Embed Size (px)
Citation preview

http://informahealthcare.com/txmISSN: 1537-6516 (print), 1537-6524 (electronic)
Toxicol Mech Methods, Early Online: 1–8! 2014 Informa Healthcare USA, Inc. DOI: 10.3109/15376516.2014.945108
ORIGINAL ARTICLE
Hesperidin restores experimentally induced neurotoxicity in Wistar rats
Mehar Naseem and Suhel Parvez
Department of Medical Elementology and Toxicology, Jamia Hamdard (Hamdard University), New Delhi, India
Abstract
Carbon tetrachloride (CCl4) is a highly toxic industrial solvent with pronounced systemic toxicityincluding brain. Neurotoxicity may be a direct result of hepatic dysfunction from CCl4intoxication. Over the years CCl4 has been used as an excellent model for studyingexperimentally induced neurotoxicity in murine models. Hesperidin (HP) is a knowncytoprotectant with comprehensive anti-oxidant and neuroprotective properties. The aim ofthe present study was to evaluate experimentally induced neurotoxicity by CCl4 and itsabrogation by using antioxidant potential of HP. CCl4 caused a significant enhancement in thelipid peroxidation (LPO) levels and protein carbonyl (PC) content. HP supplementationsignificantly restored the LPO levels and PC content. It also replenished the altered enzymaticand non-enzymatic antioxidants in brain tissues of rats. The neurotoxicity markers were alsorestored to normalcy with HP treatment. It is suggested that HP, by attenuating neuronaloxidative stress, holds promise that can ameliorate CCl4-induced neurotoxicity. HP has thepotential to be explored as a universal neuroprotectant in xenobiotically induced neurotoxicitymediated by oxidative stress.
Keywords
Biomarkers, brain, carbon tetrachloride,natural compound, oxidative stress,rodents
History
Received 14 April 2014Revised 15 June 2014Accepted 11 July 2014Published online 30 July 2014
Introduction
A large increase in reactive oxygen species (ROS) which are
produced during various cellular metabolism leads to several
damaging effects like tissue damage, DNA damage, oxidative
stress, lipid peroxidation (LPO), and cell death (Saquib et al.,
2012). ROS is implicated in pathogenesis of a number of
neurodegenerative diseases in brain, cognitive dysfunction,
cancer, and a variety of other conditions (Kovacic &
Somanathan, 2012). Brain also contains high level of
ascorbate and iron, which may be essential during brain
development involved in the production of ROS. Numerous
studies have indicated that oxidative stress is responsible for
the disruption of blood brain barrier (BBB) integrity
associated with neurodegenerative disorders as it is prone to
ROS (Freeman & Keller, 2012). Brain may deal in a different
way with oxidative stress as compared with liver or kidney
(Seminotti et al., 2012).
Carbon tetrachloride (CCl4) is a well-known hepatotoxic
agent which has also been implicated in inhibiting mitochon-
drial respiratory chain in the brain (Boer et al., 2009). Natural
compounds have been used for various purposes in medicines,
cosmetics, and also as dietary supplements (Albarracin et al.,
2012). Flavonoids are compounds, which are antioxidants
from natural origin and are present in a number of
natural sources (Kamaraj et al., 2009). Hesperidin
(HP, 3,5,7-trihydroxy flavanone-7-rhamnoglucoside) is a
biologically and pharmacologically active citrus bioflavonoid
by-product of citrus cultivation, abundantly found in sweet
orange and lemon (Gaur et al., 2011). It has been previously
reported that HP is having free radical scavenging as well as
anti-LPO properties and has been shown to be powerful
against ROS and it has also been assayed for its neuropro-
tective efficacy in diabetic rat model (Ibrahim, 2008).
It has attracted particular interest due to its several health
beneficial effects like anti-inflammatory, anticarcinogenic,
antimicrobial, and immunomodulatory effects (Nones et al.,
2010).
HP has been shown to be protective against various
neurobehavioral alterations caused (Viswanatha et al., 2012).
It has been well documented that HP is a potential
neuroprotective antioxidant, it may because of being lipo-
philic in nature, crosses the BBB easily (Salem et al., 2012).
HP deals with neurotoxicity due to its antioxidative and anti-
inflammatory properties and also protects the dopamine
depletion which enhances it neuroprotective efficiency
(Tamilselvam et al., 2013). However, protective role of HP
against CCl4 induced neurotoxicity has not been investigated
in a rodent model. The aim of the present study was to explore
modulatory effects of HP against CCl4-induced neurotoxicity
in Wistar rats.
Address for correspondence: Dr. Suhel Parvez, Department of MedicalElementology and Toxicology, Jamia Hamdard (Hamdard University),New Delhi 110062, India. Tel: +91 11 26059688x5573. Fax:+91 11 26059663. E-mail: [email protected], [email protected]
Tox
icol
ogy
Mec
hani
sms
and
Met
hods
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Mem
oria
l Uni
vers
ity o
f N
ewfo
undl
and
on 0
8/03
/14
For
pers
onal
use
onl
y.

Materials and methods
Chemicals
Sulfosalicylic acid and thiobarbituric acid (TBA) were
obtained from Hi-Media Labs (Mumbai, India). Butylated
hydroxytoluene (BHT), o-phosphoric acid (OPA), trichloro-
acetic acid (TCA), 1-Amino-2-naphthol-4-sulfonic acid
(ANSA), 2,4-dinitrophenylhydrazine (DNPH), and perchloric
acid (PCA) were purchased from Merck Limited (Mumbai,
India). Bovine serum albumin, reduced glutathione (GSH),
oxidized glutathione (GSSG), and benzylamine hydrochloride
(BAHC) were purchased from Sigma Chemicals Co.
(St. Louis, MO). Reduced nicotinamide adenine dinucleotide
phosphate (NADPH), 1-chloro-2,4-dinitrobenzene (CDNB),
5,5-dithiobis-2-nitrobenzoic acid (DTNB), ethylenediamine-
tetraacetic acid (EDTA), and acetylthiocholine iodide (ATC)
were obtained from SRL (Mumbai, India). Other routine
chemicals were obtained from Hi-Media Labs (Mumbai,
India), Merck Limited (Mumbai, India), and SRL
(Mumbai, India). CCl4 and HP were obtained from Merck
Limited (Mumbai, India) and SRL Labs (Mumbai, India),
respectively.
