Embed Size (px)
Citation preview

ARCHIVES OF BIOCHEMISTRY AND BIOPHYSICS
Vol. 256, No. 2, August 1, pp. 421-429, 1987
Hormonal Effects on Mitochondrial Respiration: Potential Role of Endogenous Lipolytic Activities’
SHARON CROST, MONICA K. MARTIN, AND JUNE W. PALMER’
Department of Pharmacology, University of Illinois College of Medicine, Chicago, Illinois 60612
Received January 6, 1987
Hormonal effects on heart mitochondrial metabolism are investigated by comparing respiratory rates, Ca2’ uptake capacity, and lipolytic activities of mitochondria isolated from control rats to those of mitochondria isolated from thyroparathyroidectomized animals. Two biochemically and morphologically distinct populations of heart mito- chondria are prepared-one derived from the region of the cell directly beneath the sarcolemma (subsarcolemmal mitochondria), the other originally between the myofibrils (interfibrillar mitochondria). Subsarcolemmal mitochondria isolated from normal rat cardiac tissue have both lower respiratory rates and Ca2+ uptake capacity than do in- terfibrillar mitochondria. However, when these mitochondrial populations are isolated from hearts from thyroparathyroidectomized rats, there is a selective increase in the maximal ability of the subsarcolemmal mitochondria to accumulate Ca2+, which is ac- companied by a proportionate increase in their maximal respiratory rates. Neither Ca2+ uptake capacity nor respiratory rates are similarly increased in the interfibrillar mi- tochondria. Cytochrome contents and mitochondrial protein recoveries are not signifi- cantly changed in either of these mitochondrial preparations. The relationship between these selective increases in respiratory properties of the subsarcolemmal mitochondria to endogenous lipolytic activities is also investigated. It was previously demonstrated that in the absence of Ca2+, both the rate and extent of formation of free fatty acids from endogenous phospholipids is greater in subsarcolemmal than interfibrillar mito- chondria (J. W. Palmer et al. (1981) Arch. Biochem. Biophys. 211,674-682). In this study it is shown that lipolysis is also more sustained in the subsarcolemmal mitochondria when Ca2+ is added. In the subsarcolemmal mitochondria isolated from thyroparathy- roidectomized rats, however, the rates of release of stearic acid and oleic acid are reduced in both the presence and absence of Ca2+. In the presence of added Ca2+, the rate of release of arachidonic acid is also decreased compared to control subsarcolemmal mi- tochondria, suggesting that the expressed activity of Ca2+-activated phospholipase A2 is lower in those mitochondria isolated from the thyroparathyroidectomized animals, in which respiratory rates and Ca2+ uptake capacity are increased. 0 1987 Academic Press, Inc
In the heart there are at least two pop- ulations of mitochondria, one located be- neath the sarcolemma (subsarcolemmal
1 This work was supported by NIH Grant PHS HL 30399 and The Chicago Heart Association. Portions have been published in abstract form (J. W. Palmer, S. A. Crost, and M. K. Martin (1984) Fed. Proc. 43, 1521).
’ To whom correspondence should be addressed.
mitochondria), another between the myo- fibrils (interfibrillar mitochondria). These morphologically distinct mitochondria may be separately isolated by, first, mechanical disruption to release subsarcolemmal mi- tochondria, followed by nagarse (proteo- lytic) digestion, which releases the inter- fibrillar mitochondria (1). Such sequential isolation now permits their biochemical properties to be selectively determined.
421 0003-9861/87 $3.00 Copyright 0 1987 by Academic Press, Inc. All rights of reproduction in any form reserved.

422 CROST, MARTIN, AND PALMER
Differences have been reported in respi- ratory rates (l), Ca2+ transport rates and uptake capacity (2-4), and lipolytic activ- ities (5). Sequential isolation is also the preferred method for ex vivo studies of mi- tochondria since selective changes in either population of mitochondria may be ob- served. Such changes would be either masked when both populations are simul- taneously isolated using nagarse or ignored when only subsarcolemmal mitochondria are isolated using mechanical disruption. For example, it has recently been demon- strated that interfibrillar, but not subsar- colemmal, mitochondria are altered in the cardiomyopathic hamster heart (6). This effect was missed in previous studies which did not separately isolate the two mito- chondrial populations.
We have chosen to study selective re- sponses of these two mitochondrial popu- lations in the hypothyroid rat heart. Hy- pothyroidism has a profound effect on car- diac performance, which may either directly or indirectly influence mitochon- drial metabolism. There are many studies of effects of altered thyroid status on iso- lated mitochondria, but most used liver mitochondria, a few used skeletal muscle mitochondria (e.g., 7, S), and the majority of studies of heart mitochondria examined thyrotoxicity. One previous study of mi- tochondria isolated from hearts of hypo- thyroid animals (9) used proteolytic diges- tion to isolate the mitochondria. While this study reported decreases in the respiratory rates and cytochromes, the study is in- complete because the contribution of the subsarcolemmal mitochondria has been masked.
Past studies of effects of low thyroid hormone levels on heart mitochondrial metabolism are additionally incomplete because they have focused upon respiratory rates and cytochrome contents. In the present study we will also monitor Ca2+ uptake capacity and lipolytic activities. Ca2+ transport properties will be deter- mined because metabolic effects of other hormones such as angiotensin II, vaso- pressin, and glucagon may be mediated by alterations in the intracellular distribution of Ca2’ (10-14) which may result in changes
in mitochondrial Ca2+ fluxes. Accordingly, the possibility that thyroid hormone im- balance may affect, either directly or in- directly, the ability of heart mitochondria to take up and retain Ca2+ will be examined.
