Embed Size (px)
Citation preview

HIV-1 can escape from RNA interference by evolving an alternative structure in its RNA genome
796–804 Nucleic Acids Research, 2005, Vol. 33, No. 2 doi:10.1093/nar/gki220
Department of Human Retrovirology, Academic Medical Center, University of Amsterdam, Meibergdreef 15, 1105 AZ Amsterdam, The Netherlands
Ellen M. Westerhout, Marcel Ooms, Monique Vink, Atze T. Das and Ben Berkhout*
Received December 10, 2004; Revised and Accepted January 13, 2005
生科系四甲 張智堯

INTRODUCTION Double-stranded RNA (dsRNA) 能誘導RNA interference (RNAi) ,利用 RNAi 的技術可以有效的抑制基因的表現。而在研究中發現, HIV-1 病毒中卻有突變種可以藉由改變自己 RNA 的結構來降低 RNAi 抑制基因的效果。因此 focus 在這些突變種的改變結構為何能夠抵抗 RNAi 的抑制,並加以驗證。

RNA interference (RNAi)
short interference RNA
a ribonuclease

RNA interference (RNAi)
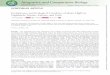
HIV
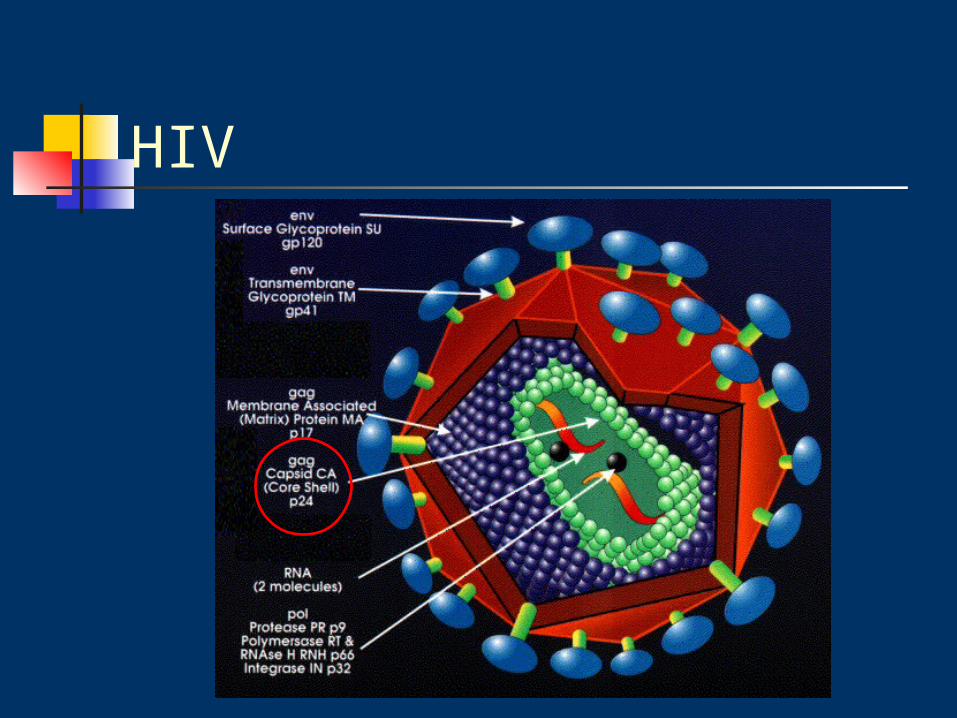
HIV genome

Materials & Methods
Cells and viruses
DNA constructs
Luciferase assay
In silico RNA analysis
RNA probing
EMSA

Cells and viruses
C33A cervix carcinoma cell
calcium phosphate
method
5 ug wild-type or mutant HIV-1 LAI transfection

Materials & Methods
Cells and viruses
DNA constructs
Luciferase assay
In silico RNA analysis
RNA probing
EMSA

