Embed Size (px)
Citation preview

High Mannose-binding Lectin with Preference for the Clusterof �1–2-Mannose from the Green Alga Boodlea coacta Is aPotent Entry Inhibitor of HIV-1 and Influenza Viruses*□S
Received for publication, December 27, 2010, and in revised form, March 22, 2011 Published, JBC Papers in Press, April 1, 2011, DOI 10.1074/jbc.M110.216655
Yuichiro Sato‡§1, Makoto Hirayama§1, Kinjiro Morimoto‡, Naoki Yamamoto¶, Satomi Okuyama§, and Kanji Hori§2
From the ‡Faculty of Pharmacy, Yasuda Women’s University, 6-13-1 Yasuhigashi, Asaminami, Hiroshima 731-0153, Japan, the§Graduate School of Biosphere Science, Hiroshima University, Kagamiyama 1-4-4, Higashi-Hiroshima 739-8528, Japan, and the¶Department of Microbiology, Yong Loo Lin School of Medicine, National University of Singapore, 5 Science Drive 2, Block MD4A,Level 5, 117597 Singapore
The complete amino acid sequence of a lectin from the greenalga Boodlea coacta (BCA), which was determined by a combi-nation of Edman degradation of its peptide fragments andcDNA cloning, revealed the following: 1) B. coacta used a non-canonical genetic code (where TAA and TAG codons encodeglutamine rather than a translation termination), and 2) BCAconsisted of three internal tandem-repeated domains, each ofwhich contains the sequence motif similar to the carbohydrate-binding site of Galanthus nivalis agglutinin-related lectins.Carbohydrate binding specificity of BCA was examined by acentrifugal ultrafiltration-HPLC assay using 42 pyridylami-nated oligosaccharides. BCA bound to high mannose-typeN-glycans but not to the complex-type, hybrid-type core struc-ture ofN-glycans or oligosaccharides from glycolipids. This lec-tin had exclusive specificity for �1–2-linked mannose at thenonreducing terminus. The binding activity was enhanced asthe number of terminal �1–2-linked mannose substitutionsincreased.Mannobiose,mannotriose, andmannopentaosewereincapable of binding to BCA. Thus, BCA preferentially recog-nized the nonreducing terminal �1–2-mannose cluster as a pri-mary target. As predicted from carbohydrate-binding propen-sity, this lectin inhibited the HIV-1 entry into the host cells at ahalf-maximal effective concentration of 8.2 nM. A high associa-tion constant (3.71 � 108 M�1) of BCA with the HIV envelopeglycoprotein gp120 was demonstrated by surface plasmon reso-nance analysis. Moreover, BCA showed the potent anti-influ-enza activity by directly binding to viral envelope hemagglutininagainst various strains, including a clinical isolate of pandemicH1N1-2009 virus, revealing its potential as an antiviral reagent.
High mannose-binding lectins are found in various taxon-omy, animals, plants, algae, and bacteria (1). In plants, monocot
mannose-binding lectins form a large family of structurally andevolutionarily related lectins, which are recently referred to asGalanthus nivalis agglutinin (GNA)3-related lectins, most ofwhich bind to high mannose glycans (2). The legume lectinfamily is another large group of plant lectins that includes thevarious highmannose-binding lectins (3). Importantly, some ofthese high mannose-binding plant lectins exhibit a strong anti-HIV activity, but others are weak or completely inactive (4, 5).Such different biological activities virtually depend on thediverse carbohydrate specificities of these plant lectins. Apartfrom the plant lectins, high mannose-binding cyanobacterial(blue-green algal) or eukaryotic algal lectins are a group ofpromising compounds for antiviral agents because of theirunique oligosaccharide binding nature and physicochemicalcharacteristics (6, 7). For example, HIV-inactivating proteinssuch as CV-N fromNostoc ellipsosporum (8, 9), scytovirin fromScytonema varium (10), MVL fromMicrocystis viridis (11), andOAA from Oscillatoria agardhii (12) are high mannose-bind-ing cyanobacterial lectins. These prokaryotic lectins share thecommon structural features, an internal multiplication of theamino acid sequences. Three-dimensional structures of theselectins commonly exhibit characteristic domain swapping (6).However, they are structurally independent from each other intheir amino acid sequences. Although many high mannose-binding lectins have been demonstrated to exhibit anti-HIVactivity (13), cyanobacterial or eukaryotic algal lectins are themost potent compounds so far reported compared with otherplant lectins, because they inhibit HIV replication with half-maximal effective concentration (EC50) values in the low nano-molar to picomolar ranges (6). Red algal lectin griffithsin(GRFT) fromGriffithsia sp. is the most potent inhibitor of HIVmultiplication, which displays the picomolar activity (14).Although red algal lectins share the common features (e.g. lowmolecular weight, cation-independent hemagglutination, andno binding propensity for monosaccharides), GRFT exception-ally binds tomonosaccharides such as glucose ormannose (14).
* This work was supported in part by a grant-in-aid for scientific research (B)from Japan Society of the Promotion of Science.
□S The on-line version of this article (available at http://www.jbc.org) containssupplemental Figs. 1 and 2 and Tables 1 and 2.
The nucleotide sequence(s) reported in this paper has been submitted to theDDBJ/GenBankTM/EBI Data Bank with accession number(s) AB604330,AB604604, and AB604605.
1 Both authors contributed equally to this work.2 To whom correspondence should be addressed: Graduate School of Bio-
sphere Science, Hiroshima University, Kagamiyama 1-4-4, Higashi-Hiro-shima 739-8528, Japan. Tel.: 81-82-424-7931; Fax: 81-82-424-7931; E-mail:[email protected].
3 The abbreviations used are: GNA, G. nivalis agglutinin; BCA, B. coacta agglu-tinin; CV-N, cyanovirin-N; ESI-MS, electron spray ionization-mass spec-trometry; GRFT, Griffithsia sp. lectin; Lys-C, lysylendopeptidase; MDCK cell,Madin-Darby canine kidney cell; MTT, 3-(4,5-dimethylthiazol-2-yl)-3,5-di-phenylformazan; MVL, M. viridis lectin; NR, neutral red; OAA, O. agardhiiagglutinin; PA, pyridylaminated; PE, pyridylethylated; RACE, rapid amplifi-cation of the cDNA end.
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 286, NO. 22, pp. 19446 –19458, June 3, 2011© 2011 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A.
19446 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 286 • NUMBER 22 • JUNE 3, 2011
by guest on July 10, 2018http://w
ww
.jbc.org/D
ownloaded from

Red algal lectin ESA-2 from Eucheuma serra is structurally andevolutionarily related to the cyanobacterial lectin OAA (15).Both lectins exclusively recognize high mannose-type N-gly-cans with extremely high affinity (association constant (KA) ��108 M�1) but do not recognize monosaccharides or small oli-gomannoses (12, 15). They also inhibit the HIV entry into thehost cells with EC50 values of low nanomolar range by directlybinding to envelope gp120 (12).In addition to inhibiting HIV, some high mannose-binding
lectins (e.g. CV-N) show a broad range of antiviral activityagainst influenza virus (16), Ebola virus (17), human herpesvi-rus 6 (18), and hepatitis C virus (19). GRFT has been demon-strated to inhibit cytotoxic effects of the corona virus thatcauses severe acute respiratory syndrome (20). Currently, theemergence of two influenza virus strains, swine-origin influ-enza virus (H1N1-2009) and the highly pathogenic avian influ-enza virus (H5N1), has become a global threat to public health.Therefore, new anti-influenza agents are in great demand toconfront the emergence of highly pathogenic mutants thatacquired the ability to transmit human to human.Previously, a novel lectin (BCA, previously declared as
boonin)was isolated from the green algaBoodlea coacta, and itsbiochemical featureswere partially characterized (21). Interest-ingly, hemagglutination activities of BCA were strongly inhib-ited by glycoproteins with highmannose-typeN-glycan but notby the monosaccharides tested. Here, we efficiently obtainedhigh purity BCA using the yeast mannan-Cellulofine affinitycolumn, and we clarified the full-length sequence of BCA byprotein sequencing and cDNA cloning. Detailed oligosaccha-ride binding specificity of BCA and its antiviral activity againsttwo global viruses, HIV and influenza virus, were evaluated.
EXPERIMENTAL PROCEDURES
Materials—The specimen of B. coacta was collected on thecoast of Kagoshima, Japan. The algal sample was immediatelytransferred to the laboratory, washed, lyophilized, and groundon a ball mill to a powder. The powdered alga, which had beenkept at �20 °C, was used for purification of the lectin BCA. Asmall portion of the alga was stored at �20 °C in RNAlater(Invitrogen) until used for the RNA extraction. Lysyl endopep-tidase (Lys-C) and endoproteinase Asp-N were obtained fromTakara Bio (Kyoto, Japan). Pyridylaminated (PA-) oligosaccha-rides were prepared as described previously (15).The following viruses were kindly provided by Dr. T. Saka-
guchi (Hiroshima University, Japan): A/WSN/33 (H1N1);A/PR8/34 (H1N1); A/FM/1/47 (H1N1); A/Kyoto/1/81 (H1N1);A/Bangkok/10/83 (H1N1); A/Beijing/262/95 (H1N1); A/Aichi/2/68 (H3N2); A/Udorn/72 (H3N2); A/Philippines/2/82(H3N2), and B/Ibaraki/2/85. A clinical isolate of the H1N1-2009 virus, A/Oita/OU1 P3-3/09, was generously provided byDr. A. Nishizono (Oita University, Japan). Recombinant glyco-sylated HIV-1 IIIB gp120 (baculovirus) was purchased fromImmunoDiagnostics (Woburn, MA). Influenza viruses weregrown in the chorioallantoic fluid of 10-day-old chicken eggs.Madin-Darby canine kidney (MDCK) cells were grown in Dul-becco’s modified Eagle’s medium (DMEM) supplemented with10% fetal bovine serum and penicillin/streptomycin.
