Embed Size (px)
Citation preview

Comparative Biochemistry and Physiology Part A 127 (2000) 237–247
Host defense function in neutrophils from the American bison(Bison bison)
Steve D. Swain, Laura K. Nelson, Angela J. Hanson, Daniel W. Siemsen,Mark T. Quinn *
Department of Veterinary Molecular Biology and Center for Bison and Wildlife Health, Montana State Uni6ersity, Bozeman,MT 59717, USA
Received 7 June 2000; received in revised form 15 August 2000; accepted 17 August 2000
Abstract
Selected host defense functions of neutrophils isolated from American bison (Bison bison) were characterized andcompared with those of cattle (Bos taurus). Bison neutrophils had a robust chemotactic response to both IL-8 and LTB4,with maximal responses occurring at 10−7 M (IL-8) and 10−8 M (LTB4). The magnitude of the chemotactic responseto IL-8 was similar in bison and bovine neutrophils (except at 10−7 M IL-8, where bison had a stronger response). Inresponse to LTB4, bison neutrophils had a much stronger chemotaxis at both 10−8 and 10−7 M than did bovine cells.Production of reactive oxygen species (ROS) in response to phorbol myristate acetate (PMA) and opsonized zymosan(OpZ) was similar between bison and bovine neutrophils. However, the production of ROS in bison neutrophilsstimulated with OpZ was primarily intracellular, while extracellular release of ROS was evident in bovine neutrophilsstimulated with OpZ. Like bovine neutrophils, bison neutrophils did not generate a respiratory burst in response tofMLF. Granules prepared from bison neutrophils had potent direct killing action on the Gram-negative bacteriaEscherichia coli but failed to kill the Gram-positive bacteria Staphylococcus aureus and, at intermediate doses, actuallyhad a permissive effect for this bacteria. Thus, bison neutrophils have potent host defense capabilities similar in qualityto those of bovine neutrophils; however, unique differences are present, which may allow bison neutrophils to respondto the distinct immunological challenges that bison encounter. © 2000 Elsevier Science Inc. All rights reserved.
Keywords: Bison; Chemotaxis; Granule; Host defense; Neutrophil; Superoxide anion; Bovine; Interleukin-8; Leukotriene B4
www.elsevier.com/locate/cbpa
1. Introduction
Host defense against infectious organisms re-quires a coordinated response by both innate andacquired immune defense mechanisms (Tizard,1997). Among the cells involved in innate immu-nity, the polymorphonuclear leukocyte or neu-trophil plays a major role (Boxer et al., 1985).
These phagocytic cells travel throughout the ani-mal in search of foreign pathogens or damagedtissue and respond to a wide variety of inflamma-tory factors (Smith, 1994; Bokoch, 1995). Once apathogen is encountered, the neutrophil utilizesan impressive array of microbicidal weapons todestroy and remove the infectious agent (Cohen,1994). Neutrophils possess a number of antimi-crobial mechanisms, which are generally classifiedas oxidative or non-oxidative in nature. Oxidativemechanisms involve the production of reactiveoxygen species (ROS) which can be microbicidal,
* Corresponding author. Tel.: +1-406-9945721; fax: +1-406-9944303.
E-mail address: [email protected] (M.T. Quinn).
1095-6433/00/$ - see front matter © 2000 Elsevier Science Inc. All rights reserved.PII: S 1095 -6433 (00 )00264 -6

S.D. Swain et al. / Comparati6e Biochemistry and Physiology, Part A 127 (2000) 237–247238
while non-oxidative mechanisms include mostother neutrophil functions, such as chemotaxis,release of proteolytic enzymes, and release ofproteins with direct bactericidal actions, such asdefensins (Cohen, 1994; Anderson, 1995). Al-though, the essential role of neutrophils in defend-ing the host is well conserved among species(Styrt, 1989), important species differences do ex-ist in how these cells respond to various stimuli.These include differences in the production ofROS, both in the initiation and the magnitude ofa response (Brown and Roth, 1991; Watson et al.,1995), differences in the relative amounts of vari-ous proteolytic enzymes contained in the cell(Styrt, 1989; Sahoo et al., 1998), and the presenceof different groups of proteins with direct bacteri-cidal actions (Gennaro et al., 1991). One group ofmammals that has been shown to possess uniquedifferences in neutrophil function is the rumi-nants. As an example, chemotactic peptides suchas N-formyl-methionine-leucine-phenylalanine( fMLF) strongly activate the production of ROSby human (Jesaitis et al., 1988), equine (Brazil etal., 1998), rabbit (Ye et al., 1993), and rodentneutrophils (Marasco et al., 1983); yet, neu-trophils from cattle (Brown and Roth, 1991; Wat-son et al., 1995), sheep (Buchta, 1990), and pigs(Fletcher et al., 1990) are essentially unresponsiveto these peptides. Bovine neutrophils also possessunique forms of antibacterial proteins and pep-tides (e.g. bactenecins) not found in any othernon-ruminant species studied (Gennaro et al.,1991).
Currently, the majority of studies on neutrophilfunction have been performed on human cells,while the remaining research in this area hasfocused on neutrophils obtained from laboratoryanimals and a few domestic livestock species only.In contrast, very little is known about the func-tion of neutrophils in wild ruminant species. TheAmerican Buffalo (Bison bison) is a good candi-date for examining possible differences in neu-trophil function in ruminants, for two reasons.First, because bison are a wild, indigenous rumi-nant species, it would be of interest to examinewhether the unique characteristics seen in neu-trophils from domestic cattle and sheep are com-mon to a wild ruminant species, which may havefunctional and evolutionary implications. Second,in some areas bison are semi-domesticated and inconstant contact with man and livestock; for ex-ample, in Yellowstone National Park and on
ranches in the western United States where bisonare raised as a source of meat. In these areas,there is a great deal of controversy over thedegree to which bison represent a disease vectorto commercial livestock, in particular as a sourceof the intracellular Gram-negative pathogen Bru-cella abortus (Tessaro, 1986). Scientific efforts toexamine this problem have focused on acquiredimmune responses, with the goal of producingappropriate vaccines that would reduce the inci-dence of Brucellosis in bison (Davis et al., 1991;Peterson et al., 1991; Olsen et al., 1998). In con-trast, virtually nothing is known about the basicfunction of neutrophils, or other innate immuneresponses in bison. To begin to address thesequestions, we have characterized a number ofbasic host defense functions, both oxidative andnon-oxidative, in bison neutrophils.
2. Methods and materials
2.1. Materials
Leukotriene B4 (LTB4) and recombinant humaninterleukin-8 (IL-8) were purchased from Cal-biochem (La Jolla, CA). Dulbecco’s phosphate-buffered saline (DPBS) was from Gibco/BRL(Grand Island, NY). Injectable grade water, ob-tained from Abbott Laboratories (North Chicago,IL), was used in all solutions made for isolatingneutrophils. Unless otherwise indicated, all otherreagents, including Histopaque and Percoll wereobtained through Sigma Chemical Co. (St. Louis,MO).
