Embed Size (px)
Citation preview

Review
10.1517/13543780802715792 © 2009 Informa UK Ltd ISSN 1354-3784 161All rights reserved: reproduction in whole or in part not permitted
Heat shock protein inhibitors and vaccines as new agents in cancer treatment Eleni M Karapanagiotou , Kostas Syrigos & Muhammad Wasif Saif † † Yale University School of Medicine, Medical Oncology, 333 Cedar Street, FMP: 116, New Haven, CT 06520, USA
Background : Heat shock proteins (HSP) play an essential role as molecular chaperones by assisting correct holding and folding in human cells. At the same time they present implications in tumor cell proliferation, differentiation, matrix invasion, angiogenesis, metastasis and cell death. They also possess the ability to present tumor molecules to the immune system and elicit an immune response. New agents targeting HSP, as anticancer treatment, are under clinical evaluation. Objective : The aim of this review is to explore the role of HSP90 inhibitors as anticancer agents as well as to evaluate the benefit from the use of autologous HSP vaccines in both the adjuvant setting and first-line treatment. Methods : The latest evidence regarding the use of geldanamycin analogues or newer water-soluble and synthetics molecules that inhibit the binding of HSP90 to client proteins was reviewed. Immunization using tumor-derived HSP in several cancer types was also evaluated. Conclusions : HSP90 inhibitors represent a promising therapeutic option although further evaluation in larger clinical trials is needed. On the other hand, HSP vaccination has already an established role in our armory against cancer.
Keywords: ansamycin family , apoptosis , cancer , geldanamycin analogues , heat shock protein 90 inhibitors , heat shock protein vaccination , heat shock proteins , immunity , synthetic non-ansamycin HSP90 inhibitor , thermotolerance , tumor
Expert Opin. Investig. Drugs (2009) 18 (2):161-174
1. Introduction
Heat shock proteins (HSP) or molecular chaperones comprise a large group of products of several distinct genes families with a vital role in cell survival during stress. They were first discovered in 1962 [1] as a set of highly conserved proteins whose expression was induced by heat shock, physical stress or several chemicals in a wide range of species [2,3] . HSP are named according to the approximate relative molecular mass of their encoded proteins: HSP100, -90, -70, -60 and small HSP including HSP27 [2,4-6] . They mainly reside in the cytoplasm and nucleus of the eukaryotic cells, with the exception of HSP60, which resides in mitochondria [7] . Some members of this large family, HSP90, for example, are constitutively expressed in cells whereas others, such as HSP70 and HSP27, are highly induced by stress factors. Each chaperone class displays relatively specialized functions but the members of different classes often work together, in large multi-protein complexes that also contain accessory proteins, to perform their duties [8] .
2. HSP functions
2.1 HSP intracellular functions The main intracellular role of HSP is that of molecular chaperoning for other cellular proteins. Their strong cytoprotective role consists of two principal functions,
1. Introduction
2. HSP functions
3. HSP90 and cancer
4. Development of HSP90 inhibitors
5. Development of anticancer
agents based on extracellular
functions of HSP
6. Conclusions
7. Expert opinion
Exp
ert O
pin.
Inv
estig
. Dru
gs D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsita
ets-
und
Lan
desb
iblio
thek
Due
ssel
dorf
on
09/1
9/13
For
pers
onal
use
onl
y.

Heat shock protein inhibitors and vaccines as new agents in cancer treatment
162 Expert Opin. Investig. Drugs (2009) 18(2)
namely, ‘protein holding’ and ‘protein folding’, aimed at balancing protein conformation under stress conditions. The main ‘holding proteins’ belong to the HSP70 and HSP90 families, which bind to unfolded sequences in polypeptide substrates showing preference for hydrophobic regions. They play a role in preventing premature self-associations in the nascent proteins during mRNA translation and protein degradation and accumulation during heat shock as well as in stabilizing unstable tertiary structures [9-11] . HSP90 and HSP70 act in association with accessory proteins in mediating substrate selection. After completion of their molecular chaperone function, they are released from protein substrates by means of their intrinsic ATP-ase domains [4] . Protein folding follows protein holding. This procedure needs the involvement of the HSP60 “chaperonin” family and other related chaperonin proteins. Chaperonins self-associate to form large folding chambers in which the substrate protein can undergo the appropriate intramolecular interactions required to attain its correct tertiary structure in an ATP-dependent process [12,13] . HSP70 might play a chaperonin role, as well [10] . One more type of HSP, the HSP-27, is also involved in ‘protein holding and folding’ using an ATP-independent mechanism [14] .
HSP gene transcription is regulated by transcription factors belonging to the Heat shock factor (HSF) family that ensure prompt transcriptional activation in stress and equally precipitous switch-off after recovery. The HSF family includes hsf1, the molecular coordinator of the heat shock response, as well as two less characterized heat shock factors hsf2 and hsf4 [15-19] . HSP transcription requires activated hsf1. The most accepted model for hsf1 function proposes that under unstressed conditions, the protein exists as a cytosolic monomer in latent complexes containing HSP70 and HSP90 [20-22] . Under stress conditions, hsf1 is liberated from HSP complexes and subsequently forms phosphorylated homotrimers in the nucleus that lead to HSP upregulation and production [21-23] . As expected mice deficient in hsf1 lack the ability to mount a classical heat shock response [24] .
2.2 HSP extracellular functions Besides to intracellular expression, HSP have been found to be located in the extracellular space or on the plasma membrane. Their presence in the serum is associated with stress conditions, including inflammation and infections, although the pathway that leads HSP to the plasma membrane remains unknown. It is suggested that cytosolic HSP are transported to the plasma membrane in concert with other proteins possessing transmembrane domains that fulfill shuttle functions or by a direct interaction of HSP with lipid components [25,26] .
Interestingly, necrotic but not apoptotic cells release HSP. This HSP release might act as a signal able to activate the immune system to recognize ‘dangerous’ situations [27] . HSP represent the carriers for several peptides. HSP 60, HSP70, HSP90 and HSP gp96 interact with peptides
from tumors or from cells containing tumors and form HSP–peptide complexes that enter antigen presenting cells (APCs) through specific surface receptors such as CD14 and Toll-like receptors (TLR) TLR-2 and TLR-4. After uptake and processing, the peptides are represented through MHC I molecules to CD8 + T cells, which differentiate to cytotoxic T lymphocytes [28-30] .
Even in the absence of immunogenic peptides, members of the HSP70 group provide danger signals for the host’s immune system [31] . Several studies have shown interactions of peptide-free HSP70 with CD14, a glycosylphosphatidylinositol (GPI)-anchored receptor, and TLR 2 /4 on APCs initiate the release of proinflammatory cytokines with a role in stimulating innate immune response, including TNF- α , IL-1 β , IL-12, IL-6 and GM-CSF. This process is triggered by the translocation of NF- κ B into the nucleus [32-34] .
One more extracellular function of HSP is their ability to stimulate the natural killer (NK) cells. NK cells play a central role in innate immunity and also modulate subsequent adaptive responses by means of cytokine production such as IFN- γ . NK cells present antitumor activity as well. They express inhibitory and activating receptors that interact with MHC class I molecules on target cells. Depending on intracellularly immunoreceptor tyrosine-based inhibitory or activatory motifs, these receptors mediate NK cell activation or tolerance towards self-antigens [35,36] . After stress, an HSP60-associated signaling peptide competes with human leukocyte antigen (HLA) leader peptides for binding to HLA-E. The inhibitory receptor complex CD94/NKG2A no longer recognizes HLA-E/HSP60–peptide complexes and thus these stressed cells provide targets for the cytolytic attack of NK cells. Moreover, HSP70 seems to act as a trigger factor for NK cells with a high cell surface density of CD94. Antibody blocking studies also suggested that CD94 is important in the interaction of NK cells with HSP70 presented on tumor cells. Surface expression of HSP70 on tumor membranes stimulates migratory and cytolytic activity of NK cells and induces apoptosis of tumor cells [37,38] . Apart from the earlier-mentioned chaperones, a variety of other HSPs were found to be present on the plasma membrane of tumor cell lines. NK-HSP-mediated lysis of tumor cells is not very clear. Strbo et al. demonstrated an essential role for perforin, the protein produced by NK cells in the activation of innate and adaptive immunity by gp96 complexes, which disrupts membranes of target cells and induces apoptosis [39] . Gross et al. demonstrated that the mechanism of lysis of HSP70 membrane positive tumor cells was biochemically characterized as grazyme B mediated but perforin-independent apoptosis [40] .
3. HSP90 and cancer
Under normal conditions HSP-90 represent 1 – 2% of cytosolic proteins whereas this expression is increased ≤ 3 times under stress conditions. There are two isoforms, HSP90a and
Exp
ert O
pin.
Inv
estig
. Dru
gs D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsita
ets-
und
Lan
desb
iblio
thek
Due
ssel
dorf
on
09/1
9/13
For
pers
onal
use
onl
y.

Karapanagiotou, Syrigos & Saif
Expert Opin. Investig. Drugs (2009) 18(2) 163
HSP90 β , with no differences in basic biochemical functions [41] . These proteins contain an N-terminal ATPase domain, a middle domain and a C-terminal domain containing protein–protein interaction and dimerization motifs. Like the N-terminal domain, the C-terminal domain may also contain an ATP binding site but the role of this site in regulating HSP90 function is not well understood. HSP90 performs in a large complex incorporating more core proteins, namely, immunophilins, HSP70–HSP40 complex that mediates formation of the HSP90-substrate interaction, the scaffold protein Hop, the p23 protein that mediates substrate choice and Cdc37 that facilitates the interactions with the substrate [42] . Both the middle and C-terminal regions are thought to contain binding sites for co-chaperones and client proteins. These interactions are regulated by means of HSP90 intrinsic ATPase activity [20] .
