Embed Size (px)
Citation preview

[CANCER RESEARCH 46. 5738-5742, November 1986]
Growth-dependent Expression of Human Mr 53,000 Tumor Antigen MessengerRNA in Normal and Neoplastic Cells1
Bruno Calabretta,2 Leszek Kaczmarek, Licia Selleri, Giuseppe Torelli, Pen-Ming L. Ming, Si-Chun Ming, and
W. Edward MercerDepartment of Pathology. Temple University Medical School. Philadelphia. Pennsylvania 19140 [B. C., L. K., P-M. L. M., S-C. M., W. E. M.J, and Centre forExperimental Hematology. Modena School of Medicine, Modena, Italy [L. S., G. T.]
ABSTRACT
We have investigated the expression of M, 53,000 protein (p53) intotal RNA isolated from human peripheral blood mononuclear cellsstimulated by phytohemagglutinin, in serum-stimulated human diploidfibroblasts, and in normal and tumor cells of human epithelial colontissue. We have found that the expression of p53 messenger RNA isgrowth regulated in human cells following kinetics similar to that previously shown in mouse 3T3 cells, and is increased in the large majority ofcolon adenocarcinomas in comparison to adjacent normal mucosa andadenoma. This increased expression of p53 is accompanied by a nearlyproportional increase in the expression of (listone H3. As the expressionof histone H3 is restricted to the S phase of the cell cycle and thereforemeasures the growth fraction of a given population, we suggest that theincreased expression of p53 observed in the large majority of colontumors simply reflects the increased number of cycling cells frequentlyfound in a neoplastic tissue. At variance with these findings a trueoverexpression of p53 was detected in one SV40-transformed humanfibroblasts cell line.
INTRODUCTION
The term p533 is used to identify a family of proteins ranging
in molecular weights from 48,000 to 55,000 that are found inhigh amount in mammalian cells transformed by a variety ofagents such as DNA tumor viruses (1, 2), RNA tumor viruses(3), and chemical agents (2-4). It was also shown that the p53protein level was higher in transformed cell lines derived fromhuman tumors than in nontransformed cell lines (5). Morerecent studies have suggested that p53 may play a direct role inthe process of neoplastic transformation (6-8). Of particularimportance, in this regard, is the observation that cotransfectionof a gene encoding the mouse p53 with an activated human rasgene yields transformed foci of primary embryonic cells (6-8).
A second direction of research on p53 function deals withthe role that this protein might have in the control of normalcell proliferation. It has been shown that an increase of p53protein and mRNA precedes the synthesis of DNA in mouse3T3 fibroblasts moving from a growth-arrested state to anactively dividing state (9), and that there is an increase of p53protein in PHA-stimulated human peripheral blood T-lympho-cytes (10), and Con A-stimulated mouse splenocytes (11).
In three significant functional studies it was shown thatmicroinjection of monoclonal antibodies against p53 proteininhibited the entry into S phase of serum-stimulated Swiss 3T3fibroblasts (12, 13), and that microinjection of active p53 encoding sequences stimulated DNA synthesis in quiescent mouse3T3 cells (14).
Received 2/17/86; revised 7/17/86; accepted 7/21/86.The costs of publication of this article were defrayed in part by the payment
of page charges. This article must therefore be hereby marked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
1This work was supported by Grant BRSG S07-RR05417 to B. C. and W. E.M., and a grant from the Leukemia Research Foundation. Chicago. IL. to B. C.
2To whom requests for reprints should be addressed.3The abbreviations used are: p53, M, 53,000 protein; PHA, phytohemagglu
tinin; PBM cells, peripheral blood mononuclear cells.
These two distinct and yet intermingling facets of p53 (rolein normal cell proliferation and involvement in neoplastic transformation), made this gene a valuable molecular tool to investigate the impairment of proliferative control that is a key factorin the origin and the progression of human neoplasia (15). Alegitimate theory of neoplastic transformation is that an increased activity of some key genes is actively involved in transforming a normal cell into a neoplastic one (16). Therefore, wehave chosen to compare the expression of p53 in normal andin adjacent tumorous colon tissue of the same patients.