2.2. Animal care and blood collection
A group of nine North American bison calves(age 4–16 months) and four Holstein bull calves(age 4–9 months) were obtained from local herdsand maintained in our approved large animal carefacility. Blood was obtained by jugular venouspuncture and collected into 20 ml siliconizedVacutainer tubes containing 5 mM ethylene di-amine tetraacetic acid (EDTA). Blood from threeto four different animals was used in all experi-ments except for the preparation of neutrophilgranule extracts described below, in which bloodfrom two bison was used. For comparison insome experiments, peripheral blood was also ob-tained from healthy adult human volunteers.

S.D. Swain et al. / Comparati6e Biochemistry and Physiology, Part A 127 (2000) 237–247 239
2.3. Neutrophil isolation
Neutrophils were obtained from bison bloodfollowing methods previously developed in ourlab for isolating bovine neutrophils (Davis et al.,1998; Soltys et al., 1999). After removing erythro-cytes by H2O lysis, the white cells were resus-pended in DPBS and fractionated on two-stepHistopaque gradients (Davis et al., 1998) or pre-formed continuous Percoll gradients (Soltys et al.,1999). The purified neutrophil bands were col-lected and washed two times with approximatelyten volumes of DPBS to remove the gradientmedia. The oxidative burst assays described belowused cells obtained from both types of gradients.Since no differences in response were seen be-tween cells prepared by either method, all otherexperiments used cells obtained via the two-stepHistopaque gradient method. All reagents used inthe isolation were endotoxin free, and the entireprocedure was done at room temperature to avoidundue isolation-related priming effects. Humanand bovine neutrophils were isolated using theHistopaque method as described previously(DeLeo et al., 1996; Davis et al., 1998).
2.4. Measurement of oxidati6e burst
The production of ROS by isolated bison neu-trophils was measured using both luminol-en-hanced chemiluminescence and cytochrome cbased assays (Dahlgren and Karlsson, 1999).Briefly, chemiluminescence was performed inwhite Fluoronunc 96-well microtiter plates (NalgeNunc International, Rochester, NY) in a totalvolume of 200 ml per well. Neutrophils (5×105)were added to DPBS with 1mM CaCl2 and 150mM luminol. After a 5 min pre-incubation at37°C, activators were added in a volume of 20 mlper well, and chemiluminescence was measuredfor up to 25 min in a Fluoroskan Ascent FLmicrotiter plate reader (Lab Systems, Helsinki,Finland), with a 10 s per well reading interval(Swain et al., 1998). Activating agents used werephorbol myristate acetate (PMA; 100 ng ml−1
final concentration), fMLF (1 mM final concentra-tion), and opsonized zymosan (OpZ; 0.5 mg ml−1
final concentration). OpZ was prepared by incu-bating 10 mg ml−1 of zymosan in autologousbison plasma for 40 min at 37°C while rotating.
Superoxide anion (O2−) production was mea-
sured in clear Immulon 2 HB 96-well microtiter
plates (Dynex Technologies, Chantilly, VA) in atotal volume of 200 ml per well. Neutrophils (2×105) were added to DPBS with 1 mM CaCl2 and100 mM cytochrome c. Pre-incubations and acti-vation were performed as described above, andabsorbance at 550 nm was measured continuouslyfor 15 min in a THERMOMax microtiter platereader (Molecular Devices, Sunnyvale, CA)(DeLeo et al., 1996). Duplicate wells containing80 mg ml−1 superoxide dismutase (SOD) wereincluded to allow for subtraction of any non-spe-cific reduction of cytochrome c. Rates of SOD-in-hibitable O2
− production are expressed as nmolesO2
− min−1 per 107cells, assuming o=21×103
M−1 cm−1 for cytochrome c.
2.5. Chemotaxis
Chemotaxis of bison, bovine, and human neu-trophils was measured in Falcon 24-well platescontaining transwell inserts (3 mm pore size) asdescribed previously (Soltys et al., 1999). Briefly,transwell inserts containing 250 ml purified cells(106 cells per ml) were placed in wells containingbuffer (control) or chemoattractant. The chemoat-tractants used were the chemokine IL-8 and theleukotriene LTB4, over the concentration range10−10–10−7 M. After incubating the plates for120 min at 37°C in a humidified 5% CO2 incuba-tor, the transwell inserts were removed, and theamount of lactate dehydrogenase (LDH) releasedby migrated cells after cell lysis was determinedusing a colorimetric assay (CytoTox 96 Non-Ra-dioactive Cytotoxicity Assay; Promega, Madison,WI). Briefly, migrated cells were lysed with 1%Triton-X 100 for 30 min, and the plates werecentrifuged at 250×g for 12 min. The superna-tant were transferred to an Immulon 96-well mi-crotiter plate, and the plates were read at 490 nmwith a THERMOmax microtiter plate reader. TheLDH measurements were then converted to abso-lute cell numbers by comparison of the valueswith standard curves created by aliquotting arange of control cell standards (25, 12.5, 6.25, and3.125×104 cells) into wells containing no tran-swell inserts and processing identically to the wellswith inserts.
2.6. Preparation of neutrophil granule extracts
Granule extracts were prepared by differentialcentrifugation (Fujita et al., 1987) of homogenized

S.D. Swain et al. / Comparati6e Biochemistry and Physiology, Part A 127 (2000) 237–247240
bison neutrophils. Purified cells were treated with3 mM diisopropylfluorophosphate for 15 min,washed, and resuspended in cavitation buffer (10mM HEPES, 100 mM KCl, 10 mM NaCl, pH7.4) at 108 cells per ml. The cells were thenhomogenized by N2 cavitation (450 psi for 15 minat 4°C), the cavitate was collected into EGTA ata final concentration of 1.25 mM, and centrifugedat 850×g for 10 min at 4°C to remove unbrokencells, nuclei, and large debris. The supernatantwas then centrifuged at 14 000×g for 10 min at4°C. The resulting pellet containing the granulefraction was resuspended in a minimal volume ofcavitation buffer. The supernatant was also col-lected and centrifuged at 170 000×g for 45 minat 4°C to obtain the neutrophil plasma membrane(pellet) and cytosol (supernatant) fractions(Bunger et al., 2000).
2.7. Bacteria killing assay
Bison and bovine granule fractions were ana-lyzed for bactericidal activity following previouslydescribed methods (Gennaro et al., 1983). Es-cherichia coli B (Clowes and Hayes, 1968) andStaphylococcus aureus (ATCC c25923) weregrown in LB media (GIBCO/BRL, Grand Island,NY) until cultures reached an OD450 of approxi-mately 0.2 (�107 ml−1). The bacteria were thenwashed twice in DPBS and resuspended in DPBSat approximately 108 ml−1. At the time of theassay, granule aliquots were disrupted with threecycles of freeze–thawing (liquid N2/room temper-ature), protein concentration was determined us-ing the Bradford method (BioRad, Hercules, CA),and the indicated amounts of granule proteinwere mixed with 107 bacteria and DPBS to a finalvolume of 400 ml per tube. After incubation withshaking for 30 min at 37°C, the samples werediluted at concentrations ranging from 1:100 to1:100 000, and 50 ml aliquots were plated on LBagar. After overnight incubation at 37°C, bacte-rial numbers were determined by colony counting.
