Embed Size (px)
Citation preview

HIBERNATION-HYPOTHERMIA AND METABOLISM IN HEDGEHOGS. CHANGES IN WATER AND
ELECTROLYTES
KAMAL S. AL-BADRY* and HODA M. TAHA
Department of Zoology. Faculty 01‘ Sc~cnce, Cairo University. Egypt
Abstract I. Hcdgchogs seem not to suffer water loss during hlbcrnation or hypothermia probabl) through water shifts from the intcrstitium to the cells and.or protein catabolism III tissues.
7. Higher potassium concentrations were retained in hedgehog’s tissues, sodium and chloride ions rose in plasma. but ionic gradients were slightly nffectcd. Probable rexons for increklsed calcium. magnesium and inorganic phosphorus wcrc discussed.
3. Changes recorded during late fall indicated the predisposition of antmals to hibernate. while changes during spontaneous ;tnd full ;trousals showed il reversed phenomenon in the hibcrnntion cycle.
4. .Artiliciall! hypothermic ammals in summer showed results similar to those 81‘ nnturnlly hypo- thermic in \+lntcr. although lower in magnitude. suggesting il specific role of tempcmture in regulating the hibernation cycle.
Ever since Suomalainen (1939) mvestigated the mag- nesium content of hibernating hedgehog’s serum. man! subsequent studies followed the same trend in investigating different ion levels in various mam- malian hibernators during hibernation and hypo- thermia (c’f: Riedesel & Folk, 1954: Zimny & Gregory, 1958: Suomalainen. 1960: Pengellcy & Fisher. 1961; Raths. 1962; Willis. I966: Fisher & Manery. 1967; Edwards & Munday. 1969. 1973: Ferren LV trl.. 1971; Willis (‘I trl.. 1971: Senturia t’f trl.. 1971: Soivio & Kristoffersson, 1973). The aim of their investigations was to find answers to one or more of some detined questions: (I ) how do the main plasma and tissue ions behave during the hibernation cycle a&or under induced artificial hypothermia’! (2) could a trigger of hibernation. or arousal. be found among them’! and finall> (3) do these ions maintain their gradients under hibernation and hypothermic conditions’?
Magnesium received the full attention of most studies (Kalter & Folk. 1979). whereas the available data on other electrolytes in hibernating animals are very unsatisfactory. Large variations in plasma ionic concentrations associated with hibernation have been reported by Fisher & Manery (1967). The lack of con- sistency between the available data on blood ions and water balances correlated wjith their changes during the hibernation cycle was dso pointed out by Pengel- le! & Kully (lY67). Nevertheless. very little infor- mation is available on electrolyte concentration and their fluctuation in tissues. other than blood. of hiber- nators.
In an earlier work. Al-Badry & Taha (1982) studied the plasma and tissue levels of amino acids and some defined organic components. during hibernation cycles and induced hypothermia, in the oriental hedge-
* Present address: Biolog) Department. Faculty of Science, Sana’a University. Sana’a. Yemen Arab Republic.
hog Herni~chirl~rs ouritlrs. In the present study. the work has been extended to investigate water and iI,- organic ions, in the same hedgehog species. in an attempt to elucidate their possible role during hiber- nation and/or hypothermia.
hlATERIALS AUD \llXHODS
The xumal used in the present investigation 1s the orlcn- tal long-enred hedgehog Hwzrwhi,zfr.s cluritus ocy~‘pt~r~z
Fischer. Sixty individuals. almost all adult males. were col- lected during e~lrly summer, reared in the laboratory .md d;lily provided with food and water. These ammals were tcstcd in groups:
Group I. Active animals in summer (Temperature 2s 30 C).
Group 2. Active animals in autumn (Temperature 7@25 C).
Group 3. Hypothermic animals during their midhiberna- tion (Temperature 5.0 i 0.2 C).
Group 4. Artificially hypothermic anim~ds in summer (Temperature 5.0 k 0.1 C).
Group 5. Awlike animals. during spontnneous xrouwl. in \Qinter (Temperature 15 70 Cl.