Normally, cell division in the colonie mucosa is restricted tothe deeper two-thirds of the crypt. Neoplastic transformationis accompanied by the loss of such restriction in cell growth.This change is visualized by the ability of the cells along theentire length of the crypt, including the surface cells, to synthesize DNA and divide (17).
We reasoned that the correct assessment of the level ofexpression of a growth-regulated gene like p53 in normal andtumor tissues requires the evaluation of the growth fraction ofboth cell populations. The expression of histone H3 gene, likethe other core histones, is finely regulated (18,19) and is limitedto the S phase of the cell cycle. Therefore, it is a good indicatorof the growth fraction (20) of different cell populations.
By comparing the expression of a growth-regulated gene toa S-phase gene and by measuring this ratio in normal and tumorcells, we can determine whether a growth-regulated gene is trulyoverexpressed in neoplastic cells.
Before studying the expression of p53 in human colon neoplasia, we wanted to be sure that the mRNA level of p53 istruly growth regulated in human cells. For this purpose humanperipheral blood mononuclear cells and diploid fibroblast cellstrains were used. In addition, cell lines derived by transformation with SV40 virus were included for comparison in thestudies reported here.
MATERIALS AND METHODS
Cells and Culture Conditions. The human diploid fibroblast cell strainWI-38 and an SV40-transformed cell line VA13-2RA derived from WI-38 were both kindly provided by Dr. R. Baserga (Temple UniversitySchool of Medicine). The human diploid cell strains IMR-90 andGM1142, and SV40-transformed derivatives SV1MR-90 andSVGM1142 were kindly provided by Dr. W. Nichols (Merck, Sharpand Dohme). They were grown in Eagle's minimum essential medium
with vitamins, antibodies, in the presence of 10% (v/v) fetal calf serum.Peripheral blood mononuclear cells were isolated by Ficoll-Hypaquegradients from venous blood collected from healthy volunteers, andcultured as described by Maizel et al. (21). The cells were washed withHanks' solution and then cultured in plastic Falcon tissue culturesflasks for the desired periods of time at a concentration of 10' cells/mlof RPMI 1640 supplemented with penicillin, streptomycin, L-gluta-iiiinc. 5% heat-inactivated fetal calf serum, and PHA (Sigma ChemicalCo.) at 10 Mg/ml- Each culture was monitored for the level of stimulation by determining autoradiographically the percentage of cells thatincorporate [3H]thymidine during 66-72 h in culture. After culture,
cells were harvested by centrifugation, washed twice in 0.9% NaCI
5738
on June 5, 2018. © 1986 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

EXPRESSION OF HUMAN p53 TUMOR ANTIGEN mRNA
solution, and total cellular RNA was then extracted. Human fibroblastsWI-38 (22) were cultured in plastic Petri dishes in 10% fetal calf serumfor 14 days until the cells become quiescent and were then used for theexperiments. Fibroblasts were either stimulated with fresh medium for1,6, 16, and 24 h, or were replaced at dilution 1:20 and after 3 dayssynchronized in S phase with 2.5 IHM hydroxyurea. After overnightexposure to hydroxyurea the drug was removed by extensive washingand cells were next harvested 4, 9, 3, 18, and 23 h later. Totalcytoplasmic RNA was extracted (23) from quiescent as well as fromstimulated and hydroxyurea block-released cells.
Tissue Samples for RNA Extraction. Eleven colon specimens wereobtained immediately after surgical resection and processed on ice.They were opened, and the contents were rinsed off with cold phosphate-buffered saline. Tissue samples were taken from the tumor aswell as from the normal areas. Ulcerated and necrotic tissue wasdissected off the tumor tissue, and the submucosa and muscularis wereremoved from normal tissue. Thus only the intact tumor and normalmucosa were used for RNA extraction. Samples were taken from 11carcinomas, 4 adenomas, and 11 normal colons. They were immediatelystored in liquid nitrogen until use in about 1 week.
