Embed Size (px)
Citation preview

Histology of the Reproductive Organs of Cryptobranchus alleganiensis (Caudata:Cryptobranchidae) in Missouri
Chris A. Ingersol; Robert F. Wilkinson; Chris L. Peterson; Robert H. Ingersol
The Southwestern Naturalist, Vol. 36, No. 1. (Mar., 1991), pp. 60-66.
Stable URL:http://links.jstor.org/sici?sici=0038-4909%28199103%2936%3A1%3C60%3AHOTROO%3E2.0.CO%3B2-G
The Southwestern Naturalist is currently published by Southwestern Association of Naturalists.
Your use of the JSTOR archive indicates your acceptance of JSTOR's Terms and Conditions of Use, available athttp://www.jstor.org/about/terms.html. JSTOR's Terms and Conditions of Use provides, in part, that unless you have obtainedprior permission, you may not download an entire issue of a journal or multiple copies of articles, and you may use content inthe JSTOR archive only for your personal, non-commercial use.
Please contact the publisher regarding any further use of this work. Publisher contact information may be obtained athttp://www.jstor.org/journals/swan.html.
Each copy of any part of a JSTOR transmission must contain the same copyright notice that appears on the screen or printedpage of such transmission.
The JSTOR Archive is a trusted digital repository providing for long-term preservation and access to leading academicjournals and scholarly literature from around the world. The Archive is supported by libraries, scholarly societies, publishers,and foundations. It is an initiative of JSTOR, a not-for-profit organization with a mission to help the scholarly community takeadvantage of advances in technology. For more information regarding JSTOR, please contact [email protected].
http://www.jstor.orgMon Nov 26 21:04:24 2007

HISTOLOGY OF THE REPRODUCTIVE ORGANS OF CRYPTOBRANCHUS ALLEGANIENSIS
(CAUDATA: CRYPTOBRANCHIDAE) IN MISSOURI
Department of Biology, Mount Vernon Nazarene College, Mount Vernon, OH 43050 Department of Biology, Southwest Mtssouri State University, Springfield, M O 65804
1635 Lee Street, Sprtngfield, M O 65803 Department of Biology, Drury College, Springfield, M O 65802
ABSTRACT-Seasonal changes in the histology of testes and ovaries were studied in Cryptobranchus alleganiensis from Niangua River, Missouri. Spermatogenesis occurred in summer. Vitellogenic activity was greatest from May to mid-September. Breeding occurred in late September. Spermatogonia occupied the testicular lobules from October through June. Interstitial cells were most evident after breeding. Follicular atresia was evident in females that had retained mature ovarian oocytes after the breeding period. Yolk elements of atretic follicles were phagocytized by cells of the follicular epithelium.
The spermatogenetic and oogenetic cycles of few amphibians have been studied in detail (Lofts, 1984). Studies of the interstitial cells of the testis and the rate of follicular atresia in amphibians are limited (Pudney and Callard, 1984; Pan- charatna and Saidapur, 1985). Consequently, we examined the histology of the reproductive organs of the hellbender, Cryptobranchus alleganiensis, an aquatic salamander.
McGregor (1899) and Humphrey (1921) found that spermatozoa were present in the testes of eastern hellbenders only during summer. Baker (1963), however, found that spermatozoa were present from mid-August to early December in the testes of hellbenders caught in Spring River, Arkansas. This population breeds in January (Peterson et al., 1989), whereas all other known populations breed from late August to early Oc- tober (Nickerson and Mays, 1973). Thus, we determined whether hellbenders in Missouri ex- hibit the same spermatogenetic cycle as eastern populations or a cycle more like hellbenders in Spring River, Arkansas.
