Embed Size (px)
Citation preview

Growth of native tree species planted in montanereforestation projects in the Colombian and EcuadorianAndes differs among site and species
Matthew C. Bare1 • Mark S. Ashton1
Received: 9 April 2015 / Accepted: 28 October 2015� Springer Science+Business Media Dordrecht 2015
Abstract The tropical Andes in Ecuador and Colombia are a biodiversity hotspot that has
endured centuries of forest clearance and degradation. Forest restoration has been identi-
fied as a regional conservation priority; in recent decades, native species reforestation
projects have proliferated, but little information exists on growth performance of com-
monly planted tree species in relation to site and soil nutrient status. This study analyzed
growth of seven common native species (Alnus acuminata, Baccharis bogotensis, Cedrela
montana,Myrica pubesens, Quercus humboltii, Sambucus nigra, Smallanthus pyramidalis)
on 12 montane forest sites across the northern region of the tropical Andes. Andean alder
(A. acuminata) was the most commonly planted species, and grows at a mean annual
diameter increment (MAI-d) of 1.81 cm y-1 and a mean annual height increment (MAI-h)
of 0.95 m y-1. S. pyramidalis, a short lived pioneer of the Asteraceae family, also
exhibited fast growth rates of 1.64 cm MAI-d and 1.21 m MAI-h. Andean oak (Q. hum-
boltii) was the second-most commonly planted species, growing with an MAI-d of 0.99 cm
and MAI-h of 0.56 m. Soil magnesium and potassium were significant predictors of MAI-d
and MAI-h for A. acuminata, while soil nitrogen, phosphorous, sodium, and calcium were
negatively associated with growth (p\ .001). We speculate that A. acuminata did not
grow as well on soils richer in calcium and phosphorus because they were less conducive
to nitrogen symbiosis common to this species. Soil magnesium and calcium were signif-
icant predictors (p\ .05) of diameter growth for Q. humboltii. For both species, we
attribute growth responses to soil nutrients as a result of the variable nature of fertility in
the complex and variable soils that make up the volcanic and surficial geological landscape
of the northern Andes. Results indicate that native species can grow in a variety of soil
conditions, and exhibit growth rates comparable to non-native species. However, our
results suggest native species are site restricted for best growth and should be planted on
particular soils. We make recommendations for reforestation for the species in this study.
& Matthew C. [email protected]
1 Yale School of Forestry and Environmental Studies, 360 Prospect Street, New Haven, CT 06511,USA
123
New ForestsDOI 10.1007/s11056-015-9519-z

Keywords Restoration � Tropical Andes � Growth rates � Alnus acuminata � Quercushumboltii � Nitrogen fixation � Smallanthus pyramidalis � Soil fertility
Introduction
Worldwide, the area of degraded forest and abandoned agricultural land is growing, and
both passive and active forms of restoration have the potential to conserve biodiversity,
stabilize eroded landscapes, and sequester carbon (Silver et al. 2000). Across the tropics,
large areas of abandoned and marginal agricultural land have potential for establishment of
second-growth forest (Chazdon 2008), and restoration through enrichment planting (Millet
et al. 2013) or native tree plantations (Wishnie et al. 2007; Lamb 2011; Rodrigues et al.
2011). Such restoration activities can facilitate forest recovery and augment forest tree
diversity (Parrotta et al. 1997; Holl et al. 2000; Ashton et al. 2001; Lamb et al. 2005).
The tropical Andes biodiversity hotspot, stretching from Venezuela to Chile, is an
important ecological region with great potential for forest restoration (Conservation
International 2014). Within the tropical Andes, the northern Andean montane forests of
Colombia and Ecuador (Dinerstein et al. 1995) hold high rates of species richness and
endemism (Gentry 1992), but have lost significant areas of natural forest (Etter et al. 2008).
However, many areas of the northern montane forest are experiencing forest regrowth:
between 2000 and 2010, Colombia gained approximately 23,773 square kilometers of forest
and Ecuador 5867 square kilometers, both largely in the montane regions (Aide et al. 2013).
Worldwide studies find that approximately an equal amount of forest area is undergoing
reforestation or natural regrowth as is being deforested (200,000–300,000 km2 y-1), and
that much of the regrowth area is in hilly and mountainous regions on marginal soils and
steep slopes such as in the tropical Andes (Asner et al. 2009).
Within the northern Andean montane forest, conservation of native forest fragments and
forest restoration are conservation priorities (Conservation Internacional Colombia 2014).
Fragments and native tree plantings in these forests are found to be an important source of
plant and bird biodiversity (Kattan et al. 1994; Murcia 1997; Gilroy et al. 2014). One
commonly planted montane tree is Andean alder (Alnus acuminata Kunth.), a fast growing
nitrogen-fixing tree, useful for soil enrichment in silvo-pastoral systems. A comparison of a
native A. acuminata plantation with natural forest regeneration in the Colombian Andes
found that A. acuminata plantings are taller than natural forest re-growth with a similar
basal area, although natural forest regrowth had higher total plant richness (Murcia 1997).
Studies in southern Ecuador have found that A. acuminata can achieve fast growth rates,
even greater than exotics such as Eucalyptus saligna and Pinus patula, especially in gaps
(Weber et al. 2008; Gunter et al. 2009).
Despite the importance of forest restoration in the northern Andes, few studies exist on
the growth rates of common native species in diverse sites. Most research examines the
growth of A. acuminata (Cornelius et al. 1996; Rıos et al. 2004; Medina et al. 2008; Weber
et al. 2008; Gunter et al. 2009); these studies test the growth of A. acuminiata of different
seed provenances, or different strains and application methods of the nitrogen-fixing
Frankia bacteria associated with Alnus. In southern Ecuador, a few studies have evaluated
the growth of other native species such as Heliocarpus americanus, Piptocoma discolor,
Cedrela montana, and Juglans neotropica in experimental plots; these studies find that
native early successional trees such as A. acuminiata, H. americanus, and P. discolor grow
New Forests
123

fast in planted sites (Weber et al. 2008; Gunter et al. 2009). These studies have also found
that seed selection, provenance, and quality are important determinants of planting success,
in addition to silvicultural treatments such as weed control (Stimm et al. 2008; Weber et al.
2008). In this site, Wilcke et al. (2008a) showed that tree growth is also affected by soil
fertility, altitude, and land use history; studies in other tropical montane ecosystems make
similar conclusions (Grubb 1977; Bautista-Cruz and del Castillo 2005). In the Neotropics
more generally, numerous studies have measured growth rates of commonly planted
lowland trees (Worbes 1999 in Venezuela; Hooper et al. 2002, Griscom et al. 2005, and
Wishnie et al. 2007 in Panama; and Haggar et al. 1997 and Piotto et al. 2003 in Costa
Rica). Elsewhere in the tropics, studies have shown that selected native species can achieve
growth rates comparable or superior to those of fast growing exotics (McNamara et al.
2006; Schneider et al. 2014).
Nevertheless, although a few studies provide information on growth of selected species
in experimental conditions, growth data from a wider range of field conditions and soil
nutrient concentrations is necessary for a better understanding of commonly planted spe-
cies in the northern Andes. In the tropics generally, old and lowland tropical soils are P
limited whereas young, highland tropical soils are more likely to be N limited (Vitousek
and Farrington 1997; Davidson et al. 2004). In the northern Andes, however, soil condi-
tions are highly variable, often a result of volcanic activity and landslides (Bussmann et al.
2008; Wilcke et al. 2008b). There is very little grey literature and no published data on
many tree species that have been more recently planted, such as Smallanthus pyramidalis
Triana., a fast growing Asteraceae tree commonly used for erosion control and live fences.
Another important tree of montane forest, both ecologically and for timber, is Andean oak
(Quercus humboltii Bonpl.). This tree is found in Panama, Venezuela, Colombia, and
Ecuador, and dominates montane forest across much of its range in Colombia (Leon et al.
2009), but few published data are available on its growth rates and soil affinities.
Restoration practice is growing in the tropical Andes, especially in Colombia and
Ecuador (Murcia et al. 2015). State ministries have directed large-scale reforestation
projects with exotic timbers since the middle twentieth century, and since the 1990s, non-
governmental organizations, international donors, and research institutions have become
engaged in numerous small projects focused on ecological restoration (CESA-Intercoop-eration Suiza 1992; Endo 1994; Meza et al. 2006; Andrade-Perez 2007; Farley 2007). Most
recently, private sector financing has become more available for restoration via payments
for watershed services, climate change mitigation, and mining offsets (Bare 2014).
However, while many agencies publish general guides, little specific data are available on
native tree plantings.
Colombia has recently prepared a national plan for restoration (Ospina Arango and
Vanegas Pinzon 2010), largely based on the Society for Ecological Restoration primer
(Society for Ecological Restoration 2004). This plan describes restoration of forest
ecosystems with goals of providing habitat connectivity, biodiversity, and ecosystem
functions like water provision, however, it offers little guidance on species selection and
background of established field projects in Colombia. In Ecuador, detailed information
exists about a study site in the southern Andes, but little information is available about
plantings elsewhere in the country.
The first objective of our study was to build on what little has been published and to
evaluate the growth performance of a range of native tree species planted in forest
restoration projects in the northern Andean montane forests. Our second objective is to
compare growth of the most commonly selected tree species in relation to soil nutrition on
reforestation sites.
New Forests
123
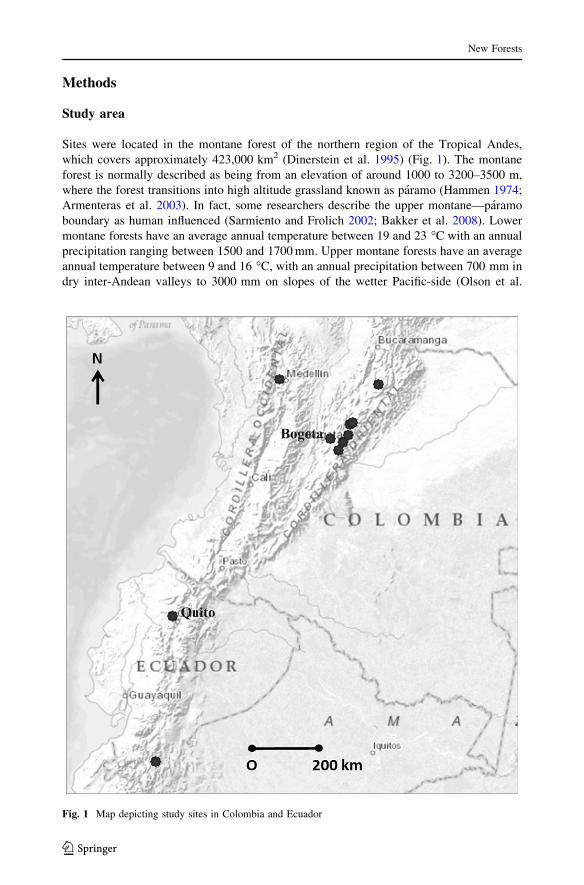
Methods
Study area
Sites were located in the montane forest of the northern region of the Tropical Andes,
which covers approximately 423,000 km2 (Dinerstein et al. 1995) (Fig. 1). The montane
forest is normally described as being from an elevation of around 1000 to 3200–3500 m,
where the forest transitions into high altitude grassland known as paramo (Hammen 1974;
Armenteras et al. 2003). In fact, some researchers describe the upper montane—paramo
boundary as human influenced (Sarmiento and Frolich 2002; Bakker et al. 2008). Lower
montane forests have an average annual temperature between 19 and 23 �C with an annual
precipitation ranging between 1500 and 1700mm. Upper montane forests have an average
annual temperature between 9 and 16 �C, with an annual precipitation between 700 mm in
dry inter-Andean valleys to 3000 mm on slopes of the wetter Pacific-side (Olson et al.
Fig. 1 Map depicting study sites in Colombia and Ecuador
New Forests
123