2.8. Electrophoresis
Granule fractions of bison and bovine neu-trophils were separated by sodium dodecyl sul-fate-polyacrylamide gel electrophoresis (SDS-PAGE) on 4–22% polyacrylamide gradient gelsand stained with GELCODE Blue Stain (PierceChemical Co., Rockford, IL) as described previ-
ously (Swain et al., 2000). For reference,prestained molecular weight standards (Amer-sham Pharmacia, Piscataway, NJ) were includedon all gels.
3. Results
3.1. Bison neutrophil isolation
We have adapted methods developed by ourlab for bovine neutrophil isolation to successfullypurify neutrophils from bison blood. These isola-tion procedures resulted in preparations of bisoncells that were \89% neutrophils, as determinedby Wright staining and microscopic analysis. Thislevel of purity is comparable with what is nor-mally achieved with bovine neutrophil prepara-tions (Kremer et al., 1992; Soltys et al., 1999). Theremaining cells typically consisted of eosinophils(�7%) and mononuclear leukocytes (B4%). Cellviability, as determined by trypan blue exclusion,was greater than 99%. Cells from both types ofseparation gradients (Percoll and Histopaque)gave similar results in functional tests, so theHistopaque gradients were used routinely becauseof their ease of use.
3.2. Bison neutrophil chemotaxis
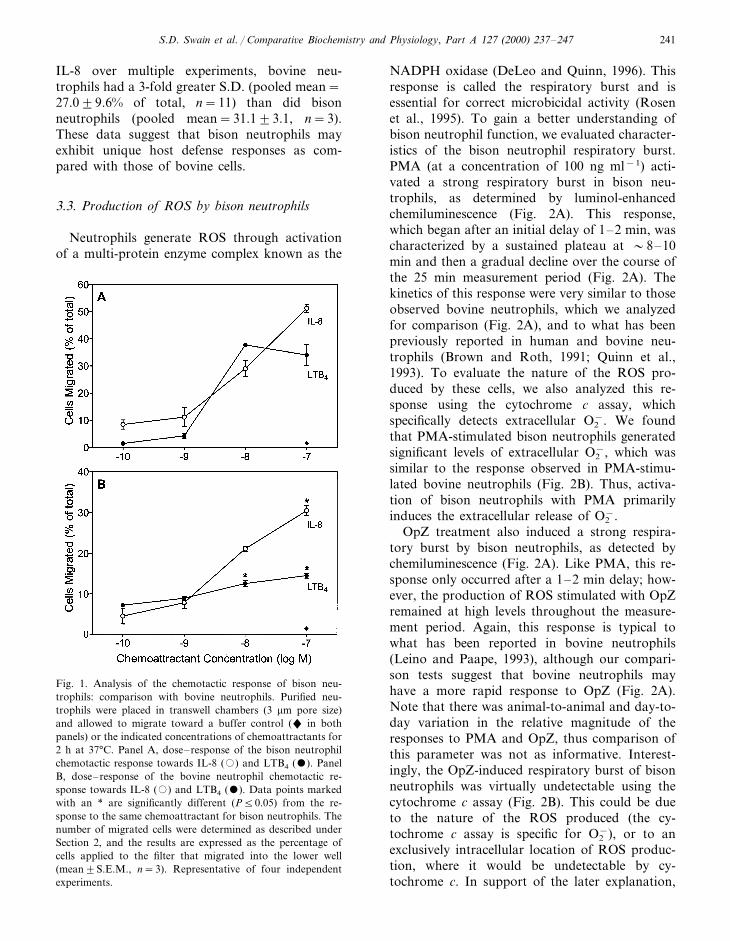
Bison neutrophil chemotaxis was evaluated inresponse to two chemoattractants that have beenshown to be important inflammatory mediators(IL-8 and LTB4) for both human and bovine cells(Persson et al., 1993; Bokoch, 1995; Caswell et al.,1999). As shown in Fig. 1A, bison neutrophilsexhibited a strong, dose-dependent migration to-wards both IL-8 and LTB4. The magnitude of thechemotactic response toward IL-8, when ex-pressed in terms of the percentage of applied cellsthat migrated, was similar to that of bovine neu-trophils analyzed under identical assay conditions(Fig. 1B), with the exception that bison neu-trophils exhibited slightly greater chemotaxis tothe highest dose of IL-8 tested (10−7 M). Incontrast, chemotaxis of bison neutrophils towardLTB4 was much greater than that seen in bovineneutrophils, at both 10−8 and 10−7 M (Fig. 1B).It was also notable that bovine neutrophils exhib-ited much more day-to-day variation in chemotac-tic response than did bison neutrophils. Forexample, looking only at the response to 10−8 M
S.D. Swain et al. / Comparati6e Biochemistry and Physiology, Part A 127 (2000) 237–247 241
IL-8 over multiple experiments, bovine neu-trophils had a 3-fold greater S.D. (pooled mean=27.099.6% of total, n=11) than did bisonneutrophils (pooled mean=31.193.1, n=3).These data suggest that bison neutrophils mayexhibit unique host defense responses as com-pared with those of bovine cells.