The principal role of this molecular chaperone is to stabilize the fragile structures of many of the receptors, protein-kinases and transcriptions factors that lie along with the pathways of normal cellular growth [43] . The HSP90-based complex is also required to maintain signaling proteins in an active conformation that can be rapidly triggered by growth signals [11,43] . Moreover, this complex is needed to maintain transformed proteins owing to overexpression or mutation in active conformation. Transformed proteins such as v-Src, BCR-ABL and p53 gain a stable configuration owing to HSP90 chaperoning function [44,45] . It seems HSP90 plays a critical role in tumor progression as it facilitates the autonomous function of unstable growth signaling molecules as well as serves as a biochemical ‘buffer’ within the growing pool of mutant proteins ( Figure 1 ).
HSP90 may inhibit apoptosis through several interactions. It has been reported that HSP90 binds directly to Apaf-1 and inhibits its oligomerization, recruitment of procaspase-9 and further apoptosis [46] . HSP90 also presents interactions
with phosphorylated serine/threonine kinase Akt/PKB, a protein that generates a survival signal in response to growth factor stimulation. Phosphorylated Akt can phosphorylate the Bcl-2 family protein Bad and caspase–9 leading to their inactivation and cell survival [47] . Besides, Akt can inhibit NF- κ B-mediated apoptosis through Ι κ B kinase phosphorylation [48] . Other routes through which HSP90 inhibits NF- κ B-mediated apoptosis are either via stabilization of the receptor inter-acting protein (RIP) [49] or by interactions within IκB kinase (IKK) complex, which include HSP90 and Cdc37 in its structural components [50] . One more implication of HSP90 in cell survival is its inhibitory action on the calcium-dependent proteases calpains [51] .
Increased expression of HSP90 has been implicated in resistance to senescence. All somatic cells possess replicative checkpoints that place limits on the number of permitted cell divisions. To escape senescence and undergo unlimited growth, tumor cells must bypass that point at which the telomeres on chromosomes have shortened enough to prevent successful future cell divisions. HSP90 may play an essential role in telomerase stability [52] .
HSP90, apart from HSP70, has a role in angiogenesis owing to its stabilizing properties on the transcription factor HIF1- α . Moreover, VEGF and nitric oxide synthase, two basic players in angiogenesis, require HSP90 for their induction and stability in vascular endothelial cells. These factors interact in a complex HSP90-dependent signaling network that ensures the assembly of new capillaries in the tumors. Several studies have shown that tumor angiogenesis present close relations with HSP90 overexpression, and HSP90 targeted drugs inhibit angiogenesis [53,54] .
Finally, HSP90 may possess a role in tumor invasion and metastasis. Tumor cells that overexpress HSF1 and HSP show an increased tendency to invade their microenviroment and to spread to distant organs [55] . HSP90 binds to matrix
Unlimitedreplication potential
Telomerase
MMP-2 MET
Self-sufficientgrowth factor signaling
Raf
Cyclin D
AKTIGF-1R
CDK4
ErbB-2/HER-2
Many of the molecular determinants oftumor growth and survival are HSP90 clients
HIF-1αVEGFR
Sustainedangiogenesis
Tissue invasionand metastases
Insensitivity toanti-growth signals
Avoidance ofapoptosis
HSP90
Figure 1 . HSP90 clients involved in tumor growth and survival.
Exp
ert O
pin.
Inv
estig
. Dru
gs D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsita
ets-
und
Lan
desb
iblio
thek
Due
ssel
dorf
on
09/1
9/13
For
pers
onal
use
onl
y.

Heat shock protein inhibitors and vaccines as new agents in cancer treatment
164 Expert Opin. Investig. Drugs (2009) 18(2)
metalloprotein 2 and this action represent the first step of extracellular invasion. Interestingly, HSP90 inhibitors have been implicated in bone metastasis in breast cancer. The mechanism involved is not fully explained [56] . The principal functions of HSP90 in normal and malignant cell are described in Table 1 .
4. Development of HSP90 inhibitors
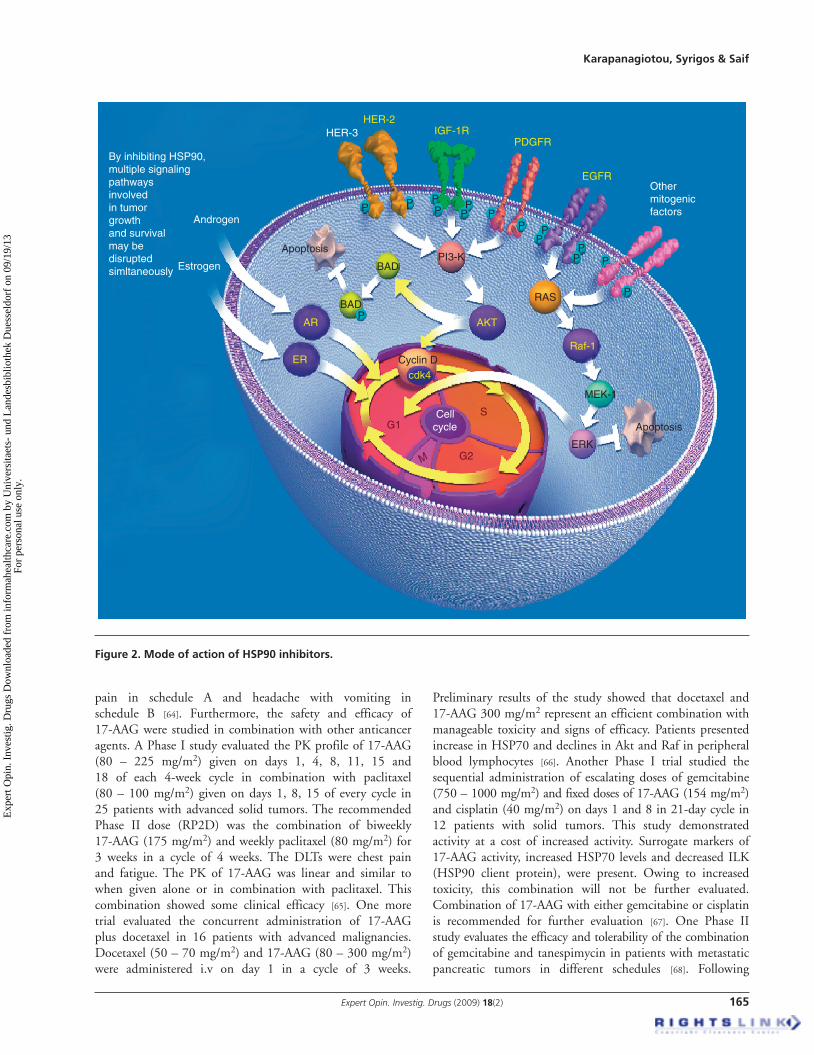
Initially, the development of HSP inhibitors was not such an attractive idea, as molecular chaperones work with substrates in a stoichiometric rather than a catalytic manner. But the ATPase domain of HSP90 has been effectively targeted and a very promising family of anticancer drugs has been produced. Although it might be anticipated that HSP90 inhibitors attacking proteins expressed in both normal and malignant cells would lack specificity and destroy both normal and cancerous tissues, this is not true, as these drugs selectively inhibit tumor cells. It has been observed that anti-HSP90 agents present with a protective role in normal cells, probably owing to the potent induction of other cytoprotective HSPs including HSP70. The principal antitumor activity of HSP90 inhibitors is the degradation of proteins required for autonomous growth as well as the inhibition of increased concentration of HSP90 substrates found in tumor cells ( Figure 2 ). Using HSP90 inhibitors, there is a fear of increasing resistance to cancer therapy caused by the selection of resistant traits, as HSP90 serves as a biochemical buffer in tumor cells, maintaining phenotypic normalcy in a highly stressful environment.
Some years ago, the discovery that the natural products geldanamycin and radicilol bind to HSP90 and result in client protein destabilization drew the interest for the potential use of these agents in anticancer treatment.
Table 1 . HSP90 functions in normal and malignant cells.
HSP90 functions in normal cell
Prevents proteins from unfolding during heat shock
Stabilizes the fragile structures of many of the receptors/protein kinases/transcription factors
Maintains signal proteins in an active conformation
Regulates mitogenesis (mediator of proliferation), gene transcription/cell regulation
HSP90 functions in malignant cell
Inhibits apoptosis via inhibition of several pathways
Stabilizes the conformation of mutant proteins (capacitor of evolution)
Sustains angiogenesis
Enhances invasion and metastasis
Permits limitless replicative potential
HSP: Heat shock proteins.
4.1 Geldanamycin Geldanamycin, a benzoquinone ansamycin, is able to compete with ATP at the nucleotide binding site in the NH2-terminal domain of HSP90 [57] . Inhibition of the essential ATPase activity of HSP90 leads to the degradation of several client proteins by the ubiquitin–proteasome pathway [58] resulting in cell cycle arrest and apoptosis [59] . In preclinical studies geldanamycin showed promising activity but in animal models it was found to be too hepatotoxic and could not be tested in humans [60] .
Following these results extensive research work led to new synthetic chemotypes and semisynthetic derivatives of the known natural products.
4.2 AAG 17-Allylamino-17-demethoxygeldamycin (17-AAG) is struc-turally similar to geldanamycin, maintains its antitumor activity and has a better toxicity profile and therapeutic index. This is the first anticancer drug of its class to enter clinical trials.
