Embed Size (px)
Citation preview

Zoologisches Institut der Universitat Erlangen-Nurnberg, Germany
A possible evolutionary pathway to insect flight starting from lepismatid organization
I. HASENFUSS
Abstract
Starting from the hypothesis that flight in Pterygota evolved from lepismatid organization of their ancestors, the functional anatomy of the thoraxwas studied in Lepisma saccharina Linnaeus, 1758, and a Ctenolepisma sp. in regard to both the adaptations to the adaptive zone of Lepismatidaeand to pre-adaptations for the evolution of Pterygota. Well-preserved parts of three subcoxal leg segments were found in the pleural zoneparticipating in leg movement. The lepismatid strategy of escaping predators by running fast and hiding in narrow flat retreats led to a dorso-ventrally flattened body which enabled gliding effects when dropped, followed by flight on the ground. The presumed exploitation of soft tissue atthe tips of low growing Devonian vascular plants opened a canalized pathway to the evolution of the flying ability. Locomotion to another plantwas facilitated by dropping. It is possible that threat by spider-like predators favoured falling and gliding as escape reactions by selection. Fallingexperiments with ‘lepismatid’ models revealed a narrow ‘window’ for gliding, with optimum dimensions of 8 mm body length and 8 mg weight.An equation was derived which describes the glide distance as function of weight, area of the horizontal outline, the specific glide efficiency of thebody, and a non-linear function of the falling height. Improved gliding was made possible by enlarging thoracic paratergites into broad wing-likeextensions of light-weight organization. The disadvantage of the lateral lobes for locomotion on the ground could be minimized by tilting themvertically when running and horizontally when gliding. This movability could be attained by the intercalation of a membranous strip betweentergite and paratergite and the utilization of the pre-existing muscular system and the articulation between the two most basal subcoxal sclerites asa pivot. The dorsal part of the most basal subcoxa was thus integrated into the wing. Initiation of active flight was possible by flappingmovements during gliding. Morphological, ontogenetic and ecological aspects of the origin of Pterygota are discussed.
Key words: functional anatomy – locomotion – evolution – flight – Lepismatoidea – Pterygota – regeneration
Introduction
Evolution, apart from phenomena of neutral evolution, can be
regarded as a succession of adaptive solutions to problemswith which consecutive generations of organisms were con-fronted. The solutions are always based on the previously
achieved organization, including the ontogenetic potentialitiesof varying the organizational features. The adaptations arerealized by alterations which are continuously favoured by
selection within the limitations of environmental and organ-izational constraints. The correct reconstruction of the mainaspects of evolutionary events depends largely on informationabout the ancestral precursor organization, especially on
insight into its complex structural and functional relations.There is a good chance of adequate reconstruction if theplesiomorphic organization is well maintained in cladistically
related organisms. The corresponding traits of these can thenbe reliably assigned to the ancestor in question.There are no doubts that the cladistically nearest relatives of
Pterygota are the Zygentoma as represented by the extantLepismatoidea (see Hennig 1969; Kristensen 1975, 1981). Thelepismatid organization is formed by the corresponding
adaptive zone and according to Hennig’s ‘Deviationsregel’(Hennig 1953) it is probable that the preflight ancestor ofPterygota was likewise of this organization and inhabited thesame adaptive zone. This hypothesis is especially justified if
traits of this organization can be regarded as pre-adaptive forthe evolution of flight and if one or other of the pterygotanfeatures can be traced back to specific lepismatid adaptations.
The conclusion is almost inevitable, if the Zygentoma areparaphyletic in terms of Pterygota as taken into account byBoudreaux (1979) and Staniczek (2000). The relict Tricholepi-
dion gertschi Wygodzinsky (1961) (Lepidotrichidae) has main-tained the ligament between the transversal mandibularmuscles typical for the basal insect lineages, whereas in both
the other Zygentoma and Pterygota the ligament has disap-
peared and the corresponding muscle insertions shifted partlyto the anterior tentorial arms and partly to the hypopharynx(see Staniczek 2000 for discussion). Recently, Furst von Lieven
(2000) and Staniczek (2000) found that the larvae of Epheme-roptera retained the lepismatoid mandible working mechanism(in the imago the mouth parts are atrophied) and that both the
Odonata and the Neoptera are in this respect synapomorphousand constitute the monophylon Metapterygota. This contri-butes additional support to the assumption that Pterygotaevolved by transformation from a lepismatid stage.
The aim of the present work was to study and understandthe features of the lepismatid organization as adaptations tothe corresponding adaptive zone and to find an evolutionary
pathway to pterygotan organization with the aid of anappropriate hypothetical scenario. It seems sound to arguethat from all thinkable hypotheses about a historical evolu-
tionary process that one will come next to the real event whichis most conclusive, namely, which explains a maximum ofobservable evolutionary results, found in fossil or extant
organisms, by necessitating a minimum of speculative assump-tions. A scenario which meets these conditions as far aspossible is presented.It was necessary to study the structural and functional
features of the skeleto-muscular system of the lepismatidthorax on preserved and living specimens because the existingreferences do not satisfy the demands of this work. The
description of the skeleto-muscular patterns by Barlet (1951,1953, 1954) does not permit insight into the movements thatactually occur and the analysis of the locomotory system by
Manton (1972) is, at least for the basal parts of the legs, notdetailed enough. As pointed out by Ellington (1991) allaerodynamic theories postulate some form of gliding as thefirst step in the evolution of flight. Falling and gliding
U.S. Copyright Clearance Center Code Statement: 0947–5745/2002/4002–0065$15.00/0 www.blackwell.de/synergy
J. Zool. Syst. Evol. Research 40 (2002) 65–81� 2002 Blackwell Verlag, BerlinISSN 0947–5745
Received on 13 July 2001

experiments with models offered the opportunity to obtaininformation on the aerodynamic potentialities of lepismatidinsects as the presumed starting point for the evolution of
active flight.
Materials and methods
The morphological, functional and behavioural studies were made onLepisma saccharina Linnaeus, 1758, captured in my home, and on anundetermined Ctenolepisma species from Tenerife living outdoor understones. The latter species was sampled by lifting stones and holdingthem above a plastic film, the disturbed animals soon dropped downand were not able to run away on the smooth film. The capturedanimals were either fixed and preserved in a mixture of 2-propanol,acetic acid and formaldehyde recommended by Smith (1970) or heldalive for months in glass vials with cork stoppers that were moisteneddaily. The vials contained some crumpled paper and little food (rolledoats, dry catfood).The spatial relationships of the pterothoracic skeleto-muscular
system were studied in both thick sections made with a razor bladeand in complete series of histological sections. The thick sections werefaintly stained with diluted picro-indigocarmine, embedded in cedar-wood oil and examined microscopically, partly with polarized light anda first-order-red compensator plate. The histological sections werestained with Cason’s trichrome (see Bock 1989), a simple modificationof Mallory’s triple stain which enables discrimination between endoc-uticle, mesocuticle and exocuticle. The numbering of muscles areaccording to Matsuda (1970).The movements of the cuticular parts during leg-moving cycles were
studied on living specimens where the dorsal side was stuck to a smallmetallic plate. The very rapid leg movements were observed visuallywith a stereo microscope and documented with flash-light photogra-phy (see Fig. 3A–C); the series of phographs permitted the recon-struction of the different phases of movement. The maximal number ofleg moving cycles per second was estimated from stride length and themaximal running speed. The speed was roughly estimated by compar-ison with reference speeds and the stride length was measured from thedistance between footprints left on a smoky glass plate.The postures and attitudes of freely falling animals were photo-
graphed in the light of a digital stroboscope (Novadirect, Kehl,Germany) with 194 flashes per second against a remote darkbackground.The gliding abilities of lepismatid insects were tested by experiments
with models imitating the insects in shape, horizontal outline and mass.The models were cut out of stiff paper or thin cardboard, and the masswas adjusted by sticking suitable pieces of metal to the underside of the‘thorax’ (see Fig. 6). They were either used as ‘flat’ models or stuckwith suitable pieces of foamed plastic to imitate the spatial shape ofLepisma more closely (see f in Fig. 6b). Finally, the models were bentin a fashion for optimal gliding. Starting from size and mass of living
Lepisma (body length L ¼ 8 mm, planform area A ¼ 12 mm2, weight8 mg), models of different size were made by isometric variation. Themodels were dropped from various heights H and the glide distance Xwas measured on the ground as the distance between the landing pointand the point below the point of falling. Each experiment wasrepeated n ¼ 20 times. For details see legends to Figs 6–8 and Tables 1and 2.
Results and discussion
Lepismatid adaptive zone and organization
The lepismatid adaptive zone is characterized by the combi-nation of two strategies to avoid mortality by predators. (1)
Seeking refuge in narrow flat spaces (e.g. below stones) intowhich predators cannot follow. (2) Running away as fast aspossible when disturbed or when in the open. The corres-
ponding selection caused extreme fleetness and a dorso-ventrally flattened, extremely smooth and lithe body withbackwardly directed flattened thoracic legs (Figs 1 and 3A).
The effectiveness of these strategies is obvious from the factthat in Tenerife, for example, abundant populations of aCtenolepisma sp. coexist with the small gecko Tarentoladelalandii Dumeril & Bibron, 1836, in the same habitat.
That living or refuge-seeking within flat spaces is anancestral feature of all extant Lepismatoidea (i.e. Lepismati-dae, Nicoletiidae and Lepidotrichidae) is obvious from the
peculiar form and posture of the palpus labialis which is thesame in all Lepismatoidea. For reasons of space, the palpus iscurved backwards behind the mouthparts and ends in a
widened, dorso-ventrally flattened terminal segment (Fig. 3A).In Nicoletiidae, the body is less dorso-ventrally flattened thanin Lepismatidae, apparently because the Nicoletiidae are
secondarily subterraneous. Lepismatidae are especially well-adapted to flat spaces: the posterior sternal border bears aflange overlapping and protecting the basal median part of thecoxae (s in Fig. 3A). The sternal flange is missing in both the
Nicoletiidae and the Lepidotrichidae (see Wygodzinsky 1961).
Subcoxal parts
Adaptation to life in flat spaces resulted in a uniquely
organized skeleto-muscular system of the thorax, as shownin Figs 1–4 for the pterothorax of Lepismatidae. The presentstudies led to the conclusion that there are three distinct
subcoxal leg segments proximal to the coxa in spite of the factthat these segments are only antero-dorsally sclerotized; the
Table 1. Glide distance X of nearly isometric, flat lepismatid models of different size
Model size I II III IVL 4 mm 8 mm 16 mm 32 mmA 3 mm2 12 mm2 50 mm2 200 mm2
M 2 mg 8 mg 65 mg 500 mgX SD max X SD max X SD max X SD max
Glide distance (cm) 11.2 7.2 23 39.1 15.4 59 19.9 11.3 37 10.3 4.5 19Glide distance (cm) 14.5 11.5 31 45.3 10.1 58 20.6 6.8 32 10.5 2.1 14Glide distance (cm) – – – 50.1 11.2 65 21.6 7.1 30 – – –Glide distance (cm) – – – ~53.5 ~6.3 ~62 26.8 5.2 33 – – –Expected X (cm) 45 45 23 12
Definition of L, A, M and the models’ shape and posture as in Fig. 6. The values of L, M, A of model size II are the same as in adult Lepismasaccharina. The models were dropped from height H ¼ 1.9 m, the ‘head’ pointing downward. Each mean X is the result of 20 glide experimentswith the same individual model; SD, standard deviation; max, maximal value (all in cm). In the series marked with ~ only straight or nearlystraight flights were taken into account. The expected values of X were calculated by using the formula in Fig. 6 and the constant F · f(H) ¼ 300(mg/mm).
66 HASENFUSS

