Embed Size (px)
Citation preview

Physiological Psychology1974, Vol. 2 (4), 523·528
Hippocampal lesions during rearing and transferof perceptual learning in rats"
BARRY s.ANTONtUniversity ofPuget Sound, Tacoma, Washington 98416
and
THOMAS L. BENNETTColorado State University , Fort Collins, Colorado 8052 1
Bilateral lesions of the hippocampus in humans result in severe deficits of recent memory whichconsist of an inability to recall events a few minutes after they have occurred. On the other hand , suchpatients demonstrate normal abilities to recall events that transpired before the operation. Studiesemploying animals as subjects have produced contrary evidence concerning a memory-consolidation viewof hippocampal functioning because hippocampal animals are not deficient in learning a variety of tasks.In the present research, the recent memory hypothesis was reexamined employing the early experienceparadigm of Gibson and Walk (1956). This methodological variation provided a testing situation moreanalogous to the human testing situation in that animals were required to utilize salient cues from theirenvironment and to transfer this information to a subsequent discrimination task. It was found thatanimals receiving bilateral hippocampectomy before exposure to the to-be-discriminated stimuli fared nobetter in a subsequent discrimination test than animals not exposed to the stimuli. In addition, animalsundergoing hippocampal ablation after exposure to the forms demonstrated significantly betterperformance in the test situation than nonexposed animals. The findings of the present research providesome support for the recent memory hypothesis of hippocampal functioning .
Recent interest in the hippocampus and its possiblefunctions in associative processes was first generated byhuman clinical observations which indicated thatbilateral damage to this structure resulted in severedeficits in recent memory, that is, in the ability to addnew information to long-term memory . These effectsseemed to be specific to the short-term memorymechanism, because general intelligence and the abilityto recall information acquired prior to surgery weregenerally unaffected. Further, the extent of this deficitwas found to vary directly with the amount of bilateralhippocampal involvement produced by the ablation.These observations led Penfield and Milner (1958) tosuggest that the initial encoding of memory traces wasaccomplished by hippocampal mechanisms. For a reviewof these findings, the reader is referred to Milner (1966).
Attempts to find support for and elucidate the basisof this learning deficit using infrahuman subjects havebeen discouraging (see Douglas, 1967 ; Kimble, 1968).While lower animals with hippocampal damage arecertainly behaviorally different from normal animals,particularly with regard to their perseverative behaviorpatterns, the animal-lesion literature has failed tosubstantiate or refute the recent memory hypothesis ofhippocampal functioning. Lower animals can learn avariety of tasks in the absence of an intact hippocampus.
*This article is based on a dissertation submitted by the firstauthor in partial fulfillment of the requirements for the Ph.D.degree at Colorado State University.
tRequests for reprints should be sent to : Barry S. Anton.Department of Psychology, University of Puget Sound, Tacoma,Washington, 98416.
It appears plausible that the lack of supportingevidence for the recent memory hypothesis wheninfrahumans have served as experimental subjects hasbeen the product of methodological differences in thehuman vs lower animal inquiries. The almost exclusiveuse of two-choice discrimination problems in theanimal-lesion studies may have resulted in hippocampalinvestigators failing to tap the relevant dimensionsnecessary to evoke evidence of a recent memory deficit.In the majority of the human studies , the testingsituation involved recall of chains of verbal items orcomplex events from the subject's daily life.
An alternative technique for examining the recentmemory hypothesis in rodents, in a condition which isperhaps more analogous to the testing regimen used withthe human hippocampal patients, is to require theinfrahuman to use information from its pastenvironmental experiences in a transfer paradigm.Specifically , one could examine the effects ofhippocampal lesions on transfer of perceptual learningusing the early experience model of Gibson and Walk(1956). We have used this paradigm in a number ofstudies in recent years to examine the influence of earlyexperience on later discrimination learning (e.g., Bennett& Ellis, 1968; Bennett, Anton, & Levitt, 1971).
