Embed Size (px)
Citation preview

INTRODUCTION
• HDX is often used to probe protein structure and dynamicsusing a variety of detection techniques (NMR, ESI MS, etc.)
• HDX is a complex process whose kinetics depends on bothprotein dynamics and intrinsic amide hydrogen exchange rates
• Two distinct HDX regimes (EX1 and EX2) are commonlyidentified depending on the rates of protein refolding (kcl) andintrinsic exchange (kint)
• HDX under native conditions almost always follows the EX2kinetics
• It is often assumed that denaturing conditions favor the EX1regime (unless kint is reduced under such conditions evenmore than kcl)

k1 k2 k-1
EX1: kint>>kcl: kHDX=kcl
k1 k2 k-1
EX2: kcl>>kint: kHDX=kintKunfold

• All of the above formalism has been initially developed tointerpret results of HDX NMR experiments and kcl refers to re-protection of individual hydrogen atoms, not collective motions
• When this formalism is applied to interpret the results of HDXESI (or MALDI) MS experiments, an implicit assumption ismade that kcl refers to a collective motion (e.g., transition froman unprotected state to a structured conformation)
• Most proteins, however, are not simple two-state systemsand transitions within different pairs of states may result indifferent HDX kinetics
• In order to interpret the results of HDX MS experimentcorrectly, better understanding is needed on how complexHDX kinetics reflects protein dynamics and structure

CASE STUDY 1: A TWO-STATE PROTEIN(HDX kinetics of Chymotrypsin Inhibitor II)
• Chymotrypsin Inhibitor II (CI2) is asmall, single domain protein• CI2 folds in a simple two-stateprocess under native conditions• Non-native partially structuredstates of CI2 (equilibriumintermediates) have been detectedonly under extreme conditions [1]• Only two states (Native andUnfolded) can be populated underthe conditions used in this work [2]• The vast body of knowledge on CI2dynamics (NMR, time-resolvedspectroscopy, etc.) makes it anattractive model protein

HDX under native conditions: Two-Phase EX2 Kinetics
HDX was monitored across a wide range of solutiontemperatures. It follows a simple biphasic kinetics up to 45oC.Only one phase can be confidently measured at 60oC.

• The two phases correspond to ca. 15 fast exchanging amides(often assigned to local fluctuations) and 30-41 slowlyexchanging amides (often assigned to global unfolding)• The two rate constants can be used to calculate ∆G values ofthese two processes• Since ∆Gslow corresponds to a N → U transition, it can be usedto estimate free energy of protein folding• Calculated ∆Gslow value is consistent with previous non-MSdata
Technique used ∆Gslow (room T)___________________________________________________
fluorescence(GdmCl denaturation) 7.03±0.16fluorescence(thermal denaturation) 7.18±0.43HDX NMR (slow) 7.6HDX MS (slow) 7.1___________________________________________________
HDX NMR (fast) 4.95HDX MS (fast) 4.6

Minimalistic energy surface diagram for a two-state proteinMonte-Carlo simulation of a particle movement on a 2-D energy surface
Infrequent events ofglobal unfolding (particleleaves the potential wellfor short periods of time)
Frequent events of local fluctuations(particle climbs up the wall of the potentialwell and reaches the “increased flexibilityregion” for short periods of time)

• Elevation of free energy above the ground state by ∆Gfastresults in some increase of the chain flexibility, although theprotein remains in the potential well of the Native state
• Elevation of free energy above the ground state by ∆Gslowresults in a transition to an Unfolded state, providingsignificantly more conformational freedom (random coil)
• Protein excursions from the bottom of the potential well arevery short, since the reverse activation energy barriers areeither small (U→N) transition or non-existent (localfluctuations)
• Such behavior leads to EX2 type exchange kinetics, sincekcl>>kint for both processes