1995). Vegetation changes rapidly along the altitudinal gradient. Lower elevation forests
are dominated by palms (Arecaceae), and the angiospermous families Melastomataceae,
Lauraceae, Rubiaceae, Malvaceae, and Fabeaceae. Higher elevation forests transition to
Myricaceae, Fagaceae, Clusiaceae, Ericaceae, and Asteraceae, while palms and Melas-
tomataceae remain common (Armenteras et al. 2003; Homeier et al. 2008). Much of the
higher montane forest on wet slopes in Colombia is dominated by Andean oak (Q. hum-
boltii) (Leon et al. 2009). Higher elevation forests have lower total species richness but
normally higher rates of endemism, particularly among vascular epiphytes (Gentry 1992).
Most of the northern Andes as a whole are volcanic ash derived soils (Andisols). These
soils typically display high amounts of soil carbon, dense networks of micropores, high
water retention, and low bulk density (Tonneijck et al. 2010). Aluminum–humus com-
plexes are common in acidic soils (Zehetner et al. 2003). Other areas in the Andes shaped
by erosion and landslides or that were not derived from volcanic ash deposits are classified
as Entisols or Inceptisols (Cambisols) (Wilcke et al. 2008b). Throughout much of the
northern Andes, landslides are common and play a significant role in the soil heterogeneity
and shaping forest vegetation (Bussmann et al. 2008).
The northern Andean montane ecosystem is the most densely settled area of Ecuador
and Colombia, and agriculture has been the primary driver of land use change over the past
several centuries (Sarmiento and Frolich 2002; Etter et al. 2008). In Colombia, approxi-
mately 75 % of the population lives in the Andean region; in Ecuador it is approximately
50 % (Colombia Censo General 2005; Ecuador en Cifras Resultados 2015). Land use
activity in the higher areas is most often associated with potato cultivation, and coffee and
fruit trees in the lower lying areas, although cattle pasture is common throughout the region
and is replacing some agricultural crops (Etter et al. 2006; Guhl 2008). Coffee cultivation
is focused on areas of better access and soil fertility, whereas cattle pasture and estab-
lishment of natural second-growth is more likely to occur on less fertile soils and on steep,
remote mountain areas (Etter et al. 2006; Asner et al. 2009; Aide et al. 2013). Across the
region, forest fragments are spread throughout the agricultural landscape (Etter et al. 2006;
Murgueitio et al. 2011; Lerner et al. 2014). Mining is practiced throughout the region, but
is a minor driver of forest clearing compared to agriculture (Etter et al. 2008).
While most forest clearing in the Andean montane ecosystem occurred over the past
centuries, some clearing continues today. In Colombia nationwide, Hansen et al. (2013)
calculate a rate of forest loss of 3.0 % from 2000 to 2012 or an average annual loss of
0.25 % (in the forest cover[25 % threshold). The areas with highest amounts of forest loss
were the departments of Caqueta, Meta, and Antioquia; all located primarily in montane—
lowland forest transition zones of the Andes. In Ecuador, Hansen et al. (2013) calculate a
2.7 % forest loss from 2000 to 2012 or 0.23 % average annual rate of forest cover loss. The
areas with the highest amounts of forest loss were the lowland Amazon and coastal regions,
although other studies find high continued rates of forest loss in montane regions of
southern Ecuador (Tapia-Armijos et al. 2015).
Site selection
Our study sites were focused on projects with a native forest restoration goal. Timber
plantations using exotic trees were excluded. Forest restoration projects were identified
using expert interviews and internet search, followed up with interviews with key infor-
mants. Project managers from various sectors (academic, government, NGO) were con-
tacted for background information, project reports, and interviews. Seventy projects,
primarily in the montane region of the two countries, were identified and catalogued.
New Forests
123

Project data was triangulated when possible using written reports, interviews with project
managers, and third party interviews with other restoration practitioners in the region.
Of seventy projects catalogued as the most promising based on our initial survey and
interviews, sixteen were selected for field visits and data collection. These sites were
selected because they had an implementation record of [5 years (established prior to
2008), availability of information about date of planting and species planted, and a basic
record of site protection (not necessarily maintenance of the planted trees, but merely
protection of the site from fire or re-conversion to pasture or agriculture). Of the 16 sites
where data were collected, four were discarded after site survey because of site aban-
donment or unclear planting data; the remaining 12 were finally used for data analysis. The
goals of these restoration projects were for a mix of purposes including scientific research,
conservation and landscape connectivity, watershed protection, and agroforestry (live
fences, nutrient retention). Most sites were fenced to prevent cattle grazing, but not fer-
tilized and only minimally weeded (see Appendix 1 for complete site details and
objectives).
Sampling design
Because sites were derived from a variety of sources, planting design and species varied
greatly. In the study sites, trees varied between 5 and 15 years of age and were spaced
between 2 and 5 m. Study sites were grouped into large ([2 ha) and small (\2 ha). Small
sites were sampled via census (all trees were measured); in large sites, three line transects
crossing the entirety of the planting site (50–500 m) were selected randomly, and all trees
within 5 m of the center line were measured. In all sites, planted trees with height greater
than diameter at breast height (dbh) (1.3 m) were identified to species, measured for dbh,
and ocular estimates of tree canopy height made to the nearest half meter, using a survey
rod for calibration. Data was collected in 2013 (see Appendix 1 for details).
Soil samples were collected from 3 to 5 random locations at each restoration site;
approximately 100 g were collected from the A horizon, approximately 2–4 cm below the
mineral soil surface. Composite samples from each site were analyzed in the field for
texture and then air dried and shipped to the U.S. for chemical analysis. Air-dried samples
were analyzed for pH at the Yale School of Forestry and Environmental Studies soil
laboratory (Greeley Memorial Laboratory). Soil carbon, organic matter, cation exchange
capacity, and additional nutrients (N, P, K, Na, Mg, and Ca) were analyzed at the
University of Georgia Department of Crop and Soil Sciences Lab for Environmental
Analysis. Dissolved carbon was determined using a Shimadzu TOC-5050A Total Organic
Carbon Analyzer. Analysis was based on loss of ignition (David 1988). All other elements
were determined by spectrophotometric methods and ion chromatography using a DIO-
NEX DX500 modular system (University of Georgia University of Georgia Laboratory for
Environmental Analysis 2015).
Data analysis
Based on the data gathered, a total of seven species were identified for data anlysis: A.
acuminata Kunth. (Betulaceae, known locally as aliso, or Andean alder in English), B.
bogotensis Kunth. (Asteraceae, known locally as chilco), C. montana Moritz ex Turcz.
(Meliaceae, known locally as cedro de montana), Myrica pubesens Humb. & Bonpl. ex
Willd. (Myricaceae, known locally as laurel de cera), Q. humboltii Bonpl. (Fagaceae,
known locally as roble, or Andean oak in English), S. nigra L. (Adoxaceae, known in
New Forests
123