MATERIALS of hellbend- AND M ~ T ~ o ~ s - S a m p l e s ers were collected by hand during daylight one to three times a month from September 1979 to October 1980 in Niangua River, Laclede Co., Missouri. They were killed within 24 h of capture by immersion in a solution of tricaine, and total length (TL, 11 mm) was re-
corded. Reproductive organs were removed and placed in an alcohol-formalin-acetic acid fixative for 24 h be- fore being stored in 70% ethanol. Ovaries, oviducts, testes, and Wolffian ducts were dehydrated separately in alcohol, cleared in methyl salicilate, and embedded in Paraplast. Ovaries were sectioned at 8 to 15 pm and other tissues at 8 pm. Tissues were stained by the modified Schorr procedure or the Mallory-Heidenhain rapid one-step method (Humason, 1972). Five stages of oocyte development were identified (Wischnitzer, 1966) and recorded for each specimen. Measurements of oocyte diameters were made with an ocular microm- eter. All means are presented k 1 SE.
RESULTS-Mature males ( n = 45) in Niangua River ranged from 344 to 579 mm T L (Table I). One immature male (318 mm TL) was col- lected. Each seminiferous lobule (Lofts, 1984) was cone-shaped, and the apical end was con- nected by a short duct to the longitudinal col- lecting duct. The entire testis was involved in the annual spermatogenetic cycle and in the release of spermatozoa. Development within the testis occurred as a wave that progressed from the cau- dal to the cephalic end.
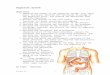
Primary spermatogonia occupied the apical end of each lobule all year. Regenerative changes for a were evident as as 25 September when spermatozoa still occupied some lobules (Fig. 1 A). Spermatogonia had pro- liferated in one third to one half of the length of

March 1991 Ingersol et a1.-Reproductive histology of Cryistobranchus allegan~ensts 61
TABLEI-Number and range of total length (TL) of mature Cryptobranchus alleganiensis for each sample.
Mature females Range
of mean diameterMature males of largest
Date Num- Range of T L Predominant cell Num- Range of TL Oocyte stages oocytes3 collected ber (mm) type in lobules ber' (mm) presentZ (mm)
10 January 3 442-524 Spermatogonia 3 430-497 111, IV 2.3-2.9 22 February 1 470 Spermatogonia 3 448-484 111, IV 2.3-2.6 27 March 1 480 111, IV 2.9 23 April 1 396 Spermatogonia 8 393-569 111, IV, V 2.4-2.8
6 May 4 474-518 Spermatogonia 3 419-519 IV, V 2.8-3.1 5 June 6 344-451 Spermatogonia 2 469-494 IV, V 3.4-3.5 7 July 1 380 Spermatocytes 4 390-458 V 3.8-4.4
14 July 4 477-579 Spmeratids 25 July 2 41 3-41 5 Spermatids 2 460-479 V 5.0-5.2 11 August 2 380-445 Spermatozoa 3 420-458 V 5.2-5.4 25 August 2 438-495 Spermatozoa 3 475-512 V 5.5-5.7
8 September 3 345-529 Spermatozoa 3 417-525 V 5.8-6.3 17 September 9 408-525 V 5.7-6.3 25 September 2 499-532 Spermatogonia 2 493-494 Oviducal ova
9 October 3 455-489 Spermatogonia 2 450-490 111, IV 2.2-2.5 22 October 2 456-489 Spermatogonia 3 438-470 111, IV 1.9-2.4 15 November 6 390-470 Spermatogonia 5 457-540 111, IV 2.3-2.6 10 December 3 392-488 Spermatogonia 4 425-479 111, IV 2.2-2.6
' Does not include 13 females that had retained entire clutch after breeding season. Every mature female had oocyte stages I and 11. Maximum standard error for oocyte diameter was 0.09 mm for any female.
