Embed Size (px)
Citation preview

Helping is Costly to Young Birds in Cooperatively Breeding White-Winged ChoughsAuthor(s): Robert Heinsohn and Andrew CockburnSource: Proceedings: Biological Sciences, Vol. 256, No. 1347 (Jun. 22, 1994), pp. 293-298Published by: The Royal SocietyStable URL: http://www.jstor.org/stable/50175 .
Accessed: 07/05/2014 21:32
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
The Royal Society is collaborating with JSTOR to digitize, preserve and extend access to Proceedings:Biological Sciences.
http://www.jstor.org
This content downloaded from 169.229.32.136 on Wed, 7 May 2014 21:32:48 PMAll use subject to JSTOR Terms and Conditions

Helping is costly to young birds in cooperatively breeding white-winged choughs
ROBERT HEINSOHN AND ANDREW COCKBURN
Evolutionary Ecology Group, Division of Botany and Zoology, Australian National University, Canberra, ACT 0200, Australia
SUMMARY
Cooperative breeding among birds is at its most extreme in white-winged choughs (Corcorax melanorhamphos). Choughs have never been observed to breed successfully without helpers, and reproductive success increases linearly across all group sizes (maximum = 16). Further, helpers contribute to every aspect of reproduction, including nest building and incubation. Here we show that the contribution of young helpers (one year old and less) to incubation depends on the group in which they live. In small groups (3-5 birds), young helpers contribute as much to incubation as older birds, but in large groups they contribute little. In large groups, help increases sharply with age. Old birds contribute equally, regardless of group size. Although choughs generally do not lose body mass over incubation, young helpers lose mass in proportion to the amount of incubation they perform, independent of any effect of group size. This provides evidence that helpers in cooperatively breeding birds suffer costs from providing help additional to the costs incurred from remaining philopatric. It also demonstrates that the needs of the group influence whether young birds provide help.
1. INTRODUCTION
Cooperative breeding in birds is usually portrayed as the consequence of two decisions made by young birds (Brown 1987; Emlen 1991): (i) whether to defer dispersal and remain in the territory of the parents; and (ii) whether to provide help to their parents as they attempt to raise one or more subsequent broods. Attempts to answer the first question have focused on ecological factors which constrain dispersal, or on benefits of occupancy of the natal territory which outweigh any gains to be made from moving elsewhere (Brown 1987; Stacey & Ligon 1987; Emlen 1991). By contrast, the question of whether philopatric young should provide help has attracted much speculation but little empirical evidence. Emlen et al. (1991) enumerate may benefits of helping, and these are usually set against the cost of philopatry, the loss of reproduction incurred by not moving elsewhere. Variation between individuals in the extent of help is interpreted in the light of these benefits. For example, helping may be more valuable to a helper closely related to the birds it helps (Emlen & Wrege 1988). Costs directly resulting from the act of helping have attracted little attention. This is surprising as helping is often portrayed as altruistic, which by definition involves some cost. There is some evidence supporting a cost of helping in fish (reduced growth in Lamprologus richardi (Taborsky 1984) and mammals (mass loss in Marmota marmota (Arnold 1990)). However, in the birds, which have dominated the study of cooperative breeding, there is little relevant evidence. Eden's (1987) experimental demonstration that moorhen Gallinula chloropus were more likely to help when provisioned
with food certainly suggests that helping only occurs when costs are tolerable. A measurable cost of helping has only been suggested for pied kingfishers, Ceryle rudis (Reyer 1984). Helpers in this species are of two sorts: primary helpers, related to the dominants, and secondary helpers, unrelated to and recruited by the dominants. Primary helpers work as hard as dominants and have lower survival than other birds, whereas secondary helpers work less and have survival com- parable with dominants. Thus the relation between work rate and survival is still not clear.
Indeed, one recent line of argument suggests that helping is an inevitable consequence of a decision to stay on the parental territory (Jamieson & Craig 1987; Jamieson 1989, 1991). This exposes young birds to a stimulus (e.g. begging nestlings) which produces a response to feed or otherwise assist in all birds. If we assume that helping has no direct cost, there can be no selection against provisioning behaviour. This hypothesis can only be disentangled where birds show variation in the extent or presence of help.