Purification of B. coacta Lectin—The powdered alga (25 g)was stirred at 4 °C overnight with 10 volumes (w/v) of 20 mM
phosphate buffer (pH 7.0) containing 0.15 M NaCl (PBS). Themixture was centrifuged at 13,000 � g for 30 min, and thesupernatant was recovered. The residues were extracted oncemore with 150 ml of PBS in the same way. To both extractscombined, solid ammonium sulfate was added to attain a finalconcentration of 75% saturation. The mixture was kept over-night at 4 °C and centrifuged at 13,000 � g for 30 min. Theresulting precipitatewas dissolved in a small amount of distilledwater and then dialyzed thoroughly against PBS. After the non-dialysate was centrifuged at 13,000 � g for 30 min, the super-natant was collected as a salting-out fraction.A 3-ml portion (23.2 mg of protein) of the salting-out frac-
tion was applied to a yeast mannan-immobilized column (10 �100mm,Vt� 7.85ml, 1.19mg of ligand/ml of gel) equilibratedwith PBS. The affinity column was prepared as described pre-viously (22). The column was thoroughly washed with 20 mM
phosphate buffer (pH 7.0) containing 1 MNaCl and then elutedwith absolute ethylene glycol. The flow rate of 0.2 ml/min wasmaintained during the chromatography. Fractions of 1ml werecollected and measured for absorbance at 280 nm and forhemagglutination activity. The active factions showing thehemagglutination activity (�25) were pooled, thoroughlydialyzed against distilled water, and further applied to aYMC PROTEIN-RP column (6.0 � 250 mm) (YMC, Kyoto,Japan) equilibrated with 10% acetonitrile in 0.05% trifluoro-acetic acid (TFA). The column was washed with the startingsolvent and then eluted at a flow rate of 1.0 ml/min by alinear gradient (10–70%) of acetonitrile in 0.05% TFA. Theeluate was monitored by absorption at 280 nm and hemag-glutination activity. The active fractions were pooled anddialyzed against distilled water.Analytical Methods—Protein concentration was quantitated
by the Lowry method (23) using bovine serum albumin as thestandard. SDS-PAGE (24) was performed using a 15% (w/v) gel.Staining for carbohydrate was carried out using the G. P. Sen-sor, a carbohydrate-detection kit (J-OILMILLS, Tokyo, Japan),as described previously (25), except that fetuin was used as areference glycoprotein.Hemagglutination Assay—Hemagglutination assay was per-
formed using a 2% (v/v) suspension of trypsin-treated rabbiterythrocytes, as described previously (21). Briefly, rabbit bloodpreparation was washed three times with 50 volumes of saline,and the packed cells were suspended in saline to give a 2% (v/v)suspension of native erythrocytes. One-tenth volume of 0.5%trypsin in saline was added to a 2% native erythrocyte suspen-sion, and the mixture was incubated at 37 °C for 60 min. Afterwashing three times with saline, a 2% trypsin-treated erythro-cyte suspension was prepared in saline. Hemagglutinationactivity was expressed as a titer, the reciprocal of the highest2-fold dilution exhibiting positive hemagglutination, or as aminimum hemagglutination concentration, the protein con-centration of the highest lectin dilution exhibiting positivehemagglutination.Enzymic Digestion and Separation of Peptides—BCA was
subjected to S-pyridylethylation according to the methoddescribed previously (15). Two hundred �g of S-pyridyleth-
Antiviral Activity of High Mannose-binding Green Algal Lectin
JUNE 3, 2011 • VOLUME 286 • NUMBER 22 JOURNAL OF BIOLOGICAL CHEMISTRY 19447
by guest on July 10, 2018http://w
ww
.jbc.org/D
ownloaded from

ylated (PE-) BCAwas digestedwith Lys-C (enzyme/substrate�1:100 (w/w)) in 50 mM Tris-HCl (pH 8.5) at 37 °C for 24 h.Asp-N digestion (enzyme/substrate � 1:100 (w/w)) was per-formed using the same amount of PE-BCA in 50 mM Tris-HCl(pH7.5) at 37 °C for 18 h. For the isolation of peptide fragments,each digest was separated by reverse-phase HPLC on a TSKgelODS-120T column (4.6� 250mm) (Tosoh, Tokyo, Japan) witha linear gradient (5–65%) of acetonitrile in 0.1% TFA.Amino Acid Sequence Analysis—The N-terminal amino acid
sequences of intact protein and peptides generated by enzymicdigestion were determined by an automated protein sequencer(Applied Biosystems 477A) connected to the 3-phenyl-2-thio-hydantoin-amino acid analyzer (120A) (Applied Biosystems).Molecular Weight Determination of Protein and Peptides—
The molecular weights of native BCA, PE-BCA, and peptidefragments were determined by Finnigan LCQ electron sprayionization (ESI)-mass spectrometry (MS) (Finingan, CA).cDNA Cloning of BCA—Total RNA was extracted from the
RNAlater-treated alga using the plant RNA isolation reagent(Invitrogen). mRNA purification from the total RNA was per-formed using a NucleoTrapmRNA purification kit (Macherey-Nagel, Duren, Germany). Full-length cDNAs were synthesizedfrom 200 ng of mRNA using a GeneRacer kit (Invitrogen)according to the manufacturer’s instruction. The first PCRfor rapid amplification of the cDNA 5� end (5�-RACE) was ini-tiated by adding to each 0.2 �l of 10-fold diluted synthesizedcDNA to 8 tubes of a 9.8-�l solution containing 6 pmol ofGeneRacer_5�_Primer, 50 pmol of a degenerated primerBCA_5�_RACE_R1 designed from the partial BCA sequence(see supplemental Table 1 for the primer sequences), 1 �l of10� Blend Taq buffer (Toyobo, Osaka, Japan), 2 nmol each ofdNTP, and 0.25 units of Blend Taq DNA polymerase (Toyobo).The reaction with T Gradient Thermocycler (Biometra, Got-tingen, Germany) consisted of denaturation at 94 °C for 3 min,followed by 35 cycles consisting of denaturation at 94 °C for30 s, annealing at gradient temperature of 50–64 °C (2 °Cincrements) for 30 s, and extension at 72 °C for 1 min, and thefinal extension step at 72 °C for 5 min. The PCR products in 8tubes were pooled and then diluted to 100-fold. The nestedPCR was performed by the same method, except that 0.2 �lof the dilution was used as a template and 2 pmol ofGeneRacer_5�_Nested_Primer and 50 pmol of a degeneratedprimer BCA_5�_RACE_R2 as a primer pair (supplementalTable 1). Nested PCR products were subcloned into pGEM-TEasy vector (Promega). Cycle sequencing reaction was per-formed using a BigDyeTerminator cycle sequencing kit version3.1 and ABI 3130xl DNA sequencer (Applied Biosystems).3�-RACE was performed in the same way as 5�-RACE asdescribed above, except the use of 2 pmol each ofGeneRacer_3�_Primer and BCA_F1 (supplemental Table 1)designed from the sequence obtained by the 5�-RACE. At last,following the manufacturer’s instruction, the full-length BCAcDNA was amplified using a high fidelity DNA polymeraseKOD FX Neo (Toyobo) and a primer pair of BCA_5�_End_Fand BCA_3�_End_R (supplemental Table 1), which weredesigned from the 5�- and 3�-terminal sequences of BCA cDNAobtained by 5�- and 3�-RACE and subcloned into pCR-BluntII-TOPO vector (Invitrogen).
Sequence Data Processing—Homologous sequences wereidentified with the basic local alignment search tool program.The internal tandem repeat regions were compared with eachother using ClustalW2 (26). Signal peptide region was pre-dicted with SignalP 3.0 (27).Centrifugal Ultrafiltration-HPLC Method—The oligosac-
charide binding activity of BCAwas determined using a centrif-ugal ultrafiltration-HPLCmethod as described previously (15).Briefly, 90�l of 500 nM BCA in 50mMTris-HCl (pH 7.0) and 10�l of 300 nM PA-oligosaccharide were mixed and kept at roomtemperature for 1 h. Subsequently, unreacted PA-oligosaccha-rides were recovered by centrifugation (10,000 � g, 30 s) withNanospin Plus (Gelman Science, MI). An aliquot of the filtratewas applied to TSKgel ODS-80TM column (4.6 � 150 mm,Tosoh) and eluted with 10%methanol in 0.1 M ammonium ace-tate buffer at a flow rate of 1.0ml/min in a column oven (40 °C).The eluate was monitored at an excitation wavelength of 320nm and an emission wavelength of 400 nm, and then unboundPA-oligosaccharide (Ounbound) was quantified. The amount ofbound PA-oligosaccharide (Obound) was obtained by followingformula: Obound �Oadded �Ounbound, where Oadded representsthe amount of added PA-oligosaccharide determined from thefiltrate of reaction solution without a lectin. The binding activ-ity (Obound/Oadded) was calculated as a ratio of the amount ofbound PA-oligosaccharide to that added.Anti-HIV Activity of BCA—In vitro evaluation of anti-HIV
activity of BCA was performed by a colorimetric assay asdescribed previously (28). Briefly, 3-(4,5-dimethylthiazol-2-yl)-3,5-diphenylformazan (MTT, Sigma) was used to detect theviability of both HIV-1 (HTLV-IIIB strain)- and mock-infectedMT-4 cells in the presence of a test compound at variousconcentrations.Interaction of BCA with gp120—Direct interaction of BCA
with the HIV envelope gp120 was analyzed by surface plas-mon resonance analysis using a BIAcore 2000 system (GEHealthcare) as described previously (12), except that therecombinant glycosylated HIV-1 IIIB gp120 (baculovirus,ImmunoDiagnostics) was immobilized to give 300 reso-nance units on a carboxymethylated dextran-coated sensorchip (CM5, GE Healthcare). The data were fit globally to asimple Langmuir 1:1 binding model with local Rmax (maxi-mum response) using BIAevaluation 3.1 software.Anti-influenza Activity of BCA—Evaluation of anti-influenza
activity was performed by the neutral red (NR) dye uptakeassay. Various concentrations of the lectin were prepared withDMEMcontaining 10�g/ml trypsin in a 96-wellmicroplate. Toeach well, virus was added as a multiplicity of infection of�0.001 infectious particles per cell. After incubating at 37 °Cfor 48 h, 100�l of NR dye (150�g/ml inDMEM)was added andfurther incubated for 2 h. NR dye incorporated into the cellswas extracted by the addition of 100 �l of 1% acetic acid, 50%ethanol. The color intensity of the dye absorbed by and subse-quently eluted from the cells was measured at 540 nm with amicroplate reader (1420 multilabel counter, PerkinElmer LifeSciences) as a factor of surviving from the virus infection.Immunofluorescence Microscopy—Immunofluorescence stain-
ing was performed to visualize and evaluate BCA inhibition ofinfluenza virus infection as described previously (29). Briefly,
Antiviral Activity of High Mannose-binding Green Algal Lectin
19448 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 286 • NUMBER 22 • JUNE 3, 2011
by guest on July 10, 2018http://w
ww
.jbc.org/D
ownloaded from
MDCK cells grown on cover glass were infected withA/Udorn/72 at a multiplicity of infection of �0.001 infectiousparticles per cell, in the presence or absence 1 �M BCA inDMEM containing 10 �g/ml trypsin. After 24 h post-infection,the infected cells were fixed and visualized by incubating withmouse monoclonal anti-hemagglutinin antibody (HyTest,Turku, Finland) at 37 °C for 1 h followed by fluorescein isothio-cyanate (FITC)-conjugated goat anti-mouse IgG antibody(Anticorps Secondaires, Compiegne, France) at 37 °C for 1 h.The cells were mounted using Vectashield with 4�,6-di-amidino-2-phenylindole (DAPI) (Vector Laboratories, Burlin-game, CA) andwere observed under a fluorescencemicroscope(OLYMPUS BX51, Olympus, Japan).Enzyme-linked Immunosorbent Assay (ELISA)—Direct inter-
action between BCA and viral hemagglutinin was analyzedusing an ELISA as described previously (29). BCA (5 �g/ml) incarbonate buffer (pH 9.6) was immobilized on ELISA plates(BD Biosciences). Following washing with phosphate-bufferedsaline (pH 7.4), containing 0.1% Tween 20 (PBST), the wellswere blocked with 3% skimmilk for 1 h at 37 °C. After a furtherPBST washing, the wells were incubated with various concen-trations of an influenza vaccine preparation (Astellas, Tokyo,Japan) enriched for virus hemagglutinin for 1 h at 37 °C. Afterwashing with PBST, the wells were incubated with mouse anti-hemagglutinin monoclonal antibody (HyTest) for 1 h at 37 °Cfollowed by incubation with horseradish peroxidase (HRP)-conjugated goat anti-mouse IgG antibody (GE Healthcare) for1 h at 37 °C. After washing with PBST, 3,3�,5,5�-tetramethyl-benzidine substrate (Sigma) was added. The reaction wasstopped using 3,3�,5,5�-tetramethylbenzidine stop reagent(Sigma), and absorbance at 450 nm was measured using amicroplate reader (1420 multilabel counter, PerkinElmer LifeSciences). The sameELISAwas performed to test the inhibitoryeffect of yeast mannan on the interaction between BCA andhemagglutinin, except that yeast mannan was added to theplate coated with BCA prior to incubation with hemagglutinin(3 �g/ml).
