Embed Size (px)
Citation preview

Yol. 142:247-259, lS96MARINE ECOLOGY PROGRESS SERIES
Mar Ecol Prog SerPublished October 24
Declines in submersed n.rcrophyte populations havebeen documented at many locations rvorldrvide duringthe past several decades, Frequently, potential causes
are identified by comparing the existing environ-mental conditions of formerly vegetated sites eitherto nearby areas that have remained vegetated or tohistorical records. In this manner, significant losses ofvegetation have often been attributed to excessive
anthropogenic inputs of suspended particulate mate-rial, dissolved nutrients, or both (e.9, clen Harlog &
E-mail: moore@vims,eduPresent address: US Geological Survey, Biological Re"
sources Division, i2201 Sunrise Valley Drive, Nlail Stop300, Reston, Vilginia 21092, USA
O Inter-Research 1996
Resale ol {ull arlicle not permitterl
Zostera marina (eelgrass) growth and survivalalong a gradient of nutrients and turbidity
in the lower Chesapeake Bay
Kenneth A. Moore*, Hilary A. Neckles*', Robert J. Orth
The Virginia Instilute of Marine Science, School of Marine Science, College of William and Mary, Cloucester Point,Virginia 23062, USA
ABSTRACT: Survival of transplanted Zostera narina L. (eelgrass), Z. marina growth. and environ-mental conditions u'ere studied concurrently at a number ol sites in a southrvestern tributary of theChesapeake Bay to elucidate the factors liniting nracrophyte distribulion in this region. Consistentdifferences in survival of lhe transplants \^'ere observed, wilh no long-term sun,ival at any of the sltesthat \^/ere formerly vegetated \dith this species but that currently remain unvegetated. Therefore, thecurrent distribution ol Z. marina likely lepresents lhe extent of suitable environmental conclittons in theregion, and the lack of recovcry into histoncally vtrgetated sites is not solely due to lack of propagules.Poor long-term survival rvas related to seasonally high Ievels of water column light attenuation. Falltransplants died by the end of summer follorving exposuro to levels of high spring turbidity (Ku > 3.0).
Accurnulation of an epiphyte matnx durrng the late spring [0.36 to 1.14 g g rdry rvtl mav a]so havecontributed to this stress. Dtfferences in lvater column nutrient levels among sites during the fall andrvinter {10 to 15 UN4 dissolved inorganic nitrogen and 1 pM dissolved inorganic phosphates) had noobservable effect on epipbyte accumulatior] or macroph)'te growth. Salinity effects $,ere ntinor andthere were no symptoms of disease. Although summertin'le conditions resulted in clepresstons itrgrorvth, they did not alone limrt Iong-term survival, It is suggestecl thdt water quallty conditionserrhancrng adequate seagrass growth during the spring may be key to long-terrn Z. marina survivaland successful recolonization in this region.
KEY IVORDS: Chesapeake Bay . Zostera marina ' Seagrass ' Grorvth Surviva.l Epiphytes Waterqr.rality Inorganic nuttients Turbidity
INTRODT.JCTION Polderman 1975, Phil[ps et al, 1978, Davis & Carey1981, Kemp et al. 1983, Orth & Moore 1983, Giesen etal. 1990, Stevenson et al. 1993)
In order to relate persistent lack of vegetation to un-suitable habitat, environrnental conditions and in situplant grorvth and sulvival must be studied concur-rently. Fol example, Jupp & Spence (19?7) used recip-rocal transplants to determine the importance of waveactlon and sediment nutrient concenirations in limitingmacrophyte recolonization and growth rn a eutrophiclake. Similarly, Cambridge et al. (1986) concludedfrom transplant experiments that the conditions ini-tially causing the loss of seagrasses from an Australiansound still existed in that region Without such infor-mation, poor recruitment because of an insufficientsupply of propagules remains an alternative hypo-thesis to explain persistent lack of vegetation.

248
Zostera maina is the domrnant submersed macrophytein the mesohaline and polyhaline regions of ChesapeakeBay. Historically, extensive seagrass beds covered theshoal areas of less than 2 m depth along ihe bay and theeastern and rvestern shore tributaries. Declines in abun-dance of Z. marina occurred throughout the bay in theearly 1970s (Ort-h & Moore 1983, 1984). Losses weregleatest in the upriver sections of the western tributariesand the deeper, channelrvard areas of macrophyte dis-tribution. N'lany areas of lower Chesapeokc Boy thotonce supported dense seagrass beds currently remainunvegetated.
Here we describe a series of studies designed toelucidate the factors lirniting submersed macrophytedistribution in one southwestern tributary of Chesa-peake Bay, the York River. Zoslera marina populationsdeclined precipitously from the upriver and deeperareas of the York River by 1974, and many areas renraindevoid of vegetation (Batiuk et al. 1992). We usedboth field manipulations and observations to explorethe relationships between macrophyte distribution andenvironmental conditions in the York River: (1) ivetested the hypothesis that environmental quality, ratherthan macrophyte recruitment, restdcts macr:ophytedistribution lo a subset of its former range; (2) weexperimentally evaluated the potential for differencesin macrophl,te gro\^/th at currently and formerly vege-tated sites; and (3) rve quantified differences in rvaterquality betrveen currently and formerly vegetated sitesthat rnay be influencing paiterns of Z. marina abun-dance. Our results demonstrate environmental contiolof plant distribution and suggest those variables con-tribr-rting to persistent lack of vegetation in the regiou.
STUDY SITES
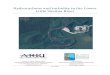
Study sites were established in the York River, Vir-ginia, tlSA, extending from the mouth of the tributaryto the histor:jc upriver limits of macrophyte distribution(Fig. i). We selected sites in areas that had been or arecLrrrently vegetated wilh Zostera maina (Marsh 1970,
1973, Crth 1973, Crth et al. 1979). In this re$.on Z.
marina is most abundant at depths of 80 to 110 cmbelow mean sea level (MSL) and Ruppia maritima L.
(sensu lato) occurs at shallolver depths [Orth & Moore19BB). All stations rvere therefore located at approxi-nately B0 cm below MSL to permit our conclusions to
be related to the majority of potential Z. marina habitatin this region.
The first station in this York River estuadne transect,Y0, (Guinea Marsh; 0 km) is located at the mouth of thetributary and supports Zostera marina beds that havedecreased only moderalely in area since 1937 [Orthet al. 1979). The second station, Y1 1, (Gloucester Point;
Mar Ecol Prog Ser 142; 241-259, '1996
Fig. 1 York River, Virginia, USA, study area shorving studysites and submersed rnacrophyte clistributions in 1970 and 1987
11 km) is located approximately 11 km upriver and isat the upriver limit of the current Z. ntarina distribu-tion. Populations disappeared from this area by 1974,
and have since regrorvn slightly from both transplant-ing and natural recruitment. The last 3 stations, Y12(Mumfort Island; 12 km), Y1B (Catlett Island; 1B km),and Y26 (Claybank; 26 km) lie sticcessively upriver.Ixiensive beds of Z. ntarina disappeared conrpletelyfrom these 3 sites by 1974. All sites are characterizedby shallow flats [<2 rn be]olv MSI-) extendlng land-r.vard from a narro\,v but much deeper (>10 m belowMSL) mid-channel reqion. Sediments in the shoalareas are principally fine sands.
METHODS
Transplant experiments. We used transplant 'gar-
dens' to test the hypolhesis that enrrironmental condi-tions ultimately limit distribution of Zostera marjna in
f\IN
I.,:.(
_9r,
":,1
.J
.)
-)
L-.lri"
a
,' \
,l
2 ;!
