. One would expect that as parasite density is decreased still further, a density would be reached a t which percentage parasit- ism would begin to decline. Therefore, to determine if the maximum degree of parasitism occurs a t a relatively low or high level, the range of hosts per adult parasite was extended further than in the previous experiments.
Can
. J. Z
ool.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
MIC
HIG
AN
ST
AT
E U
NIV
ER
SIT
Y o
n 12
/05/
14Fo
r pe
rson
al u
se o
nly.
BURNETT: HOST-PARASITE POPULATION 747
Method Medium-sized host larvae (late second and third instar) were exposed on
tomato plants for 18 hours to adults of E. formosa a t ratios of 1 ,3 ,6 ,9 , 12, and 15 hosts per parasite. The number of hosts exposed during the summer months a t naturally varying temperatures ranged in the 16 replicates of each host- parasite ratio from 160 to over 600. After the parasites were removed from the tomato plants, the host larvae were reared until all black scales were formed and then the numbers of hosts parasitized were recorded. The numbers of hosts that were almost immediately killed were also recorded; as the results confirmed the previous observation that killing of hosts occurs a t high para- site densities, they are not presented.
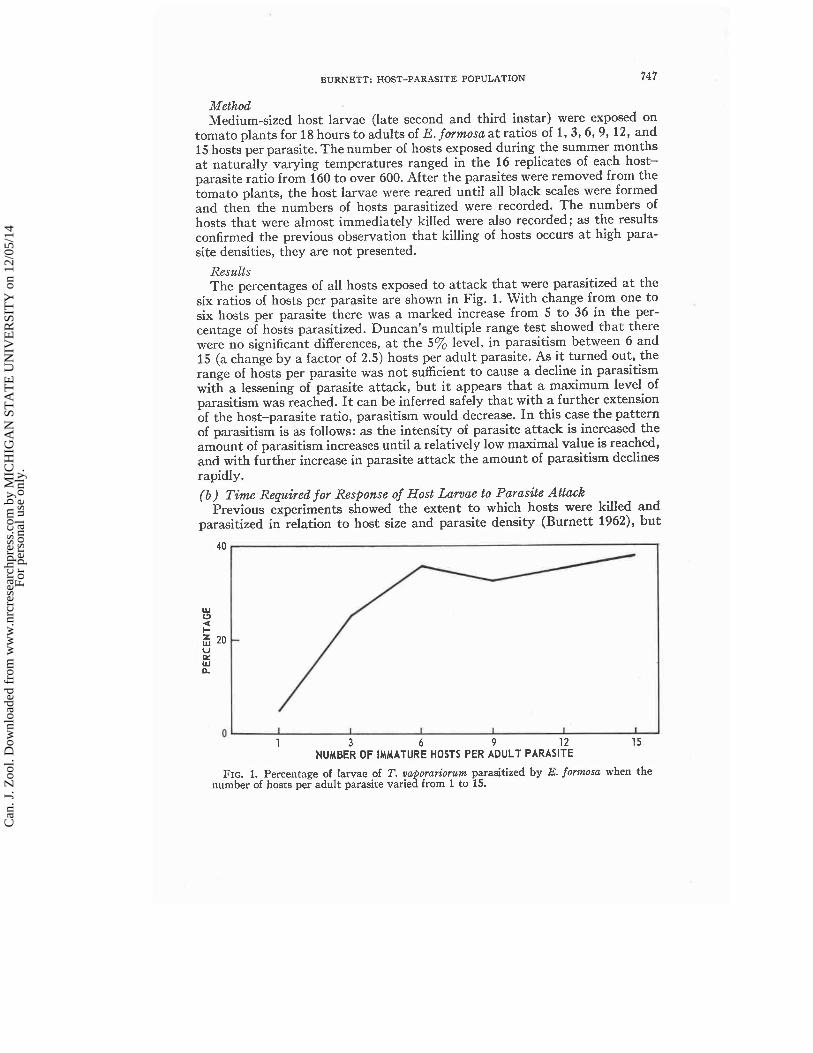
Results The percentages of all hosts exposed to attack that were parasitized a t the
six ratios of hosts per parasite are shown in Fig. 1. With change from one to six hosts per parasite there was a marked increase from 5 to 36 in the per- centage of hosts parasitized. Duncan's multiple range test showed that there were no significant differences, a t the 5% level, in parasitism between 6 and 15 (a change by a factor of 2.5) hosts per adult parasite. As i t turned out, the range of hosts per parasite was not sufficient to cause a decline in parasitism with a lessening of parasite attack, but i t appears that a maximum level of parasitism was reached. I t can be inferred safely that with a further extension of the host-parasite ratio, parasitism would decrease. In this case the pattern of parasitism is as follows: as the intensity of parasite attack is increased the amount of parasitism increases until a relatively low maximal value is reached, and with further increase in parasite attack the amount of parasitism declines rapidly.
( b ) Time Required for Response of Host Larvae to Parasite Attack Previous experiments showed the extent to which hosts were killed and
parasitized in relation to host size and parasite density (Burnett 1962), but
I I 1 I I 1
1 3 6 9 12 15 NUMBER OF IMMATURE HOSTS PER ADULT PARASITE
FIG. 1. Percentage of larvae of T. vaporariorum parasitized by E. formosa when the number of hosts per adult parasite varied from 1 to 15.
Can
. J. Z
ool.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
MIC
HIG
AN
ST
AT
E U
NIV
ER
SIT
Y o
n 12
/05/
14Fo
r pe
rson
al u
se o
nly.

748 CANADIAN JOURNAL OF ZOOLOGY. VOL. 42, 1964
they did not indicate the time necessary for the two types of mortality to become apparent following attack by adult parasites. As size of host larva was a factor in determining the percentage of hosts killed, it may also have influenced the rate of dying, i.e., the larger the host, the longer i t survived after attack. Also, as the size of host attacked may modify the rate a t which parasite larvae develop (Takahashi 1957), the rate of development of E. formosa larvae may depend, to some extent, on whether the adults oviposit in small, medium, or large host scales. The two aspects of the response of host larvae to parasite attack were examined in the following experiment.
Method The responses of different sizes of host larvae to parasite attack were
determined by separately exposing small, medium, and large larvae infesting tomato plants to adults of E. formosa a t a ratio of one parasite to four hosts for 18 hours a t naturally varying temperatures. There were four replicates, the numbers of hosts exposed ranging from 255 to 393. The numbers of small, medium, and large hosts that died and the number of black scales formed were recorded daily.
Results Because the experiments were conducted a t naturally varying temperatures,
there was some variation among the replicates but conditions were comparable within each experiment. Nearly all the small larvae were found dead when the parasites were removed from the plants and many of the medium and large larvae had also perished. By the end of 48 hours all of the deaths among the three size classes of larvae attributable to adult parasite attack had occurred. By the 10th day after attack a second form of death resulted from the un- successful development of the parasite larvae; as shown in Table I, from 3 t o 28% of all the hosts exposed to parasite attack died along with the develop- ing parasite larvae they contained. More large than small hosts died from this second type of mortality because fewer of the former were killed in the original parasite attack. The median time for development of black scales is shown in Table I to range from 7 to 12 days and to vary with the size of host para- sitized. Thus, the smaller the host larva, the sooner it died and the longer i t took for the development of black scales following parasite attack.
( 6 ) Rate of Development of Host and Parasite The duration of the various stages of the whitefly was estimated by observ-
ing, individually, the development of large numbers of hosts in a population reared in the greenhouse a t 72-76 O F . As it was not possible to identify the four larval stages of T. vaporariorum during the sampling of the experimental
TABLE I Effects of host size on development of parasite larvae
yo hosts killed by unsuccessful No. days from parasitization to Larval size development of parasite larvae black scale formation
Small 3-1 1 11-12 Medium 1-12 9-10 Large 21-28 7-8
Can
. J. Z
ool.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
MIC
HIG
AN
ST
AT
E U
NIV
ER
SIT
Y o
n 12
/05/
14Fo
r pe
rson
al u
se o
nly.