RESULTS
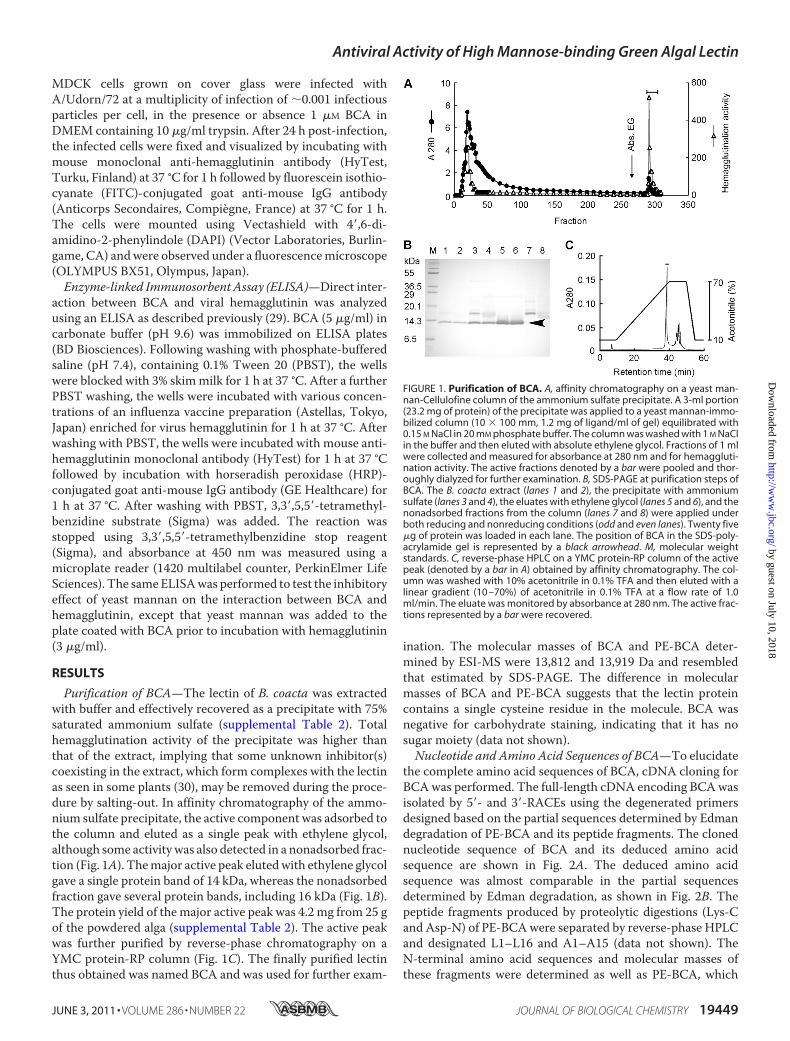
Purification of BCA—The lectin of B. coacta was extractedwith buffer and effectively recovered as a precipitate with 75%saturated ammonium sulfate (supplemental Table 2). Totalhemagglutination activity of the precipitate was higher thanthat of the extract, implying that some unknown inhibitor(s)coexisting in the extract, which form complexes with the lectinas seen in some plants (30), may be removed during the proce-dure by salting-out. In affinity chromatography of the ammo-nium sulfate precipitate, the active component was adsorbed tothe column and eluted as a single peak with ethylene glycol,although some activity was also detected in a nonadsorbed frac-tion (Fig. 1A). Themajor active peak elutedwith ethylene glycolgave a single protein band of 14 kDa, whereas the nonadsorbedfraction gave several protein bands, including 16 kDa (Fig. 1B).The protein yield of themajor active peak was 4.2 mg from 25 gof the powdered alga (supplemental Table 2). The active peakwas further purified by reverse-phase chromatography on aYMC protein-RP column (Fig. 1C). The finally purified lectinthus obtained was named BCA and was used for further exam-
ination. The molecular masses of BCA and PE-BCA deter-mined by ESI-MS were 13,812 and 13,919 Da and resembledthat estimated by SDS-PAGE. The difference in molecularmasses of BCA and PE-BCA suggests that the lectin proteincontains a single cysteine residue in the molecule. BCA wasnegative for carbohydrate staining, indicating that it has nosugar moiety (data not shown).Nucleotide and Amino Acid Sequences of BCA—To elucidate
the complete amino acid sequences of BCA, cDNA cloning forBCAwas performed. The full-length cDNA encoding BCAwasisolated by 5�- and 3�-RACEs using the degenerated primersdesigned based on the partial sequences determined by Edmandegradation of PE-BCA and its peptide fragments. The clonednucleotide sequence of BCA and its deduced amino acidsequence are shown in Fig. 2A. The deduced amino acidsequence was almost comparable in the partial sequencesdetermined by Edman degradation, as shown in Fig. 2B. Thepeptide fragments produced by proteolytic digestions (Lys-Cand Asp-N) of PE-BCAwere separated by reverse-phase HPLCand designated L1–L16 and A1–A15 (data not shown). TheN-terminal amino acid sequences and molecular masses ofthese fragments were determined as well as PE-BCA, which
FIGURE 1. Purification of BCA. A, affinity chromatography on a yeast man-nan-Cellulofine column of the ammonium sulfate precipitate. A 3-ml portion(23.2 mg of protein) of the precipitate was applied to a yeast mannan-immo-bilized column (10 � 100 mm, 1.2 mg of ligand/ml of gel) equilibrated with0.15 M NaCl in 20 mM phosphate buffer. The column was washed with 1 M NaClin the buffer and then eluted with absolute ethylene glycol. Fractions of 1 mlwere collected and measured for absorbance at 280 nm and for hemaggluti-nation activity. The active fractions denoted by a bar were pooled and thor-oughly dialyzed for further examination. B, SDS-PAGE at purification steps ofBCA. The B. coacta extract (lanes 1 and 2), the precipitate with ammoniumsulfate (lanes 3 and 4), the eluates with ethylene glycol (lanes 5 and 6), and thenonadsorbed fractions from the column (lanes 7 and 8) were applied underboth reducing and nonreducing conditions (odd and even lanes). Twenty five�g of protein was loaded in each lane. The position of BCA in the SDS-poly-acrylamide gel is represented by a black arrowhead. M, molecular weightstandards. C, reverse-phase HPLC on a YMC protein-RP column of the activepeak (denoted by a bar in A) obtained by affinity chromatography. The col-umn was washed with 10% acetonitrile in 0.1% TFA and then eluted with alinear gradient (10 –70%) of acetonitrile in 0.1% TFA at a flow rate of 1.0ml/min. The eluate was monitored by absorbance at 280 nm. The active frac-tions represented by a bar were recovered.