.\:
'/,
nrtO. t.: .-h\t"-
' )t:'
l
L
Yto;+
i::::. ,,r,j z' marina - 1970
l r.marina - 1987
? . ,, ,, ? ---. ,19Kilometers

the York River, We transplanted Z, marina to currentlyand fonnerly vegetated sites to determjne the presentcapacity oI various sites to support lnacrophyte grolvth.Pre',,ious transplanting efforts in this reqion have de-termined that fall is the best season to ensure trans-plant sr,rccess (Fonseca et al. 1985, K. Moore & R. Onthunpubl. deita), therefore transplanting rvas undertakenin Septernber and October of 1984, 1985, and 1986
Plants r,vere collected from the established bed at Y0,
tlansferred [o transplant sites, and responses mea-sured; the designs of the transplant experiments aresuinmarized in Table i, In 1984, planting units con-srsted of sods (20 cm x 20 cm) rvith intact sediments.During subsequent years tlie shoots were washed freeof sediments, and plantrng units consisted of 10 to i5shoots bunclled together rvith a metal trvist tie similarto methods of Fonseca et al. (1982, 1985) for ease oftransplanting. No apparent differences have been ob-served in the survival rate of transplants in this regionusinq these 2 methods (Fonseca et al. 1985, K. Moore &
R. Onth unpubl. data). All vegetation rn'as transplantedwithin 24 h of removal frorn the donor site. Piantingunits were spaced at 2 m or 0.5 m centers (Table 1)in3 to 4 replicate 5 x 5 arrays of 25 planting units at eachsite, Survivorship rvas monitored each year (Table 2) atmonthly to bimonthly intervals until either no plantsremained at a site or the planting units had coalesced.Sr-rrvivorship was calctilated as the percent of originalplanting units remaining tn individual replicate arrays.
During 1984 and i985, 4 similar arrays of plantingunits rvere established adjacent to the survivorshipplots at each transplant site to provide material fordestructive sampling. The additional macrophyte re-sponses measured are suntmarized in Table 1. Plantstransplanted rn 1984 were sampled in November 1984
and January, March, May, and .luly 1985. On eachsample ciate, 3 to 5 core samples of 0.33 m2 were takenfrom the natural seaqrass bed at Y0 and 5 arbitrarilyselected planting units rvere excavated from the de-structive sampling arrays at each transplant site formacrophyte biomass determination. The plants werewashed gently in the field to r€move sediment and
r"loore et d.: Zoslera tldl'ina gro\vth ancl sut'r,tval )AQ
transported immediately to the laboratory. Leaveswere separated from roots and rhizomes and all plantrrraterial rvas dried at 55oC. Five separate samples con-sisting of 5 large terminal shoots each were collected ateach site for epiphyte samplirrg to quantily drfferencesin epiphyte Ioads betrveen presently and f ormerly\regetated sites that may be affecting macrophyte sur-rrival. Shoots, rvhich consisted of all leaf material abovethe meristematic region (Sand-Jensen 1975), weleseparated from the remainder oi the plant and sruitledseveral times in a beaker of filtered seawater toremove loosely adhering rnaterial. The leaves in eachsample were separated into leaf age classes, and theepiphytic material was scraped into filtered seawaterrvith the edge of a glass n-ricroscope slide. Mobileepifauna were discarded. Epiphytic material was col-lected on pre-combusted glass fiber filters (Gelman,Type A/E), dried at 55"C, and cornbusted at 500'C for5 h. The area of leaf substrate for each sample wasdetermined using a Li-Cor Model 31 area meter andleal dry u'eight and ash-free weight were determined.
Plants transplanted in 1985 were sampled in March,May, June, and July 1986. At each site, 5 to 7 plantingunits n'ere arbitrarily collected, from whrch 5 sub-samples containing 5 large terminal shoots each wereformed. Epiphlzti6 mass was determined as describedpreviously. The areas ol leaves !vere measurecl and dly."veight and ash-free weight rvere determined. The bio-mass of remaining leaves was then calculated from thelinear regression of leaf weight on leaf area. Below-ground biomass was determined irom 3 of the samples.The rhizomes were separated into individual intern-odes for dry weight and ash-free weight measure-ments. The roots from all internodes in a sample werecombined for analyses.
Growth experiments. Although the transplant ex-periments yielded information on patterns of macro-phyte survival and biomass allocation, ambient turbid-it.y prevented us frorn measunng actual rnacrophytegrowth in sittt. Therefore, to evaluate the effect ofrvater. quality on nacrophyte growth at currently andformerly vegetated sites, we reloc.rted turfs of Zostera
Table 1. Design of transplant experiments
Time of N4ethodtransplantrnq (spacing)
Tra nsplantsr tes
Y11, Y26
Y0, Y11, V12,Y18, Y26
Y11,Y12, Y1B, Y26
Response measured
Transplant sulvrvorshipEntire sods collected Ior macrophyte biomass'Individual shoots collected for epiphl,ti6 rnaterial'Transplant survivorshipIndividual shoots collected for macrophyte biomass and epiphlrtic malerial
Transplant sun,ivorship
Fall 1984
Fall 1985
Fall 1986
Sods (2 m)
Bundles (0.5 m)
Bundles (0.5 nr)
uBecause no plants were transplanted to Y0, samples were taken from natulal Zostera marjna bed

250 Mar Ecol Prog Ser 142:247^259,1996
Table2. Zosterantarina.Percentsurvivalattransplantsiles.Valuesareback-transformedmeansof arcsine.squarerootlransformeddata. Unilke letters denote signrfrcant differences (p < 0.05) anonq srtes on each sample date. bd: transplanted plantrng unitscoalesced lvith one another or new recruits be1,6n6 deternrinatjon. E: water column turbtdily preclrrded sun ivorship deternrination
Transplantperiod
Site Sample dateMay 1985Nov 1984 Mar 1985 JLrl 1985 Aug l9B5 Oct l9B5
Fall 1984 Y11Y26
100 "91 b
8236
82"gl,
64'0b
64' bd0b 0
Aug 1986 Oct 1986Oct 1985 Mar 1986 Mav 1986 Jun 1986 Jul 19BG
Fall 1985 YO
Yl1v12YlBv26
100'100 d
100 u
100 a
100 u
100 "60 I'
64b62b60b
bd60,64'56060'
bd60064044h34'
bd60012"Bb0'
bd60
000
bdbd000
b
l)
b
Oct 1986 Apr 1987 May 1987 Jul 1987 Aug 1987 Oct 1987
Fall 1986 Y11v1.2Y1BY26
i00 "100 d
100.100 u
80d87b91tl'95'
80087b91 r'.'
95'
47
EEE
47u6u0b0b
bd00
0
mailna from the stable grassbed at Y0 to sites Y11 andY26. We measured in situ macrophyte grolvth fromApril 1985 to July 19B6 using a modified leaf markingtechnique (Sand-Jensen 1975). Whole turfs of Z.
marina, including roots, rhizomes, and undisturbedsediments to a depth of 20 cm, were obtairled fromthe grass bed at Y0, placed in polyethylene boxes (40 x
60 x 20 cm), and 1 box placed at Y11 and 1 at Y26.After a 2 wk acclimation period, three 15 cm diameterrings were arbitrarily located within each box, Eachshoot within each circular quadrat was tagged with a
numbered, monel metal band placed around its base.
The younqest leaf was marked with a srnall notch andthe ieaf lengths and widths were recorded. At approx-imately r,veekly intervals the boxes were retrieved,placed in a seawater bath, and the length and width ofall leaves on tagged shoots recorded. The number ofnew leaves on each shoot rvas recorded, any newshoots within the quadrats were tagged, and theyoungest leaf on all shoots was marked. Thus, indi-vidual Ieaves could be uniquel)' identified and mea'sured from formation through loss. Leaf grorvth wasdetermined as changes in leaf length. Dry weight andash-free \^/eights at each sampling period rvere derivedusing leaf weight to area relationships determinedfrorr the experimental transplants for each penod.Specif ic rates of biomass change were calculatedfor each marking inter:val as leaf production or loss
divided by initial biomass. Boxes at the srtes were dis-turbed periodically, generally through the burrorvingaclion of crabs or fish. Therefore, when excavationoccurred in a box at either site, boxes at both sites werereplaced with others that had been acclimating at the
respective sites for identical periods of time, generallyranging from 3 to 4 wk. Plants in boxes were not usedfor survivorship measurements.