In addition, lipolytic activity will be measured because both mitochondrial res- piration and Ca2+ transport may be influ- enced by the phospholipid composition of the inner mitochondrial membrane. Free fatty acids and lysophospholipids formed by the action of mitochondrial phospholi- pases on endogenous phospholipids (15-20) may disrupt the membrane potential, re- sulting in uncoupling of respiration and premature release of accumulated Ca2+. There are also recent reports that oxida- tion/reduction components of the mito- chondrial electron transport chain are not immobilized in a rigid protein lattice, but are rather free to diffuse in the highly fluid lipid component of the membrane (21-23). Thus lipolytic products, such as free fatty acids, may affect maximal respiratory ef- ficiency by altering diffusion rates of mo- bile membrane components such as cyto- chrome c or ubiquinone.
To repeat, subsarcolemmal and interfi- brillar mitochondria differ in all mito- chondrial properties to be measured in this study, namely, respiratory rates, Ca2+ up- take capacity, and lipolytic activities. By selectively monitoring these properties in these distinct mitochondrial types, we should be able to more precisely assess the mitochondrial status in the hypothyroid rat heart.
MATERIALS AND METHODS
Preparation and characterization of expe+nental animals. Male Sprague-Dawley rats (200-250 g) pur- chased from King Animal Labs, Inc., Oregon, Wis- consin, were housed in plastic cages on sanicel bedding. The hypothyroid state was produced by surgical thy- roparathyroidectomy. The anesthetic used was ket- amine (0.8%), acepromazine (0.2%), 0.8 ml/kg body wt. Sham operations consisted of all operational steps including retraction. Thyroidectomized animals were given 2 ml calcium gluconate (1% w/v) po on the morning of surgery and in their drinking water thereafter as previously described (B-26). The ex- perimental animals were maintained on Purina low iodide diet and all control animals received normal

HORMONAL EFFECTS ON HEART MITOCHONDRIA
TABLE I
CHARACTERISTICSOFEXPERIMENTALANIMALS
423
Increase in body weight (%) Heart weight/body weight (% ) Serum T4 (wg/dl) Serum calcium (mM)
Sham-operated control Thyroidectomized
47.5 + 2.6 11.0 *1.1* 0.28 -+ 0.004 0.23 f 0.003* 6.47 f 0.22 0.57 *0.10* 5.15 kO.27 4.28 f 0.31*
Note. Male Sprague-Dawley rats were thyroidectomized or sham operated and metabolic parameters were determined as described under Materials and Methods. Preoperative to time of sacrifice weight gains, heart weights, and serum T4 levels were determined on 25 control and 37 thyroidectomized rats. Serum calcium was measured on serum obtained by decapitation on 30 control and 26 thyroidectomized rats. Data are expressed as means f SE.
* Means which are significantly different; P < 0.05, Students t test.
Purina rat chow. Total body weights were recorded weekly and wet weight of the ventricles was deter- mined following sacrifice of the animals.
Total thyroxine levels were measured on tail vein serum samples obtained approximately 4 weeks after surgery using solid phase radioimmunoassay kits purchased from Diagnostic Products Corp., Los An- geles, California. Thyroidectomized animals whose thyroxine levels were greater than 1 pg/dl were not used for mitochondrial isolations. Ca*+ levels were routinely estimated from serum obtained upon de- capitation by measuring the increase in absorbance at 507-511 nm (Aminco DW2 spectrophotometer) us- ing murexide (0.1 mM) as indicating dye. Accuracy of this method was verified using atomic absorption spectrophotometry (Allied Analytical Systems, An- dover, MA).
Attainment of the hypothyroid state is documented by the characterization of these animals as reported in Table I. The increase in total body weight of the thyroidectomized animals is significantly less than the weight gain of the control animals, whereas the ratio of heart weight to total body weight is maintained. Circulating thyroxine (TJ3 levels are reduced 90% at this time. Hypocalcemia was prevented by addition of calcium gluconate to the drinking water of the thy- roidectomized animals, yet the serum calcium levels of these animals are still slightly, but significantly, lower than in control animals.
Mitochondrial isolation and incubation Subsarco- lemma1 and interfibrillar heart mitochondria were isolated as described previously (1) except that the
a Abbreviations used: T,, thyroxine; Hepes, 4-(2- hydroxyethyl)-1-piperazineethanesulfonic acid; EGTA, ethylene glycol bis(&aminoethyl ether)-N,A”- tetraacetic acid; C16:0, palmitic acid; Cl&O, stearic acid; Cl&l, oleic acid; C18:2, linoleic acid; C20:4, arachidonic acid.
low ionic strength medium (220 mM mannitol, 70 mM sucrose, 3 mM Hepes) was used for all preparative procedures except the proteolytic (nagarse) digestion of the low speed Polytron pellet. Use of the normal ionic strength buffer (110 mM KCI, 50 mM Hepes, pH 7.4,2 mM EGTA, plus 0.2% albumin) was retained for proteolytic digestion, since it facilitated the separation of the mitochondria from myofibrillar protein. Protein was determined by the biuret method following solubilization of the mitochondria with deoxycholate (27). Cytochrome contents were determined by the procedure described by Williams (28). Glutamate de- hydrogenase, which was used as a mitochondrial marker, was assayed by published procedures (29).
