Embed Size (px)
Citation preview

Introduction
Although many insects survive sub-zero temperatures by utilizing strategies of freeze tolerance and freeze avoidance, the majority of insects are chill-susceptible and die at tempera-tures above the freezing point of their body fluids ( Bale, 2002; Sinclair et al. , 2003 ). Nevertheless, the short-term response of chill-susceptible insects to low temperatures can be plastic: low temperature survival can be improved by both acclimation at mild temperatures in the order of > 12 h ( Rako & Hoffmann, 2006 ) and rapid cold-hardening (RCH) at more severe temperatures over periods of minutes to a few hours ( Lee et al. , 1987 ). The mechanisms of each of these types of plasticity are still being elucidated. Because the mechanisms are unknown, Sinclair & Roberts (2005) suggest that it is premature to assume that the processes associated with acclimation and RCH are the same. Several other stud-ies ( Powell & Bale, 2005; Shintani & Ishikawa, 2007 ) in
aphids and beetles show that RCH persists even after accli-mation, which implies that the mechanisms of each are independent.
Drosophila melanogaster is a popular model organism for studies of acclimation and hardening at both high and low temperatures ( Hoffmann et al. , 2003; Rako & Hoffmann, 2006; Kristensen et al. , 2008 ). However, the responses of D. melanogaster to low temperatures are not always consistent among developmental stages ( Rako & Hoffmann, 2006; Rajamohan & Sinclair, 2008 ). Rako & Hoffmann (2006) report the effects of rearing, hardening and low temperature acclimation in adult D. melanogaster , and find that acclimation at 12 °C still improves acute cold survival in flies that are reared at a low temperature (19°C); however, they do not comment on whether acclimated flies still show an RCH response. Knowledge of the interactions between hardening and acclimation effects on cold toler-ance in D. melanogaster is important for interpreting and designing future mechanistic experiments. In addition, ther-mal responses vary among lines of D. melanogaster ( Rajamohan & Sinclair, 2008 ) and it not known whether there are differences in acclimation – hardening interactions among genetic backgrounds.
As part of a broader programme investigating the feasibil-ity of cryopreservation of D. melanogaster larvae, the extent
Hardening trumps acclimation in improving cold tolerance of Drosophila melanogaster larvae
A R U N R A J A M O H A N and B R E N T J . S I N C L A I R * Department of Biology, The University of Western Ontario, London, Ontario, Canada
Abstract . Chill-susceptible insects are able to improve their survival of acute cold exposure over both the short term (i.e. hardening at a relatively severe temperature) and longer term (i.e. acclimation responses at milder temperatures over a longer time frame). However, the mechanistic overlap of these responses is not clear. Four larval stages of four different strains of Drosophila melanogaster are used to test whether low temperature acclimation (10 °C for 48 h) improves the acute cold toler-ance (LT 90 , ~ 2 h) of larvae, and whether acclimated larvae still show hardening responses after brief exposures to nonlethal cold or heat, or a combination of the two. Acclimation results in increased cold tolerance in three of four strains, with variation among instars. However, if acclimation is followed by hardening pre-treatments, there is no improvement in acute cold survival. It is concluded that short-term thermal responses (e.g. hardening) may be of more ecological relevance to short-lived life stages such as larvae, and that the mechanisms of low temperature hardening and acclimation in D. melanogaster may be antagonistic, rather than complementary.
Key words . Acclimation , chilling injury , cold tolerance , cross tolerance , heat shock , rapid cold-hardening .
* Current address: Department of Entomology, North Dakota State University, Fargo, North Dakota, U.S.A.
Correspondence: Brent J. Sinclair, Department of Biology, The University of Western Ontario, London, ON, N6G 1L3, Canada. Tel.: +1 519 661 2111; fax: +1 519 661 3935; e-mail: [email protected]
Physiological Entomology (2009) 34, 217–223 DOI: 10.1111/j.1365-3032.2009.00677.x
© 2009 The AuthorsJournal compilation © 2009 The Royal Entomological Society 217

to which the natural chilling tolerance of D. melanogaster larvae can be improved through alteration of handling and pre-treatment conditions is being explored. There are signifi-cant differences in cold tolerance between larval instars and between wild-type strains, and larvae of most stage/strain combinations show an increased tolerance to acute cold exposure when both the heat shock response and the RCH response are induced by pre-treatment ( Rajamohan & Sinclair, 2008 ). In addition, recovery from cold exposure is highest when larvae are removed directly to room tempera-ture, rather than a slow or stepwise rewarming ( Sinclair & Rajamohan, 2008 ). In the present study, the role of low tem-perature acclimation in improving acute cold tolerance (i.e. for exposures to intense cold for less than 3 h; Sinclair & Roberts, 2005 ), and the interaction between acclimation and rapid hardening processes are investigated. The acclimation treatment (48 h at 10 °C) represents a significant portion of the post-embryonic development in D. melanogaster , and as long a duration as is practicable for analysis within a single instar. Two hypotheses are tested. The first hypothesis is that acute cold tolerance of D. melanogaster larvae is improved after an acclimation pre-treatment. However, in keeping with observations of RCH and other hardening pre-treatments, it is hypothesized that different wild-type strains of D. melanogaster will show differing magnitudes of response to acclimation, reflecting an effect of the overall genetic background on the organism’s ability to acclimate. The second hypothesis is that acclimation and hardening are independent processes. Thus, it is predicted that larvae subjected to low temperature acclimation will still display improved cold tolerance (over and above anything imparted by acclimation) in response to hardening pre-treatments.