DNA constructs R1-R9 mutants :
cellular DNA PCR with 5’ Env primer tTAI-AD and 3’
U5 primer CN1
digested with XhoI and BspEI and cloned into Blue-3’L
TR
XhoI and BspEI fragment cloned into wild-type LAI clone
R1-R9 mutant clones
pGL3-Nef
(firefly luciferase expression vector) :
Nef fragment
PCR with primer EW1 and EW3
digested with XbaI and cloned into pGL3
pGL3-Nef
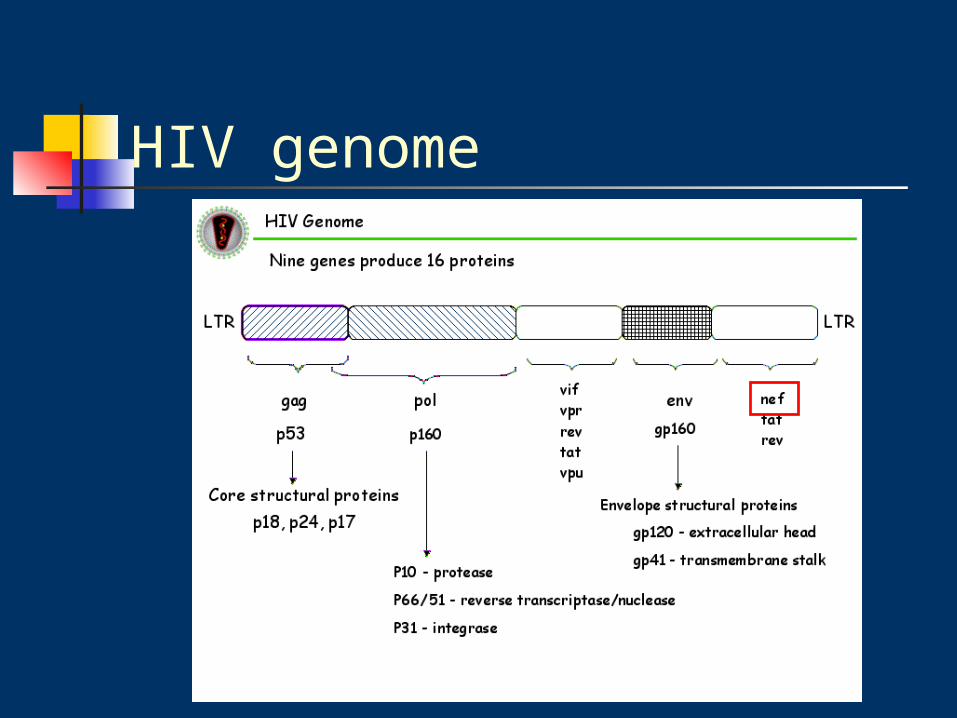
DNA constructs
pBS-siRNA-Nef :pRetro-SUPER-shNef vector
(express siRNA-Nef)
digested with EcoR1 and XhoI
H1 RNA polymerase III promoter
ligated into EcoR1/ XhoI site of pBluescriptII
建構成

DNA constructs
R8 escape virus 在 siRNA-Nef target 區域部分卻沒有沒有產生 mutant ???

CA-24 level by ELISA
SupT1 T-cells transduced with pRetro-SUPER cultured in RPMI 1
640 medium.
HIV-1 LAI infect (1ng of CA-p24)
CA-24 level by ELISA
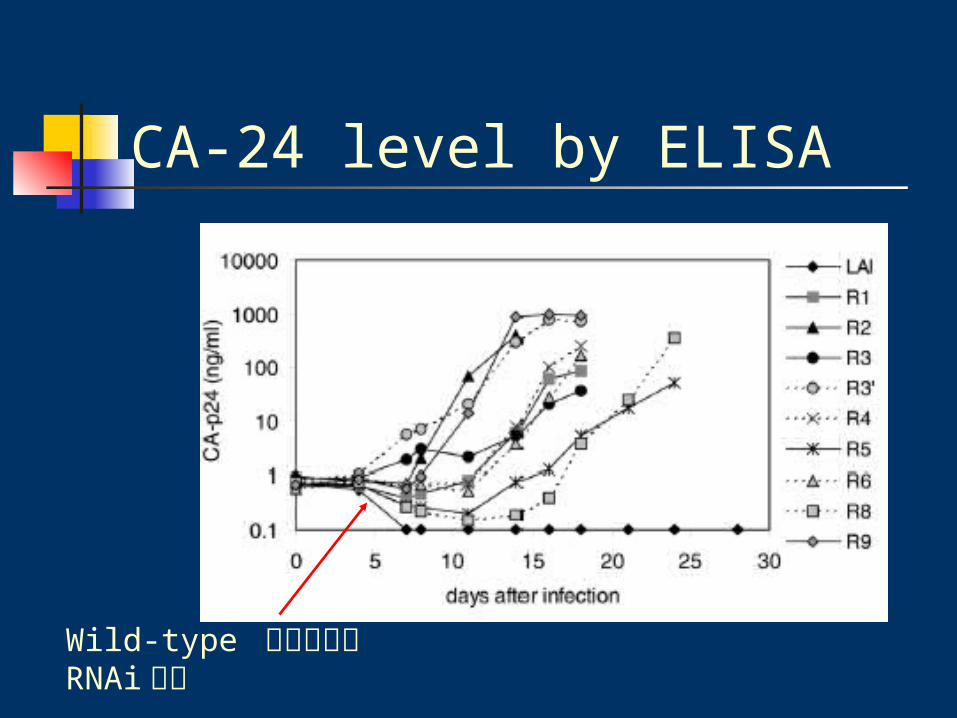
CA-24 level by ELISA
Wild-type 如預期的被 RNAi 抑制