BURNETT: HOST-PARASITE POPULATION 749
populations, the scales of the host were classified arbitrarily as small, medium, and large. There was considerable variability in the duration of each stage of the host insect, some of which may be attributed to variation among the host plants, but average durations were estimated as follows: egg stage, 5 or 6 days; small larval stage, 5 days; medium larval stage, 8 days; large larval stage, 2 days; pupal stage, 4 days; and total life cycle, about 25 days.
In populations reared a t 72-76 OF the average time from formation of black scales to emergence of the adult female parasite was 10 days. But as the period from parasitization to formation of black scales is affected by the size of the host parasitized, there is considerable variation in the developmental period of E. formosa. However, when the rate of development of the parasite was studied a t a series of constant temperatures from 65 to 86 O F , the period from parasitization of third and fourth larval instars to formation of black scales was about one-half of the total developmental period (Burnett 1949). At 72-76 OF, therefore, this period is approximately 9 or 10 days for medium larvae and the length of development of the parasite 19 or 20 days.
Methods Two populations were reared in adjacent 14 by 17 ft sections of a greenhouse
in a temperature range of 72-76 O F . Each population was propagated on 64 tomato plants arranged in 16 groups of 4 plants each. The first plant in each group of four was placed in its position 2 weeks before the second plant, 4 weeks before the third plant, and 6 weeks before the fourth plant. Each plant had five or six leaves when placed in the section. In 4 weeks i t had about 12 leaves and after this time all further new growth was removed. Eight weeks after introduction into the section, the plant was placed under the greenhouse benches for 2 additional weeks to permit the emergence of any adult parasites from black scales on its leaves and i t was replaced by a new plant. By this procedure one-quarter of the plants in each section were renewed every 2 weeks and, as the whitefly usually deposits its eggs on new foliage, satisfactory conditions for the reproduction of the host insect were maintained.
The first population, called population R, was started by infesting the plants with 80 adults of T. vaporariorum on each of 10 occasions (a total of 800 adults) between the 1st and 33rd day and releasing 64 adults of E. formosa on 10 occasions (a total of 640 adults) from the 15th to 47th day of the ex- periment. The infestation schedule of the second population, population S, was the same as that of the first, except that three times as many adults of the host and of the parasite were released a t each infestation.
Sampling of Adult Hosts and Parasites For sampling purposes, the 64 plants in each greenhouse section were
divided into four lots, each lot containing four groups of 4 plants each. All the adults of T. vaporariorum and of E. formosa were counted without dis- turbance, twice a week, on the 4 plants of one group chosen randomly from each lot of 16 plants. By this technique, the variations in the abundance of host and parasite adults were estimated, at four sampling points, by counting the adults on one-quarter of the host plants.
Can
. J. Z
ool.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
MIC
HIG
AN
ST
AT
E U
NIV
ER
SIT
Y o
n 12
/05/
14Fo
r pe
rson
al u
se o
nly.

750 CANADIAN JOURNAL OF ZOOLOGY. VOL. 42. 1964
Sampling of Immature Stages of T . vaporariorum For the sampling of the immature stages, a second group of 4 plants was
chosen randomly from each lot of 16 plants. All the immature stages of T. vaflorariorum were counted, twice a week, by means of a binocular micro- scope on four leaflets of each of the four plants in a group. On each plant, two of the leaflets were on one leaf and the other two were on the leaf immediately above the first. The data for each leaflet were recorded separately. The numbers of yellow and black eggs, of living (white) and dead (brown) larvae, of living and dead pupae, and of black scales were counted on each of the 64 leaflets. The four larval stages of T. vaporariorum were divided arbitrarily into small (first and early second instars), medium (late second and third instars), and large (fourth instar) size classes. In addition, the numbers of adult hosts and parasites found on each leaflet were recorded.
Estimates Derived from Sampling Records The numbers of hosts killed between sampling periods were taken as the
differences between the numbers of dead hosts on two consecutive counts. Some dead hosts, particularly small ones, eventually drop off the leaf; thus the mortality could be underestimated. As the adult whiteflies oviposit on a given leaflet for about a week only, however, the increase in the numbers of dead scales can be checked, after 10 days, against the decrease in the numbers of living ones.
Usually, host mortality is estimated from the numbers of hosts exposed to the parasite a t one period and the numbers killed within a following period. In the present case, however, as small host larvae died soon after attack, many hatched from eggs, settled down on the leaflets, and were attacked and killed before they were observed. Thus the numbers of hosts counted as dead in one sampling period were sometimes greater than the numbers recorded as exposed to attack a t the previous sampling period. Therefore, the numbers of hosts exposed to the parasites between two sampling periods were estimated to be the numbers of living scales counted a t the second period plus the numbers killed between the two periods. All the living scales were not exposed for the full 3- or 4-day period, as many small larvae had eclosed 1 or 2 days before counting.
The increase in the number of black scales between two sampling periods gave the number of hosts successfully parasitized 9 or 10 days before, three counts previously.
The emergence of whiteflies was estimated from the numbers of pupae formed rather than from the numbers of empty shells from which the adults emerged, because in these experiments the empty shells fell off the leaflets after a short period. The numbers of black scales formed and the numbers of pupae killed were subtracted from the numbers of pupae in the previous sampling period. The remaining values were halved because each pupa was counted usually a t two consecutive sampling periods.
Neither yellow (1-day-old) nor black eggs proved to be satisfactory stages for an estimation of the reproductive rate of the host. Yellow eggs were not observed a t many sampling periods; usually they were seen when the adults were numerous. The stalked eggs were,easily counted but the empty shells
Can
. J. Z
ool.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
MIC
HIG
AN
ST
AT
E U
NIV
ER
SIT
Y o
n 12
/05/
14Fo
r pe
rson
al u
se o
nly.
NU
MB
ERS
(IN
THO
USA
ND
S)
OF
AD
ULT
H
OST
S AN
D PA
RA
SITE
S IN
SA
MPL
E
Can
. J. Z
ool.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
MIC
HIG
AN
ST
AT
E U
NIV
ER
SIT
Y o
n 12
/05/
14Fo
r pe
rson
al u
se o
nly.
752 CANADIAN JOURNAL OF ZOOLOGY. VOL. 42. 1964
remained in place on the leaf after the larvae had eclosed. In addition, the numbers of adult hosts counted on the sampling leaflets varied greatly from one sampling period to the next. In this case, the number of adult hosts re- corded on thc 64 tomato plants nt one sampling period was divided into the number of small larvae rccorded thrce periods lntcr to give a measure of host reprmluction. As the size of the: sample For adults was much larger than that for the immature stageses, as small larvae were often counted twice (at two consecutive smnplinp periods), and as the sex ratio of the whitefly is approxi- mately one to one, th i s measure is only an index ol the reproductive rate b u t it can be used to compare reprocluctioiz between sarnplin~ periods.
The number of adult parasites (all iemales) rccorded on the 64 tomato plants a t one sampling period was divided into the number 01 immature hosts (either total number or number of small, medium, and large larvae) recorded a t the same period to give an index of the intensity of parasite attack on larval hosts. The gcneral trcnd of an index obtained by averaging two consecutive counts of adult numbers differs veq- little from the one, based on a singIe count, cmploycd.