Materials and methods
Four wild-type strains of D. melanogaster (Berlin K, Canton S and Oregon R from the Bloomington Stock Center, and W5N collected in Washington State in 2005 by Dr Michael Dillon) were reared in the laboratory at 21 °C under an LD 14 : 10 h photoperiod and approximately 70% relative humid-ity (RH). Flies were reared in 250-mL bottles (approximately 150 eggs per bottle) or 50-mL vials (50 – 75 eggs per vial) on a cornmeal-yeast medium [1.7% active yeast (w/v), 1% soy flour, 7.0% cornmeal, 4.5% malt extract, 0.5% agar, 7.5% (v/v) corn syrup and 0.5% propionic acid]. Flies were transferred to population cages and eggs and larvae were collected as previously described by Rajamohan & Sinclair (2008) . Four stages of larvae (first, second and third instars and wandering stage third instars) were used.
Preliminary experiments indicated that an acclimation regime of 10 °C for 48 h was appropriate for these experi-ments because survival was high; larvae did not develop sig-nificantly during the time period and the 48-h period is long enough to ensure that responses are likely to be distinct from the acute responses observed in RCH ( Sinclair & Roberts, 2005 ). For acclimation, larvae (or eggs, for acclimating first
instars) were transferred to food vials, incubated at 21 °C (until the appearance of larvae of the appropriate stage), and the vials moved to a Sanyo MIR-153 incubator (Sanyo Scientific, Bensenville, Illinois) at 10 °C and approximately 60% RH, and under the same LD 12 : 12 photoperiod used for rearing. Stage was verified by examining the mouthparts of a subsample of larvae at the time of cold exposure ( Demerec, 1965 ), and only live larvae were used for cold exposure experiments.
Prior to cold exposure, both acclimated and non-acclimated larvae were assigned to groups, placed in foil packets (width 15 mm, height 10 mm) and exposed to one of four pre-treatments: heat shock (36.5 °C for 1 h followed by 1 h at 21 °C), RCH (1 h at 0 °C) and heat shock followed by RCH (‘HS + RCH’;1 h at 36.5 °C, 1 h at 21 °C and 1 h at 0 °C), or no pre-treatment (21 °C for 1 h). Further details of pre-treatments are provide elsewhere ( Rajamohan & Sinclair, 2008 ).
After acclimation and/or pre-treatment, acute cold toler-ance was assessed by placing the packets into 2-mm thick slots in an aluminum block cooled by circulating fluid from a refrigerated circulator [either VWR1157p (VWR, Canada) or Lauda Proline 3530c (Fisher Scientific, Canada)]. Larvae were exposed to cold for a temperature and time that is close to 2 h, and does not result in freezing of the larvae, but has previously been determined to result in approximately 90% mortality (for details of these times and temperatures for each stage-strain combination, see Rajamohan & Sinclair, 2008 ). Larvae were removed immediately to 21 °C (previ-ously shown to maximize survival, Sinclair & Rajamohan, 2008 ) and transferred to a Petri dish containing cornmeal-yeast medium. Larvae were considered ‘alive’ if they were moving and feeding and showed no sign of necrosis after 24 h. Third-instar and wandering larvae that pupariated suc-cessfully and showed no signs of necrosis were considered to be alive.
Survival data (as proportions) were approximately normal after arcsine-square root transformation, and thus trans-formed data were analysed separately for each strain using a general linear model with type three sums of squares using PROC GLM in SAS, version 9.1 (SAS Institute Inc., Cary, North Carolina). Least squares means (which represent the values for selected levels when taking into account variation sourced from other terms in the model) and Tukey’s HSD were used to visualize interaction effects and make post-hoc comparisons, respectively.
Results
Most stages and strains showed good survival of the acclima-tion treatment, with the exception of third instars of W5N and Canton S ( Table 1 ). No sex bias in survival was observed in groups of larvae that were observed up to eclosion (data not shown).