Materials & Methods
Cells and viruses
DNA constructs
Luciferase assay
In silico RNA analysis
RNA probing
EMSA

Luciferase assay100 ng wild-type or mutant pGL3-Nef
+0.5ng pRL-CMV
+0.5-500 ng pBS-siRNA-Nef
completed with pBluescriptII 1ug (total), 15ul water
Mixed 25ul 2XHBS, 10ul 0.6MCaCl2 to cultrue medium, incubated room te
mperature 20 mins
mixed
refreshed 16h
24h
lysed in 150ul Passive Lysis Buffer (PLB) shaking 20 mins at roo
m temperature
centrifuged
10ul supernatant to measure luciferase
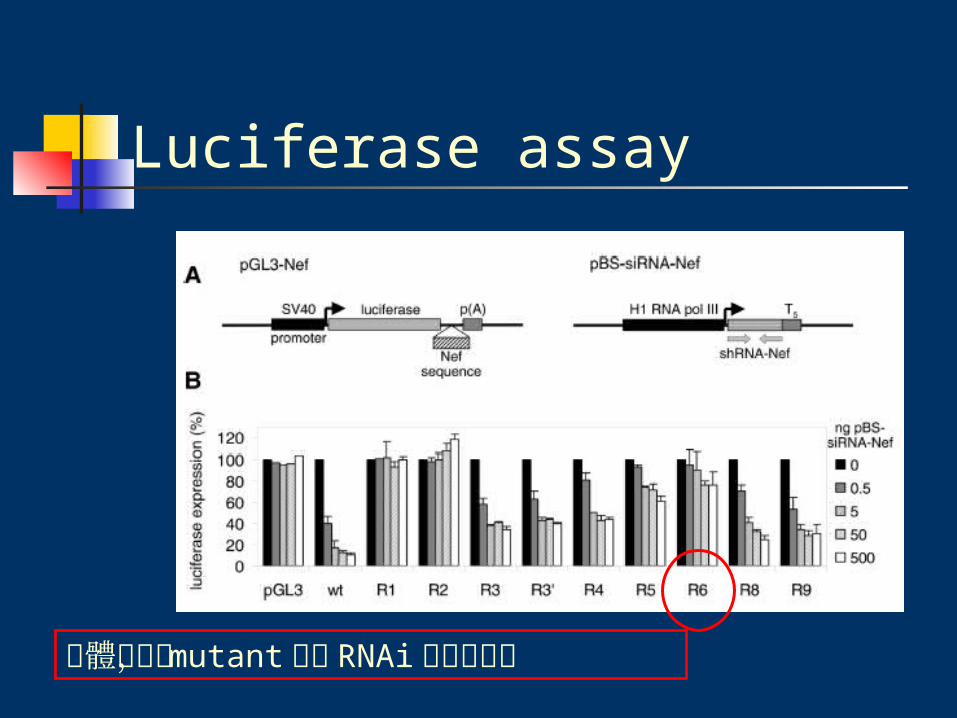
Luciferase assay
總體來說, mutant 皆對 RNAi 有抵抗作用