Incubation conditions were chosen to maximize respiration (1, 34), CaZ+ uptake (4), or lipolysis (5). Oxygen utilization was monitored using a Clark ox- ygen electrode and Yellow Springs Instruments mon- itor. The incubation medium contained 80 mM KCl, 50 mM Hepes, pH 7.4, 5 mM phosphate (K+), 1 mM EGTA, and 1 mg/ml defatted, dialyzed bovine serum albumin. Calcium uptake was monitored by dual wavelength spectroscopy (Aminco DW2) using mu- rexide (0.1 mM) as indicating dye and monitoring ab- sorbance changes at 533-507 nm. For these measure- ments, mitochondria (0.67 mg protein/ml) were in- cubated at 30°C in medium containing 220 mM mannitol,70 mM sucrose,5 mM Hepes,pH 7.4,1.7 mM phosphate (K+), 2.5 pg rotenone, and 1.7 mM succinate (K+) as respiratory substrate. Ca’+, 1 pmol/mg protein or 0.67 mM, was added 2 min after the addition of mitochondria when Ca2+ uptake was measured. To maximize fatty acid release, the mitochondria (0.67 mg protein/ml) were incubated under the same con- ditions except that the succinate concentration was 10 mM, phosphate was omitted, and 67 nmol Ca’+/mg protein was added.
Lipid analyses. Contents of endogenous free fatty acids and those released during incubation of the mi- tochondria were determined by gas chromatographic
424 CROST, MARTIN, AND PALMER
analysis of the fatty acid methyl ester derivatives. Lipid extracts were prepared by the Bligh and Dyer procedure (30) from 4-ml aliquots of mitochondrial incubations containing 0.67 mg of mitochondrial pro- tein/ml. The free fatty acids were derivatized with diazomethane (15) and purified by silicic acid chro- matography (31) as follows: columns were prepared in Pasteur pipets using silica gel (Sigma chromato- graphic grade, 100-200 mesh), washed with petroleum ether, and activated overnight at 100°C. Samples were added in 0.2 ml petroleum ether and the columns were rinsed with a total of 2 ml petroleum ether followed by 2 ml chloroform. The methyl esters were eluted with 2 ml petroleum etherldiethyl ether (l/l). The purified methyl esters were dissolved in 10 ~1 carbon disulflde for gas chromatographic analysis on a Schi- madzu GC 5A, equipped with a Hewlett-Packard 3390A integrator. Compounds are separated on a 1.5 m, 3 mm diameter, glass column packed with 10% SP 2330 on 100/120 mesh chromosorb. Peak area values were normalized compared to that of an internal standard (heptadecanoic acid) which was carried throughout the procedure to correct for losses during isolation and analysis.
Data are presented as means f SE. Statistical analyses were performed by the Student’s t test. Un- less otherwise indicated, a probability of co.05 was considered significant.
RESULTS
Respiratory Properties of Mitochondria from Thyroidectomized Animals
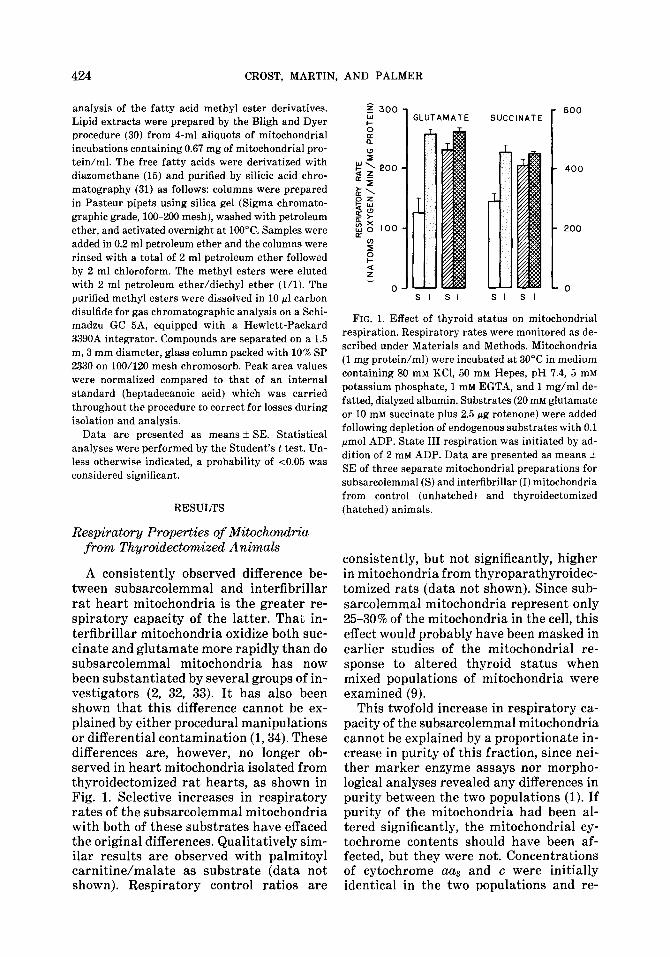
A consistently observed difference be- tween subsarcolemmal and interfibrillar rat heart mitochondria is the greater re- spiratory capacity of the latter. That in- terfibrillar mitochondria oxidize both suc- cinate and glutamate more rapidly than do subsarcolemmal mitochondria has now been substantiated by several groups of in- vestigators (2, 32, 33). It has also been shown that this difference cannot be ex- plained by either procedural manipulations or differential contamination (1,34). These differences are, however, no longer ob- served in heart mitochondria isolated from thyroidectomized rat hearts, as shown in Fig. 1. Selective increases in respiratory rates of the subsarcolemmal mitochondria with both of these substrates have effaced the original differences. Qualitatively sim- ilar results are observed with palmitoyl carnitine/malate as substrate (data not shown). Respiratory control ratios are
$ 300
1 600 I- GLUTAMATE SUCCINATE
SI SI SI SI
FIG. 1. Effect of thyroid status on mitochondrial respiration. Respiratory rates were monitored as de- scribed under Materials and Methods. Mitochondria (1 mg protein/ml) were incubated at 30°C in medium containing 80 mM KCl, 50 mM Hepes, pH 7.4, 5 mM
potassium phosphate, 1 mM EGTA, and 1 mg/ml de- fatted, dialyzed albumin. Substrates (20 mM glutamate or 10 mM succinate plus 2.5 pg rotenone) were added following depletion of endogenous substrates with 0.1 pmol ADP. State III respiration was initiated by ad- dition of 2 mM ADP. Data are presented as means + SE of three separate mitochondrial preparations for subsarcolemmal (S) and interfibrillar (I) mitochondria from control (unhatched) and thyroidectomized (hatched) animals.