HDX under mildly denaturing conditions: Mixed EX1/EX2 Kinetics

• The EX2 component of HDXkinetics appear to reflect rare andshort global unfolding events (verylarge apparent ∆G; accurateestimations are difficult due tounavailable data on kint in thepresence of MeOH).
• The EX1 component of HDXkinetics reflects “prolonged” globalunfolding events (proteins becometrapped in the U-state for extendedperiods of time). The kinetic data canbe used to evaluate activation energyof protein unfolding
kHDX= kop= 0.04 min-1
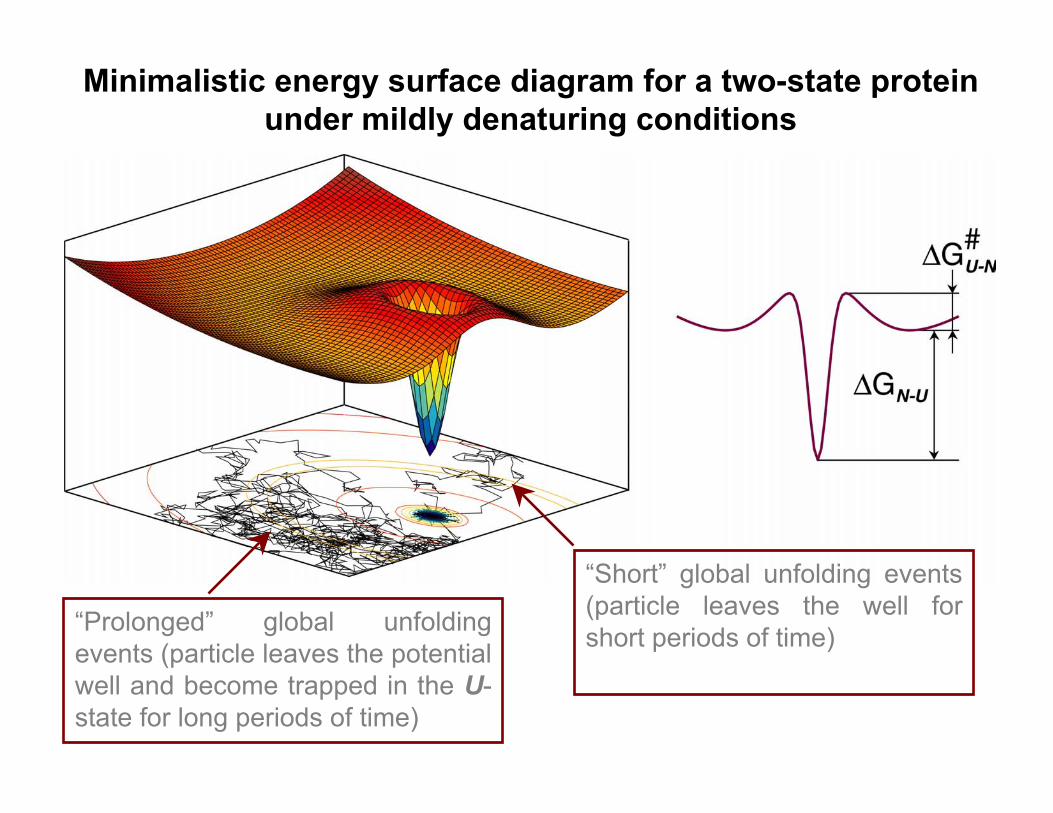
Minimalistic energy surface diagram for a two-state proteinunder mildly denaturing conditions
“Prolonged” global unfoldingevents (particle leaves the potentialwell and become trapped in the U-state for long periods of time)
“Short” global unfolding events(particle leaves the well forshort periods of time)
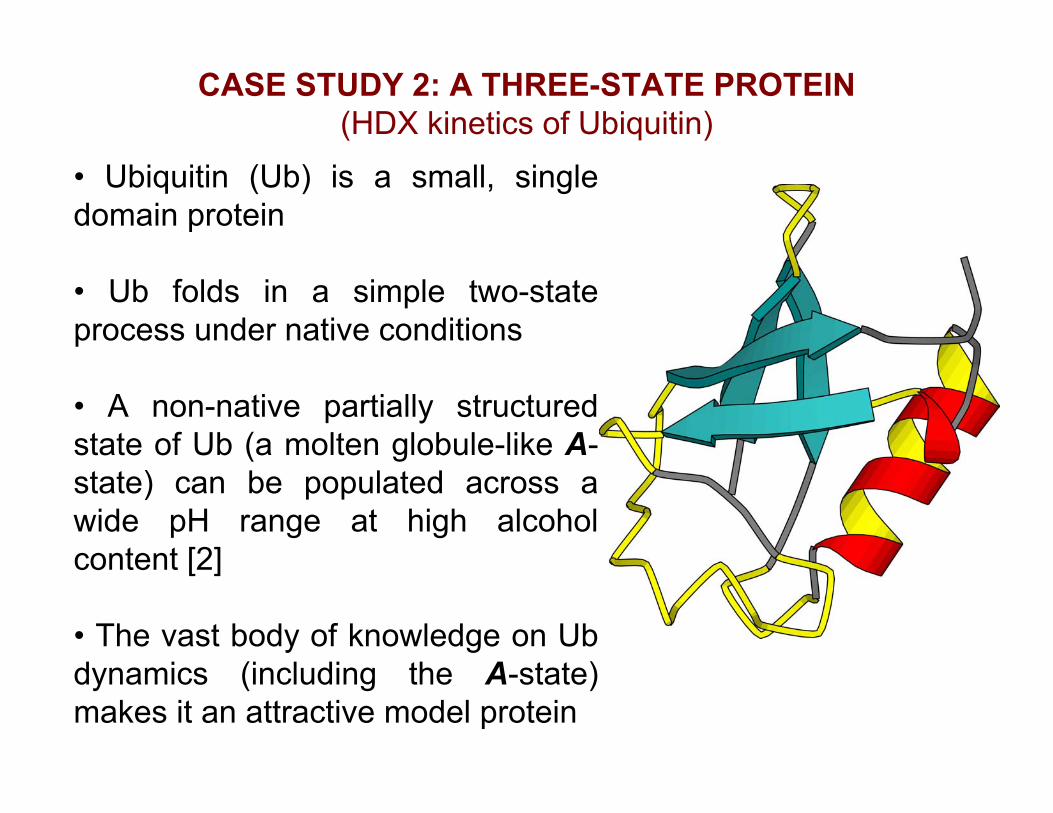
CASE STUDY 2: A THREE-STATE PROTEIN(HDX kinetics of Ubiquitin)
• Ubiquitin (Ub) is a small, singledomain protein
• Ub folds in a simple two-stateprocess under native conditions
• A non-native partially structuredstate of Ub (a molten globule-like A-state) can be populated across awide pH range at high alcoholcontent [2]
• The vast body of knowledge on Ubdynamics (including the A-state)makes it an attractive model protein