Antiviral Activity of High Mannose-binding Green Algal Lectin
JUNE 3, 2011 • VOLUME 286 • NUMBER 22 JOURNAL OF BIOLOGICAL CHEMISTRY 19449
by guest on July 10, 2018http://w
ww
.jbc.org/D
ownloaded from

resulted in elucidation of the partial sequences that correspondto 118 amino acid residues of the lectin molecule (Fig. 2B). Inthe sequencing, some peptides (A1, A4, A5, L4, and L12) weregenerated by nonspecific cleavage. The 30 N-terminal aminoacid sequence of PE-BCA was determined as GAF(Q/K)AI-
SGESGKYLSHAFAKIWLQNGYQGL, at the fourth residue ofwhich two amino acids, glutamine and lysine, were identified,suggesting the presence of the isoforms. Comparison of cDNAand peptide sequences of BCA elucidated that the general stopcodons of TAA and TAG, but not TGA, encoded glutamine
FIGURE 2. Nucleotide and amino acid sequences of BCA. A, nucleotide and deduced amino acid sequences of the cDNA encoding BCA. The stop codon TGAis shown as an asterisk. Glutamine residues coded by TAA and TAG codons are represented in boldface. The italicized and nonitalicized numbers represent thepositions of nucleotides and amino acids. The nucleotide sequence data appear in the DDBJ, EMBL, and GenBankTM databases under accession numberAB604330. B, amino acid sequence of BCA. The partial amino acid sequences obtained by Edman degradation of PE-BCA and its peptide fragments (repre-sented with arrows) are compared with the complete amino acid sequence deduced from the BCA cDNA. The peptide fragments produced by cleavage ofPE-BCA with Lys-C or Asp-N were separated by reverse-phase HPLC. Amino acids identified by sequential Edman degradation are indicated by solid lines, andunidentified amino acids are indicated by dashed lines. The numbers above the sequence represent the position of amino acids. Numeric values below the linesindicate the molecular masses (Da) of peptides determined by ESI-MS, and the values in parentheses indicate the calculated ones from the sequences. Twopeptides were included in the fraction of L8 (L8-1 and -2). L, Lys-C peptides; A, Asp-N peptides. The putative signal peptide and C-terminal propeptide areshown as the underlined sequence and the double underlined sequence, respectively. The signal peptide region was defined with SignalP 3.0.
Antiviral Activity of High Mannose-binding Green Algal Lectin
19450 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 286 • NUMBER 22 • JUNE 3, 2011
by guest on July 10, 2018http://w
ww
.jbc.org/D
ownloaded from

residues (Gln-56 and Gln-132) (Fig. 2A), as reported for someproteins in several other species of green algae (31–33). Thus,BCA cDNA was composed of 1,023 bp containing 117 bp of5�-untranslated region (UTR), 363 bp of 3�UTR, and 540 bp ofopen reading frame (ORF), which encoded 180 amino acids,including the signal peptide of 19 residues (1–19 amino acids), 3internal tandem-repeated domains (20–144 amino acids), andthe C-terminal region of 36 residues (145–180 amino acids)(Fig. 2B). The three tandem-repeated domains, each of whichconsisted of 40–43 residues, showed sequence identity of47.6–62.8% to each other (Fig. 3A). No homologous gene can-didates that showed high sequence similarities with BCA werefound by in silico search. BCA showed no significant sequencesimilarity with other highmannose-type glycan-binding lectinssuch as CV-N, OAA, and ESA-2. Strikingly, however, BCA par-tially possessed the carbohydrate-binding motif of GNA-re-lated lectins, despite having no overall sequence similarity. Thesequence alignment of both BCA and GNA shows obvious dis-ruption of GNA-subdomain III in BCA, whereas the sequencesof BCA at GNA-subdomains I and II were partially conserved(Fig. 3B).Carbohydrate Binding Specificity of BCA—The carbohydrate
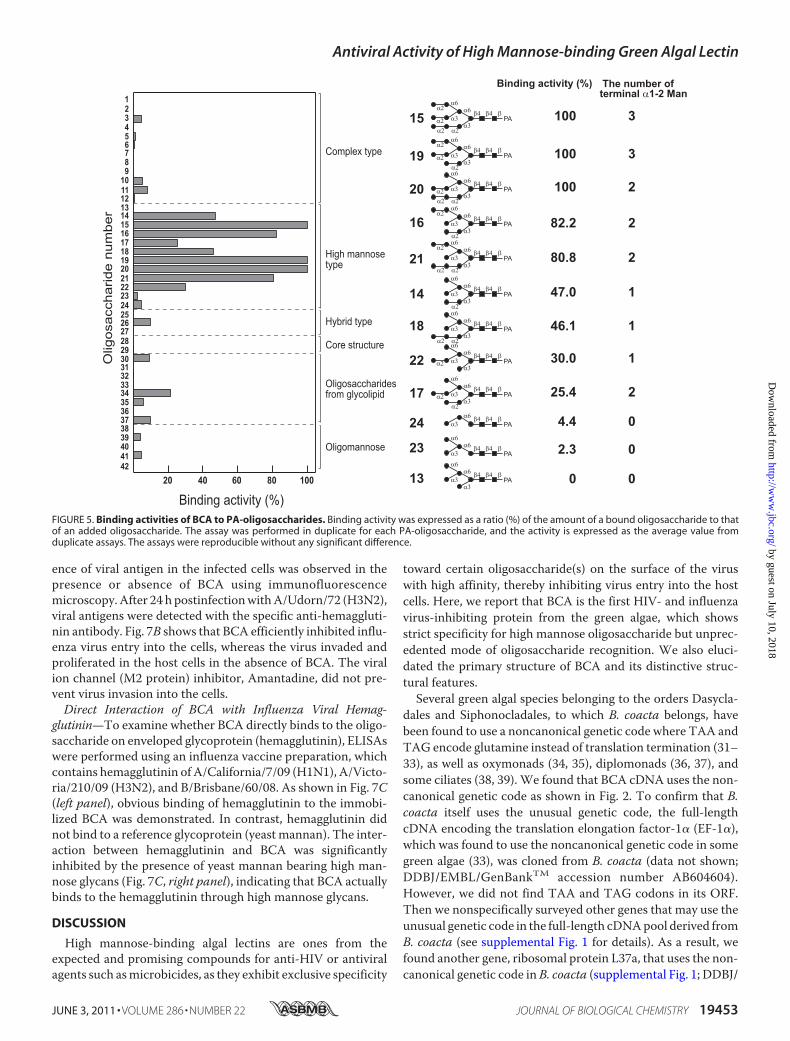
binding specificity of BCA was investigated by a centrifugalultrafiltration-HPLCmethod. Fig. 4 shows the structure of PA-oligosaccharides used in this study. Of the 42 kinds of PA-oli-gosaccharides tested, BCA selectively recognized 9 carbohy-drate structures (oligosaccharides 14–22) that are categorizedinto high mannose-typeN-glycans (oligosaccharides 13–24) asshown in Fig. 5. The complex typeN-glycans (oligosaccharides1–12), hybrid-type N-glycans (oligosaccharides 25–27), thecore structures of N-glycans (oligosaccharides 28 and 29), andoligosaccharides from glycolipid (oligosaccharides 30–37)were incapable of binding to BCA. The binding ability of BCAto high mannose-type N-glycans differed depending on thestructure of branched carbohydrate moiety. BCA showed thepreference for the oligosaccharides bearing terminal �1–2-
linkedmannose(s), and the activity was increased in proportionto the increased number of �1–2 substitutions at nonreducingend. This observation was evident from the fact that BCAbound completely to the oligosaccharides 15 and 19 bearingthree fully exposed �1–2-linked mannoses (Fig. 5). As for theoligosaccharides having two terminal �1–2-mannoses, twoclosely proximate �1–2-mannoses were likely to be preferredfor BCA compared with two sterically distant �1–2-mannoses,because BCA completely bound to oligosaccharide 20, and theactivities for oligosaccharides 16 and 21 were somewhatdecreased (about 80%). Activity toward oligosaccharide 17 wasmuch lower (25.4%) despite having two terminal �1–2-linkedmannoses. With the oligosaccharides having only one terminal�1–2-mannose (oligosaccharides 14, 18, and 22), the bindingactivity of BCA was further decreased showing the activityaround 30–47%. In contrast, this lectin did not interact withhigh mannose-type N-glycans that are devoid of terminal�1–2-mannose as seen in oligosaccharides 13, 23, and 24.These results indicate that primary targets of BCA are the ter-minal �1–2-linked mannose(s). Moreover, no interaction wasobserved inmannobioses 38–40, mannotriose 41, andmanno-pentaose 42 that are the main constituents of branched moietyof high mannose-type N-glycans. Interestingly, even �1–2-linked mannobiose 38 lacks the binding ability to BCA. Theresults suggest that BCA exclusively recognizes nonreducingterminal �1–2-linked mannose and the clustering of those res-idues might contribute to enhance BCA affinity.Anti-HIV Activity of BCA—In vitro evaluation of potent anti-
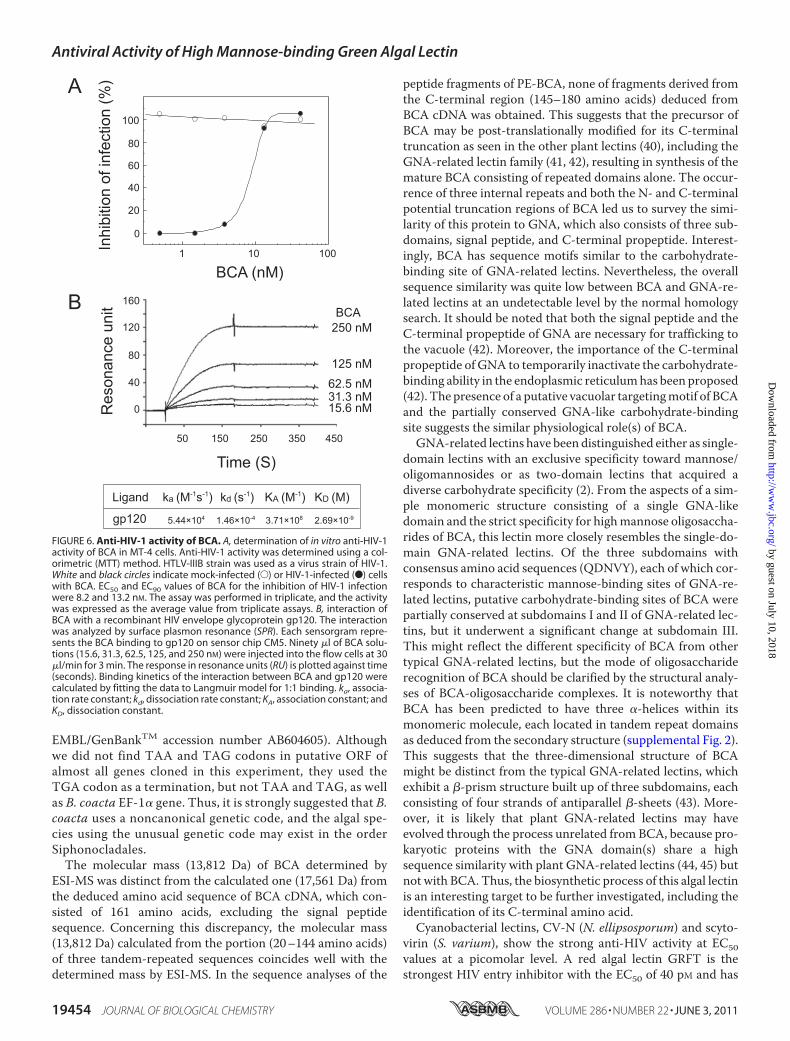
HIV activity of BCAwas determined by the conventional MTTassay using MT-4 cells. BCA inhibited the HIV-1 infectiondose-dependently, with an EC50 of 8.2 nM (Fig. 6A). Cell viabil-ity was not affected up to 100 nM, the highest dose in thisexperiment.Direct Interaction of BCA with gp120—To evaluate the
molecular basis of anti-HIV activity of BCA, we tested thedirect interaction of BCA with a recombinant HIV envelope
FIGURE 3. Structural features of BCA. A, primary structure of BCA. The sequence alignment of repeated domain was generated with ClustalW2. Identicalresidues are shaded. B, comparison of amino acid sequences between BCA and GNA. The alignment was carried out using ClustalW2 and then manuallyadjusted. The structural motif of three mannose-binding sites of GNA and corresponding sequences in BCA are represented in boldface. The subdomainnumbers of GNA are indicated above each binding site. Signal peptide and C-terminal propeptide regions are represented in lowercase letters.