Populations of Different Densities ( a ) Numbers of Adults
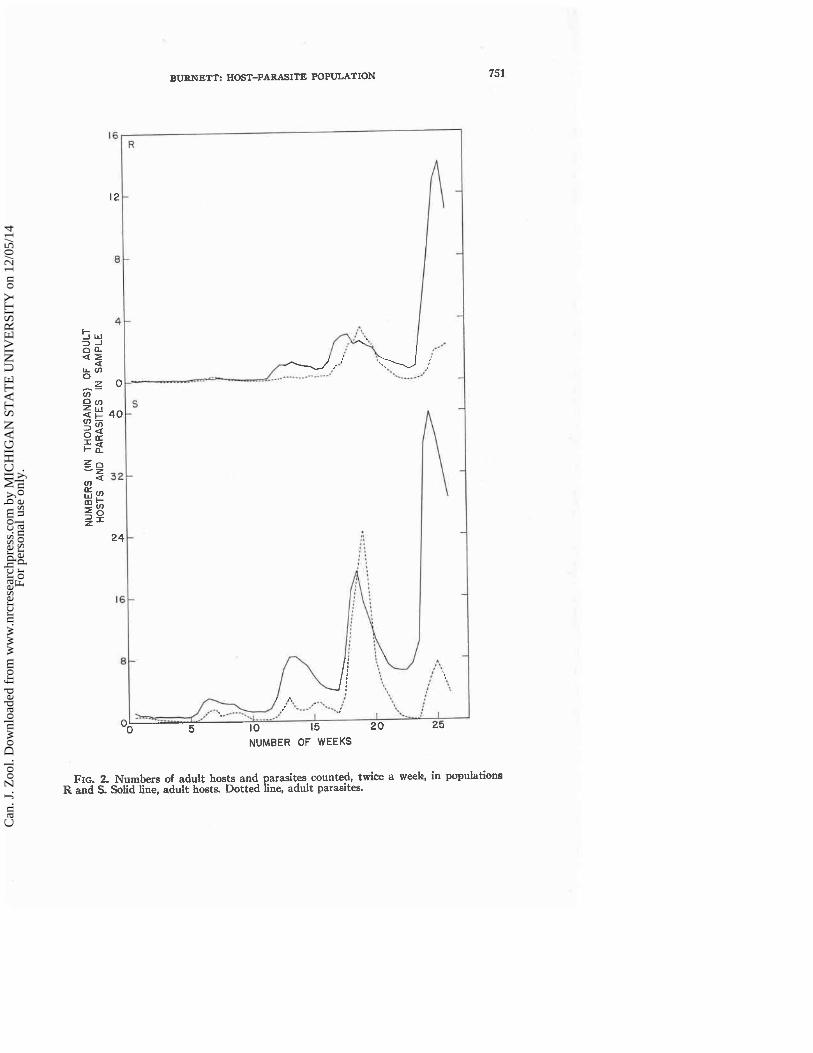
The numbers of adults of T. vaporariorum and of E. formosa counted twice weekly for 26 weeks in populations R and S, after the infestation programs were completed, are shown in Fig. 2. The variation in the numbers of adult hosts followed the usual pattern of fluctuations with increasingly greater
10 15 20 25 WEEKS
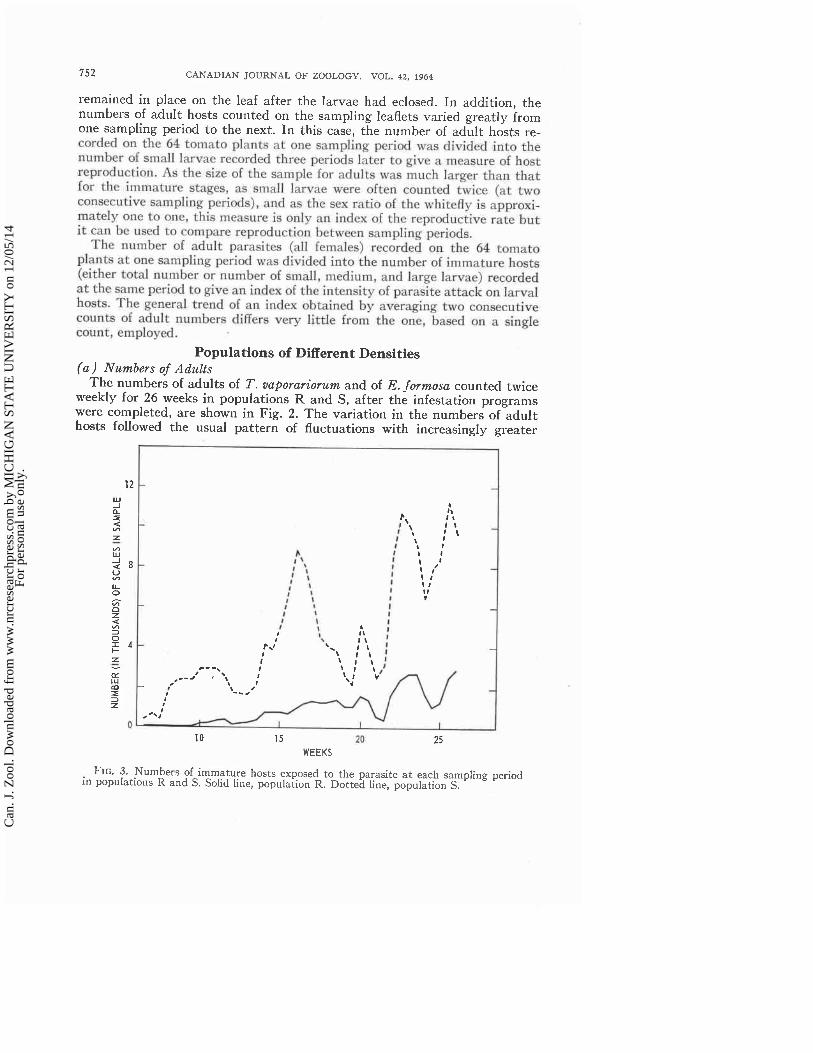
FIG. 3. Numbers of immature hosts exposed to the parasite a t each sampling period in populatioils R and S. Solid line, population R. Dotted line, population S.
Can
. J. Z
ool.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
MIC
HIG
AN
ST
AT
E U
NIV
ER
SIT
Y o
n 12
/05/
14Fo
r pe
rson
al u
se o
nly.
BURNETT: HOST-PARASITE POPULATION 753
maximal and minimal vaIues. The increases and decreases of the parasite were related to thosc of the host, bu t the numbers of adult parasites dropped very quickly to a very low Ievcl follou,ing the maximum near the 20th week. The fluctuations occurred at the same t ime in hoth populations indicating tha t thc rates of development were similar in hoth cases. The numbers rccorded in population S were usuallv about three times as great as those in population R a t comparable sampling periods. And within both populations the magni- tude and phase of t he fluctun~ions in host and parasite numbers were similar at each of the four sampling points.
( b ) LarvaJ Mortality The numbers of larvae of T. vaporariorum cnunted on the 64 Ieaflets at each
sampling period in populations R and S are shnwn in Fig. 3. As some tirnc was required to develop the sampling technique for the i r n m a t u ~ hosts, the counts of living and dead scales of T. aaporarior~lm hegan on the sixth week after
NO. OF WEEKS
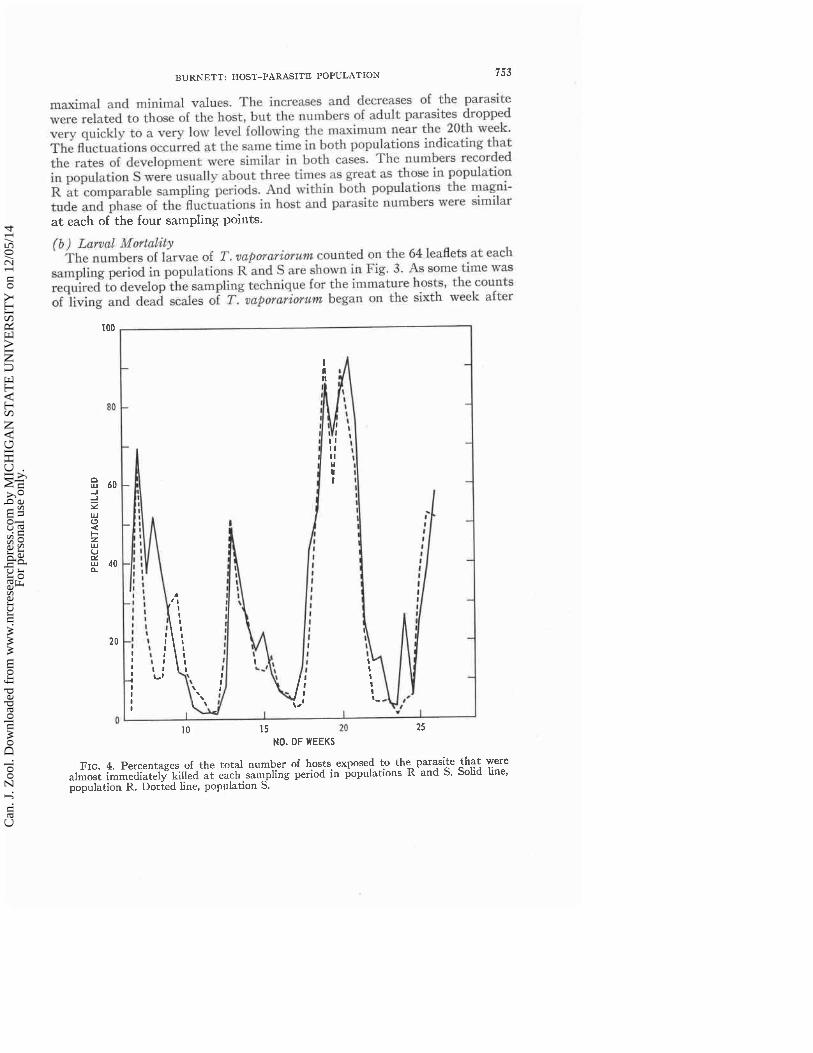
FIG. 4. Percentages of the total number of hosts exposed to the parasite that were almost immediately killed a t each sampling period in populations R and S. Solid line, population R. Dotted line, population S.