Using growth information derived from the markedplants, rhizome production rates of the plants trans-planted to Y11 and Y26 in the fall of 1985 were esti-mated. It was assurned that on average, the individualrhizome internodes lvere formed at the same rate as
leaves (Sand-Jensen 1975, Jacobs 1979, Aioi et al.1981). Using the calculated leaf formation rates, theages of individual internodes were thus determined foreach of the transplant samples obtained in March,May, June, and July 1986. Rhizome productrorl wasthen calculated by summing the biomass of rhizomeinternodes (including roots) produced betrveen sampledates
Environmental monitoring. \,\rorldwide declines ot'
submersed macrophyte populations have been vari-ously attributed to increases in water column turbidityand to increases in dissolved nutrient concentrationsand consequent epiphyle accumulaiion Therefore, todetermine rvhether rvater quality differences may beinlluencing patterns of Zostera marina abun.da.nce inthe York River, rve monitored water quality at the trans-plant sites frorn January 1985 through December 1987.
We collected triplicate subsurface rvater samples ap-proxrmately every 14 d at each of the sites. All sampleswere obtained sequentially on the same day over a 2to 4 h period beginning with the mosl downrjver site;samples vrere stored in the dark on ice for up to 4 hwhile being transporled to the laboratory and wereanalyzed immediately on arrival. Nitrite, nitrate, andammonium were determined spectrophot.ometrically

follorving the methods of Parsons et al.(1984) and inorganic phosphorus fol-lowing the methods of USEPA (1979).
Suspended matter wds collected onprecombusted, Celman Type A/E glossfiber filters, dried to constant !veight at55'C and combusted at 500"C for 5 h.(lhlorophyll a (chl a) was collected onWhatn-ran GF/F glass fiber filters, ex-tracted in a solvent mixture of acetone,dimethyl sulfoxide and 1% diethyl-amine (45 :45: 10 by volume) and deter-mined fluorometricaily (Shoaf & Lium1 976). Chlorophyll concentrations wereuncorrected for phaeoplgments. Salin-ity lvas measured with a refractometer.
We measured diffuse downwellingphotos),nthetically active radiation(PAR) from triplicate, water colunrnprofiles of photosynthetic photon fluxclensity (PPFD) using an underwater 2r,cosine-corrected sensor (LI-COR, Inc,,LI-192SA). These data were obtained concurrentlywith the rvater sarnples. Measurements of PPFD oneach sample date were summarized as the attenuationof dor.snwelling PAR. The downrvelling attenuation co-efficient (K,r) was calcuiated according to Beer's Law.
Statistical analysis. Macrophyte and epiphyte re-sponse variables (rnd envrronmental measurementswere analyzed using 2-lvay analysis of variance withmain effects of site and date (SPSSx subprogramMANOVA, SPSS, Inc. 1986), Experimental units werereplicate arrays for survivorship measurements, sam-ples for macrophyt.e and epiphyte biomass measure-ments, quadrats for growth measurements, and watersamples or light profiles for environmental measure-ments. Residual analysis was used to check modelassumptions and log transformations were appliedq,here necessary (Neter & Wasserman 1974). N4eanswere compared among sltes rvithin sample dates usingTukey or Bonferroni Multrple comparisons with a
family confidence coefficient of 0.95.
RESULTS
Transplant experiments
Survival of Zostera marina transplants differed con-sistently between sites upriver and dorvnriver of Y11dttring all 3 yr of transplanting (Table 2). At Y11andY0, after some initial losses during the winter, thetransplants became well established and persistent. AtY26, loss of tlansplants occurred during the spring andearly summer, so that by August no veg€tation re-
Moore et al.. Zostera marina growth and survival 251
Table 3. Zostera ntarina. Shoot biomass, 1984 to 1985. Biomass Values are back-transiormed from nreans of log tiansformed data. Unlike letters denote siq-nrficant differences (p < 0.05) among means on each sample date. S/R: shoot to
rool-rhizome ra[io. nsi no survival at Y26 by Ju] 1985
mained. At Y12 and Y18, although the plants survivedfor a longer peliod through the sr-rmmer than Y26 theyalso clied out completely by the encl of August.
Initially no significant differences in shoot biomassmeasurements of 1984 transplants were observedamong srtes (Table 3). By January, however, Y26shoots had lor,ver below qround biornass, resulting tn a
significantly higher shoot to root/rhizome (S/R) ratio. InMarch, S/R ratios of ,plants at Y26 remained hrgherthan of those at Y 1 i . By May, increases in growth wereevidenI at all sites. The greatest leaf biomass occurredat Y0. No biomass differences occurred between Y11and Y26. By July, no living plants remained at Y26,although dead, blackened rhizomes provided evi-dence of recent, viable plants.
Sampling of the 1985 transpiants revealed a similarpattern of S/R ratios along the river axis (Table 4). InMarch 1986, only the S/R ratios at Y26 were signifi-cantly higher than at Y0; by June, the S/R ratio in-creased with distance upriver. By.luly all plants at Y26were gone.
Various measu.res of epiphytic density (dry or ash-free mass of epiphytes per unit area or mass of leaftissue) yielded similal patterns among sites, and re-sponses to sites were similar arTlong leaf age classes.Therefore, results are expressed only as dry weightratios calculated on a whole shoot basis (Table 5), Theepiphytlc material included diatoms such as Nitzschiasp. and Ltcmophora sp- as well as heterotrophic flagel-lates and bacteria, ar-rd attached debris (Neckles et al,1994). Macroalgae (e,9. Enteromorpha sp.) formed a
smail proportion (<5%) of the total mass and wereexcluded from analysis. The highest epiphyte mass
Date Site n Shoot(mg dry mass sh-r;
Root-rhizom.e S/R(mg dry nass sh-r)
Nov 1984
Jan 1985
Mar 1985
May 1985
Jul 1985
YO
Yi1y26
YOY11Y26
YO
Yl1Y26YO
Y11Y26
YOY11Y26
555
555
3
55
355
3
5
ns
38,BO O
26.14 "38.13 d
15.90 d
r2.7 5 a
14.58 "
18.79 " b
1331"27 .40 b
105.03 "39.08 "53.20 a
119.65 "42.48 o
ns
40.39 "39.78'.
50.35 d
68.80 .15.93 b
28.90 d
41 18.24.85 a
61.66 "4530"35.36 "
75.49 "36.58',
ns
1.37 "0.6s "0.95 a
0.32 "0.19 a
0.92 b
0.65 " b
0.32 "1.10 b
1.70 a
0.86 "i.50 i1.58 a
7.74 a
ns

252 Mar Ecol Prog Ser 142.247 259, 1996
Table 4. Zostera ntarina. Shoot bromass, 1985 to 1986. Bionrass values are back-transformecl from means ol Iog transfornl(:d datd. Unlike letters denot.e siqtrifi-cant differenct:s (p < 0.05) among means on crrch sample date. S,jR: shoot to
root-rhizonre ratio. ns: no survival at Y26 by July 1986
occurred on the Y11 transplants rn November i984.Each year, densities were signilicantly higher at Y26than at the other 2 sites immediately before the Y26transplants disappeared.
Although no forn'ral rneasures of the incic.lence of dis-ease were taken, the plants were observed throughoutthe study for evidence of infection such as might becaused by Labyrinthula sp. associated with the eel-glass wasting disease fMuehlstein et al. 19BB), Typi-cally, the older leaves on the plants had occasionaldark patches of damaged tissue rvhtch covered nomore than 5%" of. Lhe leaf tissue as recently describedby Burdick et al. (1993). There was no evidence ofnecrosis on the younger leaves holvever, and no evi-
dence of the charactetistic infection ofyounger leaves from adjacent olderlearres as has been docurnented (cf.
