Embed Size (px)
Citation preview

HISTOCHEMICAL CHANGES IN MUSCLE NECROSIS AND REGENERATION
BARBARA SMITH Department of Pathology,' St Bartholomew's Hospital, London
IT has been suggested (Engel, 1962) that histochemistry and cyto- chemistry have a great deal to offer in the diagnosis of muscle disease. If this is to be generally true and not just appropriate to a few extremely rare metabolic disorders it is necessary to know the changes that occur in the simple processes of trauma and denervation before one can assess the more complicated problems. This paper is devoted to the changes that occur in simple trauma and regeneration in rats, in lesions produced in suckling mice by the Coxsackie virus (Godman, Bunting and Melnick, 1952~) and in the lesions produced in rabbits by the injection of corticosteroids (Ellis, 1956).
MATERIAL AND METHODS
Muscle trauma was produced in 25 SpragueDawley rats weighing 200-250 g. They were anaesthetised with ether and an incision was made exposing the tibialis anterior. Either a small pointed piece of dry ice was placed in contact with the muscle for about ten seconds, or two parallel longitudinal cuts were made and the muscle between pinched two or three times with an artery forceps. The wounds were sutured; healing was uneventful. The animals were killed at intervals between 16 hr and 3 wk later and the damaged muscle removed for study.
Twelve newborn mice received intracerebral and subcutaneous injections of Coxsackie virus; five of them received Coxsackie B and seven Coxsackie A. The animals were killed between 3 and 7 days later.
Eight rabbits were given cortisone acetate 25 mg. per kg. and four were given friamcinolone (9-fluoro- ldhydroprednisolone) 3 mg. per kg. intramuscularly each day for 16 days. The latter dosage is half that used in a previous study demon- strating the changes in corticosteroid myopathy (Smith, 1964). The animals lost some weight, but all recovered when the drug was stopped. Muscle biopsies were taken at the end of their course of injections and again 2-3 wk later when they had completely recovered.
Some muscle was quenched in a mixture of dry ice and acetone in preparation for cryostat sections and some was fixed in cold calcium-formol overnight and then left in cold gum-sucrose for 3-7 days (Holt, 1959).
The methods used on the cryostat sections were those for DPN and TPN diaphorase, succinate and non-nucleotide-dependent a-glycerophosphate dehydro- g e ~ ~ e s and the TPN-dependent glucose-dphosphate and isocitrate dehydrogenases. Methods similar to those described by Nachlas, Walker and Seligman (1958) and Pearse (1960) were used. Menadione was added to the incubation media where succinate or a-glycerophosphate were substrates (Wattenberg and Long, 19601, and phenazine methosulphate to those where glucose-&phosphate or isocitrate were substrates (Conklin, Dewey and Kahn, 1962). Phosphorylase (Takeuchi and Kuriaki, 1955) and adenosine triphosphatase (ATPase) (Padykula and Herman,
1. PATR. BACT.-VOL. 89 (1965) 139

140 BARBARA SMITH
1955) were studied in cryostat sections. The fixed blocks were used for the acid phosphatase technique (Holt), the specificity of the technique being checked by inhibiting the enzyme with 0 . 1 ~ sodium fluoride. Cryostat sections post-fixed in cold calcium-form01 for 5 min. and then washed, were used for the alkaline phosphatase method (Gomori, 1946).
As doubt has been expressed (Godman, Bunting and Melnick, 19526) about the specificity of the Gomori acid phosphatase technique, preparations were made with the diazonium coupling technique (Burstone, 1958).
RFSULTS The normal tibialis anterior of the rat contains a random mixture
of “ red” and “white” fibres. The “ red” fibres contain a large number of mitochondria; the “ white ” fibres contain very few mito- chondria, but are rich in phosphorylase (Dubowitz and Pearse, 1960-62). When examined under low power the sections show a checkerboard appearance of light and dark fibres whether incubated for phosphorylase or for mitochondrial enzymes.