3.3. Production of ROS by bison neutrophils
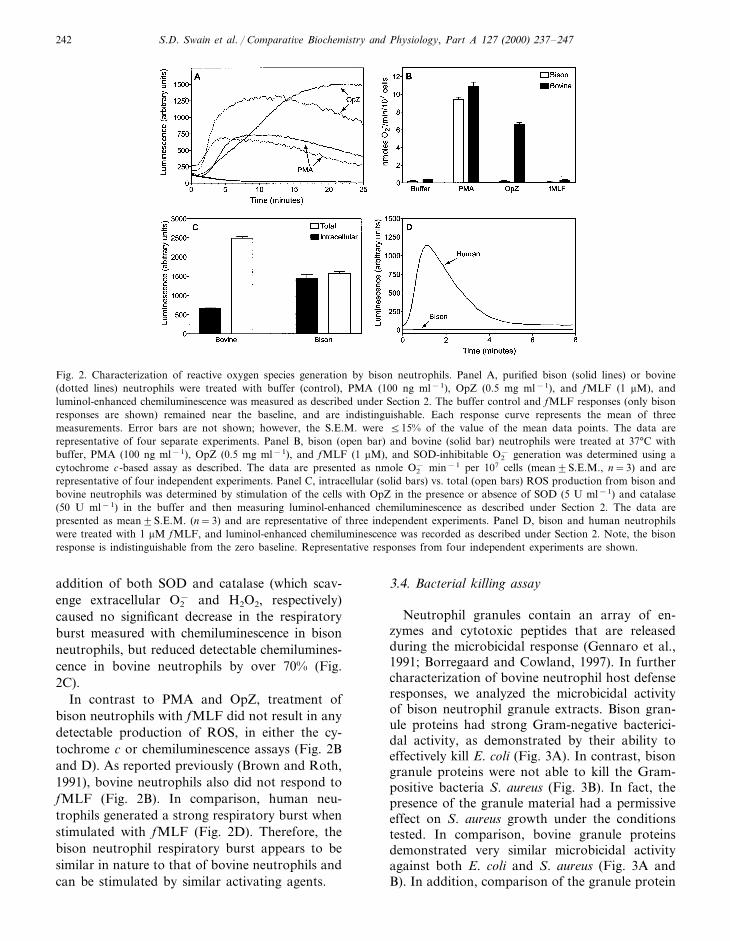
Neutrophils generate ROS through activationof a multi-protein enzyme complex known as the
NADPH oxidase (DeLeo and Quinn, 1996). Thisresponse is called the respiratory burst and isessential for correct microbicidal activity (Rosenet al., 1995). To gain a better understanding ofbison neutrophil function, we evaluated character-istics of the bison neutrophil respiratory burst.PMA (at a concentration of 100 ng ml−1) acti-vated a strong respiratory burst in bison neu-trophils, as determined by luminol-enhancedchemiluminescence (Fig. 2A). This response,which began after an initial delay of 1–2 min, wascharacterized by a sustained plateau at �8–10min and then a gradual decline over the course ofthe 25 min measurement period (Fig. 2A). Thekinetics of this response were very similar to thoseobserved bovine neutrophils, which we analyzedfor comparison (Fig. 2A), and to what has beenpreviously reported in human and bovine neu-trophils (Brown and Roth, 1991; Quinn et al.,1993). To evaluate the nature of the ROS pro-duced by these cells, we also analyzed this re-sponse using the cytochrome c assay, whichspecifically detects extracellular O2
−. We foundthat PMA-stimulated bison neutrophils generatedsignificant levels of extracellular O2
−, which wassimilar to the response observed in PMA-stimu-lated bovine neutrophils (Fig. 2B). Thus, activa-tion of bison neutrophils with PMA primarilyinduces the extracellular release of O2
−.OpZ treatment also induced a strong respira-
tory burst by bison neutrophils, as detected bychemiluminescence (Fig. 2A). Like PMA, this re-sponse only occurred after a 1–2 min delay; how-ever, the production of ROS stimulated with OpZremained at high levels throughout the measure-ment period. Again, this response is typical towhat has been reported in bovine neutrophils(Leino and Paape, 1993), although our compari-son tests suggest that bovine neutrophils mayhave a more rapid response to OpZ (Fig. 2A).Note that there was animal-to-animal and day-to-day variation in the relative magnitude of theresponses to PMA and OpZ, thus comparison ofthis parameter was not as informative. Interest-ingly, the OpZ-induced respiratory burst of bisonneutrophils was virtually undetectable using thecytochrome c assay (Fig. 2B). This could be dueto the nature of the ROS produced (the cy-tochrome c assay is specific for O2
−), or to anexclusively intracellular location of ROS produc-tion, where it would be undetectable by cy-tochrome c. In support of the later explanation,
Fig. 1. Analysis of the chemotactic response of bison neu-trophils: comparison with bovine neutrophils. Purified neu-trophils were placed in transwell chambers (3 mm pore size)and allowed to migrate toward a buffer control (" in bothpanels) or the indicated concentrations of chemoattractants for2 h at 37°C. Panel A, dose–response of the bison neutrophilchemotactic response towards IL-8 (�) and LTB4 (). PanelB, dose–response of the bovine neutrophil chemotactic re-sponse towards IL-8 (�) and LTB4 (). Data points markedwith an * are significantly different (P50.05) from the re-sponse to the same chemoattractant for bison neutrophils. Thenumber of migrated cells were determined as described underSection 2, and the results are expressed as the percentage ofcells applied to the filter that migrated into the lower well(mean9S.E.M., n=3). Representative of four independentexperiments.
S.D. Swain et al. / Comparati6e Biochemistry and Physiology, Part A 127 (2000) 237–247242
Fig. 2. Characterization of reactive oxygen species generation by bison neutrophils. Panel A, purified bison (solid lines) or bovine(dotted lines) neutrophils were treated with buffer (control), PMA (100 ng ml−1), OpZ (0.5 mg ml−1), and fMLF (1 mM), andluminol-enhanced chemiluminescence was measured as described under Section 2. The buffer control and fMLF responses (only bisonresponses are shown) remained near the baseline, and are indistinguishable. Each response curve represents the mean of threemeasurements. Error bars are not shown; however, the S.E.M. were 515% of the value of the mean data points. The data arerepresentative of four separate experiments. Panel B, bison (open bar) and bovine (solid bar) neutrophils were treated at 37°C withbuffer, PMA (100 ng ml−1), OpZ (0.5 mg ml−1), and fMLF (1 mM), and SOD-inhibitable O2
− generation was determined using acytochrome c-based assay as described. The data are presented as nmole O2
− min−1 per 107 cells (mean9S.E.M., n=3) and arerepresentative of four independent experiments. Panel C, intracellular (solid bars) vs. total (open bars) ROS production from bison andbovine neutrophils was determined by stimulation of the cells with OpZ in the presence or absence of SOD (5 U ml−1) and catalase(50 U ml−1) in the buffer and then measuring luminol-enhanced chemiluminescence as described under Section 2. The data arepresented as mean9S.E.M. (n=3) and are representative of three independent experiments. Panel D, bison and human neutrophilswere treated with 1 mM fMLF, and luminol-enhanced chemiluminescence was recorded as described under Section 2. Note, the bisonresponse is indistinguishable from the zero baseline. Representative responses from four independent experiments are shown.
addition of both SOD and catalase (which scav-enge extracellular O2
− and H2O2, respectively)caused no significant decrease in the respiratoryburst measured with chemiluminescence in bisonneutrophils, but reduced detectable chemilumines-cence in bovine neutrophils by over 70% (Fig.2C).
In contrast to PMA and OpZ, treatment ofbison neutrophils with fMLF did not result in anydetectable production of ROS, in either the cy-tochrome c or chemiluminescence assays (Fig. 2Band D). As reported previously (Brown and Roth,1991), bovine neutrophils also did not respond tofMLF (Fig. 2B). In comparison, human neu-trophils generated a strong respiratory burst whenstimulated with fMLF (Fig. 2D). Therefore, thebison neutrophil respiratory burst appears to besimilar in nature to that of bovine neutrophils andcan be stimulated by similar activating agents.