Group 6. Awxke animals ;Ifter full ;~rousal in c:irl> sprlnp (Temperature 30 25 C).
Animals were treated xcording to the methods pre- viously described by Al-Budry & Taha (1982).
In all cxes. animals were sacrifced by dec;lpitatlon. Blood was collected III hcparinizcd sample or centrifuge tubes. Plasma aas obtamed by centrlfugmg blood for 20 mln at 3000 r.p.m. in :L refrigerated centrifuge. it W:LS then quickly frozen xnd kept for analysis.
Immedilltely after decapitation. the brains were excised from the skulls. similarly liver slices were tnken as rapidly as possible. Blood and tissue samples were directly dried in an own at 103 k 7 C until constant weight. Water content aas estimated in dried blood and tissue samples, after fat extraction. and \%a expressed as a percentage of fat-free wet weight.
The dried tissues Mere dgested in 50”,, mtrlc xid. brought to a knoan volume using distilled water and then ancdyscd for electrolytes.

4 i 6 KAMAL S. AL-BADRY and HODA M. TAHA
Sodium and potassium m plasma and tissues. as well as calcium ,Ind magnesium in tissues were determined by the Pyc Unlcnm atomic absorption Model SP 1900. Tissue inorgamc phosphorus was determined by the method of Fiske & Subbarow (1925). Plasma calcium. magnesium and inorganic phosphorus were determined using the bio Llerieux reagent kits. folloslng their instruction manuals. Concentraticyns of plasma chloride were estimated by the method described by Schales & Schales (1941). the data \\crc checked ngamst the Corning chloride meter Model 920 and range of values did not exceed a maximum error of I cl II
RESLYLTS AND DISCUSSION