In general, second-instar larvae were most tolerant of acute cold exposure ( Fig. 1 and Table 2 ) and the HS + RCH and RCH pre-treatments resulted in increased survival relative to
218 A. Rajamohan and B. J. Sinclair
© 2009 The Authors
Journal compilation © 2009 The Royal Entomological Society, Physiological Entomology, 34, 217–223
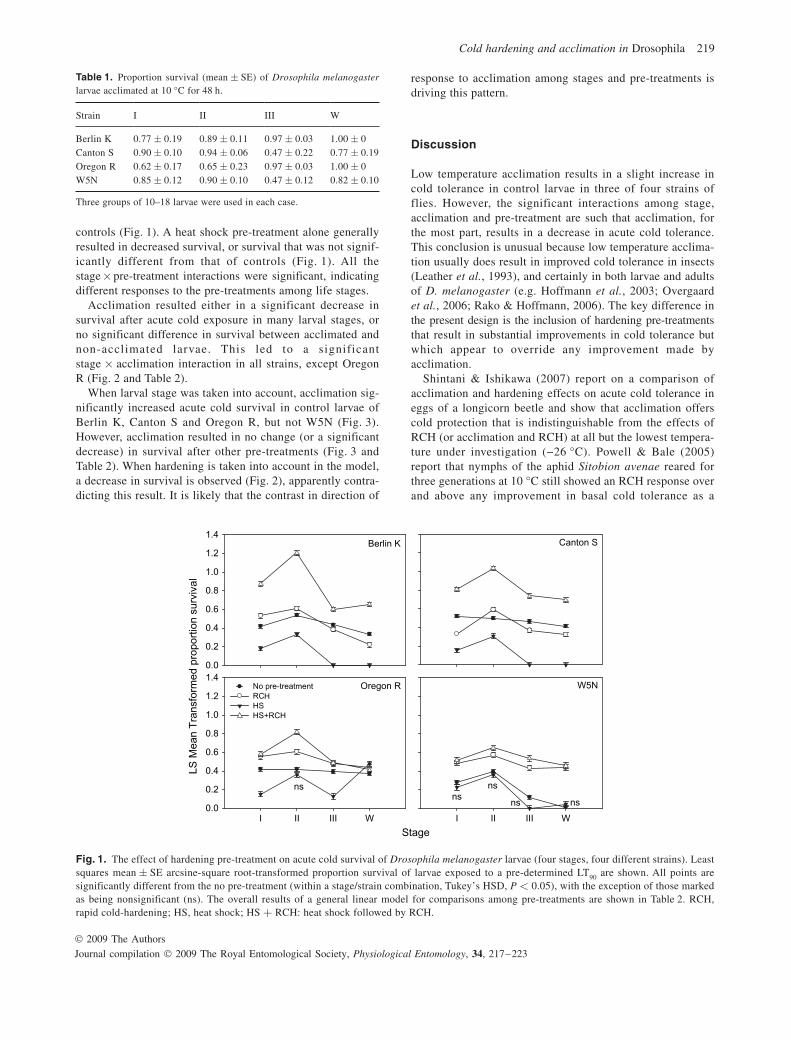
controls ( Fig. 1 ). A heat shock pre-treatment alone generally resulted in decreased survival, or survival that was not signif-icantly different from that of controls ( Fig. 1 ). All the stage × pre-treatment interactions were significant, indicating different responses to the pre-treatments among life stages.
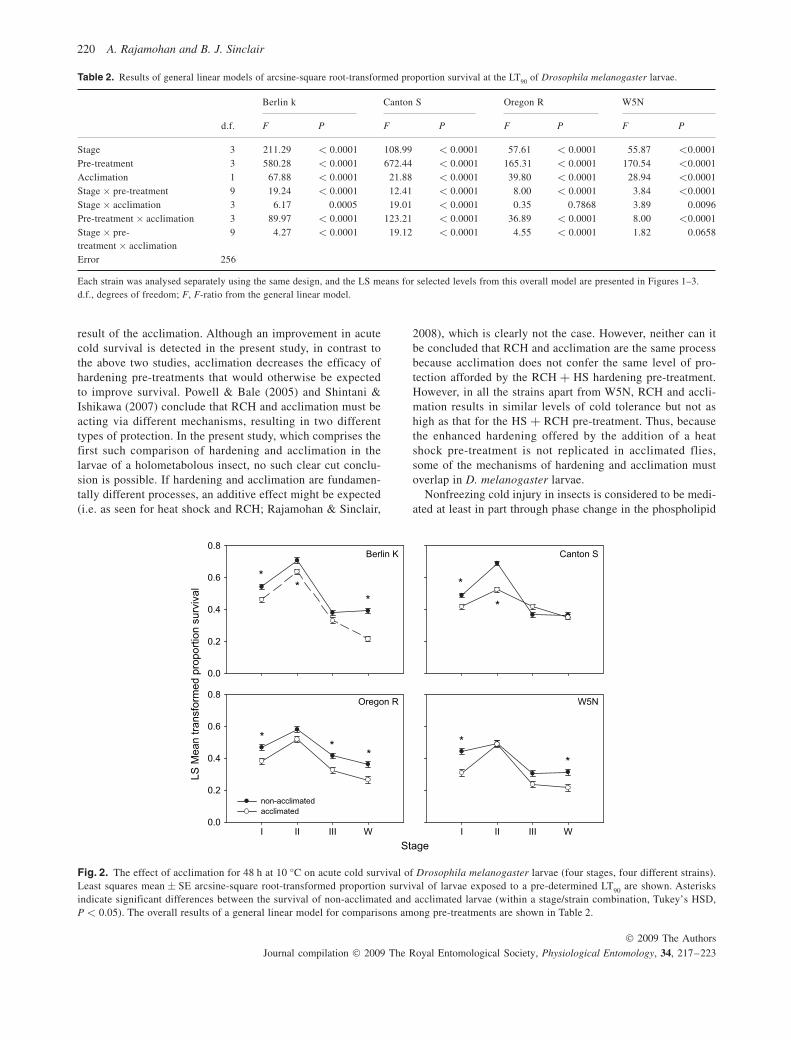
Acclimation resulted either in a significant decrease in survival after acute cold exposure in many larval stages, or no significant difference in survival between acclimated and non-acclimated larvae. This led to a significant stage × acclimation interaction in all strains, except Oregon R ( Fig. 2 and Table 2 ).