The early experience paradigm follows a procedure inwhich experimental animals are exposed to theto-be-discriminated forms (e.g., circles and triangles)during rearing. At maturity , the animals are required tolearn to discriminate between these stimuli as a test ofthe early experience conditions. It has generally been
523

524 ANTON AND BENNETT
found that preexposed animals learn the discriminationproblem significantly faster than do nonexposed controlsubjects. This paradigm requires animals to acquireinformation in their daily living environments and latertransfer this information to a discrimination problem. Itwas hypothesized that the amount transferred to thesubsequent discrimination problem byhippocampectomized animals might provide both asensitive index of the effects ofhippocampectomy and amore analogous testing situation to that employed in thehuman investigations. As a result, the procedure mightprovide a better framework for assessing the validity ofthe recent memory hypothesis for infrahuman subjects.
METHOD
Experimental DesignAlbino rats were reared in a visuaIly sparse environment from
birth until the completion of discrimination training. Stimulusplacement procedures were begun at 50 days of age for theanimals receiving preexposure to the to-be-discriminated forms,and placement procedures were continued for the duration ofthe experiment. Half of the animals received early experiencewith the forms and half did not. Within each of theearly-experience conditions, the subjects received one of fourkinds of surgical treatment : no lesion, cortical control lesion,early hippocampal ablation, or late hippocampal ablation. Thecortical control animals were run because aspiration lesions ofthe hippocampus also destroy the cortex. A detailed descriptionof the resulting groups and the surgical procedures utilized toeffectuate the lesions are presented below. At 100 days of age,the subjects were placed on a 23-h food deprivation schedule inpreparation for the beginning of discrimination pretraining at110 days old. FoIlowing four sessions of pretraining, the animalsof all groups were given a circle-triangle discrimination problemas a test of the effects of the experimental manipulations.
SubjectsThe subjects were male albino rats, with eight animals assigned
to each of the eight groups.
Experimental GroupsThe conditions of early exposure and surgical treatment
resulted in eight experimental groups as foIlows: (a) InGroup NNE (normal nonexposed), the animals were unoperatedand not preexposed to the stimuli. (b) In Group NE (normalexposed), the animals were also unoperated but were preexposedto the to-be-discriminated stimuli. (c) In Group CNE (corticalnonexposed), the animals received bilateral lesions of theneocortex overlying the hippocampus, and experience with theto-be-discriminated forms was not provided. (d) In Group CE(cortical exposed), the animals were identicaIly treated surgicaIlyas Group CNE and were preexposed to the stimuli. (e) InGroup EHNE (early hippocampal nonexposed), the rats receivedbilateral aspiration of the hippocampus at age 45 days and noearly experience with the shapes. (f) In Group EHE (earlyhippocampal exposed), the animals also received bilateralaspiration of the hippocampus at 45 days but foIlowing 5 daysrecovery, experience with the to-be-discriminated stimuli wasintroduced. (g) In Group LHNE (late hippocampal nonexposed),the animals received bilateral destruction of the hippocampus at105 days of age but no early experience with the forms. (h) InGroup LHE (late hippocampal exposed), animals receivedexperience with the to-be-discriminated stimuli from age 50days, and at age 105 days the hippocampus was bilateraIlyremoved through aspiration. During the course of theexperiment, one animal died from the CNE and LHE groups.
RearingThe animals were reared (three to five per cage) in Hoeltge
smaIl-animal cages, measuring 9 x 9 x 15 in., which were placedin compartments measuring 2 ft square. The sides, bottoms, andtops of the compartments were painted flat white . The openingof each compartment was two feet from the white waIl of theroom in which the animals were housed. Food and water wereavailable ad lib. Each of the compartments was iIluminated from7:00 a.m. to 6:00 p.m, daily by a 25-W bulb attached to thecompartment ceiling and shielded by a square piece of whiteflannel. Thus, during the rearing period, the visual field of theanimals consisted of the cage, the compartment walls and ceiling,the water bottle, the cagemates, and the stimuli for thepreexposure groups.
Stimulus placement procedures were introduced at age 50days. For the groups that received early experience with forms,the stimuli consisted of two circles, 3 in. in diarn, and twoequilateral triangles, 3-%in. per side. These shapes wereconstructed from 1/8-in.-thick black Plexiglas. The stimuli weresuspended from the cage ceiling by means of a wire and placedso that the subjects could obtain both visual andtactual-kinesthetic feedback from the forms. There was onecutout to each waIl, and their positions were rotated weeklyaround the cage so that their spatial proximity to food and watercould be controIled.