Antiviral Activity of High Mannose-binding Green Algal Lectin
JUNE 3, 2011 • VOLUME 286 • NUMBER 22 JOURNAL OF BIOLOGICAL CHEMISTRY 19451
by guest on July 10, 2018http://w
ww
.jbc.org/D
ownloaded from

glycoprotein gp120 by surface plasmon resonance analysis. Asshown in Fig. 6B, BCA dose-dependently bound to the gp120immobilized on a sensor chip CM5. From the kinetic analysis,the association constant (KA) of BCA-gp120 interaction wascalculated to be 3.71 � 108 M�1.Anti-influenza Virus Activity of BCA—The anti-influenza
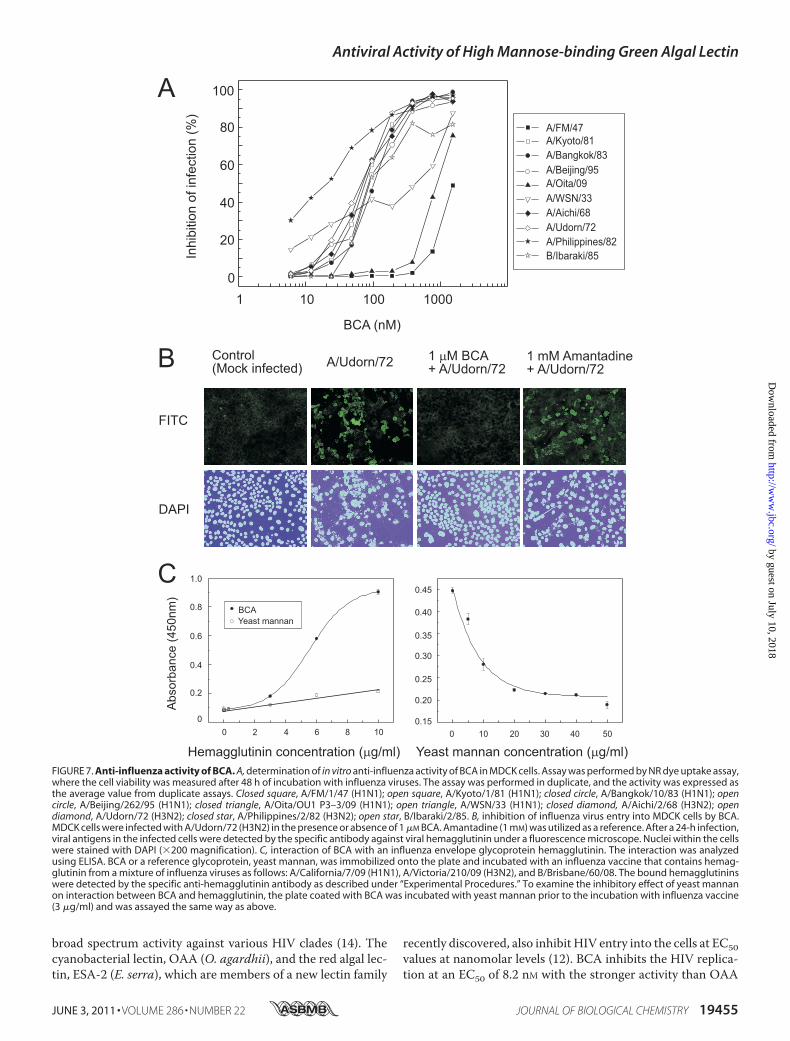
virus activity of BCAwas investigated with respect to inhibitingreplication in MDCK cells by the NR dye uptake assay. Teninfluenza A virus strains, including laboratory-adapted strainsand one influenza virus B strain, were tested for their BCAsensitivity (Fig. 7A). As shown in Table 1, BCA showed antiviralactivity for most of influenza virus strains tested, even thoughthe EC50 varied depending on the strains. Philippines/2/82
(H3N2) was the most sensitive strain to BCA. Interestingly,BCA showed stronger inhibition against H3N2 subtypes atEC50 values of 18.8–74.2 nM, whereas it was much weaker forH1N1 subtypes showing EC50 values of 79.3–1590.2 nM. Theclinical isolates of the recent pandemic strain, swine-origininfluenza virus, A/Oita/OU1 P3-3/09 (H1N1), was also suscep-tible to BCA, although the degree of susceptibility (EC50 of 820nM) was much lower compared with the H3N2 strains. A labo-ratory-adapted strain, A/PR8/34 (H1N1), was the only strainthat is insensitive to BCA. This lectin was also effective againstinfluenza B strain B/Ibaraki/2/85 at a moderate level.Immunofluorescence Microscopy—To explore the mecha-
nism of inhibition of influenza virus infection by BCA, the pres-
FIGURE 4. Structures of PA-oligosaccharides used in this study. Open circle, galactose; open square, N-acetylgalactosamine; closed diamond, glucose; closedsquare, N-acetylglucosamine; closed circle, mannose; open triangle, fucose; open pentagon, xylose.
Antiviral Activity of High Mannose-binding Green Algal Lectin
19452 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 286 • NUMBER 22 • JUNE 3, 2011
by guest on July 10, 2018http://w
ww
.jbc.org/D
ownloaded from
ence of viral antigen in the infected cells was observed in thepresence or absence of BCA using immunofluorescencemicroscopy.After 24 hpostinfectionwithA/Udorn/72 (H3N2),viral antigens were detected with the specific anti-hemaggluti-nin antibody. Fig. 7B shows that BCA efficiently inhibited influ-enza virus entry into the cells, whereas the virus invaded andproliferated in the host cells in the absence of BCA. The viralion channel (M2 protein) inhibitor, Amantadine, did not pre-vent virus invasion into the cells.Direct Interaction of BCA with Influenza Viral Hemag-
glutinin—To examine whether BCA directly binds to the oligo-saccharide on enveloped glycoprotein (hemagglutinin), ELISAswere performed using an influenza vaccine preparation, whichcontains hemagglutinin of A/California/7/09 (H1N1), A/Victo-ria/210/09 (H3N2), and B/Brisbane/60/08. As shown in Fig. 7C(left panel), obvious binding of hemagglutinin to the immobi-lized BCA was demonstrated. In contrast, hemagglutinin didnot bind to a reference glycoprotein (yeast mannan). The inter-action between hemagglutinin and BCA was significantlyinhibited by the presence of yeast mannan bearing high man-nose glycans (Fig. 7C, right panel), indicating that BCA actuallybinds to the hemagglutinin through high mannose glycans.
DISCUSSION
High mannose-binding algal lectins are ones from theexpected and promising compounds for anti-HIV or antiviralagents such asmicrobicides, as they exhibit exclusive specificity
toward certain oligosaccharide(s) on the surface of the viruswith high affinity, thereby inhibiting virus entry into the hostcells. Here, we report that BCA is the first HIV- and influenzavirus-inhibiting protein from the green algae, which showsstrict specificity for high mannose oligosaccharide but unprec-edented mode of oligosaccharide recognition. We also eluci-dated the primary structure of BCA and its distinctive struc-tural features.Several green algal species belonging to the orders Dasycla-
dales and Siphonocladales, to which B. coacta belongs, havebeen found to use a noncanonical genetic code where TAA andTAG encode glutamine instead of translation termination (31–33), as well as oxymonads (34, 35), diplomonads (36, 37), andsome ciliates (38, 39). We found that BCA cDNA uses the non-canonical genetic code as shown in Fig. 2. To confirm that B.coacta itself uses the unusual genetic code, the full-lengthcDNA encoding the translation elongation factor-1� (EF-1�),which was found to use the noncanonical genetic code in somegreen algae (33), was cloned from B. coacta (data not shown;DDBJ/EMBL/GenBankTM accession number AB604604).However, we did not find TAA and TAG codons in its ORF.Then we nonspecifically surveyed other genes that may use theunusual genetic code in the full-length cDNApool derived fromB. coacta (see supplemental Fig. 1 for details). As a result, wefound another gene, ribosomal protein L37a, that uses the non-canonical genetic code inB. coacta (supplemental Fig. 1; DDBJ/
FIGURE 5. Binding activities of BCA to PA-oligosaccharides. Binding activity was expressed as a ratio (%) of the amount of a bound oligosaccharide to thatof an added oligosaccharide. The assay was performed in duplicate for each PA-oligosaccharide, and the activity is expressed as the average value fromduplicate assays. The assays were reproducible without any significant difference.
