Embed Size (px)
Citation preview

H. T. Kusumo á L. D. Druehl
Variability over space and time in the genetic structureof the winged kelp Alaria marginata
Received: 30 September 1998 /Accepted: 26 October 1999
Abstract The genetic structure of Alaria marginataPostels & Ruprecht was investigated spatially and sea-sonally using ampli®ed-fragment length polymorphisms(AFLP). Using one primer, 206 scoreable bands wereproduced. Individual plants that were separated by aslittle as a few decimeters to >100 km could be distin-guished, and followed an isolation-by-distance model.Genetic similarity (average April data) ranged from 76%for patches (a few decimeters in diameter), to 71% forindividual kelp stands (15 m across) and 67% for agroup of stands separated by 185 km. Greater geneticsimilarity of patches occurred at a wave-sheltered sitethan at a wave-exposed site. The lower wave action maylimit meiospore dispersal, resulting in patches of relatedsporophytes. Genetic similarities between A. marginatastands indicated a pattern of gene ¯ow that is consistentwith local currents. In one stand, genetic similaritieswere markedly di�erent between seasons. This seasonalpattern may be the result of di�erent age groups domi-nating the sampled stands over time. The genetic struc-ture of A. marginata comprises local scale (patch andwithin-stand) heterogeneity and larger scale (between-stands) homogeneity.
Introduction
Genetic di�erentiation re¯ects microevolutionary andecological processes. An understanding of patterns ofgenetic structure in seaweed populations provides im-portant information about factors in¯uencing gene ¯ow,such as mode and distance of dispersal and populationsize, and environmental factors in¯uencing natural se-lection, including di�erential survival or local extinction(Endler 1977, 1986; Palumbi 1994). Genetic di�erentia-tion among geographic locations has been shown forpopulations of many organisms (see reviews by Avise1994 and Palumbi 1994). Re®nement of our under-standing of genetic structure of natural populationsunder di�erent conditions is a continuing challenge(Avise 1994).
Little is known about the genetic structure ofpopulations of marine algae. Earlier studies have usedelectrophoretically detectable enzyme variation to as-sess di�erentiation among populations of marinebenthic algae, e.g. Cheney and Babbel (1978); Miuraet al. (1979); Innes (1987, 1988); Sosa and Garcia-Reina (1992); Lindstrom (1993); Pearson and Murray(1997). These studies, based on isozyme data, werelimited in sensitivity due to the limited number of thegenetic markers available for analyses and the factthat di�erences at the DNA level may not be ex-pressed at the enzyme level (McDonald and McDer-mott 1993).
DNA analyses have the potential of resolving ®negenetic structure; most studies have concentrated onlarger-scale genetic structure associated with biogeo-graphic distance (e.g. van Oppen et al. 1995; Albertoet al. 1997; Coyer et al. 1997). These studies found arelatively wide range of genetic diversity in seaweedpopulations. Pearson and Murray (1997), in an isozymeanalysis of the red alga Lithothrix aspergillum, suggestedthat the genetic variation of seaweed populations is re-lated to reproductive traits, including reproductive modeand dispersal ability. Life-cycle characteristics may
Marine Biology (2000) 136: 397±409 Ó Springer-Verlag 2000
Communicated by M. H. Horn, Fullerton
H. T. Kusumo (&)1 á L. D. DruehlDepartment of Biological Sciences,Simon Fraser University,Burnaby, British Columbia V5A 1S6, Canada
H. T. Kusumo á L. D. DruehlBam®eld Marine Station,Bam®eld, British Columbia V0R 1B0, Canada
Present address:1University of Chicago,Department of Ecology and Evolution,1101 E. 57 Street,Chicago, Illinois 60637-1503
Fax: 001 (0)773 702-9740e-mail: [email protected]

in¯uence dispersal. For example, low-level genetic dif-ferentiation was observed in exclusively sexual popula-tions ofHalidrys dioicaGardner, an obligate outcrossingseaweed with broad dispersal ability (Lu and Williams1994). In contrast, a high level of genetic di�erentiationwas found in populations of Pelvetia fastigiata (J. Ag.)De Toni, which is monoecious, with limited dispersal ofgametes and zygotes (Williams and Di Fiori 1996).Dispersal by kelp is by meiospores and, for kelp withpnematocysts or hollow stipes, dislodged macrophytesmay transport spore-bearing tissue. In Alaria esculenta(L.) Greville in Norway, meiospores were releasedwithin a 5 m radius of the sporophyte (Sundene 1962).Meiospores from a transplanted sporophyte of Macro-cystis pyrifera C. Agardh developed into gametophyteswhich produced a new generation of sporophytes withina 5 m radius of the transplant (Anderson and North1966). By clearing patches of mussels at varying dis-tances from established clumps of Postelsia palma-eformis Ruprecht, Dayton (1973) showed that dispersalof meiospores and subsequent recruitment of newsporophytes by P. palmaeformis was limited to within3 m of the clump. This ®nding was supported by Coyeret al. (1997) who, using a multilocus ®ngerprintingtechnique, found genetic similarities ranging from 77 to100% within clumps of P. palmaeformis. These studiesindicate that the dispersal distance of meiospores is verylimited in kelp.
Other studies, however, suggest that kelp may dis-perse over long distances. Long-distance dispersal in therange of kilometers was reported from colonizationstudies of Alaria sp. and other kelp, and red and greenalgae at the island of Surtsey, Iceland (reviewed byDruehl 1981a). It was not, however, resolved whetherthe dispersal was by means of spores and gametes, ordrift macrophytes. Reed et al. (1988) conducted anin situ experiment and con®rmed the existence of long-distance dispersal for Macrocystis pyrifera andPterygophora californica Ruprecht. They observed thatturbulent water ¯ow associated with winter stormscontributed to resuspension of kelp spores occurringnear the bottom and allowed them to be transportedgreater distances by prevailing currents. Reed et al.(1992) determined that the length of time thatM. pyrifera and P. californica spores could swim undervarious conditions was a maximum of 72 h. The spores,however, could continue to exist and even photosyn-thesize for longer periods. Similarly, Kain (1964) dem-onstrated the ability of Laminaria hyperborea (Gunn.)Fosl. spores to persist for periods up to 30 d in the dark.These studies suggest that kelp spores may survive long-distance transport.
Previous studies have used molecular techniques todiscern large-scale patterns of genetic di�erentiationamong kelp populations. Bhattacharya et al. (1990) as-sessed genetic variation, using restriction-fragmentlength polymorphism (RFLP) analysis of the ribosomalcistron, in sporophytes of the kelp Costaria costata(C. A. Agardh) Saunders from several locations in the
northeast Paci®c. They found evidence for genetic sub-structure among stands as close as 2 km. Mro z (1989)investigated variation in the ribosomal cistron of Alariaspp. (including A. marginata) in northern Washingtonand southern British Columbia using RFLP analyses.Based on one or a few specimens from each of severalwidely separated Alaria spp. stands, he concluded therewas essentially no intraspeci®c variation. In both thesestudies, only one or a few sporophytes were sampledfrom each location. Bhattacharya and Druehl (1989)attempted to correlate morphological and genetic vari-ation in the kelp C. costata, again employing very smallsample sizes.