the regressing lobules in three males collected on dominant. They were also present during M a y 9 October (Fig. 1B). and June prior to spermatogenic activity, but they
Spermatozoa were present in the distal portion developed from spermatogonia rather than sper- of isolated lobules that were filled predominantly matocytes at this time. with spermatogonia in five of six males on 15 Interstitial cells were most abundant imme-November. Spermatozoa filled the Wolffian ducts, diately after the spermatozoa had left the lobules. but not the testes, of the smallest (390 mm T L ) During winter they were evident primarily in the of the six males. Spermatozoa were retained in periphery of the testes between distal ends of isolated lobules in two of three males on 10 De- adjacent lobules. Interstitial cells were rare or cember, in one of three on 10 January, and in a absent from July to early September. single male during February. Spermatozoa were Secondary spermatocytes were predominant in never observed in testes or Wolffian ducts ob- the single specimen collected on 7 July. Sper- tained from April through early July. matids also were present in the caudal end of the
Secondary spermatogenic activity sometimes testis. Formation of primary spermatocytes prob- occurred at the end of the normal spermatogenetic ably began during late June because only sper- cycle, resulting in secondary spermatocytes that matogonia were evident in five hellbenders col- subsequently degenerated into spherical struc- lected on 5 June. Approximately equal numbers tures of unknown function (Fig. 1C). These of secondary spermatocytes and spermatids char- spherical structures were recognized as early as acterized sections from four males collected on 14 25 September. Frequently, they were evident in July (Fig. ID) . One male had a few isolated the lumina of the distal ends of lobules during lobules of spermatozoa in the caudal portion of the annual cycle when spermatogonia were pre- the testis. Secondary spermatocytes were predom-

FIG. 1-Seasonal testicular changes of Cryptobranchw alleganiensis. A. September, vacated lobules, lobules with spermatozoa, and development of spermatogonia, scale = 175 pm. B. October, regression after breeding of germinal epithelium in distal half of lobules and repopulation of spermatogonia in proximal half of lobules and repopulation of spermatogonia in proximal half of lobules, scale = 175 pm. C. Spherical structures; interstitial cells evident in winter and early spring, scale = 70 pm. D. July, spermatogenesis; interstitial cells not evident, scale = 35 pm. ET = vacated seminiferous lobules, IC = interstitial cells, SC = spermatocytes, SD = spermatids, SG = spermatogonia, SS = spherical structures, and SZ = spermatozoa.

March 1991 Ingcrsol et a1.-Reproductive histology of Cryptobranchw alleganimsis 63
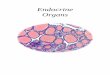
FIG. 2-Spermatozoa of Cryptobranchus alleganiensis. A. August, spermatozoa located peripherally in lobule, scale = 35 pm. B. September, spermatozoa in glomerular capsule of renal corpuscle in epididymis, scale = 70 pm. G = glomerulus, ST = Sertoli cells, and SZ = spermatozoa.
inant in the anterior portion of the testis with terior portion of the opisthonephros (Ratcliff, spermatids in the posterior portion.
Spermiogenesis was evident in two males col- lected on 25 July. Spermatozoa were observed along the periphery of the lobules with their heads oriented toward the lumen, which was filled with Sertoli cells (Fig. 2A). Anterior sections of these testes were populated predominantly with sper- matids but frequently contained secondary sper- matocytes in the periphery of the distal parts of the lobules. The proximal two-thirds of the cau- dal lobules had spermatids, and the distal one- third had spermatozoa.
Lobules of seven males collected between 11 August and 8 September were filled with sper- matozoa, but spermatozoa were not present in the longitudinal canals of the testes or Wolffian ducts. Spermatozoa were not found in the testes of two males collected on 25 September.
Spermatozoa were absent from the epididy- mides and Wolffian ducts during July and August and on 8 September when they were abundant in the testes. The Wolffian duct of one hellbender collected on 25 September was full of sperma- tozoa. A section through the epididymis, or an-
1965), bf this individual revealed a nephric unit with the glomerular capsule containing sperma- tozoa (Fig. 2B). A few spermatozoa were evident in the epididymides of three of five males collected in October, but they were absent in sections of Wolffian ducts near the cloaca.
Mature females (n = 73) ranged from 390 to 569 mm TL. Eight immature females, 230 to 41 5 mm TL, were collected. Stage I oocytes were the principal oocytes of immature specimens and con- stituted the largest proportion of oocytes in adults. These irregularly shaped, previtellogenic oocytes were surrounded by a single follicular epithelial layer and a theca interna. The zona pellucida was absent in this stage. Stage I oocytes ranged from 30 to 860 pm (x = 400 + 30) in diameter.