Animals
Adult male Wistar rats weighing 250–300 g were used for the
study. The experimental rats were acclimatized for a week
prior to the commencement of the dose regime. The animals
were housed under standard laboratory conditions where
temperature was maintained at 22 ±2 �C with a relative
humidity of 65 ± 10% and a photoperiod of 12 h light/dark
cycle. All the experiments were carried out according to the
standard guidelines of Institutional Animal Ethics Committee
(IAEC) of Jamia Hamdard (Hamdard University).
Experimental design
The animals were randomly divided into four groups, and
each group consisted of seven animals. The description about
the different groups is as follows:
Group 1: Control group. The animals were treated with a
vehicle for HP (i.e. normal saline) by oral route for eight
consecutive days and a single subcutaneous (s.c.) injection of
olive oil on the 7th day.
Group 2: HP group. In this group, rats received HP (200 mg/
kg b.w., oral) treatment for a period of 8 d.
Group 3: CCl4 group. The animals were administered
normal saline for 8 d and were exposed to CCl4(2 ml/kg b.w., s.c., 40% v/v in olive oil) on the 7th day of
the treatment.
Group 4: HP (200 mg/kg b.w., oral) + CCl4 (2 ml/kg
b.w., s.c.). Rats were given HP treatment for 8 d and a
single injection of CCl4 on the 7th day along with HP.
The dose of CCl4 and HP used in our study was based on
previously published research reports (El-Sayed et al., 2008;
Jayakumar et al., 2008). At the end of the experimental period
of 8 d, the animals were anesthetized and sacrificed by
decapitation. The brain was immediately dissected, and
washed in chilled phosphate buffer to remove blood. The
effect of CCl4 and role of HP on the oxidative stress
biomarkers were studied in the brain tissue.
Sample preparation
Post-mitochondrial supernatant (PMS) and homogenate
preparation
The brain of the animals was promptly excised and washed
with chilled phosphate-buffered saline for the removal of
blood. The brain was homogenized 1:10 w/v in 0.1 M sodium
phosphate buffer (pH 7.4) with a Potter–Elvehjem homogen-
izer. The homogenates obtained were used to estimate brain
TBARS. The homogenates were centrifuged at 10 500 g for
20 min at 4 �C to get the PMS, using a REMI C-24 centrifuge
(Remi Sales and Engineering, Ltd., Mumbai, India). The
PMS supernatant obtained was used for the analysis of
oxidative stress biomarkers, non-enzymatic antioxidants,
enzymatic antioxidants, and neurotoxicity biomarkers.
Biochemical estimation
Oxidative stress biomarkers
Determination of LPO. The extent of LPO was measured
using the procedure of Tabassum et al. (2007). The concen-
tration of thiobarbituric acid reactive substances (TBARS)
was determined using an extinction coefficient of
1.56� 105 M�1 cm�1 and results were expressed as nmol
TBARS formed/h/mg protein.
Determination of protein oxidation. Protein oxidation in brain
PMS was measured as a concentration of protein carbonyls
(PC) formed. PC level was determined using DNPH assay
according to the procedure of Chaudhary & Parvez (2012).
The results were expressed as nmol DNPH incorporated/mg
protein based on the molar extinction coefficient of
2.1� 104 M�1 cm�1.
Non-enzymatic antioxidants
Estimation of non-protein thiol (NP-SH). The NP-SH was
measured according to the method described by Govil et al.
(2012). The molar extinction coefficient of 13 100M�1 cm�1 at
412 nm was used for the determination of the NP-SH content.
The values were expressed as nmol of the NP-SH/mg protein.
Estimation of reduced glutathione (GSH). The GSH content
was estimated according to the method of Chaudhary &
Parvez (2012). The GSH concentration was calculated as
nmol GSH/mg protein using a molar extinction coefficient of
1.36� 104 M�1 cm�1.
Enzymatic antioxidants
Activity of glutathione peroxidase (GPx). The activity of GPx
was assayed according to the method of Haque et al. (2003).
The assay mixture consisted of sodium phosphate
buffer (0.1 M, pH 7.4), EDTA (1 mM), sodium azide (1 mM),
GSH (1 mM), NADPH (0.02 mM), H2O2 (0.25 mM), and 0.1 ml
of PMS of brain in a total volume of 2 ml. Oxidation of NADPH
was recorded kinetically at 340 nm. The enzyme activity
was calculated as nmol NADPH oxidized/min/mg protein,
using a molar extinction coefficient of 6.22� 103 M�1 cm�1.
Activity of glutathione reductase (GR). The activity of GR was
assayed by the method of Haque et al. (2003). The assay
2 M. Naseem & S. Parvez Toxicol Mech Methods, Early Online: 1–8
Tox
icol
ogy
Mec
hani
sms
and
Met
hods
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Mem
oria
l Uni
vers
ity o
f N
ewfo
undl
and
on 0
8/03
/14
For
pers
onal
use
onl
y.
system consisted 0.1 M phosphate buffer (pH 7.4), 0.5 mM
EDTA, 1 mM GSSG, 0.1 mM NADPH, and 0.3 ml super-
natant of brain tissue in a total volume of 2 ml. The enzyme
activity was quantitated at 25 �C by measuring the disappear-
ance of NADPH at 340 nm, and was calculated as nmol
NADPH oxidized/min/mg protein using a molar extinction
coefficient of 6.223� 103 M�1 cm�1.
Activity of glutathione-s-transferase (GST). The activity of
GST was assayed using the method of Haque et al. (2003) by
the catalytic reaction of CDNB with the sulfhydryl group of
glutathione and was measured kinetically at 340 nm. The
assay system consisted of 0.1 M phosphate buffer (pH 7.4),
10 mM GSH, 10 mM CDNB, and 0.1 ml of PMS of rat brain.
The enzymatic activity was calculated as nmol CDNB
conjugate formed/min/mg protein using a molar coefficient
of 9.6� 103 M�1 cm�1.