After trauma the damaged muscle loses almost all glycolytic and oxidative activity for about two days. The muscle fibres then begin to sprout and a number of small fibres with central nuclei appear in the damaged area. These fibres contain large amounts of oxidative enzymes and ATPase, but no phosphorylase at all. In particular they contain large quantities of TPN diaphorase and the two TPN-dependent dehydrogenases, isocitrate and glucose-6-phosphate dehydrogenase (fig. 1). This activity persists until the fibres become histologically normal, a process taking about 17 days. The nuclei become peripheral and the checkerboard appearance both with the phosphorylase and mitochondrial enzymes returns. Any remaining small fibres contain no phosphorylase. The inflammatory cells initially include a number of polymorphonuclear leucocytes, which are positive for alkaline phosphatase (fig. 2), as are the neighbouring capillaries. There appears to be both a vascular proliferation and an increase in activity of the vessels extending some distance from the lesion. Later, as the macro- phages predominate, these cells are seen to have a high concentration of TPN-dependent enzymes (Smith and Rubinstein, 1962). The new collagen associated with the repair process also contains alkaline phosphatase.
Normal muscle in the newborn mouse shows high and uniform mitochondrial activity, but the phosphorylase content is rather low and patchy. In spite of this the latter is demonstrably absent in the severely damaged areas of Coxsackie-infected mice. There is little change in mitochondrial activity except that the more eosinophilic fibres are darker in mitochondrial enzyme preparations (fig. 3). Necrosis is nowhere severe enough to produce loss of mitochondrial activity. The most striking finding in these animals is the intense acid phosphatase activity in the damaged muscle (fig. 4), shown by both Holt and Burstone methods. Serial sections stained with HE do not show obvious
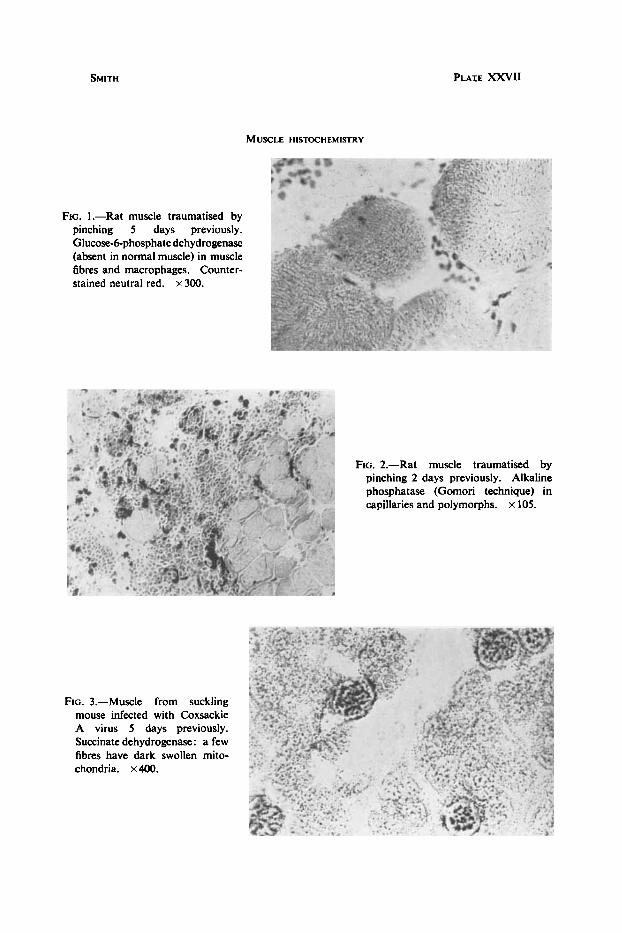
SMITH PLAIE XXVII
MUSCLE HISTOCHEMISTRY
FIG. 1.-Rat muscle traumatised by pinching 5 days previously. Glucose-6-phosphate dehydrogenase (absent in normal muscle) in muscle fibres and macrophages. Counter- stained neutral red. x 300.
FIG. 2.-Rat muscle traumatised by pinching 2 days previously. Alkaline phosphatase (Gomori technique) in capillaries and polymorphs. x 105.
FIG. 3.-Muscle from suckling mouse infected with Coxsackie A virus 5 days previously. Succinate dehydrogenase: a few fibres have dark swollen mito- chondria. x400.

SMITH
MUSCLE HISTOCHEMISTRY
PLATE XXVIlI
FIG. 4.-Muscle from suckling mouse infected with Coxsackie A virus 6 days previously. Coupled diazonium method for acid phosphatase. x 300.