consistently, but not significantly, higher in mitochondria from thyroparathyroidec- tomized rats (data not shown). Since sub- sarcolemmal mitochondria represent only 2.530% of the mitochondria in the cell, this effect would probably have been masked in earlier studies of the mitochondrial re- sponse to altered thyroid status when mixed populations of mitochondria were examined (9).
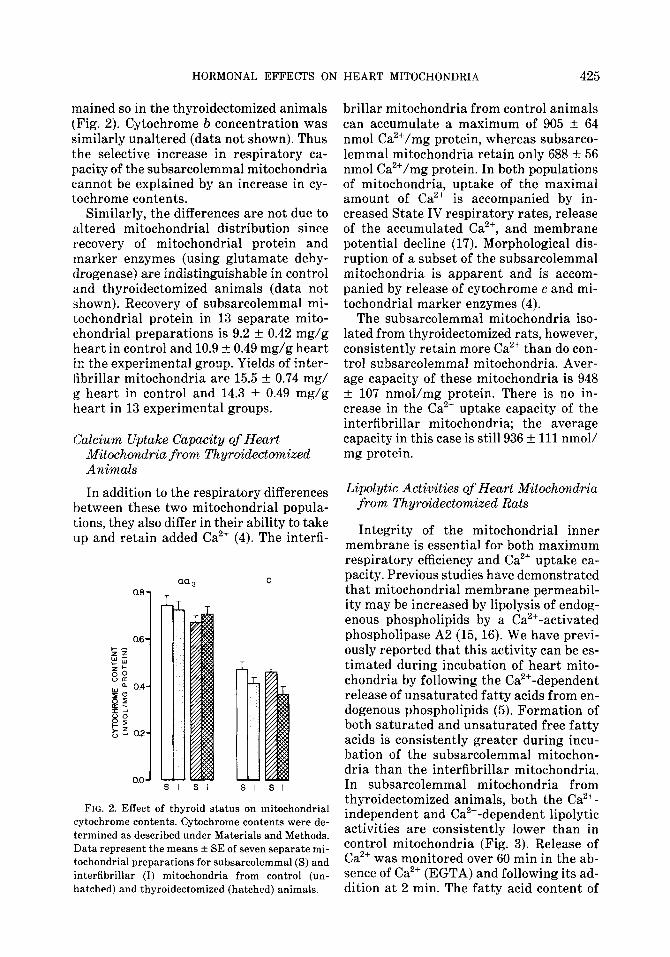
This twofold increase in respiratory ca- pacity of the subsarcolemmal mitochondria cannot be explained by a proportionate in- crease in purity of this fraction, since nei- ther marker enzyme assays nor morpho- logical analyses revealed any differences in purity between the two populations (1). If purity of the mitochondria had been al- tered significantly, the mitochondrial cy- tochrome contents should have been af- fected, but they were not. Concentrations of cytochrome aa and c were initially identical in the two populations and re-
HORMONAL EFFECTS ON HEART MITOCHONDRIA 425
mained so in the thyroidectomized animals (Fig. 2). Cytochrome b concentration was similarly unaltered (data not shown). Thus the selective increase in respiratory ca- pacity of the subsarcolemmal mitochondria cannot be explained by an increase in cy- tochrome contents.
Similarly, the differences are not due to altered mitochondrial distribution since recovery of mitochondrial protein and marker enzymes (using glutamate dehy- drogenase) are indistinguishable in control and thyroidectomized animals (data not shown). Recovery of subsarcolemmal mi- tochondrial protein in 13 separate mito- chondrial preparations is 9.2 + 0.42 mg/g heart in control and 10.9 + 0.49 mg/g heart in the experimental group. Yields of inter- tibrillar mitochondria are 15.5 + 0.74 mg/ g heart in control and 14.3 + 0.49 mg/g heart in 13 experimental groups.
Calcium Uptake Capacity of Heart Mitochondria from Thywidectomized Animals
In addition to the respiratory differences between these two mitochondrial popula- tions, they also differ in their ability to take up and retain added Ca2+ (4). The interfi-
aa, 0.61 T
FIG. 2. Effect of thyroid status on mitochondrial cytochrome contents. Cytochrome contents were de- termined as described under Materials and Methods. Data represent the means f SE of seven separate mi- tochondrial preparations for subsarcolemmal (S) and interfibrillar (I) mitochondria from control (un- hatched) and thyroidectomized (hatched) animals.
brillar mitochondria from control animals can accumulate a maximum of 905 +- 64 nmol Ca2+/mg protein, whereas subsarco- lemma1 mitochondria retain only 688 * 56 nmol Ca2+/mg protein. In both populations of mitochondria, uptake of the maximal amount of Ca2+ is accompanied by in- creased State IV respiratory rates, release of the accumulated Ca2+, and membrane potential decline (17). Morphological dis- ruption of a subset of the subsarcolemmal mitochondria is apparent and is accom- panied by release of cytochrome c and mi- tochondrial marker enzymes (4).