In a Phase I clinical trial 17-AAG was intravenously administered on days 1, 8 and 15 of a 28-day cycle in patients with advanced solid tumors. A total of 21 patients were treated. The maximum-tolerated dose (MTD) of weekly 17-AAG was 308 mg/m 2 where dose-limiting toxicities (DLTs) at 431 mg/m 2 were hepatic dysfunction, anemia, nausea, vomiting and myalgias. Its t 1/2 was 4.15 h. No tumor responses were seen. 17-AAG increased HSP70 levels in peripheral blood mononuclear cells (PBMCs) indicating that HSP90 has been affected [61] . One more Phase I clinical trial explored the pharmacokinetic–pharmacodynamic (PK–PD) profile of 17-AAG in 30 patients with advanced malignancies. The recommended dose for Phase II trials was 450 mg/m 2 /week. The DLTs were diarrhea and hepatotoxicity. Some clinical activity was observed as well as c-RAF-1 inhibition, CDK4 depletion and HSP70 induction using post-treatment biopsies [62] . A Phase I clinical trial explored 17-AAG as a 1-h infusion daily for 5 consecutive days in a cycle of 21 days in 19 patients with solid tumors. The MTD was 40 mg/m 2 /day whereas the main side effect was hepatic transaminitis. Less than 3% of the daily dose was excreted into the urine and clearance did not correlate with body surface area. 17-AAG biologic activity was suggested by increased levels of HSP70 and decreased levels of Raf1 [63] . On the basis of in vitro data showing increased activity of 17-AAG when it is administered at short intervals, Ramanathan et al. evaluated two schedules in 44 patients with advanced refractory tumors. Patients in schedule A received 17-AAG i.v. twice a week for 3 consecutive weeks and patients in schedule B twice a week for 2 consecutive weeks. Cycles were repeated after a 1-week rest. Both regimens underwent evaluation in combination with other antineoplastic drugs at the recommended Phase II dose of 17-AAG in schedule A of 150 mg/m 2 and in schedule B of 200 mg/m 2 . The DLTs included thrombocytopenia and abdominal
Exp
ert O
pin.
Inv
estig
. Dru
gs D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsita
ets-
und
Lan
desb
iblio
thek
Due
ssel
dorf
on
09/1
9/13
For
pers
onal
use
onl
y.
Karapanagiotou, Syrigos & Saif
Expert Opin. Investig. Drugs (2009) 18(2) 165
pain in schedule A and headache with vomiting in schedule B [64] . Furthermore, the safety and efficacy of 17-AAG were studied in combination with other anticancer agents. A Phase I study evaluated the PK profile of 17-AAG (80 – 225 mg/m 2 ) given on days 1, 4, 8, 11, 15 and 18 of each 4-week cycle in combination with paclitaxel (80 – 100 mg/m 2 ) given on days 1, 8, 15 of every cycle in 25 patients with advanced solid tumors. The recommended Phase II dose (RP2D) was the combination of biweekly 17-AAG (175 mg/m 2 ) and weekly paclitaxel (80 mg/m 2 ) for 3 weeks in a cycle of 4 weeks. The DLTs were chest pain and fatigue. The PK of 17-AAG was linear and similar to when given alone or in combination with paclitaxel. This combination showed some clinical efficacy [65] . One more trial evaluated the concurrent administration of 17-AAG plus docetaxel in 16 patients with advanced malignancies. Docetaxel (50 – 70 mg/m 2 ) and 17-AAG (80 – 300 mg/m 2 ) were administered i.v on day 1 in a cycle of 3 weeks.
Preliminary results of the study showed that docetaxel and 17-AAG 300 mg/m 2 represent an efficient combination with manageable toxicity and signs of efficacy. Patients presented increase in HSP70 and declines in Akt and Raf in peripheral blood lymphocytes [66] . Another Phase I trial studied the sequential administration of escalating doses of gemcitabine (750 – 1000 mg/m 2 ) and fixed doses of 17-AAG (154 mg/m 2 ) and cisplatin (40 mg/m 2 ) on days 1 and 8 in 21-day cycle in 12 patients with solid tumors. This study demonstrated activity at a cost of increased activity. Surrogate markers of 17-AAG activity, increased HSP70 levels and decreased ILK (HSP90 client protein), were present. Owing to increased toxicity, this combination will not be further evaluated. Combination of 17-AAG with either gemcitabine or cisplatin is recommended for further evaluation [67] . One Phase II study evaluates the efficacy and tolerability of the combination of gemcitabine and tanespimycin in patients with metastatic pancreatic tumors in different schedules [68] . Following
P
P
P PP P
P PP
PP
P P
P
P
RAS
Raf-1
AKT
PI3-KBAD
BAD
AR
ER
cdk4Cyclin D
MEK-1
ERK
G1
G2
SCellcycle
M
Apoptosis
Apoptosis
HER-3HER-2
IGF-1RPDGFR
EGFROthermitogenicfactors
Androgen
Estrogen
By inhibiting HSP90,multiple signalingpathwaysinvolvedin tumorgrowthand survivalmay bedisruptedsimltaneously
Figure 2 . Mode of action of HSP90 inhibitors.
Exp
ert O
pin.
Inv
estig
. Dru
gs D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsita
ets-
und
Lan
desb
iblio
thek
Due
ssel
dorf
on
09/1
9/13
For
pers
onal
use
onl
y.

Heat shock protein inhibitors and vaccines as new agents in cancer treatment
166 Expert Opin. Investig. Drugs (2009) 18(2)
Phase I clinical trial results, 17-AAG was evaluated for efficacy besides safety in Phase II clinical trials. A total of 15 eligible patients with hormone refractory metastatic prostate cancer and increased prostate serum antigen (PSA) levels received 300 mg/m 2 i.v. weekly for 3 weeks in 4-week cycles. No PSA response was shown. Unfortunately, study stopped recruitment and concluded that no further evaluation of this dose at this subpopulation is needed [69] . Positive results of Phase I study evaluating the concurrent administration of tanespimycin (Tan) (17-AAG in chremophor to increase water solubility or in suspension) and transtuzumab (T) in pretreated HER 2 positive patients with advanced solid tumors [70] led to a Phase II clinical trial that enrolled 29 patients with HER+ breast cancer who progressed on receiving transtuzumab. Women received standard weekly doses of transtuzumab followed by 450 mg/m 2 Tan. The results of this study were encouraging as 5 partial response (PR) and 5 stable disease (SD) were observed at a manageable toxicity. This study is still open [71] .
Owing to the fact that 17-AAG formulation is problematic, new formulations have been tested in clinical trials.
4.3 CNF1010 CNF1010, a lipid formulation of 17-AGG, has been evaluated in 30 patients with advanced solid tumors as 1-h i.v. infusion, twice a week, 3 weeks out of 4. There were no DLTs at doses ≤ 175 mg/m 2 whereas drug side effects included gastrointestinal toxicities, reversible hepatic enzyme elevation, hyperbilirubinemia, fatigue anemia and hyperglycemia. Plasma CNF1010 PK appeared dose-proportional whereas t 1/2 was 5.2 h and was dose independent. At doses ≥ 83 mg/m 2 CNF1010 showed efficacy that translated into minor tumor regressions associated with increases in HSP70 observed in PBMCs and decreases in plasma HER 2 ectodomain. Dose escalation of CNF1010 continues at 225 mg/m 2 [72] .
4.4 17-DMAG A more water-soluble geldanamycin analogue 17-dimethyl-aminoethylamino, 17-demethoxygeldanamycin (17-DMAG) has also entered clinical trials. Pharmacokinetic and PD studies of oral alvespimycin (KOS-1022; 17-DMAG) was performed in 28 patients with advanced malignancies. Two different schedules, one with daily administration and another with every other day for 4 out of 6 weeks, were studied without reaching any DLT. Drug-related toxicities included fatigue, nausea, anorexia, proteinuria and peripheral edema. Induction in HSP70 as well as tumor responses were present. Dose escalation continues in both schedules so that an RP2D is defined [73] .
4.5 IPI-504 One more water-soluble HSP90 inhibitor, IPI-504, has been evaluated in patients resistant to tyrosine-kinase inhibitor (TKI) metastatic gastrointestinal stromal tumors (GIST). On the basis of laboratories studies showing that inhibition
of the HSP90 results in selective destruction of the mutated KIT kinase in human GIST cell lines, IPI-504 was administered intravenously in patients with metastatic, TKI-resistant GIST or advanced/metastatic soft tissue sarcoma in 21-day cycles either twice weekly for 2 weeks followed by 1 week off treat-ment or twice weekly continuously for 3 weeks. IPI-504 was well-tolerated and the MTD for twice weekly for 2 weeks with 1 week off treatment is 400 mg/m 2 [74] . Following these results, 21 patients with progressive GIST after both previous imatinib and sunitib were enrolled in escalating dosage of i.v. IPI-504 on days 1, 4, 8 and 11 of a 21-day cycle. IPI-504 has been well tolerated with decreased fluorodeoxyglucose (FDG) avidity of GIST lesions, as well as the induction of SD in patients with previous progression. Owing to reactivation of positron emission tomography (PET) signal in lesions during drug breaks, new dosage schedules are under evaluation [75] . Recently, IPI504 was evaluated for efficacy and tolerability in a Phase II study in patients with unresectable stage IV or refractory melanoma as an intravenous infusion twice weekly in 3-week cycles. This study was prematurely closed [76] .
Owing to problematic bioavailability of geldanamycin and its derivates as well as increased hepatotoxicity that seems to result from the quinine ring, several groups are developing new completely synthetic HSP90 inhibitors. These include the synthesis of purine-based compounds that target the HSP90 nucleotide binding site with high affinity while demonstrating good aqueous solubility and oral bioavailability.
4.6 SNX-5422 SNX-5422 is a synthetic inhibitor of HSP90 entering clini-cal trials. It is an oral prodrug of SNX-2112, which seems to be more potent than 17-AAG and 17-DMAG in most of the cell lines tested. A Phase I dose escalation trial accrued 11 patients with solid malignancies. Patients were given SNX-5422 every other day for 21 days followed by 7 days of rest. No DLTs were observed and the drug was well tolerated. The t 1/2 was estimated at 8 – 10 h. The trial continues to enroll patients at a dose of 21.28 mg/m 2 [77] . More clinical Phase I trials are recruiting patients with relapsed solid malignancies and lymphomas. Patients receive SNX-5422 p.o. twice a week until disease progression while the primary outcome for both the MTD and the PK profile is studied [78] .
4.7 BIIB021 One more synthetic inhibitor of HSP90 is BIIB021 (CNF2024). BIIB021 is a fully synthetic non-anamycin HSP90 inhibitor, which is under investigation for solid and hematological malignancies. In a Phase I trial BIIB021 was evaluated in 23 patients with solid or hematological malignancies. Patients received 800 mg p.o. twice weekly. Preliminary results of the study showed that this oral HSP90 inhibitor is well tolerated at a dose of 800 mg p.o. twice weekly whereas biologic activity has been confirmed with elevations of serum and intracellular HSP70 and decreases in serum Her-2
Exp
ert O
pin.