Tissue Samples for Microscopy. Whole thickness tissue slices weretaken from the tumor as well as from normal areas for microscopy.They were fixed in buffered formalin, embedded in paraffin, and routinely stained with hematoxylin and eosin for histológica!diagnosis.
RNA Isolation. Total cellular RNA was purified from culture cellsand colon tissue according to Frazier et al. (24). Briefly, the tissue washomogenized in a Waring blender in the extraction buffer [75 mMNaCl; 20 mM EDTA; 10 mM Tris-HCl (pH 8.0); 0.2% sodium dodecylsulfate] mixed 1:1 with buffer-saturated phenol. The aqueous phase wasrecovered by centrifugation, reextracted with an equal volume of phenoland chloroform:isoamyl alcohol (25:24:1), and once again with chlo-roform:isoamyl alcohol (24:1). The nucleic acids were precipitated withethanol, and DNA was removed by treatment with DNase I andprecipitation with 3 M sodium acetate (pH 5.5). The integrity and thecorrect amount of the electrophoresed RNA samples were monitoredby ethidium bromide staining of agarose formaldehyde gels.
Northern Blot Analysis. Total cellular RNA was denatured with 6.3%formaldehyde and 50% formamide and then size fractioned on a 1.2%agarose gel containing 6.6% formaldehyde. Blotting of RNA to nitrocellulose was done as described by Thomas (25). Nick translation (26,27) of the plasmid DNA at high specific activity was performed asdescribed by Lai et al. (28). Prehybridization, hybridization, and post-hybridization washes were essential as described by Wahl et al. (29).Filters were exposed to Kodak X-ray films using intensifying screen at-70°C. Densitometer readings of the films were performed with the
aid a Zeineh soft laser densitomer (Biomed Instruments, Inc., Fullerton,CA).
Plasmids. Plasmids carrying the gene probes used in these studieswere: php53c-l kindly provided by Dr. D. Givol (The WeizmannInstitute of Science) containing a full-length human p53 cDNA described by Zakut-Houri et al. (30), and pFo 422 carrying a histone H3gene (kind gift of Dr. G. Stein, University of Florida).
RESULTS
Growth-dependent Expression of p53 RNA in Human Cells.Previous work from our laboratory has shown that the p53protein recognized by monoclonal antibody PAbl22 is notdetectable in resting (G0) human peripheral blood mononuclearcells; however, following PHA stimulation it is detectable at18-24 h and reaches a maximum level of synthesis between 42and 48 h after stimulation (10). We wanted to establish thatthe increased p53 protein synthesis observed in PHA-stimu-lated PBM cells is paralleled by an increase in the steady-statelevels of p53 mRNA. Fig. \A shows a Northern blot of RNAisolated from PBM cells at different times after PHA stimulation, hybridized with a human p53 cDNA probe. The resultspresented in Fig. \A show that p53 mRNA levels increase at18 h (Fig. IA, Lane b), reach the highest level at 40 h (Fig. IA,
abed
Babode f g h ¡
p 53
H3
Fig. 1. A, expression of p53 in PHA-stimulated human T-lymphocytes. TotalRNA was isolated, and 15 /ig/lane were electrophoresed on a 1.2% agarose-formaldehyde gel and subsequently transferred to nitrocellulose as described byThomas (25). Hybridization was carried out with the php53c-l nick translated toa specific activity of 4-5 x 10* cpm/fjg. The size of p53 RNA band is approxi
mately 3.0 kilobases. The sources of the RNAs in the various lanes are as follows:Lanes a, b. c. and d, human peripheral blood mononuclear cells cultures stimulatedwith PHA for 0 h in Lane a, 18 h in Lane b, 40 h in Lane c, and 66 h in Lane d.B, cell cycle-dependent expression of p53 gene in human diploid fibroblasts.Human diploid fibroblasts (WI-38) were plated at the density 1 x 104/cm2 ofplastic Petri dishes in Earle's minimal essential medium supplemented with 10%