Few studies have attempted to assess genetic varia-tion in kelp on small spatial scales. DNA ®ngerprintingmethods provide the opportunity of exploring the ge-netic structure of kelp sporophytes separated by only afew centimeters. The only studies conducted to date toestimate within-patch and local genetic variation in kelpwere on Macrocystis pyrifera (Coyer et al. 1994) andPostelsia palmaeformis (Coyer et al. 1997). Coyer et al.(1994) concluded that multilocus M13 ®ngerprinting canmeasure genetic variation within populations ofM. pyrifera because of the relatively small genetic vari-ation of the meiospores compared to variation observedamong plants within the population. By using multi-locus M13 DNA ®ngerprinting and RAPD (randomlyampli®ed polymorphic DNA) methods, Coyer et al.(1997) resolved genetic variation among individualsporophytes of P. palmaeformis on the scale of centi-meters (within clumps) and over 250 km (among popu-lations).
We elected to study the ®ne genetic populationstructure within and between populations of the wingedkelp Alaria marginata Postels & Ruprecht, one of thedominant seaweeds in coastal British Columbia waters.Does genetic di�erentiation vary as a function of bio-geographic scale (adjacent vs widely spaced plants)?Is there any seasonal genetic structural shift in theA. marginata populations?. Our study tests these ques-tions by examining genetic di�erentiation in the wingedkelp A. marginata as a function of its spatial and tem-poral distribution. In addition, our study explores therole of gene ¯ow in in¯uencing the genetic structure ofA. marginata populations.
The macroscopic generation of Alaria marginata is alarge (up to 2 m long) sporophyte that alternates withmicroscopic, haploid, dioecious gametophytes that de-velop from bi¯agellated meiospores (Druehl et al. 1989).The egg remains attached to the settled female gameto-phyte, where it is fertilized and subsequently developsinto the new sporophytic generation. A. marginata isconsidered biennial, with new macrophyte recruitmentmostly restricted to December/January (Widdowson1964; Lee 1965). We have, however, observed meiosporeproduction throughout the year. A. marginata is anintertidal kelp inhabiting moderately sheltered to fullywave-exposed coastal habitats. The plants are arrangedin patches, usually a few decimeters in diameter. Several
398

such patches usually make up a kelp stand, often re-stricted to the head of a small bay.
Our approach was to de®ne genetic di�erentiation atthe intra- and inter-patch level among three kelp standsup to 185 km apart. This genetic di�erentiation wasassessed using DNA markers generated by the poly-merase chain reaction (PCR)-based ®ngerprinting tech-nique, ampli®ed-fragment length polymorphisms(AFLP; Zabeau and Vos 1993).
The variety of markers available to assess genetic dif-ferentiation has been greatly expanded through PCR-based ®ngerprinting technology (Avise 1994). Recently,Zabeau and Vos (1993) developed AFLP, a PCR-based®ngerprinting technique. As a combination of classicaland PCR-based ®ngerprinting methods, AFLP is areliable, e�cient, and reproducible molecular markertechnique that can be used to genetically di�erentiateclosely related organisms such as bacterial strains(Huys et al. 1996; Janssen et al. 1996), fungal types(Mueller et al. 1996), rice (Mackill et al. 1996), soybean(Maughan et al. 1996), lentil (Sharma et al. 1996), and thered alga Chondrus crispus Stackhouse (Donaldsonet al. 1998).
As reviewed by Janssen et al. (1996), most PCR-based genomic ®ngerprinting varies in ampli®cation ef-®ciency because of small temperature changes (Meunierand Grimont 1993; Penner et al. 1993; He et al. 1994).In addition, for arbitrary primed segments of genomicDNA (RAPD), primer- and DNA concentration(Kernodle et al. 1993; MacPherson et al. 1993; Davin-Regli et al. 1995), the type of DNA polymerase used(Schierwater and Ender 1993), and DNA templatequality (Kernodle et al. 1993; Micheli et al. 1994) appearto be crucial factors that a�ect reproducibility. It is notknown to what extent mismatches between primer andtemplate are tenable, and the use of degenerate primersmay possibly cause small variations in the bandingpatterns. Further, van Oppen et al. (1996) and Coyeret al. (1997) observed that RAPD data are robust onlyfor the identi®cation of large-scale biogeographic pat-terns in populations that are hundreds to thousands ofkilometers apart.
In contrast, AFLP primers perfectly match theirtarget site because the ®rst few cycles of the reaction arecarried out under high stringency conditions (Zabeauand Vos 1993). Vos et al. (1995) reported that AFLP isinsensitive to variations in the template DNA concen-tration. Furthermore, the AFLP technique does not re-quire prior knowledge of genomic sequences and can beused on any DNA irrespective of its origin or complexity(Zabeau and Vos 1993; Vos et al. 1995). AFLP generates50 to 100 genomic fragments per reaction, of which upto 50% may be potential markers (Breyne et al. 1997).Most importantly, AFLP has been shown to be highlyreproducible after being tested in nine di�erent Euro-pean laboratories (Jones et al. 1997).
Previously, Coyer et al. (1997) pioneered explorationof the detailed genetic di�erentiation of a natural sea-weed population using DNA markers. Our investigation
paralleled this earlier study by exploring the relationshipbetween biogeographic distance and genetic di�erentia-tion in Alaria marginata. Our study expanded on that ofCoyer et al. by following seasonal changes in geneticdi�erentiation. Further, due to features of AFLP tech-nology, we were able to di�erentiate genetically adjacentplants and plants separated by 185 km using the sameDNA markers.
Materials and methods
Sampling
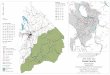
Samples of Alaria marginata Postels & Ruprecht were collectedfrom stands at Execution Rock (48°48¢N; 125°09¢W) and Kelp Bay(48°51¢N; 125°07¢W) in Barkley Sound, British Columbia, Canada(Fig. 1). Samples were also collected from Rosario Beach (48°25¢N;122°40¢W), Whidbey Island, Washington State, USA. The twoBarkley Sound stands, 6.5 km apart, were selected to representlocal extremes of an environmental gradient inhabited by A. mar-ginata. Execution Rock represented a more oceanic, wave-exposed,environment, and Kelp Bay a more wave-sheltered, estuarinecondition. A. marginata is continuously distributed between thesetwo stands (Druehl and Elliott 1996). The third stand, at RosarioBeach, 185 km distant from the Barkley Sound stands, was selectedto represent an outlying A. marginata stand.
Sporophytes of Alaria marginata were gathered from 3 to 7haphazardly selected patches across an A. marginata stand. Patchesvaried in diameter from 30 to 50 cm and sporophytes were sepa-rated in the patches by a few centimeters to 50 cm. All maturesporophytes (indicated by the presence of spore patches on spor-ophylls) were collected. Distance between patches selected in astand was '5 m, and as much as possible samples were collected ata similar intertidal level. The samples were collected in January,April and September in 1995 from Execution Rock and Kelp Bay,and in April 1995 from Rosario Beach. Samples pressed and driedon herbarium paper served as sources for DNA extraction andvoucher specimens.
For each sampling date at each stand, two patches were chosen(except the September collection at Kelp Bay, for which only onepatch was selected). The ®rst patch was chosen using randomnumbers; the second patch sampled was the third patch away fromthis, '15 m distant. As there were only three patches collected fromKelp Bay in September, only one was sampled. A total of 71sporophytes were used for genetic analyses. The number of maturesporophytes collected from the patches ranged from 4 to 7 for allstands and months.