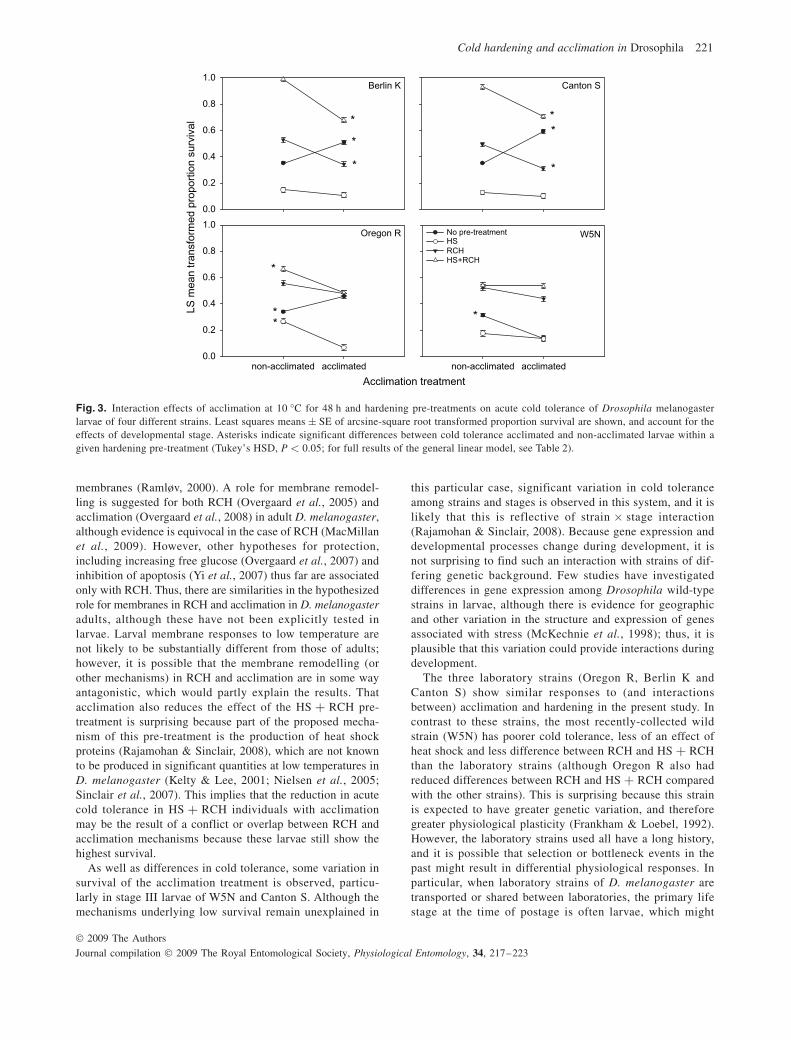
When larval stage was taken into account, acclimation sig-nificantly increased acute cold survival in control larvae of Berlin K, Canton S and Oregon R, but not W5N ( Fig. 3 ). However, acclimation resulted in no change (or a significant decrease) in survival after other pre-treatments ( Fig. 3 and Table 2 ). When hardening is taken into account in the model, a decrease in survival is observed ( Fig. 2 ), apparently contra-dicting this result. It is likely that the contrast in direction of
response to acclimation among stages and pre-treatments is driving this pattern.