Neurotoxicity biomarkers
Activity of monoamine oxidase (MAO). The activity of MAO
was measured by using the method of Chaudhary & Parvez
(2012). The enzyme activity was calculated as mmol BAHC
hydrolyzed/min/mg protein using a molar extinction coeffi-
cient of 7.6925 M�1 cm�1.
Activity of acetylcholinesterase (AChE). The activity of AChE
was estimated by using the method of Chaudhary & Parvez
(2012). The enzyme activity was calculated as nmol ATC
hydrolyzed/min/mg protein using a molar extinction coeffi-
cient of 1.36� 104 M�1 cm�1.
Activity of total ATPase. The activity of total ATPase was
measured as the release of inorganic phosphate (Pi) by the
method of Chaudhary & Parvez (2012). The activity was
measured as mg Pi liberated/min/mg protein.
Determination of protein
Estimation of protein contents in biological fractions of brain
was done using bovine serum albumin as standard by the
method of Lowry et al. (1951).
Statistical analysis
The results obtained were expressed as mean ± standard error
(SE). All data were analyzed using analysis of variance
(ANOVA) followed by Tukey’s test. Values of p50.05 were
considered as significant. All the statistical analyses were
performed using graph pad prism 5 software (Graph Pad
Software, Inc., San Diego, CA).
Results
No mortality and significant alteration in the body weight of
different groups of subject/animals were documented during
and after the treatment (data not shown).
Oxidative stress biomarkers
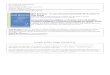
Effect on LPO
Treatment of animals with CCl4 (2 ml/kg b.w.) showed a
significant (p50.001) elevation in the LPO content of brain
homogenate when compared with the control group
(Figure 1A). HP (200 mg/kg, oral) pre-treatment revealed a
significant (p50.01) depletion in the LPO content of brain
homogenate as compared with the CCl4-treated group. HP
alone did not show any significant change in the LPO content
of animals beyond control values.
Effect on protein oxidation (PC)
Exposure to CCl4 resulted in a significant (p50.001)
enhancement in the activity of protein oxidation in the brain
PMS of CCl4-treated animals as compared with the control
group (Figure 1B). In HP pre-treated group, there was a
significant (p50.01) depletion in the PC content of brain
PMS. However, no significant effect in the PC content of HP
treated group was observed when compared with control.
Non-enzymatic antioxidant status
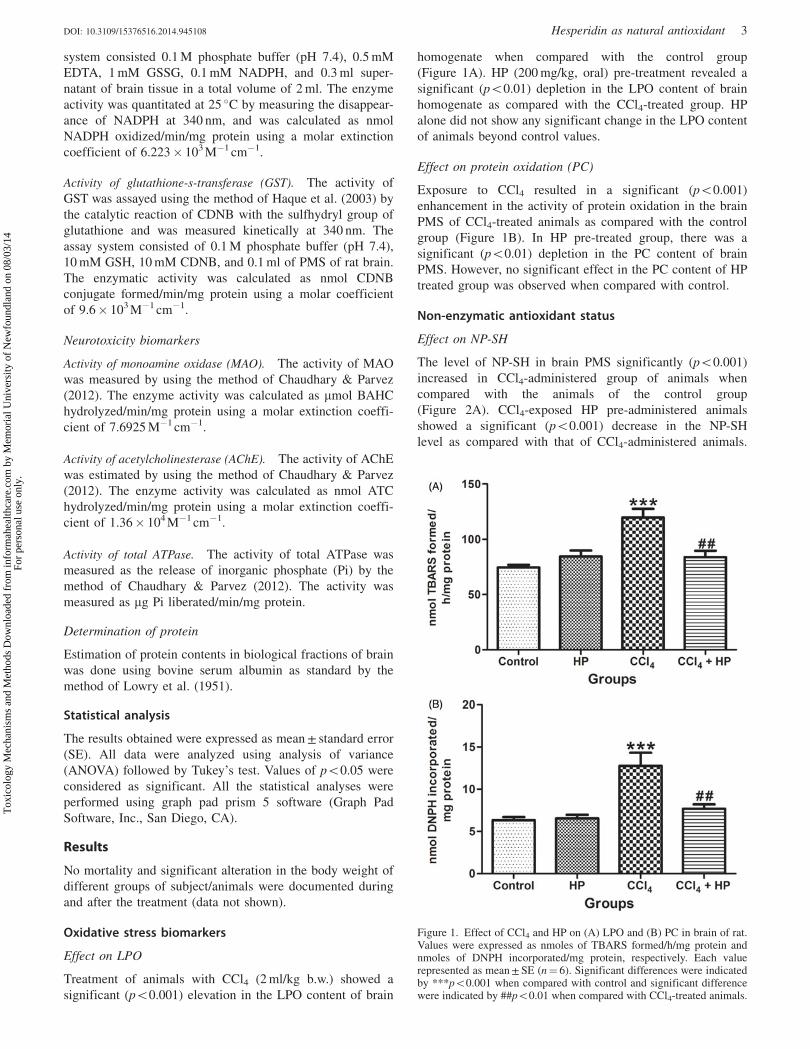
Effect on NP-SH
The level of NP-SH in brain PMS significantly (p50.001)
increased in CCl4-administered group of animals when
compared with the animals of the control group
(Figure 2A). CCl4-exposed HP pre-administered animals
showed a significant (p50.001) decrease in the NP-SH
level as compared with that of CCl4-administered animals.
Figure 1. Effect of CCl4 and HP on (A) LPO and (B) PC in brain of rat.Values were expressed as nmoles of TBARS formed/h/mg protein andnmoles of DNPH incorporated/mg protein, respectively. Each valuerepresented as mean ± SE (n¼ 6). Significant differences were indicatedby ***p50.001 when compared with control and significant differencewere indicated by ##p50.01 when compared with CCl4-treated animals.
DOI: 10.3109/15376516.2014.945108 Hesperidin as natural antioxidant 3
Tox
icol
ogy
Mec
hani
sms
and
Met
hods
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Mem
oria
l Uni
vers
ity o
f N
ewfo
undl
and
on 0
8/03
/14
For
pers
onal
use
onl
y.
HP-treated animals showed no any significant alteration in the
NP-SH level in brain PMS when compared with control group
of animals.