Short et al. 1988, Burdick et al. 1993).As the production of nerv leavesslorvecl during the summer, especiallyat sites upriver of Yl1, older leaveswere gradually lost and the nr:mbersof leaves per sh.oot decreased. Eventu-ally, rnanv shoots were composed ofonly several small leaves that hadceased eLongating, lvltb no evidenceof infected spots or patches.
Growth experiments
At both YL l and Y26 highest growthrates occurred each spring and a
second period of increased growth oc-curred in ihe fall (Fig. 24). Leaf grorvthwas lolv during the summer and winter(Fiq. 2A). Siqnificant differences be-tween the sites were observed onlyduring the spring ancl fall periods of
rapid growth. Tl:re rate of leaf formation (Fig. 2C) rvassignificantly greater at Y11 than at Y26 during earlySeptember 1985 and during Aplil and May 1986. Ratesof leaf loss were highest at both sites during late sum-rner (Fig. 2D). Hortever, leaf loss increased earlier inthe season at Y26 than at Y11 (Fig.2D), resulting in a
significantly greater rate upriver, from April throughJuly 1986. The rate of leaf growth rvas greater at Y11throughout the spring and fall periods (Fig. 2A). Differ-ences in leaf replacement and growth resulted in con-siderable seasonal difierences in shoot size betweensites. For example, the mean shoot biomass at Y11 inMay 1986 rvas 45 mg compared to 11 mg at Y26. Sin-rilarsite differences of lesser magnrtude occurred in the fall
Tabte 5. Zostera nlarina. Epiphvt.ic density tg g ' dry ma.ss '; for 19B4 anci 1985. Datd are back-transfornred from means of logtransformed data. Unhke letters denote significant differenc,.rs (p < 0 0.5) among srtes on each sample date. ns: no survival at Y26
by July 1985 and July 1986
Date Site n Shoot(niq dry mass sh-r)
Root-rhrzonrr,:(nrg dry mass sh 11
S/R
Mar 1986
ir'lay 1986
Jun 1986
Jul 1986
YOY11Y12Y1BY26
YOY11Y12Y1Bv26YOY11Y72Y18v26YOYllY12Y18Y26
5
55
5
55555
555
5
55
5
NS
22.03'29.85 "25.76 ^
40.50 h
47.42n
66.37 ',7 r.28 h
90.99 u,b
151 I "t t'
r04.47 t'
68.08'117.49 h
1 02.56 ', b
168.65 "136.14 t'
69.34 ..r'
210.38'95.50 I'
66.07 ',
NS
30.41 ''
33.81 ',
26.00 "
30.97 "
85 70'.91 62"'
182.39 I185.35 !'
95.06 :'
r31.52 " t'
195.BB I'
104 95 ',:''!72.58 b
63.09'95.50 "
234.42 h
106.91 " "6823'
NS
o.l')'088',b0.99 "
r'
1.10 "i'.1.54 h
o 77 "b0.79 " I'
0.50 n
0.82'.i',1.10 "0.52'0.60'b0.98 b
0.98 b
Lt6'
072^0.90 "0.89'0.97 "
NS
Transplant Siteperiod Nov'l 984 Jan 1985
Sample daleMar 1985 N4ay 1985 Jul 19U5 Oct 1985
Fall 1984 YO
YliY26
7.17 '0.74 "0.06 b
0.06 "o. 1g I'
1 14'-
0.29 "0.54 !
NS
\. iar 1986 \1:r.,' 1986 Jrin 1986 Jul 1986
007.o.28 t'
034 "
Fall 1985 YOY11Y26
0.67 "7.03 h
T.B2 "
0.85 "0s8"1.06 '
007,7 .021'027"
0.13 0
008"0.36 b
0.'ia "0.65 '
ns
1.15 "1.'t 1 ',

T!r\o-l(d
I
oocaEoE
0.3
0.2
0.1
0.0
0.8
0.6
0.4
o.2
0.0
4.0
3.0
2.0
1.0
0.0
4.O
3.0
2.0
1.0
0.0
Production
Loss
Production
lvloore et aI.. Zostera mar-ina glrou.th and sun,ival
A,
253
Below-ground rhizorne production (Table 6) r,vas
similar at Y11 and Y26 from November to March, dur-ing lvhich time rates at both sites rvere qr-rite low. FromN4arch until l.he die-off ol vegetation at Y26 in July,rates were significantly greater at Y11. lvlaximunr pro-dlrction occurred at both sites between Vlalch andMay.
Environmental monitoring
Environmental variables were compared among siteswithin each sampling date. The spatial and temporaldistribution of water quality parameters were consis-tent {rom year to year, so data are presented graphi-cally as monthly means fronr 1985 to 1987. For clarity,only data from Y0, Y11, and Y26 are included. Levelsof environmental parameters at Y12 and Yl-B weregenerally intermediate between Y11 and Y26.
Water ternperatures were similar at all sites withannual minima approachinq 0"C in late January andmaxima near 30'C in August (Fig. 3,l\). Salinity de-creased approxirnately 5%' from Y0 to Y26 (Fig. 38).Minima and maxima were during January and August,respectively, and paralleled river infiow into the baysystem.
Concentrations of total suspended solids {TSS) rverevariable among sites but r-rsually increased with dis-tance upriver (Fig.3C). Consistently, each spring[April to June) concentrations at Y26 were significant]ygreater than at downriver sites. The suspended loadconsisted principally of inorganic particles; organiccontent of the seston was usually less than 30%. Thispercentage decreased with distance upriver.
Patterns of increasing light attenuation 1K,,) withdistance upriver paralleled those observed for thesuspended particles (Fig. 3D). Step-wise, multipleregression of K3 on the principal measured compo-nents of attenuation Ifilterable inorganic matter (FIM),filterable organic matter (FOM), and chl a| revealed
C
I
(!-Tooaoq)
(gc)
" Y11. Y26
T*, 7'\.***" I
n-/n! \-.&|ar(]
Loss
Jun Aug Oct Dec Feb Apr Jun Aug
Dare (198s-1986)
Fig.2. Zostera marina. Results of gro\,vth experiments. Meanrates of (A) shoot production, {B) shoot loss, (C) leaf produc-tion, and {D) leaf loss for Z. marina turls contained jn boxes
at sites Y1 1 and Y26. Bars are 1 SE
as r,vell, This difference in shoot size contributed to a
greater rate of total biomass loss at Yl1 cluring thespring and fall (Frg. 28), although the mean daily netchange in biomass remained higher at this site dur-ing these periods. Mass specific rates of leaf biomassaccumulation and loss at each site followed the samegeneral patterns as did shoot-specific leaf growth.
D(0.s)
Apr
Table 6. Zostera marina. Belowground productron for 1985 to 198tj. Produclion data are back-transformecl from means of logtransformed data. Unlike letters denole significant differences (p < 0.05) betrveen sites during each period. na: data not available
due to complete mortality at Y26 by 21 July 1986
Site Period Days n lr4ean no. ofsegments formed
Production{mg drv mass sh t d-r1
15 Nov 1985 to 1B Mar 19864 Nov 1985 to 9 Mar 1986
24 Mar 1986 lo 9 May l98620 Mar 1986 to 13 May 1986
8 May 1986 to 9 Jun 1986B May 1986 to 10 Jun 1986
10 Jun 1986 to 21 Jul 198610 Jun 1986 to 21 Jul I 986
).:) 4
5
I3
32
3
na
0.06,0.07 n
205',0.63 b
1.18 u
0.26 b
0.65na
25
z525
Y11Y26
Y11y26
Y11v26Y11Y26
124724
4747
3334
4242
25
25na

254
Eov
30)4201510
0
Mar Ecol Prog Ser 1,42:247 -259, 1996
Jan Mar May Jul Sep Nov
Highest levels of dissolved inorganic nitrogen (DIN)occurred during the fall and winter periods (Septenr-ber to February; Fig 4A). At this time, DIN speciesconsisted principally of ammonium aLthough nitlitecomprised approximately 50% of DIN by December,especially at Y26. Concentrations of DIN were signifi-cantly higher at Y26 than at the downriver sites duringthe fall and winter. During the summer (June toAugust; Fig AA) ammonium accounted for greaterthan B0ol, of DiN and therp wpre generaily no differ-ences in DIN levels among the stations. Nitrateaccounted for approximately 5 lo 15o/" of DIN at all sta-tions throughout the year.