Can
. J. Z
ool.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
MIC
HIG
AN
ST
AT
E U
NIV
ER
SIT
Y o
n 12
/05/
14Fo
r pe
rson
al u
se o
nly.
754 CANADIAN JOURNAL OF ZOOLOGY. VOL. 42. 1964
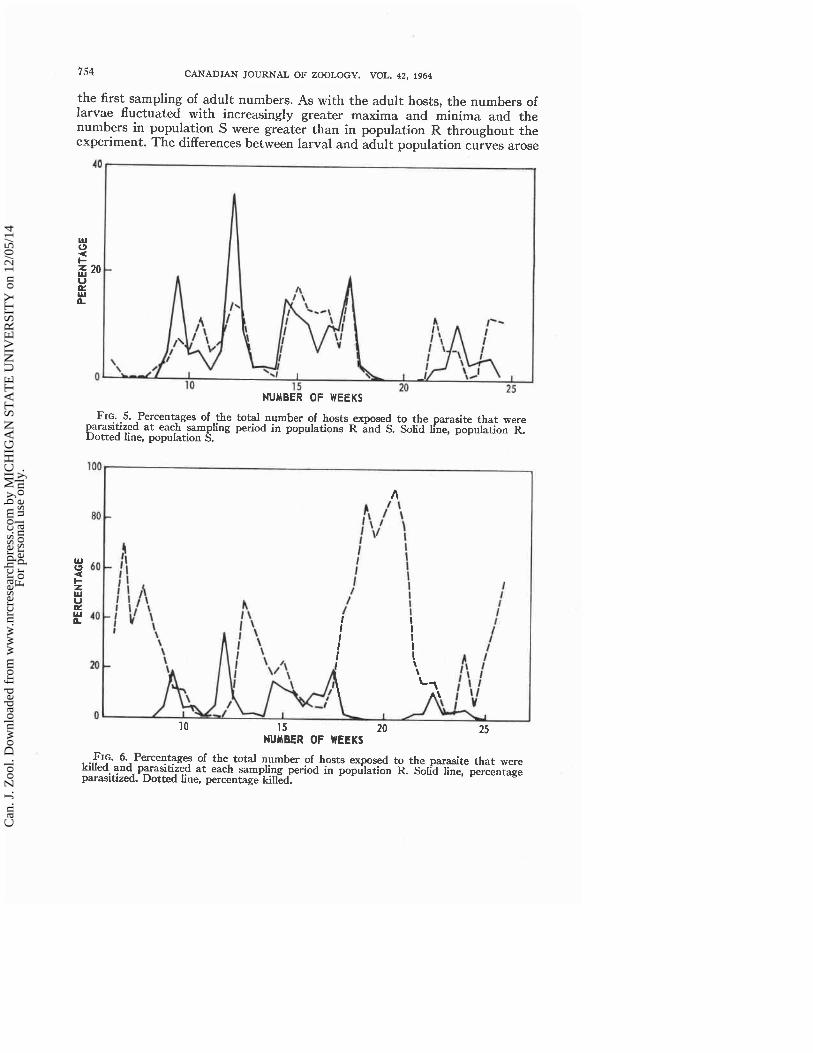
the first sampling of adult numbers. As with the adult hosts, the numbers of larvae fluctuated with increasingly greater maxima and minima and the numbers in population S were greater than in population R throughout the experiment. The differences between larval and adult population curves arose
FIG. .5. Percentages of the total number of hosts exposed to the parasite that were parashzed a t each sampling period in populations R and S. Solid line, population R. Dotted line, population S.
FIG. 6. Percentages of the total number of hosts exposed to the parasite that were kiied and parasitized a t each sampling period in population R. Solid line, percentage parasitized. Dotted line, percentage killed.
Can
. J. Z
ool.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
MIC
HIG
AN
ST
AT
E U
NIV
ER
SIT
Y o
n 12
/05/
14Fo
r pe
rson
al u
se o
nly.

BURNETT: HOST-PARASITE POPULATION 755
from the differences in the duration of each stage as well as from variation in mortality of the immature hosts.
The percentages of all the larval hosts exposed to adult parasite attack in each population that were almost immediately killed a t half-weekly intervals are shown in Fig. 4 and the percentages of all hosts exposed that were parasi- tized are shown in Fig. 5. The percentages killed in populations R and S are nearly identical a t any given time and the percentages parasitized in the two populations are very similar to each other. Also, the fluctuations in per- centages of hosts killed and in percentages of hosts parasitized were replicated, with some slight variation, a t each of the four sampling points within each population.
Figure 6 shows a comparison of the percentages of larval hosts killed with those parasitized in population R throughout the sampling period. Many more hosts were killed than were parasitized; the total percentage mortality of the host larvae was not usually much greater than that of hosts killed. For the most part, parasitism occurred only when the killing of host scales by adult parasite attack was a t a low level.
Aspects of the Interaction between Host and Parasite These experiments were specifically designed to compare variation in
mortality between two populations of different densities. Despite a number of limitations (such as the seasonal and daily variations in the physical environ- ment, difference in size of samples of adult and immature populations, and discontinuities in the sampling of time segments of the developing immature populations because of gaps between one set of four leaflets on a host plant and a second set of four leaflets on the next youngest plant), the experiments yield additional information on the interaction of T. vaporariorum and E. formosa.