Three recent lines of evidence suggest that the decision to help is also influenced by the behaviour of adult birds, rather than just decisions made by the helper. First, in pied kingfishers, recruitment of secondary helpers appears to depend on the energetic requirements of the dominants (Reyer & Westerterp 1985). Second, Emlen & Wrege (1992) have shown that dominant white-fronted bee-eaters (Merops bullock- oides) harass young helpers which attempt to breed independently, increasing the likelihood that they will abandon attempts at reproduction and instead remain at their parent's nest and help. Third, Mulder & Langmore (1993) have shown that temporary removal
Proc. R. Soc. Lond. B (1994) 256, 293-298 293 (C 1994 The Royal Society Printed in Great Britain
20-2
This content downloaded from 169.229.32.136 on Wed, 7 May 2014 21:32:48 PMAll use subject to JSTOR Terms and Conditions

294 R. Heinsohn and A. Cockburn A cost of helping behaviour in birds
of helper superb fairy-wrens (Malurus cyaneus) leads to their being attacked by the dominant male when they return to the territory. However, the intensity and duration of this attack is context dependent. The most severe escalation occurs when the group is feeding nestlings or fledglings, suggesting the attacks are 'punishment' for neglect of helping duties. These studies contain a series of implications. In pied kingfishers, there are circumstances where birds would contribute help but are only accepted by adults under certain contexts. By contrast, the data from bee-eaters suggest that there are circumstances where birds attempt to breed independently but are instead harassed, leading them to stay at home and help. Last, dominant fairy-wrens behave in a manner which enforces help among subordinate birds, suggesting that there are circumstances where those subordinates would decline to help in the absence of 'punishment'.
Among cooperatively breeding birds, group living and helping behaviour are most important in white- winged choughs. Apart from occasional cases of inter- group transfer (Heinsohn 1991 a), birds of both sexes live their lives in the groups in which they are born, so group members are likely to be close relatives. Birds are apparently incapable of reproduction until 4 years old (Rowley 1978). However, group members of all ages contribute to every aspect of reproduction, including both nest building and incubation. Reproduction is extremely sensitive to group size. In two independent studies, pairs of choughs were never successful at fledging offspring, and trios were only rarely successful (Rowley 1978; Heinsohn 1991 b). Choughs thus appear to be obligatorily cooperative (Heinsohn 1991 c, 1992). Further, reproductive success continues to increase linearly across all group sizes, from 3 to 16 (Heinsohn 1992).
In this paper we show that one form of help (incubation) is sensitive to group size. Clutches of this species are incubated constantly, with group members sharing the effort. First-year birds contribute little incubation in group sizes of six or more, yet contribute as much as older birds in smaller groups. By contrast, the contribution of old birds (5 years or more) is not affected by group size. Like the studies described above, this suggests that helping is not inevitable, and may in part be influenced by parental needs. Further, we also show that, although incubation does not affect the mass of older birds, young birds lose mass in proportion to the amount of incubation they perform. This suggests that, in addition to the needs of dominants, direct costs of helping behaviour could influence whether subordinates help or not. This trade- off between direct costs and benefits of helping should be incorporated into the broader theory attempting to explain cooperative breeding in birds.
2. METHODS
A colour-marked population of white-winged choughs on Black Mountain, Australian Capital Territory, 1490 15' E 360 05' S was studied from 1985 until 1989 (see H-einsohn (1992) for a description of reproduction in this population). Choughs are found in permanent cohesive groups of 2-16
Proc. R. Soc. Lond. B (1994)
which initiate nesting in September. They build a mud nest at 5-15 m above the ground in Eucalyptus trees, and typically lay a clutch of four eggs. All group members may contribute to nest building and incubation. Choughs are ideal for studying age-related behaviour as they are easily aged by eye colour until they are 5 years old (Rowley 1974). They have an extended adolescence of 4 years before reaching sexual maturity (Rowley 1978), making the only potential breeders in a group those over 4 years old. Clutch size in this study showed little variation. Most clutches had four eggs. Incubation takes between 18-21 days, and duration is independent of group size. Eggs are never left uncovered; there is always one individual sitting on the nest. Thus incubation demands are the same in total for all groups, but require a larger average effort for members of small groups.