Materials & Methods
Cells and viruses
DNA constructs
Luciferase assay
In silico RNA analysis
RNA probing
EMSA

In silico RNA analysisMfold program
Hybridization can be predicted
Thermodynamic stability (△G) can be caculated
RNA structures can be predicted
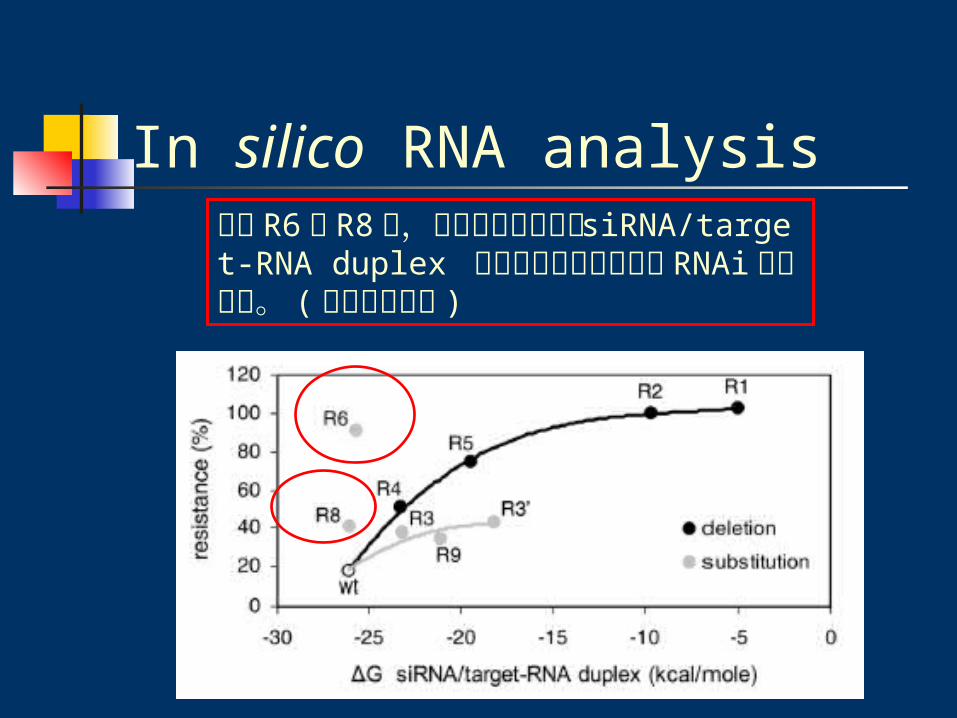
In silico RNA analysis除了 R6 與 R8 外,所有突變種皆隨著 siRNA/target-RNA duplex 穩定性的減少而增加對 RNAi 的抵抗力。 (趨近線性關係 )

In silico RNA analysisQuestion為何只有 R6 與 R8 沒有隨著 siRNA/target-RNA duplex 穩定性的減少而增加對 RNAi的抵抗力 ?? ( 事實上他們的 siRNA/target-RNA duplex 穩定性較高,卻有較高抵抗力 )
尤其 R8 的突變位置不在 siRNA target 的區域,卻對 RNAi 有較高的抵抗力 ??

In silico RNA analysis
sensitive hairpin
Resistant hairpin
Stem loop
△G 上升

In silico RNA analysis
△G 下降

In silico RNA analysis
-15.0
-13.0
-17.9
-15.1
-21.3
-17.9
R6 mutant:(R) Hairpin more stable
R8 mutant:(S) Hairpin less stable
Note : △ G 越小,越 stable ; △ G 越大,越不穩定
所以 R8 & R6 因此形成較穩定的 (R) hairpin loop 結構

In silico RNA analysis
=
Mutant position at –26/-7
Note :
△△G 大於 0 ,表示 (R) hairpin loop△ G 較低,所以其較(R) hairpin loop stable 。