English as elderberry, native to Europe), and S. pyramidalis Triana. (Asteraceae, known
locally as arboloco). Mean annual increment for height and diameter were calculated by
taking measurements in 2013 and averaging growth over the age of the tree. Two-way
analysis of variance (ANOVA) was performed to test significance of species, site, and
interaction effects on mean annual increments of diameters and height (MAI-d, MAI-h) of
the seven common species in the study. Mean annual increments were compared among
sites and among species for statistical significance using a Tukey test. Multiple regression
analysis (backwards stepwise, Minitab 16) was used on soil nutrient variables for the MAI-
d and MAI-h of the two most commonly found trees, A. acuminata and Q. humboltii. Soil
texture, pH, organic matter, soil carbon, cation exchange capacity, aluminum, and site
elevation showed no significant effects and were removed from further analysis. Sampling
design and data analysis draw significantly on a similar study conducted by (Schneider
et al. 2014).
Results
A total of 802 trees were measured of 45 different species. Statistics were calculated on
the seven species most commonly found across the 12 sites. A. acuminata was by far the
most commonly planted species, being a popular restoration tree for its fast growth and
nitrogen-fixing ability. A. acuminata represented 44 % of all trees sampled across
reforestation sites, planted at seven sites. Q. humboltii was found at four sites and S.
pyramidalis, a fast-growing, short-lived Asteraceae tree common to disturbed areas, was
found at three sites. Mid-successional timber trees such as C. montana, Tabebuia rosea
(Bignoniaceae), J. neotropica (Juglandaceae), and Podocarpus spp. (Podocarpaceae)
were planted at several sites, but many individuals of these species had died due to
intense competition with pasture grass (see Appendix 2 for a partial list of species
planted).
Growth rates between species were significantly different, as were growth rates between
sites (Table 1). The interaction effect between site and species was significant, indicating
that species differed in their growth rates and changed ranking in relation to each other
across the different restoration sites and soils. Tukey studentized t test comparisons of
growth rates among species revealed A. acuminata had the greatest diameter growth,
averaging 1.81 cm y-1, along with S. pyramidalis at 1.63 cm y-1 (Fig. 2). Growth was
significantly greater in these two species compared to all other species measured except C.
Table 1 Two-way ANOVA of mean annual increment for diameter (cm) and height (m) by sites, speciesand interaction between sites and species
Source N df Mean SD F p R2 (adj)
MAI-d (cm)
Site 12 11 1.40 0.47 5.303 \.0001 .553
Species 7 6 1.21 0.41 4.094 .001
Site 9 species 17 2.944 .001
MAI-h (m)
Site 12 11 0.82 0.26 5.368 \.0001 .573
Species 7 6 0.71 0.32 8.504 \.0001
Site 9 species 17 7.357 \.0001
New Forests
123
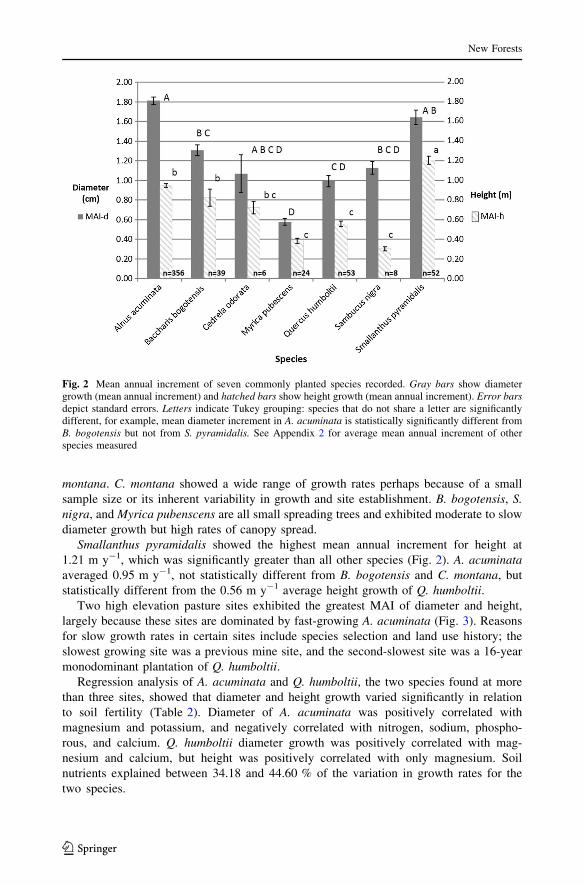
montana. C. montana showed a wide range of growth rates perhaps because of a small
sample size or its inherent variability in growth and site establishment. B. bogotensis, S.
nigra, and Myrica pubenscens are all small spreading trees and exhibited moderate to slow
diameter growth but high rates of canopy spread.
Smallanthus pyramidalis showed the highest mean annual increment for height at
1.21 m y-1, which was significantly greater than all other species (Fig. 2). A. acuminata
averaged 0.95 m y-1, not statistically different from B. bogotensis and C. montana, but
statistically different from the 0.56 m y-1 average height growth of Q. humboltii.
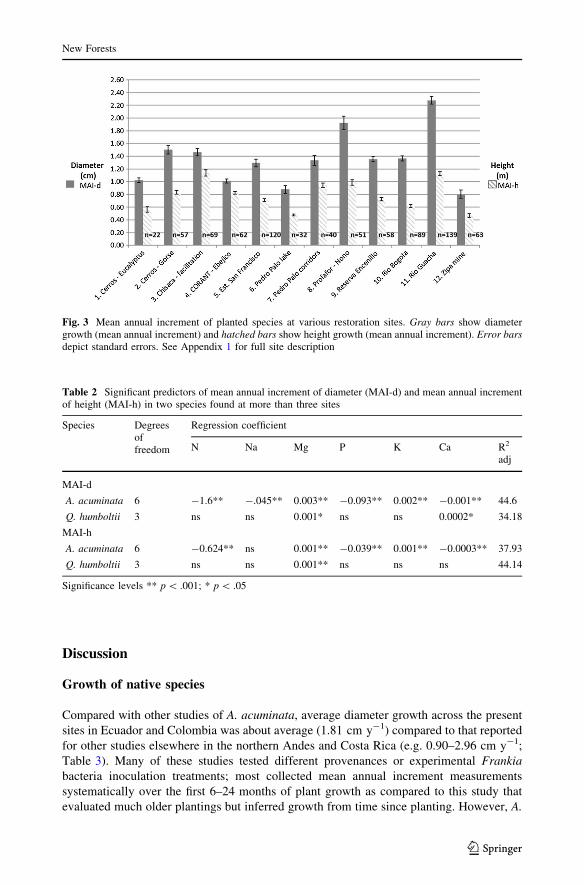
Two high elevation pasture sites exhibited the greatest MAI of diameter and height,
largely because these sites are dominated by fast-growing A. acuminata (Fig. 3). Reasons
for slow growth rates in certain sites include species selection and land use history; the
slowest growing site was a previous mine site, and the second-slowest site was a 16-year
monodominant plantation of Q. humboltii.
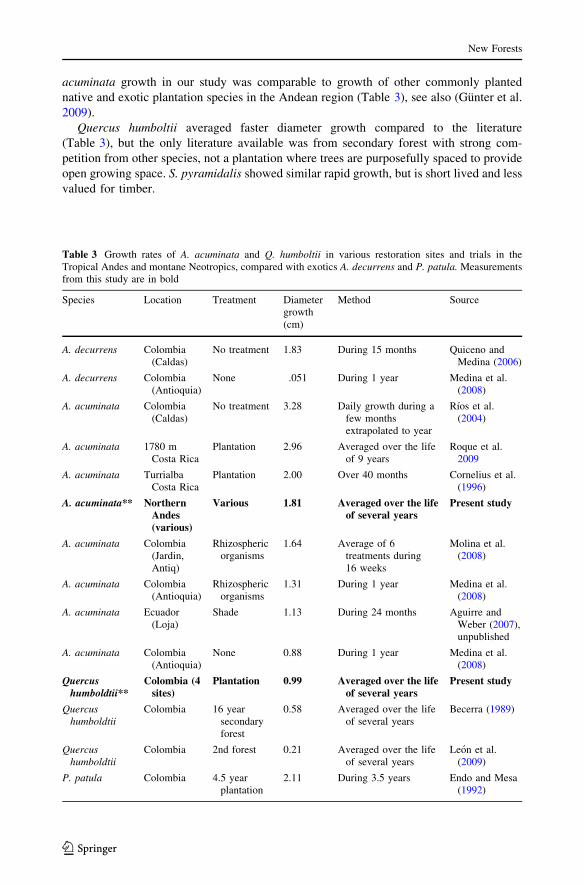
Regression analysis of A. acuminata and Q. humboltii, the two species found at more
than three sites, showed that diameter and height growth varied significantly in relation
to soil fertility (Table 2). Diameter of A. acuminata was positively correlated with
magnesium and potassium, and negatively correlated with nitrogen, sodium, phospho-
rous, and calcium. Q. humboltii diameter growth was positively correlated with mag-
nesium and calcium, but height was positively correlated with only magnesium. Soil
nutrients explained between 34.18 and 44.60 % of the variation in growth rates for the
two species.
Fig. 2 Mean annual increment of seven commonly planted species recorded. Gray bars show diametergrowth (mean annual increment) and hatched bars show height growth (mean annual increment). Error barsdepict standard errors. Letters indicate Tukey grouping: species that do not share a letter are significantlydifferent, for example, mean diameter increment in A. acuminata is statistically significantly different fromB. bogotensis but not from S. pyramidalis. See Appendix 2 for average mean annual increment of otherspecies measured
New Forests
123
Discussion
Growth of native species
Compared with other studies of A. acuminata, average diameter growth across the present
sites in Ecuador and Colombia was about average (1.81 cm y-1) compared to that reported
for other studies elsewhere in the northern Andes and Costa Rica (e.g. 0.90–2.96 cm y-1;
Table 3). Many of these studies tested different provenances or experimental Frankia
bacteria inoculation treatments; most collected mean annual increment measurements
systematically over the first 6–24 months of plant growth as compared to this study that
evaluated much older plantings but inferred growth from time since planting. However, A.
Fig. 3 Mean annual increment of planted species at various restoration sites. Gray bars show diametergrowth (mean annual increment) and hatched bars show height growth (mean annual increment). Error barsdepict standard errors. See Appendix 1 for full site description
Table 2 Significant predictors of mean annual increment of diameter (MAI-d) and mean annual incrementof height (MAI-h) in two species found at more than three sites
Species Degreesoffreedom
Regression coefficient
N Na Mg P K Ca R2
adj
MAI-d
A. acuminata 6 -1.6** -.045** 0.003** -0.093** 0.002** -0.001** 44.6
Q. humboltii 3 ns ns 0.001* ns ns 0.0002* 34.18
MAI-h
A. acuminata 6 -0.624** ns 0.001** -0.039** 0.001** -0.0003** 37.93
Q. humboltii 3 ns ns 0.001** ns ns ns 44.14
Significance levels ** p\ .001; * p\ .05
New Forests
123
acuminata growth in our study was comparable to growth of other commonly planted
native and exotic plantation species in the Andean region (Table 3), see also (Gunter et al.
2009).
Quercus humboltii averaged faster diameter growth compared to the literature
(Table 3), but the only literature available was from secondary forest with strong com-
petition from other species, not a plantation where trees are purposefully spaced to provide
open growing space. S. pyramidalis showed similar rapid growth, but is short lived and less
valued for timber.
Table 3 Growth rates of A. acuminata and Q. humboltii in various restoration sites and trials in theTropical Andes and montane Neotropics, compared with exotics A. decurrens and P. patula. Measurementsfrom this study are in bold
Species Location Treatment Diametergrowth(cm)
Method Source
A. decurrens Colombia(Caldas)
No treatment 1.83 During 15 months Quiceno andMedina (2006)
A. decurrens Colombia(Antioquia)
None .051 During 1 year Medina et al.(2008)
A. acuminata Colombia(Caldas)
No treatment 3.28 Daily growth during afew monthsextrapolated to year
Rıos et al.(2004)
A. acuminata 1780 mCosta Rica
Plantation 2.96 Averaged over the lifeof 9 years
Roque et al.2009
A. acuminata TurrialbaCosta Rica
Plantation 2.00 Over 40 months Cornelius et al.(1996)
A. acuminata** NorthernAndes(various)
Various 1.81 Averaged over the lifeof several years
Present study
A. acuminata Colombia(Jardin,Antiq)
Rhizosphericorganisms
1.64 Average of 6treatments during16 weeks
Molina et al.(2008)
A. acuminata Colombia(Antioquia)
Rhizosphericorganisms
1.31 During 1 year Medina et al.(2008)
A. acuminata Ecuador(Loja)
Shade 1.13 During 24 months Aguirre andWeber (2007),unpublished
A. acuminata Colombia(Antioquia)
None 0.88 During 1 year Medina et al.(2008)
Quercushumboldtii**
Colombia (4sites)
Plantation 0.99 Averaged over the lifeof several years
Present study
Quercushumboldtii
Colombia 16 yearsecondaryforest
0.58 Averaged over the lifeof several years
Becerra (1989)
Quercushumboldtii
Colombia 2nd forest 0.21 Averaged over the lifeof several years
Leon et al.(2009)
P. patula Colombia 4.5 yearplantation
2.11 During 3.5 years Endo and Mesa(1992)
New Forests
123