Antiviral Activity of High Mannose-binding Green Algal Lectin
JUNE 3, 2011 • VOLUME 286 • NUMBER 22 JOURNAL OF BIOLOGICAL CHEMISTRY 19453
by guest on July 10, 2018http://w
ww
.jbc.org/D
ownloaded from
EMBL/GenBankTM accession number AB604605). Althoughwe did not find TAA and TAG codons in putative ORF ofalmost all genes cloned in this experiment, they used theTGA codon as a termination, but not TAA and TAG, as wellas B. coacta EF-1� gene. Thus, it is strongly suggested that B.coacta uses a noncanonical genetic code, and the algal spe-cies using the unusual genetic code may exist in the orderSiphonocladales.The molecular mass (13,812 Da) of BCA determined by
ESI-MS was distinct from the calculated one (17,561 Da) fromthe deduced amino acid sequence of BCA cDNA, which con-sisted of 161 amino acids, excluding the signal peptidesequence. Concerning this discrepancy, the molecular mass(13,812 Da) calculated from the portion (20–144 amino acids)of three tandem-repeated sequences coincides well with thedetermined mass by ESI-MS. In the sequence analyses of the
peptide fragments of PE-BCA, none of fragments derived fromthe C-terminal region (145–180 amino acids) deduced fromBCA cDNA was obtained. This suggests that the precursor ofBCA may be post-translationally modified for its C-terminaltruncation as seen in the other plant lectins (40), including theGNA-related lectin family (41, 42), resulting in synthesis of themature BCA consisting of repeated domains alone. The occur-rence of three internal repeats and both the N- and C-terminalpotential truncation regions of BCA led us to survey the simi-larity of this protein to GNA, which also consists of three sub-domains, signal peptide, and C-terminal propeptide. Interest-ingly, BCA has sequence motifs similar to the carbohydrate-binding site of GNA-related lectins. Nevertheless, the overallsequence similarity was quite low between BCA and GNA-re-lated lectins at an undetectable level by the normal homologysearch. It should be noted that both the signal peptide and theC-terminal propeptide of GNA are necessary for trafficking tothe vacuole (42). Moreover, the importance of the C-terminalpropeptide of GNA to temporarily inactivate the carbohydrate-binding ability in the endoplasmic reticulumhas been proposed(42). The presence of a putative vacuolar targetingmotif of BCAand the partially conserved GNA-like carbohydrate-bindingsite suggests the similar physiological role(s) of BCA.GNA-related lectins have been distinguished either as single-
domain lectins with an exclusive specificity toward mannose/oligomannosides or as two-domain lectins that acquired adiverse carbohydrate specificity (2). From the aspects of a sim-ple monomeric structure consisting of a single GNA-likedomain and the strict specificity for highmannose oligosaccha-rides of BCA, this lectin more closely resembles the single-do-main GNA-related lectins. Of the three subdomains withconsensus amino acid sequences (QDNVY), each of which cor-responds to characteristic mannose-binding sites of GNA-re-lated lectins, putative carbohydrate-binding sites of BCA werepartially conserved at subdomains I and II of GNA-related lec-tins, but it underwent a significant change at subdomain III.This might reflect the different specificity of BCA from othertypical GNA-related lectins, but the mode of oligosacchariderecognition of BCA should be clarified by the structural analy-ses of BCA-oligosaccharide complexes. It is noteworthy thatBCA has been predicted to have three �-helices within itsmonomeric molecule, each located in tandem repeat domainsas deduced from the secondary structure (supplemental Fig. 2).This suggests that the three-dimensional structure of BCAmight be distinct from the typical GNA-related lectins, whichexhibit a �-prism structure built up of three subdomains, eachconsisting of four strands of antiparallel �-sheets (43). More-over, it is likely that plant GNA-related lectins may haveevolved through the process unrelated from BCA, because pro-karyotic proteins with the GNA domain(s) share a highsequence similarity with plant GNA-related lectins (44, 45) butnot with BCA. Thus, the biosynthetic process of this algal lectinis an interesting target to be further investigated, including theidentification of its C-terminal amino acid.Cyanobacterial lectins, CV-N (N. ellipsosporum) and scyto-
virin (S. varium), show the strong anti-HIV activity at EC50values at a picomolar level. A red algal lectin GRFT is thestrongest HIV entry inhibitor with the EC50 of 40 pM and has
FIGURE 6. Anti-HIV-1 activity of BCA. A, determination of in vitro anti-HIV-1activity of BCA in MT-4 cells. Anti-HIV-1 activity was determined using a col-orimetric (MTT) method. HTLV-IIIB strain was used as a virus strain of HIV-1.White and black circles indicate mock-infected (E) or HIV-1-infected (F) cellswith BCA. EC50 and EC90 values of BCA for the inhibition of HIV-1 infectionwere 8.2 and 13.2 nM. The assay was performed in triplicate, and the activitywas expressed as the average value from triplicate assays. B, interaction ofBCA with a recombinant HIV envelope glycoprotein gp120. The interactionwas analyzed by surface plasmon resonance (SPR). Each sensorgram repre-sents the BCA binding to gp120 on sensor chip CM5. Ninety �l of BCA solu-tions (15.6, 31.3, 62.5, 125, and 250 nM) were injected into the flow cells at 30�l/min for 3 min. The response in resonance units (RU) is plotted against time(seconds). Binding kinetics of the interaction between BCA and gp120 werecalculated by fitting the data to Langmuir model for 1:1 binding. ka, associa-tion rate constant; kd, dissociation rate constant; KA, association constant; andKD, dissociation constant.
Antiviral Activity of High Mannose-binding Green Algal Lectin
19454 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 286 • NUMBER 22 • JUNE 3, 2011
by guest on July 10, 2018http://w
ww
.jbc.org/D
ownloaded from
broad spectrum activity against various HIV clades (14). Thecyanobacterial lectin, OAA (O. agardhii), and the red algal lec-tin, ESA-2 (E. serra), which are members of a new lectin family
recently discovered, also inhibit HIV entry into the cells at EC50values at nanomolar levels (12). BCA inhibits the HIV replica-tion at an EC50 of 8.2 nM with the stronger activity than OAA
FIGURE 7. Anti-influenza activity of BCA. A, determination of in vitro anti-influenza activity of BCA in MDCK cells. Assay was performed by NR dye uptake assay,where the cell viability was measured after 48 h of incubation with influenza viruses. The assay was performed in duplicate, and the activity was expressed asthe average value from duplicate assays. Closed square, A/FM/1/47 (H1N1); open square, A/Kyoto/1/81 (H1N1); closed circle, A/Bangkok/10/83 (H1N1); opencircle, A/Beijing/262/95 (H1N1); closed triangle, A/Oita/OU1 P3–3/09 (H1N1); open triangle, A/WSN/33 (H1N1); closed diamond, A/Aichi/2/68 (H3N2); opendiamond, A/Udorn/72 (H3N2); closed star, A/Philippines/2/82 (H3N2); open star, B/Ibaraki/2/85. B, inhibition of influenza virus entry into MDCK cells by BCA.MDCK cells were infected with A/Udorn/72 (H3N2) in the presence or absence of 1 �M BCA. Amantadine (1 mM) was utilized as a reference. After a 24-h infection,viral antigens in the infected cells were detected by the specific antibody against viral hemagglutinin under a fluorescence microscope. Nuclei within the cellswere stained with DAPI (�200 magnification). C, interaction of BCA with an influenza envelope glycoprotein hemagglutinin. The interaction was analyzedusing ELISA. BCA or a reference glycoprotein, yeast mannan, was immobilized onto the plate and incubated with an influenza vaccine that contains hemag-glutinin from a mixture of influenza viruses as follows: A/California/7/09 (H1N1), A/Victoria/210/09 (H3N2), and B/Brisbane/60/08. The bound hemagglutininswere detected by the specific anti-hemagglutinin antibody as described under “Experimental Procedures.” To examine the inhibitory effect of yeast mannanon interaction between BCA and hemagglutinin, the plate coated with BCA was incubated with yeast mannan prior to the incubation with influenza vaccine(3 �g/ml) and was assayed the same way as above.