DNA constructs
m1-m4 mutants :pGL3-Nef
mutagenesis PCRwith mutagenic primers EWmut1 、 EWmut2 、 E
Wmut3 and general primers EW1 、 EW2 、 EW3
m1-m4 mutants
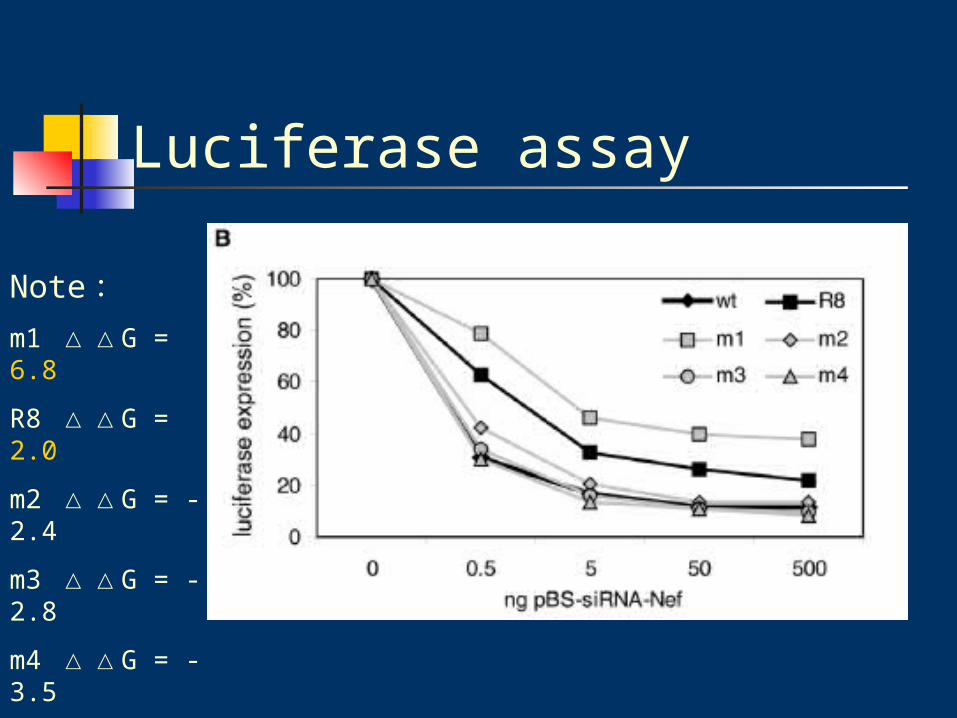
Luciferase assay
Note:m1 △△G = 6.8
R8 △△G = 2.0
m2 △△G = -2.4
m3 △△G = -2.8
m4 △△G = -3.5
wt △△G = -2.8
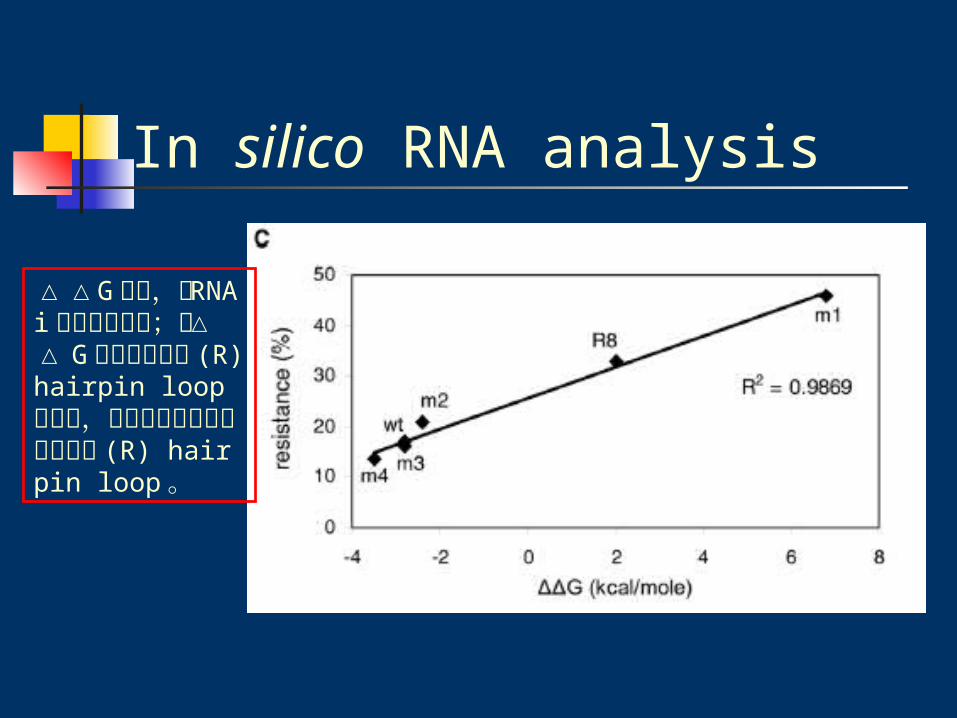
In silico RNA analysis
△△G 越高,對 RNAi 的抵抗力越強;而△△ G 越高也表示其 (R) hairpin loop 越穩定,會使結構趨向形成較穩定的 (R) hairpin loop 。