We observed systematically the contribution of birds of varying age (to the nearest half year; individuals 5 years and older were pooled) to incubation in 16 groups ranging in size from 2 to 16 individuals. It was not possible to observe the contributions of all group members, as in some groups not all birds were colour banded. In total, data were available for 98 individuals. Observations of each group totalled between 2 and 11 hours per group (X= 6.2 h+0.8 s.e.), in bouts of 1 hour, spread equally over the incubation period. Many change-overs occurred each hour, allowing partitioning of time to be estimated with confidence in small groups, as all birds had bouts of incubation. We watched some larger groups for longer periods to confirm our suspicion that some of the birds were not contributing to incubation at all. Because the total observation time varied, contributions by individuals were converted to estimated proportions of incubation effort. These values are useful because they are directly comparable between groups. Data as proportions typically follow a binomial distribution, necessitating the use of logistic regression to assess the influence of parameters of interest (group size, age). Logistic regression relies on analysis of deviance. If the mean residual deviance (residual deviance divided by degrees of freedom) is equal to one, changes in residual deviance caused by the addition of factors or covariates approximate the x2 distribution (Dobson 1983). This was not the case in our analysis, so we used instead a more conservative measure, the deviance ratio, which approximates the F distribution (Dobson 1983). Logistic regressions were calculated by using GENSTAT (GENSTAT 5 Committee 1987).
To assess the impact of incubation, we measured body masses of birds by using remote-controlled scales. The scales were modified from a Bonso Model 322 from which the control panel had been removed and re-attached with 15 m of cable, enabling monitoring of mass from that distance. Choughs were attracted to the scales with cheese. We obtained paired masses of 11 juvenile (first year) helpers just before the start of incubation and close to the time of hatching (the time between measurements had a range of 24-33 days). The 11 juveniles came from 9 different groups. Some of these were from different groups from those used in the preceding analysis, as we had many juveniles marked in groups with only a few banded adults. It proved more difficult to attract older birds. Unpaired masses were also obtained for 49 individuals of various ages either before (n= 26) or after incubation (n = 23).
3. RESULTS
(a) Contributions to incubation
The proportion of time an individual devoted to incubation was affected by age (FE 97 = 75.8,
This content downloaded from 169.229.32.136 on Wed, 7 May 2014 21:32:48 PMAll use subject to JSTOR Terms and Conditions

A cost of helping behaviour in birds R. Heinsohn and A. Cockburn 295
(a) (b) (c) 0.6
0
,0.4 EI 0.2X _; 0
v - 0 X0 0 0.2 0 0 000 ~0 8 3
o9 0
00
~o.o _ _ _ _ _ _ _ _8 9 0
0 1 2 3 4 5+ 0 1 2 3 4 5+ 0 1 2 3 4 5+
age / years age I years age I years
Figure 1. The dependence of incubation effort on age and group size. (a) Birds in pairs (filled squares; there are no young birds in pairs) or in groups of 3-5 (circles); (b) birds from groups of 6-9 birds; (c) birds from groups of 12-16 birds. There are no young birds in pairs. Logistic regression equations are also depicted for one of the group sizes represented in each figure: (a) 4 birds; (b) 6 birds; (c) 12 birds. The full set of regression equations is presented in table 1.
Table 1. Regression coefficients and standard errors for nine group sizes in the logistic regression of proportion of incubation performed by an individual (pi) against age of individuals
(Birds of 5 years and older were pooled for analysis. fl0 and AX are the regression coefficients in the equation: logit (pi) = flo + l (age). There is no evidence that pi is affected by age for groups of 5 or less, but in larger groups incubation effort increases sharply with age.)
group size flo s.e. AX s.e.
2 1.02 1.01 -0.21 0.21 3 - 0.63 1.04 -0.02 0.22 4 -0.98 1.01 - 0.04 0.21 5 - 1.47 1.01 0.04 0.21 6 -2.38 1.01 0.30 0.21 7 - 3.29 1.02 0.53 0.21 9 - 2.98 1.01 0.31 0.21 12 -4.68 1.01 0.70 0.21 16 -4.28 1.02 0.55 0.21
p < 0.001), group size (EF889 = 172.1, p < 0.001) and their interaction (F8,81 = 5.5, p < 0.05). Intercepts, slopes and standard errors for the full set of regression equations are given in table 1. In small groups, age did not affect the amount of time devoted to incubation (figure 1, table 1). By contrast, incubation effort increased sharply with age in groups with six or more birds (table 1). Old birds contributed similar amounts of incubation, as the logistic curves converge for ages greater than 4 years (figure 1).