Molecular methodology
The genetic analysis employed the ampli®ed-fragment lengthpolymorphism (AFLP) technique. The AFLP assays were per-formed with the GIBCO±BRL AFLP Analysis System II, AFLPSmall Genome Primer Kit (Catalogue 10717-015, GIBCO±BRL),as recommended by the manufacturer. Preliminary analyses weredone to determine the primer pair appropriate for the selectiveampli®cation step, based on the number of ®ngerprinting bandsand level of variations between samples. EcoRI-AG and MseI-CAG (GIBCO±BRL) was the primer pair selected for theanalysis.
DNA extraction
Various DNA-extraction methods were tried, such as the protein-ase K (Mayes et al. 1992), CTAB (Stewart and Via 1993), LiCl(Palumbi 1996) and CsCl (Fain et al. 1988) methods. For reasons
399

of reproducibility and consistency, the CsCl method was selected toextract the DNA. The CsCl DNA-extraction method followed theprotocols of Fain et al., which were scaled down to 1 g dry tissuefor each sample. The CsCl extraction method gave reproduciblebanding patterns. This was con®rmed by running the same sampleswith variations on AFLP procedures and concentrations of DNAtemplate.
Gel analyses
Each sample of 3 lL was loaded on 6% polyacrylamide gel (20:1acrylamide: bis; 7.5 M urea; 1´ TBE bu�er) with a total of up to 30samples run on each gel. Each of the 71 samples was run two orthree times on separate gels. For alignment purposes, at least twostandard ®ngerprint-samples were run on each gel. These gels wereelectrophoresed at 60 W for 2 h (until the xylene cyanol/slower dyewas two-thirds down the length of the gel). The gel was dried onBio Rad Gel Dryer Model 583, and exposed to x-ray Kodak XAR®lm at ±70 °C for 48 h. The ®lm was developed using Kodak de-veloper and ®xer.
Data analyses
Two approaches were taken to assess genetic relationships. The®rst, using a similarity coe�cient, was used to view relationshipsbetween individuals and within, as well as between, patches andstands. The second, based on an analysis of molecular variance(AMOVA), was used to determine the genetic structure of popu-lations (Exco�er et al. 1992).
Fingerprinting patterns on the X-ray ®lm were analyzed man-ually. Each ®ngerprinting band was denoted as either present orabsent. For statistical analysis, genetic similarity was de®ned as acorrelation coe�cient (Upholt 1977; Nei and Li 1979). This coef-®cient was designed to calculate the proportion of fragments sharedin DNA digestion pro®les of di�erent plants. The correlation co-e�cient (S) was calculated as:
S � 2Nxy
Nx � Ny;
where: Nx, Ny, and Nxy are the number of bands observed in PlantsX, Y, and shared by X and Y, respectively.This genetic similarity
Fig. 1 Locations of collectionstands of Alaria marginata atExecution Rock and Kelp Bay(Barkley Sound, Vancouver Is-land, British Columbia, Cana-da), and Rosario Beach(Washington State, USA) [Ar-rows show net current direction(Doe 1952); Enlarged areaBarkley Sound]
400

correlation-coe�cient has been used by others to evaluate AFLPdata (Sharma et al. 1996; Roa et al. 1997).
The coe�cient-similarity matrices obtained were used in mul-tidimensional-scaling (MDS) analyses to present sample diversitygraphically.
The AMOVA was employed to elucidate spatial and temporalgenetic-structure of Alaria marginata. AMOVA estimates genetic-structure indices using information on the allelic content ofhaplotypes as well as their frequencies (Exco�er et al. 1992). Theinformation on the di�erences in allelic content between haplotypesis entered as a matrix of euclidean squared distances (Schneideret al. 1997). For the spatial genetic structure, variance was parti-tioned into three levels: among plants within patches, betweenpatches within stands, and among stands. For the temporal geneticstructure, variance was partitioned into among plants within col-lection times, and among collection times. Signi®cance values wereassigned to variance components on the basis of a set of null dis-tributions generated by a non-parametric permutation processwhich drew 16 000 independent samples from the distance matrix,and randomly reallocated plants into stands and patches (forspatial-variability analysis), or into collection times (for temporal-variability analysis). These procedures were done using ArlequinVersion 1.1 software package (Schneider et al. 1997).
Spatial distribution data were employed in four sets of analyses:(1) January data for Execution Rock and Kelp Bay; (2) April datafor Execution Rock and Kelp Bay; (3) September data for Exe-cution Rock and Kelp Bay; (4) April data for Execution Rock,Kelp Bay, and Rosario Beach. Temporal distribution data con-sisted of two sets of analyses: all Execution Rock data and all KelpBay data.
Results
Molecular method
The primer pair generated a total of 206 score-able ®n-gerprinting bands from the AFLP analysis of Alariamarginate. Duplicate gels and triplicate gels for each ofthe 71 samples rendered identical bands in every case(Fig. 2).
Genetic variation
Pairwise genetic comparisons
AFLP technology genetically di�erentiated betweenadjacent plants and also between plants separated by185 km. An example of the genetic similarity coe�cientmatrices generated from the 71 macrophytes analyzed ispresented in Fig. 3. These coe�cients varied widely,ranging from 28% between the April collection (Fig.3)from Execution Rock [Patch 1, Plant No. 5 (E15)] andRosario Beach [Patch 1, Plant No. 4 (R14)] to 100%between an Execution Rock and a Kelp Bay plant in theSeptember collection (data not shown). Of the twoplants (E15, R14: Fig. 3) with the lowest genetic simi-larity coe�cient (28%), R14 consistently demonstratedthe lowest values when paired with other members of itspatch and with all members from the other patches andstands sampled in April. One of the two plants that gaverise to the highest genetic similarity coe�cient (100%)
consistently elevated the coe�cients when paired withother members of its patch (Kelp Bay).
Within-stand and -patch comparisons
Genetic similarities generally increased as the geographicsize of the assessed area decreased (Table 1). For in-stance, the genetic similarity of April collections was67% for Execution Rock, Kelp Bay and Rosario Beachcombined (encompassing a distance of 185 km); it in-creased to 69% for Execution Rock and Kelp Baycombined (encompassing a distance of 6.5 km); it fur-ther increased to 71 and 74% for Rosario Beach andKelp Bay, respectively, when considered separately (en-compassing a distance of �15 m), but for ExecutionRock it decreased to 68%; ®nally it increased to 74, 81,and 74% for within-patch variations at Execution Rock,Kelp Bay, and Rosario Beach, respectively. Similarpatterns were observed for January, September, and allcollections combined (Table 1).
MDS analyses are visualized as clusters based ongenetic similarity coe�cients (Fig. 4). Clusters weretighter within the Kelp Bay stand (re¯ecting highersimilarity coe�cients) than within Execution Rock andRosario Beach stands. Generally, within-patch similaritywas greater for Kelp Bay (mean 78%) than for Execu-tion Rock (mean 69.7%), with some exceptions (Fig. 4).There were di�erent degrees of clustering betweenpatches within each stand for each sampling date. Forexample, in January, Kelp Bay Patch 3 was looselyclustered relative to Patch 6 (Fig. 4). Comparison ofgenetic similarities between patches within each standfor each sampling date suggested di�erent degrees ofclustering. Close examination of MDS data revealedthat Patch 3 January samples for Kelp Bay were looselyclustered relative to Patch 6 samples, which was tightlyclustered (Fig. 4).