Inv
estig
. Dru
gs D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsita
ets-
und
Lan
desb
iblio
thek
Due
ssel
dorf
on
09/1
9/13
For
pers
onal
use
onl
y.

Karapanagiotou, Syrigos & Saif
Expert Opin. Investig. Drugs (2009) 18(2) 167
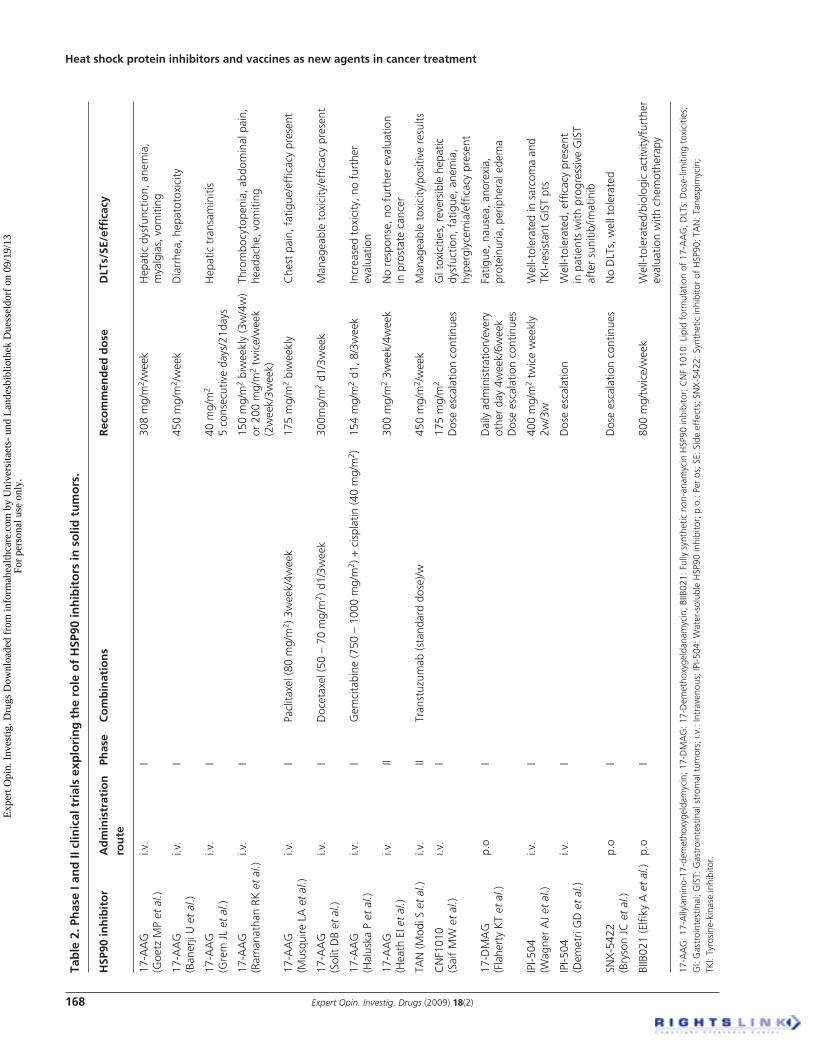
ECD. Further studies of BIIP021 in combination with chemotherapy and targeted agents are planned [79] . One more synthetic HSP90 inhibitor is STA-9090. This is a small molecule with a novel chemical structure, unrelated to geldanamycin or its family compounds. It has shown a potency that is 10 – 100 times greater than that of geldanamycin and its analogues in a wide range of cancer types. In in vivo models STA-9090 has shown activity even in cancers resistant to multikinase inhibitors. Two studies are recruiting patients with refractory to standard therapy solid malignancies to determine the MTD of STA-9090 administered as 1-h infusion once a week or twice a week for 3 consecutive weeks followed by a 1-week dose interval [78] . The results of Phase I and II clinical trials exploring the role of HSP90 inhibitors in solid tumors are seen in Table 2 .
5. Development of anticancer agents based on extracellular functions of HSP
Heat shock proteins can be used either as autologous vaccines using patient-specific antigens derived from the tumor of an individual patient or in vaccines using well-defined recombinant tumor antigens.
5.1 HSPPC-96; vitespen The first autologous HSP vaccine, Oncophage ® (Antigenics, Inc., New York, NY, USA)/vitespen, is an HSP–peptide complex (HSPPC)-96, a gp96 HSP peptide complex derived from resected tumor and then formulated for intradermal or subcutaneous administration. The HSP complexes bind to the CD91 receptor on antigen-presenting cells (APCs) and are taken into the cells. The APCs then travel to the lymph nodes, where they re-present the antigenic peptides on their surface. This action represents the trigger for a cancer immune response.
A large, multicenter, open-label, randomized Phase III study evaluated the efficacy, in terms of recurrence free survival (RFS), of the autologous, tumor-derived HSPPC-96 (vitespen) as adjuvant treatment in patients at high risk of recurrence after resection of locally advanced renal cell carcinoma (RCC). Patients were randomly assigned to receive either vitespen (n = 409) or observation alone. Vitespen was administered intradermally once a week for 4 weeks, then every 2 weeks until vaccine depletion. After a median follow-up of 1.9 years in the intention to treat (ITT) population, recurrence events were reported in 136 (37.7%) patients in the vitespen group and in 146 (39.8%) in the observation group (HR: 0.923, 95% confidence interval, CI, 0.729 – 1.169, p = 0.506). The patients continue to be followed up for survival. No serious side effects were present except one event of autoimmune thyroiditis grade 2 in the vitespen group. Injection-site erythema and injection-site induration were common adverse events in this group. Although no difference in RFS was seen between vitespen and observation group, a trend in favor of vitespen was observed
in patients with disease stage I and II (HR: 0.576, 95% CI 0.324 – 1.023, p = 0.056) [80] . Following this study, vitespen was approved in Russia for the treatment of kidney cancer patients at intermediate risk of recurrence after surgery.
One more study evaluated the clinical efficacy as determined by time to progression (TTP) and response rate (RR) of autologous vitespen (formerly HSPPC-96; Oncophage) with and without the administration of IL-2 (IL-2; Prloleukin: Chiron ® , Emoryville, CA, USA) in stage IV metastatic RCC patients undergoing nephrectomy. Vaccines were administered at weeks 1, 2, 3, 4, 6, 8 followed by restaging at around week 10. Patients who had stable or responsive disease continued to receive vaccine, with four more vaccinations biweekly. Patients who had progressive disease received four consecutive 5-day-per-week courses of 11*10(6) U of IL-2 s.c. with four doses of vitespen at 2-week intervals. At the next evaluation, patients with a complete response (CR) received two further cycles of vitespen (with IL-2 if also received during previous cycle) or until vaccine supply was exhausted. Patients with SD or PR repeated their previous cycle of therapy. Disease progressors who had not yet received IL-2 began IL-2 treatment, and progressors who had already received IL-2 came off study. Study results demonstrated that 2 patients showed CR, 2 showed PR, 7 showed SD and 33 progressed out of 60 evaluable patients. Treatment with vitespen did not result in a remarkable benefit in most of the patients with metastatic RCC [81] .
The second large, open-label, multicenter, Phase III clinical study evaluated the antitumor activity of vitespen, in terms of overall survival (OS), in patients with previous untreated melanoma stage IV. A total of 322 patients were randomly assigned 2:1 to receive vitespen or physician’s choice containing one or more of the following: dacarbazine, temozolamide, IL-2 or complete tumor resection. Patients on the vitespen arm received variable number of injections owing to the autologous nature of vitespen therapy. The OS was indistin-guishable between the two arms, although patients with M1a and M1b substages receiving a larger number of vitespen immunizations survived longer than those receiving fewer such treatments. This result is in line with the immunologic mechanism of action of vitespen indicating delayed onset of clinical activity after exposure to the vaccine. Further studies are needed to clarify the effectiveness of vitespen in these subpopulations [82] .
A smaller Phase II study evaluated the immunogenicity and antitumor activity of autologous, tumor-derived HSPPC-96 (Oncophage) given with GM-CSF and IFN- α in pretreated metastatic melanoma patients. Patients underwent surgical resection of metastatic lesions for HSPPC-96 production. The vaccine was administered s.c. in 4-weekly intervals along with GM-CSF s.c. at the same site at days -1, 0, +1 and IFN- α (3 MU) s.c. at a different site at days +4 and +6. Patients who responded received four extra injections in 2-week intervals or more depending on available vaccine.
Exp
ert O
pin.
Inv
estig
. Dru
gs D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsita
ets-
und
Lan
desb
iblio
thek
Due
ssel
dorf
on
09/1
9/13
For
pers
onal
use
onl
y.
Heat shock protein inhibitors and vaccines as new agents in cancer treatment
168 Expert Opin. Investig. Drugs (2009) 18(2)
Tab
le 2
. Ph
ase
I an
d II
clin
ical
tri
als
exp
lori
ng
th
e ro
le o
f H
SP90
inh
ibit
ors
in s
olid
tu
mo
rs.
HSP
90 in
hib
ito
r A
dm
inis
trat
ion
ro
ute
Ph
ase
Co
mb
inat
ion
s R
eco
mm
end
ed d
ose
D
LTs/
SE/e
ffi c
acy
17-A
AG
(G
oetz
MP
et a
l. )i.v
.I
308
mg/
m 2 /
wee
kH
epat
ic d
ysfu
nctio
n, a
nem
ia,
mya
lgia
s, v
omiti
ng
17-A
AG
(B
aner
ji U
et
al .)
i.v.
I45
0 m
g/m
2 /w
eek
Dia
rrhe
a, h
epat
otox
icity
17-A
AG
(G
rem
JL
et a
l. )i.v
.I
40 m
g/m
2 5
cons
ecut
ive
days
/21d
ays
Hep
atic
tra
nsam
initi
s
17-A
AG
(R
aman
atha
n RK
et
al. )
i.v.