3.4. Bacterial killing assay
Neutrophil granules contain an array of en-zymes and cytotoxic peptides that are releasedduring the microbicidal response (Gennaro et al.,1991; Borregaard and Cowland, 1997). In furthercharacterization of bovine neutrophil host defenseresponses, we analyzed the microbicidal activityof bison neutrophil granule extracts. Bison gran-ule proteins had strong Gram-negative bacterici-dal activity, as demonstrated by their ability toeffectively kill E. coli (Fig. 3A). In contrast, bisongranule proteins were not able to kill the Gram-positive bacteria S. aureus (Fig. 3B). In fact, thepresence of the granule material had a permissiveeffect on S. aureus growth under the conditionstested. In comparison, bovine granule proteinsdemonstrated very similar microbicidal activityagainst both E. coli and S. aureus (Fig. 3A andB). In addition, comparison of the granule protein
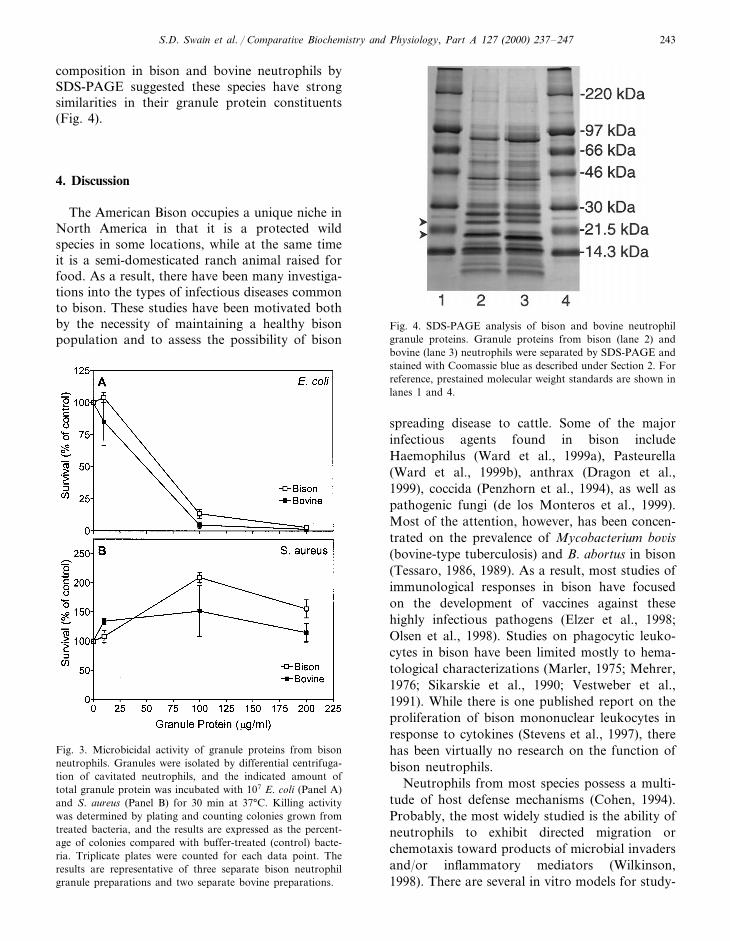
S.D. Swain et al. / Comparati6e Biochemistry and Physiology, Part A 127 (2000) 237–247 243
composition in bison and bovine neutrophils bySDS-PAGE suggested these species have strongsimilarities in their granule protein constituents(Fig. 4).
4. Discussion
The American Bison occupies a unique niche inNorth America in that it is a protected wildspecies in some locations, while at the same timeit is a semi-domesticated ranch animal raised forfood. As a result, there have been many investiga-tions into the types of infectious diseases commonto bison. These studies have been motivated bothby the necessity of maintaining a healthy bisonpopulation and to assess the possibility of bison
Fig. 4. SDS-PAGE analysis of bison and bovine neutrophilgranule proteins. Granule proteins from bison (lane 2) andbovine (lane 3) neutrophils were separated by SDS-PAGE andstained with Coomassie blue as described under Section 2. Forreference, prestained molecular weight standards are shown inlanes 1 and 4.
Fig. 3. Microbicidal activity of granule proteins from bisonneutrophils. Granules were isolated by differential centrifuga-tion of cavitated neutrophils, and the indicated amount oftotal granule protein was incubated with 107 E. coli (Panel A)and S. aureus (Panel B) for 30 min at 37°C. Killing activitywas determined by plating and counting colonies grown fromtreated bacteria, and the results are expressed as the percent-age of colonies compared with buffer-treated (control) bacte-ria. Triplicate plates were counted for each data point. Theresults are representative of three separate bison neutrophilgranule preparations and two separate bovine preparations.
spreading disease to cattle. Some of the majorinfectious agents found in bison includeHaemophilus (Ward et al., 1999a), Pasteurella(Ward et al., 1999b), anthrax (Dragon et al.,1999), coccida (Penzhorn et al., 1994), as well aspathogenic fungi (de los Monteros et al., 1999).Most of the attention, however, has been concen-trated on the prevalence of Mycobacterium bo6is(bovine-type tuberculosis) and B. abortus in bison(Tessaro, 1986, 1989). As a result, most studies ofimmunological responses in bison have focusedon the development of vaccines against thesehighly infectious pathogens (Elzer et al., 1998;Olsen et al., 1998). Studies on phagocytic leuko-cytes in bison have been limited mostly to hema-tological characterizations (Marler, 1975; Mehrer,1976; Sikarskie et al., 1990; Vestweber et al.,1991). While there is one published report on theproliferation of bison mononuclear leukocytes inresponse to cytokines (Stevens et al., 1997), therehas been virtually no research on the function ofbison neutrophils.
Neutrophils from most species possess a multi-tude of host defense mechanisms (Cohen, 1994).Probably, the most widely studied is the ability ofneutrophils to exhibit directed migration orchemotaxis toward products of microbial invadersand/or inflammatory mediators (Wilkinson,1998). There are several in vitro models for study-

S.D. Swain et al. / Comparati6e Biochemistry and Physiology, Part A 127 (2000) 237–247244
ing leukocyte chemotaxis. For human neutrophils,the most common approach is to measure neu-trophil movement through filters toward a solu-tion containing the chemoattractant peptidefMLF (Schiffmann et al., 1975). However, be-cause neutrophils from most ruminants that havebeen tested are apparently non-responsive tofMLF (Styrt, 1989), the chemoattractant ofchoice for bovine neutrophils in assays of thistype is the cytokine IL-8 (Caswell et al., 1999;Galligan and Coomber, 2000). Not only dobovine neutrophils exhibit chemotaxis to bothrecombinant human and bovine IL-8 in vitro, butthere are also reports suggesting that IL-8 plays arole in vivo during conditions where a local accu-mulation of neutrophils occurs (e.g. mastitis,Persson et al., 1993; Persson-Waller et al., 1997;Barber and Yang, 1998). IL-8 is produced byseveral cell types (neutrophils, monocytes, en-dothelial cells) in response to a number of inflam-matory mediators, such as lipopolysaccharide,interleukin-1, and tumor necrosis factor-a (Lind-ley, 1998). In addition, IL-8 is becoming widelyviewed as a key inflammatory stimulus resultingin early influx of neutrophils to sites of potentialinfection (Harada et al., 1994). For these reasons,we chose to examine chemotaxis of bison neu-trophils towards IL-8. We report here that bisonneutrophils demonstrated a dose-dependentchemotaxis towards IL-8 over the concentrationrange of 1–100 nM. The chemotactic response ofbison neutrophils to IL-8 is very similar to whathas been reported for bovine neutrophils (Caswellet al., 1999), although this group tested IL-8concentrations as high as 10 mg ml−1 and re-ported increased chemotaxis at these concentra-tions. In contrast, Galligan and Coomber (2000)reported that higher numbers (\60%) of bovineneutrophils migrated towards low doses of recom-binant human IL-8, with a plateau of the responseat 5 ng ml−1. In our comparisons of bison andbovine neutrophil chemotaxis with recombinanthuman IL-8, under identical conditions, neu-trophils from both species exhibited similar re-sponses, although, neutrophil response to 10−7 MIL-8 was slightly stronger in bison. Possible expla-nations for this discrepancy in bovine responsesare different measurement systems used by differ-ent investigators, and the large animal-to-animalvariation that is characteristic of bovine cells (Sal-gar et al., 1991).