(b) Juvenile mass loss
Mass for all choughs increased with age (F1,69 = 92. 1, p < 0.001), as reported by Rowley ( 1978). However, in an analysis of covariance, there was no evidence that mass changed over the incubation period (F1,67 = 1.5). Thus there is no evidence that incubation causes loss of body mass across all age groups.
However, for the 11 juveniles weighed both before and after incubation, there were several interrelated trends with group size. Although body masses for these
Proc. R. Soc. Lond. B (1994)
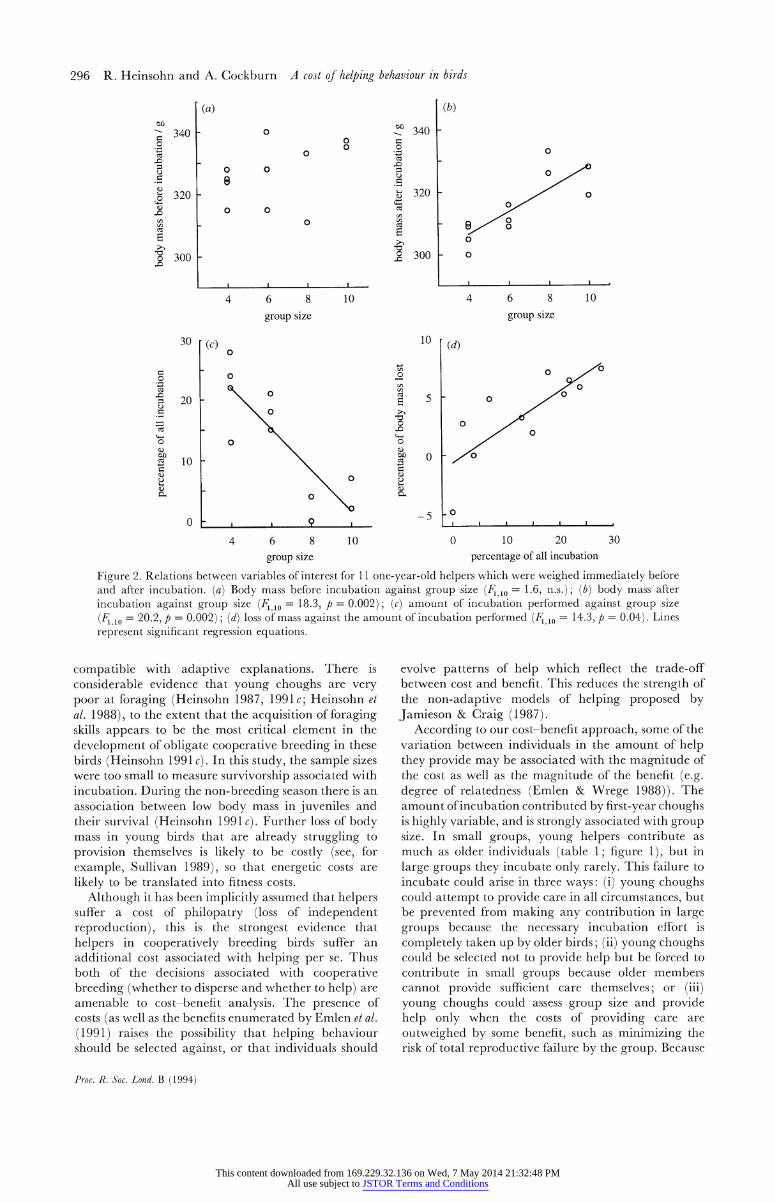
individuals were not dependent on group size before incubation (F110 = 1.6; figure 2a), mass after in- cubation increased with group size (F1 l0 = 18.3, p = 0.002; figure 2 b). The percentage of all incubation performed decreased as group size increased (FI,10 = 20.2, p = 0.002; figure 2 c), and the percentage of body mass lost increased with the proportion of incubation performed (F110 = 14.3, p = 0.04; figure 2d). Thus it is clear that juveniles in small groups performed a higher proportion of the incubation effort and lost more mass, but we cannot tell from these analyses whether it is the higher proportion of incubation performed or some other attribute of group size which determines mass loss.