The subsarcolemmal mitochondria iso- lated from thyroidectomized rats, however, consistently retain more Ca2+ than do con- trol subsarcolemmal mitochondria. Aver- age capacity of these mitochondria is 948 + 107 nmol/mg protein. There is no in- crease in the Ca2+ uptake capacity of the interfibrillar mitochondria; the average capacity in this case is still 936 + 111 nmol/ mg protein.
Lipolytic Activities of Heart Mitochondria from Thyroidectomized Rats
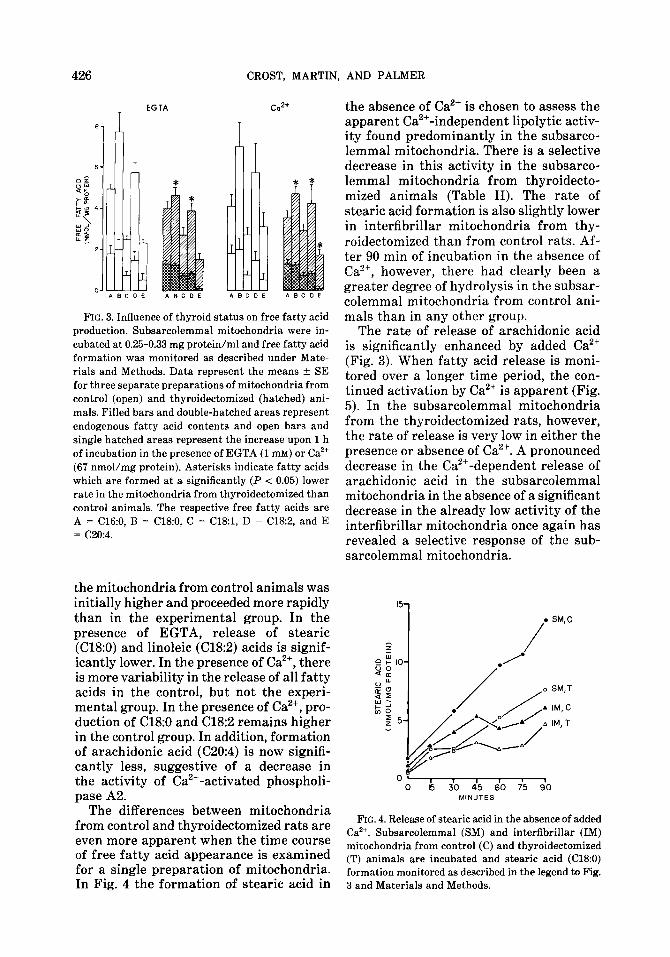
Integrity of the mitochondrial inner membrane is essential for both maximum respiratory efficiency and Ca2+ uptake ca- pacity. Previous studies have demonstrated that mitochondrial membrane permeabil- ity may be increased by lipolysis of endog- enous phospholipids by a Ca2+-activated phospholipase A2 (15,16). We have previ- ously reported that this activity can be es- timated during incubation of heart mito- chondria by following the Ca2+-dependent release of unsaturated fatty acids from en- dogenous phospholipids (5). Formation of both saturated and unsaturated free fatty acids is consistently greater during incu- bation of the subsarcolemmal mitochon- dria than the interfibrillar mitochondria. In subsarcolemmal mitochondria from thyroidectomized animals, both the Ca2+- independent and Ca2+-dependent lipolytic activities are consistently lower than in control mitochondria (Fig. 3). Release of Ca2’ was monitored over 60 min in the ab- sence of Ca2+ (EGTA) and following its ad- dition at 2 min. The fatty acid content of
426 CROST, MARTIN, AND PALMER
EGTA c.2+
FIG. 3. Influence of thyroid status on free fatty acid production. Subsarcolemmal mitochondria were in- cubated at 0.25-0.33 mg protein/ml and free fatty acid formation was monitored as described under Mate- rials and Methods. Data represent the means f SE for three separate preparations of mitochondria from control (open) and thyroidectomized (hatched) ani- mals. Filled bars and double-hatched areas represent endogenous fatty acid contents and open bars and single hatched areas represent the increase upon 1 h of incubation in the presence of EGTA (1 mM) or Ca2+ (67 nmol/mg protein). Asterisks indicate fatty acids which are formed at a significantly (P < 0.05) lower rate in the mitochondria from thyroidectomized than control animals. The respective free fatty acids are A = C16:0, B = Cl&O, C = C18:1, D = C18:2, and E = C20:4.
the mitochondria from control animals was initially higher and proceeded more rapidly than in the experimental group. In the presence of EGTA, release of stearic (Cl&O) and linoleic (C18:2) acids is signif- icantly lower. In the presence of Ca’+, there is more variability in the release of all fatty acids in the control, but not the experi- mental group. In the presence of Ca2+, pro- duction of C18:O and Cl&2 remains higher in the control group. In addition, formation of arachidonic acid (C20:4) is now signifi- cantly less, suggestive of a decrease in the activity of Ca2+-activated phospholi- pase A2.