Between-stand and -patch comparisons
Genetic similarity between stands and patches also re-¯ected the geographic distance between the populationunits compared (Figs. 5 and 6). The Execution Rockwithin-stand genetic similarity was generally lower thanthe similarity between Execution Rock and Kelp Baystands and the Kelp Bay within-stand similarity wasgreater than the between-stand genetic similarity. Be-tween-stand genetic similarity decreased with increasinggeographic distance. Rosario Beach had genetic simi-larities of 63 and 65% between Execution Rock andKelp Bay, respectively, whereas Execution Rock andKelp Bay had a genetic similarity between them of 68%for the April collection (Fig. 5). This trend was followedfor other collection times (Fig. 5).
Between-patch genetic similarities were lower thaneither Execution Rock and Kelp Bay within-patch ge-netic similarities in three cases; in the remaining three
401

cases the between-patch genetic similarities were higherthan the Execution Rock and lower than the Kelp Baywithin-patch similarities (Fig. 6).
Genetic structure
AMOVA analyses revealed signi®cant di�erences in ge-netic structure among plants within patches and amongplants between patches, but not among plants betweenstands. Spatial genetic variation of Alaria marginata wasmainly due to variation of plants within patches (67 to88%), variance between patches within stands accountedfor 15 to 39%, and variance among stands accounted for0 to 7% (Table 2). F-statistics revealed signi®cant dif-ferences between patches within stands (FSC; p < 0.01)but there was no signi®cant di�erence between plantsamong stands (FCT; p � 0.01). When individuals wererandomly compared across the whole study area without
regard to their original position, signi®cant structuraldi�erence was indicated (FST; p < 0.01). These resultswere consistent for all three collection occasions inBarkley Sound as well as for the single Barkley Soundplus Rosario Beach collection (Table 2).
AMOVA analysis of temporal genetic variability in-dicated signi®cant seasonal di�erences in genetic struc-ture for Kelp Bay but not for Execution Rock (Table 3;p � 0.01). At Kelp Bay, 29% of the genetic variance ob-served was due to di�erent collection times, and at Exe-cution Rock only 6% of the variance was accounted for.
Discussion and conclusions
Our results revealed the following noteworthy ®ndings.Genetic similarity coe�cients increased consistently asthe geographic units being considered decreased in
Fig. 2 Alaria marginata. Sam-ple autoradiograph depictingampli®ed fragment lengthpolymorphisms constructedusing single PCR-primer com-bination (EcoRI±AG andMseI±CAG). Plants weresampled from Execution Rock(E) and Kelp Bay (K) [STDstandard ®ngerprint samples;numbers following site abbre-viations collection time(1 January, 2 April, 3 Sep-tember), patch number, andplant number, respectively(bands below arrow are thoseused in present analysis)]
402

scale, with the greatest similarity occurring within Al-aria marginata patches and the lowest genetic similar-ity occurring among all stands together. There wassigni®cant di�erence in genetic structuring amongplants in patches and among patches within stands,but there was no signi®cant di�erence between plantsof di�erent stands. There was a signi®cant temporalchange in the genetic structuring of the A. marginatastands. These areas will be the focus of the followingdiscussion.
AFLP approach
AFLP revealed genetic di�erentiation among adjacentstands, patches and even within patches of Alaria mar-ginata. The 206 scoreable bands generated from a primerpair provided a powerful tool for resolving populationstructure at the levels investigated. To our knowledge,this is the ®rst comprehensive application of AFLPtechnology to a marine population study. Coyer et al.(1997) required two genetic marker systems (M13minisatelite ®ngerprinting and RAPD) to resolve geneticdi�erentiation in the kelp Postelsia palmaeformis over asimilar geographic scale. Donaldson et al. (1998) suc-
cessfully explored the application of AFLP technologyin resolving genetic di�erences among six plants of a redalga from three locations.
Fig. 3 Alaria marginata. Genetic similarity coe�cients (%) frompairwise comparisons of stands in Execution Rock (E), Kelp Bay (K)and Rosario Beach (R) in April 1995. Patch number and plantnumber follow each site abbreviation, in that order
Table 1 Alaria marginata. Summary of genetic similarity coe�-cients (%, �SE) for stands, patches and months for ExecutionRock (E), Kelp Bay (K), and Rosario Beach (R) (All coe�cientscalculated for all 3 mo combined) Coe�cients derived from in-dividual pairwise values presented in Fig. 3. (N = 10 to 14 forstands, except for Kelp Bay in September, where N = 5, andN = 4 to 7 for patches)
Sampling site Jan Apr Sep All
StandsE, K, R 67 � 0.5E, K 67 � 0.7 69 � 0.9 63 � 1.4 64 � 0.3E 65 � 1.8 68 � 1.9 58 � 1.5 64 � 1.1K 71 � 1.0 74 � 1.5 82 � 4.7 73 � 0.9R 71 � 1.7
PatchesExecution Rock 62 � 3.3 78 � 2.3 62 � 4.1
75 � 2.3 70 � 6.4 67 � 2.1(av) (69) (74) (66) (70)
Kelp Bay 65 � 1.7 76 � 2.8 82 � 4.780 � 1.4 86 � 1.7
(av) (73) (81) (82) (79)
Rosario Beach 63 � 3.185 � 1.0
(av) (74) (74)
403

Genetic variation
Pairwise genetic comparisons
The lowest genetic similarity value (28%) resulted fromgenetic pairing of a Rosario Beach and Execution Rockplant. The possibility exists that the Rosario Beach plantgenetically represented a second species, Alaria tenuifoliaSetchell. Widdowson (1971) described plants fromshores adjacent to Rosario Beach as being morpho-logically intermediate between A. marginata andA. tenuifolia. A review of our voucher specimens for theRosario beach collection did not suggest apparentmorphological deviation of this plant from the otherRosario Beach plants. The possibility exists, however,that this plant may be a hybrid. Laboratory-producedmorphological hybrids and, in one case, a naturallyoccurring hybrid, are well documented between kelpspecies and genera (see Lewis 1996 for review). In oneinstance, parentally-distinct DNA sequences were iden-ti®ed in a laboratory cross between two kelp species[Lessoniopsis littoralis (Tilden) Reinke x A. marginata:Liptack and Druehl 1999].
A pairing of a Kelp Bay and Execution Rock plantresulted in a 100% genetic similarity coe�cient. We do
not consider that this similarity coe�cient representedan identical genetic match, but rather an identical matchof the 206 employed genetic markers. It is possible thatthis 100% value was a ``book-keeping'' error. However,we consider this unlikely, as other pairwise comparisonsbetween members of the same Kelp Bay patch revealedhigh similarity values. There have been other reports of100% genetic similarity. Identical ®ngerprint-bandingpatterns have been described for an entire population ofChannel Island fox (Gilbert et al. 1990) and for blackpoplar trees (Win®eld et al. 1998). Using M13 ®nger-printing data, Coyer et al. (1997) identi®ed three plantsof the kelp Postelsia palmaeformis in one cluster, andtwo plants from each of two other clusters with geneticsimilarity coe�cients of 100%. Coyer et al.'s (1997)clusters di�ered from our patches in that the clusteredP. palmaeformis had coalesced holdfasts, whereas in theAlaria marginata patches the plants were distinct fromeach other. Due to the close proximity of the identicalP. palmaeformis plants, there is a greater possibility thatthey represented identical genotypes. There are ways inwhich genetically identical kelp sporophytes may beproduced. Parthenogenesis has frequently been observed
Fig. 4 Alaria marginata. Multidimensional scaling of genetic similar-ities at Execution Rock and Kelp Bay (First number patch number;second number plant number) Fig. 5 Alaria marginata. Genetic similarity coe�cients (% � SE)
within stands at Execution Rock, Kelp Bay and Rosario Beach within(ovals) and between (boxes) stands (N = 10 to 14 for monthlysamples, except for Kelp Bay in September, where N = 5)
404

under laboratory conditions, with the potential of morethan one parthenosporophyte arising from the samegametophyte (Lewis 1996). Also, identical sporophyteshave been produced in the laboratory by crossing clonedgametophytes (Druehl and Boal 1981). Presumably, aparallel process could occur under ®eld conditions.However, the distance (6.5 km) between the pairedplants which gave rise to a 100% similarity coe�cient inour study suggests that this was unlikely.