HDX under native conditions: Two-phase EX2 kinetics
fast phase,ΔGfast=3.1 kcal/mol
slow phase,ΔGslow=5.4 kcal/mol
HDX follows a simple biphasic kinetics at room temperature.Calculated free energy of protein folding (ΔGslow) agrees withearlier denaturation measurements (ΔG=5.0 kcal/mol, [5])

Minimalistic energy surface diagram for a three-state proteinMonte-Carlo simulation of a particle movement on a 2-D energy surface
Infrequent events of globalunfolding (particle leaves thepotential well for short periods oftime)Frequent N→A transitions (low reverse
activation energy barrier ∆G‡A-N
prevents particle trapping in the A-statefor long periods of time)

HDX of Ub under mildly denaturing conditions
• N → A: mixed EX1/EX2• N, A → U: EX2

Minimalistic energy surface diagram for a three-state proteinunder mildly denaturing conditions
Both “prolonged” and short conformational switches(long- and short-term excursions of the particle fromthe potential well N to A-basin of attraction)
“Short” global unfolding events(particle stays outside of both N-and A-wells for short periods oftime) eventually returning to eitherN- or A-basin of attraction

CONCLUSIONS
• Protein amide HDX reactions have very convoluted kineticswhich is reflective of multiple dynamic process
• HDX of small model proteins under native conditions revealstwo major types of dynamic events,fast local fluctuation andslow global unfolding
• Free energies of both of these processes calculated basedon the kinetic data are consistent with that derived from othersources

• Despite extensive efforts, no “pure” EX1 exchangekinetics was ever observed even for small model proteins
• Mixed EX1/EX2 HDX kinetics of a two-state proteinreflects multiplicity of transitions between the native stateand the random coil
• Analysis of such kinetics yields valuable information onthe energetics of transitions between the native state andthe random coil
• Mixed EX1/EX2 HDX kinetics of a three-state proteinalso reflects presence of multiple transitions between thepairs of states (native state to non-native intermediate,native state to random coil, etc.)

REFERENCES1. Silow, M., Oliveberg, M. J. Mol. Biol. 2003, 326, 263.2. Mohimen A., Hoerner, J., Dobo, A., Kaltashov, I.A. Anal.
Chem. 2003, in press.3. Itzhaki, L.S.; Neira, J.L.; Fersht, A. R. J. Mol. Biol. 1997,
270, 89.4. Jackson, S.E.; el Masry, N.; Fersht, A.R. Biochemistry
1993, 32, 11270.5. Vijay-Kumar, S., Bugg, C.E., Cook, W.J. J. Mol. Biol.
1987, 194, 531.
ACKNOWLEDGEMENTSNIH R01 GM61666
Dr. Andre Melcuk (UMass)

OVERVIEWPURPOSEGain better understanding of how protein dynamics is reflectedby hydrogen exchange kinetics under various conditions.
METHODFT ICR MSAmide Hydrogen Exchange (HDX)
RESULTS• Protein amide HDX reactions have very convoluted kineticswhich is reflective of multiple dynamic process occurring onvarious time scales within the protein
• Despite its extreme complexity, HDX can be used to mapprotein energy surfaces and obtain quantitative information onthe energetics of protein structural dynamics





