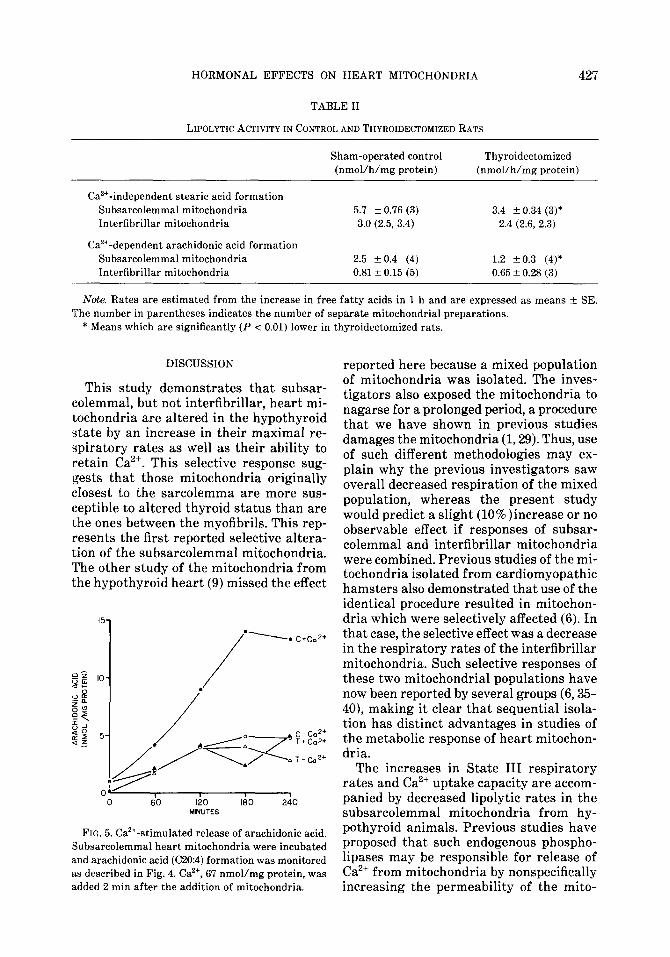
The differences between mitochondria from control and thyroidectomized rats are even more apparent when the time course of free fatty acid appearance is examined for a single preparation of mitochondria. In Fig. 4 the formation of stearic acid in
the absence of Ca2’ is chosen to assess the apparent Ca2+ -independent lipolytic activ- ity found predominantly in the subsarco- lemma1 mitochondria. There is a selective decrease in this activity in the subsarco- lemma1 mitochondria from thyroidecto- mized animals (Table II). The rate of stearic acid formation is also slightly lower in interfibrillar mitochondria from thy- roidectomized than from control rats. Af- ter 90 min of incubation in the absence of Ca2+ however, there had clearly been a grea;er degree of hydrolysis in the subsar- colemmal mitochondria from control ani- mals than in any other group.
The rate of release of arachidonic acid is significantly enhanced by added Ca2+ (Fig. 3). When fatty acid release is moni- tored over a longer time period, the con- tinued activation by Ca2+ is apparent (Fig. 5). In the subsarcolemmal mitochondria from the thyroidectomized rats, however, the rate of release is very low in either the presence or absence of Ca2+. A pronounced decrease in the Ca2+-dependent release of arachidonic acid in the subsarcolemmal mitochondria in the absence of a significant decrease in the already low activity of the interfibrillar mitochondria once again has revealed a selective response of the sub- sarcolemmal mitochondria.
‘5-l
I I 1 f 0 15 30 45 60 75 90
MINUTES
FIG. 4. Release of stearic acid in the absence of added Ca’+. Subsarcolemmal (SM) and interfibrillar (IM) mitochondria from control (C) and thyroideetomized (T) animals are incubated and stearic acid (Cl&O) formation monitored as described in the legend to Fig. 3 and Materials and Methods.
427 HORMONAL EFFECTS ON HEART MITOCHONDRIA
TABLE II
LIPOLYTIC ACTIVITY IN CONTROL AND THYROIDECTOMIZED RATS
Sham-operated control Thyroidectomized (nmol/h/mg protein) (nmol/h/mg protein)
Ca2+-independent stearic acid formation Suhsarcolemmal mitochondria 5.7 f 0.76 (3) 3.4 f 0.34 (3)* Interfibrillar mitochondria 3.0 (2.5, 3.4) 2.4 (2.6, 2.3)
Ca’+-dependent arachidonic acid formation Subsarcolemmal mitochondria 2.5 f 0.4 (4) 1.2 kO.3 (4)* Interfibrillar mitochondria 0.81 f 0.15 (5) 0.65 f 0.28 (3)
Note. Rates are estimated from the increase in free fatty acids in 1 h and are expressed as means + SE. The number in parentheses indicates the number of separate mitochondrial preparations.
* Means which are significantly (P < 0.01) lower in thyroidectomized rats.