Pairwise similarity values obtained in earlier studiesusing the AFLP technique fall within the range of ourvalues (28 to 100%). Genetic similarity in cultivars oflentil ranged from 83 to 94% (Sharma et al. 1996). Therange of genetic similarity coe�cients within and be-tween wild and cultivated species of soybean was 60 to99% (Maughan et al. 1996). For cassava, intraspeci®cand interspeci®c variation in similarity coe�cients ran-ged from 70 to 80% and 26 to 70%, respectively (Roaet al. 1997).
Within-stand and -patch comparisons
Genetic variation in Alaria marginata, expressed as ge-netic similarity, was consistent insofar as the similarityincreased as the geographic units compared decreased inscale (Table 1). This is a reasonable trend, suggesting adecreasing genetic similarity, most probably associated
Fig. 6 Alaria marginata. Genetic similarity coe�cients (% � SE)within patches at Execution Rock, Kelp Bay and Rosario Beachwithin (ovals) and between (boxes) patches (N � 4 to 7)
Table 2 Alaria marginata. Spatial analysis of variance (AMOVA)of kelp collected from Execution Rock (E), Kelp Bay (K) andRosario Beach (R) (p probability of having more extreme variancecomponent and F-statistic than observed values by chance alone).FCT (®xation index representing di�erences between individualsamong stands) and s2CT (sum of variances arising from di�erencesbetween individuals among stands) were tested under randompermutation of whole populations across stands. FSC (®xation in-dex representing di�erences between individuals among patcheswithin stands) and s2SC (sum of variances arising from di�erences
between individuals among patches within stands) were tested un-der random permutation of individuals across patches but withinsame stand. FST (®xation index representing di�erences betweenindividuals among patches regardless of stands) and s2ST (sum ofvariances arising from di�erences between individuals among pat-ches regardless of stands) were tested under random permutation ofindividuals across stands regardless of source-stands or patch(N = 10 to 14 for stands, except for Kelp Bay in September, whereN = 5, and N = 4 to 7 for patches)
Variance component Observed partition p F-statistics
variance % total
E, K (Jan)among stands s2CT = )0.32 )3.00 0.3380 FCT = )0.030among patches/stands s2SC = 1.58 15.02 0.0025 FSC = 0.146
within patches s2ST = 9.23 87.98 0.0002 FST = 0.120
E, K (Apr)among stands s2CT = )0.67 5.84 0.3390 FCT = )0.058among patches/stands s2SC = 4.43 38.55 0.0003 FSC = 0.364
within patches s2ST = 7.73 67.29 0.0000 FST = 0.327
E, K (Sep)among stands s2CT = )1.82 16.18 0.3355 FCT = )0.162among patches/stands s2SC = 3.83 34.08 0.0081 FSC = 0.293
within patches s2ST = 9.23 82.10 0.0008 FST = 0.179
E, K, R (Apr)among stands s2CT = 0.83 6.77 0.1139 FCT = 0.068
among patches/stands s2SC = 3.25 26.47 0.0000 FSC = 0.284
within patches s2ST = 8.21 66.76 0.0000 FST = 0.332
405

with increased di�culty in gene transport with increas-ing distance. Our study revealed relatively lower geneticsimilarity in A. marginata populations than the study onthe kelp Postelsia palmaeformis (Coyer et al. 1997).Coyer et al. determined within-patch variation andvariation between patches 250 km apart. Within patch-es, the genetic similarity coe�cient ranged from 77 to100% (5 out of 6 patches were 90%): between distantstands (250 km separation) it was 56%. Two featuresmay be responsible for the di�erent genetic similaritycoe�cients recorded in Coyer et al.'s study and our own:dispersal mechanisms and environmental conditions.P. palmaeformis releases its spores while the plant isemerged; thus, the spores from the parent sporophytehave the opportunity of becoming concentrated andattached at very close proximity to the sporulatingsporophyte (Dayton 1973; Paine 1979, 1988). When andhow A. marginata sporulates is not known. On the basisof transplant studies, Sundene (1962) suggested, thatdispersal of A. esculenta (L.) Greville was restricted to5 m, and that because of this kelp patches could containnumerous siblings.
The Postelsia palmaeformis stands studied by Coyeret al. (1997) occupied fully wave-exposed habitats thatmay be more environmentally uniform than our studyareas, which were distinguished by varying degrees ofwave exposure (Druehl 1981b). However, the di�erencesbetween Coyer et al.'s study and our own may alsopartially result from sensitivity di�erences between themarkers used (M13 ®ngerprinting and RAPD vs AFLP).
The degree of genetic di�erentiation in Kelp Bayplants was consistently higher than in Execution Rockplants for all three sampling periods. The MDS analysisshowed that plants from Kelp Bay clustered into small,genetically-distinct groups, whereas those from Execu-tion Rock did not cluster as strongly into genetically-distinct patches. Within-stand, between-plant clustervariation in genetic similarity exhibited heterogeneity ingenetic cluster structure. Thus, whereas Kelp Bay tendedto be tightly clustered genetically, Execution Rock wasrelatively loosely clustered. Plant patches could varyfrom the stand norm, which may have re¯ected the smallwithin-patch sample size.
These genetic di�erences might be explained by theenvironmental di�erences between the two stands. Waveaction, which was relatively high at Execution Rock,could improve local spore dispersal, resulting in a moreuniform distribution of genotypes at Execution Rock.The relatively low wave action at Kelp Bay, could resultin limited spore dispersal and the establishment of ge-netically distinct patches. Many brown algae have beenshown to possess relatively limited dispersal abilities(Sundene 1962; Anderson and North 1966; Dayton1973; Norton 1981; Kendrick and Walker 1991).
Between-stand and -patch comparisons
The pattern of between-stand genetic similarities sug-gests there may be di�erent degrees of gene ¯ow betweenExecution Rock and Kelp Bay (Fig. 5). The geneticsimilarities between these two areas were either higher orequal to the within-Execution Rock similarities and al-ways lower than within-Kelp Bay similarities. Thissuggests a net gene ¯ow from Execution Rock to KelpBay. Conversely, the genetic similarities between Rosa-rio Beach and either Execution Rock or Kelp Bay werelower than within-Rosario Beach, Execution Rock, orKelp Bay genetic similarities, suggesting no substantialnet gene ¯ow over this greater distance (185 km). Theappearance of a net gene ¯ow may be the result of other,unmonitored, gene pools impacting on the study sites.
Between-patch genetic similarity coe�cients suggest-ed in three cases a net gene ¯ow between patches. Thiswas indicated by the between-patch genetic similaritycoe�cient being higher than one of the within-patchsimilarity coe�cients) (for example, see the Januarysamples for Execution Rock and Kelp Bay: Fig. 6). Inthe three remaining cases, there was no evidence for netgene ¯ow.