To remove the effects of group size we used the residuals from the incubation-group size regression (figure 2 c) in a multiple regression analysis. The body mass (in grams) of an individual after incubation was proportional to its mass before incubation (F19 = 6.9, p = 0.03), but also decreased as relative incubation effort increased (Fj8 = 6.6, p = 0.03), giving the regression equation:
massafter = 38.5 + 0.9 (massbefore) -1.5 (residual).
Thus mass loss increases with actual incubation effort, independent of any effect of group size.
4. DISCUSSION
First-year white-winged choughs lose mass in pro- portion to the amount of incubation they contribute. This effect is sustained even when initial mass and effects of group size on the probability of incubation are considered explicitly. Older choughs appear un- affected by incubation.
It has been suggested that mass loss during re- production may be adaptive in birds (Freed 1981; Norberg 1981; Houston 1993). This argument has strong empirical support in some species (e.g. least auklets, Aethia pusilla (Jones 1994), but in others there is evidence that mass loss is costly (e.g. swifts, Apus apus (Martins & Wright 1993)). Because mass loss in white- winged choughs is confined to young birds which incubate, we do not believe that the data are
This content downloaded from 169.229.32.136 on Wed, 7 May 2014 21:32:48 PMAll use subject to JSTOR Terms and Conditions
296 R. Heinsohn and A. Cockburn A cost of helping behaviour in birds
(a) (b)
340 0 -.340 a 0 Da t 0 a 0 ~~~~~~~~~~~~~~~0 o ~~0 0 0
; 320 , 320 o 0 o , 0
0
0 o 300 , 300 o
4 6 8 10 4 6 8 10
group size group size
30 (c) 10 (d)
0 0
o 00 20 ,
- _ 0
0~~~~~~~~~~~ 0~~~~~~~~~
0~~~
0 0 o~~~~~~~
0 r 9 5 0
4 6 8 10 0 10 20 30
group size percentage of all incubation
Figure 2. Relations between variables of interest for 11 one-year-old helpers which were weighed immediately before and after incubation. (a) Body mass before incubation against group size (F1,10 = 1.6, n.s.); (b) body mass after incubation against group size (F1010 = 18.3, p = 0.002); (c) amount of incubation performed against group size
(F1 10 = 20.2, p = 0.002); (d) loss of mass against the amount of incubation performed (F1,10 = 14.3, p = 0.04). Lines represent significant regression equations.
compatible with adaptive explanations. There is considerable evidence that young choughs are very poor at foraging (Heinsohn 1987, 1991 c; Heinsohn et al. 1988), to the extent that the acquisition of foraging skills appears to be the most critical element in the development of obligate cooperative breeding in these birds (Heinsohn 1991 c). In this study, the sample sizes were too small to measure survivorship associated with incubation. During the non-breeding season there is an association between low body mass in juveniles and their survival (Heinsohn 1991 c). Further loss of body mass in young birds that are already struggling to provision themselves is likely to be costly (see, for example, Sullivan 1989), so that energetic costs are likely to be translated into fitness costs.
Although it has been implicitly assumed that helpers suffer a cost of philopatry (loss of independent reproduction), this is the strongest evidence that helpers in cooperatively breeding birds suffer an additional cost associated with helping per se. Thus both of the decisions associated with cooperative breeding (whether to disperse and whether to help) are amenable to cost-benefit analysis. The presence of costs (as well as the benefits enumerated by Emlen et al. (1991) raises the possibility that helping behaviour should be selected against, or that individuals should
Proc. R. Soc. Lond. B (1994)
evolve patterns of help which reflect the trade-off between cost and benefit. This reduces the strength of the non-adaptive models of helping proposed by Jamieson & Craig (1987).