arched sclerites are separated by infolded arthrodial membrane(Figs lc, 2, 3E,F, 4b). The surrounding pleural side wall andthe posterior sternal parts of the segments are membranous.The subcoxae, or their sclerites, are marked here as I, II and
III, beginning with the most basal one. For functional reasonsdiscussed below, the most basal subcoxal sclerite is subdividedby a narrow joint into a dorsal and a ventral part (Id and Iv in
Figs lc, 4b).The sclerites I, II and III are known in the literature as
anapleurite, catapleurite and trochantin, respectively. The
assumption that the pleura and part of the sternal area arederived from basal leg segments is an old idea. However, it isnormally thought that only one – or at most two – subcoxal
segments are integrated (for review see Matsuda 1970; Boud-
reaux 1979; Kukalova-Peck 1983, 1987). The view that thereare three subcoxal segments is supported by the followingfacts. (1) Each subcoxal sclerite has muscles of its own, oneinsertion is on the sclerite itself, the other either on a body part
(tergite, endosternum) or on another leg segment (Fig. 4b). (2)Within each joint, there is at least one articulation between thesclerites (Fig. 1c). As described below, the whole subcoxal
complex is moved together with the coxa; the leg base is freelymovable within the membranous sidewall of the body. (3) Thesubcoxal sclerites and the basis of coxa are somewhat
telescoped. All these parts bear groups of proprioceptive setaeventrally near their proximal margin suggesting serial homol-ogy (Fig. 2). It should be noted in passing that three subcoxal
and six distal segments (coxa, trochanter, femur, tibia, tarsus
Table 2. Glide distances and esti-mated relative glide efficiencies of atype IIB model (Fig. 8) with plasticfoam added, straight ‘abdomen’and stepwise shortened ‘winglength’ (WL). Models weredropped in gliding orientationfrom height H ¼ 1.0 m
B WL A M X *F *F¢ X¢
4 0 20 11.3 0.237 134 126.8 0.2246 1 26 11.8 0.244 111 117.3 0.2588 2 32 12.2 0.271 103 107.8 0.28312 4 44 13.0 0.325 96 93.5 0.31616 6 56 13.9 0.254 63 69.7 0.28020 8 68 14.7 0.250 54 50.7 0.253
B, total breadth in the ‘thoracic’ region (mm);WL, wing length (mm); A, area of horizontal outline (mm2);M, mass (mg); X, measured glide distance (m), each value is the mean of n ¼ 20 experiments;*F ¼ F · f(H), relative glide efficiency (g/m) for H ¼ l.0 m, estimated with the equation in Fig. 6;*F ¢ ¼ F ¢ · f(H), expected relative glide efficiency for H ¼ 1.0 m, estimated from the linear regression*F ¢ ¼ 93.5 – 4.76 · (B–11) with best fit to the experimental data; X¢ ¼ expected glide distance estimatedby using X¢ ¼ *F ¢ · A/M.
Fig. 1. Mesothoracic legs of Ctenolepisma sp. (a,b) and Lepisma saccharina (c–g), double headed arrows indicate possible movements. (a) Lateralview. (b) Dorsal view of leg. (c,d) Movability of coxae, covered contours dashed, membranes punctured. (e) Articulation III/cxv. (f). ArticulationIII/cxd. (g) Articulation II/III. a–a¢, b–b¢, axes of rotary movements; p–p¢, parasagittal plane; Id, Iv, II, III, subcoxal sclerites; Id/II, II/III, III/cxd, IIIcxv, articulations; cx, coxa; fe, femur; pta, pretarsus; ti, tibia; tr, trochanter; ta, tarsus
Evolutionary pathway to insect flight 67

and pretarsus) make up a total of nine leg segments which
might be, according to Paulus (1985), the ancestral number ofEuarthropoda.
Leg movements
The transformation to backwardly directed legs necessitatedconsiderable reorganization of the leg movement system. Theobserved movability of the more distal leg joints is shown inFig. la,b. The propulsive backstroke occurs now by abduction
of the trochanter, the forward stroke by its adduction (insteadof remotion or promotion of the coxa, respectively).The very fast leg movements were studied by a series of
photographs made with flash light (Fig. 3A–C). In thephotographs, the phase of the leg movement cycles could beidentified by the known features of hexapodan leg coordina-
tion. According to Manton (1972), the legs of both sides of asegment normally work in opposite phase and the phasedifference or time delay of succeeding legs is one-third of thepace duration. The relative durations of forward and back-
strokes of each leg are approximately in the ratio 2 : 1, and thepropulsive backstroke of a leg terminates nearly at the pointwhen the leg behind has finished its forward stroke and started
the backstroke.Three main movements of the basal leg parts were observed.
(1) Latero-median abduction and adduction of the coxae
already described by Manton (1972) – different phases ofmovement are shown in Fig. 3A,B. (2) The leg base is movedanteriorly during the forward stroke (right body side in
Fig. 3B) and posteriorly during the backward stroke (left bodyside). (3) Rotary movements on the longitudinal axis of the
coxa. The lateral margin of right coxa is turned ventrally in
Fig. 3C whereas it is lifted in Fig. 3B.Movement (2) contributes to the stride length and thus
improves the running speed. Movement (1) helps to keep the
leg tip at the same comparatively short distance from thesagittal plane of the body – a fact which is indispensable forwalking. The coxae of both body sides are simultaneouslyadducted to the median when the legs are in the middle of
forestroke or backstroke (see forelegs in Fig. 3A). The coxaespread laterally when the legs are moved out of this position inanterior or posterior directions. The coxae move back to the
median when the reverse occurs. The rotary movement on thelongitudinal axis of the coxa is responsible for lift and touchdown of the leg tip as described below.
Skeleto-muscular system of the pterothorax
Latero-median abduction of coxae must be caused by thecontraction of muscles 101 and 102 (Figs 3 and 4b; i.h inFig. 3E,F), adduction by muscles connecting the lateral
margin of coxae with the endosternite (n in Fig. 3E, omittedin Fig. 4). The leg base is moved slightly backwards during thepropulsive backstroke of the leg by two muscles inserting on
the sclerites of subcoxa I: muscle 92 and the large muscle 25(Fig, 4b; e.f in Fig. 3E,F). Lever effects between the anteriorsclerite Iv and and the dorsal sclerite Id are avoided by a
narrow membranous zone between both. (According to thepresent hypothesis that both belong to subcoxa I, this jointmust be a secondary adaptation.) Sclerite Id is a flat triangular
structure with thickened anterior margins, it occupies thebottom of a flat horizontal invagination (Fig. 3E,F). Poster-
Fig. 2. Lepisma saccharina, ante-ro-ventral view of left sidemetathoracic leg base showingsubcoxal sclerites (Iv, II, III), coxa(cx), groups of proprioceptive setae(arrows) and muscle insertions.Covered contours are dashed orpunctuated, for numbers of mus-cles see Fig. 4. III/cxv – ventralarticulation of coxa
68 HASENFUSS

iorly, it offers a place for the insertion of muscle 92 whichoriginates within the paratergite (Fig. 4a,b; e in Fig. 3E,F).Theinvagination of Id is doubtless the result of the dorso-ventraldepression of the body and was possible because the lateral
longitudinal muscles above the invagination prevented con-flicts with the dorso-ventral muscles (Figs 3E and 4a). Theopposite movement during the forestroke is executed mainly
by contraction of the muscles 61 and 31. Muscle 61 arises fromthe tergite of the same segment (Fig. 4b; d in Fig. 3E) and 31from the endosternite of the foregoing segment (q in Fig. 3E;
omitted in Fig. 4b). Both muscles insert on the coxal apodeme(Ca in Fig. 4b) and pull the coxa with the whole subcoxalcomplex in the anterior direction. (Forceful subcoxal pulling
muscles would be inadequate because the segments aretelescoped to some degree. Accordingly, muscles 82 and 71are very slender – Figs 3 and 4b).The articulations distal to the coxa are not suitable to lift
the leg tip during the forward stroke and touch it downduring the backstroke. Observations showed that this occurs
by slight oscillating rotary movements of the coxa on itslongitudinal axis (b–b¢ in Fig. lc). To avoid up and downmovements of the median side of the coxae which wouldconflict with the need for flatness, one basal coxa articulation
(III/cxv – in Figs lc,d, 2) has to be located near the medianside of the coxa (Fig. 1d). A second opposed articulation (III/cxd) permits the transmission of rotary movement of the
sclerite II on axis a–a¢ to the coxa by the independentlymovable sclerite III (Fig. lc). The axes a–a¢ and b–b¢ areinclined to each other. If the coxa is not permitted to execute
rotary movement on axis a–a¢ by muscles, it is forced to moveon axis b–b¢ and sclerite III works like the movableconnecting piece in a cardan shaft in which the axis of
movement by articulations with one adjacent part is trans-verse to that with the other. Accordingly, the axis ofmovement between sclerite II and III (articulation II/III) istransverse to the axis of movement between sclerite III and
coxa (articulations III/cxv and III/cxd). The subcoxal scleritesand their articulations are functionally important and this
Fig. 3. (A–C) Ctenolepisma sp. ventral side, flash photographs during ‘rapid running’, phases of leg movement. A, Overview. B, C, Metathorax.D, Ctenolepisma sp., micrograph of the mesothoracic trochanter, dorsal view, polarized light with first-order-red compensator plate. E, F,Lepisma saccharina, micrographs of histological cross sections of the left side of metathorax: E, overview, F, region of subcoxal articulation Id/II(F); large arrow points at the articulation Id/II; small arrow points at the tendon which connects muscle 53–55 with the dorsal end of sclerite Iv;for muscle numbers and for location of section planes see Fig. 4b (a-a¢ for E, b-b¢ for F). Id, II, III, subcoxal sclerites; ab, abductores trochanteriwith cuticular tendon; ad, adductores trochanteri; ar, coxo-trochanteral articulation; cr, crop; cx, coxa; ec, endocuticle; fe, femur; mc,mesocuticle; p, paratergite; pl, palpus labialis; t, tergite; tf, trochantero-femoral muscles; tr, trochanter; s, sternal flange. Scale: 0.1 mm. Musclesof E, F: a, longitudinal medio-dorsal; b, longitudinal latero-dorsal; c, 91/93; d, 61; e, 92; f, 25; g, 53–55; h, 102; i, 101; j, 72/73; k, 81; m,longitudinal ventral; n, coxo-endosternal extending to lateral coxal margin; o, 71; q–31
Evolutionary pathway to insect flight 69