Discrimination TrainingThe testing apparatus was a Grice discrimination box as
modified by Gibson and Walk (1956). At the age of 110 days,the animals were given 4 days of pretraining identical to themethods outlined by Gibson and Walk (1956). Pretrainingconsisted of training the animals to push open the panels toobtain a food reward in the absence of visual forms attached tothe panels.
After pretraining, discrimination training was begun foIlowingprocedures described by Bennett and Ellis (1968) and Gibsonand Walk (1956) . The subject could obtain a pinch of wet mashfrom one of the two stimulus panels, a black circle or a blacktriangle on a white background, by pushing open the correctstimulus door. Half the subjects were trained to circle positiveand half to triangle positive. The side of presentation of thepositive stimulus was derived from FeIlow's (1967) chancestimulus sequences for discrimination tasks. The criterion fortask mastery was 18 out of 20 correct responses with the last 10consecutive responses correct. The animals were run until thiscriterion was achieved or until they had been run on thediscrimination task for a maximum of 300 trials. The rats wereadministered 12 trials per day.
Surgery and HistologyFollowing anesthetization via sodium pentobarbital
(65 mg/kg body weight) preceded by 0.1 cc (0.2 mg) atropinesulfate to reduce respiratory distress, neocortical andhippocampal ablations were accomplished via aspiration(Isaacson, Douglas, & Moore, 1961). For the hippocampalablations, the neocortical tissue covering thehippocampus was aspirated to expose the hippocampus, whichwas then removed as completely as possible. Care was taken toavoid injury to the underlying thalamus. The cortical-lesiongroups had the hippocampus exposed but not removed. Forthese latter animals, the extent of the cortical ablation wasincreased in an attempt to equate the total amount of braintissue removed for both hippocampal and cortically lesionedsubjects. FoIlowing completion of surgery, each rat received0.1 cc Mikedimide (0.1 mg), a barbiturate antagonist,intraperitoneaIly. Tetracycline was added to the animal's watersupply for several days foIlowing surgery as a prophylacticagainst infection.
FoIlowing completion of behavior testing, the lesionedanimals were kiIled by a lethal dose of sodium pentobarbital and
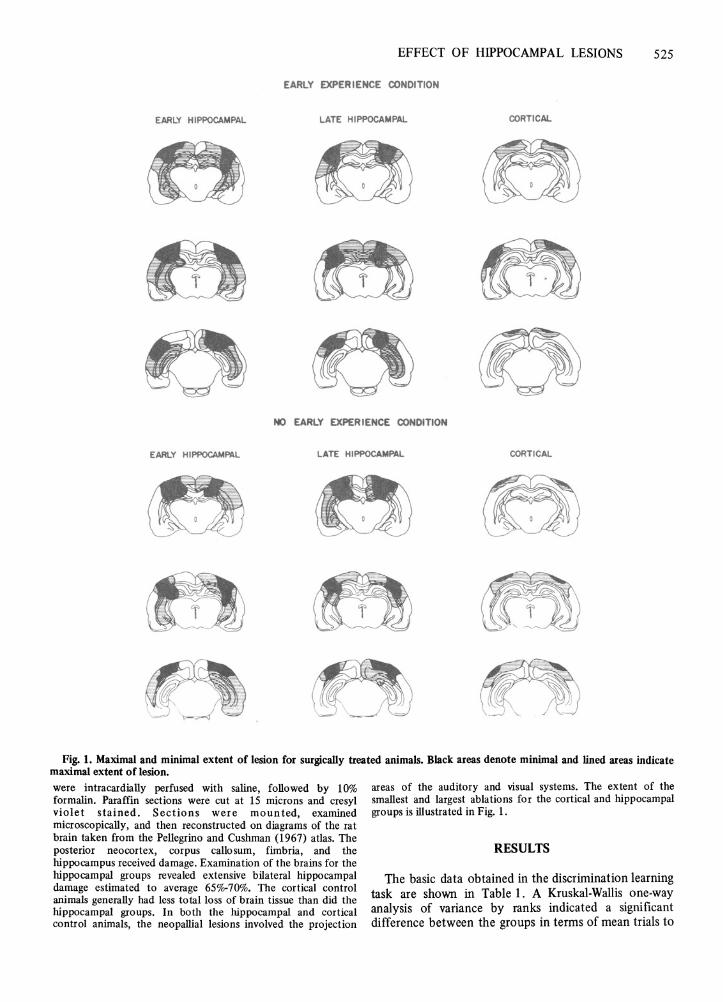
EFFECT OF HIPPOCAMPAL LESIONS 525
EARLY EXPERIENCE CONDITION
EARLY HIPPOCAMPAL
EARLY HIPPOCAMPAL
LATE HIPPOCAMPAL
NO EARLY EXPERIENCE CONDITION
LATE HIPPOCAMPAL
CORTICAL
CORTICAL
RESULTS
areas of the auditory and visual systems. The extent of thesmallest and largest ablations for the cortical and hippocampalgroups is illustrated in Fig. 1.