Discussion
Low temperature acclimation results in a slight increase in cold tolerance in control larvae in three of four strains of flies. However, the significant interactions among stage, acclimation and pre-treatment are such that acclimation, for the most part, results in a decrease in acute cold tolerance. This conclusion is unusual because low temperature acclima-tion usually does result in improved cold tolerance in insects ( Leather et al. , 1993 ), and certainly in both larvae and adults of D. melanogaster (e.g. Hoffmann et al. , 2003; Overgaard et al. , 2006; Rako & Hoffmann, 2006 ). The key difference in the present design is the inclusion of hardening pre-treatments that result in substantial improvements in cold tolerance but which appear to override any improvement made by acclimation.
Shintani & Ishikawa (2007) report on a comparison of acclimation and hardening effects on acute cold tolerance in eggs of a longicorn beetle and show that acclimation offers cold protection that is indistinguishable from the effects of RCH (or acclimation and RCH) at all but the lowest tempera-ture under investigation (−26 °C). Powell & Bale (2005) report that nymphs of the aphid Sitobion avenae reared for three generations at 10 °C still showed an RCH response over and above any improvement in basal cold tolerance as a
Table 1. Proportion survival (mean ± SE) of Drosophila melanogaster larvae acclimated at 10 °C for 48 h.
Strain I II III W
Berlin K 0.77 ± 0.19 0.89 ± 0.11 0.97 ± 0.03 1.00 ± 0 Canton S 0.90 ± 0.10 0.94 ± 0.06 0.47 ± 0.22 0.77 ± 0.19 Oregon R 0.62 ± 0.17 0.65 ± 0.23 0.97 ± 0.03 1.00 ± 0 W5N 0.85 ± 0.12 0.90 ± 0.10 0.47 ± 0.12 0.82 ± 0.10
Three groups of 10 – 18 larvae were used in each case.
Berlin K
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4Canton S
Oregon R
StageI II III W
LS M
ean
Tra
nsfo
rmed
pro
port
ion
surv
ival
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4No pre-treatmentRCHHSHS+RCH
W5N
I II III W
nsns
ns
ns
ns
Fig. 1. The effect of hardening pre-treatment on acute cold survival of Drosophila melanogaster larvae (four stages, four different strains). Least squares mean ± SE arcsine-square root-transformed proportion survival of larvae exposed to a pre-determined LT 90 are shown. All points are significantly different from the no pre-treatment (within a stage/strain combination, Tukey’s HSD, P < 0.05), with the exception of those marked as being nonsignificant (ns). The overall results of a general linear model for comparisons among pre-treatments are shown in Table 2 . RCH, rapid cold-hardening; HS, heat shock; HS + RCH: heat shock followed by RCH.
Cold hardening and acclimation in Drosophila 219
© 2009 The Authors
Journal compilation © 2009 The Royal Entomological Society, Physiological Entomology, 34, 217–223
result of the acclimation. Although an improvement in acute cold survival is detected in the present study, in contrast to the above two studies, acclimation decreases the efficacy of hardening pre-treatments that would otherwise be expected to improve survival. Powell & Bale (2005) and Shintani & Ishikawa (2007) conclude that RCH and acclimation must be acting via different mechanisms, resulting in two different types of protection. In the present study, which comprises the first such comparison of hardening and acclimation in the larvae of a holometabolous insect, no such clear cut conclu-sion is possible. If hardening and acclimation are fundamen-tally different processes, an additive effect might be expected (i.e. as seen for heat shock and RCH; Rajamohan & Sinclair,
2008 ), which is clearly not the case. However, neither can it be concluded that RCH and acclimation are the same process because acclimation does not confer the same level of pro-tection afforded by the RCH + HS hardening pre-treatment. However, in all the strains apart from W5N, RCH and accli-mation results in similar levels of cold tolerance but not as high as that for the HS + RCH pre-treatment. Thus, because the enhanced hardening offered by the addition of a heat shock pre-treatment is not replicated in acclimated flies, some of the mechanisms of hardening and acclimation must overlap in D. melanogaster larvae.
Nonfreezing cold injury in insects is considered to be medi-ated at least in part through phase change in the phospholipid
Table 2. Results of general linear models of arcsine-square root-transformed proportion survival at the LT 90 of Drosophila melanogaster larvae.
d.f.
Berlin k Canton S Oregon R W5N
F P F P F P F P
Stage 3 211.29 < 0.0001 108.99 < 0.0001 57.61 < 0.0001 55.87 <0.0001 Pre-treatment 3 580.28 < 0.0001 672.44 < 0.0001 165.31 < 0.0001 170.54 <0.0001 Acclimation 1 67.88 < 0.0001 21.88 < 0.0001 39.80 < 0.0001 28.94 <0.0001 Stage × pre-treatment 9 19.24 < 0.0001 12.41 < 0.0001 8.00 < 0.0001 3.84 <0.0001 Stage × acclimation 3 6.17 0.0005 19.01 < 0.0001 0.35 0.7868 3.89 0.0096 Pre-treatment × acclimation 3 89.97 < 0.0001 123.21 < 0.0001 36.89 < 0.0001 8.00 <0.0001 Stage × pre-treatment × acclimation
9 4.27 < 0.0001 19.12 < 0.0001 4.55 < 0.0001 1.82 0.0658
Error 256
Each strain was analysed separately using the same design, and the LS means for selected levels from this overall model are presented in Figures 1 – 3 .d.f., degrees of freedom; F , F -ratio from the general linear model.