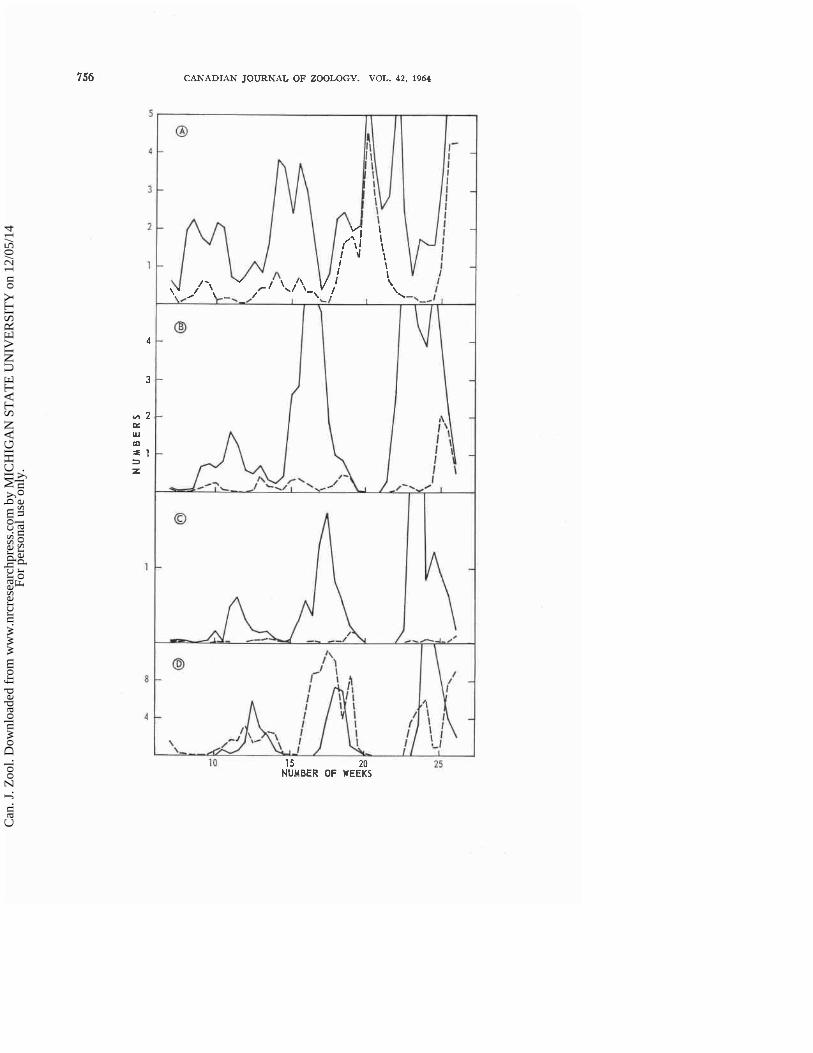
( a ) Mortality of Immature Hosts The numbers of small, medium, and large larval hosts exposed to and
killed by the parasite adults are shown in Fig. 7, A, B, and C respectively for the period of sampling of population S. The numbers of small, medium, and large larvae exposed to the parasite and the numbers of small and medium larvae killed by adult attack fluctuated with peaks of increasing amplitude. The decrease in numbers of small larvae a t the peak of each fluctuation (weeks 10, 15, and 22) was caused by the sampling technique; adult hosts were chang- ing to new foliage that was located (in time) between the sampling leaflets. The medium larvae appear a t times to have been more abundant than the small larvae from which they developed, but this resulted from the fact that individuals spent more time as medium than as small scales. And fewer large larvae are shown in Fig. 7, C than were exposed to the parasites because some passed through this stage between sampling periods. There were large differ- ences in the numbers of small, medium, and large larvae almost immediately killed: of all hosts killed, 76% were small, 22% were medium, and 2% were large. Some of the large larvae died from the unsuccessful development of parasite larvae rather than from attack by adult parasites.
Can
. J. Z
ool.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
MIC
HIG
AN
ST
AT
E U
NIV
ER
SIT
Y o
n 12
/05/
14Fo
r pe
rson
al u
se o
nly.
CANADIAN JOURNAL OF ZOOLOGY. VOL. 42, 1964
Can
. J. Z
ool.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
MIC
HIG
AN
ST
AT
E U
NIV
ER
SIT
Y o
n 12
/05/
14Fo
r pe
rson
al u
se o
nly.

BURNETT: HOST-PARASITE POPULATION 757
Figure 7, D shows the numbers of black scales that were formed and the numbers of hosts tha t were estimated to emerge on the sampling leaflets between satnpling perids. 'The numbers af hlack scales fluctuated and accur- ately predicted the abundance ni adult parasites emerging 9 or 10 days later, except for the marked discrepancy in t h e 18th meek. The fluctuations in the appearance of new black scales in the 13th, lPth, and 25th weeks cannot be attributed to errors of sampling because the decrease ancl increase in the numbers of black scales rorined occurred mostly on the same set of leaflets, An indication of the jncidencc nf parasitization and of killing of host larvae is given by the fact that 10,776 blacli scales were formed on the sampled leaflets throughout the sampling period, whereas 9,450 medium-sized srales were killecl. The estimate ol the emergence of adult hosts predicted closely the phasing and relative abundance of the variation in numbers of adult T. tuporarkrlrn.
Fi'qre T indicates t h a t the parasitization and the kitling of the various stages of T. vapotariarum lollowed a more or less well-defined sequence. This sequence is best illustrated in relation to the fluctuation jn numbers of adult hosts by comparing Figs. 2 and 7. The host scales were parasitized when t11e numbers of adults were at the end of a decline in one fluctuation and beginning t o increase in numbers in the following fluctuation. A11 three larval stages could have been parasitized at this time because some of each were exposed; but most of the hosts were medium and large forms at this period in the fluctuation of immature stages and the majority of black scales probably dcveIoped from these groups. Following thc successful parasi tization oE the scales, first medium and then large larvae were Icilled. The combination of parasitization and killing of medium and large host larvae resulted in the cessation of increase in numbers of adult hosts. Immediately after the host aduIts reached their maximum abundance in a fluctuation the small larvae were killed, and with further decline in lumbers of adult hosts the medium- sizecl host larvae were killecl. After this, parasitization began and the cycle of mortality started again. In these experiments, there were two periods when medium scales were killed and one period each far small and large hosts during each flucbtation in thc numbers ol adult hosts. As parasitization and killing 01 immature Ilosts can occor simultanenusly, there was a gndation from one aspect of larval mortality tn the other rather than successive cliscretc periods of parasitis~n and death of small, medium, and large hosts.
( b ) Age Structures of Host and Parasite Populations Figures 2 and 7, A, B, and C show that the abundance of each stage of T.
vaporariorum increased and decreased a t more or less regular intervals, that the peaks of abundance of one stage followed those of the next youngest stage,
FIG. 7. A. Numbers, in thousands, of small Inrvae of T. wfioru~orua exposed to and killed by adults of E. formosa on sample lcnfl~ts of pnpulntion S. ?%!id line, numbers exposer!, Dolled line, rlun~hers killed. H. Nutrrhers, i l l thousands, oi mediunl lsrvac exposed and killed. Solid tine. rlurnbcrs expn~erl. Unt ted line, nrlni hers killcd. C. Zliirnl~ers, iri thot~sa~lrls, o I large 1.lrvne cxposd and killccl, Snlicl Iit~e, numbers exposed. Dol.terl line, n ~ ~ r n l ~ e r s killed. D. Numbers, i r k hunrlrrtls, of I~lack scatcc fortrlecl and nf xtlulc hosts esti- rnatcd to havc en~ergerl in popt~lntior~ 5. Solid line. adirlt hosts. Dotted line, black scales.
Can
. J. Z
ool.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
MIC
HIG
AN
ST
AT
E U
NIV
ER
SIT
Y o
n 12
/05/
14Fo
r pe
rson
al u
se o
nly.
758 CANADIAN JOURNAL OF ZOOLOGY. VOL. 42, 1964
and that killing and parasitization of immature hosts followed a well-defined sequence. I t would appear, therefore, that the compositions of the host and parasite populations varied in a regular manner as each species passed through fluctuations in abundance.
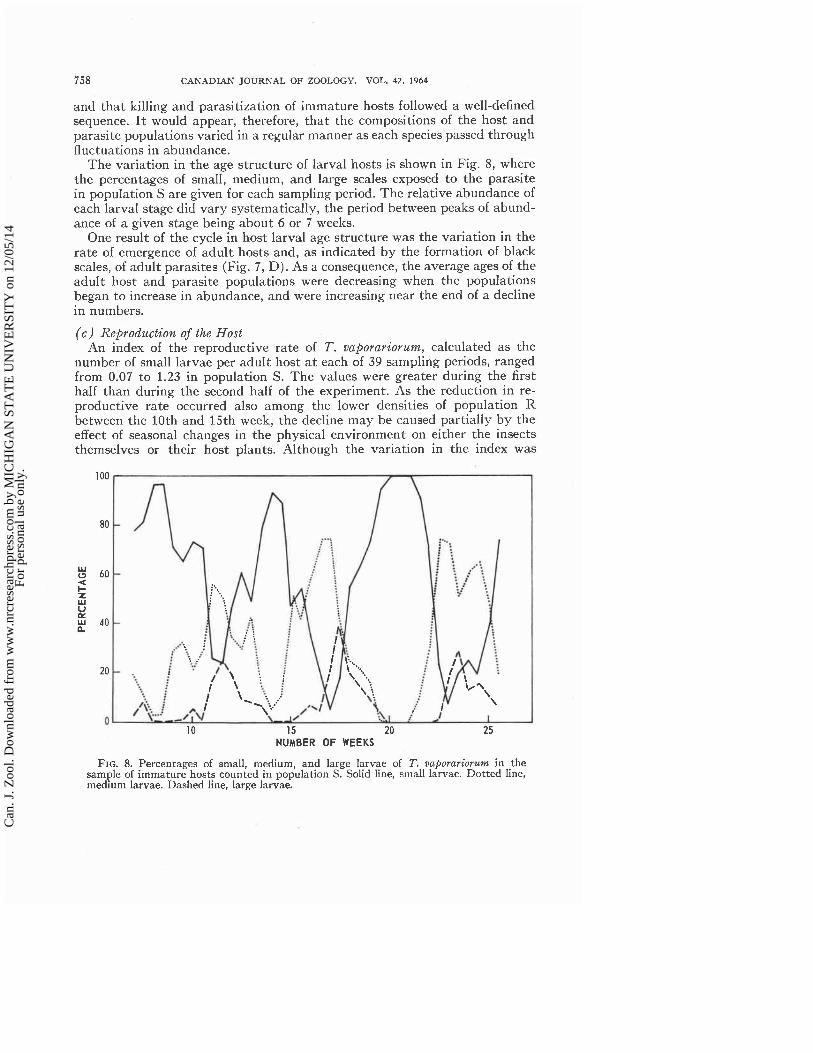
The variation in the age structure of larval hosts is shown in Fig. 8, where the percentages of small, medium, and large scales exposed to the parasite in population S are given for each sampling period. The relative abundance of each larval stage did vary systematically, the period between peaks of abund- ance of a given stage being about 6 or 7 weeks.