Materials & Methods
Cells and viruses
DNA constructs
Luciferase assay
In silico RNA analysis
RNA probing(Lead-induced cleava
ge)
EMSA

RNA structure probing (Lead-induced cleavage)
GAA
AC
ACA
CUU
loopstem
所以與預測結構相符合Note : Lead-induced cleavage 主要是切單股結構

RNA structure probing (Lead-induced cleavage)
G
A
G
G
stem
loop
CU AA
loop
所以 R8 也與預測結構相符合
RNA structure probing (Lead-induced cleavage)
經由 Lead-induced cleavage 可以證明由Mfold program 預測出來的結構是正確的。
HIV-1 RNA structure 的改變確實避開了 RNAi 作用的區域。

Materials & Methods
Cells and viruses
DNA constructs
Luciferase assay
In silico RNA analysis
RNA probing(Lead-induced cleava
ge)
EMSA

EMSA (Electrophoretic mobility shift assay)
The siRNA-Nef antisense oligonucleotide was 5’ end labled in the present of 1ul of [γ32P]
ATP

EMSA (Electrophoretic mobility shift assay)
R8 mutant bound siRNA明顯較少, free siRNA 明顯較多
Note:
Duplex formation(%) bound siRNA
=
free + bound siRNA
不超過 39%

Conclusion
mutant Nef gene 確實可以抵抗 siRNA-Nef 作用。
可以藉由 RNA nucleotide substitution or deletion 使 siRNA binding mismatch 。
經由 luceferase assay 、 in silico analysis預測結構、 RNA probing 證實預測結構的正確性、 EMSA ,證明 HIV-1 mutant 型確實藉由局部 (local) 改變 RNA 結構 escape RNA interference 。

Discussion經由 RNAi 這個強大的基因治療工具雖然確實可以抑制 HIV-1 的病毒複製。但是其mutant 型卻可以 escape RNAi 的抑制作用;或許可以設計一個可以 overlap 整個 open reading frames 的 siRNA 來使病毒無從閃躲RNAi 的抑制作用。在藥物設計上也要開始考量 mutant 型的病毒會 escape RNAi-mediated inhibition 。

Thank you

RNA structure probing (Lead-induced cleavage)
wild type and R8 (20pmol) denature 60ul water 85 ℃ 3mins
snap cooling on ice
20ul 4XMO buffer incubated 30 mins 37 ℃
incubated with lead(II) acetate at room temperature
stop cleavage by 3ul 1M EDTASamples (15ul) 0, 5, 15, 25 mins
RNA structure probing (Lead-induced cleavage)
3pmol 32p-labled oligonucleotide was annealed to 3pmol of the lead(II)-treated RNA by incubation at 85 ℃ 3mins
slow cooling 60 ℃ 1h
20ul gel-loading bufferII
samples heated 95℃
10ul samples analized on denatured 6% arylamide

EMSA (Electrophoretic mobility shift assay)
wild type and R8 denature in 30ul water at 85℃ 3 mins
snap cooling
renatured with 10ul 4X MO buffer 37 ℃ 30mins
the transcripts were diluted in 1X MO buffer
final concentration 0 to 7.5 M in MO buffer

EMSA (Electrophoretic mobility shift assay)
取 20ul sample + 2.6nm the 5’-labled oligonucleotide(labled with kinaseMax kit in the present of [γ32P]ATP)
incubated 30mins, room temperature
4ul non-denaturing loading buffer
Analyzed on 4% acrylamide gel
electrophoresis