Deposition of yolk platelets began in the pe- ripheral cytoplasm of stage I1 oocytes, and the zona pellucida became evident. A yolkless band of cytoplasm separated the follicular cells and newly formed zona pellucida from the newly de- posited small yolk platelets. Diameters of stage I1 oocytes ranged from 600 to 1,300 pm (x = 1,030 + 30). Like stage I oocytes, stage I1 oocytes

64 The Southwestern Natural~st vol. 36, no. 1
were observed in all ovaries of mature females each month.
Stage 111 oocytes had only the peripheral cy- toplasm filled with yolk. Diameters of stage I11 oocytes ranged from 1,100 to 2,100 pm (2= 1,450 + 40). They were absent from May through September.
Yolk filled all but the perinuclear zone in stage IV oocytes. Diameters of these oocytes ranged from 1,200 to 2,300 pm (2= 1,760 + 70). They were absent from most ovarian sections during May and June and from all sections July through September. As in stage I11 oocytes, small yolk granules were deposited adjacent to the zona pel- lucida, and larger granules were deposited toward the nucleus.
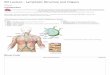
Stage V oocytes were completely filled with yolk. Smaller yolk granules were present adjacent to the zona pellucida and follicular epithelial lay- er in the peripheral cytoplasm (Fig. 3A). The nucleus migrated from its central position to the periphery of the cytoplasm as the oocyte increased in size. The nucleus was central in May, ec- centrical during June, and nearly peripheral by August. Stage V oocytes occurred predominantly from May through September, the same period when secretory products accumulated in oviducts. Ova were spawned in mid-September to early October at a mean diameter of 6,010 & 70 pm.
Immature females exhibited a relationship be- tween developmental stage of oocytes and total length (Table 2). However, the three largest im- mature females (383 mm, 385 mm, 41 5 mm T L ) seemed to be approaching sexual maturity. They were assumed to be immature because they had oocyte stages IV or V that were fewer in number and smaller in diameter than those of mature females collected during the same month. Also the oviducts of these females were smaller than those of mature females. For example, the di- ameter of the oviduct of the largest immature female (415 mm TL) was 1.8 mm compared to 5.3 to 8.4 mm for the mature females collected in May. Peterson et al. (1989) reported size of oviducts and gonads relative to body size of hell- benders in this study.
Thirteen mature females collected after the breeding period retained their entire clutch of eggs: three in January and December; two in May and October; one in February, June, and November. Except for the two in October, the ova of these females were undergoing atresia and presented a chronological sequence in the re-
gression of an atretic oocyte. The earliest stages consisted of the hypertrophy of the follicular ep- ithelium and the disappearance of the zona pel- lucida. The epithelium became highly vacuolated and protruded into the yolk-filled cytoplasm (Fig. 3B). Follicular cells functioned as phagocytes in the removal of yolk elements (Fig. 3C). Blood vessels were prominent in the thecal tissue. Dur- ing May there was abundant coalescence of yolk elements not yet phagocytized by the follicular epithelium. Liquefaction of yolk elements also seemed to occur at this time. The follicular epi- thelium became highly vascularized in May.
A stage IV oocyte in the process of regression was observed in a hellbender captured in May. A large number of neutrophils had invaded the atretic oocyte. However, neutrophils were not as- sociated with mature oocytes that were retained.
D I S C U S S I O N - T ~ ~ ~ ~ ~of the spermatogenetic cycle of hellbenders in Niangua River was similar to that of eastern (collection sites not reported) hellbenders examined by McGregor (1 899) and Humphrey (1921); however, Wikramanayake and Dryden (1985) collected two males in Pennsyl- vania during May that contained spermatids. This is more than a month earlier than when sper- matids were present in males collected in this study. Spermatogenesis in males in Niangua Riv- er was completed in late July, whereas Baker (1963) first observed spermatozoa in mid-August in hellbenders in Spring River, Arkansas. Sper- matozoa remained abundant in males in Arkan- sas until early December. In contrast, few males in Niangua River had abundant spermatozoa af- ter September. Spawning in Niangua River oc- curs in September (Dundee and Dundee, 1965; Peterson, 1988; Peterson et al., 1989). The male collected on 15 November with spermatozoa in the Wolffian ducts may have failed to breed.