According to our cost-benefit approach, some of the variation between individuals in the amount of help they provide may be associated with the magnitude of the cost as well as the magnitude of the benefit (e.g. degree of relatedness (Emlen & Wrege 1988)). The amount of incubation contributed by first-year choughs is highly variable, and is strongly associated with group size. In small groups, young helpers contribute as much as older individuals (table 1; figure 1), but in large groups they incubate only rarely. This failure to incubate could arise in three ways: (i) young choughs could attempt to provide care in all circumstances, but be prevented from making any contribution in large groups because the necessary incubation effort is completely taken up by older birds; (ii) young choughs could be selected not to provide help but be forced to contribute in small groups because older members cannot provide sufficient care themselves; or (iii) young choughs could assess group size and provide help only when the costs of providing care are outweighed by some benefit, such as minimizing the risk of total reproductive failure by the group. Because
This content downloaded from 169.229.32.136 on Wed, 7 May 2014 21:32:48 PMAll use subject to JSTOR Terms and Conditions

A cost of helping behaviour in birds R. Heinsohn and A. Cockburn 297
our data are based on correlations, we cannot distinguish these alternatives directly. None the less, the behaviour of choughs when incubation is rotated between group members provides some clues as to the direction of causation. Eggs are incubated at all times. An incubation bout by an individual usually terminates in two ways: (i) by the arrival of a group member or members at the nest, this inevitably prompts the immediate departure of the incubating bird, which resumes foraging, and the new arrival then takes over incubating; or (ii) some birds are unprepared to wait for relief, and call from the nest, prompting the arrival of another group member. Although egg laying and the start of incubation often entails considerable interest by all choughs, young birds in large groups never show unsolicited interest in an established nest. The universal immediate departure by the current incubator suggests that any chough which 'wanted' to incubate would be allowed to do so. The only instance when a nest was left unattended was when a young chough abandoned incubation and started foraging without soliciting a replacement. This suggests that choughs are not 'frustrated' helpers, and that their failure to incubate in large groups is either because they are forced to do so, or because they assess that help is unnecessary. We believe that foraging constraints (Heinsohn et al. 1988) and mass loss may be the source of reluctance to provide care.
Coupled with the observations that birds are more likely to help when provided with supplemental food (in moorhens), that adults intervene to stop birds that might help from doing so (in pied kingfishers), that adults frustrate attempts by their offspring to breed independently, so they remain at home and help (in bee-eaters), and punish birds that do not provide care (in superb fairy-wrens), our data suggest a richer interpretation of the dynamics of cooperative breeding societies than is conventionally applied. In addition to the view that cooperative breeding is driven by benefits accruing to the non-dispersers, cooperative societies are also influenced by the needs of the adults (or group) and the costs suffered by the helpers from which care is obtained. Further empirical and theoretical attention to these possibilities is required.
We thank Peter Dunn, David Green, Elsie Krebs, Kate Lessells, Rob Magrath, Penny Olsen and Linda Whitting- ham for comments on the manuscript, and the Australian National Botanic Gardens for permission to conduct research. John Grant suggested the use of remote-controlled scales, and Bruce Barrie built them.
REFERENCES
Arnold, W. 1990 The evolution of marmot sociality. I. Why disperse late? Behav. Ecol. Sociobiol. 27, 229-237.
Brown, J. L. 1987 Helping and communal breeding in birds: ecology and evolution. Princeton University Press.
Dobson, A. J. 1983 An introduction to statistical modelling. London: Chapman & Hall.
Emlen, S. T. 1991 Evolution of cooperative breeding in birds and mammals. In Behavioural ecology: an evolutionary
Proc. R. Soc. Lond. B (1994)
approach, 3rd edn (ed. J. R. Krebs & N. B. Davies), pp. 301-337. Oxford: Blackwell Scientific Publications.
Emlen, S. T. & Wrege, P. H. 1988 The role of kinship in helping decisions among white-fronted bee-eaters. Behav. Ecol. Sociobiol. 23, 305-315.
Emlen, S. T. & Wrege, P. H. 1992 Parent-offspring conflict and the recruitment of helpers among bee-eaters. Nature, London. 356, 331-333.
Emlen, S. T., Reeve, H. K., Sherman, P. W., Wrege, P. H., Ratnieks, F. L. W. & Shellman-Reeve, J. 1991 Adaptive versus nonadaptive explanations of behavior: the case of alloparental helping. Am. Nat. 138, 259-270.
Freed, L. A. 1981 Loss of mass in breeding wrens: stress or adaptation? Ecology 62, 1179-1186.