Berlin K Canton S
Oregon R
StageI II III W
LS M
ean
tran
sfor
med
pro
port
ion
surv
ival
0.0
0.2
0.4
0.6
0.8
0.0
0.2
0.4
0.6
0.8
non-acclimatedacclimated
W5N
I II III W
*
**
**
*
**
**
Fig. 2. The effect of acclimation for 48 h at 10 °C on acute cold survival of Drosophila melanogaster larvae (four stages, four different strains). Least squares mean ± SE arcsine-square root-transformed proportion survival of larvae exposed to a pre-determined LT 90 are shown. Asterisks indicate significant differences between the survival of non-acclimated and acclimated larvae (within a stage/strain combination, Tukey’s HSD, P < 0.05). The overall results of a general linear model for comparisons among pre-treatments are shown in Table 2 .
220 A. Rajamohan and B. J. Sinclair
© 2009 The Authors
Journal compilation © 2009 The Royal Entomological Society, Physiological Entomology, 34, 217–223
membranes ( Ramløv, 2000 ). A role for membrane remodel-ling is suggested for both RCH ( Overgaard et al. , 2005 ) and acclimation ( Overgaard et al. , 2008 ) in adult D. melanogaster , although evidence is equivocal in the case of RCH ( MacMillan et al. , 2009 ). However, other hypotheses for protection, including increasing free glucose ( Overgaard et al. , 2007 ) and inhibition of apoptosis ( Yi et al. , 2007 ) thus far are associated only with RCH. Thus, there are similarities in the hypothesized role for membranes in RCH and acclimation in D. melanogaster adults, although these have not been explicitly tested in larvae. Larval membrane responses to low temperature are not likely to be substantially different from those of adults; however, it is possible that the membrane remodelling (or other mechanisms) in RCH and acclimation are in some way antagonistic, which would partly explain the results. That acclimation also reduces the effect of the HS + RCH pre-treatment is surprising because part of the proposed mecha-nism of this pre-treatment is the production of heat shock proteins ( Rajamohan & Sinclair, 2008 ), which are not known to be produced in significant quantities at low temperatures in D. melanogaster ( Kelty & Lee, 2001; Nielsen et al. , 2005; Sinclair et al. , 2007 ). This implies that the reduction in acute cold tolerance in HS + RCH individuals with acclimation may be the result of a conflict or overlap between RCH and acclimation mechanisms because these larvae still show the highest survival.
As well as differences in cold tolerance, some variation in survival of the acclimation treatment is observed, particu-larly in stage III larvae of W5N and Canton S. Although the mechanisms underlying low survival remain unexplained in
this particular case, significant variation in cold tolerance among strains and stages is observed in this system, and it is likely that this is reflective of strain × stage interaction ( Rajamohan & Sinclair, 2008 ). Because gene expression and developmental processes change during development, it is not surprising to find such an interaction with strains of dif-fering genetic background. Few studies have investigated differences in gene expression among Drosophila wild-type strains in larvae, although there is evidence for geographic and other variation in the structure and expression of genes associated with stress ( McKechnie et al. , 1998 ); thus, it is plausible that this variation could provide interactions during development.
The three laboratory strains (Oregon R, Berlin K and Canton S) show similar responses to (and interactions between) acclimation and hardening in the present study. In contrast to these strains, the most recently-collected wild strain (W5N) has poorer cold tolerance, less of an effect of heat shock and less difference between RCH and HS + RCH than the laboratory strains (although Oregon R also had reduced differences between RCH and HS + RCH compared with the other strains). This is surprising because this strain is expected to have greater genetic variation, and therefore greater physiological plasticity ( Frankham & Loebel, 1992 ). However, the laboratory strains used all have a long history, and it is possible that selection or bottleneck events in the past might result in differential physiological responses. In particular, when laboratory strains of D. melanogaster are transported or shared between laboratories, the primary life stage at the time of postage is often larvae, which might
Berlin K Canton S
Oregon R
Acclimation treatmentnon-acclimated acclimated
LS m
ean
tran
sfor
med
pro
port
ion
surv
ival
0.0
0.2
0.4
0.6
0.8
1.0
0.0
0.2
0.4
0.6
0.8
1.0W5N
non-acclimated acclimated
No pre-treatment
RCHHS
HS+RCH
***
*
*
**
*
*
*
Fig. 3. Interaction effects of acclimation at 10 °C for 48 h and hardening pre-treatments on acute cold tolerance of Drosophila melanogaster larvae of four different strains. Least squares means ± SE of arcsine-square root transformed proportion survival are shown, and account for the effects of developmental stage. Asterisks indicate significant differences between cold tolerance acclimated and non-acclimated larvae within a given hardening pre-treatment (Tukey’s HSD, P < 0.05; for full results of the general linear model, see Table 2 ).