I15
0 m
g/m
2 bi
wee
kly
(3w
/4w
) or
200
mg/
m 2
twic
e/w
eek
(2w
eek/
3wee
k)
Thro
mbo
cyto
peni
a, a
bdom
inal
pai
n,
head
ache
, vom
iting
17-A
AG
(M
usqu
ire L
A e
t al
. )i.v
.I
Pacl
itaxe
l (80
mg/
m 2 )
3w
eek/
4wee
k17
5 m
g/m
2 bi
wee
kly
Che
st p
ain,
fat
igue
/effi
cac
y pr
esen
t
17-A
AG
(S
olit
DB
et a
l. )i.v
.I
Doc
etax
el (5
0 –
70 m
g/m
2 ) d
1/3w
eek
300m
g/m
2 d1
/3w
eek
Man
agea
ble
toxi
city
/effi
cac
y pr
esen
t
17-A
AG
(H
alus
ka P
et
al. )
i.v.
IG
emci
tabi
ne (7
50 –
100
0 m
g/m
2 ) +
cis
plat
in (4
0 m
g/m
2 )15
4 m
g/m
2 d1
, 8/3
wee
kIn
crea
sed
toxi
city
, no
furt
her
eval
uatio
n
17-A
AG
(H
eath
EI e
t al
. )i.v
.II
300
mg/
m 2
3wee
k/4w
eek
No
resp
onse
, no
furt
her
eval
uatio
n in
pro
stat
e ca
ncer
TAN
(Mod
i S e
t al
. )i.v
.II
Tran
stuz
umab
(sta
ndar
d do
se)/w
450
mg/
m 2 /
wee
kM
anag
eabl
e to
xici
ty/p
ositi
ve r
esul
ts
CN
F101
0 (S
aif
MW
et
al. )
i.v.
I17
5 m
g/m
2 D
ose
esca
latio
n co
ntin
ues
GI t
oxic
ities
, rev
ersi
ble
hepa
tic
dysf
uctio
n, f
atig
ue, a
nem
ia,
hype
rgly
cem
ia/e
ffi c
acy
pres
ent
17-D
MA
G
(Fla
hert
y K
T et
al .)
p.o
ID
aily
adm
inis
trat
ion/
ever
y ot
her
day
4wee
k/6w
eek
Dos
e es
cala
tion
cont
inue
s
Fatig
ue, n
ause
a, a
nore
xia,
pr
otei
nuria
, per
iphe
ral e
dem
a
IPI-5
04
(Wag
ner
AJ
et a
l .)i.v
.I
400
mg/
m 2
twic
e w
eekl
y 2w
/3w
Wel
l-tol
erat
ed in
sar
com
a an
d TK
I-res
ista
nt G
IST
pts
IPI-5
04
(Dem
etri
GD
et
al. )
i.v.
ID
ose
esca
latio
nW
ell-t
oler
ated
, effi
cac
y pr
esen
t in
pat
ient
s w
ith p
rogr
essi
ve G
IST
afte
r su
nitib
/imat
inib
SNX
-542
2 (B
ryso
n JC
et
al .)
p.o
ID
ose
esca
latio
n co
ntin
ues
No
DLT
s, w
ell t
oler
ated
BIIB
021
(Elfi
ky A
et
al. )
p.o
I80
0 m
g/tw
ice/
wee
kW
ell-t
oler
ated
/bio
logi
c ac
tivity
/fur
ther
ev
alua
tion
with
che
mot
hera
py
17-A
AG
: 17-
Ally
lam
ino-
17-d
emet
hoxy
geld
amyc
in; 1
7-D
MA
G: 1
7-D
emet
hoxy
geld
anam
ycin
; BIIB
021:
Ful
ly s
ynth
etic
non
-ana
myc
in H
SP90
inhi
bito
r; C
NF
1010
: Lip
id f
orm
ulat
ion
of 1
7-A
AG
; DLT
s: D
ose-
limiti
ng t
oxic
ities
; G
I: G
astr
oint
estin
al; G
IST:
Gas
troi
ntes
tinal
str
omal
tum
ors;
i.v.
: Int
rave
nous
; IPI
-504
: Wat
er-s
olub
le H
SP90
inhi
bito
r; p
.o.:
Per
os; S
E: S
ide
effe
cts;
SN
X-5
422:
Syn
thet
ic in
hibi
tor
of H
SP90
; TA
N: T
anes
pim
ycin
; TK
I: Ty
rosi
ne-k
inas
e in
hibi
tor.
Exp
ert O
pin.
Inv
estig
. Dru
gs D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsita
ets-
und
Lan
desb
iblio
thek
Due
ssel
dorf
on
09/1
9/13
For
pers
onal
use
onl
y.

Karapanagiotou, Syrigos & Saif
Expert Opin. Investig. Drugs (2009) 18(2) 169
A total of 38 patients were enrolled but only 20 were evalu-able. Clinical activity was limited to SD, as 11 patients showed SD. Both tumor-specific T-cell-mediated and NK cell responses were generated in a proportion of patients [83] . The groundwork for the previous study was laid by a study that examined the immunogenicity and antitumor activity of HSPPC-96 as monotherapy in metastatic melanoma patients. A total of 42 patients were able to receive the vaccine and 39 were assessable after one cycle of vaccination divided in 4-weekly injections. In 21 patients, a second cycle of 4-weekly injections was given because no progression occurred. At the end of follow-up 2 patients presented with CR, and 3 with SD out of 28 enrollments with measurable disease. Researchers concluded that Oncophage vaccine induces clinical and tumor-specific T-cell responses in a significant minority of patients [84] . Finally, Oncophage has been granted fast track and orphan drug designations from the US FDA for kidney cancer and metastatic melanoma. Oncophage has orphan drug status for kidney cancer from the European Medicine Agency.
HSPPC-96 (Oncophage; vitespen) was also administered in 14 patients with NSCLC stage IB to IIIA who underwent surgery with complete resection of the tumor and no evidence of metastatic disease. The vaccine was administered subcutaneously 6 – 8 weeks postsurgery once weekly for 4 weeks and then every 2 weeks until vaccine depletion. Vaccine production feasibility was the primary end point of the study whereas secondary end points were RFS, OS and safety. This study showed that the manufacturing success of HSPPC-96 for this indication was 100%, and the RFS ranged from 24 to 33 months.
This type of vaccine was also administered in 10 patients with adenocarcinoma of the pancreas after complete resection of the tumor. A dose of 5 µg of autologous HSPPC-96 was administered weekly for four doses. There were no DLTs and an estimated OS of 2.2 years. Besides, no correlation between immune response and prognosis was observed.
Oncophage was also evaluated in small cohorts of patients with gastric carcinoma. In a Phase I trial, 21 patients underwent R 0 or R 1 resection for gastric carcinoma and received intradermal injections at four different dose levels. This study showed no toxicity and encouraging results [85] . This vaccine was also evaluated in patients with metastatic colorectal cancer to liver. A total of 29 patients were vaccinated with the vaccine manufactured from resected liver metastases. The first cycle consisted of 4 weekly s.c. injections and the second one of 4 biweekly s.c. injections given after a 4-week break. An immune response was identified by a significant increase in T cells post-vaccination. Statistical analysis showed T-cell immune response to be an independent factor for prognosis for both good and bad prognostic group patients [86] . A continuing study evaluates the safety and efficacy of HSPPC-96 in patients with recurrent glioma.
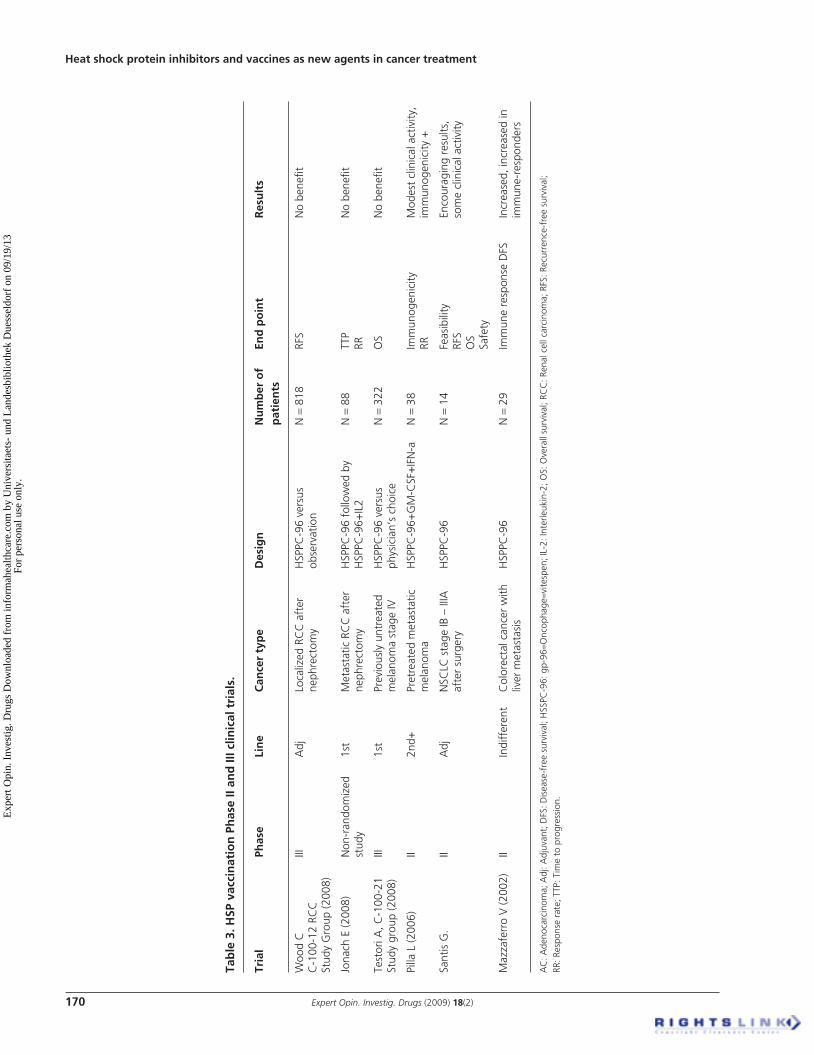
The results of Phase II and III clinical trials exploring the role of HSP vaccination in solid tumors are seen in Table 3 .