Another important chemotactic agent for hu-man neutrophils, both in vitro and in vivo, is theeicosanoid LTB4 (Crooks and Stockley, 1998). Inbovines, LTB4 has been shown to have modest invitro chemotactic effects on neutrophils, whilehaving little effect on neutrophil accumulation inthe in vivo teat cistern model (Persson et al.,1993). In our experiments, bison neutrophils ex-hibited a consistent pattern of chemotaxis towardsLTB4, with a peak response at 10−8 M, andgradually decreasing levels of chemotaxis at bothlower and higher concentrations. Our compari-sons with bovine neutrophils showed an identi-cally shaped dose-response curve, although themagnitude of the response to LTB4 was greater inbison than in cattle (�3-fold). Overall, theseresults indicate that bison neutrophils respond tophysiological inflammatory mediators that fit theexpected pattern of host defense for neutrophils.
Another vital neutrophil function is the genera-tion of ROS, which play an important anti-micro-bial role in defense against bacteria or fungi(Rosen et al., 1995). We investigated this responsein bison neutrophils using two complementarytechniques. The cytochrome c assay allows for thequantitation of O2
− produced, which facilitatesdirect comparison with published reports of neu-trophils from other species. The luminol chemilu-minescence technique, while not quantitative,provides for easier examination of the kinetics ofthe oxidative burst and also takes into accountROS produced inside the cell but not released.Overall, the oxidative burst of bison neutrophilswas similar to that of bovine neutrophils in manyrespects. First, bison neutrophils did not respondto fMLF treatment with a respiratory oxidativeburst. Second, the magnitude of the PMA-in-duced respiratory burst was quantitatively lower(at least with respect to extracellular O2
− release)than what was seen with human neutrophils.Third, bison neutrophils exhibited a considerableamount of animal-to-animal and day-to-day vari-ation in their oxidative burst response to PMA,and especially to OpZ, as has been reported incattle (Salgar et al., 1991). What was potentiallyinteresting about bison neutrophils was the highratio of intracellular to extracellular ROS produc-tion in response to OpZ. Whether this has anyfunctional implications is still unclear; however,sequestering of the ROS in phagocytic vesicles inresponse to particulate stimuli could provide effi-cient killing action towards phagocytosed patho-

S.D. Swain et al. / Comparati6e Biochemistry and Physiology, Part A 127 (2000) 237–247 245
gens without damage to host tissue that couldresult from the release of large amounts of extra-cellular ROS. A more detailed study with a largersample size and other types of particulate stimuliwill be necessary, however, to determine whetherthis is a unique feature of bison neutrophils.
Another component of the potent anti-micro-bial strategy of neutrophils is the production ofsmall, cationic proteins or peptides that can di-rectly kill bacteria by disrupting bacterial mem-brane integrity (Ganz, 1999). There are two majorgroups of these molecules, the defensins and thecathelicidins (Gudmundsson and Agerberth,1999); however, there is a great deal of hetero-geneity of these molecules, with different bio-chemical features and species distributions(Spitznagel, 1990). Cattle, for example, seem to bethe only species described to date that have theneutrophil cathelicidin proteins bac7 and bac5(Zanetti et al., 1990; Gennaro et al., 1991). Be-cause there are such species differences in thecomplement of antibacterial proteins, we wereespecially interested in comparing bison neu-trophil granule antibacterial capabilities withthose of bovine. The pattern of bacterial killing byneutrophil granules that we present here appearsto be very similar in bison and cattle. This wouldsuggest that the major anti-bacterial proteins inbison neutrophils are related to the cathelicidinproteins found in other ruminants, since theseproteins potently kill Gram-negative, but notGram-positive bacteria. In addition, comparisonof bison and bovine granule proteins by SDS-PAGE suggests that there are only minor differ-ences in the molecular weights of the majorgranule proteins (see Fig. 4, arrowheads).Whether these differences would correspond todifferent potency against other pathogens is un-known. One limitation of these assays is that theyare performed with laboratory strains of commonbacteria. A more interesting comparison might beto utilize bacteria that are potentially pathogenicto the animal in its natural ecology.
Overall, bison neutrophils appear to have po-tent host defense capabilities similar in quality tothat of bovine neutrophils. Bison neutrophils mi-grate effectively toward the physiologicalchemoattractants IL-8 and LTB4, they produceROS in response to stimulation by the pharmaco-logical activator PMA and the quasi-physiologicalstimulus of opsonized zymosan, and they haveefficient non-oxidative killing of Gram-negative
bacteria, probably due to granule derived antibac-terial proteins. It should be noted, however, thatunique and subtle differences do appear to bepresent in the bison system that may allow bisonneutrophils to respond to the distinct host defensechallenges that bison encounter. Further analysisof these differences may eventually lead to abetter understanding of the innate immune re-sponse in bison.
Acknowledgements
We would like to thank Dr David Pascual forproviding bison for this study. We would also liketo thank Jim Thompson and Kerri Rask forexpert animal care and sampling. This work wassupported in part by USDA/NRICGP 99-03600,USDA/NRICGP 99-03508, USDA 99-02950,USDA Animal Health Formula Funds, and theMontana State University Agricultural Experi-mental Station. Dr Quinn is an Established Inves-tigator of the American Heart Association. This ismanuscript 2000-40 from the Montana Agricul-tural Experiment Station, Montana StateUniversity-Bozeman.
References
Anderson, R., 1995. The activated neutrophil-Formidable forces unleashed. South Afr. Med. J.85, 1024–1028.
Barber, M.R., Yang, T.J., 1998. Chemotactic activitiesin nonmastitic and mastitic mammary secretions:presence of interleukin-8 in mastitic but not non-mastitic secretions. Clin. Diagn. Lab. Immunol. 5,82–86.