Local current ¯ow was an environmental factor thatmay have contributed to the patterns of genetic between-stand similarities observed for Execution Rock and KelpBay plants. Doe (1952) showed that the net current ¯owin Barkley Sound was counter-clockwise from TrevorChannel, across the head of Barkley Sound to ImperialEagle Channel, and out to the open Paci®c (Fig. 1).Thus, the net current between the two stands wouldtravel from Execution Rock to Kelp Bay. This currentdirection may favor spore dispersal or gene ¯ow fromExecution Rock to Kelp Bay. Being down-current to thenet ¯ow could account for the genetic distinctivenessbetween the Kelp Bay plants in contrast to the Execu-tion Rock plants. This simple explanation, however,does not take into account local back-eddies and tidalcurrents which could homogenize spore dispersal in bothdirections between Execution Rock and Kelp Bay. Thepossibility of long-range dispersal (km vs m) could beenhanced and biased towards an Execution Rock toKelp Bay transport by the potential for prolonged sporesuspension in the more wave-exposed seas of ExecutionRock. Reed et al. (1988) suggested that such a mecha-
Table 3 Alaria marginata. Temporal analysis of variance (AMO-VA) of genetic diversity of kelp collected in January, April andSeptember from Execution Rock and Kelp Bay (N within collectiontimes =10 to 14, except for Kelp Bay in September, where N = 5;N among collection times =33 for Execution Rock and 26 for KelpBay)
Variance component Observed partition p F-statistics
variance % total
Execution Rockamong collection times 0.71 6.01 0.0123 0.060within collection times 11.07 93.99
Kelp Bayamong collection times 3.85 29.39 0.0000 0.294w ithin collection times 9.26 70.61
406

nism may be important in the spore transport of otherkelp species.
Genetic structure
The pattern of genetic structure variation in Alariamarginata in this study, which was characterized by ge-netic heterogeneity at the local scale of patch and stand,and genetic homogeneity at the larger scale of betweenstands has been observed for a limpet and for a seaurchin (Johnson and Black 1984; Watts et al. 1990).Two possible explanations, post-recruitment selectionand pre-recruitment selection combined with the e�ectsof di�erent patches having di�erent histories of re-cruitment have been advanced to explain this structuralpattern (Watts et al. 1990).
If post-recruitment selection were the main force indetermining genetic structure of Alaria marginata pop-ulations, and selection pressure were assumed to berelatively constant at a particular patch/stand, one couldexpect a genetic shift in allele frequencies between gen-erations (Watts et al. 1990). Based on Lee's (1965) ob-servations on A. marginata recruitment, the mature(with sporophylls) plants we collected in January andApril represented an age class extending from the pre-vious year. By September, this age class would have beensupplemented by ®rst-year plants recruited earlier in theyear that have become mature. The temporal datashowed signi®cant di�erences between collection timesfor Kelp Bay, but not for Execution Rock. The absenceof temporal genetic variation at Execution Rock sug-gests that the genetic variation of A. marginata could bemore a response to pre-recruitment selection and/or thee�ect of di�erent patches having di�erent histories ofrecruitment than to post-recruitment selection.
The low genetic structural variation among standswas probably due to e�ective long-distance spore dis-persal, which has been demonstrated for other kelpspecies (Reed et al. 1988) and marine organisms withfree-¯oating spore/larvae stages (Scheltema 1971; Berger1973; Winans 1980; Johnson and Black 1984; Waples1987; Watts et al. 1990). Allendorf (1983) estimated thatgenetic exchange of roughly one individual betweenpopulations per generation would eliminate any geneticdi�erentiation.
Our data suggest that water movement may be par-amount in determining genetic variation at the geo-graphic scales we investigated. At the microscale (withinand between patches, cm to m), the degree of turbulence,resulting from wave action, may dictate the pattern ofmeiospore dispersal; at the mesoscale (tens to thousandsof meters), local currents may dominate. This assumesthat the plants in question are totally passive in theirdispersal. Norton (1992) has noted that, once released,seaweed propagules have little ``control over their des-tiny''. However, the plants may have strategies to opti-mize their dispersal. Meiospores may be released duringtimes of calm or turbulence or during ¯ooding or ebbing
tides. The intertidal Postelsia palmaeformis releases itsspores when the tide is out, thereby avoiding the tur-bulence associated with its open-ocean habitat (Dayton1973; Paine 1979, 1988). Such behavioral options maychange dispersal distances and directions markedly(Downes and Keough 1998). To understand the relativeroles of environment and behavior in dispersal will re-quire speci®cally designed studies. Our study has dem-onstrated that the degree of genetic resolution requiredfor such studies can be achieved through AFLP tech-nology.
Statistical consideration
Our study has provided a data set that allowed us tore¯ect on genetic variation as a function of space andtime. We were, however, only able to suggest environ-mental relationships based on consistent trends. Forexample, the genetic di�erences we observed between thewave-exposed Execution Rock and the wave-shelteredKelp Bay plants apply only to those speci®c stands. Theactual cause of the observed genetic structuring is notknown. To de®ne more exactly the impact of variousparameters on the observed genetic structuring wouldrequire su�cient replication of stand types (Hurlbert1984).
Acknowledgements We wish to thank Drs. A.T. Beckenbach,R.E. deWreede, A.R. Palmer, M.J. Smith, D. R. Levitan, andR. Rochette for their helpful comments on this manuscript.Ms K. Beckenbach and Dr. D.L. Baillie provided critical assistanceat various stages of this research. We also acknowledge the signi-®cant contributions of the anonymous reviewers. The Higher Ed-ucation Project ADB Loan No. 1253-INO (The Ministry ofEducation and Culture, Directorate General of Higher EducationRepublic of Indonesia), Simon Fraser University, and Bam®eldMarine Station provided scholarship funds to HTK. This researchwas conducted at the Bam®eld Marine Station and the Institute ofMolecular Biology and Biochemistry, Simon Fraser University,and funded by a NSERC Canada operating grant to LDD.All experimental procedures conducted comply with the laws ofCanada.