5.2 HSP110 complexed to GP100 A second approach to HSP vaccination is the manufacturing of a vaccine using recombinant tumor antigens. A vaccine consisting of HSP110 complexed to the intracellular domain (ICD) of the breast cancer antigen HER-2/neu that elicits both a strong antigen-specific type I cellular immunity and humoral responses was produced [87] . This vaccine stimulates CD8 + and CD4 + T-cell responses against ICD in mice [88] . Immunization with HSP110 complexed to gp100 (a human melanoma-associated antigen) protected mice against subsequent challenge with human gp100-transduced B16 melanoma. Vaccination also significantly suppressed the growth of established tumors in a therapeutic model.
6. Conclusions
Heat shock proteins possess a very exciting role in cells being overexpressed under stress conditions. Overexpression of HSP is present in a variety of tumors and is associated with poor prognosis and therapy resistance. For many years inhibition of HSP was not an attractive target as these proteins act in a stoichiometric manner rather than in a catalytic manner. The breakthrough was made when it was found that geldanamycin and its analogues could inhibit the ATP-ase domain of HSP90 and cause inhibition of the binding of HSP90 to the client proteins. Since then, many strides have been made in manufacturing new agents that are more potent and less toxic. These new HSP90 inhibitors are under clinical evaluation in Phase I and II clinical trials in solid tumors as monotherapy or in combination with other chemotherapeutic or biologic agents. HSP90 inhibitors present a satisfactory toxicity profile whereas their efficacy remains to be further evaluated. So far, the findings of clinical trials have been encouraging. Moreover, the role of HSP as carriers that present tumor molecules to the immune system has been evaluated in large clinical trials. The immunization is carried out with tumor-derived HSP, which bring attached the specific tumor peptides. The HSPPC-96 has successfully tested in large Phase III clinical studies in patients with RCC and melanoma. Efficacy using the autologous HSPPC-96 vaccine has been seen in a variety of solid tumors. As the inhibition of HSP leads to further inhibition of several pathways of tumorigenesis, further exploration of HSP inhibitors is needed.
7. Expert opinion
Very promising strides have been made in the development of HSP inhibitors. Using the latest information on the role of HSP in the intracellular as well as in the extracellular domain inhibitors of HSP90 have been developed and have entered clinical trials. Ansamycin family drugs were first used as HSP90 inhibitors, as they could target ATPase domain of
Exp
ert O
pin.
Inv
estig
. Dru
gs D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsita
ets-
und
Lan
desb
iblio
thek
Due
ssel
dorf
on
09/1
9/13
For
pers
onal
use
onl
y.
Heat shock protein inhibitors and vaccines as new agents in cancer treatment
170 Expert Opin. Investig. Drugs (2009) 18(2)
Tab
le 3
. HSP
vac
cin
atio
n P
has
e II
and
III c
linic
al t
rial
s.
Tria
l Ph
ase
Lin
e C
ance
r ty
pe
Des
ign
N
um
ber
of
pat
ien
ts
End
po
int
Res
ult
s
Woo
d C
C
-100
-12
RCC
St
udy
Gro
up (2
008)
IIIA
djLo
caliz
ed R
CC
aft
er
neph
rect
omy
HSP
PC-9
6 ve
rsus
ob
serv
atio
nN
= 8
18RF
SN
o be
nefi t
Jona
ch E
(200
8)N
on-r
ando
miz
ed
stud
y1s
tM
etas
tatic
RC
C a
fter
ne
phre
ctom
yH
SPPC
-96
follo
wed
by
HSP
PC-9
6+IL
2N
= 8
8TT
P RR
No
bene
fi t
Test
ori A
, C-1
00-2
1 St
udy
grou
p (2
008)
III1s
tPr
evio
usly
unt
reat
ed
mel
anom
a st
age
IVH
SPPC
-96
vers
us
phys
icia
n’s
choi
ceN
= 3
22O
SN
o be
nefi t
Pilla
L (2
006)
II2n
d+Pr
etre
ated
met
asta
tic
mel
anom
aH
SPPC
-96+
GM
-CSF
+IF
N-a
N =
38
Imm
unog
enic
ity
RRM
odes
t cl
inic
al a
ctiv
ity,
imm
unog
enic
ity +
Sant
is G
.II
Adj
NSC
LC s
tage
IB –
IIIA
af
ter
surg
ery
HSP
PC-9
6N
= 1
4Fe
asib
ility
RF
S O
S Sa
fety
Enco
urag
ing
resu
lts,
som
e cl
inic
al a
ctiv
ity
Maz
zafe
rro
V (2
002)
IIIn
diff
eren
tC
olor
ecta
l can
cer
with
liv
er m
etas
tasi
sH
SPPC
-96
N =
29
Imm
une
resp
onse
DFS
Incr
ease
d, in
crea
sed
in
imm
une-
resp
onde
rs
AC
: Ade
noca
rcin
oma;
Adj
: Adj
uvan
t; D
FS: D
isea
se-f
ree
surv
ival
; HSS
PC-9
6: g
p-96
= Onc
opha
ge = v
itesp
en; I
L-2:
Inte
rleuk
in-2
; OS:
Ove
rall
surv
ival
; RC
C: R
enal
cel
l car
cino
ma;
RFS
: Rec
urre
nce-
free
sur
viva
l; RR
: Res
pons
e ra
te; T
TP: T
ime
to p
rogr
essi
on.
Exp
ert O
pin.
Inv
estig
. Dru
gs D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsita
ets-
und
Lan
desb
iblio
thek
Due
ssel
dorf
on
09/1
9/13
For
pers
onal
use
onl
y.

Karapanagiotou, Syrigos & Saif
Expert Opin. Investig. Drugs (2009) 18(2) 171
HSP90 and further inhibit HSP90 chaperoning function. These agents were abandoned owing to poor solubility and increased hepatotoxicity. Geldanamycin water-soluble derivatives such as 17-AAG and 17-DMAG with an increased potency to inhibit HSP90 showed a better bioavailability and safer toxicity profile. Their use in Phase I and II clinical trials guided us to a better understanding of PK and PD profile of these agents. Increased hepatotoxicities as well as gastroinitestinal symptoms represent the principal drawbacks to be solved. In an attempt to overcome these obstacles new, more water-soluble formulations such as tanespimycin or synthetic geldanamycin analogues have been developed by several research groups. Moreover, new fully synthetic and non-anamycin molecules have been developed such as BIIB021. The results from Phase I and II clinical trials exploring the role of these new agents as anticancer drugs are encouraging. These new molecules have shown activity in many different types of solid tumors including breast cancer, metastatic sarcomas and relapsed GIST previously treated with imatinib and sunitinib. In spite of some encouraging results, HSP90 inhibitors did not show any efficacy in prostate cancer. The combina-tions of HSP90 inhibitors with other chemotherapeutic or biologic agents need further evaluation. Some studies show that these combinations are safe and efficient
whereas some others conclude that these combinations are toxic and inefficient.
Regarding vaccination using the HSPPC-96, a gp96 HSP peptide complex derived from resected tumor and then formulated for intradermal or subcutaneous administration has made vast progress. Clinical efficacy of HSPPC-96 has been evaluated in large-scale Phase III clinical trials in patients with renal cancer and melanoma. Vitespen (Oncophage, HSPPC-96) has showed some degree of efficacy as an adjuvant treatment in subpopulations of advanced renal cancer patients treated with surgery and in metastatic melanoma patients. It presents a satisfactory toxic profile but its main limitation is the need of autologous tissue. This vaccination is only available for patients under-going some kind of surgery. The autologous HSPPC-96 vaccination has been evaluated in many other solid tumors including NSCLC, pancreatic cancer, gastric cancer and colorectal cancer with liver metastates in an adjuvant setting with encouraging results.
Declaration of interest
The authors state no conflict of interest and have received no payment in preparation of this manuscript.
Bibliography Papers of special note have been highlighted as either of interest (•) or of considerable interest (••) to readers.
1. Ritossa P. Problems of prophylactic vaccinations of infants. Riv Ist Sieroter Ital 1962 ; 37 : 79 -108
2. Lindquist S, Craig EA. The heat-shock proteins. Annu Rev Genet 1988 ; 22 : 631 -77
3. Georgopoulos C, Welch WJ. Role of the major heat shock proteins as molecular chaperones. Annu Rev Cell Biol 1993 ; 9 : 601 -34
4. Bukau B, Horwich AL. The Hsp70 and Hsp60 chaperone machines. Cell 1998 ; 92 : 351 -66
5. Tang D, Khaleque MA, Jones EL, et al. Expression of heat shock proteins and heat shock protein messenger ribonucleic acid in human prostate carcinoma in vitro and in tumors in vivo. Cell Stress Chaperones 2005 ; 10 : 46 -58
6. Easton DP, Kaneko Y, Subjeck JR. The hsp110 and Grp1 70 stress proteins: newly recognized relatives of the Hsp70s. Cell Stress Chaperones 2000 ; 5 : 276 -90
7. Rachel A, Tyson JR, Stirling CJ. A novel subfamily of Hsp70s in the endoplasmic reticulum. Trends Cell Biol 1997 ; 7 : 277 -82
8. Nollen EA, Morimoto RI. Chaperoning signaling pathways: molecular chaperones as stress-sensing ‘heat shock’ proteins. J Cell Sci 2002 ; 115 : 2809 -16
9. Wegele H, Muller L, Buchner J. Hsp70 and Hsp90--a relay team for protein folding. Rev Physiol Biochem Pharmacol 2004 ; 151 : 1 -44
10. Mayer MP, Bukau B. Hsp70 chaperones: cellular functions and molecular mechanism. Cell Mol Life Sci 2005 ; 62 : 670 -84
11. Pratt WB, Toft DO. Regulation of signaling protein function and traffi cking by the hsp90/hsp70-based chaperone machinery. Exp Biol Med (Maywood) 2003 ; 228 : 111 -33
12. Spiess C, Meyer AS, Reissmann S, et al. Mechanism of the eukaryotic chaperonin: protein folding in the chamber of secrets. Trends Cell Biol 2004 ; 14 : 598 -604
13. Young JC, Agashe VR, Siegers K, et al. Pathways of chaperone-mediated protein folding in the cytosol. Nat Rev Mol Cell Biol 2004 ; 5 : 781 -91
14. Arrigo AP. Heat shock proteins as molecular chaperones. Med Sci (Paris) 2005 ; 21 : 619 -25
15. Nakai A, Tanabe M, Kawazoe Y, et al. HSF4, a new member of the human heat shock factor family which lacks properties
of a transcriptional activator. Mol Cell Biol 1997 ; 17 : 469 -81
16. Rabindran SK, Giorgi G, Clos J, et al. Molecular cloning and expression of a human heat shock factor, HSF1. Proc Natl Acad Sci USA 1991 ; 88 : 6906 -10
17. Schuetz TJ, Gallo GJ, Sheldon L, et al. Isolation of a cDNA for HSF2: evidence for two heat shock factor genes in humans. Proc Natl Acad Sci USA 1991 ; 88 : 6911 -5
18. Sorger PK, Pelham HR. Yeast heat shock factor is an essential DNA-binding protein that exhibits temperature-dependent phosphorylation. Cell 1988 ; 54 : 855 -64
19. Wu C. Heat shock transcription factors: structure and regulation. Annu Rev Cell Dev Biol 1995 ; 11 : 441 -69
20. Nadeau K, Das A, Walsh CT. Hsp90 chaperonins possess ATPase activity and bind heat shock transcription factors and peptidyl prolyl isomerases. J Biol Chem 1993 ; 268 : 1479 -87
21. Baler R, Dahl G, Voellmy R. Activation of human heat shock genes is accompanied by oligomerization, modifi cation, and rapid translocation of heat shock transcription factor HSF1. Mol Cell Biol 1993 ; 13 : 2486 -96
22. Sarge KD, Murphy SP, Morimoto RI. Activation of heat shock gene transcription by heat shock factor 1 involves
Exp
ert O
pin.