The oriental hedgehog species, used in the present investigation, displayed the phenomenon of hiber- nation. It became torpid in November, passed through 4-5 short homeothermal periods that divided the season into hibernation cycles. During the season, the cycles lengthened at first but became constant after about six weeks with a constant 15-20 day heter- othermal period. Accordingly. water and electrolytes were analysed in six animal groups, the first was tested in late autumn before hibernation started, four others while at different phases of the hibernation cycle: whereas the sixth was tested under deep hypo- thermia during summer. To indicate if the ionic gradients across hepatic and cerebral cells were main- tained during hibernation and arousal, the ratios between ion concentrations in tissues and plasma were considered (Edwards & Munday, 1969; Bito & Roberts. 1974).
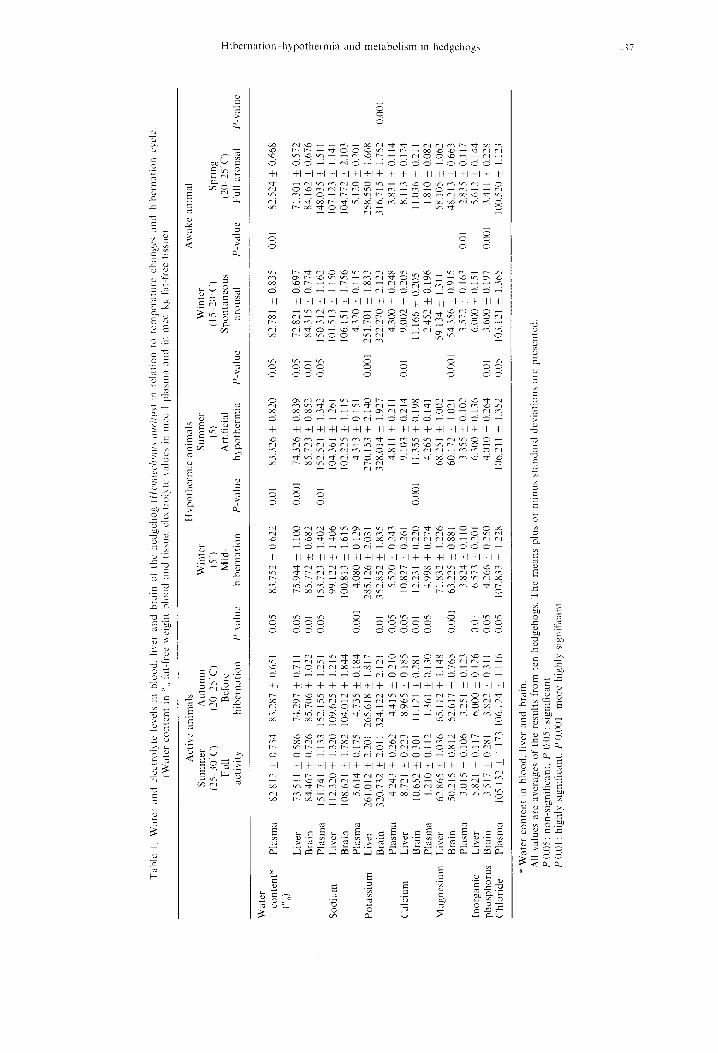
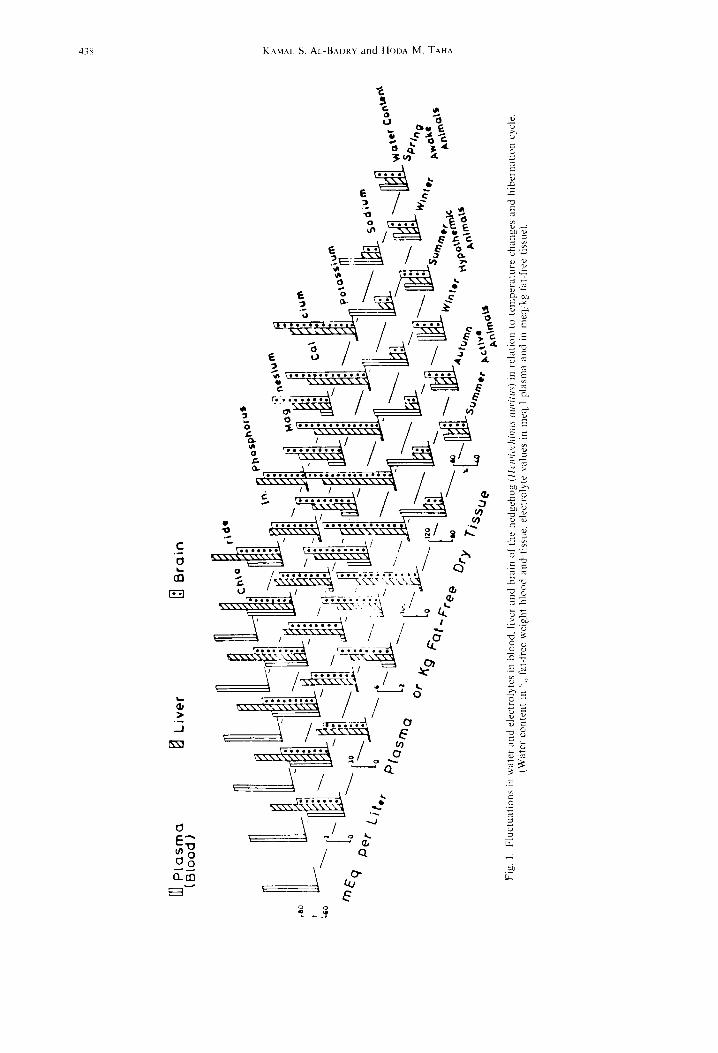
Statistical analysis of the data was carried out using the student’s r-test and the means + standard devi- ations are presented in Table 1. The data of animals active in autumn. hypothermic in winter and summer were compared with those of animals active during summer: whereas the data of aroused animals were compared with those deeply hypothermic during mid- winter. For the sake of clarity all results were also graphically depicted in Fig. 1.
Water and electrolyte levels lie within the normal mammalian range (Senturia et al., 1972). Our results revealed higher w,ater, potassium and calcium con- tents in brain than in liver; whereas magnesium and inorganic phosphorus were consistently lower. How- ever. sodium levels were almost similar in both tissues.