Effect on GSH
Significant (p50.001) higher level of the GSH content was
observed in brain PMS of CCl4-exposed animals when
compared with the control group (Figure 2B). HP pre-
treatment prior to CCl4 injection significantly (p50.001)
declines the GSH content and prevented these elevations.
Animals treated with HP did not show any significant change
when compared with the control group.
Enzymatic antioxidants activities
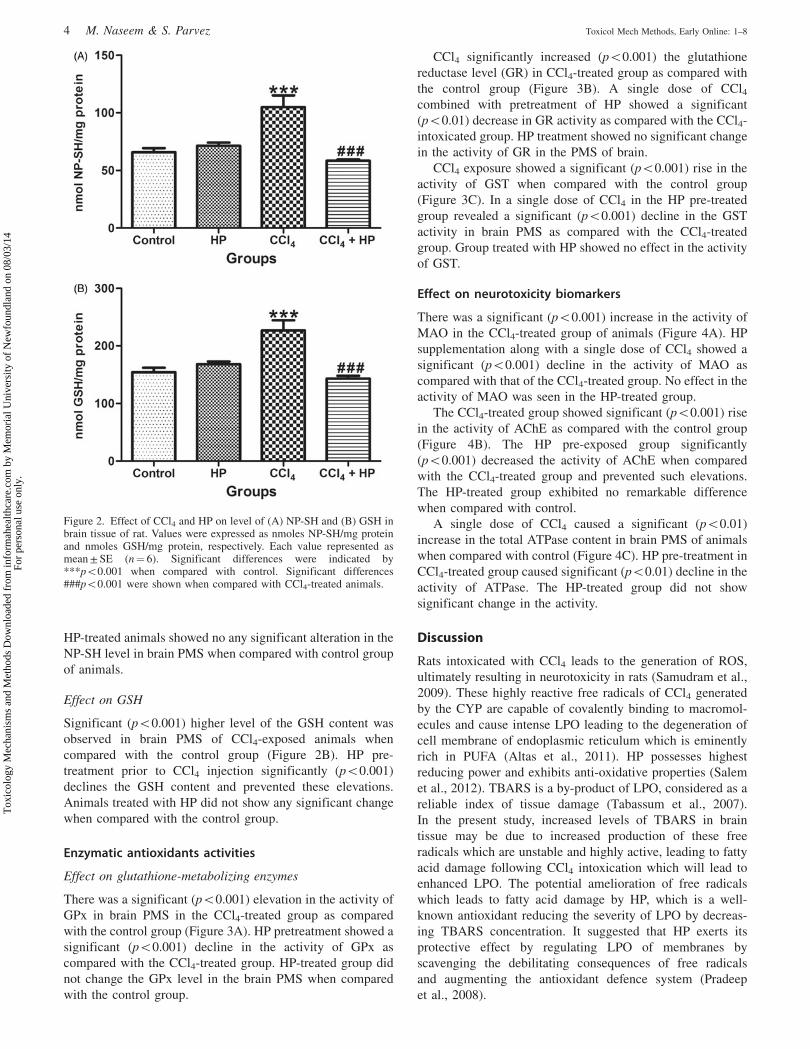
Effect on glutathione-metabolizing enzymes
There was a significant (p50.001) elevation in the activity of
GPx in brain PMS in the CCl4-treated group as compared
with the control group (Figure 3A). HP pretreatment showed a
significant (p50.001) decline in the activity of GPx as
compared with the CCl4-treated group. HP-treated group did
not change the GPx level in the brain PMS when compared
with the control group.
CCl4 significantly increased (p50.001) the glutathione
reductase level (GR) in CCl4-treated group as compared with
the control group (Figure 3B). A single dose of CCl4combined with pretreatment of HP showed a significant
(p50.01) decrease in GR activity as compared with the CCl4-
intoxicated group. HP treatment showed no significant change
in the activity of GR in the PMS of brain.
CCl4 exposure showed a significant (p50.001) rise in the
activity of GST when compared with the control group
(Figure 3C). In a single dose of CCl4 in the HP pre-treated
group revealed a significant (p50.001) decline in the GST
activity in brain PMS as compared with the CCl4-treated
group. Group treated with HP showed no effect in the activity
of GST.
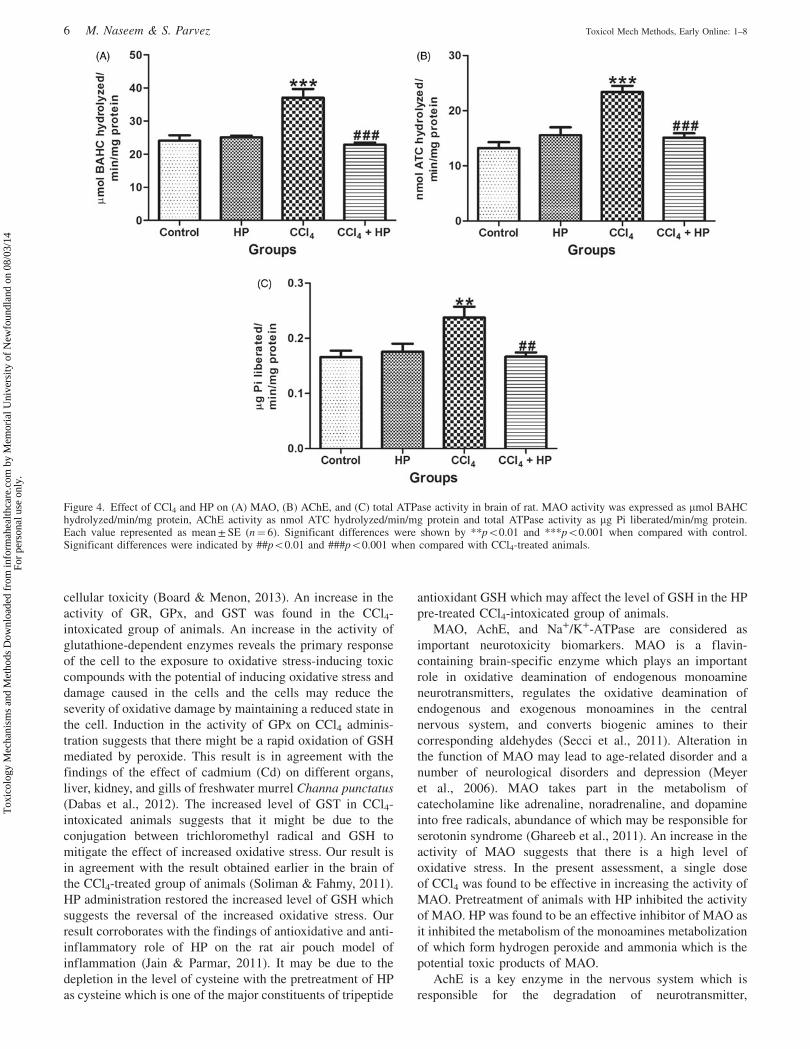
Effect on neurotoxicity biomarkers
There was a significant (p50.001) increase in the activity of
MAO in the CCl4-treated group of animals (Figure 4A). HP
supplementation along with a single dose of CCl4 showed a
significant (p50.001) decline in the activity of MAO as
compared with that of the CCl4-treated group. No effect in the
activity of MAO was seen in the HP-treated group.