Cold hardening and acclimation in Drosophila 221
© 2009 The Authors
Journal compilation © 2009 The Royal Entomological Society, Physiological Entomology, 34, 217–223

explain why an effect is apparent in the larval stages. Notably, W5N is the only strain to show a consistent negative effect of acclimation when stage is taken into account. This suggests that there may be no advantage to acclimation responses to low temperatures in larvae in the wild, most likely because low temperature events signal the onset of winter and most evidence suggests that D. melanogaster overwinter in an adult reproductive diapause ( Saunders et al. , 1989; Izquierdo, 1991 ). Alternately, the advantages to induction of RCH at ecologically relevant cooling rates, as observed in adults ( Kelty & Lee, 1999 ), may render longer-term responses redundant when larvae are unlikely to experience prolonged cold periods. This contrast in the direction of responses between W5N and other strains may provide a tool for the future dissection of the mechanisms of acclimation in D. melanogaster larvae, and could also provide a system suitable for the systematic investigation of hypotheses con-cerning the evolutionary advantages of acclimation.
Conclusions
Larvae of different wild-type strains of D. melanogaster respond markedly differently to acclimation, which corre-sponds well to previous comparisons of responses to harden-ing ( Rajamohan & Sinclair, 2008 ). Although acclimation does improve survival, it has a significant negative interac-tion with hardening pre-treatments, most likely reflecting the relative importance of short- and long-term responses to cold in a relatively short-lived life stage. Overall, HS + RCH appears to be the most effective means of increasing chilling tolerance of D. melanogaster larvae.
Acknowledgements
We thank Heather Tarnowski for technical assistance, and Heath MacMillan and Katie Marshall and two anonymous referees for their constructive comments on an earlier version of the manuscript. This research was supported by the Canadian Foundation for Innovation and the Ontario Research Fund, and by grant number RR022885 to B.J.S. from the National Center for Research Resources (NCRR), a component of the National Institutes of Health (NIH). Its contents are solely the responsibility of the authors and do not necessarily represent the official views of NCRR or NIH.
References
Bale , J.S . ( 2002 ) Insects and low temperatures: from molecular biology to distributions and abundance . Philosophical Transactions of the Royal Society of London Series B, Biological Sciences , 357 , 849 – 861 .
Demerec , M . ed . ( 1965 ) Biology of Drosophila . Hafner , New York, New York .
Frankham , R. & Loebel , D.A . ( 1992 ) Modeling problems in conser-vation genetics using captive Drosophila populations – rapid genetic adaptation to captivity . Zoo Biology , 11 , 333 – 342 .
Hoffmann , A.A. , Sorensen , J.G. & Loeschcke , V . ( 2003 ) Adaptation of Drosophila to temperature extremes: bringing together quanti-tative and molecular approaches . Journal of Thermal Biology , 28 , 175 – 216 .
Izquierdo , J.I . ( 1991 ) How does Drosophila melanogaster overwin-ter . Entomologia Experimentalis Et Applicata , 59 , 51 – 58 .
Kelty , J.D. & Lee , R.E. Jr ( 1999 ) Induction of rapid cold hardening by ecologically relevant cooling rates in Drosophila melanogas-ter . Journal of Insect Physiology , 45 , 719 – 726 .
Kelty , J.D. & Lee , R.E. Jr ( 2001 ) Rapid cold-hardening of Drosoph-ila melanogaster (Diptera: Drosophilidae) during ecologically based thermoperiodic cycles . Journal of Experimental Biology , 204 , 1659 – 1666 .
Kristensen , T.N. , Hoffmann , A.A. , Overgaard , J . et al . ( 2008 ) Costs and benefits of cold acclimation in field-released Drosophila . Proceedings of the National Academy of Sciences of the United States of America , 105 , 216 – 221 .
Leather , S.R. , Walters , K.F.A. & Bale , J.S . ( 1993 ) The Ecology of Insect Overwintering . Cambridge University Press , U.K .
Lee , R.E. Jr , Chen , C.-P. & Denlinger , D.L . ( 1987 ) A rapid cold-hardening process in insects . Science , 238 , 1415 – 1417 .
MacMillan , H.A. , Guglielmo , C.G. & Sinclair , B.J . ( 2009 ) Mem-brane remodelling and glucose in Drosophila melanogaster : a test of rapid cold-hardening and chilling tolerance hypotheses . Jour-nal of Insect Physiology , 55 , 243 – 249 .