seems to be the reason for the less-reduced condition of the
subcoxal leg segments in Lepismatidae.During the propulsive backstroke the whole leg complex is
moved backwardly by contraction of both the very forcefulmuscle 25 (f in Fig. 3E,F) and muscle 92 (Fig. 4a,b). Simul-
taneously, the trochanter swings backwards by the contractionof muscles 50, 53–55 and 52 (Figs 2,3 and 4b), and the lateral
margin of the coxa is pressed down by the contraction of
muscles 25 and 53–55. Muscles 25 and 53–55 pull the postero-dorsal margin of sclerite Iv (and indirectly the anterior marginof II) inwards and thus cause the slight rotary movement onaxis a–a¢ (f and g in Fig. 3F). Contraction of muscles 74 and 63raise the lateral coxa margin during the forward stroke. Thisskeleto-muscular arrangement allowed the formation of a
Fig. 4. Metathoracic muscles ofLepisma saccharina, left side, in-tegument shown as transparent,border of the muscle insertionspunctured. Numbering of musclesaccording to Matsuda (1970). AC,antecosta. (a) Dorsal view withtergal longitudinal muscles, sitesof dorso-ventral muscle insertionsas in (b), outline of sclerite Idpunctured; full circle, location ofarticulation Id/II; dashed line,ventral border of the paratergite.Anterior insertion of muscle 92 ison sclerite Id. (b) Lateral view withmuscles of the leg base, coxo-endosternal and trochantero-en-dosternal muscles not shown; a–a¢and b–b¢ mark the planes of sec-tions in Fig. 3E,F), respectively.Sclerotized leg parts marginallypunctured; Id, Iv, II, III, scleritesof subcoxal segments; CX, coxa;Ca, coxal apodeme (dashed line).Muscle insertions: pt on paraterg-ite, cx on coxa, en on endosternite;troch abd, abductores trochanteri.Muscles 83–86 said to connect thetergite with sclerite II were notfound, the corresponding tergalinsertions were observed to belongto muscle 61. Muscle 31 not drawn,it connects the coxal apodeme Cawith the endosternite of the pre-ceding segment; together with 61 itprotracts the leg base
70 HASENFUSS

sternal integumental fold which covers and mechanicallyprotects the ventral side of the leg basis (s in Fig. 3A).In Archaeognatha, the structures corresponding to the
articulations III/cxv and III/cxd are on the lateral dorsalmargin of the vertically arranged coxa, III/cxd being behindIII/cxv (see Manton 1972: Fig. 31). It is probable that, starting
from such an arrangement, the coxae were tilted backwardlyby shifting the anterior articulation III/cxv in antero-ventralposition. This would mean that this location of III/cxv is asynapomorphy of Lepismatidae (or even Lepismatoidea?) and
Pterygota (compare Figs 4b and 10).Judging from Fig. 52 in Boudreaux (1979), Tricholepidion
exhibits all subcoxal sclerites but seems to be less stringently
adapted to flat spaces. The coxa is more vertical andarticulation II/III is close to articulation III/cxd, a featureindicating that the movement system is somewhat different
from that in Lepismatidae. Tricholepidion is found in compar-atively weak material like leaf litter or under bark, not understones as are the Lepismatidae (Wygodzinsky 1961).
Autotomy and regeneration of legs
Other solutions to the problem of lifting and setting down thefoot tips by utilizing more distal leg joints would be hard toenvisage: the dicondylous coxo-trochanter joint with its now
nearly vertical rotary axis is fully engaged in forward andbackstroke, the trochantero-femur joint is specialized forautotomy and the more distal leg segments are acting nearly
in the same plane (Fig. 1). Autotomy was observed when a legof a Lepisma or Ctenolepisma specimen was prevented fromrapid movement, e.g. by holding on the distal part with
forceps. The proximal end of the femur is somewhat telescopedinto the trochanter and fits tightly into it, like a stopper withina bottle-neck (Fig. 3D). Numerous small retractor muscle
fibres hold the femur in position, and only when external forceis large enough to overcome the muscular tension does thefemur work itself loose and break off. Legs lost to autotomy inCtenolepisma regenerated readily and appeared with the next
moult after 6–8 weeks. The regenerated legs were fullyfunctional, in colour slightly paler but otherwise similar tothe previous ones. Other lost parts (scales, parts of antennae
and caudal filaments) regenerate fully on the same occasions.
Fleetness
Lepismatidae are extremely fleet – when disturbed they run
away very fast and stop immediately on finding a suitableretreat. The stride length is somewhat more than half the bodylength and the pace duration is, according to Manton (1972),
‘of the order of 0.02 s or less’. Hence, the maximal frequency ofleg movement is 50 Hz or somewhat more. The presentobservations on Lepisma and Ctenolepisma are in accordancewith this statement. By measuring the footprints on a smoky
glass plate, the stride length was found to be 6 mm for aLepisma specimen of 11 mm body length and 8 mm for aCtenolepisma of 13 mm body length. The corresponding
running speeds were roughly estimated by comparison withreference speeds and found to be at least 0.3 and 0.4 m/s,respectively.
If a glass plate is held perpendicularly in the path of a fastrunning lepismatid, the animal does not bump into it with itshead but turns aside and continues running along the plate.Assuming that the reaction is caused by contact with the
antennae which are held in a V-shape in front of the head, the
reaction time is of the order of 20 or 30 ms. The reactions arefast enough for correction movements even during free fall.
Posture during falling
If a lepismatid is dropped in any orientation from any height(even from few cm), it lands safely on the ventral side and isable to run away without time delay. The posture during the
free fall was photographically documented by using a strobo-scope with nearly 200 flashes per second against a darkbackground. The photographs show that during free fall the
legs are stretched out latero-ventrally as far as possible andthat the caudal filaments and antennae are spread as shown inFig. 5. It seems that the spread appendages improve parachu-ting, reduce velocity and thus minimize the risk of injurious
landing. At the beginning, the abdomen is curved dorsallycausing the animals to turn until the ventral side faces theground (Fig. 5a,b). This happens because the point of gravity
is sufficiently distant from the point of drag. Thereafter theabdomen is held straight (Fig. 5c). No gliding was observed –it seems that gliding is prevented by drag of the extended legs
and diverging antennae. However, as shown by experimentswith models, gliding should occur when the antennae and theflattened legs are pressed back against the body.It is of interest for the discussion below that spiders dropped
in any orientation likewise always land on the feet and quicklyget away. As documented by photographs made with strobo-scope, spiders somewhat raise their distal leg parts during
falling; they thus raise the point of drag above the point ofgravity and turn until the ventral side is facing the ground.
Experiments with models
As lepismatids do not utilize gliding themselves the principalgliding abilities of insects of lepismatid organization werestudied on models imitating the lepismatid body in shape, size
and mass. Legs were omitted in the models, imitating a posturewith the flattened legs closely applied to the body and thusforming an aerodynamically effective ventral plane. Themodels were dropped from height H and the glide distance X
was measured on the ground (Fig. 6). As in the fallingexperiments with living lepismatids, the models landed safelyon the ventral side when the ‘caudal filaments’ were spread
laterally and the ‘abdomen’ was curved dorsally as inFig. 6a,b. Additionally, in this posture, the ventral plane ofthe ‘thorax’ is inclined optimally for gliding.
The complicated aerodynamic equations which are used indiscussing physics of gliding (Kingsolver and Koehl 1985,1994; Ellington 1991; Wootton and Ellington 1991) can be
replaced, for the present purpose, by a very simple equation inwhich forces and velocities are eliminated. It is derived fromthe basic aerodynamic equations, valid for Reynolds numberslarger than one, and is more convenient for the exploitation of
gliding experiments because it describes explicitly the glidedistance X as a function of certain relevant features of thegliding object and of a non-linear function of height [f(H)]
from which the object is dropped (equation in Fig. 6). Therelevant features of the object are: the mass M, the horizontalplanform area A, and a factor of proportionality which can be
regarded as the gliding efficiency F of the object. The efficiencyF and the function f(H) are a priori unknown, however, theproduct F · f(H) can be estimated from experimental data byusing the equation in Fig. 6. For experiments with the same
height H, the values of the products are measures of the
Evolutionary pathway to insect flight 71