One result of the cycle in host larval age structure was the variation in the rate of emergence of adult hosts and, as indicated by the formation of black scales, of adult parasites (Fig. 7, D). As a consequence, the average ages of the adult host and parasite populations were decreasing when the populations began to increase in abundance, and were increasing near the end of a decline in numbers.
(c) Reproduction of the Host An index of the reproductive rate of T. vaporariorurn, calculated as the
number of small larvae per adult host a t each of 39 sampling periods, ranged from 0.07 to 1.23 in population S. The values were greater during the first half than during the second half of the experiment. As the reduction in re- productive rate occurred also among the lower densities of population R between the 10th and 15th week, the decline may be caused partially by the effect of seasonal changes in the physical environment on either the insects themselves or their host plants. Although the variation in the index was
NUMBER OF WEEKS
FIG. 8. Percentages of small, medium, and large larvae of T. vaporariorum in the sample of immature hosts counted in population S. Solid line, small larvae. Dotted line, medium larvae. Dashed line, large larvae.
Can
. J. Z
ool.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
MIC
HIG
AN
ST
AT
E U
NIV
ER
SIT
Y o
n 12
/05/
14Fo
r pe
rson
al u
se o
nly.

BURNETT: HOST-PARASITE POPULATION 759
very great, low values occurred near the end of a decline in numbers of adult hosts; possibly, as females aged their reproductive rate declined.'
( d ) Factors Affecting Mortality of Host Larval Populations As supplementary experiments (Burnett 1962) showed that the percentages
of host larvae almost immediately killed and the percentages parasitized were influenced by the numbers of adult parasites per immature host and by the size of the larvae attacked, the effects of these two variables on host mortality were examined in population S.
Killing of Host Larvae Figure 7 shows that greater proportions of the small, medium, and large
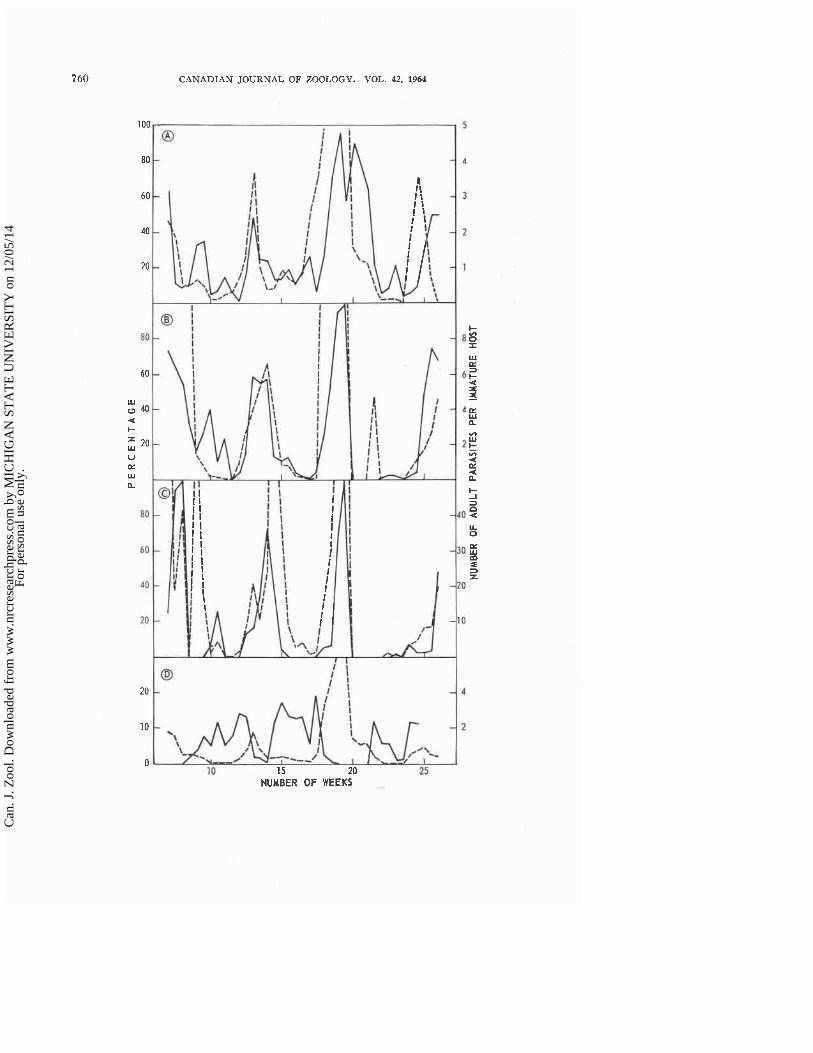
larvae exposed to the parasite were killed on some occasions than on others. This variation is indicated in Fig. 9, A, B, and C, which shows, respectively, the percentages of small, medium, and large larvae killed in population S a t each sampling period. I t is apparent from a comparison of Figs. 7 and 9 that, although both numbers and percentages of hosts killed varied within each fluctuation of the abundance of immature hosts, the ~ e a k values of these two measures of mortality did not occur a t the same timi. There were only slight differences in the times when the percentages of small, medium, and large larvae killed began to increase and reached their peaks, as shown in Fig. 9. But, as this variation was taking place in a population of changing age structure (Fig. 8), the maximal percentages for small larvae occurred when the relative abundance of this stage was near a maximum, whereas the maximal per- centages for medium and large larvae occurred when the relative abundance of these later stages was approaching minimal values. I t is unlikely that the fluctuations of percentages killed, in contrast to those of numbers killed, were of increasing amplitude; the first fluctuations of small and medium larvae were not complete but the first fluctuation of large larvae, which occurred later than those of the first two stages, was as high as any subsequent peak. At some sampling periods all the hosts exposed to the parasite were killed, whereas a t others less than 5y0 of the hosts died.
The relation between the percentages of small, medium, and large larval hosts killed and the intensity of parasite attack, as measured by the numbers of adult parasites per immature host, is also shown in Fig. 9, A, B, and C. As the counts of adult parasites on the sample leaflets varied greatly from one census to the next, the regular sampling of adult parasites was used as a measure of parasite abundance. In this connection, the values obtained in the supplementary experiments, reported above, do not apply directly to the experimental populations; in the supplementary experiments the ratio of adult parasites to host larvae varied from 1: 1 to 1 : 4 or from 1: 1 to 1: 15 for an 18-hour period, whereas in the population experiments the ratio on the sample leaflets varied from 1 : 18 to 1 : 3600 and the parasites were in contact with the host scales continually. Figure 9 shows that in the experimental populations parasite density was a factor influencing host mortality because, for each larval size, the percentages of hosts killed tended t o vary as the ratio of adult parasites to immature hosts varied. When the parasite density was lowest, about 5y0 of the immature population died. As this general value was
Can
. J. Z
ool.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
MIC
HIG
AN
ST
AT
E U
NIV
ER
SIT
Y o
n 12
/05/
14Fo
r pe
rson
al u
se o
nly.