Inv
estig
. Dru
gs D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsita
ets-
und
Lan
desb
iblio
thek
Due
ssel
dorf
on
09/1
9/13
For
pers
onal
use
onl
y.

Heat shock protein inhibitors and vaccines as new agents in cancer treatment
172 Expert Opin. Investig. Drugs (2009) 18(2)
oligomerization, acquisition of DNA-binding activity, and nuclear localization and can occur in the absence of stress. Mol Cell Biol 1993 ; 13 : 1392 -407
23. Trinklein ND, Murray JI, Hartman SJ, et al. The role of heat shock transcription factor 1 in the genome-wide regulation of the mammalian heat shock response. Mol Biol Cell 2004 ; 15 : 1254 -61
24. McMillan DR, Xiao X, Shao L, et al. Targeted disruption of heat shock transcription factor 1 abolishes thermotolerance and protection against heat-inducible apoptosis. J Biol Chem 1998 ; 273 : 7523 -8
25. Schmitt E, Gehrmann M, Brunet M, et al. Intracellular and extracellular functions of heat shock proteins: repercussions in cancer therapy. J Leukoc Biol 2007 ; 81 : 15 -27
26. Arispe N, Doh M, Simakova O, et al. Hsc70 and Hsp70 interact with phosphatidylserine on the surface of PC12 cells resulting in a decrease of viability. FASEB J 2004 ; 18 : 1636 -45
27. Todryk SM, Melcher AA, Dalgleish AG, et al. Heat shock proteins refi ne the danger theory. Immunology 2000 ; 99 : 334 -7
28. Singh-Jasuja H, Toes RE, Spee P, et al. Cross-presentation of glycoprotein 96-associated antigens on major histocompatibility complex class I molecules requires receptor-mediated endocytosis. J Exp Med 2000 ; 191 : 1965 -74
29. Schild H, Rammensee HG. gp96 – the immune system’s Swiss army knife. Nat Immunol 2000 ; 1 : 100 -1
30. Syrigos KN, Harrington KJ, Karayiannakis AJ, et al. Clinical signifi cance of heat shock protein-70 expression in bladder cancer. Urology 2003 ; 61 : 677 -80
31. Matzinger P. The danger model: a renewed sense of self. Science 2002 ; 296 : 301 -5
32. Asea A, Kraeft SK, Kurt-Jones EA, et al. HSP70 stimulates cytokine production through a CD14-dependant pathway, demonstrating its dual role as a chaperone and cytokine. Nat Med 2000 ; 6 : 435 -42
33. Wang Y, Kelly CG, Singh M, et al. Stimulation of Th1-polarizing cytokines, C-C chemokines, maturation of dendritic cells, and adjuvant function by the peptide binding fragment of heat shock protein 70. J Immunol 2002 ; 169 : 2422 -9
34. Lehner T, Wang Y, Whittall T, et al. Functional domains of HSP70 stimulate
generation of cytokines and chemokines, maturation of dendritic cells and adjuvanticity. Biochem Soc Trans 2004 ; 32 : 629 -32
35. Moretta A, Bottino C, Vitale M, et al. Activating receptors and coreceptors involved in human natural killer cell-mediated cytolysis. Annu Rev Immunol 2001 ; 19 : 197 -223
36. Long EO. Regulation of immune responses through inhibitory receptors. Annu Rev Immunol 1999 ; 17 : 875 -904
37. Gastpar R, Gehrmann M, Bausero MA, et al. Heat shock protein 70 surface-positive tumor exosomes stimulate migratory and cytolytic activity of natural killer cells. Cancer Res 2005 ; 65 : 5238 -47
38. Chen X, Tao Q, Yu H, et al. Tumor cell membrane-bound heat shock protein 70 elicits antitumor immunity. Immunol Lett 2002 ; 84 : 81 -7
39. Strbo N, Oizumi S, Sotosek-Tokmadzic V, et al. Perforin is required for innate and adaptive immunity induced by heat shock protein gp96. Immunity 2003 ; 18 : 381 -90
40. Gross C, Koelch W, DeMaio A, et al. Cell surface-bound heat shock protein 70 (Hsp70) mediates perforin-independent apoptosis by specifi c binding and uptake of granzyme B. J Biol Chem 2003 ; 278 : 41173 -81
41. Sreedhar AS, Kalmar E, Csermely P, et al. Hsp90 isoforms: functions, expression and clinical importance. FEBS Lett 2004 ; 562 : 11 -5
42. Pratt WB, Galigniana MD, Harrell JM, DeFranco DB. Role of hsp90 and the hsp90-binding immunophilins in signalling protein movement. Cell Signal 2004 ; 16 : 857 -72
43. Neckers L, Ivy SP. Heat shock protein 90. Curr Opin Oncol 2003 ; 15 : 419 -24
44. Neckers L, Neckers K. Heat-shock protein 90 inhibitors as novel cancer chemotherapeutic agents. Expert Opin Emerg Drugs 2002 ; 7 : 277 -88
45. Nimmanapalli R, O’Bryan E, Bhalla K. Geldanamycin and its analogue 17-allylamino-17-demethoxygeldanamycin lowers Bcr-Abl levels and induces apoptosis and differentiation of Bcr-Abl-positive human leukemic blasts. Cancer Res 2001 ; 61 : 1799 -804
46. Pandey P, Saleh A, Nakazawa A, et al. Negative regulation of cytochrome c-mediated oligomerization of Apaf-1 and
activation of procaspase-9 by heat shock protein 90. EMBO J 2000 ; 19 : 4310 -22
47. Cardone MH, Roy N, Stennicke HR, et al. Regulation of cell death protease caspase-9 by phosphorylation. Science 1998 ; 282 : 1318 -21
48. Ozes ON, Mayo LD, Gustin JA, et al. NF-kappaB activation by tumour necrosis factor requires the Akt serine-threonine kinase. Nature 1999 ; 401 : 82 -5
49. Lewis J, Devin A, Miller A, et al. Disruption of hsp90 function results in degradation of the death domain kinase, receptor-interacting protein (RIP), and blockage of tumor necrosis factor-induced nuclear factor-kappaB activation. J Biol Chem 2000 ; 275 : 10519 -26
50. Chen G, Cao P, Goeddel DV. TNF-induced recruitment and activation of the IKK complex require Cdc37 and Hsp90. Mol Cell 2002 ; 9 : 401 -10
51. Bando Y, Katayama T, Kasai K, et al. GRP94 (94 kDa glucose-regulated protein) suppresses ischemic neuronal cell death against ischemia/reperfusion injury. Eur J Neurosci 2003 ; 18 : 829 -40
52. Workman P. Altered states: selectively drugging the Hsp90 cancer chaperone. Trends Mol Med 2004 ; 10 : 47 -51
53. Sun J, Liao JK. Induction of angiogenesis by heat shock protein 90 mediated by protein kinase Akt and endothelial nitric oxide synthase. Arterioscler Thromb Vasc Biol 2004 ; 24 : 2238 -44
54. Pfosser A, Thalgott M, Buttner K, et al. Liposomal Hsp90 cDNA induces neovascularization via nitric oxide in chronic ischemia. Cardiovasc Res 2005 ; 65 : 728 -36
55. Hoang AT, Huang J, Rudra-Ganguly N, et al. A novel association between the human heat shock transcription factor 1 (HSF1) and prostate adenocarcinoma. Am J Pathol 2000 ; 156 : 857 -64
56. Price JT, Quinn JM, Sims NA, et al. The heat shock protein 90 inhibitor, 17-allylamino-17-demethoxygeldanamycin, enhances osteoclast formation and potentiates bone metastasis of a human breast cancer cell line. Cancer Res 2005 ; 65 : 4929 -938
57. Grenert JP, Sullivan WP, Fadden P, et al. The amino-terminal domain of heat shock protein 90 (hsp90) that binds geldanamycin is an ATP/ADP switch domain that regulates hsp90 conformation. J Biol Chem 1997 ; 272 : 23843 -50
Exp
ert O
pin.
Inv
estig
. Dru
gs D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsita
ets-
und
Lan
desb
iblio
thek
Due
ssel
dorf
on
09/1
9/13
For
pers
onal
use
onl
y.