relative gliding efficiencies of the objects, and for experimentswith the same object and different heights H the products, aswell as the glide distances themselves, permit impression of thefunction f(H) – for effects of different heights see Fig. 8.
The influence of the ratio A/M was tested by experimentswith isometric flat models of different sizes, similar tolepismatids in outline and mass (Fig. 6). The measured glide
distances exhibited considerable variance, mainly dependingon whether the glide path was more or less straight within avertical plane or curved with stochastic turns. Nevertheless,
except for the smallest model I, the results shown in Table 1(see p. 66) are in good accord with the equation. Withincreasing body size the distance X decreases drastically
because the mass increases with the cube and the area onlywith the square of a linear measure. The violence of the impacton ground increased considerably with the size of the models.The glide distance of the small model I is much reduced. It
appears that, in contrast to the larger models, the inertial massof this model is too small to stabilize gliding. The turbulent airstream relative to the falling object pulls the object out of its
gliding attitude again and again, it staggers and goes more orless directly down. To determine the starting point of thiseffect, the weight of models of the optimal size II was altered
by several steps and tested as before (Fig. 7). In the range of 8–16 mg, the mean glide distance follows the equation – thereduction begins immediately below 8 mg mass. The resultsmean that the gliding abilities are optimal for lepismatid
insects of approximately 8 mm body length and 8 mg mass –and this is just the size of adult Lepisma. Larger and smallerinsects should achieve smaller glide distances. The further
experiments were therefore made with size II models.
It was supposed that increased glide distance might beattained by enlarging the ‘thoracic’ area around the point ofgravity. Unexpectedly, the addition of slender ‘lateral wings’
(of the same form and size as in models IIB and IIC in Fig. 8)did not improve the gliding abilities if they were added tomodel II with dorsally bent abdomen and spread ‘caudal
filaments’. The glide distance was equal or even smaller thanfor model II without ‘wings’. The extensions improvedparachuting instead of gliding, indicated by slower descent.These undesired effects could be avoided by changing the
‘posture’ of the model II to model IIA with a nearly straightabdomen and the ‘caudal filaments’ put together to anunpaired projection (Fig. 8; IIA.b). The model IIA glided
only when it was dropped in the gliding orientation as shownin Fig. 8b, but the glide path was nearly always in a verticalplane, in contrast to the often curved glide path of model II. If
only the less curved events were taken into account for modelII, the mean glide distances were the same as for model IIA(compare the X-values for II and IIA in Fig. 8).
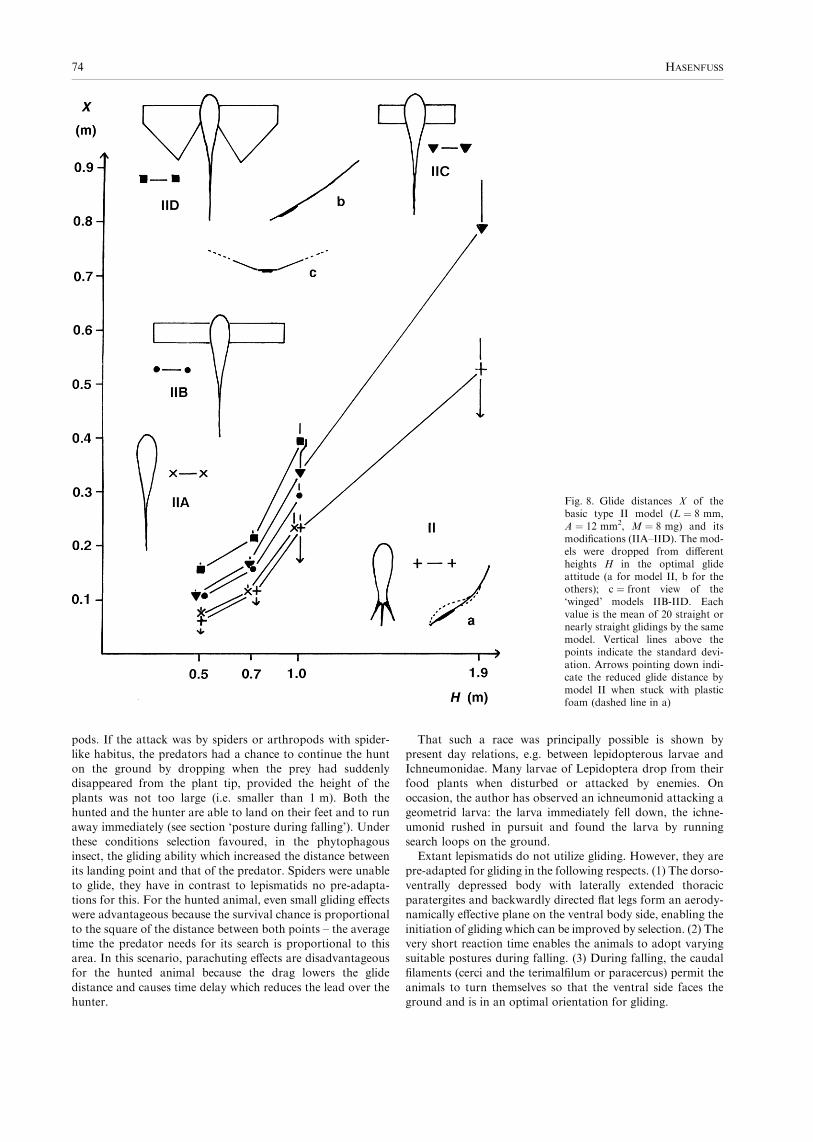
Unlike model II, the addition of wings to the type IIA modelsignificantly increased the glide distance as is evident fromFig. 8. The Mann–Whitney U-test showed that all X-values ofmodels IIB, IIC, IID are different from the corresponding
values of model IIA to the 0.001 probability level. Theexperiments with models IIB, IIC and IID showed thatenlarging the wings increases the glide distance in particular
if the wings are at least as broad as long. The X-values ofmodel IID are significantly larger than the correspondingvalues of model IIC and IIB. Elongation of narrow wings as in
model IIB does not improve the glide distance: for H ¼ 1.0 mthe distance for model IIB is even smaller than for model IIC(according to the Mann–Whitney test this difference is
significant to the 0.01 probability level). This indicates thatinitially the wings had to be short and broad and that longslender wings are adaptations in the context of active flight. Itshould be noted that the addition of wings improved the
attitude stability if they were somewhat raised as shown inFig. 8c; the models glided if dropped in any orientation.The product F · f(H) for the same heights H is a relative
measure of the gliding efficiency F. The estimates of F · f(H)for the models in Fig. 8 and height H ¼ 1.0 m were (in g/m)160 for II and IIA, 93 for IIC, 58 for IIB, and 54 for IID.
Remarkably, the wingless model of lepismatid shape has thegreatest gliding efficiency! It is apparent that F depends on thebreadth B of the models (i.e. the distance between their mostlateral points in the region of the ‘thorax’). Elongation of the
anterior edge raises drag and reduces velocity, and the glidedistance is reduced because it is proportional to the square ofthe velocity. This drag effect of small slender wings was already
observed by Kingsolver and Koehl (1985) by their measure-ments on models in a wind tunnel. Increased glide distance istherefore only possible if, by adding wings, the quotient A/M is
large enough to over-compensate for the reduced glidingefficiency. This is especially true when the added wings arenearly as broad as they are long or even broader.
This hypothesis was tested by experiments with a type IIBmodel stuck with plastic foam and equipped with 8 mm longand 3 mm broad wings which were stepwise shortened bycutting away parts of the tip (Table 2, see p. 67). The
somewhat greater mass of the model improved its glidingperformance: it glided perfectly straight. The glide distancesmeasured were in accordance with the above hypothesis. They
increased with the wing length until the wings were as long as
Fig. 5. Posture during falling, drawings after photographs made withstroboscope at 194 flashes per second. (a) Lepisma saccharina,drawings at intervals of 5 ms, middle drawing is 14 cm below thepoint of drop. (b,c) Ctenolepisma sp., distance below the point of drop:16 cm (b), 30 cm (c)
72 HASENFUSS

they were broad, and decreased when the wings became longer.
As expected, the estimated relative gliding efficiencies F · f(H)show, in rough approximation, linear regression in regard tobreadth B.
Discussion of the evolutionary pathway
Pre-adaptations and scenario
The dicondylous mandibles of Lepismatoidea and Pterygota
permitted the exploitation of wide range of food material
formerly not accessible to insects. It might therefore well bethat the presumed lepismatid-like early ancestors of Pterygotawere able to feed on living standing vascular plants. The basalparts of the stems were probably not suitable for this. [Lesions
in fossil axes of Rhynia plants (Lower Devonian) are of a sortas made by arthropods with piercing mouthparts (Kevan et al.1975).] More probably, the ancestors of Pterygota were to
climb the plants and gnaw at the soft tissue of the shoot tips.Feeding in this way demanded leaving the already exploitedshoots and climbing neighbouring ones. Every alteration
which facilitated this change was of selective value until theoptimal solution of the problem was achieved, namely activeflight from one plant tip to another.
One of the first achievements seems to be the ability torecognize plant stems in the environment. Since the eyes ofPterygota are generally well developed, it must be assumedthat the early pterygotan ancestor was diurnal and probably
evolved the ability to search actively for suitable plants bywalking in the direction of visually perceived vertical struc-tures. That such an ability is within the scope of insect
organization is shown by the analogous ability of the carabidbeetle Calosoma sycophanta Linnaeus, 1758, which entersshrubs and trees in search for lepidopterous larvae and pupae.
When walking on the ground the beetles are able to locateshrubs, and they have been observed to run in straight lines tothem from a distance of half a dozen metres or so. Occasion-ally, it may happen that a Calosoma runs to and climbs a
human person standing motionless in the landscape (observa-tion made by the author). In contrast to the pterygotan line,the extant Lepismatoidea are active only in the dark and the
eyes are more or less reduced – certainly a secondary situation.Another possible improvement was leaving exploited shoots
by dropping as a mode of fast, energetically inexpensive
vertical descent. This was especially advantageous, when theshoot was densely covered with upwardly directed spine-likeprotrusions which hindered the descent, and/or when the
phytophagous animals were attacked by carnivorous arthro-
Fig. 6. (a,b) ‘Lepismatid’ models used for glide experiments. (a) Horizontal outline, L, ‘body’ length. (b) Side view in glide attitude, f, plasticfoam stuck to some of the flat models, m, piece of metal. (c) Graph of the path of an object dropped from height H to the ground with horizontaltravel X ( ¼ glide distance) due to aerodynamic effects; the equation describes the glide distance as function of features of the gliding object andof a non-linear function of height f(H)
Fig. 7. Mean glide distances X of flat models type II (L ¼ 8 mm, A ¼12 mm2) with different mass achieved by sticking metal pieces ofappropriate weight to the underside (m in Fig. 6b). Experiments asexplained in Table 1 (H ¼ 1.9 m). The curve is calculated from theequation in Fig. 6 for the constant F · A · f(H) ¼ 312 (mg · cm)
Evolutionary pathway to insect flight 73
pods. If the attack was by spiders or arthropods with spider-like habitus, the predators had a chance to continue the hunton the ground by dropping when the prey had suddenlydisappeared from the plant tip, provided the height of the
plants was not too large (i.e. smaller than 1 m). Both thehunted and the hunter are able to land on their feet and to runaway immediately (see section ‘posture during falling’). Under
these conditions selection favoured, in the phytophagousinsect, the gliding ability which increased the distance betweenits landing point and that of the predator. Spiders were unable
to glide, they have in contrast to lepismatids no pre-adapta-tions for this. For the hunted animal, even small gliding effectswere advantageous because the survival chance is proportional
to the square of the distance between both points – the averagetime the predator needs for its search is proportional to thisarea. In this scenario, parachuting effects are disadvantageousfor the hunted animal because the drag lowers the glide
distance and causes time delay which reduces the lead over thehunter.
That such a race was principally possible is shown bypresent day relations, e.g. between lepidopterous larvae andIchneumonidae. Many larvae of Lepidoptera drop from theirfood plants when disturbed or attacked by enemies. On
occasion, the author has observed an ichneumonid attacking ageometrid larva: the larva immediately fell down, the ichne-umonid rushed in pursuit and found the larva by running
search loops on the ground.Extant lepismatids do not utilize gliding. However, they are
pre-adapted for gliding in the following respects. (1) The dorso-
ventrally depressed body with laterally extended thoracicparatergites and backwardly directed flat legs form an aerody-namically effective plane on the ventral body side, enabling the
initiation of gliding which can be improved by selection. (2) Thevery short reaction time enables the animals to adopt varyingsuitable postures during falling. (3) During falling, the caudalfilaments (cerci and the terimalfilum or paracercus) permit the
animals to turn themselves so that the ventral side faces theground and is in an optimal orientation for gliding.
Fig. 8. Glide distances X of thebasic type II model (L ¼ 8 mm,A ¼ 12 mm2, M ¼ 8 mg) and itsmodifications (IIA–IID). The mod-els were dropped from differentheights H in the optimal glideattitude (a for model II, b for theothers); c ¼ front view of the‘winged’ models IIB-IID. Eachvalue is the mean of 20 straight ornearly straight glidings by the samemodel. Vertical lines above thepoints indicate the standard devi-ation. Arrows pointing down indi-cate the reduced glide distance bymodel II when stuck with plasticfoam (dashed line in a)
74 HASENFUSS