FIG. 5.-Erector spinae muscle from rabbit given triamcinolone for 16 days: biopsy on 17th day. Small basophilic fibres contain more RNA than the rest. Paraffin section, cresyl violet. x 250.
FIG. 6.-Cryostat section of same muscle TPN diaphorase: small as in fig. 5.
fibres very dark. x 300.

MUSCLE HISTOCHEMISTR Y 141
necrosis although there was a mild inflammatory reaction and some fibres were small.
The histochemical changes occurring in corticosteroid myopathy in rabbits, namely, the loss of phosphorylase, the increased mito- chondrial activity in " white " fibres and the presence of lysosomes, have been fully discussed elsewhere (Smith). The animals studied in the present experiment received smaller and non-fatal doses so that the enzyme changes during the recovery process could be studied. After 16 days the animals given the smaller dosage show a myopathy, a striking feature of which is the presence of a number of small basophilic and pyroninophilic fibres (fig. 5). These fibres show intense activity with TPN diaphorase and the two TPN-dependent dehydrogenases (fig. 6). Biopsies done after .recovery show that the phosphorylase activity returns, the lysosomes disappear and the muscle becomes histo- logically and histochemically normal.
DISCUSSION The three types of experimental procedure described were chosen to
illustrate different patterns of necrosis and regeneration. The speed at which the necrosis occurred, the presence or absence of an in- flammatory reaction and the amount of regeneration that took place were responsible for the histological and histochemical pictures.
One of the earliest and most definite signs of muscle damage was loss of phosphorylase activity. Muscle injury seemed to change the energy-producing mechanism from a very specialised anaerobic type to the more usual oxidative phosphorylation. In the traumatised muscle the phosphorylase activity disappeared very quickly. Normal suckling mice have incompletely developed skeletal muscle and its phosphorylase content is low. In the Coxsackie-injected animals it disappeared. In the corticosteroid-treated rabbits the phosphorylase activity was reduced very early in single fibres and it was the first noticeable abnormality. It must be emphasised that these fibres still had normal oxidative enzymes and a normal ATPase activity so that their mito- chondria and myofibrils were intact : the phosphorylase activity was not absent because the fibres were dead and disrupted. Regenerating fibres contained no phosphorylase until they became histologically normal, when it reappeared.
Mitochondria1 enzyme activity, in particular succinate dehydro- genase' and DPN diaphorase, disappeared in rapid necrosis, but was little altered in the early stages of the slower process. In the Coxsackie- injected mice a few fibres stood out from the rest because their mito- chondria were swollen and darker than those of the neighbouring fibres. This was almost certainly due not to an increase of enzyme content but to mitochondrial damage. It is much easier to detect enzyme activity in abnormal than normal mitochondria (Ziegler and Linnane, 1958). There was an increase in all mitochondrial enzymes in

142 BARBARA SMITH
the regenerating fibres. Acid phosphatase is known to be a con- stituent of lysosomes, which are extremely scanty in skeletal muscle (Anderson-Cedergren, 1959). One of the most striking features of necrotic muscle fibres treated by the Gomori-type acid-phosphatase method was the increase in activity of this enzyme, which would appear to indicate an increase in the number of lysosomes. However, if the enzyme was inhibited by adding sodium fluoride to the incubation mix- ture the amount of lead sulphide deposited was diminished, but not abolished. It appeared likely therefore that the necrotic tissue had an affinity for lead and that some of the staining was non-specific, as was suggested by Godman et al. (19523). The investigations were repeated using Burstone’s method for acid phosphatase, which is based on a differ- ent principle but is also not so sensitive. Activity was present, but it was not as extensive or as great as that seen using the Gomori technique. One can conclude therefore that acid phosphatase was present in the necrotic fibres, but that the Gomori technique is not specific and only the difference in activity between the sections incubated with and without added sodium fluoride is significant.