Soil nutrients
Regressions showed A. acuminata diameter and height growth was positively correlated
with magnesium and potassium concentrations and negatively correlated with nitrogen,
sodium, phosphorous, and calcium. Interestingly, our study suggests that magnesium and
calcium are strong positive and negative predictors respectively of tree growth for the two
species tested. This may be related to the large difference in fertility of northern Andean
soils of volcanic (often fertile, high pH, high cation exchange capacity, high calcium) and
non-volcanic glacial origin (often low fertility, low pH, low cation exchange capacity, low
calcium). In particular, the nitrogen fixing species A. acuminata did not grow as well on
soils richer in calcium and phosphorus (we suggest these soils are Andisols), perhaps
because they were less conducive to nitrogen symbiosis. We speculate that species growth
could also be negatively affected by other interacting site factors that we did not measure
such as wind, animal browse, site treatment (use of herbicides, weeding) or soil moisture
availability. In this study, sites practicing silvo-pastoral management protected planted
trees with fencing, but animal browsing still could have occurred from smaller native
herbivores.
The finding that A. acuminata is negatively associated with soil nitrogen is interesting,
as it is a nitrogen-fixing tree. In sites with dense stands of A. acuminata, one would expect
higher growth to correspond with higher soil nitrogen, not necessarily because the tree
responds to nitrogen fertility but because the tree enriches the soil with N fixation and
litterfall. However, it is also possible that A. acuminata grows slower in high nitrogen sites
because its association with nitrogen-fixing bacteria is absent (the symbiotic relation is not
necessary) and because of greater competition with grass and other surrounding nitrogen-
demanding vegetation, particularly when sites are not maintained (Davidson et al. 2004).
A. acuminata is also ectomycorrhizal, suggesting that it can grow well in low-nutrient
environments (Becerra et al. 2005). Similarly, a study of Alnus nepalensis in India found
that seedlings with mycorrhizal fungi grew slower in conditions of higher soil fertility (Jha
et al. 1993).
Quercus humboltii diameter growth was positively correlated with increased amounts of
soil magnesium and calcium. Height growth was correlated with magnesium. Again, like
Alnus, Quercus is an ectomycorrhizal genus. Other studies have shown that the symbiotic
plant-fungal relationship makes Quercus more nutrient-use efficient, particularly for
phosphorous and magnesium (Lehto and Zwiazek 2011; Simard et al. 2003). Other studies
have found that ectomycorrhizal presence is a stronger predictor of tree growth than soil
fertility (Zangaro et al. 2007), suggesting that parameters other than soil fertility (e.g. soil
water availability) could explain the growth of these species. Lastly, studies in the Andes
found that soil texture was an important predictor of plantation tree growth, specifically,
that trees grew better in clay loam (Henri 2001); while eroded soil significantly reduces
growth (Carpenter et al. 2004). In this study, soil texture was not correlated with growth of
planted species.
Finally, it is important to note that the height and diameter growth measured in this
study represents only one form of plant response to soil fertility. Growth allocation above-
and below-ground can differ among species. Early-successional species in fertile envi-
ronments usually allocate growth to above-ground biomass, whereas mid and late-suc-
cessional species often allocate more growth below-ground, especially in nutrient poor and
drought-stressed soils (Poorter 2001; Zangaro et al. 2007). In this study, elevation was not
found to be a significant predictor of tree growth, possibly because elevation effects were
New Forests
123

modified by species selection or site history. Some low elevation sites were slow growing
because they contained mid-successional trees instead of fast growing pioneers (sites 4 and
6 in Appendix 1), while some high elevation sites were fast growing because they con-
tained fast growing A. acuminata (sites 8 and 9 in Appendix 1). Moreover, many low and
mid-elevation sites suffered a more intense land use history with greater erosion (sites 1, 2,
4, 10, and 12), whereas the high elevation sites experienced a lighter land use history of
low intensity pasture. Finally, an additional factor not evaluated in this study is the effect
of seed provenance and quality, as well as silvicultural treatments after planting, factors
which have been found to influence tree survival and growth (Stimm et al. 2008; Weber
et al. 2008).
Management applications
Not surprisingly, mid-successional species such as Q. humobltii and C. montana showed
slower diameter and height growth than pioneer species A. acuminata and S. pyramiadlis.
In this study, C. montana exhibited slow growth but with significant variability; other
studies that monitored plantings of the related Cedrela odorata in tropical lowland sites
of Central America report similar levels of variability (Wishnie et al. 2007). In this
study, other mid-successional species, Juglans neotropica, T. rosea, and Podocarpus
spp., were planted at five sites but all were \1 m in height and many were in poor
health, with few leaves and slow growth (personal observation). These slower-growing
species likely had trouble competing with the pasture grass present in almost all sites, a
factor that has been identified to inhibit tree recovery of J. neotropica and C. montana in
southern Ecuador (Gunter et al. 2009), as well as for other mid-successional species in
other tropical montane sites (Aide and Cavelier 1994; Sarmiento 1997; Holl et al. 2000)).
Additionally, a study in one of the same field sites used in this study (Estacion Cientıfica
San Francisco, southern Ecuador) finds that lack of mycorrhizal association may be a
limiting factor for reforestation, and that inoculation can assist seedling establishment
(Urgiles et al. 2009).
In this study, short-lived pioneer trees such as S. pyramidalis, Verbesina crasirramea,
and Ochroma pyramidalis, as well as longer-lived A. acuminata may be effective at
facilitating site conditions for mid-successional species. Some site managers practice this
facilitation intentionally by first planting pioneers and later enrichment planting the mid-
successional species. Other site managers practice facilitation unintentionally by planting
all species at once, and letting the more rapidly growing pioneer trees facilitate site
conditions for the mid-successional species. In southern Ecuador, shrubs and ferns were
found to facilitate the growth of certain mid-successional species (Gunter et al. 2009),
while in a meta-study of various restoration sites, facilitation has been found to assist
tree growth in mixed species plantings (Piotto 2008). Similarly, in this study, many sites
with dense infestations of the invasive leguminous shrub Ulex europaeus (gorse, or
retamo espinosa in Spanish, a species native to Europe but common throughout the
northern Andes) are treated with mechanical removal of gorse and broadcast seeding of
the bi-annual herbaceous plant Lupinus bogotensis to provide growing space and sunlight
for pioneer tree species and hopefully prevent re-colonization by the gorse. Use of nurse
plants such as Lupinus for restoration has been described widely in the literature
(Callaway and Walker 1997; Gomez-Aparicio et al. 2004; Blanco-Garcıa et al. 2011;
Reyes and Rıos 2011). Although it is practiced for site amelioration in areas of Ulex
New Forests
123

