Embed Size (px)
Citation preview

Heat Acclimation and Heat Stress Have Different Effects on Cholinergic
Muscarinic Receptors PAVEL KASPLER AND MICHAL HOROWITZ"
Division of Physiology Hadassah Schools of Dental Medicine and Medicine
The Hebrew University POB 12272
Jerusalem 91 120, Israel
BIPHASIC ALTERATIONS IN CELLULAR AND INTEGRATIVE RESPONSES DURING HEAT ACCLIMATION
Previous studies from our laboratory on the dynamics of heat acclimation provided evidence that an integral part of an animal's acclimation repertoire is biphasic alterations in cellular responses. These biphasic alterations coincide with a biphasic pattern of changes in the regulation of the heat dissipation effectors.]q2 For example, in the evaporative cooling system of the rat (submaxillary salivary gland) decreased glandular responsiveness to neurotransmitters at the initial phases of heat acclimation is replaced in later phases of the acclimation process by increased glandular responsiveness. Concomitantly, the rate of excitability of the autonomic outflow innervating the gland increased and decreased in the early and late phases of heat acclimation re~pectively.~
Our data indicated that changes in several steps along the cholinergic musca- rinic signal transduction pathway for water secretion are responsible for the bipha- sic altered glandular responsiveness ob~erved . ' .~ Causal relationships between changes in muscarinic receptor density and affinity and the altered glandular responsiveness suggest that membranal receptors are the initial mediators in the decreasedhncreased effector organ responsiveness. Additional intracellular alter- ations apparently amplify these effects. We hypothesize that upon heat stress in acclimating animals these concerted temporal peripheral changes modify the preacclimation rate of autonomic excitation of this effectorial loop in an opposite manner as illustrated in FIGURE 1.
AUTONOMIC VS MEMBRANALICELLULAR COMPENSATORY STRATEGIES
The postsynaptic desensitization at the initiation of heat acclimation coincides with two membranal events, muscarinic receptor (MR) upregulation and musca- rinic binding affinity downregulation. The observed MR upregulation, in the face of persistent accelerated excitation, contradicts the acknowledged concept of the
Corresponding author. Fax: 972 2 439736; E-mail: [email protected]. 620

KASPLER & HOROWITZ: CHOLINERGIC MUSCARINIC RECEPTORS 621
HEAT ACCLIMATION
SHORT TERM LONG TERM
AUTONOMIC EXCITATION
PERIPHERAL RESPONSE
FIGURE 1. Central peripheral interaction during heat acclimation. During short-term heat acclimation an abrupt rise in body temperature leads to impaired membrane function and autonomic accelerated excitation. During long-term heat acclimation increased effector effi- ciency reduces the need for accelerated autonomic discharge. For further details see text.
mechanisms underlying receptor upregulation (Le., denervation). On the contrary, prolonged exposure to the transmitter leads to desensitization which is usually explained by either receptor downregulation or changes in receptor-G protein in- teraction.
The occurrence of decreased binding affinity in response to heat acclimation was measured in atrial and intestinal muscarinic receptors, as well as in cardiac /3-adrenoreceptors.' This implies a non organ-selective membranal response.
One reason for the modulation of receptor affinity is changes in the cellular membrane lipid environment. Membrane lipids form an important solvent compart- ment for membrane bound proteins and thus changes in their physical state or composition may modulate their activity. Both a rise in body temperature and thermal acclimation have been reported to affect the lipid environment of the cell membrane.6 This, together with the facts that 1. initiation of heat acclimation is usually accompanied by an abrupt rise in body temperature, and 2. resting temperature in long-term heat acclimation is 1 "C higher than the preacclimation state,' suggest that temperature is the likely candidate to affect membrane proper- ties of the acclimating animal.
In contrast to receptor affinity, receptor upregulation may be controlled by processes other than changes in membrane properties ( e .g . , protein synthesis). Thus, our findings of increased receptor density at the early phase of heat acclima- tion can be attributed to a cellular compensatory response to a handicapped cellular secretory signal.