Though fluctuations in blood and tissue water con- tents, within the different phases of hibernation, were slight they were significant in hypothermic and aroused animals. Thus. blood water increased by l.lil”,, during hibernation but returned to normal lcvcls during arousal. Regarding tissue water. notice- able changes were recorded in hepatic water that in- creased by 3.3 I Y,‘, during hibernation but decreased by 4.1 I and 6.1 I”,, during spontaneous and full arousals respectively. It seems probable that brain tissue could tolerate anhydremia than liver tissue. Among the few reports dealing with brain water changes during hypothermia. Saarikoski (1970) indicated that the water content of cerebral cortex and brain stem slices from hypothermic rats and hedgehogs was almost unchanged. In this connection, the water potential or osmolarity of brain tissue was reasonably assumed to be the same as that of blood. This assumption was
based upon the relatively free movement of water between blood and brain and the lack of active trans- port of water in mammalian brains (Pappius. 1963).
A similar pronounced rise in the liver water con- tent, during hibernation was reported by Bintr 61 Rie- desel (1967) as being unique to mammalian hiber- nators. They attributed their results to water storage in the liver and/or reduced metabolic activity of this organ during hibernation. However. these authors reported a concordant fall in blood water. In this con- nection, Denyes & Hassett (1960) could not demon- strate any statistically significant difference in the dry liver weight of awake. cold-exposed golden ham- sters. compared with hibernating ones. /II vitro studies demonstrated that wjater content of several tissues in homeotherms increased with tissue cooling (Fisher & Manery. 1967).
Generally, our data suggest capability of hibernat- ing and hypothermic hedgehogs to maintain their tissue water probably through water shifts from the interstitium to the cells during hibernation. Catah- olism of proteins in tissues. illustrated in the present hedgehog species (Al-Badry & Taha. unpublished work) may be important for water balance. Our results are concordant with those of Kristoffersson PI trl. (1965), Ciausen & Storesund (1971) and BintL (It rrl. (1979) who showed that hedgehogs and squirrels did not suffer water loss during hibernation and or star- vation.
The electrolyte pattern, presented here, indicates slight and non-significant rise in plasma sodium and chloride ions during hibernation (1.3”,, and 2.5”,, re- spectively); during arousal. a slight but significant de- crease was encountered. The changes in tissue sodium were more pronounced with an opposite pattern to that of plasma. The maintenance of sodium ion balance, during the hibernation cycle. was slightly dis- turbed, since the tissue:plasma ratio changed from 0.740. during activity, to 0.674. during hibernation. but almost returned normal after full arousal.
Different results were encountered in potassium ion drifts between activity. hibernation and arousal. The data show that plasma levels gradually declined with onset of the hibernation cycle. reached minimum dur- ing mid-hibernation. then eventually recovered after arousal. The opposite pattern was recorded for liver and brain potassium levels; where a trend to increase during hibernation and to decrease at arousal WIS
noticed. However. changes in potassium gradients were more remarkable than in sodium, during deep hypothermia. but similarly returned to normal levels in arousal.
The results indicate that an essential feature of the hedgehog’s tissues, similar to that of other mammalian hibernators (Willis tit ul., 1971). was their ability to retain higher potassium concentrations and to ex- clude sodium at the low temperatures characteristic of deep hibernation. The observed changes in the ionic gradients give an indication that the active processes maintaining the ionic gradients across the cells. and subsequently the electric potentials of cell membranes in the hedgehog’s liver and brain tissues were slight11 affected. Nevertheless, they could maintain the func- tionally essential ion gradients and the excitability of membranes more than hypothermic non-hibernators (Saarikoski, 1970).
Hlhernation~h~pothermla and metahollsm in hedgehogs
KAMAI. S. AL-BADKY and HODA M. TAHA

Hibernation hypotherrma and metabolism in hedgehogs 439
Our present data contradict those given by Fisher & Manery (1967) who postulated that lowering the temperature of mammalian tissues would induce ion diffusion and reduce the activity of the involved trans- port enzymes thus leading to a lowered active trans- port of sodium out of the cells and potassium into the cells. However. the same authors argued that their opinion was based upon an iu vitro study of hypo- thermic tissues. They offered no guarantee that the same is true for in GUO study. They also pointed out that their results were in disagreement with other irl ri~o results on bats and hedgehogs.