As pointed out by Magdefrau (1968), Smart and Hughes(1972), and Kevan et al. (1975), terrestrial vascular plantsappeared in the Upper Silurian and were well developed in the
Lower Devonian. Initially they exceeded not more than a fewdecimetres in height and reached 1 m height at the end ofLower Devonian. During this period the plants were of suitable
size for the supposed evolutionary processes. Some of the plantshoots were densely covered with upwardly directed spine-likeprotrusions. In the further course of the Devonian the plantsevolved to trees.
The fossil record shows clearly that carnivorous arthropodsexisted at the time in question, suggesting that the typicallepismatid adaptive zone was already realized. A very diver-
sified fauna of fossil spiders is known from the Carboniferous,and arthropods of spider habitus had already appeared in theLower Devonian (Petrunkevitch 1955; Kevan et al. 1975).
Even today insects are the main prey of spiders and it may wellbe that this relation has been maintained since the origin ofPterygota.
Considering the optimal gliding size (8 mm body length) itseems that a glide distance at least in the order of 20 cm issufficient to escape hunting enemies. This distance can beattained from heights of 1 m or more (Fig. 8). Selection for
improving the gliding ability therefore occurred only when theheight was smaller. Thus, improved gliding was in principlepossible almost from the very beginning of the appearance of
vascular plants. Shortening the way on ground or even landingon the next stem by improved gliding ability was advanta-geous. Once initiated, gliding was therefore further optimized
by selection.
Presumed adaptations for gliding
The falling experiments with models showed that there is a
‘window’ for gliding in insects of lepismatid shape with theoptimal size of approximately 8 mm body length and 8 mgmass. Insects with smaller than the critical mass are excludedfrom gliding (in contrast to active flight, which is possible for
extremely small insects). Thus, the smaller juvenile instars werenot able to participate in gliding and remained bottom-dwellers. The lepismatid glide window has remained undetec-
ted because the former attempts to reconstruct the selectivefactors initiating insect wing evolution started from stick-likemodels (Flower 1964), much larger models with wings
(Wootton and Ellington 1991) or were confined to measure-ments on larger models in wind tunnel (Kingsolver and Koehl1985; for review see Ellington 1991; Kingsolver and Koehl
1994). However, Flower (1964) had already concluded fromsuch experiments that, in accordance with the present findings,insects of approximately 1 cm length had the highest chance ofevolving wings.
Gliding is improved when the mass is reduced as far aspossible in the range above the critical mass and the thoracicparatergites are enlarged so that the lateral extensions are at
least as broad as long. Utilization of this effect demandschanges in posture during falling. At first, the abdomen mustbe curved dorsally and the tail bristles spread laterally to
achieve gliding attitude. Thereafter the abdomen must bestraight and the tail bristles put together. The fallingexperiments with living lepismatids and the extremely shortreaction times observed show that such abilities are within the
scope of lepismatid organization. Furthermore, it seemsprobable that gliding could be utilized to shorten the way
on the ground to the next recognized plant by evolving theability to control the glide direction by appropriate move-ments of the abdomen. (Walking to visually perceived plant
stalks was important for effective exploitation of plants – seescenario.) Further optimizations concerns the improvement oflanding (perhaps by reducing the velocity by spreading the
tail bristles?).The inevitable increase of the thoracic breadth in the course
of gliding optimization has the disadvantage that the lateralextensions interfere with vertical environmental structures
when the animals were running on the ground. Within theassumed scenario, selection thus promoted increased glidingability through enlarged thoracic paratergites on the one hand
and favoured unimpeded running over the ground on theother. The problem could be solved by turning the paratergitesupwards when running and turning them horizontally when
gliding. Because the zone between the paratergites and the legbase is membranous in lepismatoid insects, this effect couldeasily be attained via a strip of weakness within the tergite
between the area of the insertions of the dorso-ventral musclesand the point above the articulation Id/II (Figs 3E.F, 4a and9). [That this is possible in principle is shown by analogy inoribatid mites which evolved movable paratergal plates
(Woodring 1962)]. The successive contractions of the dorso-ventral muscles during running then pulled the median part ofthe tergite ventrally, pressed the lateral tergal parts against the
subcoxal articulations Id/II, which now acted as pivots, andforced the paratergites into a vertical posture (Fig. 9). On theother hand, when the animal was falling, the dorsal longitudi-
nal muscles were contracted and bent the terga so that theparatergites were forced into a more horizontal posture thatwas apt for gliding. This event explains the existence of the
indirect flight muscles in Pterygota and why some of the dorso-ventral muscles are bimodal, i.e. engaged in both wing and legmovement. The thoracic paratergal movements must havebeen simultaneous. It may well be that the somewhat disturb-
ing paratergal muscles (1, 2, 3 and 92 in Fig. 4a) were reducedbefore this evolutionary step as a result of body massreduction.
Once in place, the initially somewhat crude mechanism wasoptimized. The zone of tergal weakness was transformed into ahinge with a straight narrow strip of flexible cuticle. This hinge
separates the parts of the wing from the tergal differentiationswhich evolved later, and is well seen as a straight structure inPaleozoic fossils (e.g. h in Kukalova-Peck 1978: Fig. 45). Theleg base movements in the antero-posterior direction had to be
abandoned by reducing the muscle 92 (Fig. 4a.b) and by insome way fixing the sclerite Iv to the anterior margin of thetergite as well as to the sternite. This initiated the evolution of
the rigid thoracic box. The other leg movements had to bemaintained together with the ability to appress the stillflattened legs to the body during gliding. That part of Iv
bearing the insertions of muscles 25 and 53–55 must thereforehave been separated from the remainder by arthrodial mem-brane. The separated sclerite later became the basalare
(BA – Fig. 10); this conclusion is supported by the probablehomology of the trochanteral muscles 53–55 and p-tr2(compare Figs 4a and 10). The pivot point of the paratergite(articulation Id/II) was now fixed and became the pleural wing
process (WP – Fig. 10). The invaginated sclerite Id wasautomatically moved together with the paratergite and becamein some way integrated into the wing (Fig. 9). Especially the
sole pterale on the underside of the wing (pterale 2) seem to be
Evolutionary pathway to insect flight 75