References
Alberto F, Santos R, LeitaÄ o JM (1997) DNA extraction andRAPD markers to assess the genetic similarity among Gelidiumsesquipedale (Rhodophyta) populations. J Phycol 33: 706±710
Allendorf FW (1983) Isolation, gene ¯ow, and genetic di�erentia-tion among populations. In: Schonewald-Cox CM, ChambersSM, MacBryde B, Thomas L (eds) Genetics and conservation.Benyamin/Cummings, London, pp 51±65
Anderson EK, North WJ (1966) In situ studies of spore productionand dispersal in the giant kelp Macrocystis. Proc int SeaweedSymp 5: 73±86 [Young EG, McLochlan JL (eds) PergamonPress, Oxford]
Avise JC (1994) Molecular markers, natural history and evolution.Chapman & Hall, New York
Berger E (1973) Gene-enzyme variation in three sympatric speciesof Littorina. Biol Bull mar biol Lab, Woods Hole 145: 83±90
Bhattacharya D, Baillie DL, Druehl LD (1990) Population analysisof the kelp Costaria costata (Phaeophyta) using a polymorphicribosomal DNA probe. Pl Syst Evolut 170: 177±191
407

Bhattacharya D, Druehl LD (1989) Morphological, DNA sequencevariation in the kelp Costaria costata (Phaeophyta). Mar Biol102: 15±23
Breyne P, Boerjan W, Gerats T, Van Montagu M, Van Gysel A(1997) Applications of AFLP in plant breeding, molecular bi-ology and genetics. Belg J Bot 129: 107±117
Cheney DP, Babbel GR (1978) Biosystematic studies of the redalgal genus Euchema. I. Electrophoretic variation amongFlorida populations. Mar Biol 47: 251±264
Coyer JA, Olsen JL, Stam WT (1997) Genetic variability andseparation in the sea palm kelp Postelsia palmaeformis(Phaeophyceae) as assessed with M13 ®ngerprints and RAPDS.J Phycol 33: 561±568
Coyer JA, Robertson DL, Alberte RS (1994) Genetic vari-ability within a population and between diploid/haploidtissue of Macrocystis pyrifera (Phaeophyceae). J Phycol 30:545±552
Davin-Regli A, Abed Y, Charrel RN, Bollet C, de Micco P (1995)Variations in DNA concentrations signi®cantly a�ect the re-producibility of RAPD ®ngerprinting patterns. Res Microbiol146: 561±568
Dayton PK (1973) Dispersion, dispersal, and persistence of theannual intertidal alga, Postelsia palmaeformis Ruprecht. Ecol-ogy 54: 433±438
Doe LAE (1952) Currents and net transport in Loudoun Channel,April 1950. J Fish Res Bd Can 9: 42±64
Donaldson S, Chopin T, Saunders GW (1998) Ampli®ed fragmentlength polymorphism (AFLP) as a source of genetic markers forred algae. J appl Phycol 10: 365±370
Downes BJ, Keough MJ (1998) Scaling of colonization process instreams: parallel and lessons from marine hard substrata. AustJ Ecol 23: 8±26
Druehl LD (1981a) Geographical distribution. Bot Monogr 17:306±325
Druehl LD (1981b) The development of an edible kelp culturetechnology for British Columbia. British Columbia. MarineResources Branch, Ministry of Environment, Victoria, BC(Fish Dev Rep No. 32)
Druehl LD, Boal R (1981) Manipulation of the laminarialean lifecycle and its consequences for kombu mariculture. Proc intSeaweed Symp 10: 575±580 [Levring T(ed) Walter de Grayter,Berlin]
Druehl LD, Elliott CTJ (1996) Parks Canada Barkley Sound. Kelpdistribution. Part 2. Park Canada kelp distribution. Bam®eldMarine Station, Bam®eld, British Columbia
Druehl LD, Robertson BR, Button DK (1989) Characterizing andsexing laminarialean meiospores by ¯ow cytometry. Mar Biol101: 451±456
Endler JA (1977) Geographic variation, speciation, and clines.Princeton University Press, Princeton, New Jersey
Endler JA (1986) Natural selection in the wild. Princeton Univer-sity Press, Princeton, New Jersey
Exco�er L, Smouse PE, Quattro JM (1992) Analysis of molecularvariance inferred from metric distances among DNA haplo-types: application to human mitochondrial DNA restrictiondata. Genetics, Austin, Tex 131: 479±491
Fain SR, Druehl LD, Baillie DL (1988) Repeat and single copysequences are di�erentially conserved in the evolution of kelpchloroplast DNA. J Phycol 24: 292±302
Gilbert DA, Lehman N, O'Brien SJ, Wayne RK (1990) Genetic®ngerprinting re¯ects population di�erentiation in the Califor-nia Channel Island fox. Nature, Lond 344: 764±767
He Q, Viljanen M, Mertsola J (1994) E�ects of thermocyclers andprimers on the reproducibility of banding patterns in randomlyampli®ed polymorphic DNA analysis. Molec Cell Probes 8:155±160
Hurlbert SH (1984) Pseudoreplication and design of ecological ®eldexperiments. Ecol Monogr 54: 187±211
Huys G, Coopman R, Janssen P, Kersters K (1996) High-resolu-tion genotypic analysis of the genus Aeromonas by AFLP ®n-gerprinting. Int J system Bact 46: 572±580
Innes DJ (1987) Genetic structure of a sexually reproducing En-teromorpha linza (Ulvales: Chlorophyta) in Long Island Sound.Mar Biol 94: 459±467
Innes DJ (1988) Genetic di�erentiation in the intertidal zone inpopulations of the alga Enteromorpha linza (Ulvales: Chloro-phyta). Mar Biol 97: 9±16
Janssen P, Coopman R, Huys G, Swings J, Bleeker M, Vos P,Zabeau M, Kersters K (1996) Evaluation of the DNA ®nger-printing method AFLP as a new tool in bacterial taxonomy.Microbiology 142: 1881±1893
Johnson MS, Black R (1984) Pattern beneath the chaos: the e�ectof recruitment on genetic patchiness in an intertidal limpet.Evolution 38: 1371±1383
Jones CJ, Edwards KJ, Castaglione S, Win®eld MO, Sala F, van deWiel C, Bredemeijer G, Vosman B, Matthes M, Daly A,Brettschneider R, Bettini P, Buiatti M, Maestri E, MalcevschiA, Marmiroli N, Aert R, Volckaert G, Rueda J, Linacero R,Vazquez A, Karp A (1997) Reproducibility testing of RAPD,AFLP and SSR markers in plants by a network of Europeanlaboratories. Molec Breedg 3: 381±390
Kain JM (1964) Aspects of the biology of Laminaria hyperborea.III. Survival and growth of gametophytes. J mar biol Ass UK44: 415±433
Kendrick GA, Walker DI (1991) Dispersal distances for propagulesof Sargassum spinuligerum (Sargassaceae, Phaeophyta) mea-sured directly by vital staining and venturisuction sampling.Mar Ecol Prog Ser 79: 133±138
Kernodle SP, Cannon RE, Scandalios JG (1993) Concentrationof primer and template qualitatively a�ects products inrandom-ampli®ed polymorphic DNA PCR. BioTech 14:362±364
Lee RKS (1965) Development of marine algal communities on theJuan de Fuca Strait, British Columbia. Ph. D. thesis. Universityof British Columbia, Vancouver
Lewis RJ (1996) Hybridization of brown algae: compatibility andspeciation. In: Chaudhadry BR, Agrawal SB (eds) Cytology,genetics, and molecular biology of algae. SPB Academic bv,Amsterdam, pp 275±289
Lindstrom SC (1993) Inter- and intrapopulation genetic variationin species Porphyra (Rhodophyta: Bangiales) from British Co-lumbia and adjacent waters. J appl Phycol 5: 53±62
Liptack MK, Druehl LD (1999) Molecular evidence for an inter-familial laminarialean cross. (In preparation)
Lu TT, Williams SL (1994) Genetic diversity and genetic structurein the brown alga Halidrys dioica (Fucales: Cystoseiraceae) inSouthern California. Mar Biol 121: 363±371