CANADIAN JOURNAL OF ZOOLOGY. VOL. 42, 1964
NUMBER OF WEEKS
Can
. J. Z
ool.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
MIC
HIG
AN
ST
AT
E U
NIV
ER
SIT
Y o
n 12
/05/
14Fo
r pe
rson
al u
se o
nly.

BURNETT: HOST-PARASITE POPULATION 761
obtained in other experiments (Burnett 1962) in which the host insect was reared from egg to adult in the absence of the parasite, this value may be taken as a measure of natural mortality in the absence of parasite activity.
Two features of Fig. 9 indicate that the proportions of small, medium, and large host larvae in population S were also a factor in the degree of killing inflicted by a given density of adult parasites. First, as shown by the right- hand ordinates, the larger the host larvae, the greater the index of adult parasites per immature host required to produce a comparable level of killing. Second, there were obvious exceptions to the close relationship between the percentages of hosts killed and the numbers of parasites per immature host in each category of host (and also for the total number of hosts). The most striking was probably that for percentage of small larvae killed between the 17th and 21st weeks, but others occurred in the percentages of medium and large larvae killed in the 18th week. In the 17th week there was a distinct lag in the increase in percentage of small larvae killed with increase in the index of parasite-host ratio. At this time medium and large hosts were relatively abundant and were also subject to attack; note the continuing high level of percentage of parasitism (Fig. 9, D). In the 20th week, on the other hand, a much lower ratio resulted in high percentages of mortality because almost all hosts exposed were small larvae.
The influence of parasite density and of host size on the almost immediate killing of host larvae may be determined, to some extent, by relating the varia- tion in killing of small larvae (because these were the most abundant of the three stages) to variations in parasite density and relative abundance of small larvae in the population. The following multiple regression equation was calculated
Y = -3.352 + 5.234X1 + 0.3705X2
where Y = percentage of small larvae killed by the adult parasites, XI = the ratio of adult parasites to small host larvae, and XZ = percentage of small larvae in the population. The regression is highly significant (0.1% level) and the coefficients of both XI and Xz are also highly significant. An analysis of variance shows that the regression removes 50% of the variation in percentage of small larvae killed.
Examination of a graph relating percentage of small larvae killed to parasite density and percentage of small larvae in the population suggests an additional factor that may have influenced the mortality of small larvae. When large larvae were relatively abundant, the percentage of small hosts killed was less
FIG. 9. A. Percentages of small larvae of T. vaporariorum killed and numbers of adults of E. formosa per small larva in population S. Solid line, percentage killed. Dotted line, index of parasite-host ratio. B. Percentages of medium larvae killed and numbers of adults per medium larva. Solid line, percentage killed. Dotted line, index of parasite- host ratio (when there were no hosts the ratio was set a t zero). C. Percentages of large larvae killed and numbers of adults per large larva. Solid line, percentage killed. Dotted line, index of parasite-host ratio (when there were no hosts the ratio was set a t zero). D. Percentages of all larvae of T. vaporariorum parasitized by adults of E. formosa and numbers of adult parasites per larva. Solid line, percentages of hosts parasitized. Dotted line, index of parasitehost ratio.
Can
. J. Z
ool.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
MIC
HIG
AN
ST
AT
E U
NIV
ER
SIT
Y o
n 12
/05/
14Fo
r pe
rson
al u
se o
nly.

762 CANADIAN JOURNAL OF ZOOLOGY. VOL. 42, 1964
than expected. In this case, the size of host larvae may influence their sus- ceptibility to attack as well as their susceptibility to death following attack. That the size of a host influences its susceptibility to attack was shown in previous experiments (Burnett 1943, Table I) in which slightly more large larvae were attacked than would have occurred if the parasite attacked a mixture of large and medium larvae in proportion to their abundance. If the alternative hosts in these previous experiments had been large and small larvae, the preference for large hosts might have been much greater. Another factor, not primarily dependent on parasite activity, has been observed in the greenhouse to affect, to some extent, the survival of host scales. When the adult whitefly population was dense, as i t was a t the peak of each fluctuation, the females laid some of their eggs on older foliage. The probability of death is greater for larvae developing on older than for those developing on newer leaves.
Parasitization of Host Larvae The percentages of all hosts exposed to the parasites that were estimated
to be parasitized between sampling periods are shown in Fig. 9, D along with an index of the numbers of adult parasites per immature host. If the percentage of parasitism is calculated using only medium and large stages, the variations are similar to those shown but, of course, the values are higher. Parasitization occurred a t low parasitehost ratios, reached a maximum with slight increase in parasite density, and quickly declined with further increase in parasite attack. This type of change in level of parasitism was suggested by Fig. 1.
Figure 9, D shows that there were marked decreases in the percentage of hosts parasitized in about the l l th , 17th, and 23rd weeks, corresponding to the variation in the formation of black scales in about the 13th, 18th, and 25th weeks (Fig. 7, D), without a marked change in the parasite-host ratio. As these decreases occurred when the numbers of adult parasites had declined to minimal values, they could be related to the scarcity of E. formosa adults. However, Fig. 7, D shows that adults did not emerge for some time prior to these periods (the reason, of course, for the decline in numbers of adults). I t is possible, therefore, that the decline in parasitism without a marked change in the parasite-host ratio was caused partially by the aging of the adult parasite population. This interpretation is supported by the fact that a t higher temperatures the parasite lays most of its eggs soon after emergence rather than ovipositing uniformly throughout its adult life.
Discussion There were three features of the propagation of populations R and S that
should be emphasized in making an analysis of the interaction of T. vapora- riorum and E. formosa. First, as the parasite is very small (about 1 mm long), the greenhouse section that contained 64 separated tomato plants enhanced any effect that dispersion of immature hosts might have on the ability of the parasite to find hosts. Second, by infesting the plants with the two species over a 6-week period, the effects of life-history phenomena on the fluctuations in abundance of host and parasite were minimized. Third, because of the differences in numbers of adult hosts and parasites about the 25th week, a fifth fluctuation, larger in amplitude than the previous four, is probable. As
Can
. J. Z
ool.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
MIC
HIG
AN
ST
AT
E U
NIV
ER
SIT
Y o
n 12
/05/
14Fo
r pe
rson
al u
se o
nly.

BURNETT: HOST-PARASITE POPULATION 763
host density would be so great a t the peak of the fifth fluctuation that T. vaporariorurn would adversely affect its food supply, the trends of the growth- forms obtained in these experiments would continue until the interaction be- tween host plant and host insect modified significantly that between the host and parasite species.
Propagation of populations R and S has illustrated three characteristics of the interaction between E. formosa and its host, T. vaporariorum, when the species were reared in the greenhouse a t 72-76 O F . First, although both populations tended to fluctuate with peaks of increasing amplitude, the den- sity of population S remained about three times greater than that of population R throughout the period of propagation. Second, the percentages of immature hosts almost immediately killed, and also of hosts parasitized, were much the same, a t a given sampling period, in both populations; a much higher density of parasites in one population did not produce a proportionately greater mor- tality in that population. Third, within each population almost identical patterns of fluctuations in numbers of adult hosts occurred a t each sampling point and similar fluctuations were recorded also in the numbers of adult parasites, in numbers of immature hosts, in percentages of hosts killed, and in percentages of hosts parasitized; rather than dispersion and dispersal be- tween leaves and plants being of importance in the interaction of host and parasite populations, the same events occurred a t approximately the same time a t approximately the same intensity in all subunits of the populations. In view of these observations i t is unlikely that either the fluctuations in abundance of the two species or the general level about which the fluctuations occurred resulted from any differential distribution in space of the two species. On the contrary, the parasite had little difficulty in finding hosts a t all times. I t should be noted that this interpretation differs from that of Pimentel and Al-Hafidh (1963), who, although they obtained, in principle, the same types of growth-forms for these two species as reported in this paper, stressed the role of the space-time structure of the environment.