Nevertheless. our results are basically in agreement with those of Suomalainen (1960); Pengelly & Kelly (1967); Clausen & Storesund (1971). The last authors reported that hibernation tended to alter the par- tition of electrolytes between tissues and plasma and resulted in conserved extracellular sodium and chlor- ide ions relative to the intracellular ions. Our findings agree also with those of Willis rt al (1971) who indi- cated that the cerebral cortex and liver tissues of hi- bernating ground squirrels and hamsters retained high potassium content and that kidney and liver might compensate for loss. due to cold, in other tissues. In this connection. Edwards & Munday (1974) correlated the increased plasma sodium and de- creased potassium in hedgehogs with the increased :Lctivity of the zona glomeruloza. in winter. resulting in altered renal excretion of these two ions.
Our results on chloride ion shifts in plasma. though almost non-significant. are consistent with those of Senturia l’r (I/. (1972). The fuctuations of this ion sccmcd to follow those of sodium. However. Soivio & Kristoffersson (lY74) reported constant levels of sodium and chloride in hedgehog’s plasma during hi- bernation. but they presented similar falls in potas- sium level.
The calcium. magnesium and inorganic phosphate contents of plasma and tissues. during hibernation. \\crc high but returned normal after full arousal. Among these electrolytes. magnesium received plenty of studies in hedgehogs as well as in other mam- malian hibernators (for reviews. ~1’ Suomalainen, 1960). This investigator suggested that the increase in magnesium was ;I typical feature of hibernation in man) heterotherms. Furthermore. various investiga- tors linked magnesium ion to the regulation of body temperature in these animals (cf. Platner & Hosko. 195.1: Edwards & Mundaq. 1974). In this connection. Hlto & Roberts (1974) pointed out that the role of magnesium in thcrmoregulation of mammalian hiber- nators could be expected to involve central nervous function: its level in brain and plasma must be con- sidered. On the other hand. the role of magnesium as a narcotizing agent has been established by Pengellq & Fisher (lY61) and confirmed by Pengelley & Chnf- fee ( 1966).
The present results rcveal considerable fluctuations in plasma and tissue magnesium levels between active. h! pothermic and aroused hedgehogs concurred with significant shifts in tissuc:plasma ratios. Thus, plasma magnesium was 3 I.?“,, higher during hibernation than in active state, while hepatic and cerebral magnesium increnscd only by 14.2 and 25.Y”,,. Subsequent shifts in tissue: plasma ratios from 51 .Y5 to 14.37 in liver and from 41.5 to 12.65 in brain were encountered. On the
other hand. aroused vs hibernating animals showed significant decline in magnesium concentrations (63.7”” in plasma, 19.1”, in liver and X.7”;, in brain). Thus concentrations and ionic gradients tended to return to normal.
The highly remarkable elevation of plasma magnc- sium during hibernation may suggest certain tissue sources for magnesium release, at that time. Pengellel & Chaffee (1966) postulated that erythrocytes were the immediate source of magnesium for plasma. How- ever Edwards & Munday (1969) suggested that innc- tivc muscles might be the source. Ferren er trl.. (1071) showed that higher serum magnesium. found in hiber- nators. could occur as a result of an exchange \vith heart and skeletal muscle magnesium. The present results may recall the assumption of Kalter &L Folk (1979) that the fluctuating magnesium levels could be considered a trigger in the hibernation cycle of mam- malian hibernators. The results of Soivio & Kriatof- fcrson (lY74) may support this assumption. These authors showed that plasma magnesium in hedgehogs was highest at the moment when the arousal process was triggered. HoNever. relevant data concerning this assumption are still lacking.
Our results also shou that calcium and inorganic phosphate contents. like magnesium. were higher dur- ing hibernation. The) increased by 30.0 and 76.X”,, in plasma. by 24.1 and ILY”,, in liver. and bq 14.8 and 21 .2”,, in brain. Ho&ever. the changes in their tis- SLX: plasma ratios here very slight and non-signili- cant.