a derivative of Id. This hypothesis is a modified version ofRasnitsyn’s notion (Rasnitsyn 1981) that the wings arecomposites of tergal and pleural parts. Furthermore, itexplains the ontogenetical relations between leg base and
wings put forward by Kukalova-Peck (1983) and recent studiesof gene expressions (Averof and Cohen 1997). As far as thesearguments are conclusive they support the notion that sclerite
Id is really derived from a former leg segment. [For criticism ofthe conclusions of Averof and Cohen (1997) see Wagele 2000:p. 151.]
Further possible optimizations were: (1) the uncoupling ofthe wing movements from leg movements by appropriateneuronal reorganization; (2) progress in reducing weight by
evolving membranous parts and stiffening nervature on thebasis of pre-existing blood lacunae within the wings; (3)beginning of further differentiations of the wing articulationsby which the transduction of the tergal bending to the wing
movements was improved. This included improved bendingfeatures in the tergum and the reduction of now unnecessarylongitudinal tergal muscles. Finally, only 5 (¼ t14) and 6
(¼ t12) were retained as main flight muscles.It seems inevitable that during the evolutionary phase of
gliding, all thoracic segments were engaged in a similar way,
and that not only the paratergites of the pterothorax but alsothose of the prothorax were movable. It is therefore notsurprising that signs of prothoracic paratergite movabilitywere found in Paleozoic Pterygota (see Kukalova-Peck 1978).
Presumed adaptations for active flight
With the above optimizations for gliding, a pre-adaptiveplateau was attained from which active flight was accessible.
The possible selection by predators was not eliminated by theoptimizations because, for low falling heights, gliding was notsufficient to bring the animal out of the danger zone. Initiation
of active flight was therefore of selective advantage. Activeflight from one plant tip to another was so advantageous thatits evolution went on even after the threat by predators ceased.
Evolution was therefore proceeding without interruptiontowards active flight and, since there was no adaptive zonein which an intermediate stage of this process was optimal, nointermediate stage had the chance to be preserved.
The wings used for active flight evolved into elongated, moreslender extensions and were restricted to the pterothorax; the
prothoracic appendages became immovable. This immobiliza-tion was probably due to the fact that the prothorax is too faraway from the point of gravity and that the moments exerted
by prothoracic wings would be too large. Furthermore,vibration caused by movable prothoracic wings are thus keptaway from the head with its sensory organs.
Initiation of active flight was possible by neuroethologicalvariation: flapping the wings up and down during gliding wasapt to increase the glide distance. Adaptation for light weightfavoured the development of somewhat passively flexible wing
planes with more rigid leading anterior margins. During thedown-stroke the wing thus became automatically pronatedand during the up-stroke supinated generating components of
lift and drive. Additionally, pronation could be supported bythe contraction of the basalar muscles and supination by themuscles of the later evolved subalare.
During active flight, the legs no longer had to be flattenedand appressed to the body and the subcoxal sclerites II and IIIcould now be integrated into the thoracic box improving the
mechanical stability of the flight apparatus. The dorso-ventrally depressed body became more stick-like. Since thelepismatid muscular system of the leg base was, as it appears,at least partly transformed into the pterygotan musculature
(compare Figs 4 and 10), the original spatial arrangement ofthe subcoxal parts had to be maintained. The axis of the basalarticulations of the coxa remained unchanged (its ventral
articulation is normally still in the antero-ventral instead ofventral position – Fig. 10) and the distal parts of the legs werevertically erected by appropriate torsion within the coxa.
Except for the pterygotan basalare, subalare and trochant-inus, the lepismatid sternal and pleural membranous partsbecame sclerotized throughout, thus forming the rigid frame-
work for the flight apparatus. Sclerites II and III were fusedtogether and the part between the articulations Id/II and III/cxd was stiffened by the pleural ridge (PL in Fig. 10).As already stated in the previous section, that part of sclerite
Iv which bears the anterior insertions of the muscles 53–55 and25 had to be separated from the remainder by flexible cuticleand became the basalare (compare Figs 4 and 10). This is
because the main part of Iv was integrated firmly into the rigidsegmental framework whereas the ability of the subcoxalsclerites II and III to move was still important. With the
reduction of the endosternite, the posterior insertion of muscle25 shifted to the ventral part of the body wall near the anteriormargin of the segment (p23 and p-s12 in Fig. 10). This waspossible because the posteriorly directed movement of the leg
base was no longer needed. Muscle 53–55 maintained itsinsertion on the trochanter abductor tendon, thus indicatingthe homology with the pterygotan p-tr2.
The basalar muscles are direct flight muscles and they aresuitable for controlling the pronation of wings. Antagonisticsupination of wings is possible by contraction of the subalar
muscle t-cx8 (Fig. 10). In contrast to the basalar muscles itsorigin is not yet clear. One possibility among others is that thelespismatid precursor muscle of t-cx8 was engaged in the wing
upstroke and located so far behind articulation Id/II that itcaused supination at the same time. Candidates for this arethe lepismatid posterior dorso-ventral muscles, probablymuscle 63 (Fig. 4b). This hypothesis means that the effectivity
of this muscle was improved by separating its dorsal insertionfrom the tergite on a sclerite of its own (the subalare) andshifting this sclerite firstly behind and then below the wing
base.
Fig. 9. Schematic section illustrating movability of paratergite andsclerite Id after the hypothetical intercalation of the unsclerotized zoneuns. The pre-existing articulation Id/II is the pivot for thesemovements. Id, II – subcoxal sclerites
76 HASENFUSS

The trochantinus appears as the anterior part of sclerite IIIwhich is either completely integrated into the thoracic box (as
in all paleopterous insects) or separated by membrane fromboth the sclerite II and the caudal part of sclerite III (manyNeoptera – TN in Fig. 10). Muscles t-ti1 and t-ti1,2,3 whichconnect the tip of the trochantinus and the tergite tilt the axis
going through both basal articulations of the coxa. This seemsto be advantageous for crawling in the narrow spaces typicalfor the neopterous adaptive zone. In the paleopterous adaptive
zone, this ability to move is useless and the trochantinus isintegrated into the rigid thoracic box to increase stability. Ifthe presented scenario is correct, the adults of early Pterygota
were paleopterous and the movable trochantinus of the flyingstage of Neoptera was secondary. However, contrary to theassumption of Boudreaux (1979), this does not necessarily
mean that it is a completely new structure.As pointed out in the ecological section, neoptery is
probably due to the adaptation of the imago to nymphalfood. Since in early evolution the nymphs were living in
narrow habitats, the nymphs retained lepismatid characterswithin their locomotor system. For the ancestor of Neoptera,it was therefore easy to secondarily carry over the trochantinal
ability to move from the nymph to the adult and to combine it
with the imaginal thoracic box. That this was possible isshown by the fact that, in adult Plecoptera for example, the
nymphal muscles are retained in the adult with few exceptions(Wittig 1955). Thus both views: that the separated trochant-inus is a synapomorphy of adults in Neoptera and that it ishomologous with the anterior part of sclerite III in Lepis-
matidae, are valid and do not exclude each other. The firstapproach exclusively compares the imaginal stages, thesecond one considers the complete organization, i.e. the
holomorphe.In the pterothorax, nymphs and adults of the neopterous
Plecoptera exhibit trochantina which are similar to that drawn
in Fig. 10. However, in the nymphal prothorax, the sclerite III(called coxopleurite) is separated from sclerite II (anapleurite)by flexible cuticle as in Lepismatidae (see Snodgrass 1935:
Fig. 90). In particular the articulations corresponding to III/cxd, III/cxv and II/III are in the same positions as inLepismatidae: the articulation II/III is far away from thearticulation III/cxd (as shown in Fig. 1c) and not close to it as
in Tricholepidion. It seems that the lepismatid pattern ofsubcoxal parts was maintained in nymphal Plecoptera becausethe nymphs lived permanently in habitats with narrow spaces.
The still separated prothoracic sclerites III and II support the
Fig. 10. Typological scheme ofpterygotan pterothorax, lateralview with lateral and dorsal mus-cles. Muscle names according toMatsuda (1970), names of theprobably homologous lepismatidmuscles (see Fig. 4a,b) below thepterygotan names. III/cxd, III/cxv– basal articulations of coxa; BA –basalare; CX – coxa; P3 – pterale 3;PH – phragma; PL – pleural ridge;SA – subalare; TN – trochantinus,WP – pleural wing process
Evolutionary pathway to insect flight 77

view that the pterygotan pleuron really evolved by fusingsubcoxal sclerites.Summing up, it seems that the evolution of flight, once
initiated with improvement of gliding, was canalized untilactive flight was achieved. The present synthesis supports theclassical hypothesis that the evolution of wings started with
gliding via paratergites. It presents a solution to the problem ofhow the paratergites became movable by utilizing lepismatidpre-adaptations. Furthermore, it supports the idea of Hinton(1963) that there were biotic factors (vascular plants, preda-
tors) which were responsible for selection which caused thecorresponding transformations.
Evidence for the presumed transformations
The following facts can be taken as special evidence of thetransformation from lepismatid to pterygotan organization.(1) A considerable part of the lepismatid muscle system seems
to be homologous with muscles of Pterygota (see Matsuda1970 and Fig. 10). In particular the existence of the pterygotanmuscle p-tr2 which connects the basalare with the trochantinus
supports strongly the transformation in question. (2) Com-parative studies of motoneurone patterns supplying dorsallongitudinal muscles revealed a common pattern in Lepismaand Pterygota which is quite different from that in Chilopoda
(Heckmann and Kutch 1995). (3) The two-bundled appearanceof muscle t-p5,6 is explained by its origin from two differentmuscles: muscle 87/88 of subcoxa I and 81 of subcoxa II
(compare Figs 4b and 10). (4) The antero-ventral articulationof the coxa (III/cxv) is in the same position in Lepismatidaeand Pterygota (compare Figs 1c and 10). (5) The movable
trochantinus of Neoptera is undoubtedly the homologue of theanterior part of sclerite III in Lepismatidae, moved byhomologous muscles (compare Figs 4b and 10). (6) The
separated subcoxal sclerites II and III in the prothorax oflarvae of Plecoptera indicate that the pterygotan pleura arereally derived from lepismatid subcoxal parts.
Ontogenetic aspects
Experiments have shown that insects with a mass smaller than8 mg (and smaller than about 8 mm body length) were notable to glide. Thus, the smaller juvenile instars of the
pterygotan ancestors were excluded from using the lateralextensions for gliding and had to remain substrate-dwellers.The diverging adaptive zones of non-gliding juvenile instars
and adult instars capable of gliding necessitated differentialadaptations: adaptations for different food, a need for extremelight-weight construction in adults which is of less importance
to juvenile stages, etc. The problem could be solved by utilizingan ‘ontogenetic switch’ which enabled the independent accu-mulation of juvenile (nymphal/larval) and adult (imaginal)adaptations by a phase of organismic reorganization, i.e.
metamorphosis. This switch is achieved in Pterygota by areduced concentration of juvenile hormone during ecdysis. Theamount of ontogenetic reorganization can be very different,
beginning with very slight alterations (as in nearly ‘ametabo-lous’ post-embryonal development). However, the groundorganization for both is generally built up during embryonal
development.The lepismatoid insects are ametabolous and this means
that, at the beginning of the evolution of Pterygota, theadaptations for the adults could be only attained via
morphogenetic alterations during embryonal development.
It is therefore to be expected that the extensions used forgliding by early Pterygota must already have been present inthe youngest instars, formed as if they were functional.
However, for the juvenile stages living on the ground suchextensions were disadvantageous, as they were in the adultswhen running on the ground. Improvement was possible by
the same adaptations as in adults: by the ability to move thelateral extensions up and down. Finally, however, the mostsuccessful solution for the juvenile wings was to reduce thesize of the lateral lobes, apply them to the body backwards
and immobilize them by fusion with the tergite. This finalstage is generally found in extant non-endopterygotaninsects.
These considerations are in accordance with the fossil recordpresented by Kukalova-Peck (1978). She found that inPaleozoic nymphs the wings were laterally directed thoracic
outgrowth with signs of actual or former ability to move, i.e.more or less pronounced hinge structures with pteralia,even – as residuals – on the prothorax. In Paleozoic nymphs
of Ephemeroptera and Megasecoptera, the wings were largelateral outgrowths considerably exceeding the lateral border ofthe trunk, but streamlined by being curved backwards (seeKukalova-Peck 1978: Figs 28 and 31). Possibly, such exten-
sions were engaged in secondary roles. In other lines, theoutgrowths did not or did only little exceed the lateral borderof the trunk, nevertheless they still exhibited signs of former
ability to move (Kukalova-Peck 1978: Figs 22, 24, 33 and 35).Nymphs with movable wings must have existed for many
million years and there arises the question why juvenile instars
did not likewise achieve active flight which is physicallypossible even for very small individuals? The ultimate reasonfor this is probably the same as for the other animals which
evolved the ability to fly. Juvenile Chiroptera begin flyingshortly before they attain the terminal size, however, theycannot fly perfectly until they are fully adult (Kleiman andDavis 1979; Corbet and Harris 1991). In Aves the situation is
similar. This suggests that body growth and active flightcannot be successfully combined, perhaps because active flightdemands extremely sophisticated organization, which cannot
be achieved when the tissues are growing. There seems to exista ‘rule of incompatibility of growth and flight’. Thus it can betaken for granted that, from the very beginning, active flight
was exclusively achieved by adult individuals of the ultimatesize, i.e. imagines. In early Pterygota, the ability to moult in theimaginal stage was still present as is indicated by thesubimaginal moult in Ephemeroptera. The fact that all other
extant Pterygota do not moult in the imaginal stage isprobably a result of selection for light weight. Moultingdemands fully functional epidermis cells even within the wings.
As pointed out by Maiorana (1979), degeneration of theepidermis in wings is a means of improving the wing efficiencywith the result that, except for Ephemeroptera, functional
wings are confined to only one instar: the last and reproductivestage. Further she pointed out that the subimaginal moult inEphemeroptera was apparently retained because it is needed to
complete the elongation of caudal filaments and forelegs ofadults. These elongations are special features of the reproduc-tive system of mayflies and cannot develop during nymphal life.Accordingly, the subimagines exhibit very low flight activity:
they fly only from the water surface where they emerge to thebank or shore and start to moult to the definitive imago.The avoidance of mechanical interference with the environ-
ment caused the exopterygotan nymphs to develop their
78 HASENFUSS