DISCUSSION
This study demonstrates that subsar- colemmal, but not interfibrillar, heart mi- tochondria are altered in the hypothyroid state by an increase in their maximal re- spiratory rates as well as their ability to retain Ca2+. This selective response sug- gests that those mitochondria originally closest to the sarcolemma are more sus- ceptible to altered thyroid status than are the ones between the myofibrils. This rep- resents the first reported selective altera- tion of the subsarcolemmal mitochondria. The other study of the mitochondria from the hypothyroid heart (9) missed the effect
MWTES
FIG. 5. Ca’+-stimulated release of arachidonic acid. Subsarcolemmal heart mitochondria were incubated and arachidonic acid (C20:4) formation was monitored as described in Fig. 4. Ca’+, 67 nmol/mg protein, was added 2 min after the addition of mitochondria.
reported here because a mixed population of mitochondria was isolated. The inves- tigators also exposed the mitochondria to nagarse for a prolonged period, a procedure that we have shown in previous studies damages the mitochondria (1,29). Thus, use of such different methodologies may ex- plain why the previous investigators saw overall decreased respiration of the mixed population, whereas the present study would predict a slight (lO%)increase or no observable effect if responses of subsar- colemmal and interfibrillar mitochondria were combined. Previous studies of the mi- tochondria isolated from cardiomyopathic hamsters also demonstrated that use of the identical procedure resulted in mitochon- dria which were selectively affected (6). In that case, the selective effect was a decrease in the respiratory rates of the interfibrillar mitochondria. Such selective responses of these two mitochondrial populations have now been reported by several groups (6,35- 40), making it clear that sequential isola- tion has distinct advantages in studies of the metabolic response of heart mitochon- dria.
The increases in State III respiratory rates and Ca2+ uptake capacity are accom- panied by decreased lipolytic rates in the subsarcolemmal mitochondria from hy- pothyroid animals. Previous studies have proposed that such endogenous phospho- lipases may be responsible for release of Ca2+ from mitochondria by nonspecifically increasing the permeability of the mito-

428 CROST. MARTIN, AND PALMER
chondrial inner membrane (15-18). In- creases in steady-state concentrations of chaotropic agents, such as fatty acids, in microenvironments of the inner membrane may result not only in release of accumu- lated Ca’+, but also in increased proton permeability resulting in decreased max- imal respiratory rates. Accordingly, the increased respiratory rates and Ca2+ up- take capacity of the subsarcolemmal mi- tochondria are accompanied by decreased lipolysis. Fewer free fatty acids are being formed in those mitochondria with greater respiratory rates and Ca2+ uptake capacity.
The mitochondrial changes presented in this study may represent an adaptive re- sponse secondary to primary effects of thyroid hormone in the cell. There are well- defined nuclear thyroid hormone receptors which regulate synthesis of many proteins and enzymes, including plasma membrane Na+/K+ ATPase and a myosin ATPase iso- zyme (41).In the hypothyroid state, de- creased activity of ATPases results in de- creased turnover of ATP. The present study suggests that the mitochondria respond to this lowered energy utilization by an in- creased rate and efficiency of respiration. Despite this compensatory action, heart rate and contractility still decrease in the hypothyroid heart. The overall decreases are probably due to decreases in several enzyme activities, substrate levels, ADP, and phosphate concentrations, despite the rise in mitochondrial maximal respiratory rates.
Free fatty acids are the major fuel for energy in the heart. Thyroid hormones control lipolytic activities in many tissues, including the heart, resulting, in the hy- pothyroid state, in decreased supply of fatty acids for energy. The mitochondrial lipases reported here may thus serve a metabolic role in addition to the suggested regulation of mitochondrial membrane in- tegrity. In the hypothyroid state, there may be decreased lipolysis, fewer fatty acids available for fuel, and also more efficient ATP production. As a consequence, there would also be less heat production, in agreement with the known calorigenic ef- fect of thyroid hormone.
Decreased circulating levels of parathy-
roid hormone, calcitonin, or growth hor- mone which result from surgical thyro- parathyroidectomy may also affect cardiac mitochondrial function. For example, re- cent studies have shown that parathyroid hormone may increase the beating rate of isolated heart cells (42). In addition, these investigators have shown a direct effect of parathyroid hormone on respiratory rates of isolated heart mitochondria (43).
The present study suggests that there is a correlation between the ability of heart mitochondria to accumulate Ca2+, their maximal respiratory rates, and the activity of endogenous phospholipases. Hormonal modulation of mitochondrial Ca2+ homeo- stasis has been described by many inves- tigators; in particular, rapid effects of glu- cagon on liver mitochondrial function have been observed (44). From the results of the present study, it is tempting to propose that the activity and/or synthesis of the cardiac phospholipases is under hormonal control, and that the consequent altera- tions in mitochondrial function may result in altered distribution of intracellular Ca2+.
REFERENCES
1. PALMER, J. W., TANDLER, B., AND HOPPEL, C. L.
(1977) J. Biol. Chem 252.8731-8739. 2. MCMILLIN-WOOD, J., WOLKOWICZ, P. E., CHU, A.,
TATE, C. A., GOLDSTEIN, M. A., AND ENTMAN,
M. L. (1980) Biochim. Bkphys. Acta 591, 251- 265.
3. PALMER, J. W. (1978) Fed Proc. 37, 1616. 4. PALMER, J. W., TANDLER, B., AND HOPPEL, C. L.
(1984) Amer. J. Physiol. 250, H741-H748.
5. PALMER, J. W., SCHMID, P. C., PFEIFFER, D. R.,
AND SCHMID, H. H. 0. (1981) Arch. B&hem Biophys. 211,674-682.
6. HOPPEL, C. L., TANDLER, B., PARLAND, W., TUR-
KALY, J. S., AND ALBERS, L. D. (1982) J. Biol. Chem. 257,1540-1548.
7. TATA, J. R., ERNSTER, L., LINDBERG, O., ARRHENIUS,
E., PEDERSON, S., AND HEDMAN, R. (1963)
Biochem. J. 86,408-428. 8. WINDER, W. W., BALDWIN, K. M., TERJUNG, R. L.,
AND HOLLOSZY, J. 0. (1975) Amer. J. Physiol 228,1341-1345.
9. NISHIKI, K., ERECINSKA, M., WILSON, D. F., AND
COOPER, S. (1978) Amer. J. Physiol. 235, C212- c219.