TPN diaphorase and TPN-dependent isocitrate dehydrogenase are present in normal muscle but not prominent, and the TPN-dependent glucose-6-phosphate dehydrogenase cannot be detected histochemically . This last enzyme is a constituent of the hexose monophosphate shunt which appears to play no part in normal skeletal muscle metabolism. All three enzymes were found in high concentration in the sprouts and small regenerating fibres, presumably because these fibres had a high concentration of RNA and the hexose monophosphate shunt is used in the synthesis of this substance. The enzymes occurred not only in the small basophilic fibres but also in the mitochondria of neighbouring fibres. The histochemical picture was more striking in the corti- costeroid-treated animals, as the regenerating fibres were scattered randomly over the section instead of being all together. A similar increase in TPN-dependent enzymes, particularly glucose-6-phosphate dehydrogenase, has been found in mouse dystrophy (McCaman, 1960), another condition associated with marked muscle regeneration.
It has been suggested (Fenichel and Engel, 1963) that the enzyme system on which muscles are dependent for their energy is constant for a given muscle fibre. It is apparent from this study that this is not true. My earlier investigation of severe corticosteroid myopathy in rabbits showed that phosphorylase activity in random fibres is lost and that such fibres are converted into “red” fibres with large numbers of mito- chondria. In the present investigation animals that had a great loss of phosphorylase at the end of their treatment had a normal histochemical pattern 3 wk later.
SUMMARY Muscle necrosis produced experimentally in four different ways is
associated with loss of glycolytic activity and some increase of acid

MUSCLE HISTOCHEMISTR Y 143
phosphatase. Rapid muscle necrosis is associated with loss of mito- chondrial enzymes, though in slow muscle necrosis there may be an apparent increase due to mitochondrial damage.
Muscle regeneration is associated with an increase of mitochondrial activity and particularly of hexose monophosphate shunt enzymes associated with ribonucleic acid synthesis, but phosphorylase does not reappear until the process is complete.
The mechanism of energy production in any given fibre can change from glycolytic to mitochondrial and back again.
I am grateful to Dr R. B. Heath for injecting the Coxsackie virus, and to Mr P. Crocker for the photographs.
This work was assisted by a grant from the Muscular Dystrophy Group of Great Britain.
REFERENCES ANDERSON-CEDERGREN, E. . . . BURSTONE, M. S.. . . . . . CONKLIN, J. L., DEWEY, M. M., AND
DUBOWITZ, V., AND PEARSE, A. G. E. ELLIS, J. T. . . . . . . . ENGEL, W. K. . . . . . . FENICHEL, G. M., AND ENGEL, W. K. GODMAN, G. C., BUNTING, H., AND
GOMORI, G. . . . . . . .
KAHN, R. H.
MELNICK, J. L. 99 ,, ,* ,>
HOLT, S. J. . . . . . . . MCCAMAN, M. W. . . . . . NACHLAS, M. M., WALKER, D. G.,
PADYKULA, HELEN A., AND HERMAN,
PEARSE,A.G.E. . . . . .
AND SELIOMAN, A. M.
EDITH
SMITH, BARBARA . . . . . . SMITH, BARBARA, AND RUBINSTEIN,
TAKEUCHI, T., AND KURIAKI, H. . L. J.
WATTENBERG, L. w., AND LEONG, J. L.
A. W. ZIEQLER, D. M., AND LCNNANE,
1959. 1958. 1962.
1960-62. Histochemie. 2. 105.
J. Ultrastruct. Res., Suppl. 1. J. Nut. Cancer Inst., 21, 523. Amer. J. Anat., 110, 19.
1956. 1962. 1963. 1952~.
19526. 1946.
1959. 1960. 1958.
1955.
1960.
1964. 1962.
1955. 1960.
1958.
Amer. J . Path.; 32, 993. Neurology (Minneap.), 12, 778. Ibid., 13, 1059. Amer. J. Path., 28, 223.
Ibid., 28, 583. Amer. J. Clin. Path., 16, Techn.
Exp. Cell Res., Suppl. 7 , p. 1. Science, 132, 621. J. Biophys. Biochem. Cytol., 4,29.
Bull., vii, p. 177.
J. Histochem., 3, 170.
Histochemistry, theoretical and
Neurology (Minneap.), 14, 857. This Joiirnul, 83, 572.
J. Histochem., 3, 153. Ibid., 8, 296.
applied, 2nd ed., London.
Biochim. biophys. Acta (Amst.), 30,53.





