europeus colonization, some studies have found that nitrogen-fixing plants such as
Lupinus can actually facilitate weed invasion in habitats of native plants (Maron and
Connors 1996). Elsewhere, numerous researchers have recommended facilitation or site
amelioration using fast-growing species, either native or exotic (Brockerhoff et al. 2008;
Jacobs et al. 2015). In Costa Rica, Carpenter et al. (2004) recommend Pinus tecunumanii
to stabilize soil, provide habitat for seed dispersers, suppress weeds, and moderate
microclimate conditions, thus facilitating the establishment and growth of natives. In
Mexico, de la Luz Avendano-Yanez et al. (2015) find that P. patula plantations can
facilitate survival of native mid-successional trees (although inhibit growth). Parrotta
et al. (1997) and Holl et al. (2000) conclude that remnant pasture trees and fast-growing
pioneer trees are necessary to attract seed-dispersers and facilitate restoration. Similarly,
in sites with depleted soils or extreme microclimatic conditions (sun and wind), exotics
with wide ecological amplitudes may be necessary to ameliorate conditions for native
species with smaller niches (Calvo-Alvarado et al. 2007).
Other studies in the region have found nitrogen-fixing trees such as Inga spp. to be a
valuable means of site amelioration in degraded pasture in the Andes (Rhoades et al. 1998).
A. acuminata can be a valuable component of agroforestry systems, where it can serve as a
source of nitrogen fertilization for grass and as shade for cattle (Russo 1990). In southern
Ecuador, researchers have described the economic potential of agroforestry systems where
farmers rotate pasture and A. acuminata in order to reduce pressure on cutting of new
mature forest (Knoke et al. 2009). Similarly, groups in Colombia promote silvo-pastoral
systems with nitrogen-fixing A. acuminata and cattle fodder plants such as Leucaena
leucocephala, (Calle et al. 2009) as found in the sites of Pedro Palo and Rio Guacha in this
study.
Across the region, it should be noted that restoration sites observed in this study were
designed primarily for demonstration or research purposes and required significant
resource investments for planting and site maintenance. At the same time, much of the
region is undergoing a forest transition (Aide et al. 2013), where a significant amount of
land is returning from pasture and agricultural use back to forest. A recent study in
Colombia (Sanchez-Cuervo et al. 2012) found 28,000 km2 of area was reclaimed as sec-
ond-growth from 2001 to 2010, approximately twice as much land as areas that had lost
vegetative cover (deforestation). Most of the reforested areas have occurred from agri-
cultural abandonment, a finding consistent with analysis across Latin America (Aide and
Grau 2004; Rudel et al. 2009; Aide et al. 2013). Steep mountain pasture and coffee areas
are often the most marginal and are abandoned due to urbanization, low agricultural prices,
and/or rural violence (Etter et al. 2006, 2008). In a low-elevation montane forest region of
Ecuador, one study found increasing trends of spontaneous silvo-pastoral landscapes
(Lerner et al. 2014). In highly degraded sites with low nutrient levels such as the mines and
semi-urban areas observed in this study, active restoration may be needed. However, in the
mildly disturbed agricultural landscapes that comprise a significant area of the northern
Andean montane region, active restoration can be combined with agroforestry and silvo-
pastoral systems, using fast growing species such as A. acuminata. These species provide
rapid nutrient inputs to degraded soil, and frequently have greater timber value than other
early-succession trees (Knoke et al. 2009). In most cattle pasture areas, re-vegetation with
native species occurs naturally, although active management can promote the introduction
New Forests
123

of more valuable timber species (Calle et al. 2009; Murgueitio et al. 2011). In the frag-
mented agricultural landscapes that compose much of the tropical Andes, we suggest these
types of agroforestry-oriented restoration activities in order to minimize costs while
facilitating landscape connectivity.
Conclusions
This study provides valuable baseline information on commonly planted native tree
species in montane forests of the northern Andes. Restoration projects in Ecuador and
Colombia, including the ones observed in this study, are largely focused on these
montane forests, and have multiplied in the last two decades (Bare 2014; Murcia et al.
2015). Many of these projects involve silvo-pastoral landscapes (Murgueitio et al.
2011), payments for watershed services (Goldman-Benner et al. 2012; Saenz et al.
2014), and are driven by local or national governments (Bare 2014; Murcia et al.
2015). We show that native species, in particular Andean alder (A. acuminata), grows
fast in a range of restoration sites and soil conditions, but particularly those soils that
are relatively young and low in nitrogen. Our study contributes to a growing body of
literature that native species such as those studied can grow well on degraded sites in
the northern Andes (Murcia 1997; Gunter et al. 2009).
Acknowledgments The authors are grateful for financial support received from the Tropical ResourcesInstitute at the Yale School of Forestry and Environmental Studies and the Gordon and Betty MooreFoundation. Numerous colleagues have assisted with research planning, including Tina Schneider, FlorenciaMontagnini, Eva Garen, Alicia Calle, and Gillian Bloomfield of the Yale School of Forestry and Envi-ronmental Studies and the Environmental Leadership and Training Institute. In Colombia and Ecuador, theauthors are grateful for the assistance of Carolina Murcia of CIFOR, Jose Ignacio Barrera of the UniversidadJaveriana, Orlando Vargas of the Universidad Nacional, Nikolay Aguirre of the Universidad de Loja, dozensmore experts in the fields of conservation and forest restoration, and dozens more project managers,technicians, guides, ranchers, and farmers.
Appendix 1
See Table 4.
New Forests
123
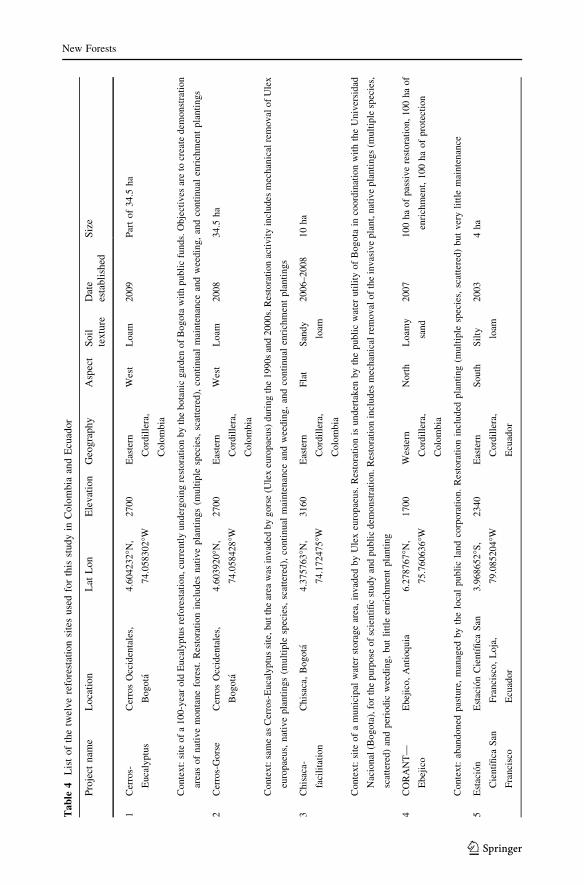
Table
4Listofthetwelvereforestationsitesusedforthisstudyin
Colombia
andEcuador
Project
nam
eLocation
Lat
Lon
Elevation
Geography
Aspect
Soil
texture
Date
established
Size
1Cerros-
Eucalyptus
CerrosOccidentales,
Bogota
4.604232�N
,
74.058302�W
2700
Eastern
Cordillera,
Colombia
West
Loam
2009
Partof34.5
ha
Context:siteofa100-yearold
Eucalyptusreforestation,currentlyundergoingrestorationbythebotanicgarden
ofBogotawithpublicfunds.Objectives
areto
createdem
onstration
areasofnativemontaneforest.Restorationincludes
nativeplantings(m
ultiple
species,scattered),continual
maintenance
andweeding,andcontinual
enrichmentplantings
2Cerros-Gorse
CerrosOccidentales,
Bogota
4.603920�N
,
74.058428�W
2700
Eastern
Cordillera,
Colombia
West
Loam
2008
34.5
ha
Context:sameas
Cerros-Eucalyptussite,butthearea
was
invaded
bygorse(U
lexeuropaeus)duringthe1990sand2000s.Restorationactivityincludes
mechanicalremovalofUlex
europaeus,nativeplantings(m
ultiple
species,scattered),continual
maintenance
andweeding,andcontinual
enrichmentplantings
3Chisaca-
facilitation
Chisaca,
Bogota
4.375763�N
,
74.172475�W
3160
Eastern
Cordillera,
Colombia
Flat
Sandy
loam
2006–2008
10ha
Context:site
ofamunicipal
water
storagearea,invaded
byUlexeuropaeus.Restorationisundertaken
bythepublicwater
utility
ofBogota
incoordinationwiththeUniversidad
Nacional(Bogota),forthepurpose
ofscientificstudyandpublicdem
onstration.Restorationincludes
mechanicalremovaloftheinvasiveplant,nativeplantings(m
ultiplespecies,
scattered)andperiodic
weeding,butlittle
enrichmentplanting
4CORANT—
Ebejico
Ebejico,Antioquia
6.278767�N
,
75.760636�W
1700
Western
Cordillera,
Colombia
North
Loam
y
sand
2007
100haofpassiverestoration,100haof
enrichment,100haofprotection
Context:abandoned
pasture,managed
bythelocalpubliclandcorporation.Restorationincluded
planting(m
ultiple
species,scattered)butverylittle
maintenance
5Estacion
Cientıfica
San
Francisco
EstacionCientıfica
San
Francisco,Loja,
Ecuador
3.968652�S,
79.085204�W
2340
Eastern
Cordillera,
Ecuador
South
Silty loam
2003
4ha
New Forests
123
Table
4continued
Project
nam
eLocation
Lat
Lon
Elevation
Geography
Aspect
Soil
texture
Date
established
Size
Context:restorationispartofan
ongoingscientificexperim
entoperated
betweenalocaluniversity,anationalNGO,anddonors.Plantingsweredonein
rowswithmultiplespecies;
area
isfencedfrom
cowsbutnotweeded
6Pedro
Palolake
LagunaPedro
Palo,Tena,
Cundim
arca
4.685656�N
,
74.386229�W
2180
Central
Cordillera,
Colombia
Flat
Loam
1998
\5ha
Context:restorationobjectiveistheenlargem
entofan
oak
forestoriginally
foundborderingalake.
Project
iscoordinated
byalocalcitizen’s
associationandfunded
byvarious
publicagencies.Restorationincluded
initialplanting(onespecies,row
planting)butlittle
maintenance;area
isfencedto
protect
plantingsfrom
cows
7Pedro
Palo
corridors
LagunaPedro
Palo,Tena,
Cundim
arca
4.685193�N
,
74.377331�W
2020
Central
Cordillera,
Colombia
Flat
Loam
2008
2–5ha
Context:restorationobjectiveis
theestablishmentofsilvo-pastoralsystem
withlivefencesandecological
corridors;project
iscoordinated
byalocalcitizen’s
associationand
funded
byvariouspublicagencies.Restorationincluded
initialplanting(m
ultiple
species,scattered)andenrichmentplantingofmid-successional
speciesbelow
thepioneer
species.Areais
fencedfrom
cowsbutnotweeded
8Profafor
Nono,Pichincha,
Ecuador
0.095844�S,
78.608061�W
2820
Western
Cordillera,
Ecuador
East
Sandy
loam
2006
3ha
Context:project
was
conducted
byanational
NGO
withfundingto
conduct
voluntary
carbonoffsetsforinternational
donors.Areawas
plantedin
rows(m
ultiple
species)
onan
abandoned
pasture,in
landmanaged
byapublicagency.Areais
notmaintained
9ReservaEncenillo
Guasca,Cundim
arca
4.794627�N
,
73.914223�W
3200
Eastern
Cordillera,
Colombia
North
Loam
y
sand
2007
\2ha
Context:landisanature
reserve,owned
byanationalNGO.Therestorationprojectaimsto
restore
asm
allarea
ofthereservethatwas
previouslyin
cattlepasture.Areawas
planted
in2007(onespecies,row
plantings),withinfrequentweeding
New Forests
123
Table
4continued
Project
nam
eLocation
Lat
Lon
Elevation
Geography
Aspect
Soil
texture
Date
established
Size
10
Rio
Bogota
Suesca,Cundim
arca
5.404071�N
,
73.781451�W
2600
Eastern
Cordillera,
Colombia
Flat
Silty clay
loam
mostly
2005–2013
\2haalongriver
corridor
Context:landistheripariancorridor(publicright-of-way)ofasm
allwaterway.Restorationisperform
edbyalocalNGO.Restorationaimsto
preventerosionandrestore
ecosystem
functions.Areais
continually
weeded
andenrichmentplantedwithmultiple
species
11
Rıo
Guacha
Rio
Guachaarea,Boyaca
6.138158�N
,
73.090559�W
2000
Eastern
Cordillera,
Colombia
South
Silty clay
loam
2008
\2halivefences
Context:restorationobjectiveistheestablishmentofasilvo-pastoralsystem
withlivefence
andecologicalcorridorsthroughoutacattlepasture
landscape;restorationisundertaken
byanational
NGO
incoordinationwithlocallandowners(onprivateland).Plantingsaredonein
rows,maintenance
israre
butplantingsarefencedoff
from
cows
12
Zipamine
OIPeldar,Zipaquira,
Cundim
arca
5.067165�N
,
73.867447�W
2760
Eastern
Cordillera,
Colombia
Flat
Sandy
loam
2003
1ha
Context:restorationproject
istherehabilitationofan
abandoned
sandmine;
conducted
byan
environmentalconsultingfirm
foraminingcompany.Restorationinvolves
re-
establishmentofsoil(available
on-site),andplantingofmultiple
species(nativeandexotic),scattered
New Forests
123
Appendix 2
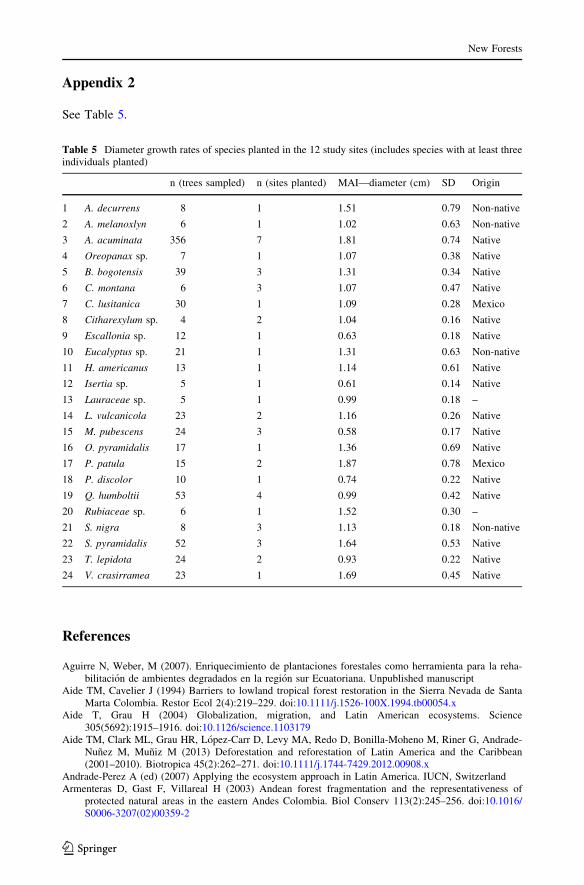
See Table 5.
References
Aguirre N, Weber, M (2007). Enriquecimiento de plantaciones forestales como herramienta para la reha-bilitacion de ambientes degradados en la region sur Ecuatoriana. Unpublished manuscript
Aide TM, Cavelier J (1994) Barriers to lowland tropical forest restoration in the Sierra Nevada de SantaMarta Colombia. Restor Ecol 2(4):219–229. doi:10.1111/j.1526-100X.1994.tb00054.x
Aide T, Grau H (2004) Globalization, migration, and Latin American ecosystems. Science305(5692):1915–1916. doi:10.1126/science.1103179
Aide TM, Clark ML, Grau HR, Lopez-Carr D, Levy MA, Redo D, Bonilla-Moheno M, Riner G, Andrade-Nunez M, Muniz M (2013) Deforestation and reforestation of Latin America and the Caribbean(2001–2010). Biotropica 45(2):262–271. doi:10.1111/j.1744-7429.2012.00908.x
Andrade-Perez A (ed) (2007) Applying the ecosystem approach in Latin America. IUCN, SwitzerlandArmenteras D, Gast F, Villareal H (2003) Andean forest fragmentation and the representativeness of
protected natural areas in the eastern Andes Colombia. Biol Conserv 113(2):245–256. doi:10.1016/S0006-3207(02)00359-2
Table 5 Diameter growth rates of species planted in the 12 study sites (includes species with at least threeindividuals planted)
n (trees sampled) n (sites planted) MAI—diameter (cm) SD Origin
1 A. decurrens 8 1 1.51 0.79 Non-native
2 A. melanoxlyn 6 1 1.02 0.63 Non-native
3 A. acuminata 356 7 1.81 0.74 Native
4 Oreopanax sp. 7 1 1.07 0.38 Native
5 B. bogotensis 39 3 1.31 0.34 Native
6 C. montana 6 3 1.07 0.47 Native
7 C. lusitanica 30 1 1.09 0.28 Mexico
8 Citharexylum sp. 4 2 1.04 0.16 Native
9 Escallonia sp. 12 1 0.63 0.18 Native
10 Eucalyptus sp. 21 1 1.31 0.63 Non-native
11 H. americanus 13 1 1.14 0.61 Native
12 Isertia sp. 5 1 0.61 0.14 Native
13 Lauraceae sp. 5 1 0.99 0.18 –
14 L. vulcanicola 23 2 1.16 0.26 Native
15 M. pubescens 24 3 0.58 0.17 Native
16 O. pyramidalis 17 1 1.36 0.69 Native
17 P. patula 15 2 1.87 0.78 Mexico
18 P. discolor 10 1 0.74 0.22 Native
19 Q. humboltii 53 4 0.99 0.42 Native
20 Rubiaceae sp. 6 1 1.52 0.30 –
21 S. nigra 8 3 1.13 0.18 Non-native
22 S. pyramidalis 52 3 1.64 0.53 Native
23 T. lepidota 24 2 0.93 0.22 Native
24 V. crasirramea 23 1 1.69 0.45 Native
New Forests
123