Humphrey (1921) believed that interstitial cells of hellbenders were not the site of production of testicular hormone. Instead, he believed the in- terstitial cells developed as a structural response to the regression of the testes after the spermato- genetic cycle was completed and spermatozoa evacuated. However, Pudney et al. (1983) re-ported that the region of differentiated Leydig cells of Necturus maculosus synthesizes steroids. As in hellbenders, hypertrophy of the interstitial cells of N. maculosus occurs after spermiation (Humphrey, 1921).
Although nephric units with glomeruli occur

March 1991 Ingersol ct a1.-Reproductive histology of Cryptobranchw alleganiensis 65
FIG. 3-Atresia of oocyte of Cryptobranchus allegan- iensis. A. Normal stage V oocyte, scale = 17 pm. B. Atretic oocyte showing hypertrophied follicular cells extended into yolk, scale = 70 pm. C. Atretic oocyte showing yolk granules phagocytized by follicular cells, scale = 35 pm. A = shrinkage artifact, FCN = follicular cell nuclei, HFC = hypertrophied follicular cells, YG = yolk granules, and ZP = zona pellucida.
TABLE 2-Relationship between total length (TL) and oocyte development of eight immature Crypto- branchus alleganiensis from Niangua River.
Date collected
5 June 22 February 22 February 23 April
5 June 8 September 7 July 6 May
Stage of largest oocyte
I I I I11 I11 IV v v
mainly in the posterior or urinary kidney of hell- benders, they are also present in the epididymis or genital kidney (Ratcliff, 1965). Noble (1931) suggested that it was a primitive condition in amphibians for vasa efferentia to develop from the embryonic kidney and to serve as a passage- way into the Wolffian duct by way of the nephron. Some anterior opisthonephric nephrons serve as an epididymis and receive vasa efferentia from the testis. The Wolffian duct retains some func- tion as a kidney-draining duct in most salarnan- ders but serves as the vas deferens in all (Williams et al., 1984). Ratcliff (1965) reported that the nephrons of the hellbender urinary kidney fail to connect with the Wolffian duct but extend in- dependently to the cloaca, permitting the Wolf- fian duct to serve exclusively as a vas deferens in these animals.
Deposition of yolk in maturing oocytes of fe- males in Niangua River occurred in a manner similar to that of hellbenders in Pennsylvania (Smith, 1912). The largest oocytes of females killed by Smith (1912) on 6 July contained yolk gran- ules throughout the cytoplasm, corresponding to stage V oocytes found in females collected from May to September in this study.
We were surprised to find an immature female with a total length of 415 mm. Taber et al. (1975) believed that females of that size in Niangua Riv- er were mature, and all other females longer than 390 mm T L were mature. Although this female may have been nonbreeding rather than imma- ture, this study and Peterson et al. (1988) indicate all mature females produce a clutch of ova each year.
Atretic follicules are commonly found in am- phibians, although the physiological significance of follicular atresia is unknown (Saidapur, 1978).

66 The Southwestern Naturalist vol. 36, no. 1
Saidapur (1978) reported that interstitial gland cells seem to arise from large previtellogenic atret- ic follicles and that the gland cells produce ste- roids. However, Lofts (1984) stated that the fol- licular cells secrete amphibian estrogens and that there is little evidence to attribute the atretic fol- licles with a steroidogenic capacity. T h e value of the retention (and subsequent atresia) of a n entire clutch of ova by some females in this study is problematical because the hellbenders did not ap- pear physically impaired. W e have collected two mature females in Spring River, Arkansas, that also had retained a full complement of eggs after the breeding season (Peterson et al., 1989).
We thank the Biology Department, Southwest Mis- souri State University, for funds and use of equipment. We appreciate the issue of scientific collecting permits by the Missouri Department of Conservation.