Karapanagiotou, Syrigos & Saif
Expert Opin. Investig. Drugs (2009) 18(2) 173
58. Schulte TW, An WG, Neckers LM. Geldanamycin-induced destabilization of Raf-1 involves the proteasome. Biochem Biophys Res Commun 1997 ; 239 : 655 -9
59. Hostein I, Robertson D, DiStefano F, et al. Inhibition of signal transduction by the Hsp90 inhibitor 17-allylamino-17-demethoxygeldanamycin results in cytostasis and apoptosis. Cancer Res 2001 ; 61 : 4003 -9
60. Supko JG, Hickman RL, Grever MR, et al. Preclinical pharmacologic evaluation of geldanamycin as an antitumor agent. Cancer Chemother Pharmacol 1995 ; 36 : 305 -15
61. Goetz MP, Toft D, Reid J, et al. Phase I trial of 17-allylamino-17-demethoxygeldanamycin in patients with advanced cancer. J Clin Oncol 2005 ; 23 : 1078 -87
62. Banerji U, O’Donnell A, Scurr M, et al. Phase I pharmacokinetic and pharmacodynamic study of 17-allylamino, 17-demethoxygeldanamycin in patients with advanced malignancies. J Clin Oncol 2005 ; 23 : 4152 -61
63. Grem JL, Morrison G, Guo XD, et al. Phase I and pharmacologic study of 17-(allylamino)-17-demethoxygeldanamycin in adult patients with solid tumors. J Clin Oncol 2005 ; 23 : 1885 -93
64. Ramanathan RK, Belani CP, Friedland D, et al. Phase I study (twice weekly schedule) of 17-allylamino-17 demethoxygeldanamycin (17-AAG, NSC-704057) in patients with advanced refractory tumours [abstract 3050]. ASCO Meetings Abstracts; 2005
65. Musquire LA, Ramalingam S, Egorin MJ, et al. Phase I and pharmacokinetic (PK) study of 17-allylamino-17 demethoxygeldanamycin (17-combination with weekly paclitaxel for advanced solid mAAG) in alignancies. 2007 ASCO Annual Meeting Proceedings (Post-Meeting Edition). J Clin Oncol 2007 ; 25 (18 Suppl)
66. Solit DB, Egorin M, Valentin G, et al. Phase 1 pharmacokinetic and pharmacodynamic trial of docetaxel and 17AAG (17-allylamino-17-demethoxygeldanamycin) [abstract 3032]. ASCO Meetings Abstracts; 2004
67. Haluska P, Toft DO, Steinmetz SM, et al. A phase I trial of gemcitabine (Gem), 17-allylaminogeldanamycin (17-AAG) and cisplatin (CDDP) in solid tumor patients [abstract 3058]. 2004 ASCO Annual
Meeting Proceedings (Post-Meeting Edition). J Clin Oncol 2004 ; 22 (14 Suppl)
68. A Phase II Trial of 17-N-Allylamino-17-Demethoxygeldanamycin (17-AAG) in Combination with Gemcitabine in Patients with Metastatic Pancreatic Adenocarcinoma. clinicaltrials.gov2008.
69. Heath EI, Hillman D, Vaishampayan U, et al. A phase II trial of 17-allylamino-17-demethoxygeldanamycin (17-AAG) in patients with hormone-refractory metastatic prostate cancerE. I [abstract 15553]. 2007 ASCO Annual Meeting Proceedings (Post-Meeting Edition). J Clin Oncol 2007 ; 25 (18 Suppl)
70. Modi S, Stopeck AT, Gordon MS, et al. Combination of trastuzumab and tanespimycin (17-AAG, KOS-953) is safe and active in trastuzumab-refractory HER-2 overexpressing breast cancer: a phase I dose-escalation study. J Clin Oncol 2007 ; 25 : 5410 -7
71. Modi S, Sugarman S, Stopeck A, et al. Phase II trial of the Hsp90 inhibitor tanespimycin (Tan) + trastuzumab (T) in patients (pts) with HER2-positive metastatic breast cancer (MBC) [abstract 1027]. J Clin Oncol 2008 ; 26
72. Saif MW, Erlichman C, Dragovich T, et al. Phase I study of CNF1010 (lipid formulation of 17-(allylamino)-17-demethoxygeldanamycin: 17-AAG) in patients with advanced solid tumors [abstract10062]. ASCO Annual Meeting. 2006 ASCO Annual Meeting Proceedings. J Clin Oncol 2006 ; 24 (18 Suppl)
73. Flaherty KT, Gore L, Avadhani A, et al. Phase 1, pharmacokinetic (PK) and pharmacodynamic (PD) study of oral alvespimycin (A; KOS-1022; 17-DMAG): Two different schedules in patients with advanced malignancies [abstract140]. ASCO Annual Meeting Proceedings. J Clin Oncol 2007 ; 25 (18 Suppl)
74. Wagner AJ, Morgan JA, Chugh R, et al. Inhibition of heat shock protein 90 (Hsp90) with the novel agent IPI-in metastatic GIST following failure of tyrosine kinase inhibitors (TKIs) or other sarcomas: Clinical results from phase I trial [abstract 10503]. ASCO Anual Meeting. J Clin Oncol 2008 ; 26
75. Demetri GD, George S, Morgan JA, et al. Inhibition of the Heat Shock Protein 90 (Hsp90) chaperone with the novel agent IPI-504 to overcome resistance to tyrosine kinase inhibitors (TKIs) in metastatic
GIST: Updated results of a phase I trial [abstract 10024]. ASCO Annual Meeting Proceedings. J Clin Oncol 2007 ; 25 (18 Suppl)
76. A Phase 2, Open-Label, Single-Arm, Multicenter Study Evaluating the Safety and Antitumor Activity of IPI-504, A Novel Small Molecule Inhibitor of Heat Shock Protein 90 (HSP90), in Patients With Metastatic Melanoma. clinicaltrials@gov2008. Ref Type: Electronic Citation
77. Bryson JC, Infante JR, Ramanathan RK, et al. A Phase 1 dose-escalation study of the safety and pharmacokinetics (PK) of the oral Hsp90 inhibitor SNX-5422 [abstract 14613]. J Clin Oncol 2008 ; 26
78. STA 9090 clinicaltrials@gov. 2008. Ref Type: Electronic Citation
79. Elfi ky A, Saif MW, Beeram M, et al. BIIB021, an oral, synthetic non-ansamycin Hsp90 inhibitor: Phase I experience [abstract2503]. ASCO Annual Meeting. J Clin Oncol 2008 ; 26
80. Wood C, Srivastava P, Bukowski R, et al. An adjuvant autologous therapeutic vaccine (HSPPC-96; vitespen) versus observation alone for patients at high risk of recurrence after nephrectomy for renal cell carcinoma: a multicentre, open-label, randomised phase III trial. Lancet 2008 ; 372 : 145 -54
81. Jonasch E, Wood C, Tamboli P, et al. Vaccination of metastatic renal cell carcinoma patients with autologous tumour-derived vitespen vaccine: clinical fi ndings. Br J Cancer 2008 ; 98 : 1336 -41
82. Testori A, Richards J, Whitman E, et al. Phase III comparison of vitespen, an autologous tumor-derived heat shock protein gp96 peptide complex vaccine, with physician’s choice of treatment for stage IV melanoma: the C-100-21 Study Group. J Clin Oncol 2008 ; 26 : 955 -62
83. Pilla L, Patuzzo R, Rivoltini L, et al. A phase II trial of vaccination with autologous, tumor-derived heat-shock protein peptide complexes Gp96, in combination with GM-CSF and interferon-alpha in metastatic melanoma patients. Cancer Immunol Immunother 2006 ; 55 : 958 -68
84. Richards J, Testori A, Whitman E, et al. Autologous tumor-derived HSPPC-96 vs. physician’s choice (PC) in a randomized phase III trial in stage IV melanoma [abstract 8002]. ASCO Annual Meeting. J Clin Oncol 2006 ; 24 (18 Suppl)
Exp
ert O
pin.
Inv
estig
. Dru
gs D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsita
ets-
und
Lan
desb
iblio
thek
Due
ssel
dorf
on
09/1
9/13
For
pers
onal
use
onl
y.

Heat shock protein inhibitors and vaccines as new agents in cancer treatment
174 Expert Opin. Investig. Drugs (2009) 18(2)
85. Hertkorn C, Lehr A, Woelfel T, et al. Phase I trial of vaccination with autologous tumor-derived gp96 (oncophage) in patients after surgery for gastric cancer [abstract 117]. ASCO Annual Meeting 21; 2002
86. Mazzafero V, Coppa JC, Carrabba MG, et al. Vaccination with autologous tumor derived heat-shock protein peptide complex-96 (Oncophage ® ) after curative resection of colorectal liver metastases [abstract 2290]. ASCO Annual Meeting 21; 2002
87. Manjili MH, Henderson R, Wang XY, et al. Development of a recombinant HSP110-HER-2/neu vaccine using the chaperoning properties of HSP110. Cancer Res 2002 ; 62 : 1737 -42
88. Manjili MH, Wang XY, Chen X, et al. HSP110-HER2/neu chaperone complex vaccine induces protective immunity against spontaneous mammary tumors in HER-2/neu transgenic mice. J Immunol 2003 ; 171 : 4054 -61
Affi liation Eleni M Karapanagiotou 1 , Kostas Syrigos 1 & Muhammad Wasif Saif † 2 MD MBBS † Author for correspondence 1 Athens Medical School, Medical Oncology, Athens, Greece 2 Professor Yale University School of Medicine, Medical Oncology, 333 Cedar Street, FMP: 116, New Haven, CT 06520, USATel: +1 2037371569 ; Fax: +1 2037853788 ;E-mail: [email protected]
Exp
ert O
pin.
Inv
estig
. Dru
gs D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsita
ets-
und
Lan
desb
iblio
thek
Due
ssel
dorf
on
09/1
9/13
For
pers
onal
use
onl
y.





