McKechnie , S.W. , Halford , M.M. , McColl , G. & Hoffmann , A.A . ( 1998 ) Both allelic variation and expression of nuclear and cy-toplasmic transcripts of Hsr-omega are closely associated with thermal phenotype in Drosophila . Proceedings of the National Academy of Sciences of the United States of America , 95 , 2423 – 2428 .
Nielsen , M.M. , Overgaard , J. , Sørensen , J.G . et al . ( 2005 ) Role of HSF activation for resistance to heat, cold and high-temperature knock-down . Journal of Insect Physiology , 51 , 1320 – 1329 .
Overgaard , J. , Sørensen , J.G. , Petersen , S.O . et al . ( 2005 ) Changes in membrane lipid composition following rapid cold hardening in Drosophila melanogaster . Journal of Insect Physiology , 51 , 1173 – 1182 .
Overgaard , J. , Sørensen , J.G. , Petersen , S.O . et al . ( 2006 ) Reorgani-zation of membrane lipids during fast and slow cold hardening in Drosophila melanogaster . Physiological Entomology , 31 , 328 – 335 .
Overgaard , J. , Malmendal , A. , Sørensen , J.G . et al . ( 2007 ) Metabolo-mic profiling of rapid cold hardening and cold shock in Drosoph-ila melanogaster . Journal of Insect Physiology , 53 , 1218 – 1232 .
Overgaard , J. , Tomcala , A. , Sørensen , J.G . et al . ( 2008 ) Effects of acclimation temperature on thermal tolerance and membrane phospholipid composition in the fruit fly Drosophila melanogaster . Journal of Insect Physiology , 54 , 619 – 629 .
Powell , S.J. & Bale , J.S . ( 2005 ) Low temperature acclimated popula-tions of the grain aphid Sitobion avenae retain ability to rapidly cold harden with enhanced fitness . Journal of Experimental Biology , 208 , 2615 – 2620 .
Rajamohan , A. & Sinclair , B.J . ( 2008 ) Short-term hardening effects on survival of acute and chronic cold exposure by Drosophila melanogaster larvae . Journal of Insect Physiology , 54 , 708 – 718 .
Rako , L. & Hoffmann , A.A . ( 2006 ) Complexity of the cold acclima-tion response in Drosophila melanogaster . Journal of Insect Phys-iology , 52 , 94 – 104 .
Ramløv , H . ( 2000 ) Aspects of natural cold tolerance in ectothermic animals . Human Reproduction , 15 ( Suppl. 5 ), 25 – 46 .
Saunders , D.S. , Henrich , V.C. & Gilbert , L.I . ( 1989 ) Induction of diapause in Drosophila melanogaster – photoperiodic regulation and the impact of arrhythmic clock mutations on time measurement .
222 A. Rajamohan and B. J. Sinclair
© 2009 The Authors
Journal compilation © 2009 The Royal Entomological Society, Physiological Entomology, 34, 217–223

Proceedings of the National Academy of Sciences of the United States of America , 86 , 3748 – 3752 .
Shintani , Y. & Ishikawa , Y . ( 2007 ) Relationship between rapid cold-hardening and cold acclimation in the eggs of the yellow-spotted longicorn beetle, Psacothea hilaris . Journal of Insect Physiology , 53 , 1055 – 1062 .
Sinclair , B.J. & Rajamohan , A . ( 2008 ) Slow or stepped rewarming after acute low-temperature exposure does not improve survival of Drosophila melanogaster larvae . Canadian Entomologist , 140 , 306 – 311 .
Sinclair , B.J. & Roberts , S.P . ( 2005 ) Acclimation, shock and harden-ing in the cold . Journal of Thermal Biology , 30 , 557 – 562 .
Sinclair , B.J. , Vernon , P. , Klok , C.J. & Chown , S.L . ( 2003 ) Insects at low temperatures: an ecological perspective . Trends in Ecology & Evolution , 18 , 257 – 262 .
Sinclair , B.J. , Gibbs , A.G. & Roberts , S.P . ( 2007 ) Gene transcrip-tion during exposure to, and recovery from, cold and desiccation stress in Drosophila melanogaster . Insect Molecular Biology , 16 , 435 – 443 .
Yi , S.X. , Moore , C.W. & Lee , R.E . ( 2007 ) Rapid cold-hardening protects Drosophila melanogaster from cold-induced apoptosis . Apoptosis , 12 , 1183 – 1193 .
Accepted 16 February 2009
Cold hardening and acclimation in Drosophila 223
© 2009 The Authors
Journal compilation © 2009 The Royal Entomological Society, Physiological Entomology, 34, 217–223
First published online 16 April 2009





