GENSTAT 5 Committee 1987 GENSTAT 5 reference manual. Oxford: Clarendon Press.
Heinsohn, R. G. 1987 Age-dependent vigilance in winter aggregations of cooperatively breeding white-winged choughs (Corcorax melanorhamphos). Behav. Ecol. Sociobiol. 20, 303-306.
Heinsohn, R. G. 1991 a Kidnapping and reciprocity in cooperatively breeding white-winged choughs. Anim. Behav. 41, 1097-1100.
Heinsohn, R. G. 1991 b Evolution of obligate cooperative breeding in white-winged choughs: a statistical approach. Acta XX Congr. Intern. Ornithol. 3, 1309-1316.
Heinsohn, R. G. 1991 c Slow learning of foraging skills and extended parental care in cooperatively breeding white- winged choughs. Am. Nat. 137, 864-881.
Heinsohn, R. G. 1992 Cooperative enhancement of re- productive success in white-winged choughs. Evol. Ecol. 6, 97-114.
Heinsohn, R. G., Cockburn, A. & Cunningham, R. B. 1988 Foraging, delayed maturity and cooperative breeding in white-winged choughs (Corcorax melanorhamphos). Ethology 77, 177-186.
Houston, A. I. 1993 The efficiency of mass loss in breeding birds. Proc. R. Soc. Lond. B 254, 221-225.
Jamieson, I. G. 1989 Behavioral heterochrony and the evolution of birds' helping at the nest: an unselected consequence of communal breeding? Am. Nat. 133, 394- 406.
Jamieson, I. G. 1991 The unselected hypothesis for the evolution of helping behavior: too much or too little emphasis on natural selection? Am. Nat. 138, 271-282.
Jamieson, I. G. & Craig, J. L. 1987 Critique of helping behaviour in birds: a departure from functional explana- tions. In Perspectives in ethology, vol. 7 (ed. P. Bateson & P. Klopfer), pp. 79-98. New York: Plenum Press.
Jones, I. L. 1994 Mass changes of least auklets Aethia pusilla during the breeding season: evidence for programmed loss of mass. J. Anim. Ecol. 63, 71-78.
Martins, T. L. F. & Wright, J. 1993 Cost of reproduction and allocation of food between parent and young in the swift (Apus apus). Behav. Ecol. 4, 213-223.
Mulder, R. A. & Langmore, N. E. 1993 Dominant males punish helpers for temporary defection in superb fairy- wrens. Anim. Behav. 45, 830-833.
Norberg, R. A. 1981 Temporary weight decrease in breeding birds may result in more fledged young. Am. Nat. 118, 838-850.
Reyer, H.-U. 1984 Investment and relatedness: a cost/ benefit analysis of breeding and helping in the pied kingfisher (Ceryle rudis). Anim. Behav. 32, 1163-1178.
Reyer, H.-U. & Westerterp, K. 1985 Parental energy expenditure: a proximate cause of helper recruitment in the pied kingfisher (Ceryle rudis). Behav. Ecol. Sociobiol. 17, 363-369.
Rowley, I. C. R. 1974 Bird life. Sydney: Collins.
This content downloaded from 169.229.32.136 on Wed, 7 May 2014 21:32:48 PMAll use subject to JSTOR Terms and Conditions

298 R. Heinsohn and A. Cockburn A cost of helping behaviour in birds
Rowley, I. 1978 Communal activities among white-winged choughs Corcorax melanorhamphos. Ibis 120, 178-197.
Stacey, P. B. & Ligon, J. D. 1987 Territory quality and dispersal options in the acorn woodpecker and a challenge to the habitat-saturation model of cooperative breeding. Am. Nat. 130, 654-676.
Sullivan, K. A. 1989 Predation and starvation: age-specific
mortality in juvenile juncos (Junco phaenotus). J. Anim. Ecol. 58, 275-286.
Taborsky, M. 1984 Broodcare helpers in the cichlid fish Lamprologus richardi: their costs and benefits. Anim. Behav. 32, 1236-1252.
Received 11 March 1994; accepted 23 March 1994
Proc. R. Soc. Lond. B (1994)
This content downloaded from 169.229.32.136 on Wed, 7 May 2014 21:32:48 PMAll use subject to JSTOR Terms and Conditions