Antiviral Activity of High Mannose-binding Green Algal Lectin
JUNE 3, 2011 • VOLUME 286 • NUMBER 22 JOURNAL OF BIOLOGICAL CHEMISTRY 19455
by guest on July 10, 2018http://w
ww
.jbc.org/D
ownloaded from

and ESA-2. All of these lectins inhibit HIV infection, based onthe same mechanism that is by the recognition of high affinitybinding to the high mannose glycans of gp120 on the virussurface. Recent studies have shown that the pattern of glycosyl-ation of HIV viral particles depends largely on the derived hostcell lines rather than the strain difference and affects the HIVinfectivity (46). Therefore, susceptibility of different host-de-rived HIV strains to BCA remains to be clarified. Besides thereason of glycomic distinctions in viral particles, the relativedifference of antiviral potency among lectins might be some-what ascribed on their diversifiedmode of highmannose oligo-saccharide recognition.Fig. 8 shows the schematic diagram of high mannose oligo-
saccharide recognized by BCA. This lectin primarily targets the�1–2-linked mannose at the nonreducing end, and clusteringof the �1–2-mannose significantly increases the binding affin-ity for BCA. It is likely that BCA recognizes the nonreducingterminal �1–2-linked mannose(s) rather than the internal one,because the binding activity of BCA was not altered betweenoligosaccharides 16 and 21, in which the only structural differ-ence is the presence of additional �1–2-linked mannose at theD1 terminal (Fig. 4). This is also applicable in comparisonbetween oligosaccharides 14 and 18,where internal�1–2-man-nose at the D1 arm did not affect the binding activity.It is known, for instance, that highmannose-binding proteins
such as human HIV-neutralizing antibody 2G12 or CV-N pre-fer the D1 and D3 arm rather than the D2 arm (47, 48). Actino-hivin, an anti-HIV lectin from the actinomycete, which binds
high mannose oligosaccharide, prefers the nonreducing termi-nal�1–2-mannose at theD1 arm and shows the highest affinityfor the oligosaccharide having a combination of D1 and D3�1–2-mannoses (49). MVL specifically recognizes 4–5 units ofthe oligomannose core (50), whereas scytovirin prefers the D3arm of Man9GlcNAc2 (51). Both ESA-2 and OAA prefer theexposed �1–3-mannose in the D2 arm (12, 15). In contrast tothese lectins, BCA did not show preference for certain arms butrather showed the preference for clustering of �1–2-mannoseresidues at the nonreducing terminus regardless of the attachedarm position. This unique oligosaccharide-binding property ofBCA, the exclusive recognition of the �1–2-mannose cluster,might be explained somewhat by the “cluster effect” of multi-valent ligands. Multivalent ligands sometimes display signifi-cant increases in functional affinity for lectins. For instance, thehepatic Gal/GalNAc receptor on the surface of mammalianhepatocytes has been shown to discriminate the cluster ofgalactose residues effectively (52). The number of Gal residues/cluster and the distance between the Gal residues are criticaldeterminants of the binding affinity. Studies on the effect ofmultivalent presentation of mannose ligands for concanavalinA showed the dramatic enhancement in functional affinity ofmannose-containing polymers relative to the monovalentderivatives (53). Thus, in biological systems, binding affinityand specificity of lectins can sometimes be enhanced by usingthe multivalent saccharide ligands, resulting in the successfulrecognition in many biological events such as cell-cell interac-tions. Unfortunately, however, the biological ligand(s) of BCAhave not been identified like other algal lectins. Very recently,one of the two oligomannose-binding sites of cyanobacteriallectin MVL has shown to have a catalytic activity to cleavage ofchitin fragments to GlcNAc (54). Therefore, it would be impor-tant to survey the catalytic activity of algal lectins, includingBCA, to gain insight into their biological significance.The mechanisms by which high mannose-binding lectins
inhibit HIV adsorption to its target cells are also applicable forother enveloped viruses. Indeed, CV-N showed a wide varietyof antiviral activity for enveloped viruses such as influenzaviruses, Ebola virus, human herpesvirus 6, and hepatitis C virus(16–19). The emergence of the recent pandemic strain, swine-origin influenza virus (H1N1-2009) led us to address the poten-tial anti-influenza virus activity of BCA, to explore the possibil-ity as a novel antiviral agent. BCA showed potent anti-influenza
FIGURE 8. Schematic representation of oligosaccharide recognized by BCA. The nonreducing terminal �1–2-mannose cluster is strongly recognized byBCA.
TABLE 1In vitro activity of BCA against various influenza strainsIC50 value of BCA was determined with MDCK cells as �3623.2 (nM).
Virus Strain EC50
Antiviralindexa
nMInfluenza A PR8/34 (H1N1) � �
FM/1/47 (H1N1) 1,590.22 � 3.85 �2.3Kyoto/1/81 (H1N1) 79.34 � 7.00 �45.7Bangkok/10/83 (H1N1) 106.47 � 8.05 �34.0Beijing/262/95 (H1N1) 96.52 � 13.25 �37.5Oita/OU1 P3-3/09 (H1N1) 819.58 � 8.33 �4.4WSN/33 (H1N1) 416.53 � 137.3 �8.7Aichi/2/68 (H3N2) 74.22 � 10.8 �48.8Udorn/72 (H3N2) 68.47 � 4.24 �52.9Philippines/2/82 (H3N2) 18.80 � 0.95 �192.7
Influenza B Ibaraki/2/85 81.82 � 9.82 �44.3aAntiviral index represents the ratio of IC50 to EC50.
Antiviral Activity of High Mannose-binding Green Algal Lectin
19456 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 286 • NUMBER 22 • JUNE 3, 2011
by guest on July 10, 2018http://w
ww
.jbc.org/D
ownloaded from

activity against most of all the influenza virus strains tested,except a laboratory-adapted strain PR/8/34 (H1N1). The recentpandemic strain, A/Oita/OU1 P3–3/09 (H1N1), was also inac-tivated by BCA, but the sensitivity wasmuch lower (EC50 of 800nM) compared with other sensitive strains. It has been reportedthat the pandemic H1N1 viruses, which have only singleN-gly-cosylation sequons on the head of hemagglutinin, were resist-ant to the innate immune proteins of the collectin superfamily,whereas the recent seasonal H1N1 possessing three to foursequons was sensitive to these proteins (55). The degree of gly-cosylation of viral hemagglutinin would be a key determinantfor BCA sensitivity because the strain PR/8/34, which is devoidofN-glycosylation sequons, was resistant to BCA. Interestingly,the inhibitory activities of BCA for H3N2 subtypes were rela-tively higher than for H1N1 subtypes. It seems that H3N2 sub-types are more sensitive to �1–2-linked mannose-bindinglectins than H1N1 subtypes, because this tendency was alsoobserved for CV-N. CV-N has been reported to neutralizeH3N2 subtype (A/Sydney/05/97) completely, whereas H1N1subtype (A/Beijing/262/95) is still infectious by the same CV-Ntreatment (16). Similarity in virus-inactivating profiles betweenBCA and CV-N might be primarily ascribed on the selectiverecognition of the �1–2-linked mannose unit.
As for influenza viruses, the hemagglutinin of glycoproteinappears to be a potential target for antiviral agents becausecertain glycosylation sites on the hemagglutinin of influenzaA viruses are highly conserved and show site specificity forattached glycans (56). It has been demonstrated that thehemagglutinin 1 (HA1) subunit has a high mannose oligo-saccharide at site 65, which contains almost exclusivelyMan9GlcNAc2, near the receptor-binding site.
As the number of amino acid sequences that encode thepotentialN-glycosylation site, referred to as sequons, in humaninfluenza A hemagglutinin H3 subtype HA1 has increased overtime, more recent strains might be more susceptible to lectins.This increase in the number of sequons is only found in thehuman H3 subtype of influenza A virus and not in H1 subtype(57). In this connection, we have observed the increased BCAsensitivity to Philippines/2/82 (H3N2) compared with the ear-lier isolated strain, Aichi/2/68 (H3N2). Similarly, H3N2 strainshave become more sensitive to human surfactant protein Dover time, in accordance with the increase of attached glycanson hemagglutinin (58). Hartshorn et al. (58) showed that sensi-tivity of H3N2 to surfactant protein D increased with multipli-cation of the glycosylation site, and the extent was greater whenthe glycosylation site was introduced in close proximity to thereceptor-binding site. Further experiments will be needed toverify our results using more recent H3N2 strains, because thenewly introduced glycosylation site does not always have highmannose oligosaccharides with �1–2-linked mannose units; inother words, increased sequons might not directly link to theBCA sensitivity.Effective viricidal agents are continuously in high demand
because the vaccine supply is sometimes not on time. Anti-body-based medicine might have some defects as the virusmutates with high frequency by antigenic drift or antigenicshift. Furthermore, they usually could not be fully active againstthe different subtypes such as H3N2 or H1N1. At this point,
carbohydratemoieties on the virus surface alternatively may begood targets for medicinal treatment because they exist in highabundance in most influenza strains, especially in the morerecent strains. Some virus strains have been developing anincreased number of glycosylation sites to evade antibody pres-sures by changing antigenicity (59). Therefore, it is advanta-geous to use lectins prophylactically, as they universally inacti-vate awide range of virus strains anddifferent subtypes. Furtherevaluation of BCA safety concerning cytotoxicity and inflam-matory activity should be required, because some lectins suchas CV-N show various side effects such as mitogenic activityand stimulation in the production of a wide variety of cytokines(60).
Acknowledgments—We are grateful to Dr. Takemasa Sakaguchi andDr. Akira Nishizono for the generous gifts of influenza viruses.
REFERENCES1. Botos, I., and Wlodawer, A. (2005) Prog. Biophys. Mol. Biol. 88, 233–2822. Van Damme, E. J., Nakamura-Tsuruta, S., Smith, D. F., Ongenaert, M.,
Winter, H. C., Rouge, P., Goldstein, I. J., Mo, H., Kominami, J., Culerrier,R., Barre, A., Hirabayashi, J., and Peumans, W. J. (2007) Biochem. J. 404,51–61
3. Barre, A., Bourne, Y., Van Damme, E. J., Peumans, W. J., and Rouge, P.(2001) Biochimie 83, 645–651
4. Barre, A., Van Damme, E. J., Peumans, W. J., and Rouge, P. (1996) PlantPhysiol. 112, 1531–1540
5. Fouquaert, E., Smith, D. F., Peumans, W. J., Proost, P., Balzarini, J., Sav-vides, S. N., andDamme, E. J. (2009)Biochem. Biophys. Res. Commun. 380,260–265
6. Ziołkowska, N. E., and Wlodawer, A. (2006) Acta Biochim. Pol. 53,617–626
7. Balzarini, J. (2007) Nat. Rev. Microbiol. 5, 583–5978. Boyd, M. R., Gustafson, K. R., McMahon, J. B., Shoemaker, R. H., O’Keefe,
B. R., Mori, T., Gulakowski, R. J., Wu, L., Rivera, M. I., Laurencot, C. M.,Currens, M. J., Cardellina, J. H., 2nd, Buckheit, R. W., Jr., Nara, P. L.,Pannell, L. K., Sowder, R. C., 2nd, andHenderson, L. E. (1997)Antimicrob.Agents Chemother. 41, 1521–1530
9. Bewley, C. A., Gustafson, K. R., Boyd, M. R., Covell, D. G., Bax, A., Clore,G. M., and Gronenborn, A. M. (1998) Nat. Struct. Biol. 5, 571–578
10. Bokesch, H. R., O’Keefe, B. R., McKee, T. C., Pannell, L. K., Patterson,G.M., Gardella, R. S., Sowder, R. C., 2nd, Turpin, J., Watson, K., Buckheit,R. W., Jr., and Boyd, M. R. (2003) Biochemistry 42, 2578–2584
11. Yamaguchi, M., Ogawa, T., Muramoto, K., Kamio, Y., Jimbo, M., andKamiya, H. (1999) Biochem. Biophys. Res. Commun. 265, 703–708
12. Sato, Y., Okuyama, S., andHori, K. (2007) J. Biol. Chem. 282, 11021–1102913. Balzarini, J. (2007) Antivir. Chem. Chemother. 18, 1–1114. Mori, T., O’Keefe, B. R., Sowder, R. C., 2nd, Bringans, S., Gardella, R., Berg,
S., Cochran, P., Turpin, J. A., Buckheit, R. W., Jr., McMahon, J. B., andBoyd, M. R. (2005) J. Biol. Chem. 280, 9345–9353
15. Hori, K., Sato, Y., Ito, K., Fujiwara, Y., Iwamoto, Y., Makino, H., andKawakubo, A. (2007) Glycobiology 17, 479–491
16. O’Keefe, B. R., Smee, D. F., Turpin, J. A., Saucedo, C. J., Gustafson, K. R.,Mori, T., Blakeslee, D., Buckheit, R., and Boyd, M. R. (2003) Antimicrob.Agents Chemother. 47, 2518–2525
17. Barrientos, L. G., O’Keefe, B. R., Bray,M., Sanchez, A., Gronenborn, A.M.,and Boyd, M. R. (2003) Antiviral Res. 58, 47–56
18. Dey, B., Lerner, D. L., Lusso, P., Boyd, M. R., Elder, J. H., and Berger, E. A.(2000) J. Virol. 74, 4562–4569
19. Helle, F., Wychowski, C., Vu-Dac, N., Gustafson, K. R., Voisset, C., andDubuisson, J. (2006) J. Biol. Chem. 281, 25177–25183
20. O’Keefe, B. R., Giomarelli, B., Barnard, D. L., Shenoy, S. R., Chan, P. K.,McMahon, J. B., Palmer, K. E., Barnett, B.W.,Meyerholz, D. K.,Wohlford-Lenane, C. L., and McCray, P. B., Jr. (2010) J. Virol. 84, 2511–2521
Antiviral Activity of High Mannose-binding Green Algal Lectin
JUNE 3, 2011 • VOLUME 286 • NUMBER 22 JOURNAL OF BIOLOGICAL CHEMISTRY 19457
by guest on July 10, 2018http://w
ww
.jbc.org/D
ownloaded from

21. Hori, K., Miyazawa, K., and Ito, K. (1986) Bot. Mar. 29, 323–32822. Okamoto, R., Hori, K., Miyazawa, K., and Ito, K. (1990) Experientia 46,
975–97723. Lowry, O. H., Rosebrough, N. J., Farr, A. L., and Randall, R. J. (1951) J. Biol.