LITERATURECITED
BAKER,C. L. 1963. Spermatozoa and spermateleosis in Cryptobranchus and Necturus. J . Tennessee Acad. Sci., 38:l-11.
DUNDEE,H. A,, AND D. S. DUNDEE. 1965. Obser-vations on the systematics and ecology of C y p t o -branchus from the Ozark Plateaus of Missouri and Arkansas. Copeia, 1965:369-370.
HUMASON,G. L. 1972. Animal tissue techniques. Third ed. W. H. Freeman and Company, San Francisco, 641 pp.
HUMPHREY,R. R. 1921. The interstitial cells of the urodele testis. Amer. J. Anat., 29:213-279.
LOFTS, B. 1984. Amphibians. Pp. 127-205, in Mar-shall's physiology of reproduction. Reproductive cy- cles of vertebrates (G. E. Lamming, ed.). Fourth ed. Churchill Livingstone, New York, 1:l-842.
MCGREGOR,J. H. 1899. The spermatogenesis of Amphiuma. J. Morphol., 15(Suppl.):57-104.
NICKERSON, NorthM. A., AND C. E. MAYS. 1973. American "giant salamanders." Milwaukee Public Mus. Publ. Biol. Geol., 1:l-106.
NOBLE, G. K. 1931. The biology of the amphibia. McGraw-Hill Book Company, New York, 577 pp.
PANCHARATNA,M., AND S. K. SAIDAPUR. 1985. Ovarian cycles in the frog Rana cyanophlyctis: a
quantitative study of follicular kinetics in relation to body mass, oviduct, and fat body cycles. J. Mor-phol., 186:135-147.
PETERSON,C. L. 1988. Breeding activities of the hellbender in Missouri. Herpetol. Rev., 19:28-29.
PETERSON,C. L., C. A. INGERSOL, R. F. WIL-AND KINSON. 1989. Winter breeding of Cryptobranchus alleganiensis bishopi in Arkansas. Copeia, 1989:1031- 1035.
PETERSON,C. L., D. E. METTER, B. T . MILLER, R. F. WILKINSON, 1988.AND M. S. TOPPING. De-mography of the hellbender Cryptobranchus alle- ganiensis in the Ozarks. Amer. Midland Nat., 119: 291-303.
PUDNEY,J., AND G. V. CALLARD. 1984. Organiza-tion of interstitial tissue in the testis of the sala- mander Necturus maculosus (Caudata: Proteidae). J . Morphol., 181:87-95.
PUDNEY,J., J. A. CANICK, P. M. MAK, AND C. V. CALLARD. 1983. The differentiation of Leydig cells, steroidogenesis and the spermatogenic wave in the testis of Necturus maculosus. General Comp. Endocrinol., 50:43-66.
RATCLIFF,M. A. 1965. The male urogenital system in Cyptobranchus. J. Tennessee Acad. Sci., 40:52- 57.
SAIDAPUR,S. K. 1978. Follicular atresia in the ova- ries of nonmammalian vertebrates. Internat. Rev. Cytol., 54:225-244.
SMITH, B. G. 1912. The embryology of Cryptobran-chus allegheniensis, including comparisons with some other vertebrates. I. Introduction; the history of the egg before cleavage. J. Morphol., 23:61-157.
TABER, C. A,, R. F. WILKINSON,JR., AND M. S. TOPPING. 1975. Age and growth of hellbenders in Niangua River, Missouri. Copeia, 1975:633- 639.
WIKRAMANAYAKE,E. D., AND G. L. DRYDEN. 1985. Body length of male Cryptobranchus alleganiensts at sexual maturity. Bull. Maryland Herpetol. Soc., 21:76-79.
WILLIAMS,A. A., R. A. BRANDON, AND J. MARTAN. 1984. Male genital ducts in the salamanders Eu-rycea lucifuga and Eurycea longicauda. Herpetolo-gica, 40:322-330.
WISCHNITZER,S. 1966. The ultrastructure of the cytoplasm of the developing amphibian egg. Adv. Morphogen., 5:131-179.