We may conclude that at the initiation of heat acclimation, avariety of processes having opposing effects interact. While thermal demands lead to accelerated persis- tent autonomic stimulation and in turn desensitization, the strained cell compen- sates for decreased affinity by receptor upregulation. Hence, the overall mem- brand response is determined by the sum of these opposing effects. In contrast to the short-term, transient events occurring upon short-term heat acclimation (STHA), following long-term acclimation the cumulative effect of moderate heat

622 ANNALS NEW YORK ACADEMY OF SCIENCES
leads to increased MR density with preacclimation binding affinity; this may involve different mechanisms.
DIFFERENTIATION BETWEEN INTRACELLULAR AND
AN EXPERIMENTAL MODEL EXTRACELLULAR INDUCED RESPONSES-
To investigate the interaction between intracellular and extracellular signals on the membranal muscarinic receptors we recently developed a new experimental model: salivary acinar cell line (HSY) undergoing simulated acclimating condi- tions. A conventional thermotolerance test (number of cells survived following heat stress of 45°C) validated the attainment of acclimation in these cells (Kaspler, Moran & Horowitz, in preparation). The study in parallel of salivary cells under- going simulated acclimating conditions and cells originating from animals subjected to heat acclimation provides a convenient experimental paradigm allowing the differentiation of intra- and extracellularly induced processes.
In this report, several examples of the data obtained are discussed. In particular, the specificity of acclimation response (vs heat stress) and evidence for a role of the direct effect of heat on the cells following short-term heat acclimation will be discussed.
CHRONIC HEAT AND MEMBRANAL CHANGES
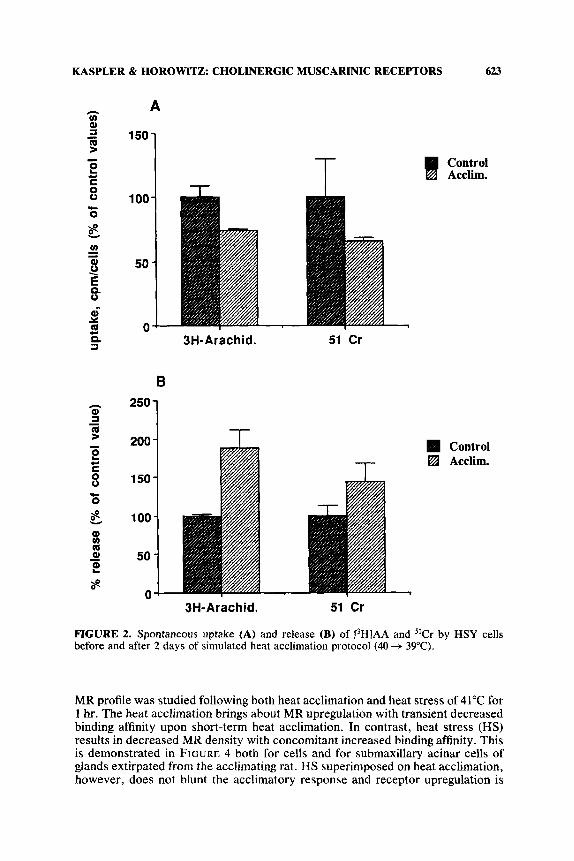
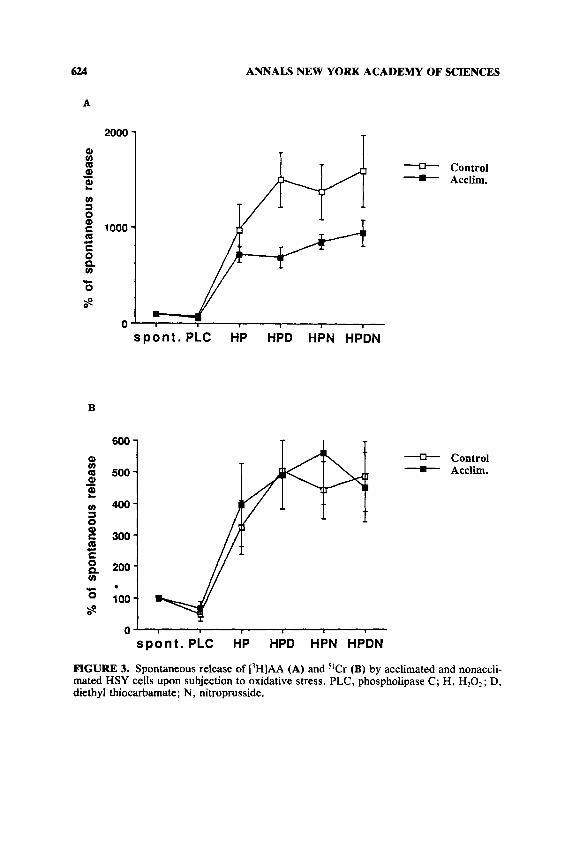
In light of the findings suggesting alteration in cell membrane as the first mediators of heat signal,' the incorporation of arachidonic acid (AA), an important component of the cell membrane, into the membrane and membrane permeability were measured using labeled [3H]AA and W r as membranal markers, respectively, upon subjection to a membranal stressor e.g. , oxidative stress, heat stress.