Bokoch, G.M., 1995. Chemoattractant signaling andleukocyte activation. Blood 86, 1649–1660.
Borregaard, N., Cowland, J.B., 1997. Granules of thehuman neutrophilic polymorphonuclear leukocyte.Blood 89, 3503–3521.
Boxer, G.J., Curnutte, J.T., Boxer, L.A., 1985. Poly-morphonuclear leukocyte function. Hosp. Pract. 40,69–90.
Brazil, T.J., Rossi, A.G., Haslett, C., McGorum, B.,Dixon, P.M., Chilvers, E.R., 1998. Priming inducesfunctional coupling of N-formyl-methionyl-leucyl-phenylalanine receptors in equine neutrophils. J.Leukocyte Biol. 63, 380–388.
Brown, G.B., Roth, J.A., 1991. Comparison of theresponse of bovine and human neutrophils to vari-ous stimuli. Vet. Immunol. Immunopathol. 28,201–218.

S.D. Swain et al. / Comparati6e Biochemistry and Physiology, Part A 127 (2000) 237–247246
Buchta, R., 1990. Functional and biochemical proper-ties of ovine neutrophils. Vet. Immunol. Im-munopathol. 24, 97–112.
Bunger, P.L., Swain, S.D., Clements, M.K., Siemsen,D.W., Davis, A.R., Gauss, K.A., Quinn, M.T.,2000. Cloning and expression of bovine p47-phoxand p67-phox : comparison with the human andmurine homologs. J. Leukocyte Biol. 67, 63–72.
Caswell, J.L., Middleton, D.M., Gordon, J.R., 1999.Production and functional characterization of re-combinant bovine interleukin-8 as a specific neu-trophil activator and chemoattractant. Vet.Immunol. Immunopathol. 67, 327–340.
Clowes, R.C., Hayes, W., 1968. Experiments in Micro-bial Genetics. Blackwell Scientific Publications,Oxford.
Cohen, M.S., 1994. Molecular events in the activationof human neutrophils for microbial killing. Clin.Infect. Dis. 2 (Suppl. 18), S170–S179.
Crooks, S.W., Stockley, R.A., 1998. Leukotriene B4.Int. J. Biochem. Cell Biol. 30, 173–178.
Dahlgren, C., Karlsson, A., 1999. Respiratory burst inhuman neutrophils. J. Immunol. Methods 232, 3–14.
Davis, A.R., Mascolo, P.L., Bunger, P.L., Sipes, K.M.,Quinn, M.T., 1998. Cloning and sequencing of thebovine flavocytochrome b subunit proteins, gp91-phox and p22-phox : comparison with other knownflavocytochrome b sequences. J. Leukocyte Biol. 64,114–123.
Davis, D.S., Templeton, J.W., Ficht, T.A., Huber, J.D.,Angus, R.D., Adams, L.G., 1991. Brucella abortusin Bison. II. Evaluation of strain 19 vaccination ofpregnant cows. J. Wildl. Dis. 27, 258–264.
DeLeo, F.R., Jutila, M.A., Quinn, M.T., 1996. Charac-terization of peptide diffusion into electropermeabi-lized neutrophils. J. Immunol. Methods 198, 35–49.
DeLeo, F.R., Quinn, M.T., 1996. Assembly of thephagocyte NADPH oxidase: molecular interactionof oxidase proteins. J. Leukocyte Biol. 60, 677–691.
de los Monteros, A.E., Carrasco, L., King, J.M.,Jensen, H.E., 1999. Nasal zygomycosis and pul-monary aspergillosis in an American bison. J. Wildl.Dis. 35, 790–795.
Dragon, D.C., Elkin, B.T., Nishi, J.S., Ellsworth, T.R.,1999. A review of anthrax in Canada and implica-tions for research on the disease in northern bison.J. Appl. Microbiol. 87, 208–213.
Elzer, P.H., Edmonds, M.D., Hagius, S.D., Walker,J.V., Gilsdorf, M.J., Davis, D.S., 1998. Safety ofBrucella abortus strain RB51 in Bison. J. Wildl. Dis.34, 825–829.
Fletcher, M.P., Stahl, G.L., Longhurst, J.C., 1990. Invivo and In vitro assessment of porcine neutrophilactivation responses to chemoattractants: flow cyto-mentric evidence for the selective absence of formylpeptide receptors. J. Leukocyte Biol. 47, 355–365.
Fujita, I., Takeshige, K., Minakami, S., 1987. Charac-terization of the NADPH-dependent superoxideproduction activated by sodium dodecyl sulfate in acell-free system of pig neutrophils. Biochim. Bio-phys. Acta 931, 41–48.
Galligan, C.L., Coomber, B.L., 2000. Effects of humanIL-8 isoforms on bovine neutrophil function invitro. Vet. Immunol. Immunopathol. 74, 71–85.
Gennaro, R., Dewald, B., Horisberger, U., Gubler,H.U., Baggiolini, M., 1983. A novel type of cyto-plasmic granule in bovine neutrophils. J. Cell Biol.96, 1651–1661.
Gennaro, R., Romeo, D., Skerlavaj, B., Zanetti, M.,1991. Neutrophil and eosinophil granules as storesof defense proteins. Blood Cell Biochem. 3, 335–368.
Ganz, T., 1999. Oxygen-independent microbicidalmechanisms of phagocytes. Proc. Assoc. Am. Physi-cians 111, 390–395.
Gudmundsson, G.H., Agerberth, B., 1999. Neutrophilantibacterial peptides, multifunctional effectormolecules in the mammalian immune system. J.Immunol. Methods 232, 45–54.
Harada, A., Sekido, N., Akahoshi, T., Wada, T.,Mukaida, N., Matsushima, K., 1994. Essential in-volvement of interleukin-8 (IL-8) in acute inflamma-tion. J. Leukocyte Biol. 56, 559–564.
Jesaitis, A.J., Allen, R.A., Parkos, C.A., 1988. Activa-tion of the neutrophil respiratory burst by chemoat-tractants: regulation of the N-formyl peptidereceptor in the plasma membrane. J. Bioenerg.Biomembr. 20, 679–707.
Kremer, W.D.J., Noordhuizen-Stassen, E.N., Henricks,P.A.J., van der Vliet, H., 1992. A procedure forparallel isolation of white blood cells, granulocyteand purified neutrophil suspensions from the pe-ripheral blood of cattle. Vet. Immunol. Im-munopathol. 31, 189–193.
Leino, L., Paape, M.J., 1993. Comparison of the chemi-luminescence responses of bovine neutrophils todifferently opsonized zymosan particles. Am. J. Vet.Res. 54, 1055–1059.
Lindley, I.J.D., 1998. Interleukin-8. In: Mire-Sluis, A.,Thorpe, R. (Eds.), Cytokines. Academic Press, SanDiego, pp. 125–140.
Marasco, W.A., Fantone, J.C., Freer, R.J., Ward, P.A.,1983. Characterization of the rat neutrophil formylpeptide chemotaxis receptor. Am. J. Pathol. 111,273–281.