Dissolved inorganic phosphate IDIP) ]evels showedIittle annual vanability (Fig. aB). Increasing levels withdistance upriver were observed during much of theyear. The highest DIP levels occurred at Y26 duringthe f all with intermediate levels at Y 1 1 .
N:P ratj.os for dissolved inorganic nutrients {Fig 4C)generally followed the patterns for DIN availability.Ratios usually exceeded 1.5 from October throughJanuary and were less than 15 from February throughSeptember. A marked increase in N:P was observed in
30z3201510
50
A,
A
Oo
lao-
B)c\
20
10
5
zll' I
lx::e,
i56):^ 50
Psoc-20310FO
c.
D5
4
32
I
0
.Y0v Y11
. Y26
Fig. 3. N,lean monthly [A) temperature, (B) salinity, [C] totalsuspended solids (TSS) and (D) light attenuatron (K6) at Y0,Y1. 1, ancl Y26 for the period of January 1985 to December
1987. Bars are 1 SE
significant effects of FIM and chl a on K,1, but no effectof FOM (Table 7). Therefore a regression equation us-ing FIM and chl a as independent variables explained46'/. of the variation in .(1. There were no consistentdifferences in chl a levels between the 2 upriver sites(Y11and Y26;Fig.4D). However, chl a concentrationswere significantly lower at Y0 than at all upriver sitesduring the early spring bloom (Fig. 4D). This seasonal,marked increase in chl a durinq Febluary and Marchhad little apparent effect on total, water coh-rmn Iightattenuation during that period (Fig 3D)
Tahle 7. Steprvrse multiple linear regressicrn of water qualitvvariables on light attenuatron (Kri. FIM: filterable inorqanicmatter Chl a: chlorophyll a. FOlr.'I; filterable organic matter
b: estimate of regression coef ficient p
z-
2.0
I r.s
-l r.oLo 05
0.0
I
B
\I\r
40
30
2Q
10
0
.Y0v Y11
co-ozo
0
5
-:--
(t)
=-
JI(_)
15 D
/-
a
Lz1-
0
I r-.Jan Mar May Jul Sep Nov
Fig. 4. Mean monthiy tA) dissolved lnorgantc nttrogrln (DlN1,(B) dissolved inorganic phosphorus (DIP), {Cl IJIN:lJlP ratios,[Dj chlorophyll a at Y0, Yi1, and Y26 for the period of
January 1985 to December 1987. Bars are 1 SE
I' b SEb p
FIMChl a
FOMConstant
0 0400 0i40.0130 636
0390460.4 6
0.0050 0040.0330 078
0.0000 0010.6900 000

April and May at Y0. This rvas principally due to aninterval of elevated nitrate (ranging from 5 to B trM)thaL was observed in 1986 at this site, with no concomi-tant change in DIP,
DISCUSSION
Distribution ol Zostera marina: propagule supply orhabitat suitability?
Distinct differences in the survival of transplantsalonq the York River inclicate there are differencesamong sites that are limiting re-colonization. Piants didnot survive dt any of the historically vegetated sitesupriver of Y11, Therefore, the lack of macrophyte re-growth into formerly vegetated areas of this estuaryhas not been dr.re simply to a lack of plopagule recruit-ment. The distribution of Zostera tnarina in the lowerChesapeake Bay at this time likely represented theextent of suitable environmental conditions in theregion. Current surveys (Orth et al. 1993) of submersedmacrophyte distribution in the York region show a con-tinued lack of plants upriver of Y11.
Transplant failure in these experiments \,vas notattributable srmply to the absence of existing vegeta-tion which might modify the local environment andprovide improved conditions for growth (Orth 1977,
Fonseca et al. 1982, Kenworthy et al. 1982). At Y11, forexample, lvhere transplants were successfully estab-lished, the littoral was largely unvegetated beforetransplanting. Differences in environmental conditionsamorlg study sites rvith varying degrees of transplantsuccess should, therefore, be related to causes of thereduced level of rnacrophyte populations found inlower Chesapeake Bay.
Transplant mortality along the river axis in the falland winter immediately follorving planting rvas similaramong sites and appeared related to physical distur-bance. Shoot biomass rvas low at all sites duringthis winter periocl and all plants lookecl healthy andvigorous. At many locations where pianting units werenrissing, rvire anchors were found proiruding out of thesediment and there was no evidence of below-groundor other naterial remaining. Il. thus appearecl thatoverwinter transplant loss r,vas mainly due to scouringactivity of storms which occurred before the plantingunits rvere additionally anchored by new root/rhizomegrowth. The lower initial loss of planting units atY0 may have been reldted to the attenuation of waveenergies by adjacent vegetation (Ward et al. 1984),
Transplant mor.tality during the summer, 1n contrast,appeared related to enviromental conditions. Althougha variety of organisms can result in great destruction toseagrass beds (Orth 1975), we found little evidence ofdisruption of the transplants by burrowing dctivities
i\'loore et aL. Zostera ntarina grorvth and survival 255
of crustaceans or fish duling the growing season. Attransplant sites upriver of Y 1 1 rvhere all the transplantseventnally died, dead rhizornes could usually be foundin the sediment at the locations of the individual plant-ing units, This confirn'red that the plants died jn sjlu,and were not sirnply uprooted or physically removed.Also, a decrease in the size and shoot abundance of theindividual planting units preceded their cornplete loss.
Results of growth experiments at Y11 and Y26 sug-gest seasonal differences in rvater quality betweenupriver and downriver sites that may have influencedtransplant success. The similarity in growth betrveensites during the rvinter provldes further evidence thattransplant loss during this period r'r'as unrelated towater quality. In contrast, differences in growth in thespring inclicate that differences in environmental suit-ability occurred during that period.
Patterns oi plant response
Patterns of Zostera marina growth and biomass allo-cation along the York River suggest potential mecha-nisms of plant response to environmental conditions.The greatest d.ifferences in plant grorvth betrveenupt'iver and downriver study sites occurred duringApril and May, when growth rates were at their arlnualntaxima; no differences were evident during the sum-mer months of June ancl July when glowth rates \^/erelorv at both sites (Fig. 2A). Mortality of experimentaltransplants at Y26 occurred throughout the spling andsummer, so that no plants remained by Ar-rgust eachyear. Transplant mortality may be attributable to inade-quate production and ensuing carbohydrate storageduring the spring. There is evidence that seasonalaccumulation of carbohydrates in seagrass rhizomesduring favorable growth periods can provide a sourceof energy for structural and respiratory reqr.rirementsduring periods of unfavorable, glowth-limiting con-ditions sucil as high temperature or low liqht (Dawes& Lawrence 1979, Titus & Adams 1979, Ott 1980,Wittman & Ott 1982, Bulthuis 1983, Drerv 1983, Pirci.985, Dau'es et ai. 1987). in the present study, trans-plants were characterized by increasing S/R biomassratios (Tables 3 & 4) and reduced beiow-ground pro-duction (Table 6) with distance upriver, suggestingthat carbohydrate storage of upriver plants may havebeen insufficient to meet metabolic den-rands duringthe summer. Chesapeake Bav is near the southern limitof Z, marina distribution, where high water tempera-tures result in high respiratory demands during sum-mer months (Evans et al. 1986J. The storage andsubsequent mobilization of photosynthate may be animportant mechanism for summertime survival of Z.marina in this region (Burke et al. 1996).