The CCl4-treated group showed significant (p50.001) rise
in the activity of AChE as compared with the control group
(Figure 4B). The HP pre-exposed group significantly
(p50.001) decreased the activity of AChE when compared
with the CCl4-treated group and prevented such elevations.
The HP-treated group exhibited no remarkable difference
when compared with control.
A single dose of CCl4 caused a significant (p50.01)
increase in the total ATPase content in brain PMS of animals
when compared with control (Figure 4C). HP pre-treatment in
CCl4-treated group caused significant (p50.01) decline in the
activity of ATPase. The HP-treated group did not show
significant change in the activity.
Discussion
Rats intoxicated with CCl4 leads to the generation of ROS,
ultimately resulting in neurotoxicity in rats (Samudram et al.,
2009). These highly reactive free radicals of CCl4 generated
by the CYP are capable of covalently binding to macromol-
ecules and cause intense LPO leading to the degeneration of
cell membrane of endoplasmic reticulum which is eminently
rich in PUFA (Altas et al., 2011). HP possesses highest
reducing power and exhibits anti-oxidative properties (Salem
et al., 2012). TBARS is a by-product of LPO, considered as a
reliable index of tissue damage (Tabassum et al., 2007).
In the present study, increased levels of TBARS in brain
tissue may be due to increased production of these free
radicals which are unstable and highly active, leading to fatty
acid damage following CCl4 intoxication which will lead to
enhanced LPO. The potential amelioration of free radicals
which leads to fatty acid damage by HP, which is a well-
known antioxidant reducing the severity of LPO by decreas-
ing TBARS concentration. It suggested that HP exerts its
protective effect by regulating LPO of membranes by
scavenging the debilitating consequences of free radicals
and augmenting the antioxidant defence system (Pradeep
et al., 2008).
Figure 2. Effect of CCl4 and HP on level of (A) NP-SH and (B) GSH inbrain tissue of rat. Values were expressed as nmoles NP-SH/mg proteinand nmoles GSH/mg protein, respectively. Each value represented asmean ± SE (n¼ 6). Significant differences were indicated by***p50.001 when compared with control. Significant differences###p50.001 were shown when compared with CCl4-treated animals.
4 M. Naseem & S. Parvez Toxicol Mech Methods, Early Online: 1–8
Tox
icol
ogy
Mec
hani
sms
and
Met
hods
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Mem
oria
l Uni
vers
ity o
f N
ewfo
undl
and
on 0
8/03
/14
For
pers
onal
use
onl
y.
PC is considered as an excellent biomarker of oxidative
protein damage as proteins are susceptible to oxidative
modification (Dalle-Donne et al., 2003). Oxidative modifica-
tion of proteins changes its configuration via aggregation,
fragmentation, and formation of cross linkages in the
polypeptide chain and also leads to secondary impairment
of biomolecules with loss of protein biological function, leads
to the generation of superoxide radical (Chen et al., 2005).
Addition of carbonyl group in proteins is directly proportional
to the protein subjected to oxidative damage and an increase
in carbonyl content is associated with various pathological
disorders (Almroth et al., 2005). A significant increase in
protein oxidation was seen in the brain of animals with the
single-dose administration of CCl4 as compared with the
control group. It might be due to increased oxidatively
modified protein in brain with the administration of CCl4. Our
result is in agreement with the findings of previous result
performed with liver (Srivastava & Shivanandappa, 2010).
Interestingly, pre-administration of HP attenuated the effect of
CCl4-induced protein damage, indicating that HP has a
protective action against CCl4-induced neurotoxicity. Our
results are in corroboration with the recent finding where HP
has been shown to be a good natural antioxidant against
cisplatin-induced oxidative damage (Sahu et al., 2013). HP
has been found to reduce the PC content in our study thereby
reducing oxidative stress and oxidative damage of protein by
scavenging the free radicals causing modification in the
protein structure.
Dietary antioxidant supplementation such as bioflavonoids
mainly protects cells and organs against ROS-induced oxida-
tive damage (Tsai et al., 2013). GSH scavenges free radicals
and provides a defence system against ROS as well as
detoxifies various xenobiotic compounds, H2O2, lipid hydro-
peroxides, and electrophilic compounds (Manna et al., 2011).
It plays crucial role in defence mechanism and also acts as a
co-enzyme for numerous enzymes involved in the cellular
defence system (Masella et al., 2005). GPx constitutes a
family of enzyme which reduces H2O2 and lipid hydroper-
oxides to the corresponding stable hydroxy compounds and
H2O utilizing GSH as a reducing substrate (Dickinson &
Forman, 2002). GR is a flavoenzyme and is represented by a
single copy gene in humans which takes part in the reduction
of GSSG (oxidized glutathione) to GSH with the help of
NADPH (Li et al., 2010). The activity of GR is very
important to maintain the level of GSH/GSSG ratio. GST is a
well-known phase-2 enzyme which plays a very crucial role in
the detoxification process of electrophilic compounds, favor-
ing the elimination from the body of the organ by conjugation
with glutathione (Afzal et al., 2013). GSH oxidizes to GSSG
by peroxide and GPx. GSH is restored from GSSG by the
enzyme GR using NADPH as an electron donor and the ratio
of GSH/GSSG within a cell is considered as the measure of
Figure 3. Effect of CCl4 and HP on (A) GPx, (B) GR and (C) GST activity in brain of rat. GPx and GR activities were expressed as nmoles of NADPHoxidized/min/mg protein whereas GST activity was expressed as nmoles of CDNB conjugate formed/min/mg protein. Each value represented asmean ± SE (n¼ 6). Significant differences were shown by ***p50.001 when compared with control. Significant differences were indicated by##p50.01 and ###p50.001 when compared with CCl4-treated animals.