The hqpercalcemla reported here agrees uith the results of Raths (1962). Eduards & Munday (IYbY). Ferren c’f rll. ( I97 I ), Senturia cut trl. (I 972) and Soivio & Kristoffersson (1974); although Suomalainen ( IYhO) and Fisher & Manery (1967) reported constant levels during the hibernation cycle. The rise in plasma cnl- cium could possibl! be attributed to the hyperactl\it> of the parathyroids and’or decalafication of bones and teeth, usually observed during hibernation (Scn- turia or al.. 1972). These authors suggested ;I probable contribution of calcitonin activity. However. Soivio & Kristoffersson (lY74.) suggested some calcium reserve other than bone that may bc responsible for such variations during hibernation.
Since calcium is an integral part of the coupling process. its rise in tissues might be expected as ;I part of the maintenance mechanism. This ma] enhance contraction of muscles and excitability of nervous tissue on arousal. However, the larger rise of calcium in liver than in brain is possibly due to the more pronounced reduction in metabolic and coupling ac- tivities in liver concomitant uith more released cal- cium ions in this organ. The altered plasma protein sq.nthesis, noticed by the present investigators (unpub- lished work). may confirm this suggestion.
.4s far as the authors arc aware, little had been published on the etl’cct of hibernation and expcrimen- tal hypothermia on inorganic phosphate content in mammalian hibernators. The elevated plasma and tissue phosphate levels. reported for the hibernating and hypothermic hedgehogs could possibly be attri- buted to reduction in metabolic activities concordant with diminished high encrgq phosphate bonds. thus resulting in a decline in ATP and creatine phosphate levels with the subsequent release of phosphate ions.

440 KAMAL S. AL-BADRY and HODA M. TAHA
an conformity with our results, Brock (1960) reported significant increase in plasma inorganic phosphate in hibernating and cold-exposed golden hamsters result- ing from diminished non-hydrolyzable phosphates, primarily phospholipids. Furthermore, she pointed out that the rise in body temperature. during arousal induced phosphorylation with concomitant utilization of inorganic phosphate into intermediate compounds.
Similar rise in liver inorganic phosphate was de- scribed by Zimny & Gregory (1958) in the 13-striped ground squirrel after I month hibernation. They related their results with the retention of high tissue glycogen levels since both phosphorus and glycogen are necessary for glycolytic activity during early arou- sal. In this connection, similar glycogen accumulation was previously reported in the oriental hedgehog dur- ing hibernation (Al-Badry & Taha, unpublished work).
The more pronounced drifts in inorganic phosphate of the hedgehog’s brain than in the plasma or liver, recorded in the present study. can contribute to the increased need of this ion for neural activities encoun- tered during spontaneous and full arousals. In this regard. Heckers & Gercken (1974) reported a similar rise in inorganic phosphate in rabbit’s brain after cer- ebral hypothermia, However. a contrastive decline in brain phosphate was described by L’vova (1972) in hibernating and hypothermic gophers, although he indicated that the phosphate needed in arousal for the formation of ATP and creatine phosphate was provided by the brain and not by other tissues.
In general, the present investigation revealed changes in water and electrolytes. within the hiber- nation cycle. contributing to the influence of tempera- ture changes encountered during the different hiber- nation phases. The group of animals, tested in late autumn before the onset of hibernation, showed results indicating the predisposition of these animals to hibernate. The group of animals spontaneously aroused during winter showed similar tendency for change, necessary for wakefulness. like fully aroused animals.
The study of artificially hypothermic animals in summer showed results similar to those obtained for naturally hypothermic in winter but of lower magni- tudes. This contradicts the findings of Edwards & Munday (1974) who recorded changes in tissue elec- trolyte levels of the European hedgehog during artifi- cial hypothermia different from those occurring naturally in winter. In the same connection. our data are also contradicting those of Mahy (1963) and Fer- ren it trl. (197 I). However. the present findings suggest that the oriental hedgehog species may be, like the golden hamster. a permissive hibernator (Hoffman, 1964) and environmental temperature presents the main extrinsic factor. among others. regulating their hibernation cycle.