256 N{ar Ecol Prog Ser 142;247 -259, 1996
Influence of environmental conditions
Salinity stress
Although Zostera sp. can tolerafe a rvide range ofsalinities, photosvnthests and respiration are inhibitedin waters u,here salinities are either hypo- or hyper-tonic (Ogata & Malsui 1965, Bieble & McRoy 1971, Kerr& Strother 1985). Although all sites used in this studyhad lristorically supported Zostera marina beds prior todie back in the 1970s, salinities do decrease rvith dis-tarrue uprr iver, suggesLilll; d pussilJle eII€ct contllbutlngto the decreased growth and sr"rrvival observed here.Evidence suggests, however, tllat the salinity effect wasminor. Salinity decreased on average approximately4 to 5%" between Y 1 1 and Y26. Using a linear relation-ship between shoot production and salinity determinedby Pinnerup (1980) for Z. marina transplants in Danishwaters during the summer, we est.intate an approximate1.07o decrease in shool production due to lower salini-ties betrveen sites Y11 ancl Y26- This compares to theapproximately 85"/" clifference in shoot productionmeasnred betrveen Y11 and Y26 cluring May and Junein the growth experiments.
Disease
Evidence has lecl rnvestigators to suggest that envi-ronmental stress may result in a weakened eelgrasshost lhat ',vould allor,r' a pathogen such as the marineslinre molcl Labyrtnthula sp. to decinrate the popula-trorrs (Rasmussen 1977, Short et al. 1988, Burdick etal. 1993) Although this is a possrble explanation forresults documented in this study, there was no evi-dence oI widespread disease symptoms in the trans-plants here. The pattern of die-off in this study alsosuggests a.n alternative explanation. Die-off here oc-curred in [he upriver stations where salinities weregenerally belotv 22%o (Fig. B). In senera]., taby-rinthula sp. tend.s to be most infective at salinitieshigher than these (Burdick et al. 1993).
Water colutrn liqht attenuation
The precipitous dlop in shoot growth in April at Y26when plant growth rates were at their annual maxima(Fig. 2A) coincided with a period of high suspendedload and reduced light (Fig 3C, D). Durincl lvlay toJune at sites Y0 artd Y11 PAR at transplant clepth wasapproximatclr'25 to 50'li, of sub-surface irradiance U0)as determined from K6 measured during that period.However for the May to June period at Y26, PARat transplant depth was only 12'r;. of ,16_ This wouldonly be marginally sufficient for growth (Duarte 1991,
Dennison et al. 19931 even given no other stressorssuch as epiphytes. Thus, lor.v light availability rvasprobably a dominant factor causing the low growthand ultimate mortality of plants at Y26. Similar rela-tions have been observed previously, r,vhere reductionsin total daily light availabiliiy in June resulted in com-plete loss of Zostera marina plants by the end of sum-mer (Dennison & Alberte i9B5). Zimmern)an et al.(i991) have suggested that extended periods of highturbidity in spring may be responsible for the limiteddepth distribution of Z. mafina in San Francisco Bay.
Dissolved nutrient concentralions
Declines of submersed macrophytes in some systemshas been attributed in part to nutrient enrichnent andconsequent increases in epiphytic accumulation thatlimits light and carbon available for leaf photosynthe-sis (e.9. Phillips et al. 1978, Trvilley et al. i985, Silber-stein et al. 1986, Hough et al. 1989). During fall periodsrvhen elevated nutrient concentrations were measuredin the formerly vegetated, uprivel sections of the YorkRiver, however, concomitantly higher epiphytic bio-mass was not observed. Thus, in this study factorsother than nutrient suppJy, such as invertebrate graz-ing activity (Howard 1982. van Montfrans et al. 1982,Cattaneo 1983, Borum 1987, Neckles et al. 1993) ortemperature (Penhale 1977, Borum & Wium-Andersen1.980, Libes 1986), limited epiphyte growrh during rhefall. Periodically bigher epiphyte loads at downriverstations (Y0 and Y11) than upriver (Y26) cluring the falland winter (Table 5) did not appear to affect transplantsurvival- Since light at the macrophyte leaf surface is afunction of both water column and epiphytic attenua-tion, lower water column turbidities (Fig. 3) at thesedorvnnver stations during this period may have miti-galed the effects of higher epiphyte loads
In the late spring (May to June) epipliytic bromasswas significantly higher at Y26 than at other sitesi thiswas immediately before the transplants disappeared.Atom.ic ratios of dissolved inorganic N:P (<10:1) ir-rdi-cated that alqal growth was likely limited bv nitrogenrather than phosphorus at this time. March to Aprilconcentrations of DIN were similar among sites upriverof Y0 (Fig. 4A), although DIN concentrations were ob-served to be significantly higher at Y26 thdn downr.iversites in May. DIP concenlrations remained consistenLl)'higher at Y26 than downriver sites throughout the year(Fig B) Although epiphytic grorvth may have beendependent upon rapid recycling of N rather thanabsolute coltcentrations, other factors may have alsocontributed to increased epiphytic densities uprlver atY26 in late spring. In turbid esluaries, consid.er:ableamounts of inorganic and organic debris may be en-

trapped by the epiphyie natrix (Kernp et al. 1983).Higher concentrations of this fouling material at Y26rnay thus reflect high springtinte concentlations of sr.rs-
pended particles at [hat site In adclitiorr, lr4urray (1983)Iound the rela[irre photosynthetrc efficiencies ol epi-Ph)rtic algae and Zostera manna to resr.rlt in increasingepiphyls;m66rophyte ratios rn,ith decreasirrg light in-tensily. Differences in the mass of this epiphytic mate-rial along the York River axis in the spring ntay thusreflect respoltses to light availabrlity. Small increasesin accumulation of this materjal rnay limit n-racrophytesurvival at high levels of Kd (Wetzel & Neckles 1986),and Z. marina appears most sensitive to eplphyte lighthmrtatron at hrgh \,vater temperatures (Neckles etal. 1993). Therefore, epiphyte biomass may have con-tributed to redr:ced macrophyte growth upriver duringthe spring turbiclity peak.
Chlonic lvater column nitrate enrichment has been re-lated to eelgrass declines tn some lnesocosm ennchmentexperiments (Burkholder et al. 1992, 1904). Athough rhemechanism is not understood, it is hypothesized thatchronic i.vater column nitrate enrichment may promoteinternal nutrient imbalances that lead to plant death. Inour study, diflerences in niirate concentrations betlveenY0 and Y26 were generally less than 1 prM, especiallyduring the spring and summer. This level of enrichmentsuggests that. nitrate toxtcity \{as not a significant con-lributor to eelgrass dechnes in ttre York River
Conclusions
The lack of regrorvth of Zostera marina into formerlyvegetated sites in a lower Chesapeake Bay tt'tbutar)' isnot simply due to lack of plopagules but can be relatedto environmental conditions, especially high levels ofturbrdity during spring periods ol potentjally maximumgrorvth and carboirydrate storage. Prolonged periodsof nitrogen enrichment drrrrng the fall and winterhad no observable effect on epiphytic accumulationsor macrophlite grOrvth, plesuntably because of Over-rtding control by other factors. However, the accumu-lation of an epiphylic matnx on t.he leaves during thespring may contribute to an injtidtlon of the seagrassdecline. Symptoms of LabyrinthtLla infection were notobserved. We suggest that insufficient grorvth duringthe spring limits Z. manna survival through the sum-mer. Although summertinte conditions may stress eel-grass poprLlations in this region, they do not alone limitlong-tenn survival. Relatively short-term stresses dur-inq certain criticai periods can therefore have iastinqelfects on seagrass popr,rlations. Water quality condi-tions enhancing adequate seagrass growth dunng thespring nay lte key to long-term Z. ntarina survival andsuccessful recolonization in this reqion.