DOI: 10.3109/15376516.2014.945108 Hesperidin as natural antioxidant 5
Tox
icol
ogy
Mec
hani
sms
and
Met
hods
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Mem
oria
l Uni
vers
ity o
f N
ewfo
undl
and
on 0
8/03
/14
For
pers
onal
use
onl
y.
cellular toxicity (Board & Menon, 2013). An increase in the
activity of GR, GPx, and GST was found in the CCl4-
intoxicated group of animals. An increase in the activity of
glutathione-dependent enzymes reveals the primary response
of the cell to the exposure to oxidative stress-inducing toxic
compounds with the potential of inducing oxidative stress and
damage caused in the cells and the cells may reduce the
severity of oxidative damage by maintaining a reduced state in
the cell. Induction in the activity of GPx on CCl4 adminis-
tration suggests that there might be a rapid oxidation of GSH
mediated by peroxide. This result is in agreement with the
findings of the effect of cadmium (Cd) on different organs,
liver, kidney, and gills of freshwater murrel Channa punctatus
(Dabas et al., 2012). The increased level of GST in CCl4-
intoxicated animals suggests that it might be due to the
conjugation between trichloromethyl radical and GSH to
mitigate the effect of increased oxidative stress. Our result is
in agreement with the result obtained earlier in the brain of
the CCl4-treated group of animals (Soliman & Fahmy, 2011).
HP administration restored the increased level of GSH which
suggests the reversal of the increased oxidative stress. Our
result corroborates with the findings of antioxidative and anti-
inflammatory role of HP on the rat air pouch model of
inflammation (Jain & Parmar, 2011). It may be due to the
depletion in the level of cysteine with the pretreatment of HP
as cysteine which is one of the major constituents of tripeptide
antioxidant GSH which may affect the level of GSH in the HP
pre-treated CCl4-intoxicated group of animals.
MAO, AchE, and Na+/K+-ATPase are considered as
important neurotoxicity biomarkers. MAO is a flavin-
containing brain-specific enzyme which plays an important
role in oxidative deamination of endogenous monoamine
neurotransmitters, regulates the oxidative deamination of
endogenous and exogenous monoamines in the central
nervous system, and converts biogenic amines to their
corresponding aldehydes (Secci et al., 2011). Alteration in
the function of MAO may lead to age-related disorder and a
number of neurological disorders and depression (Meyer
et al., 2006). MAO takes part in the metabolism of
catecholamine like adrenaline, noradrenaline, and dopamine
into free radicals, abundance of which may be responsible for
serotonin syndrome (Ghareeb et al., 2011). An increase in the
activity of MAO suggests that there is a high level of
oxidative stress. In the present assessment, a single dose
of CCl4 was found to be effective in increasing the activity of
MAO. Pretreatment of animals with HP inhibited the activity
of MAO. HP was found to be an effective inhibitor of MAO as
it inhibited the metabolism of the monoamines metabolization
of which form hydrogen peroxide and ammonia which is the
potential toxic products of MAO.
AchE is a key enzyme in the nervous system which is
responsible for the degradation of neurotransmitter,
Figure 4. Effect of CCl4 and HP on (A) MAO, (B) AChE, and (C) total ATPase activity in brain of rat. MAO activity was expressed as mmol BAHChydrolyzed/min/mg protein, AChE activity as nmol ATC hydrolyzed/min/mg protein and total ATPase activity as mg Pi liberated/min/mg protein.Each value represented as mean ± SE (n¼ 6). Significant differences were shown by **p50.01 and ***p50.001 when compared with control.Significant differences were indicated by ##p50.01 and ###p50.001 when compared with CCl4-treated animals.
6 M. Naseem & S. Parvez Toxicol Mech Methods, Early Online: 1–8
Tox
icol
ogy
Mec
hani
sms
and
Met
hods
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Mem
oria
l Uni
vers
ity o
f N
ewfo
undl
and
on 0
8/03
/14
For
pers
onal
use
onl
y.

acetylcholine (Ach), during neurotransmission which is
necessary for learning and memory deficits (Parent &
Baxter, 2004) and also plays a very important role in the
cholinergic activity (Antoniades et al., 2002). Our result is in
accordance with a recent finding in which an increase in the
activity of AchE in the region of brain has been reported with
streptozotocin treatment (Biasibetti et al., 2013). An increase
in the activity of AchE in the brain is responsible for the low
availability of acetylcholine and, therefore, it may lead to the
alteration in cholinergic transmission (Tiwari et al., 2009). HP
pre-treatment was able to reverse the altered activity of AchE
which deals with the cognitive defect by increasing cholin-
ergic neurotransmission as reported in a report with a natural
antioxidant curcumin (Agrawal et al., 2010).
The sodium potassium adenosine triphosphate (Na+/K+-
ATPase) is an enzyme which maintains neuronal excitability
and plays a major role in the export and import of three Na+
ions out and two K+ ions in of the cell by utilizing ATP across
the plasma membrane against their potential gradient in the
nervous system and Na+/K+-ATPase has also been reported as
a signal transducer (Lingrel, 2010). The Na+/K+-ATPase is an
integral membrane heterodimer belongs to a superfamily of
proteins collectively designated as P-type ATPase family
(Jaitovich & Bertorello, 2006). In the present study, a
remarkable elevation is shown in the activity of Na+/K+-
ATPase in the CCl4-intoxicated group of rats. HP pre-
treatment suppressed the elevated level of Na+/K+-ATPase
activity in brain due to its neuroprotective nature. Our result is
in agreement with the previous result reported with some
natural compounds, green tea and bamboo leaves on Na efflux
channel in ovariectomized rats (Ryou et al., 2012). Further
studies are required for deciphering cellular and molecular
mechanisms of experimentally induced neurotoxicity and the
role of HP in its attenuation.