REFEREYCES
AL-BADRY K. S. & TAHA H. M. (1982) Hibernation-- hypothermia and metabolism in hedgehogs, Changes in free umino acids and related compounds, Camp. B;CJ- (./IPI)I. Plr~.rio/. 72A, 541-547.
BlhTr G. L. & RI~-I>~SIL M. L. (1967) Water content of ground squirrel and laboratory rat tissues. C,,,,lp. B;O- ~~/~cJu. Phwiol. 22, 75-80.
BINTZ G. L., PALMER D. L., MACKIN W. W. & BLANTON F. Y. (1979) Selective tissue catabolism and water balance during starvation in Richardson’s ground squirrels. Contp. Biockem Physiol. 64, 399 403.
BITCI L. 2. & ROBERTS J. C. (1974) The effects of hiber- nation on the chemical composition of cercbrospinal and intraocular fluids, blood plasma and brain tissue of the woodchuck (Marnlotu IUOUUY). Co~lp. Biochrn~. P il~srol. 47, 173-193.
BRO(‘K M. A. (1960) Hibernation and cold storage effects on phosphates in hamster blood. AUK. J. Phxsiol. 199, 195-197.
CLALISEN G. & STORIXJNI~ A. (1971) Electrolyte distribu- tion and renal function in the hibernating hedgehog. Actcl pl~~siol. .s~~d. 83, 4-I’.
DFF~YLX A. & HASSETT J. (1960) A study of the metabolism of liver. diaphragm and kidney in cold-exposed and h- bernating hamsters. &I//. Mus. cornp. Zoo/. /lllrr. 124, 437-456.
EDWARIX B. A. & MLNVAV K. A. (1969) Electrolyte metabolism in the hibernating hedgehog (Erijl~xc,lrs etlro- pclrus). Camp, Biochen1. Physiol. 31. 329-335.
EII~ARIX B. A. & MUNI)A’~ K. A. (1974) Tissue and plasma electrolyte levels in the awake state durmg the year. in summer cold exposure and during arousal from hibernation in the hedgehog (Erimrccwa ~woptrcw.s). Camp. Bioc/w~t~. P1z)xiol. 49, 697 705.
FI:RR~N L. G.. SOITH F. E. & JA(.ORS I-1. K. (I 97 I ) Calcium and magnesium levels in tissues and serum of hihernat- ing and cold-acclimated hamsters. (‘r,+Jio/oq!, 8. 506&508.
FIS~H~.R K. C. & MAU~,RV J. F. (1967) Water and clectro- lyte metabolism in heterotherms. In Mtr~zr~trlrrrrl Hiher- niltio,l (Edited by FIS~UR K. C. et [ii.), Vol. III. pp. 235-279. American Elsevier. New York.
FISKF- C. H. & SUBIIAROM’ Y. (1925) The colourimetric de- termination of phosphorus. J. hioi. C/IUU. 66. 375 400.
H~(.K~Rs H. & GER(.UN G. (1973) Brain metabolism at Ion temperatures. J. &‘c~rochrr~ 23, 503-5 IO.
HOFFMAN R. A. (1964) Terrestrial ammals in the cold: hi- bernators. In Htrdhooh of Pl~y.~io/o~~~~, Section 4: Adap- tation to the Environment (Section editor. DILL. D. B.). pp. 379~403. American Physiological Society. Washing- ton, D.C.
KALUR V. G. & FOLK G. E. (1979) Humoral induction of mammalian hibernation. Cor~[j. Bioc,/zc,,~. Ph\~.vio/. 63. 7- 13.
KRISTOFFERSSON R., SOIVIO A. & SL~OMAL.AIUL\ P. (1965) Studies on the physiology of the hibcrnatlng hedge- hogs- 3. Changes in the hater content of various tissues in relation to seasonal and hibernation cycles. Au,I. Acc~d. .S<,i. /e,l~. A: IV 92. I 17.