* Mem- brane resistance to oxidative stress was assessed by subjection of the intact cells to a cocktail of a subtoxic level of prooxidant and H202. Experiments were carried out on HSY cells before and after two days of heat acclimation (Kaspler, Ginsburg & Horowitz, in preparation). The data (FIG. 2 ) show that the uptake of [3H]AA by the heat-acclimated cells was significantly lower than by the nonacclimated cells. Concomitantly, spontaneous release of [3H]AA was significantly greater (by 80%, FIG. 2 ) . "Cr showed a similar response. This significant change in HSY membrane properties was also observed upon subjection to oxidative stress. Accli- mated HSY cells released less [3H]AA than the nonacclimated cells (by 36%), although membrane permeability, as manifested by 51Cr release, was similar for the acclimated and nonacclimated cells (FIG. 3). Collectively, the data confirm that short-term acclimation produces significant changes in the cell membrane. We hypothesize that differences in AA metabolism may stem from changes in phospholipase A, activity or from changes in acylation rate.8 These changes are currently under investigation.
MUSCARINIC RECEPTOR PROFILE
Muscarinic receptor density and binding affinity were measured using [3H]N- methylscopolamine ([3H]NMS). To assess the specificity of heat acclimation the
KASPLER & HOROWITZ CHOLlNERGIC MUSCARINIC RECEPTORS 623
A
1501
I T T Control Acclim.
100
50
0 3H- Arachid. 51 Cr
B 250 -
200 -
150-
100-
50 -
-r Control
H Acclim.
0 3H-Arachid. 51 Cr
FIGURE 2. Spontaneous uptake (A) and release (B) of [3H]AA and "Cr by HSY cells before and after 2 days of simulated heat acclimation protocol (40 -+ 39°C).
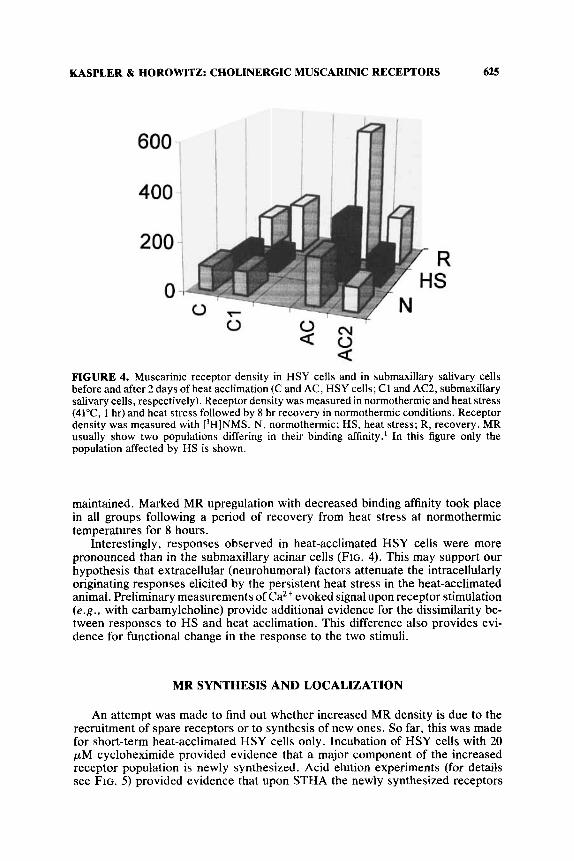
MR profile was studied following both heat acclimation and heat stress of 41°C for 1 hr. The heat acclimation brings about MR upregulation with transient decreased binding affinity upon short-term heat acclimation. In contrast, heat stress (HS) results in decreased MR density with concomitant increased binding affinity. This is demonstrated in FIGURE 4 both for cells and for submaxillary acinar cells of glands extirpated from the acclimating rat. HS superimposed on heat acclimation, however, does not blunt the acclimatory response and receptor upregulation is
624
A
2000
v) 3 0 g 1000 U C 0 % c 0
s 0
B
ANNALS NEW YORK ACADEMY OF SCIENCES
u- --t-
Control AccIim.