Declaration of interest
The authors declare that they have no conflict of interest.
University Grants Commission (UGC), Government of India,
is gratefully acknowledged for providing funding under
Faculty Research Award to S.P. M.N. was supported by a
Junior Research fellowship from UGC – Maulana Azad
National Fellowship program.
References
Afzal M, Khan R, Kazmi I, Anwar F. (2013). Hepatoprotective potentialof new steroid against carbon tetrachloride-induced hepatic injury.Mol Cell Biochem 378:275–81.
Agrawal R, Mishra B, Tyagi E, et al. (2010). Effect of curcumin on braininsulin receptors and memory functions in STZ (ICV) induceddementia model of rat. Pharmacol Res 61:247–52.
Albarracin SL, Stab B, Casas Z, et al. (2012). Effects of naturalantioxidants in neurodegenerative disease. Nutr Neurosci 15:1–9.
Almroth BC, Sturve J, Berglund A, Forlin L. (2005). Oxidative damagein eelpout (Zoarces viviparus), measured as protein carbonyls andTBARS, as biomarkers. Aquat Toxicol 73:171–80.
Altas S, Kızıl G, Kızıl M, et al. (2011). Protective effect of Diyarbakırwatermelon juice on carbon tetrachloride-induced toxicity in rats.Food Chem Toxicol 49:2433–8.
Antoniades C, Carageorgiou H, Tsakiris S. (2002). Effects of(�)deprenyl (selegiline) on acetylcholinesterase and Na+K+-ATPaseactivities in adult rat whole brain. Pharmacol Res 46:165–9.
Biasibetti R, Tramontina AC, Costa AP, et al. (2013). Green tea (�)epigallocatechin-3-gallate reverses oxidative stress and reduces
acetylcholinesterase activity in a streptozotocin-induced model ofdementia. Behav Brain Res 236:186–93.
Board PG, Menon D. (2013). Glutathione transferases, regulators ofcellular metabolism and physiology. Biochim Biophys Acta 1830:3267–88.
Boer LA, Panatto JP, Fagundes DA, et al. (2009). Inhibition ofmitochondrial respiratory chain in the brain of rats after hepaticfailure induced by carbon tetrachloride is reversed by antioxidants.Brain Res Bull 80:75–8.
Chaudhary S, Parvez S. (2012). An in vitro approach to assess theneurotoxicity of valproic acid-induced oxidative stress in cerebellumand cerebral cortex of young rats. Neuroscience 225:258–68.
Chen YR, Chen CL, Zhang L, et al. (2005). Superoxide generation frommitochondrial NADH dehydrogenase induces self-inactivation withspecific protein radical formation. J Biol Chem 280:37339–48.
Dabas A, Nagpure NS, Kumar R, et al. (2012). Assessment of tissue-specific effect of cadmium on antioxidant defense system and lipidperoxidation in freshwater murrel, Channa punctatus. Fish PhysiolBiochem 38:469–82.
Dalle-Donne I, Giustarini D, Colombo R, et al. (2003). Proteincarbonylation in human diseases. Trends Mol Med 9:169–76.
Dickinson DA, Forman HJ. (2002). Cellular glutathione and thiolsmetabolism. Biochem Pharmacol 64:1019–26.
El-Sayed el SM, Abo-Salem OM, Abd-Ellah MF, Abd-Alla GM. (2008).Hesperidin, an antioxidant flavonoid, prevents acrylonitrile-induced oxidative stress in rat brain. J Biochem Mol Toxicol 22:268–73.
Freeman LR, Keller JN. (2012). Oxidative stress and cerebral endothelialcells: regulation of the blood-brain-barrier and antioxidant basedinterventions. Biochim Biophys Acta 1822:822–9.
Gaur V, Aggarwal A, Kumar A. (2011). Possible nitric oxide mechanismin the protective effect of hesperidin against ischemic reperfusioncerebral injury in rats. Indian J Exp Biol 49:609–18.
Ghareeb DA, Hafez HS, Hussien HM, Kabapy NF. (2011). Non-alcoholic fatty liver induces insulin resistance and metabolic disorderswith development of brain damage and dysfunction. Metab Brain Dis26:253–67.
Govil N, Chaudhary S, Waseem M, Parvez S. (2012). Postnuclearsupernatant: an in vitro model for assessing cadmium-inducedneurotoxicity. Biol Trace Elem Res 146:402–9.
Haque R, Bin-Hafeez B, Parvez S, et al. (2003). Aqueous extract ofwalnut (Juglans regia L.) protects mice against cyclophosphamideinduced biochemical toxicity. Hum Exp Toxicol 22:473–80.
Ibrahim SS. (2008). Protective effect of hesperidin, a citrus bioflavonoid,on diabetes-induced brain damage in rats. J Appl Sci Res 4:84–95.
Jain M, Parmar HS. (2011). Evaluation of antioxidative and anti-inflammatory potential of hesperidin and naringin on the rat air pouchmodel of inflammation. Inflamm Res 60:483–91.
Jaitovich AA, Bertorello AM. (2006). Na+, K+-Atpase: an indispensableion pumping-signaling mechanism across mammalian cell mem-branes. Semin Nephrol 26:386–92.
Jayakumar T, Sakthivel M, Thomas PA, Geraldine P. (2008). Pleurotusostreatus, an oyster mushroom, decreases the oxidative stress inducedby carbon tetrachloride in rat kidneys, heart and brain. Chem BiolInteract 176:108–20.
Kamaraj S, Ramakrishnan G, Anandakumar P, et al. (2009). Antioxidantand anticancer efficacy of hesperidin in benzo(a)pyrene induced lungcarcinogenesis in mice. Invest New Drugs 27:214–22.
Kovacic P, Somanathan R. (2012). Redox processes in neurodegenerativedisease involving reactive oxygen species. Curr Neuropharmacol 10:289–302.
Li ZH, Zlabek V, Grabic R, et al. (2010). Modulation of glutathione-related antioxidant defense system of fish chronically treated by thefungicide propiconazole. Comp Biochem Physiol 152:392–8.