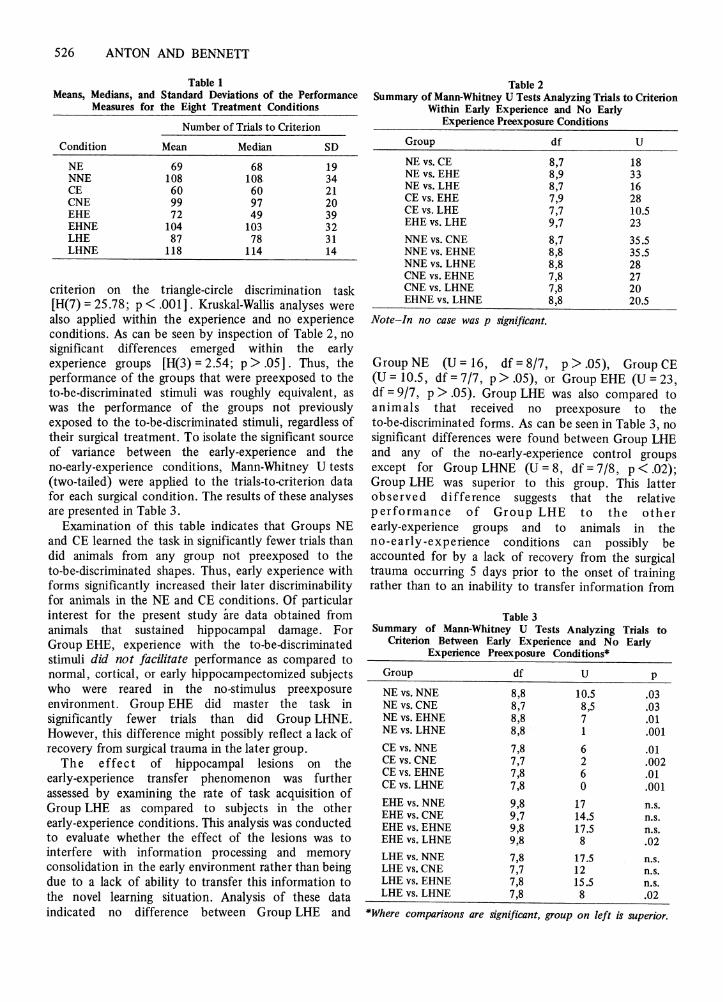
The basic data obtained in the discrimination learningtask are shown in Table 1. A Kruskal-Wallis one-wayanalysis of variance by ranks indicated a significantdifference between the groups in terms of mean trials to
Fig. 1. Maximal and minimal extent of lesion for surgically treated animals. Black areas denote minimal and lined areas indicatemaximal extent of lesion.
were intracardially perfused with saline, followed by 10%formalin. Paraffin sections were cut at 15 microns and cresylviolet stained . Sections were moun ted , examinedmicroscopically, and then reconstructed on diagrams of the ratbrain taken from the Pellegrino and Cushman (1967) atlas. Theposterior neocortex , corpus callosum, fimbria, and thehippocampus received damage. Examination of the brains for thehippocampal groups revealed extensive bilateral hippocampaldamage estimated to average 650/0-70%. The cortical controlanimals generally had less total loss of brain tissue than did thehippocampal groups. In both the hippocampal and corticalcontrol animals, the neopallial lesions involved the projection
526 ANTON AND BENNETT
Table 1Means, Medians, and Standard Deviations of the Performance
Measures for the Eight Treatment Conditions
Number of Trials to Criterion
Table 2Summary of Mann-Whitney U Tests Analyzing Trials to Criterion
Within Early Experience and No EarlyExperience Preexposure Conditions
"Where comparisons are significant, group on left is superior.