10. KESSAR, P., AND CROMPTON, M. (1981) B&hem. J, 200,379-388.

HORMONAL EFFECTS ON HEART MITOCHONDRIA 429
11. WILLIAMSON, J. R., COOPER, R. H., AND HOEK, J. B. (1981) Biochim. Biophys. Acta 639, 243- 295.
12. EXTON, J. H. (1981) MoL Cell. EndocrinoL 23,233- 264.
13. RASMUSSEN, H., AND WAISMAN, D. (1981) in Bio- chemical Action of Hormones (Litwack, G., Ed.), Vol. 8, pp. 1-115, Academic Press, New York.
14. CHEN, J. L., BABCOCK, D. F., AND LARDY, H. A. (19’78) Proc. Nat1 Acad. Sci. USA 75,2234-2238.
1.5. PFEIFFER, D. R., SCHMID, P. C., BEATRICE, M. C., AND SCHMID, H. H. 0. (1979) J. BioL Chem. 254, 11485-11494.
16. BEATRICE, M. C., PALMER, J. W., AND PFEIFFER, D. R. (1980) J. BioL Chem. 255,8663-8671.
17. PALMER, J. W., AND PFEIFFER, D. R. (1981) J. BioL Chem. 256,6742-6750.
18. PFEIFFER, D. R., PALMER, J. W., BEATRICE, M. C., AND STIERS, D. L. (1983) in The Biochemistry of Metabolic Processes (Lennon, D. L. F. et aL, eds.), pp. 67-80, Elsevier/North-Holland, New York.
19. BEATRICE, M. C., STIERS, D. L., AND PFEIFFER, D. T. (1984) J. Biol. Chem. 259,1279-1287.
20. DALTON, S., HUGHES, B. P., AND BARRITT, G. J. (1984) Biochem. .I 224,423-430.
21. HACKENBROCK, C. R. (1981) Trends Biochem. Sci. 6,151-154.
22. SCHNEIDER, H., AND HACKENBROCK, C. R. (1982) in Membranes and Transport (Martonosi, A. N., Ed.), Vol. 1, pp. 431-435, Plenum, New York.
23. SCHNEIDER, H., LEMASTERS, J. J., AND HACKEN- BROCK, C. R. (1982) J. Biol. Chem. 257, 10789- 10793.
24. LEE, Y. P., AND LARDY, H. A. (1965) J. BioL Chem. 240,1427-1435.
25. STERLING, K., MILCH, P. O., BRENNER, M. A., AND LAZARUS, J. H. (1977) Science 197,996-999.
26. RUZICKA, F. J., AND ROSE, D. P. (1981) Endocri- nology 109,664-666.
27. GORNALL, A. G., BARDAWILL, C. J., AND DAVID, M. M. (1949) J. BioL Chem. 177,751-766.
28. WILLIAMS, J. N. (1964) Arch. Biochem. Biophys. 107,537-543.
29. HOPPEL, C. L., AND TOMEC, R. J. (1972) J. BioL Chem. 247,832-841.
30. BLIGH, E. G., AND DYER, W. J. (1959) Canad. J. B&hem. Physiol. 37,911-917.
31. BROEKMEIER, K. H., SCHMID, P. C., SCHMID, H. H. O., AND PFEIFFER, D. R. (1985) J. Biol. Chem. 260,105-113.
32. MATLIB, M. A., WILSON, D., ROUSLIN, W., KRAFT, G., BERNER, P., AND SCHWARTZ, A. (1978) B&hem. Biophys. Res. &mm. 84,482-488.
33. FRIEDMANN, N., MAYEKAR, M., AND MCMILLIN- WOOD, J. (1980) Li,fe Sci 26, 2093-2098.
34. PALMER, J. W., TANDLER, B., AND HOPPEL, C. L. (1984) Arch. B&hem. Biophys. 236,691-702.
35. Kuo, T. H., GIACOMELLI, F., AND WIENER, J. (1985) Biochim. Biophys. Acta 806, 9-15.
36. WATTS, J. A., KOCH, C. D., AND LA NOUE, K. F. (1980) Amer. J. PhysioL 238, H909-H916.
37. PINSKY, W. W., LEWIS, R. M., MCMILLIN-WOOD, J., HARA, H., HARTLEY, C. J., GILLETTE, P. C., AND ENTMAN, M. L. (1981) Amer. .I PhysioL 240, H326-H335.
38. NAGAO, T., MATLIB, M. A., FRANKLIN, D., MILLARD, R., AND SCHWARTZ, A. (1980) J. Mol. Cell. Car- dial. 12, 29-43.
39. VAN JAARSVELD, H., AND LOCHNER, A. (1982) Basic Res. CardioL 77,388-403.
40. WEINSTEIN, E. S., BENSON, D. W., RATCLIFFE, D. J., MAKSEM, J., AND FRY, D. E. (1985) Arch. Surg. 120,332-338.
41. BERNAL, J., AND REFETOFF, S. (1977) Clin. End* crinol. 6, 227-249.
42. BOGIN, E., MASSRY, S. G., AND HARARY, I. (1981) J Clin. Invest. 67, 1215-1227.
43. BOGIN, E., LEVI, J., HARARY, I., AND MASSRY, S. G. (1982) Miner. Electrolyte Metab. 7, 151- 156.
44. YAMAZAKI, R. K., MICKEY, D. L., AND STORY, M. (1980) Biochim. Biophys. Acta 592, 1-12.





