L’VOVA S. P. (1972) The effect of artiliclal hypothermia and hibernation on the brain and blood content of certain macroergic phosphate in the gopher. Viol. NUU~ 15, 49-57.
PAPPIUS H. M. (1964) Water transport at cell membranes. Cw. J. 5mhm. 42, 945-953.
PENC~~LL~~ E. T. & CHAFFF~ R. R. J. (1966) Changes in plasma magnesium concentration during hibernation m the golden-mantled ground squirrel (Cirellrrv /atcwr/i,s). Camp. Biochrrn. Physioi. 17. 673%6X I,
PENOFLLLY E. T. & FISHFR K. C. (1961) Rhythmical arou- sal from hibernation in the golden-mantled ground squirrel. Cite/llr.\ iureruirs r~~suwum. Cm. .I. Zcd. 39. 105-120.

Hibernation-hypothermia and metabolism in hedgehogs 111
PI \(;I t LI \r E. T. & KELLY K. H. (1967) Plasma potassium and sodium concentrations in active and hibernating golden-mantled ground squirrels. Cirell2t.s larntr/i.s. C‘CIUI/‘. Hi~~c~/lcv,i. P/tv.\ro/. 20. 299-305.
PI ,II’\IK W. S. & Hosso M J. (1953) Mobility of serum magnesium m hqpothcrmia. ilrn. J. Ph~,.sio/. 174, 277 276
R.\IIIS I’. (1967) lJbcr das Serum Natrium. -Kalium und-Kal/lurn des uinterschlafenden und hypotheri- xhcn Ilamstcrs (Cric,c,iu\ cric,cmv L.). Z. Rio/. 113. 17% ‘04.
RII 111~ I M L 81 I-01 I( Ci. E. (1954) Serum magnesium ch,tnges in hibernation. ,.11x J. Ph~a.sid. 179. 665.
S.\,!Klhoshl J. (1970) Effect of cold and stimulation on res- pirxtwn and on potawum and water contents in bram ‘IIICCS of ;I hlhernator :md :l non-hibernator. AUU. Actid. SC I ~c,iti. I I 165. I 1.
SC ~I,XI IS 0 & SC ml IS S. (1941) A simple and accurate method I;)r the determination of chloride in bioloexal l1Uld$. .1. hir,/. C/i~~fII. 140. x79- 884.
ing comparisons with non-hibernators, guinea pig ;tnd man. Blood ions. Actu physiol. rcuutl. srcppl. 380. 1 I 35
SOIVIO A. & KRISTOFFERSSON R. (1974) Changes 1~ pl,~sm.~ main inorganic ion concentrations during the h~bcr- nation cycle in the hedgehog (Erirwc VIIY I~vc~~~~I ‘II,> I..) Am. Zoo/. Femici 11, I3 I 134.
SUOMALAIN~N P. (1939) Hibernation of the hcdgchog. \‘I. Serum magnesium and calcium. Artificial hitxrxction Also a contribution to the chemical physiology 01 the diurnal sleep. il~1~. Actrd. SC,;. /iwu. (‘4 Si) 7. I hS
SUO!V~AI.AIN~N P. (1960) Stress and neurosccret~o~~ in the hibernating hedgehog. Bu//. 12l1r.s. C’OM/I %~w/. //UVI IZJ. 27 l-283.
WILLIS J. S. (1966) Characteristics of Ion transport in Kid- ney cortex of mammalian hibernators. _I. q~‘” I’/II,\~cI/ 49. 122l-~1239.
WILLIS J. S.. GOLDMAN S. S. & Fosrt K R. F. ( 197 I I Ttsw K concentration in rektion to the role of the !adnc! in hibernation and the cause of periodic arousal. CII~II/J Bioc~/Ir,,I. PII~~.SiOl. 39. 437 445.
ZIMNV M. L. & GKI.C;OKY R. (1958) High cnrrgk phos- phates during hibernation and arousal 111 the ground squirrel. Aw. J. Ph~sioi. 195. 233 2%.





