Mackill DJ, Zhang Z, RedonÄ a ED, Colowit PM (1996) Level ofpolymorphism and genetic mapping of AFLP markers in rice.Genome 39: 969±977
Macpherson JM, Eckstein PE, Scoles GJ, Gajadhar AA (1993)Variability of the random ampli®ed polymorphic DNA assayamong thermal cyclers, and e�ects of primer and DNA con-centration. Molec Cell Probes 7: 293±299
Maughan PJ, Saghai Maroof MA, Buss GR, Huestis GM (1996)Ampli®ed fragment length polymorphism (AFLP) in soybean:species diversity, inheritance, and near-isogenic line analysis.Theor appl Genet 93: 392±401
Mayes C, Saunders GW, Tan IH, Druehl LD (1992) DNA ex-traction methods for kelp (Laminariales) tissue. J Phycol 28:712±716
McDonald BA, McDermott JM (1993) Population genetics ofplant pathogenic fungi. Electrophoretic markers give unprece-dented precision to analyses of genetic structure of populations.BioSci 43: 311±319
Meunier JR, Grimont PAD (1993) Factors a�ecting reproducibilityof random ampli®ed polymorphic DNA ®ngerprinting. ResMicrobiol 144: 373±379
Micheli MR, Bova R, Pascale E, D'Ambrosio E (1994) Repro-ducible DNA ®ngerprinting with the random ampli®ed poly-morphic DNA (RAPD) method. Nucleic Acids Res 22: 1921±1922
408

Miura W, Fujio Y, Suto S (1979) Genetic di�erentiation betweenthe wild and cultured populations of Porphyra yezoensis. To-hoku J agric Res 30: 1250±1256
Mro z LW (1989) Ribosomal DNA assessment of genetic relation-ships among Alaria Greville (Phaeophyta) stands from Van-couver Island and vicinity. MSc. thesis. Simon FraserUniversity, Burnaby, BC, Canada
Mueller UG, Lipari SE, Milgroom MG (1996) Ampli®ed fragmentlength polymorphism (AFLP) ®ngerprinting of symbiotic fungicultured by the fungus-growing ant Cyphomyrmex minutus.Molec Ecol 5: 119±122
Nei M, Li WH (1979) Mathematical model for studying geneticvariation in terms of restriction endonucleases. Proc natn AcadSci USA 76: 5269±5273
Norton TA (1981) The settlement of Sargassum muticum propa-gules in stationary and ¯owing water. J mar biol Ass UK 61:929±940
Norton TA (1992) Dispersal by macroalgae. Br phycol J 27: 293±301
Oppen MJH van, Klerk H, de Graaf M, Stam WT, Olsen JL (1996)Assessing the limit of random ampli®ed polymorphic DNAs(RAPDs) in seaweed biogeography. J Phycol 32: 433±444
Oppen MJH van, Olsen JL, Stam WT (1995) Genetic variationwithin and among North Atlantic and Baltic populations of thebenthic alga Phycodrys rubens (Rhodophyta). Eur J Phycol 30:251±260
Paine RT (1979) Disaster, catastrophe, and local persistence of thesea palm Postelsia palmaeformis. Science, NY 205: 685±687
Paine RT (1988) Habitat suitability and local population persistenceof the sea palm Postelsia palmaeformis. Ecology 69: 1787±1794
Palumbi SR (1994) Genetic divergence, reproductive isolation, andmarine speciation. A Rev Ecol Syst 25: 547±572
Palumbi SR (1996) Nucleic acids. II: the polymerase chain reaction.In: Hillis DM, Moritz C, Mable BK (eds) Molecular system-atics. 2nd edn. Sinauer Associates, Inc. Sunderland, Massa-chusetts, pp 205±247
Pearson EA, Murray SN (1997) Patterns of reproduction, geneticdiversity, and genetic di�erentiation in California populationsof the geniculate coralline alga Lithothrix aspergillum (Rho-dophyta). J Phycol 33: 753±763
Penner GA, Bush A, Wise R, Kim W, Domier L, Kasha K, La-roche A, Scoles G, Molnar SJ, Fedak G (1993) Reproducibilityof random ampli®ed polymorphic DNA (RAPD) analysisamong laboratories. PCR Meth Applics 2: 341±345
Reed DC, Amsler CD, Ebeling AW (1992) Dispersal in kelps:factors a�ecting spore swimming and competency. Ecology 73:1577±1585
Reed DC, Laur DR, Ebeling AW (1988) Variation in algal dis-persal and recruitment: the importance of episodic events. EcolMonogr 58: 321±335
Roa AC, Maya MM, Duque MC, Tohme J, Allem AC, BonierbaleMW (1997) AFLP analysis of relationships among cassava andother Manihot species. Theor appl Genet 95: 741±750
Scheltema RS (1971) Larval dispersal as a means of genetic ex-change between geographically separated populations of shal-low water benthic marine gastropods. Biol Bull mar biol Lab,Woods Hole 140: 284±322
Schierwater B, Ender A (1993) Di�erent DNA polymerases mayamplify di�erent RAPD products. Nucleic Acids Res 21: 4647±4648
Schneider S, Kue�er JM, Roessli D, Exco�er L (1997) Arlequinver. 1.1: a software for population genetic data analysis. Ge-netics and Biometry Laboratory, University of Geneva, Gene-va, Switzerland
Sharma SK, Knox MR, Ellis THN (1996) AFLP analysis of thediversity and phylogeny of Lens and its comparison with RAPDanalysis. Theor appl Genet 93: 751±758
Sosa PA, Garcia-Reina G (1992) Genetic variability and di�eren-tiation of sporophytes and gametophytes in populations ofGelidium arbuscula (Gelidiaceae: Rhodophyta) determined byisozyme electrophoresis. Mar Biol 113: 679±688
Stewart CN Jr, Via LE (1993) A rapid CTAB DNA isolationtechnique useful for RAPD ®ngerprinting and other PCR ap-plications. BioTech 14: 748±750
Sundene O (1962) The implications of transplant and culture ex-periments on the growth and distribution of Alaria esculenta.Nytt Mag Bot 9: 155±174
Upholt WB (1977) Estimation of DNA sequence divergence fromcomparison of restriction endonuclease digests. Nucleic AcidsRes 4: 1257±1265
Vos P, Hogers R, Bleeker M, Reijans M, van de Lee T, Hornes M,Frijters A, Pot J, Peleman J, Kuiper M, Zabeau M (1995)AFLP: a new technique for DNA ®ngerprinting. Nucleic AcidsRes 23: 4407±4414
Waples RS (1987) A multispecies approach to the analysis of gene¯ow in marine shore ®shes. Evolution 41: 385±400
Watts RJ, Johnson MS, Black R (1990) E�ects of recruitment ongenetic patchiness in the urchin Echinometra mathaei in WesternAustralia. Mar Biol 105: 145±151
Widdowson TB (1964) A study of variation in the genus AlariaGreville. Ph. D. thesis. University of British Columbia, Canada
Widdowson TB (1971) A taxonomic revision of the genus AlariaGreville. Syesis 4: 11±49
Williams SL, Di Fiori RE (1996) Genetic diversity and structure inPelvetia fastigiata (Phaeophyta: Fucales): does a small e�ectiveneighborhood size explain ®ne-scale genetic structure?. MarBiol 126: 371±382
Winans GA (1980) Geographic variation in the milk®sh Chanoschanos. I. Biochemical evidence. Evolution 34: 558±574
Win®eld MO, Arnold GM, Cooper F, Le Ray M, White J, Karp A,Edwards K (1998) A study of genetic diversity in Populus nigrasubsp. betulifolia in the Upper Severn area of the UK usingAFLP markers. J molec Ecol 7: 3±10
Zabeau M, Vos P (1993) Selective restriction fragment ampli®ca-tion: a general method for DNA ®ngerprinting. European Pa-tent O�ce, Munich, Germany (Publ No. 0 534 858 A1)
409























![Home - TU Graz...ã æ á á ò s æ æ æ á æ æ í ] v o ] µ v P á é](https://img.pdfslide.us/doc/110x75/611b427e79a8db487368303d/-home-tu-graz-s-v-o-v-p-.jpg)