Table 3Summary of Mann-Whitney U Tests Analyzing Trials to
Criterion Betwee n Early Experience and No EarlyExperience Preexposure Conditions*
Group df U p
NEvs.NNE 8,8 10.5 .03NE vs, CNE 8,7 8,5 .03NEvs.EHNE 8,8 7 .01NEvs. LHNE 8,8 1 .001
CE vs. NNE 7,8 6 .01CE vs. CNE 7,7 2 .002CEvs. EHNE 7,8 6 .01CE vs. LHNE 7,8 0 .001
EHEvs. NNE 9,8 17 n.s.EHE vs.CNE 9,7 14.5 n.s.EHEvs.EHNE 9,8 17.5 n.s,EHE vs, LHNE 9,8 8 .02
LHE vs. NNE 7,8 17.5 n.s,LHEvs. CNE 7,7 12 n.s.LHEvs. EHNE 7,8 15.5 n.s.LHEvs. LHNE 7,8 8 .02
Group NE (D = 16, df = 8/7, p > .05), Group CE(D = 10.5 , df= 7/7, p > .05) , or Group EHE (D = 23,df = 9/7, p > .05). Group THE was also compared toanimals that received no preexposure to theto-be-discriminated forms . As can be seen in Table 3, nosignificant differences were found between Group LHEand any of the no-early-experience control groupsexcept for Group LHNE (D = 8, df= 7/8 , p < .02) ;Group THE was superior to this group . This latterobserved difference suggests that the relativeperforman ce of Grou p THE to the otherearly -experience groups and to animals in theno-ear ly -experience conditions can possibly beaccounted for by a lack of recovery from the surgicaltrauma occurring 5 days prior to the onset of trainingrather than to an inability to transfer information from
U
1833162810.523
35.535.528272020.5
df
8,78,98,77,97,79,7
8,78,88,87,87,88,8
Group
NE vs. CENE vs. EHENE vs. LHECE vs. EHECE vs. LHEEHE vs. LHE
NNE vs.CNENNEvs.EHNENNEvs. LHNECNE vs. EHNECNEvs. LHNEEHNE vs. LHNE
Note-In no case was p significant.
Condition Mean Median SD
NE 69 68 19NNE 108 108 34CE 60 60 21CNE 99 97 20EHE 72 49 39EHNE 104 103 32LHE 87 78 31LHNE 118 114 14
criterion on the triangle-circle discrimination task[H(7) = 25.78 ; p < .00 1] . Kruskal-Wallis analyses werealso applied within the experience and no experienceconditions. As can be seen by inspection of Table 2, nosignificant differences emerged within the earlyexperience groups [H(3) = 2.54 ; p > .05] . Thus , theperformance of the groups that were preexposed to theto-be-discriminated stimuli was roughly equivalent, aswas the performance of the groups not previouslyexposed to the to-be-discriminated stimuli, regardless oftheir surgical treatment. To isolate the significant sourceof variance between the early-experience and theno-early-experience conditions, Mann-Whitney D tests(two-tailed) were applied to the trials-to-criterion datafor each surgical condition. The results of these analysesare presented in Table 3.
Examination of this table indicates that Groups NEand CE learned the task in significantly fewer trials thandid animals from any group not preexposed to theto-be-discriminated shapes. Thus, early experience withforms significantly increased their later discriminabilityfor animals in the NE and CE condit ions. Of particularinterest for the present study are data obtained fromanimals that sustained hippocampal damage. ForGroup EHE, experience with the to-be-discriminatedstimuli did not facilitate performance as compared tonormal, cortical , or early hippocampectornized subjectswho were reared in the no-stimulus preexposureenvironment. Group EHE did master the task insignificantly fewer trials than did Group LHNE.However, this difference might possibly reflect a lack ofrecovery from surgical trauma in the later group .