Ashton PMS, Gunatilleke CVS, Singhakumara BMP, Gunatilleke IAUN (2001) Restoration pathways forrain forest in southwest Sri Lanka: a review of concepts and models. For Ecol Manag 154:409–430.doi:10.1016/S0378-1127(01)00512-6
Asner GP, Rudel TK, Aide TM, Defries R, Emerson R (2009) A contemporary assessment of change inhumid tropical forests. Conserv Biol 23(6):1386–1395. doi:10.1111/j.1523-1739.2009.01333.x
Bakker J, Olivera MM, Hooghiemstra H (2008) Holocene environmental change at the upper forest line innorthern Ecuador. Holocene 18(6):877–893. doi:10.1177/0959683608093525
Bare M (2014) Forest restoration in the tropical Andes: active conservation in a biodiversity Hotspot. TropRes Bull Yale Trop Res Inst. 32–33:93–100
Bautista-Cruz A, del Castillo RF (2005) Soil changes during secondary succession in a tropical montanecloud forest area. Soil Sci Soc Am J 69(3):906–914. doi:10.2136/sssaj2004.0130
Becerra JE (1989) Estructura y crecimiento de un bosque secundario de roble (Quercus humboldtii).Colombia Forestal (Colombia) 3(3):8–22
Becerra A, Zak MR, Horton TR, Micolini J (2005) Ectomycorrhizal and arbuscular mycorrhizal colo-nization of Alnus acuminata from Calilegua National Park (Argentina). Mycorrhiza 15(7):525–531.doi:10.1007/s00572-005-0360-7
Blanco-Garcıa A, Saenz-Romero C, Martorell C, Alvarado-Sosa P, Lindig-Cisneros R (2011) Nurse-plantand mulching effects on three conifer species in a Mexican temperate forest. Ecol Eng 37(6):994–998.doi:10.1016/j.ecoleng.2011.01.012
Brockerhoff EG, Jactel H, Parrotta JA, Quine CP, Sayer J (2008) Plantation forests and biodiversity:oxymoron or opportunity? Biodiver Conserv 17(5):925–951. doi:10.1007/s10531-008-9380-x
Bussmann RW, Wilcke W, Richter M (2008) Landslides as important disturbance regimes—causes andregeneration. In: Beck E et al (eds) Gradients in a tropical mountain ecosystem of Ecuador. Springer,Berlin, pp 319–330
Callaway RM, Walker LR (1997) Competition and facilitation: a synthetic approach to interactions in plantcommunities. Ecology 78(7):1958–1965
Calle A, Montagnini F, Zuluaga AF (2009) Farmers’ perceptions of silvopastoral system promotion inQuindıo Colombia. Bois et forets des tropiques 300(2):79–94
Calvo-Alvarado JC, Arias D, Richter DD (2007) Early growth performance of native and introduced fastgrowing tree species in wet to sub-humid climates of the Southern region of Costa Rica. For EcolManag 242(2):227–235. doi:10.1016/j.foreco.2007.01.034
Carpenter FL, Nichols JD, Sandi E (2004) Early growth of native and exotic trees planted on degradedtropical pasture. For Ecol Manag 196(2):367–378. doi:10.1016/j.foreco.2004.03.030
CESA-Intercooperation Suiza (1992) Programa de reforestacion de areas marginales de la sierra ecuatori-ana. CESA-Intercooperation Suiza, Quito
Chazdon RL (2008) Beyond deforestation: restoring forests and ecosystem services on degraded lands.Science 320(5882):1458–1460
Colombia Censo General (2005). Departamento Administrativo Nacional de Estadıstica: Archivo de Datos.http://formularios.dane.gov.co/Anda_4_1/index.php/catalog/109 Accessed 26 April 2014
Conservation Internacional Colombia (2014). Conservation international http://conservation.org.co/Accessed 6 Feb 2014
Cornelius J, Mesen F, Corea E, Henson M (1996) Variation in growth and form of Alnus acuminata Kunth.grown in Costa Rica. Silvae Genet 45(1):24–30
David MB (1988) Use of loss-on-ignition to assess soil organic carbon in forest soils. Commun Soil Sci Plan19(14):1593–1599
Davidson EA, Reis de Carvalho CJ, Vieira IC, Figueiredo RDO, Moutinho P, Yoko Ishida F, Tuma Saba R(2004) Nitrogen and phosphorus limitation of biomass growth in a tropical secondary forest. Ecol Appl14(4):150–163. doi:10.1890/01-6006
de la Luz Avendano-Yanez M, Sanchez-Velasquez LR, Meave JA, del Rosario Pineda-Lopez M (2015) CanPinus plantations facilitate reintroduction of endangered cloud forest species? Landsc Ecol Eng.doi:10.1007/s11355-015-0277-z
Dinerstein E, Olson DM, Graham DJ, Webster AL, Primm SA, Bookbinder MP, Ledec G (1995) Aconservation assessment of the terrestrial ecoregions of Latin America and the Caribbean. World Bank,Washington, p 129
Ecuador en Cifras Resultados (2015) Instituto Nacional de Estadısticas y Censos. http://ecuadorencifras.gob.ec/resultados/Accessed 10 Sept 2015
Endo M (1994) CAMCORE: twelve years of contribution to reforestation in the Andean region of Colombia.For Ecol Manag 63(2):219–233. doi:10.1016/0378-1127(94)90112-0
Endo M, Mesa GV (1992) Results of a pruning trial with Pinus patula in Colombia. Ipef Int Piracicaba2:45–49
New Forests
123