Chem. 193, 265–27524. Schagger, H., and von Jagow, G. (1987) Anal. Biochem. 166, 368–37925. Sato, Y.,Murakami,M.,Miyazawa, K., andHori, K. (2000)Comp. Biochem.
Physiol. B 125, 169–17726. Larkin, M. A., Blackshields, G., Brown, N. P., Chenna, R., McGettigan,
P. A., McWilliam, H., Valentin, F., Wallace, I. M., Wilm, A., Lopez, R.,Thompson, J. D., Gibson, T. J., and Higgins, D. G. (2007) Bioinformatics23, 2947–2948
27. Bendtsen, J. D., Nielsen, H., von Heijne, G., and Brunak, S. (2004) J. Mol.Biol. 340, 783–795
28. Pauwels, R., Balzarini, J., Baba, M., Snoeck, R., Schols, D., Herdewijn, P.,Desmyter, J., and De Clercq, E. (1988) J. Virol. Methods 20, 309–321
29. Sato, Y., Morimoto, K., Hirayama, M., and Hori, K. (2011) Biochem. Bio-phys. Res. Commun. 405, 291–296
30. Rudiger, H. (1998) Acta Anat. 161, 130–15231. Schneider, S. U., Leible,M. B., andYang, X. P. (1989)Mol. Gen.Genet. 218,
445–45232. Schneider, S. U., and de Groot, E. J. (1991) Curr. Genet. 20, 173–17533. Gile, G. H., Novis, P. M., Cragg, D. S., Zuccarello, G. C., and Keeling, P. J.
(2009) J. Eukaryot. Microbiol. 56, 367–37234. Keeling, P. J., and Leander, B. S. (2003) J. Mol. Biol. 326, 1337–134935. de Koning, A. P., Noble, G. P., Heiss, A. A., Wong, J., and Keeling, P. J.
(2008) Environ. Microbiol. 10, 65–7436. Keeling, P. J., and Doolittle, W. F. (1996) EMBO J. 15, 2285–229037. Keeling, P. J., and Doolittle, W. F. (1997)Mol. Biol. Evol. 14, 895–90138. Tourancheau, A. B., Tsao, N., Klobutcher, L. A., Pearlman, R. E., and
Adoutte, A. (1995) EMBO J. 14, 3262–326739. Lozupone, C. A., Knight, R. D., and Landweber, L. F. (2001)Curr. Biol. 11,
65–7440. Mandal, D. K., Nieves, E., Bhattacharyya, L., Orr, G. A., Roboz, J., Yu,Q. T.,
and Brewer, C. F. (1994) Eur. J. Biochem. 221, 547–55341. van Damme, E. J., Smeets, K., Torrekens, S., van Leuven, F., Goldstein, I. J.,
and Peumans, W. J. (1992) Eur. J. Biochem. 206, 413–42042. Fouquaert, E., Hanton, S. L., Brandizzi, F., Peumans, W. J., and Van
Damme, E. J. (2007) Plant Cell Physiol. 48, 1010–102143. Hester, G., and Wright, C. S. (1996) J. Mol. Biol. 262, 516–531
44. Peumans, W. J., Barre, A., Bras, J., Rouge, P., Proost, P., and Van Damme,E. J. (2002) Plant Physiol. 129, 1054–1065
45. Parret, A. H., Temmerman, K., and De Mot, R. (2005) Appl. Environ.Microbiol. 71, 5197–5207
46. Krishnamoorthy, L., Bess, J. W., Jr., Preston, A. B., Nagashima, K., andMahal, L. K. (2009) Nat. Chem. Biol. 5, 244–250
47. Botos, I., O’Keefe, B. R., Shenoy, S. R., Cartner, L. K., Ratner, D. M., See-berger, P. H., Boyd, M. R., and Wlodawer, A. (2002) J. Biol. Chem. 277,34336–34342
48. Calarese, D. A., Lee, H. K., Huang, C. Y., Best, M. D., Astronomo, R. D.,Stanfield, R. L., Katinger, H., Burton, D. R., Wong, C. H., andWilson, I. A.(2005) Proc. Natl. Acad. Sci. U.S.A. 102, 13372–13377
49. Tanaka, H., Chiba, H., Inokoshi, J., Kuno, A., Sugai, T., Takahashi, A., Ito,Y., Tsunoda, M., Suzuki, K., Takenaka, A., Sekiguchi, T., Umeyama, H.,Hirabayashi, J., and Omura, S. (2009) Proc. Natl. Acad. Sci. U.S.A. 106,15633–15638
50. Williams, D. C., Jr., Lee, J. Y., Cai, M., Bewley, C. A., and Clore, G. M.(2005) J. Biol. Chem. 280, 29269–29276
51. McFeeters, R. L., Xiong, C., O’Keefe, B. R., Bokesch, H. R.,McMahon, J. B.,Ratner, D. M., Castelli, R., Seeberger, P. H., and Byrd, R. A. (2007) J. Mol.Biol. 369, 451–461
52. Lee, Y. C., Townsend, R. R., Hardy, M. R., Lonngren, J., Arnarp, J.,Haraldsson, M., and Lonn, H. (1983) J. Biol. Chem. 258, 199–202
53. Mortell, K. H.,Weatherman, R. V., and Kiessling, L. L. (1996) J. Am. Chem.Soc. 118, 2297–2298
54. Shahzad-ul-Hussan, S., Cai,M., and Bewley, C. A. (2009) J. Am. Chem. Soc.131, 16500–16508
55. Job, E. R., Deng, Y.M., Tate,M.D., Bottazzi, B., Crouch, E. C., Dean,M.M.,Mantovani, A., Brooks, A. G., and Reading, P. C. (2010) J. Immunol. 185,4284–4291
56. Mir-Shekari, S. Y., Ashford, D. A., Harvey, D. J., Dwek, R. A., and Schulze,I. T. (1997) J. Biol. Chem. 272, 4027–4036
57. Zhang, M., Gaschen, B., Blay,W., Foley, B., Haigwood, N., Kuiken, C., andKorber, B. (2004) Glycobiology 14, 1229–1246
58. Hartshorn, K. L., Webby, R., White, M. R., Tecle, T., Pan, C., Boucher, S.,Moreland, R. J., Crouch, E. C., and Scheule, R. K. (2008) Respir. Res. 9, 65
59. Abe, Y., Takashita, E., Sugawara, K.,Matsuzaki, Y.,Muraki, Y., andHongo,S. (2004) J. Virol. 78, 9605–9611
60. Huskens, D., Vermeire, K., Vandemeulebroucke, E., Balzarini, J., andSchols, D. (2008) Int. J. Biochem. Cell Biol. 40, 2802–2814
Antiviral Activity of High Mannose-binding Green Algal Lectin
19458 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 286 • NUMBER 22 • JUNE 3, 2011
by guest on July 10, 2018http://w
ww
.jbc.org/D
ownloaded from

Okuyama and Kanji HoriYuichiro Sato, Makoto Hirayama, Kinjiro Morimoto, Naoki Yamamoto, Satomi
Influenza Viruses Is a Potent Entry Inhibitor of HIV-1 andBoodlea coactafrom the Green Alga
2-Mannose−1αHigh Mannose-binding Lectin with Preference for the Cluster of
doi: 10.1074/jbc.M110.216655 originally published online April 1, 20112011, 286:19446-19458.J. Biol. Chem.
10.1074/jbc.M110.216655Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
Supplemental material:
http://www.jbc.org/content/suppl/2011/04/01/M110.216655.DC1
http://www.jbc.org/content/286/22/19446.full.html#ref-list-1
This article cites 60 references, 20 of which can be accessed free at
by guest on July 10, 2018http://w
ww
.jbc.org/D
ownloaded from





