The effect of hippocampal lesions on theearly-experience transfer phenomenon was furtherassessed by examining the rate of task acquisition ofGroup THE as compared to subjects in the otherearly-experience conditions. This analysis was conductedto evaluate whether the effect of the lesions was tointerfere with information processing and memoryconsolidation in the early environment rather than beingdue to a lack of ability to transfer this information tothe novel learning situation. Analysis of these dataindicated no difference between Group THE and

the early-experience environment to the discriminationlearning situation.
DISCUSSION
The principal finding of this study was the lack offacilitation produced by early experience with theto-be-discriminated forms on later stimulusdiscrimination learning for animals which had theirhippocampi destroyed prior to preexposure. This findingprovides at least some support for the recent memoryhypothesis of hippocampal functioning , a view positedon the basis of the extreme memory deficits which havebeen noted following hippocampectomy in humans.
On the other hand, the early-experience effect didoccur for rats that had their hippocampi removedfollowing preexposure to the shapes for 55 days whenthe learning rate of these subjects was compared tononpreexposed animals which had been subjected tosimilar surgical treatment. This latter finding isinteresting in that it points out another similaritybetween the present findings and the humanhippocampal syndrome. Although the ability toconsolidate new information is severely disrupted inhuman hippocampal patients, the ability to recall eventswhich occurred prior to surgery is generally intact(Milner, 1966). The data from the late hippocampalanimals do somewhat confound the study in that theirlesion-testing interval was different from thatexperienced by the early hippocampectomized subjects ;nevertheless , the late lesion data do provide someadditional support for the recent memory hypothesis.
While analysis of the findings demonstrated nodifference between Groups EHE and EHNE, thusimplicating the hippocampus in the mediation ofinformation processing and memory consolidation, thegenerality of this finding must be restricted in light ofother data . Intragroup comparisons of animals thatreceived experience with the to-be-discriminated formsindicated that Groups NE, CE, EHE, and LHE wereequivalent. If the recent memory hypothesis is toaccount for memory consolidation of environmentalstimuli, then Groups NE, CE, and LHE should haveperformed significantly better than Group EHE. Thisexpectation was not borne out by the present analysis.Moreover, a recent memory account would predictequivalence between Group EHE and allno-early-experience groups. However, performance ofGroup EHE was significantly better than that of one ofthe no-early-experience groups , Group LHNE. The factthat Group EHE was superior to Group LHNE canpossibly be accounted for by lack of recovery from thesurgical trauma occurring 5 days prior to the onset oftraining for the late hippocampal animals. Nonetheless,the fact that Group EHE performed as well as GroupsNE and CE does restrict the support of the presentfindings for the recent memory hypothesis .
Related research suggests two possible bases for the
EFFECT OF HIPPOCAMPAL LESIONS 527
observed deficits in the ability to profit frompreexposure to the to-be-discriminated stimuli amongthe early hippocampectomized animals. First,hippocampectomy may have resulted in decreasedattention to the cage stimuli. This seems plausible inlight of animal lesion (e.g., Jackson & Strong , 1965;Kimble & Kimble, 1970) and animal EEG (e.g., Bennett,1971 ; Bennett, Hebert, & Moss, 1973) findings whichhave implicated the hippocampus in mediating processesof attention to environmental cues. A decrease inattention to the preexposed shapes would haveaccounted for the lack of facilitation from earlyexperience observed in the hippocampectomizedanimals.
An alternative interpretation centers on the nature ofstimulus feedback required to produce theearly-experience effect. It has been shown that for thealbino rat , the subject used in the present inquiry,successful transfer from early experience is at leastpartially dependent on tactual-kinesthetic feedback fromthe forms. Bennett and Ellis (1968) found that albinorats which were allowed to manipulate the visuallypreexposed shapes learned the subsequent visualdiscrimination task in significantly fewer trials thanthose not allowed to manipulate the stimuli. Theseresults suggested that tactual-kinesthetic feedback mightaid in bringing about an ordering of the visual cues.Rickert and Bennett (1972) and Rickert , Bennett ,Anderson, Corbett, and Smith (1973) have proposedthat hippocampal ablations produce an animal that isdeficient in the ability to consolidate informationderived from response-produced feedback. If one canequate processing proprioceptive feedback duringperformance with the ability to process informationderived from tactual-kinesthetic feedback, then apossible explanation for the observed deficits obtainedin the current research emerges; namely, thehippocampus may be necessary for albino rats to processtactual-kinesthetic feedback from the preexposed visualstimuli. This information is necessary to organizestimulation derived from the visual cues. It should bepointed out that these two interpretations are notmutually exclusive.