spont. PLC HP HPD HPN HPDN
v)
0 Q) E m C 0 P v)
a
c
c 0
s
600 - 500 -
400 -
300 -
200 -
100 -
Control Acclii.
spont. PLC HP HPD HPN HPDN
FIGURE 3. Spontaneous release of [3H]AA (A) and "Cr (B) by acclimated and nonaccli- mated HSY cells upon subjection to oxidative stress. PLC, phospholipase C; H, H,O,; D, diethyl thiocarbamate; N, nitroprusside.
KASPLER & HOROWITZ: CHOLINERGIC MUSCARINIC RECEPTORS 625
FIGURE 4. Muscarinic receptor density in HSY cells and in submaxillary salivary cells before and after 2 days of heat acclimation (C and AC, HSY cells; C1 and AC2, submaxillary salivary cells, respectively). Receptor density was measured in normothermic and heat stress (41°C. 1 hr) and heat stress followed by 8 hr recovery in normothermic conditions. Receptor density was measured with ['HINMS. N, normothermic; HS, heat stress; R, recovery. MR usually show two populations differing in their binding affinity.' In this figure only the population affected by HS is shown.
maintained. Marked MR upregulation with decreased binding affinity took place in all groups following a period of recovery from heat stress at normothermic temperatures for 8 hours.
Interestingly, responses observed in heat-acclimated HSY cells were more pronounced than in the submaxillary acinar cells (FIG. 4). This may support our hypothesis that extracellular (neurohumoral) factors attenuate the intracellularly originating responses elicited by the persistent heat stress in the heat-acclimated animal. Preliminary measurements of Ca2+ evoked signal upon receptor stimulation ( e . g . , with carbamylcholine) provide additional evidence for the dissimilarity be- tween responses to HS and heat acclimation. This difference also provides evi- dence for functional change in the response to the two stimuli.
MR SYNTHESIS AND LOCALIZATION
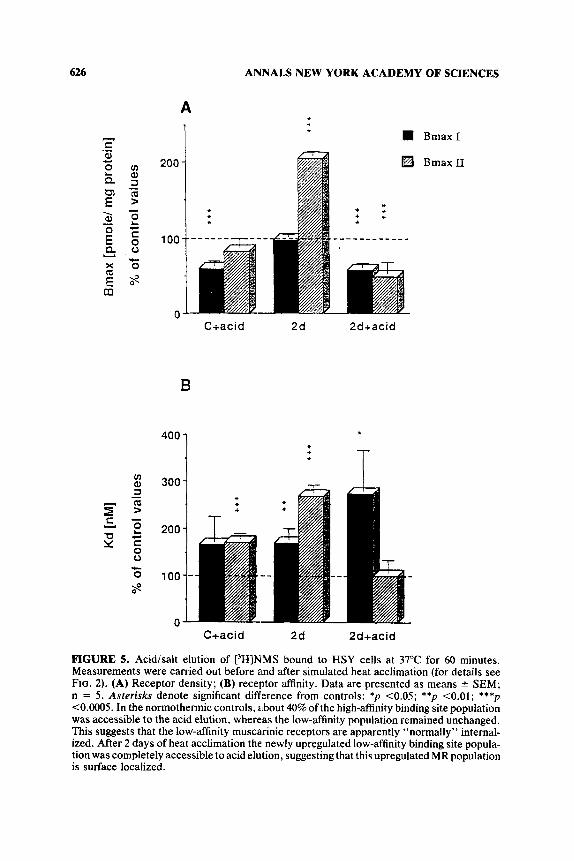
An attempt was made to find out whether increased MR density is due to the recruitment of spare receptors or to synthesis of new ones. So far, this was made for short-term heat-acclimated HSY cells only. Incubation of HSY cells with 20 pM cycloheximide provided evidence that a major component of the increased receptor population is newly synthesized. Acid elution experiments (for details see FIG. 5) provided evidence that upon STHA the newly synthesized receptors
626 ANNALS NEW YORK ACADEMY OF SCIENCES
A
BmaxI
BmaxII
1 *
C+acid 2d 2d+acid
B
4 7
*
.b T
C+acid 2d 2d+acid
FIGURE 5. Acidisalt elution of t3H]NMS bound to HSY cells at 37°C for 60 minutes. Measurements were carried out before and after simulated heat acclimation (for details see FIG. 2). (A) Receptor density; (B) receptor affinity. Data are presented as means ? SEM; n = 5 . Asterisks denote significant difference from controls: *p C0.05; **p <0.01; ***p <0.0005. In the normothermic controls, about 40% of the high-affinity binding site population was accessible to the acid elution, whereas the low-affinity population remained unchanged. This suggests that the low-affinity muscarinic receptors are apparently "normally" internal- ized. After 2 days of heat acclimation the newly upregulated low-affinity binding site popula- tion was completely accessible to acid elution, suggesting that this upregulated MR population is surface localized.