251
Acknov,ledgetnents. Thts research rvas supportecl by TheComnronu,ealth of Virgirrra, Chesalteake Bay SubnrersedAquatrc Vegetai)on lnitiative ancl a private grant f rom :\lliedSignal Foundatrolr Tl.re authors especrally thank B. Nt.rkirklor l.ll)oratory and lielcl assistanco. Thrs is contrrbutlon no.202"1 lronr the \rirryinia InstltotL' ol \.1.inne Scienr:e, School ofN'lanne Science, (iellsgs of Wrliiam ancl MarV.
LITERTVTURL C-ITED
Aroi K, Mukar H, Koike L OhtsLr N4, Hattori A (1981)Cr0rvthand organic production o[ eelqr,rss iZostera llari.na L.) inlemperate rvatels of the Pacific coast oI Japan. Il. Grorvthanalysis in winter. Aquat Bot 10:1'/5-182
Batiuk RA, Orth RJ, l.4oore KA, Dennison lVC, Sterrenson JC,Staver LW, Carter V, Rybicki NB, Hickman RE, Kollar S,Bieber S, Heasly P (1992) (lhesapeake Bay submergedaquatic vegetation habitat roquiremcnts and restorationgoals: a technical synthesis. USEpA CtsP/TRS83/92, Ches-apeake Bal, Program, Annapolis, MD
Biebl R, NlcRoy CP (1971) Plasnratlc resistance and rate ofrespiration and photosynthesis of Zosler? marjna at differ-eni salinitles and temperatures. Mar Biol B:48-56
Borunr J (1987) Dynamics of epiphyton on eelgrass (ZosterantarinaL.) leavesi relative roles of algal growth, herbivory,and substratun] turno\rer. Limnol Oceanogr 32:986*992
Bonrrn J, \,Vrum'Andersen S (1980) Biomass ancl procluctron ofepiphytes on eelgrass izosten ntarina L.J in lhe @resund,Denmark. Ophelia Suppl 1:.57-64
Bultlnris DA (1983) Elfects o[ in sjlu light reductlol] on den-sity 6n6 growth of lhe seaclrass Hetck)zostera lasmanica(trl.lrtans ex Aschers.) clen Flartog in Western Port, Vic-toria, .,\rrstraha. J Exp Ir4ar Btol Fcol 67:91-103
Br.rrdick DM, S.hort F'l Wolf J (1993) An index to assess anclmonitor the plogression of rvasting disease in eelgrassZostera ntarina. Mar Ecol Prog Ser 94:83-90
Burke N4K, Dennison WC, Moore KA (1996) Non-structuralcalboh1,6ly319 reserves of eelqrass Zostera rnanna. MarEcol Prog Ser 137:195-201
Bulkholder JM, Mason KM, Glasqorv HB Jr {1992) Water-column nitrate enrichment Jlrornotes decline of eelgrassZostera manna L.: evidence from seasonal nlesocosnexp(!rinrents. Mar Icol Prog Ser tj l : 1{j3- 1 78
Burkholder JM, Glasqorv Hll Jr, Cooke JE (1994) Compara-tlve cffects of u,ater-column nitrate enrichment on eel-grass Zosfera marjna, shoalgrass ]laloclule wriqlttit, andv,,irlgeongrass fluppia ntaritit??a. Mar Ecol prog Ser 105:121-138
Canbri<ige ML, Chiffings A\/, Brittan C, N4oore I-, McContbAJ [1986) The loss of seaqrass in Cockburn Sound,V\/estern z\ustralia. Ii. Possiblc causes of seagra-ss decline.Aquat Bot 24:269-285
Cattaneo A (19831 Grazrnq on epiphytes. Limnol Oceanollr28:124-132
Davis CJ, Cary DF (i981)Trer-rds in strbmer<;ed macrophl,lqconmunlt)es of the Currituck Sound: 1977 1979. J AquatPlant N4anage 19:3-B
Dawes (:, Chan M, Chinn R, Koch E\{, Lazar A, Tomasko D(1987) Proximate composition, photosynthe[ic and respira-tory responses of the seagrass Halophila engeJmanniiIrom Florida. Aquat Bot 27:1(15-201
Dawes CJ, Larvrence Jlvl 1 1979) Eff ecLs of blacle renroval on theproximate composition of the r'hizome of the scaqrass Iha,/assia lestuirdjnutn Banks ex Kon jcl. Aquat Bot 7:255-266
Dennison WC, Orth RJ, lvloorr: K.A, Stevenso,t JC, tlarter V,Kollar S, Bergstrom PW, Batiuk RA t1993) Assessrng
N4oorr: ert al-: Zoslera narirra ctrorvth and survival

258 N,lar Ecol Proq Ser 142.247-259, 1996
water quality rvith submersed aquatic vegetation, Biosci4 3:86-94
Dennison W(.'. .\lberte RS (1985) Role of daily hght period inthe depth distribution ol Zostera nrdflna (eelgrass). VlarEcol Prog Ser 25:51*ii1
den Harto!, D, Polderman PJG [1975) Chanr;r:s in the seagrasspopuldtions of the Dutch Waddenzor':_ Aquat Bot 1:141 .1'r7
Drerv E-'\ (1983) Sugars, cyclitols, and seagrass phylogenv.Aquat tsot 15:387-408
Duarte (-M (1991) Seagrass depth limits. Aquat Bot 40:363..377
Evans AS, Webb KL, Penhale PA (1986) Photosynthetic ter!.r-pelature acclimation in two coexisting seagrasses, Zosteramarina L and Rr"rppia maritima L. Aquat Bot 24:185-197
Fonseca MS, Fisher JS, Zieman JC, Thayer GW t1982) Influ-ence.of the seagrass, Zostera ntarina L., on current flolv.Llstuar Coas! Shelf Scr 15:351-364
Fonseca MS, Kenrvorthy lVJ, Thayer GW, Hel.ler DY, CheapKM (1985) Transplanting of the seagrasses Zostera marinaand Halodule wriqhtii for sedlment stabilization and habi-tat development on the east coast of the United States.Technical Report EL-85-9, U.S. Army Engineer Water-rvays Experiment Station, Vicksburg, MS
Giesen WIIJT, r'an Katrvljk MM, den Hartog C (i990) Ee1-grass condition and tr"rrbidity in the Dutch Wadden Sea.Aquat Bot 37:77-85
Hough RA, Fornwall MD, Negele BJ, Thompson RL, Purt DA(i9BS) Plant community dynamics in a chain of lakes:principal factors in the decline of rooted macrophytes witheutrophication- t-lydrobiologia 1?3:199-217
Howard RK (1982) Impact of feecling activrties of epibenthicamphipods on surface-fouling of eelgrass leaves. AquatBot 14:91-97
Jacobs RPL\:Nl (1979) Distribution and aspects of the produc-lion and biomass of eelqrass, ZosteQ mailna L., at Roscoff,France. AquaL Bot 7:151,,172
Jupp tsP, Spence DHN (l-977) Limitations on macrophytes rn aeulrophic lake, Loch Lr:r'en. J Ecol 65:175-186
Kemp Wlvl, Boynton WR, Twiliey RR, Stevenson JC, MeansJC t1983) The decline of submerged vdscular plants inupper Chesapeake Bay: summary of rcsu.lts concerningpossible causes. Mar Technol Soc J 17:78*89
Kenrvorthy WJ, Zieman JC, Thayer CW (1982) Evidence forthe inlluence of seagrasses on lhe benthic nitrogen in acoastal platn estuary near Beaufor[, North Carolind lUSA).Oecoloqia 54:1J2-158
Kerr EA, Strother S (1985) Effects of irradrance, temperatureand salrnily on photosynthesrs of Zostera mue11err. AquatBot 2It: 1 )i- 183
Libes M (19861 Productivity-jrradl.tnce relationship of Posido-nia oc:eantca and its epiphytes. Aquat llot 26:285*306
|.lursh GA (1970) A seasonal study of Zo,qtera epibiot(r in theYork River, Virginia. Phl) disscrtation, Colleqe of Williamancl Nlary, \,Villiamsburg, VA