This second account could be tested by examining theeffects of early hippocampal ablations on transfer ofperceptual learning for animals that do not needtactual-kinesthetic feedback for maximum transfer ofearly experience. Either hooded rats or hamsters couldbe used for such an inquiry. Both of these species have apigmented iris and superior visual acuity to the albinorat. Neither of these species require tactual-kinestheticfeedback from the early-experience shapes to maximallyprofit from preexposures to visual forms (Anton &Bennett, 1972 ; Bennett , Rickert , & McAllister, 1970).
REFERENCESAnton, B. S., & Bennett, T. L. Role of tactual-kinesthetic
feedback in transfer of perceptual learning in the golden

The effect ofof avoidancePhysiological
528 ANTON AND BENNETT
hamster. Psvchonomie Science, 1972, 28, 157-158.Bennett, T. L. Hippocampal th et a activity and behavior-A
review. Communications in Behavioral Biology, 1971, 6,37-48 .
Bennett, T. L. , Anton, B. S., & Levitt, L. Stimulus relevancy andtransfer of perceptual learning. Psycho nomic Science, 1971.25,159-160.
Bennett, T. L., & Ellis, H. C. Tactual-kinesthetic feedback frommanipulation of visual forms and nondifferentialreinforcement in transfer of perceptual learning. Journal ofExperimental Psychology. 1968,77.495-500.
Bennett, T. L ., H6bert, P. N., & Moss. D. E. Hippocampal thetaactivity and the attention component of discriminationlearning. Behavioral Biology, 1973. 8,173-181.
Bennett , T. L. , Rickert. E. J.. & McAllister, L. E. Role oftactual-kinesthetic feedback in transfer of perc eptual learningfor rats with pigmented irises. Perceptual and Motor Skills,1970.30,916-918.
Douglas. R . J. The hippocampus and behavior. PsychologicalBulletin, 1967,67.87-92.
Fellows, B. J. Chance stimulus sequences for discriminationtasks. Psychological Bulletin, 1967,67,87-92.
Gibson. E. J .. & Walk, R. D. The effect of prolonged exposure tovisually presented patterns on learning to discriminate them.Journal of Comparative and Physiological Psychology, 1956.49. 239-242.
Isaacson, R. L., Douglas. R. J .• & Moore. R. Y.radical hippocampal ablation on acquisitionresponse. Journal of Comparative and
Psychology. 1961, 54, 625-628.Jackson, W. J ., & Strong, P. N., Jr. Differential effect s of
hippocampal lesions upon sequential tasks and maze learningb y the rat. Journal of Comparative and PhysiologicalPsychology. 1969, 68,442-450.
Kimble, D. P. Hippocampus and internal inhibition.Psychological Bulletin, 1968, 70, 285-295.
Kimble. D. P.• & Kimble, R. J. The eff ect of hippocampal lesionson extinction and "hypothesis" behavior in rats. Physiologyand Behavior, 1970. 5. 735-738.
Milner. B. Amnesia following operation on the temporal lobes.In C. W. M. Whitty and O. L. Zangwill (Eds.), Amnesia.London: Butterworths, 1966. Pp , 109-133.
Pellegrino. L. J .• & Cushman, A. J . A ste r eo taxic at las of the rutbrain . New York: Appleton-Century-Crofts, 1967.
Penfield, W. , & Milner, B. Memory deficit produced by bilaterallesions in the hippocampal zo ne. A. M. A. Archives ofNeurology and Psychiatry, 1958. 79. 475-497 .
Rickert. E. J. . & Bennett, T. L. Performance ofhippocampectomized rats on discontinuously negativelycorrelated reward. Behavioral Biology. 1972. 7, 395-582.
Rickert, E. J •• Bennett, T. L .. Anderson, G. J ., Corbett. J •• &Smith, L. Differential performance of hippocarnpally ablatedrats on nondiscriminated and discriminated DRL schedules.Behavioral Biology, 1973, 8. 597-609.
(Received for publication April 11. 1974;revision received August 5, 1974.)





