Marler, R.J., 1975. Some hematologic and blood chem-istry values in two herds of American bison inKansas. J. Wildl. Dis. 11, 97–100.
Mehrer, C.F., 1976. Some hematologic values of bisonfrom five areas of the United States. J. Wildl. Dis.12, 7–13.

S.D. Swain et al. / Comparati6e Biochemistry and Physiology, Part A 127 (2000) 237–247 247
Olsen, S.C., Jensen, A.E., Palmer, M.V., Stevens, M.G.,1998. Evaluation of serologic responses, lymphocyteproliferative responses, and clearance from lymphaticorgans after vaccination of bison with Brucella abor-tus strain RB51. Am. J. Vet. Res. 59, 410–415.
Penzhorn, B.L., Knapp, S.E., Speer, C.A., 1994. Entericcoccidia in free-ranging American bison (Bison bison)in Montana. J. Wildl. Dis. 30, 267–269.
Persson, K., Larsson, I., Hallen, S.C., 1993. Effects ofcertain inflammatory mediators on bovine neutrophilmigration in vivo and in vitro. Vet. Immunol. Im-munopathol. 37, 99–112.
Persson-Waller, K., Colditz, I.G., Seow, H.F., 1997.Accumulation of leucocytes and cytokines in thelactating ovine udder during mastitis due to Staphy-lococcus aureus and Escherichia coli. Res. Vet. Sci. 62,63–66.
Peterson, M.J., Grant, W.E., Davis, D.S., 1991. Bison-brucellosis management: simulation of alternativestrategies. J. Wildl. Manage. 55, 205–213.
Quinn, M.T., Evans, T., Loetterle, L.R., Jesaitis, A.J.,Bokoch, G.M., 1993. Translocation of Rac correlateswith NADPH oxidase activation: evidence forequimolar translocation of oxidase components. J.Biol. Chem. 268, 20983–20987.
Rosen, G.M., Pou, S., Ramos, C.L., Cohen, M.S.,Britigan, B.E., 1995. Free radicals and phagocyticcells. FASEB J. 9, 200–209.
Sahoo, G., More, T., Singh, V.K., 1998. A comparativestudy on certain enzymes of the granulocyte fromdifferent ruminant species. Comp. Immunol. Micro-biol. Infect. Dis. 21, 319–325.
Salgar, S.K., Paape, M.J., Alston-Mills, B., Miller, R.H.,1991. Flow cytometric study of oxidative burst activ-ity in bovine neutrophils. Am. J. Vet. Res. 52,1201–1207.
Schiffmann, E., Corcoran, B.A., Wahl, S.M., 1975.N-formylmethionyl peptides as chemoattractants forleucocytes. Proc. Natl. Acad. Sci. USA 72, 1059–1062.
Sikarskie, J.G., Schillhorn, V., van Selm, G., Kock,M.D., 1990. Comparative blood characteristics ofranched and free-ranging American bison (Bisonbison). Am. J. Vet. Res. 51, 955–957.
Smith, J.A., 1994. Neutrophils, host defense, and inflam-mation: a double-edged sword. J. Leukocyte Biol. 56,672–686.
Soltys, J., Swain, S.D., Sipes, K.M., Nelson, L.K.,Hanson, A.J., Kantele, J.M., Jutila, M.A., Quinn,M.T., 1999. Isolation of bovine neutrophils withbiomagnetic beads: comparison with standard Per-coll density gradient isolation methods. J. Immunol.Methods 226, 71–84.
Spitznagel, J.K., 1990. Antibiotic proteins of humanneutrophils. J. Clin. Invest. 86, 1381–1386.
Stevens, M.G., Olsen, S.C., Cheville, N.F., 1997. Com-
parative effects of bovine cytokines on cattle andbison peripheral blood mononuclear cell prolifera-tion. Comp. Immunol. Microbiol. Infect. Dis. 20,155–162.
Styrt, B., 1989. Species variation in neutrophil biochem-istry and function. J. Leukocyte Biol. 46, 63–74.
Swain, S.D., Bunger, P.L., Sipes, K.M., Nelson, L.K.,Jutila, K.L., Boylan, S.M., Quinn, M.T., 1998.Platelet-activating factor induces a concentration-de-pendent spectrum of functional responses in bovineneutrophils. J. Leukocyte Biol. 64, 817–827.
Swain, S.D., Jutila, K.L., Quinn, M.T., 2000. Cell-sur-face lactoferrin as a marker for bovine neutrophildegranulation: development of a monoclonal anti-body and flow cytometric assay. Am. J. Vet. Res. 61,29–37.
Tessaro, S.V., 1986. The existing and potential impor-tance of brucellosis and tuberculosis in Canadianwildlife: a review. Can. Vet. J. 27, 119–124.
Tessaro, S.V., 1989. Review of the diseases, parasites,and miscellaneous pathological conditions of NorthAmerican Bison. Can. Vet. J. 30, 416422.
Tizard, I.R., 1997. Veterinary Immunology: An Intro-duction, vol. 5. W.B. Saunders Company, Philadel-phia, PA.
Vestweber, J.G., Johnson, D.E., Merrill, G.L., Staats,J.J., 1991. Hematological and blood chemistry profi-les of American bison grazing on Konza Prairie ofKansas. J. Wildl. Dis. 27, 417–420.
Ward, A.C., Dyer, N.W., Corbeil, L.B., 1999a. Charac-terization of putative Haemophilus somnus isolatesfrom tonsils of American bison (Bison bison). Can.J. Vet. Res. 63, 166–169.
Ward, A.C., Dyer, N.W., Fenwick, B.W., 1999b. Pas-teurellaceae isolated from tonsillar samples of com-mercially-reared American bison (Bison bison). Can.J. Vet. Res. 63, 161–165.
Watson, G.L., Slocombe, R.F., Robinson, N.E., Sleight,S.D., 1995. Definition of chemiluminescence andsuperoxide production responses of bovine neu-trophils to selected soluble and particulate stimu-lants, and comparisons with the responses toPasteurella haemolytica. Am. J. Vet. Res. 56, 1045–1054.
Wilkinson, P.C., 1998. Assays of leukocyte locomotionand chemotaxis. J. Immunol. Methods 216, 139–153.
Ye, R.D., Quehenberger, O., Thomas, K.M., Navarro,J., Cavanagh, S.L., Prossnitz, E.R., Cochrane, C.G.,1993. The rabbit neutrophil N-formyl peptide recep-tor: cDNA cloning, expression, and structure/func-tion implications. J. Immunol. 150, 1383–1394.
Zanetti, M., Litteri, L., Gennaro, R., Horstmann, H.,Romeo, D., 1990. Bactenecins, defense polypeptidesof bovine neutrophils, are generated from precursormolecules stored in the large granules. J. Cell Biol.111, 1363–1371.





