Marsh (.;A (i973) 1'he Zostcra epifaunal cornmunity in lheYork River, Vlrrlinia. (lhesapeake Scr 14:87 -97
lvluehlstein LK, Porter D, Short FT (1988) Labyrinthula sp., anrarine slime mold producing the syntlrtom_s of !v.tsltnrldlsease tn eelqrass, Zostcra marina. \,lar Biol 99;4;i5-47'2
Murray L (1983) Nletabohc and stntctural studies of severaltempercltrr seagrdss communitics, r,r,ith emphasrs on mlcro-alqal components. phD dissertatrrin, Collrclc of Williamanrl \{ary, lVillamsburg, 1'A
Neckles HA, \.\,etzcl RL, Orth RJ {1993) Relcrtive effucts ofnutrient enrtchrncnt and grazin g on epiph,!..te-m.tcrophyte(2. murna L.J clynamrcs. Oecologia 93:285-295
Neckles HA, Koepfler ET, Haas LW, Wetzel RL, Orth RJ(1994) Dynamrcs of epiphytrc photoautotrophs and het-erotrophs \n Zostera mariJl(t (eelgrdssl mlcrocosm: re-sponses to nutrient enrichrnent and grdzing. Estuarics 17:597 -60.;
Nr:ter J, Wasserman W (1974) Applied linear statisticalmodels. Rrcharcl D. Irlvin. lnc, Flonres'oocl
Ogata E, Matsui T (i9651 Photosvnthesrs in se."'eral marineplants of Japan as affected by salinity, rirying and pH, rvithattention to their growth habitats, Bot Mar 8:199-217
Orth RJ (1973) Benthic infauna of eelgrass, Zostera marina,beds. (lhesnpe,ake Sri 14.25R-?69
Orth RJ (1975) Destructlon of eelgrass, Zostera manna, bv tl.recownose ray, Rhinoptera bonasus, in the Chesapeake IJav.Chesapeake Sci 16:205 -208
Orth RJ (1977) The importance of sediment stability in sea-grass comlrunities. In: Coull BC (ed) Ecology of marinebenthos. IJniversity of South Carolina Press, Columbia,p 28 1 -300
Orth RJ, Moore KA (1983) Chesapeake Bay: an unprece-dented clecline rn submerged aquatic vegetation. Science222:51-53
Orth RJ, Moore XA (i 984) Distribu tion and abundance of sub-merged aquatic vegetation in Chesapeake Bay; an histor-ical perspective. Estuaries 7:53 I *540
Orth RJ, Moore KA (1988) Distribution of Zostera marina L.and .Ruppia nlarilima L. sensu lato along depth gradielttsin the lorver Chesapeake Bay, USA. Aquat Bot 32;291-305
Orth RJ, Moore KA, Gordon FIH (1979) Distribution andabundance of submerged aqudtic vegelation in the lolverChesapeake Ba1', Virginia. Final Report. 600/8-79-OZg/SAV1 US EPA, Annapolis, MD
Orth RJ, Nor,vak JF, Anderson CA, vVhiring JR (1993) Dis-tribution of submerged aquatic vegetation rn lhe Chesa-peake Bay and iributaries and Chincoteague Bal'"..-1g92.Final Report. USEPA, Cl'resapeake Bay Program, Anna-polis, lrlD
Olt JA (1$80) Crowth and production \n posidonia oceanica(L..) l)elile. PSZN I: Mar Ecol 1:47*64
Parsons TR, Miata Y, Lallj CM (1984) A manual of chemrcaland biological methods for searvater analysis. PerganronPress. New York
Penhale PA (1977) Macrophyte-epiphyte biomass and pro-ductivtty in an eelgrass (Zostera marina L.l comntunjty.J Exp Mar Biol Ecol 26:211-224
Phil.lips CL, Eminson D, Ivloss B (19781 A mechanism toaccount for macrophyte decltne in progressively eutrophi-cated freshrvat.ers. Aquai Bot 4:103-126
Prnncrup SP {1980) Leaf production of. Zostera ln.rrina L. atdifferent salinjties. Ophelia Suppl 1:2ir:),224
Pirc H (1985) Growth dynamics in Posidonia oceanica (L)Delile. L Seasonal changes o{ soluble carbohvdrates. starch.free amino acl.li, nitroqen and organic anlons in dilferenLpdrts of the pl.')nt. PSZN I: Mar Ecol 6:141*165
Rasnrussen E (1977) Thc rvasting drsease of eelgrass (7-osterctnaioaJ and its effecl.s on environmantal factors andlauna. I*: \lcRcry CP, Helffenc:h C (ecls) Seagrass e<_'o-
systcms: a scicntific perspective. \larcel Dekker, \ew YorkSand-Jens{rn K (197j1 Bromass, net production and cjrowth
dynamrcs in an t:olgrass lZostera marina L..l populatic,n inVellerup Vrg, De:nmark. Ophelra 14:185..20I
Shoat WT, Lrum B\U {1976) lrnproved extrdction of chloro-phl.ll .r and b from alctar. usinrl clrrnethvl sulfoxrde. LimnolOceonoqr 21:926-928
Short FT. ll:elings BlV, dt:n Ilartog C f 1988) C_'omparison ofa currr.rnt eelgrass drscase to the $.dstrng disease rn the1930s. Aquat Bot 30:295-304

lr4oore et aI.: Zostera marina grolvth and survival 255
Silberstein K, Chiffings AW, Mccomb AJ (1986) The loss ofseagrass in Cockburn Sound, Western Australia. III. Theeffect ol epiphytes on productlvity of Posidonia austraJisHook. f. Aquat Bot 24:355..37 1-
SPSS Inc (1986) SPSSx users' guicle, 2nd edn. ChicagoStevenson JC, Staver LW, Staver KW {1993) Water quality
associaLed lvith survival of submersed aquatic vegetationalong an estuarine gradienL. Estuaries 16:346-361
Trtus JE, Adarns MS (19791 Comparatrve carbohydrate stor-age and utilization patterns in the submersed macro-phytes, Myriophy)lum spicatum and Va.llrsnena dmeri-cana. Am lv'lidl Nat 102:263-271
Trvilley RR, Kemp \4rM, Staver KW, Stevenson JC, BoyntonWR {1985) Nutrient enrichment of estuarine submersedvascular plant communities. L Algal growth and effects onproduction of plants dnd assoc'iated communities. lr.larEcol Prog Ser 23:179-191
USEPA (1979) Manual of methods for chemical analysisof lvater and wastes. EPA-600-4-79-020. Environmental
This arlicle was presented by K. L. Heck Jr (Senior EditorialAdvisot), Dauphin Island, Alabama, USA
Monitoring and Suppolt Laborator,v, Cincinnati, OHvan Montfrans J, Orth RJ, Val' SA (1982) Preliminary studies
of grazing by Bittium uar-ru;n on eelgrass periphyton-Aquat Bot 14:75-89
lVard LG, Kemp Wlr4, Boyton \,VR (198a) The influenceof lvaves and seagrass communities on sr.rspendedparticlates lrl an estuanne embayment. Mar Ceol 59:85- 103
Wetzel RL, Neckles HA (1986) A model of Zostera marina L.photosynthesis and grorvth: srmulated effects of selectedph)'sical-chemrcal variables ancl ltiological interactions.Aquat Bot 26:307-323
Wiltmann KJ, Ott JA t1982) Effects of cropping on growth inthc Mediterranean seagrass Posjdonia oceanica (L.) Delile.PSZN l: Mar Ecol 3:151-159
Zimmerrnan RC, Reguzzone JL, Wyllie-Eche verria S, JosselynM, Alberte RS (1991) Assessment of environmental suit-ability for growth of Zostera marina L. (eelgrass) in SanFrancisco Bay. Aqudt Bot 39;353-366
Manuscript lirst received: July 17, 1995Revjsed version accepted: Ju)y g, 1996