Lingrel JB. (2010). The physiological significance of the cardiotonicsteroid/ouabain-binding site of the Na+,K+-ATPase. Annu RevPhysiol 72:395–412.
Lowry OH, Rosebrough NJ, Farr AL, Randall RJ. (1951). Proteinmeasurement with the Folin phenol reagent. J Biol Chem 193:265–75.
Manna P, Bhattacharyya S, Das J, et al. (2011). Phytomedicinal role ofpithecellobium dulce against CCl4-mediated hepatic oxidative impair-ments and necrotic cell death. Evid Based Complement Alternat Med2011:832805.
Masella R, Di Benedetto R, Varı R, et al. (2005). Novel mechanisms ofnatural antioxidant compounds in biological systems: involvement of
DOI: 10.3109/15376516.2014.945108 Hesperidin as natural antioxidant 7
Tox
icol
ogy
Mec
hani
sms
and
Met
hods
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Mem
oria
l Uni
vers
ity o
f N
ewfo
undl
and
on 0
8/03
/14
For
pers
onal
use
onl
y.

glutathione and glutathione-related enzymes. J Nutr Biochem 16:577–86.
Meyer JH, Ginovart N, Boovariwala A, et al. (2006). Elevated monoamineoxidase a levels in the brain: an explanation for the monoamineimbalance of major depression. Arch Gen Psychiatry 63:1209–16.
Nones J, Stipursky J, Costa SL, Gomes FC. (2010). Flavonoids andastrocytes crosstalking: implications for brain development andpathology. Neurochem Res 35:955–66.
Parent MB, Baxter MG. (2004). Septohippocampal acetylcholine:involved in but not necessary for learning and memory? Learn Mem11:9–20.
Pradeep K, Park SH, Ko KC. (2008). Hesperidin a flavanoglyconeprotects against g-irradiation induced hepatocellular damage andoxidative stress in Sprague–Dawley rats. Eur J Pharmacol 587:273–80.
Ryou SH, Kang MS, Kim KI, et al. (2012). Effects of green tea or Sasaquelpaertensis bamboo leaves on plasma and liver lipids, erythrocyteNa efflux, and platelet aggregation in ovariectomized rats. Nutr ResPract 6:106–12.
Sahu BD, Kuncha M, Sindhura GJ, Sistla R. (2013). Hesperidinattenuates cisplatin-induced acute renal injury by decreasing oxidativestress, inflammation and DNA damage. Phytomedicine 20:453–60.
Salem HRA, El-Raouf Mohamed AA, Saleh EM, Shalaby KAF. (2012).Influence of Hesperidin combined with Sinemet on genetical andbiochemical abnormalities in rats suffering from Parkinson’s disease.Life Sci J 9:930–45.
Samudram P, Vasuki R, Rajeshwari H, et al. (2009). Antioxidant andantihepatotoxic activities of ethanolic crude extract of Meliaazedarach and Piper longum. J Med Plants Res 3:1078–83.
Saquib Q, Attia SM, Siddiqui MA, et al. (2012). Phorate-inducedoxidative stress, DNA damage and transcriptional activation of p53and caspase genes in male Wistar rats. Toxicol Appl Pharmacol 259:54–65.
Secci D, Bolasco A, Chimenti P, Carradori S. (2011). The state of the artof pyrazole derivatives as monoamine oxidase inhibitors andantidepressant/anticonvulsant agents. Curr Med Chem 18:5114–44.
Seminotti B, da Rosa MS, Fernandes CG, et al. (2012). Induction ofoxidative stress in brain of glutaryl-CoA dehydrogenase deficientmice by acute lysine administration. Mol Genet Metab 106:31–8.
Soliman AM, Fahmy SR. (2011). Protective and curative effects of the15 KD isolated protein from the Peganum harmala L. seeds againstcarbon tetrachloride induced oxidative stress in brain, tests anderythrocytes of rats. Eur Rev Med Pharmacol Sci 15:888–99.
Srivastava A, Shivanandappa T. (2010). Hepatoprotective effect of theroot extract of Decalepis hamiltonii against carbon tetrachloride-induced oxidative stress in rats. Food Chem 118:411–17.
Tabassum H, Parvez S, Rehman H, et al. (2007). Nephrotoxicity and itsprevention by taurine in tamoxifen induced oxidative stress in mice.Hum Exp Toxicol 26:509–18.
Tamilselvam K, Nataraj J, Janakiraman U, et al. (2013). Antioxidant andanti-inflammatory potential of hesperidin against 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-induced experimental Parkinson’s diseasein mice. Int J Nutr Pharmacol Neurol Dis 3:294–302.
Tiwari V, Kuhad A, Bishnoi M, Chopra K. (2009). Chronic treatmentwith tocotrienol, an isoform of vitamin E, prevents intracerebroven-tricular streptozotocin-induced cognitive impairment and oxidative–nitrosative stress in rats. Pharmacol Biochem Behav 93:183–9.
Tsai CF, Hsu YW, Ting HC, et al. (2013). The in vivo antioxidant andantifibrotic properties of green tea (Camellia sinensis, Theaceae).Food Chem 136:1337–44.
Viswanatha GL, Shylaja H, Sandeep Rao KS, et al. (2012). Hesperidinameliorates immobilization-stress-induced behavioral and biochem-ical alterations and mitochondrial dysfunction inmice bymodulatingnitrergic pathway. ISRN Pharmacol 2012:479570.
8 M. Naseem & S. Parvez Toxicol Mech Methods, Early Online: 1–8
Tox
icol
ogy
Mec
hani
sms
and
Met
hods
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Mem
oria
l Uni
vers
ity o
f N
ewfo
undl
and
on 0
8/03
/14
For
pers
onal
use
onl
y.










![Evaluation of Hesperidin [Flavonoid] as a pulp capping ... · EVALUATION OF HESPERIDIN [FLAVONOID] AS A DIRECT PULP CAPPING MATERIAL Presented by: Ebtesam Osama Abo El-Mal Under supervision](https://img.pdfslide.us/doc/110x75/5d5e16ff88c9936f1b8bc77c/evaluation-of-hesperidin-flavonoid-as-a-pulp-capping-evaluation-of-hesperidin.jpg)