external wing anlagen to backward-directed, immobilizedsmall extensions. Endopterygota attained a still further step.Starting from exopterygote nymphs, the anlagen became
invaginated and disappeared completely as external append-ages. This endopterygoty made possible extremely variedannidation of the larvae, resulting in the unusually high
number of extant species. As pointed out by Hinton (1948) it isapparently not possible to realize the transition from anendopterygote stage to functioning wings during one moult. Itwas therefore inevitable to retain an exopterygote juvenile
stage at the end of larval life. Since this stage fitted neither inthe imaginal nor in the larval adaptive zone, it evolved to thequiescent pupa. That the pupa should be regarded as a
nymphal rather than as a subimaginal stage is shown by thefact that, in larvae of Lepidoptera for example, the pattern oftactile setae is maintained nearly unchanged in the pupa,
whereas the imago develops a completely different new set oftactile setae (Hasenfuss 1997). Tactile setae which are experi-mentally heat damaged in a larva, are missing in subsequent
larval instars and the pupa. In the corresponding imago,however, the patterns of sensilla develop undisturbed (Hasen-fuss, unpublished experiments).
Ecological aspects
The present hypothesis assumes that the preflight ancestors ofPterygota had biting mouth parts and fed initially in all instarson weak tissue at the tips of vascular plants (meristem, stages
of fructification) reaching new plant stems by running on theground. This implies a terrestrial mode of life on comparat-ively dry uplands in all stages. The evolution of gliding and
active flight in the adults gave rise to diverging adaptive zonesfor these and the juvenile instars (nymphs, larvae). Theimagines were fluttering from plant tip to plant tip still feeding
on weak tissue, their adaptive zone was doubtless that ofpaleopterous insects, i.e. the wings could not be folded on theback. The wing movements were primarily synchronous as isstill observed in extant Ephemeroptera (‘eosynchronous flight’
– Brodsky 1994). The juvenile instars remained substrate-dwellers and had to eat other food, probably dead, decayingplant material.
Continued damage of the plant tips caused the plants toevolve protective devices reducing the amount of food access-ible to the flying insects. The substrate-dwelling nymphs were
not affected by this. The problem could be solved via thefollowing strategies. (1) No food uptake by the imagines; thisstrategy is followed by the extant Ephemeroptera, for example.
The males ingest air in the gut, thus forming stiff support forthe long abdomen – a very effective method to reduce weight asan adaptation in the context of the nuptial dancing flight. (2)Becoming carnivorous as in Odonata. (3) Breaking through
the plant barriers mechanically by modifying the mouthpartsfor piercing and sucking. This is a very successful strategywhich possibly evolved several times independently. It is met in
the haustellate Paleozoic insects including the paleopterousPalaeodictyoptera, Megasecoptera and Permothemistida aswell as the neopterous Diaphanopterodea (see, e.g. the
reconstruction of diaphanopteroid haustellum by Kukalova-Peck 1985). Most Paraneoptera are likewise equipped withpiercing mouthparts. (4) Exploiting the same food as theflightless juveniles.
A disadvantage of the original bipartite adaptive zone is thefact that colonizing new habitats is only possible if both the
juvenile and the imaginal stages find appropriate conditions,especially suitable food. Both strategies (3) and (4) areappropriate to master this difficulty. The development of hard
piercing mouthparts enabled the animals to exploit a greatvariety of living plant material which cannot be managed usingbiting mouth parts. Equipped with the same piercing mouth-
parts, nymphs and imagines may nevertheless have quitedifferent life styles as is shown by Cicadidae for example, withsoil living nymphs and aerial imagines. In this context it is ofinterest that the nymphs of Palaeodictyoptera and Megase-
coptera possessed haustella, as did the adults, and that theywere probably terrestrial (Smart and Hughes 1972). Theonisciform shape of palaeodictyopteran nymphs (see Wootton
1972) fits well in this scenario.Strategy (4) was very successful and led supposedly to
neoptery, i.e. the ability to fold the wings on the back. Except
for Plecoptera and Endopterygota, strategy (4) is found innearly all extant Neoptera, and even within Endopterygota itwas attained by the largest group – the Coleoptera – which
protect the folded hind wings by heavily sclerotized elytrae.The utilization of the same food material by both juvenilesand adults was therefore of great selective advantage.Neoptery permitted the imagines to crawl to the same sites
as the juveniles without being impeded by the protrudingwings.Thus there are two adaptive zones in regard to wing posture:
that of neoptery in which the imagines are able to crawl withinnarrow spaces on the ground and in vegetation, and that ofpaleoptery in which the imagines are unable to do this. That
the inability to fold the wings can be advantageous is shown bybutterflies and skippers (Papilionoidea, Hesperoidea) withinthe otherwise neopterous Lepidoptera: they secondarily lost
the ability to fold the wings by secondary adaptation to theadaptive zone of paleopterous insects. From this, the questionarises as to whether the paleopterous Ephemeroptera andOdonata are primarily so or were secondarily derived from
neopterous ancestors.Recent Ephemeroptera and Odonata seem to be primarily
paleopterous. From the outgroup criterion follows that if the
Metapterygota (Odonata + Neoptera) are really monophy-letic, then the paleoptery of Ephemeroptera and Odonata ismost probably plesiomorphic in regard to Neoptera. This
hypothesis is supported by differences in adhesive deviceswhich are important for hold on smooth plant surfaces. InEphemeroptera, the adhesive pads are zones of flexiblecuticle ventrally at the base of the pretarsal claws, and in
Odonata adhesive devices are missing (Beutel and Gorb2001). However, in extant Neoptera the typical adhesivedevices are the very effective unpaired arolia between the
pretarsal claws. Judging from Fig. 33A in Kukalova-Peck(1985), even the extinct neopterous Diaphanopterodea seemto have possessed arolia. It is likely that arolia evolved in the
context of neoptery and that this happened only once. It isunlikely that Ephemeroptera and Odonata evolved from astage with arolia because in those Lepidoptera which are
secondarily incapable of folding their wings (Papilionoidea,Hesperiidae), the arolia are still present. These conclusionssupport the notion that Diaphanopterodea are cladisticallytrue Neoptera and that their haustellum evolved independ-
ently from that of paleopterous hastellate taxa. Accordingto Rasnitsyn (1981) there are no significant differences bet-ween Diaphanopterodea and Neoptera in wing articulation
structure.
Evolutionary pathway to insect flight 79