Death of scale insects normally infesting citrus has been observed to follow host feeding by adults of Metaphycus helvolus (Com.) (DeBach 1943), Aphytis lingnunensis Comp. and A. chrysomphali (Mercet) (DeBach and White 1960), Aphytis lepidosaphes Compere (DeBach and Landi 1961), and Aphytis ling- nanensis, A. fisheri De Bach, and A . melinus DeBach (DeBach and Sundby 1963). In the present experiments, only about one dozen adults of E. formosa were observed feeding on host larvae and these may have been feeding on honeydew. In spite of considerable effort, the mechanism whereby adult parasites either kill or parasitize larvae of T. vaporariorum has not been determined. But, whatever the mechanism employed, alternative types of mortality occur in a number of cases and they may be sufficiently widespread to influence the concepts of host-parasite interaction.
Conclusions The propagation, over a 26-week period, of two populations of T. vapor-
ariorum and its chalcid parasite E. formosa on tomato plants in the greenhouse a t 72-76 O F has shown that, although the rate of increase of the parasite population was greater than that of the host, the two populations fluctuated
Can
. J. Z
ool.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
MIC
HIG
AN
ST
AT
E U
NIV
ER
SIT
Y o
n 12
/05/
14Fo
r pe
rson
al u
se o
nly.

764 CANADIAN JOURNAL OF ZOOLOGY. VOL. 42. 1964
with peaks of increasing amplitude. The growth-forms of the two species were determined primarily by the variation in mortality of immature stages of the host following attack by adult parasites, by the synchronization of host and parasite populations, and by the reproduction of the host population.
The immature stages of the host were either almost immediately killed or parasitized by attack of the adult parasites. The incidence of death or parasit- ism was related to the intensity of parasite attack and to the size of the host scales attacked, but the differential susceptibility of host larvae of various sizes to attack, the age of adult parasites, and the occasional development of immature hosts on older foliage may have affected the type and degree of mortality. The pattern of mortality, in each fluctuation, followed a well- defined sequence. The chief feature of this sequence was the extensive killing of small larvae when both they and the adult parasites were most abundant. The adult parasites did not decrease in abundance, therefore, because they could not find hosts as readily as formerly, but because the adults, being short- lived (particularly as they were associated with small host larvae (Burnett 1949)), died quickly and their progeny died along with the host scales that contained them. The stages of the host that were immune to attack-eggs, pupae, and the relatively long-lived adults-carried the host population along to begin a new fluctuation. The apparent destruction of the parasite population was without doubt the primary factor in the interaction of T. vaporariorum and E. formosa because i t was a form of host protection. This protection would be especially important when environmental conditions were stringent for the host, as i t would help ensure the survival of the host-parasite system.
Synchronization of the host and parasite populations also affected, the interaction of T. vaporariorum and E. formosa. As the killing and parasitiza- tion of scales following parasite attack determined the composition of the population, the interaction established cycles in the age structures of the two populations. The cycles, on the other hand, determined that a definite composition of larval hosts was exposed for a given period to a given intensity of attack by females of E. formosa. This reciprocal relation between age struc- ture and mortality of host and parasite doubtless was the reason why the growth-forms were determined a t the beginning of the propagation of host and parasite. In the present experiments the interaction resulted in a rapid decline in parasite numbers a t the end of each fluctuation but in progressively higher minimal values of numbers of adult hosts. As the differences between numbers of adult hosts and parasites a t the end of each fluctuation became progressively greater, the successive fluctuations were of increasing amplitude.
As the parasite population did not maintain the host population a t a characteristic level of abundance, the reproduction of T. vaporariorum was an important factor in the interaction of the host and parasite species. Although the oviposition rate of T. vaporariorum was always very great, the index of reproduction varied, to some extent, with the season. As the reproductive rate of the host is very greatly altered by the succulence of the leaves of the host plant, a slight modification of the physical environment could change significantly the rate a t which host larvae enter the population. The rate of oviposition may have varied also with the density and age of adults of T. vaporariorum.
Can
. J. Z
ool.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
MIC
HIG
AN
ST
AT
E U
NIV
ER
SIT
Y o
n 12
/05/
14Fo
r pe
rson
al u
se o
nly.

BURNETT: HOST-PARASITE POPULATION 765
In these experiments the hosts were fully exposed to the parasites a t all times; the dispersal of host and parasite was not modified by barriers of any sort; the individual insects were not disturbed by the sampling technique; and once the initial conditions were established, the insects determined their own densities solely bv their natural interactions. The interactions are, how- - - ever, greatly modified by variations in the environment, particularly the physical environment. With increase in temperature above 72-76 OF, for example, the parasite's rate of increase becomes relatively greater than that of the host. Therefore, before either attempting a definitive description of the processes occurring in the interaction between T. vaporariorum and E. formosa or comparing this interaction with those reported for other host- parasite systems, for example, by Flanders and Badgley (1963), Pimentel, Nagel, and Madden (1963), Utida (1957, 1960), and Takahashi (1959), i t is necessary to propagate populations of T. vaporariorum and E. formosa under a number of conditions.
Acknowledgments The author gratefully acknowledges the assistance of Mrs. Pauline Potts
in conducting the experiments.
References BURNETT, T. 1943. The distribution of the eggs of Encarsia formosa Gahan with respect to its
host, the greenhouse whitefly Trialeurodes vaporariorum Westwood. Rev. Can. Biol. 2, 378-394.
1949. The effect of temperature on an insect host-parasite population. Ecology, 30, 113-134.
1960a. An insect host-parasite population. Can. J. Zool. 38, 57-75. 1960b. Effects of initial densities and periods of infestation on the growth-forms of a host and parasite population. Can. J. Zool. 38, 1063-1077.
1962. An effect of parasite attack on host mortality, as exemplified by Encarsia formosa and Trialeurodes vafiorariorum. Can. Entomologist, 94, 673-679.
DEBACH, P. 1943. The importance of host-feeding by adult parasites in the reduction of host populations. Econ. Entomol. 36, 647-658.
DEBACH, P. and LANDI, J. 1961. The introduced purple scale parasite, Aphy t i s lepidosphes Compere, and a method of integrating chemical with biological control. Hilgardia, 31, 459-497.
DEBACH, P. and SUNDBY, R. A. 1963. Competitive displacement between ecological homo- logues. Hilgardia, 34, 105-166.
DEBACH, P. and WHITE, E. B. 1960. Commercial mass culture of the California red scale parasite Aphytis lingnunensis. Calif. Agr. Expt. Sta. Bull. 770, 1-57.
FLANDERS, S. E. and BADGLEY, M. E. 1963. Prey-predator interactions in self-balanced laboratory populations. Hilgardia, 35, 145-183.
PIMENTEL, D. and AL-HAFIDH, R. 1963. The coexistence of insect parasites and hosts in laboratory ecosystems. Ann. Entomol. Soc. Am. 56, 676-678.
PIMENTEL, D., NAGEL, W. P., and MADDEN, J. L. 1963. Space-time structure of the environ- ment and the survival of parasite-host systems. Am. Naturalist, 97, 141-167.
SPEYER, E. R. 1927. An important parasite of the greenhouse whitefly. Bull. Entomol. Res. 17, 301-308.
TAKAHASHI, F. 1957. Synchrony between the parasitoid wasp and its hosts in their interacting system. Japan. Appl. Entomol. Zool. 1, 259-264.
1959. The effect of host finding efficiency of parasite on the cyclic fluctuation of population in the interacting system of Ephestia and Nemeritis. Japan. J. Ecol. 9, 88-93.
UTIDA, S. 1957. Population fluctuation, an experimental and theoretical approach. Cold Spring Harbor Symp. Quant. Biol. 22, 139-151.
1960. Experimental studies on the interaction between the bean weevils and their parasitic wasps. Proc. Intern. Congr. Entomol. l l t h , Vienna, 1, 731-734.
Can
. J. Z
ool.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
MIC
HIG
AN
ST
AT
E U
NIV
ER
SIT
Y o
n 12
/05/
14Fo
r pe
rson
al u
se o
nly.