KASPLER & HOROWITZ: CHOLINERGIC MUSCARINIC RECEPTORS 627
are localized on the surface of the cell membrane. This may suggest that these are active receptors.
SUMMARY AND CONCLUSIONS
In this brief report we have proved that membrand changes take place upon short-term heat acclimation. These changes may switch on a cascade of transient acclimatory compensatory responses as well as long-term processes. The changes observed in the MR profile are heat acclimation specific, and differ from those observed upon heat stress. These bring about functional changes in the signal transduction pathway for water secretion in the submaxillary salivary gland.
A recovery period from heat stress, similar to STHA, leads to MR upregulation and decreased binding affinity. This response, however, is more pronounced than that observed upon STHA. The functional importance of the phenomenon is unclear. However, it could designate a novel component of the heat shock re- sponse.
REFERENCES
1. HOROWITZ, M., P. KASPLER, Y. MARMARY & Y. ORON. 1966. Evidence for contribution of effector organ cellular responses to the biphasic dynamics of heat acclimation. J. Appl. Physiol. 80(1): 77-86.
HOROWITZ, M., D. ARGOV & R. MIZRAHI. 1983. Interrelationships between heat acclima- tion and salivary cooling mechanism in conscious rats. Comp. Biochem. Physiol. 7 4 A 945-949.
HOROWITZ, M. & U. MEIRI. 1985. Altered responsiveness to parasympathetic activation of submaxillary salivary gland in the heat acclimated rat. Comp. Biochem. Physiol.
HOROWITZ, M. 1994. Heat stress and heat acclimation: the cellular response modifier of autonomic control. I n Integrative and Cellular Aspects of Autonomic Function. K. Pleschka & R. Gerstberger, Eds. 87-97. John Libby & Co. London.
5 . HOROWITZ, M. & U. MEIRI. 1993. Central and peripheral contributions to control of heart rate during heat acclimation. Pfluegers Arch. 422 386-392.
6. COSSINS, A. R. & K. BOWLER. 1987. Temperature biology of Animals. Chapman and Hall.
7. HOROWITZ, M. 1976. Acclimatization of rats to mild heat: Body water distribution and adaptability of submaxillary salivary gland. Muegers Arch. 366: 173-176.
8. GINSBURG, I. &. R. KOHEN. 1995. Cell damage in inflammatory and infectious sites might involve a coordinated “cross-talk’’ among oxidant, microbial haemolysins and ampiphiles, cationic proteins, phospholipases, fatty acids, proteinases and cytokines (an overview). Free Radical Res. 2 2 489-517.
2.
3.
80A 57-60. 4.





