The swamp woods of the Carboniferous and early Permianoffered rich possibilities for diversification and annidation ininsects. Typically, many bottom dwelling forms or stadia were
forced to adapt to unpredictable changes of water level andbecame thus amphibious or even aquatic. This offers anexplanation as to why the supposedly primary paleopterous
Ephemeroptera and Odonata acquired aquatic nymphs – acertainly secondary mode of life. The mode of respiration isvery different in the nymphs of both taxa indicating independ-ent adaptation to aquatic life. Small appendages on legs and
other body parts described and figured by Kukalova-Peck(1987) for many Paleozoic insects possibly functioned as gills.Gill vestiges may even be retained in aerial stadia as shown by
some extant Plecoptera.It is improbable that the aquatic life style of the nymphs of
Plecoptera is due to adaptation to swamp wood conditions.
The plecopteran nymphs are generally rheobiontic and are theonly Pterygota which retain the lepismatid condition in whichsubcoxae II and III are distinct sclerites in the prothorax
(Snodgrass 1935). This suggests that the nymphs becameaquatic at an early stage of neopteran evolution and that theywere thereafter permanently living in running water, adaptedfor life either within the thin zone at the surface of the
submerged stones in which the flow velocity is low or withinthe gaps between the stones. This life in narrow spaces isprobably the reason for the maintenance of the mentioned
lepismatid feature.
Preservation of ancient features
The above discussion confirms the view that ancient charac-
teristics had especially good chances of being maintained inevolutionary lines which otherwise became very specialized. If,at an early level of organization, such characteristics were
integrated within specialized constructions, they were savedfrom further alterations and could survive even hundreds ofmillion years. Thus the specialized lepismatid type of locomo-tion within flat spaces favoured the preservation of thoracic leg
base structures which enabled the identification of threesubcoxal leg segments. In Ephemeroptera, ancient character-istics (such as the subimaginal moult, the synchronous wing
movements, the retention of three caudal filaments, etc.) couldonly be preserved because they became an integral part of anelaborate mating system with the nuptial dancing flight of
males. In Plecoptera, as noted above, ancient characters of thelocomotory system were maintained, as it seems, because theirnymphs were adapted for life in running water at an early stage
of neopteran evolution.
Acknowledgements
I thank A. S. Lawrence for revising the English.
Zusammenfassung
Ein moglicher, von der lepismatiden Organisation ausgehenderEvolutionsweg zum Flugvermogen der Insekten
Ausgehend von der Hypothese daß die Evolution des Flugvermogensder Pterygota mit der lepismatiden Organisation ihrer Ahnen begann,wurde die funktionelle Anatomie des Thorax von Lepisma saccharinaund einer Ctenolepisma sp. im Hinblick auf Adaptationen an dieadaptive Zone der Lepismatidae und im Hinblick auf Praeadaptatio-nen fur die Evolution der Pterygota untersucht. Es wurden in derPleuralzone gut erhaltene Teile von drei subcoxalen Beingliedern
gefunden, die an der Beinbewegung teilnehmen. Die Strategie derLepismatidae, vor Feinden durch rasches Laufen zu fliehen und sich inflachen Spalten zu verstecken, fuhrte zu einer dorsoventralen Abfla-chung des Korpers, die beim Herabfallen Gleiteffekte und weitereFlucht am Boden ermoglichte. Im Zusammenhang mit der vermutetenErnahrung von Triebspitzen der im Devon noch niedrigen Gefaß-pflanzen eroffnete dies einen kanalisierten Evolutionsweg zum aktivenFlug wobei moglicherweise spinnenartige Pradatoren den Selektions-druck verstarkten. Gleitversuche mit ‘lepismatiden’ Modellen zeigtenein enges Gleitfenster mit dem Optimum von 8 mg Gewicht und 8 mmKorperlange. Es wurde eine Gleichung gefunden, die die Gleitdistanzals Funktion der Gleitflache, der Masse, der Gleiteffizienz desFallkorpers und einer nicht-linearen Funktion der Fallhohe beschreibt.Vergroßerung der Gleitdistanz war moglich, wenn die Korpermasseveringert, die lateral vorragenden thorakalen Paratergite vergroßertwurden und als ‘Flugel’ nicht langer als breit waren. Die Behinderungder Flucht am Boden durch laterale ‘Flugel’ konnte durch derenVertikalstellung beim Laufen und durch Horizontalstellung beimGleiten vermindert werden. Die Beweglichkeit war durch Einfugeneines Membranstreifens zwischen Tergit und Paratergit sowie dieNutzung des vorhandenen Gelenks zwischen zwei basalen Subcoxal-gliedern mit dem vorhandenen Muskelsystem moglich. Demnach istder dorsale Teil eines ursprunglichen Beingliedes im Flugel derPterygota integriert. Auf- und Abbewegung der Flugel wahrend desGleitens ermoglichten den Beginn des aktiven Fluges. Die dafurerforderlichen Transformationen sowie deren ontogenetische undokologische Aspekte werden diskutiert.
References
Averof, M.; Cohen, S. M., 1997: Evolutionary origin of insect wingsfrom ancestral gills. Nature 385, 627–630.
Barlet, J., 1951: Morphologie du thorax de Lepisma saccharina L.(Apterygote Thysanoure). I.- Squelette externe et endosquelette.Bull. Ann. Soc. Entomol. Belg. 87, 253–271.
Barlet, J., 1953: Morphologie du thorax de Lepisma saccharina L.(Apterygote Thysanoure). II. – Musculature (ler partie). Bull. Ann.Soc. Entomol. Belg. 89, 214–236.
Barlet, J., 1954: Morphologie du thorax de Lepisma saccharina L.(Apterygote Thysanoure). II. – Musculature (2e partie). Bull. Ann.Soc. Entomol. Belg. 90, 299–321.
Beutel, R. G.; Gorb, S. N., 2001: Ultrastructure of attachmentspecializations of hexapods. Arthropoda): evolutionary patternsinferred from a revised ordinal phylogeny. J. Zool. Syst. Evol. Res.39, 177–207.
Bock, P., 1989: Romeis mikroskopische Technik. 17. Aufl. Munchen,Wien, Balltimore: Urban und Schwarzenberg, pp. 1–697.
Boudreaux, H. B., 1979: Arthropod Phylogeny with Special Referenceto Insects. New York, Chichester, Brisbane, Toronto: J. Wiley,pp. 1–320.
Brodsky, A. K., 1994: The Evolution of Insect Flight. Oxford, NewYork, Tokyo: Oxford University Press, pp. 1–229.
Corbet, G. B.; Harris, S., 1991: The Handbook of British Mammals.Bats: Order Chiroptera, 3rd edn. Oxford: Blackwell, pp. 81–145.
Ellington, C. P., 1991: Aerodynamics and the origin of insect flight.Adv. Insect Physiol. 23, 171–210.
Flower, J. W., 1964: On the origin of flight in insects. J. Ins. Physiol.10, 81–88.
Furst von Lieven, A., 2000: The transformation from monocondylousto dicondylous mandibles in the Insecta. Zool. Anz. 239, 139–146.
Hasenfuss, I., 1997: Precursor structures and evolution of Tympanalorgans in Lepidoptera (Insecta, Pterygota). Zoomorphology 117,
155–164.Heckmann, R.; Kutch, W., 1995: Motor supply of the dorsallongitudinal muscles II: comparison of motoneurone sets inTracheata. Zoomorphology 115, 197–211.
Hennig, W., 1953: Kritische Bemerkungen zum phylogenetischenSystem der Insekten. Beitr. Ent. (Sonderheft) 3, 1–85.
Hennig, W., 1969: Die Stammesgeschichte der Insekten. Frankfurt a.M: Waldemar Kramer, pp. 1–436.
Hinton, H. E., 1948: On the origin and function of the pupal stage.Trans. R. Ent. Soc. Lond. 99, 395–409.
80 HASENFUSS

Hinton, H. E., 1963: The origin of flight in insects. Proc. Roy.Entomol. Soc. Lond. (C) 28, 24–25.
Kevan, P.G.; Chaloner,W.G.; Savile, D. B.O., 1975: Interrelationshipsof early terrestrial arthropods and plants. Palaeontology 18, 391–417.
Kingsolver, J. G.; Koehl, M. A. R., 1985: Aerodynamics, thermore-gulation, and the of insect wings: differential scaling and evolution-ary change. Evolution 39, 488–504.
Kingsolver, J. G.; Koehl, M. A. R., 1994: Selective factors in theevolution of insect wings. Ann. Rev. Entomol. 39, 425–451.
Kleiman, D. G.; Davis, T. M., 1979: Ontogeny and maternal care. In:Baker, R. J.; Jones, J. K.; Carter, D. C. (eds), Biology of Bats in theNew World Family. Part III. Lubbock, Texas: Texas Tech Press,pp. 387–402.
Kristensen, N. P., 1975: The phylogeny of hexapod ‘orders’. A criticalreview of recent accounts. Z. Zool. Syst. Evol.-Forsch. 13, 1–44.
Kristensen, N. P., 1981: Phylogeny of insect orders. Ann. Rev.Entomol. 26, 135–157.
Kukalova-Peck, J., 1978: Origin and evolution of insect wings andtheir relation to metamorphosis, as documented by the fossil record.J. Morph. 156, 53–126.
Kukalova-Peck, J., 1983: Origin of the insect wing and wingarticulation from the arthropodan leg. Can. J. Zool. 61, 1618–1669.
Kukalova-Peck, J., 1985: Ephemeroid wing venation based upon newgigantic Carboniferous mayflies and basic morphology, phylogeny,and metamorphosis of pterygote insects (Insecta, Ephemerida). Can.J. Zool. 63, 933–955.
Kukalova-Peck, J., 1987: New Carboniferous Diplura, Monura, andThysanura, the hexapod ground plan, and the role of thoracicside lobes in the origin of wings (Insecta). Can. J. Zool. 65, 2327–2345.
Magdefrau, K., 1968: Palaobiologie der Pflanzen. Jena: GustavFischer, pp. 1–549.
Maiorana, V. C., 1979: Why do adult insects not moult? Biol. J. Linn.Soc. 11, 253–258.
Manton, S. M., 1972: The evolution of arthropodan locomotorymechanisms. Part 10. Locomotory habits, morphology and evolu-tion of the hexapod classes. Zool. J. Linn. Soc. 51, 203–400.
Matsuda, R., 1970: Morphology and evolution of the insect thorax.Mem. Entomol. Soc. Can. 76, 1–431.
Paulus, H., 1985: Euarthropoda. In: Siewing, R. (ed.), Lehrbuch derZoologie. Band 2 Systematik. Stuttgart: Gustav Fischer Verlag,pp. 768–791.
Petrunkevitch, A., 1955: Arachnida. In: Moore, R. C. (ed.), Treatise ofPalaeontology, Part P, Arthropoda 2. Kansas: Lawrence, pp. 32–162.
Rasnitsyn, A. P., 1981: A modified paranotal theory of insect wingorigin. J. Morph. 158, 331–338.
Smart, J.; Hughes, N. F., 1972: The insect and the plant: progressivepalaeoecological integration. In: van Emden, H. F. (ed.), Insect/Plant Relationships. Oxford: Blackwell, pp. 143–155.
Smith, E. L., 1970: Biology and structure of some Californiabristletails and silverfish (Apterygota: Macrocoryphia, Thysanura).Pan-Pacific Ent. 46, 212–225.
Snodgrass, R. E., 1935: Principles of Insect Morphology. New York:McGraw-Hill, pp. 1–667.
Staniczek, A. H., 2000: The mandible of silverfish (Insecta: Zygen-toma) and mayflies (Ephemeroptera): its morphology and phyloge-netic significance. Zool. Anz. 239, 147–178.
Wagele, J.-W., 2000: Grundlagen der Phylogenetischen Systematik.Munchen: Dr. F. Pfeil, pp. 1–315.
Wittig, G., 1955: Untersuchungen am Thorax von Perla abdominalisBurm. (Larve und Imago) unter besonderer Berucksichtigung desperipheren Nervensystems und der Sinnesorgane. Zool. Jb. Anat. 74,491–570.
Woodring, J. P., 1962: Oribatid (Acari) pteromorphs, pterogastrinephylogeny, and evolution of wings. Ann. Entomol. Soc. Am. 55,394–403.
Wootton, R. J., 1972: Nymphs of Palaeodictyoptera (Insecta) from theWestphalian of England. Palaeontology 15, 662–675.
Wootton, R. J.; Ellington, C. P., 1991: Biomechanics and the origin ofinsect flight. In: Rayner, J. M. V.; Wootton, R. J. (eds), Biomech-anics in Evolution. Cambridge: Cambridge University Press. pp. 91–112.
Wygodzinsky, P., 1961: On a surviving representative of the Lepido-trichidae (Thysanura). Ann. Entomol. Soc. Am. 54, 621–627.
Author’s address: Professor Dr Ivar Hasenfuss, Karlsbader Str9,D-91083 Baiersdorf, Germany; E-mail: [email protected]
Evolutionary pathway to insect flight 81





