Etter A, McAlpine C, Wilson K, Phinn S, Possingham H (2006) Regional patterns of agricultural land useand deforestation in Colombia. Agr Ecosyst Environ 114(2):369–386. doi:10.1016/j.agee.2005.11.013
Etter A, McAlpine C, Possingham H (2008) Historical patterns and drivers of landscape change in Colombiasince 1500: a regionalized spatial approach. Ann Assoc Am Geogr 98(1):2–23. doi:10.1080/00045600701733911
Farley KA (2007) Grasslands to tree plantations: forest transition in the Andes of Ecuador. Ann Assoc AmGeogr 97(4):755–771. doi:10.1111/j.1467-8306.2007.00581.x
Gentry AH (1992) Tropical forest biodiversity: distributional patterns and their conservational significance.Oikos. doi:10.2307/3545512
Gilroy JJ, Woodcock P, Edwards FA, Wheeler C, Baptiste BL, Uribe CAM, Haugaasen T, Edwards DP(2014) Cheap carbon and biodiversity co-benefits from forest regeneration in a hotspot of endemism.Nat Clim Change 4(6):503–507. doi:10.1038/nclimate2200
Goldman-Benner RL, Benitez S, Boucher T, Calvache A, Daily G, Kareiva P, Ramos A (2012) Water fundsand payments for ecosystem services: practice learns from theory and theory can learn from practice.Oryx 46(01):55–63. doi:10.1017/S0030605311001050
Gomez-Aparicio L, Zamora R, Gomez JM, Hodar JA, Castro J, Baraza E (2004) Applying plant facilitationto forest restoration: a meta-analysis of the use of shrubs as nurse plants. Ecol Appl 14(4):1128–1138.doi:10.1890/03-5084
Griscom HP, Ashton PMS, Berlyn GP (2005) Seedling survival and growth of native tree species inpastures: implications for dry tropical forest rehabilitation in central Panama. For Ecol Manag218(1):306–318. doi:10.1016/j.foreco.2005.08.026
Grubb PJ (1977) Control of forest growth and distribution on wet tropical mountains: with special referenceto mineral nutrition. Annu Rev Ecol Syst 8:83–107
Guhl A (2008) Coffee production intensification and landscape change in Colombia, 1970–2002. In:Millington A, Jepson W (eds) Land-change science in the tropics: changing agricultural landscapes.Springer, New York, pp 93–116
Gunter S, Gonzalez P, Alvarez G, Aguirre N, Palomeque X, Haubrich F, Weber M (2009) Determinants forsuccessful reforestation of abandoned pastures in the Andes: soil conditions and vegetation cover. ForEcol Manag 258(2):81–91. doi:10.1016/j.foreco.2009.03.042
Haggar J, Wightman K, Fisher R (1997) The potential of plantations to foster woody regeneration within adeforested landscape in lowland Costa Rica. For Ecol Manag 99(1–2):55–64. doi:10.1016/S0378-1127(97)00194-1
Hammen TVD (1974) Pleistocene changes of vegetation and climate in tropical South America. J Biogeogr.doi:10.2307/3038066
Hansen MC, Potapov PV, Moore R, Hancher M, Turubanova SA, Tyukavina A, Townshend JRG (2013)High-resolution global maps of 21st-century forest cover change. Science 342(6160):850–853. doi:10.1126/science.1244693
Henri CJ (2001) Soil-site productivity of Gmelina arborea, Eucalyptus urophylla and Eucalyptus grandisforest plantations in western Venezuela. For Ecol Manag 144(1):255–264. doi:10.1016/S0378-1127(00)00390-X
Holl KD, Loik ME, Lin EH, Samuels IA (2000) Tropical montane forest restoration in Costa Rica: over-coming barriers to dispersal and establishment. Restor Ecol 8(4):339–349. doi:10.1046/j.1526-100x.2000.80049.x
Homeier J, Werner FA, Gradstein SR, Breckle S, Richter M (2008) Potential vegetation and floristiccomposition of Andean forests in South Ecuador, with a focus on the RBSF. In: Beck E et al (eds)Gradients in a tropical mountain ecosystem of Ecuador. Springer, Berlin, pp 87–100
Hooper E, Condit R, Legendre P (2002) Responses of 20 native tree species to reforestation strategies forabandoned farmland in Panama. Ecol Appl 12(6):1626–1641. doi:10.1890/1051-0761
Jacobs DF, Oliet JA, Aronson J, Bolte A, Bullock JM, Donoso PJ, Landhausser SM, Madsen P, Peng S, Rey-Benayas JMR, Weber JC (2015) Restoring forests: what constitutes success in the 21st century? NewFor Online First. doi:10.1007/s11056-015-9513-5
Jha DK, Sharma GD, Mishra RR (1993) Mineral nutrition in the tripartite interaction between Frankia,Glomus and Alnus at different soil phosphorus regimes. New Phytol 123(2):307–311. doi:10.1111/j.1469-8137.1993.tb03740.x
Kattan GH, Alvarez-Lopez H, Giraldo M (1994) Forest fragmentation and bird extinctions: san Antonioeighty years later. Conserv Biol 8(1):138–146. doi:10.1046/j.1523-1739.1994.08010138.x
Knoke T, Calvas B, Aguirre N, Roman-Cuesta RM, Gunter S, Stimm B, Weber M, Mosandl R (2009) Cantropical farmers reconcile subsistence needs with forest conservation? Front Ecol Environ7(10):548–554. doi:10.1890/080131
New Forests
123

Lamb D (2011) Regreening the Bare hills. Tropical forest restoration in the Asia-Pacific region. Worldforests, vol VIII. Springer, Heidelberg
Lamb D, Erskine PD, Parrotta JA (2005) Restoration of degraded tropical forest landscapes. Science310(5754):1628–1632. doi:10.1126/science.1111773
Lehto T, Zwiazek JJ (2011) Ectomycorrhizas and water relations of trees: a review. Mycorrhiza21(2):71–90. doi:10.1007/s00572-010-0348-9
Leon JD, Velez G, Yepes AP (2009) Estructura y composicion florıstica de tres robledales en la region nortede la cordillera central de Colombia. Rev Biol Trop 57(4):1165–1182
Lerner AM, Rudel TK, Schneider LC, McGroddy M, Burbano DV, Mena CF (2014) The spontaneousemergence of silvo-pastoral landscapes in the Ecuadorian Amazon: patterns and processes. RegEnviron Change. doi:10.1007/s10113-014-0699-4
Maron JL, Connors PG (1996) A native nitrogen-fixing shrub facilitates weed invasion. Oecol105(3):302–312. doi:10.1007/BF00328732
McNamara S, Viet Tinh D, Erskine PD, Lamb D, Yates D, Brown S (2006) Rehabilitating degraded forestland in central Vietnam with mixed native species plantings. For Ecol Manag 233(2–3):358–365.doi:10.1016/j.foreco.2006.05.033
Medina M, Orozco H, Diez MC (2008) Establecimiento de un sistema silvopastoril mediante las especiesAlnus acuminata HBK y Acacia decurrens Willd y respuesta al empleo de organismos rizosfericos enSan Pedro (Antioquia). Livest Res Dev 20(1):Article #7
Meza A, Sabogal C, deJong W (2006) Rehabilitacion de areas degradadas en la Amazonia peruana. CIFOR,Indonesia
Millet J, Tran N, Vien Ngoc N, Tran Thi T, Prat D (2013) Enrichment planting of native species forbiodiversity conservation in a logged tree plantation in Vietnam. New For 44:369–383. doi:10.1007/s11056-012-9344-6
Molina M, Medina M, Mahecha L (2008) Microorganismos y micronutrientes en el crecimiento y desarrollodel Aliso (Alnus acuminata H.B.K.) en un sistema silvopastoril alto andino. Livest Res Rural Dev20(4):Article #54
Murcia C (1997) Evaluation of Andean alder as a catalyst for the recovery of tropical cloud forests inColombia. For Ecol Manag 99(1–2):163–170. doi:10.1016/S0378-1127(97)00202-8
Murcia C, Guariguata MR, Andrade A, Andrade GI, Aronson J, Escobar EM, Montes E (2015) Challengesand prospects for scaling-up ecological restoration to meet international commitments: Colombia as acase study. Conserv Lett. doi:10.1111/conl.12199
Murgueitio E, Calle Z, Uribe F, Calle A, Solorio B (2011) Native trees and shrubs for the productiverehabilitation of tropical cattle ranching lands. For Ecol Manag 261(10):1654–1663. doi:10.1016/j.foreco.2010.09.027
Olson DM, Graham DJ, Webster AL, Primm SA, Bookbinder MP, Ledec G (1995) A conservationassessment of the terrestrial ecoregions of Latin America and the Caribbean. World Bank, Washington,p 129
Ospina Arango OL, Vanegas Pinzon S (2010) Restauracion ecologica, rehabilitacion y recuperacion- plannacional de restauracion de ecosistemas. Republica de Colombia Ministerio de Ambiente Vivienda yDesarrollo Territorial, Bogota
Parrotta JA, Turnbull JW, Jones N (1997) Catalyzing native forest regeneration on degraded tropical lands.For Ecol Manag 99(1–2):1–7. doi:10.1016/S0378-1127(97)00190-4
Piotto D (2008) A meta-analysis comparing tree growth in monocultures and mixed plantations. For EcolManag 255(3):781–786. doi:10.1016/j.foreco.2007.09.065
Piotto D, Montagnini F, Ugalde L, Kanninen M (2003) Performance of forest plantations in small andmedium-sized farms in the Atlantic lowlands of Costa Rica. For Ecol Manag 175(1):195–204. doi:10.1016/S0378-1127(02)00127-5
Poorter L (2001) Light-dependent changes in biomass allocation and their importance for growth of rainforest tree species. Funct Ecol 15(1):113–123. doi:10.1046/j.1365-2435.2001.00503.x
Quiceno J, Medina M (2006) La Acacia decurrens Will. fuente potencial de biomasa nutritiva para laganaderıa del tropico de altura. Livest Res Rural Dev 18(12):Article #166
Reyes SP, Rıos OV (eds) (2011) Proceedings from: La restauracion ecologica en la practica: memorias del ICongreso Colombiano de Restauracion Ecologica y II Simposio Nacional de Experiencias en Res-tauracion Ecologica. Universidad Nacional de Colombia, Bogota
Rhoades CC, Eckert GE, Coleman DC (1998) Effect of pasture trees on soil nitrogen and organic matter:implications for tropical montane forest restoration. Restor Ecol 6(3):262–270. doi:10.1046/j.1526-100X.1998.00639.x
New Forests
123

Rıos W, Quiceno J, Gomez F (2004) Validacion y ajuste en sistemas silvopastoriles en la region del ejecafetero, dirigido a productores pequenos y medianos del sistema ganadero doble proposito. COR-POICA, Plan Nacional de Transferencia de Tecnologıa Agropecuaria, Manizales, Colombia, p 140
Rodrigues RR, Gandolfi S, Gustavo Nave A, Aronson J, Egydio Barreto T, Yuri Vidal C, Brancalion PHS(2011) Large-scale ecological restoration of high-diversity tropical forests in SE Brazil. For EcolManag 261:10:1605–1613. doi:10.1016/j.foreco.2010.07.005
Roque RM, Segura EE, Parada EU (2009) Aprovechamiento e industrializacion de arboles de Alnusacuminata Kuntz de plantaciones de rapido crecimiento en Costa Rica. Kuru: Revista Forestal (CostaRica) 6(16):1–11
Rudel TK, Defries R, Asner GP, Laurance WF (2009) Changing drivers of deforestation and new oppor-tunities for conservation. Conserv Biol 23(6):1396–1405. doi:10.1111/j.1523-1739.2009.01332.x
Russo RO (1990) Evaluating Alnus acuminata as a component in agroforestry systems. Agrofor Syst10(3):241–252. doi:10.1007/BF00122914
Saenz L, Mulligan M, Arjona F, Gutierrez T (2014) The role of cloud forest restoration on energy security.Ecosyst Serv 9:180–190. doi:10.1016/j.ecoser.2014.06.012
Sanchez-Cuervo AM, Aide TM, Clark ML, Etter A (2012) Land cover change in Colombia: surprising forestrecovery trends between 2001 and 2010. PLoS One 7(8):e43943. doi:10.1371/journal.pone.0043943
Sarmiento FO (1997) Arrested succession in pastures hinders regeneration of Tropandean forests and shredsmountain landscapes. Environ Conserv 24(01):14–23
Sarmiento FO, Frolich LM (2002) Andean cloud forest tree lines: naturalness, agriculture and the humandimension. Mt Res Dev 22(3):278–287
Schneider T, Ashton MS, Montagnini F, Milan PP (2014) Growth performance of sixty tree species insmallholder reforestation trials on Leyte Philippines. New For 45(1):83–96. doi:10.1007/s11056-013-9393-5
Silver W, Ostertag R, Lugo AE (2000) The potential for carbon sequestration through reforestation ofabandoned tropical agricultural and pasture lands. Restor Ecol 8(4):394–407. doi:10.1046/j.1526-100x.2000.80054.x
Simard SW, Jones MD, Durall DM (2003) Carbon and nutrient fluxes within and between mycorrhizalplants. In: van der Heijden MGA, Sanders IR (eds) Mycorrhizal ecology. Springer, Berlin, pp 33–74
Society for Ecological Restoration (2004) SER International primer on ecological restoration. Society forEcological Restoration, Washington
Stimm B, Beck E, Gunter S, Aguirre N, Cueva E, Mosandl R, Weber M (2008) Reforestation of abandonedpastures: seed ecology of native species and production of indigenous plant material. In: Beck E et al(eds) Gradients in a tropical mountain ecosystem of Ecuador. Springer, Berlin, pp 417–429
Tapia-Armijos MF, Homeier J, Espinosa CI, Leuschner C, de la Cruz M (2015) Deforestation and forestfragmentation in south Ecuador since the 1970s—losing a hotspot of biodiversity. PLoS one10(9):e0133701. doi:10.1371/journal.pone.0133701
Tonneijck FH, Jansen B, Nierop KGJ, Verstraten JM, Sevink J, De Lange L (2010) Towards understandingof carbon stocks and stabilization in volcanic ash soils in natural Andean ecosystems of northernEcuador. Eur J Soil Sci 61(3):392–405. doi:10.1111/j.1365-2389.2010.01241.x
University of Georgia Laboratory for Environmental Analysis. (2015). Laboratory for environmentalanalysis: analysis. Web accessed 19 March 2015
Urgiles N, Lojan P, Aguirre N, Blaschke H, Gunter S, Stimm B, Kottke I (2009) Application of mycorrhizalroots improves growth of tropical tree seedlings in the nursery: a step towards reforestation with nativespecies in the Andes of Ecuador. New For 38(3):229–239. doi:10.1007/s11056-009-9143-x
Vitousek PM, Farrington H (1997) Nutrient limitation and soil development: experimental test of a bio-geochemical theory. Biogeochemistry 37(1):63–75. doi:10.1023/A:1005757218475
Weber M, Gunter S, Aguirre N, Stimm B, Mosandl R (2008) Reforestation of abandoned pastures: silvi-cultural means to accelerate forest recovery and biodiversity. In: Beck E et al (eds) Gradients in atropical mountain ecosystem of Ecuador. Springer, Berlin, pp 431–441
Wilcke W, Oelmann Y, Schmitt A, Valarezo C, Zech W, Homeier J (2008a) Soil properties and tree growthalong an altitudinal transect in Ecuadorian tropical montane forest. J Plant Nutr Soil Sc171(2):220–230. doi:10.1002/jpln.200625210
Wilcke W, Yasin S, Schmitt A, Valarezo C, Zech W (2008b) Soils along the altitudinal transect and incatchments. In: Beck E et al (eds) Gradients in a tropical mountain ecosystem of Ecuador. Springer,Berlin, pp 75–85
Wishnie MH, Dent DH, Mariscal E, Deago J, Cedeno N, Ibarra D, Condit R, Ashton PMS (2007) Initialperformance and reforestation potential of 24 tropical tree species planted across a precipitationgradient in the Republic of Panama. For Ecol Manag 243(1):39–49. doi:10.1016/j.foreco.2007.02.001
New Forests
123

Worbes M (1999) Annual growth rings, rainfall-dependent growth and long-term growth patterns of tropicaltrees from the Caparo Forest Reserve in Venezuela. J Ecol 87(3):391–403. doi:10.1046/j.1365-2745.1999.00361.x
Zangaro W, Nishidate FR, Vandresen J, Andrade G, Nogueira MA (2007) Root mycorrhizal colonizationand plant responsiveness are related to root plasticity, soil fertility and successional status of nativewoody species in southern Brazil. J Trop Ecol 23(1):53–62. doi:10.1017/S0266467406003713
Zehetner F, Miller WP, West LT (2003) Pedogenesis of volcanic ash soils in Andean Ecuador. Soil Sci SocAm J 67(6):1797–1809. doi:10.2136/sssaj2003.1797
New Forests
123





