fetal calf serum, amino acids, vitamins, and antibiotics as described (22). After14 days in culture cells were quiescent and used for the experiments. Fibroblastswere either stimulated with fresh medium for 1,6, 16. and 24 h or replated atdilution of 1:20 and, after 3 days, synchronized at the G,-S-phase boundary with2.5 mM hydroxyurea. After overnight exposure to hydroxyurea the drug wasremoved by extensive washing and cells were harvested at 4, 9, 13, 18, and 23 hlater. Total cytoplasmic RNA was extracted (30) from quiescent as well as fromstimulated and hydroxyurea block-released cells. RNA (15 ><g/lane) was nextseparated on formaldehyde-containing agarose gels and transferred to nitrocellulose filter as described by Thomas (25). The filter was next consecutivelyhybridized to nick-translated histone H3 and p53 gene probes. Lane a representshybridization to RNA from quiescent W138 fibroblasts; Lanes b. c. d, and erepresent hybridization to RNA obtained from cells stimulated with fresh mediumfor (respectively) 1,6, 16, and 24 h; Lanes f, g. h, i, andj represent hybridizationto RNAs extracted from hydroxyurea-synchronized cells 4, 9, 13, 18, and 23 h(respectively) after release from the GrS-phase block.
Lane c), and remain at essentially the same high level until 66h (Fig. \A, Lane d) following PHA stimulation. Under ourculture conditions, 60% of the cells enter the S phase between40 and 66 h of PHA stimulation. Therefore, the highest levelof p53 RNA (and protein) expression in normal PBM cellsoccurs when the majority of the cells are in late G, or S phase.
We also examined the expression of p53 RNA in synchronized cultures of the human diploid fibroblasts cell strain, WI-38 (22). Histone H3 was used as a control probe because theexpression of this gene is preferentially limited to the S phaseof the cell cycle. In this study, we wanted to compare directlythe steady-state level of p53 RNA with the level of histone H3RNA in serum-stimulated fibroblasts during the G0-Gi-S-phasetransition, and also during the S-G2-M-phase transition. Forthis purpose WI-38 fibroblasts were first made quiescent byserum deprivation and subsequently were stimulated with serumto reenter the cell cycle, or exponentially growing cells werefirst blocked at the Gi-S-phase boundary by hydroxyurea treatment and subsequently were released from the block by removing the hydroxyurea-containing medium and replacing it withfresh medium.
5739
on June 5, 2018. © 1986 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

EXPRESSION OF HUMAN p53 TUMOR ANTIGEN mRNA
Fig. l B shows the levels of p53 mRNA expression in serum-stimulated WI-38 cells (Fig. 1B, Lanes a-e) and in cells releasedfrom the Gi-S-phase block (Fig. IB, Lanes b-j). The resultsshow that p53 mRNA levels increase very early after serumaddition in WI-38 cells and continue to increase, reaching thehighest level of expression after 16-24 h. The kinetics of p53expression during the G0-G|-S-phase transition in serum-stimulated fibroblasts is similar to that of PHA-stimulated lymphocytes with the highest levels occurring in both cell types in lateG,-early S phase. In WI-38 cells released from a d-S-phasehydroxyurea block during the S-G2-M transition p53 RNAexpression remains relatively constant (Fig. IB, Lanesf-j) andis considerably higher than in quiescent (G0) cells (Fig. \B,Lane a). The expression of histone H3 mRNA correspondswell with the entry into S phase of the cells following serumstimulation from quiescence (Fig. Iß,Lane e) and in cellsreleased from the Gi-S-phase hydroxyurea block (Fig. Iß,Lanes/and y'). It is noteworthy to mention that in the latter system,
at 4 h after hydroxyurea release (Fig. IB, Lane f), a largeproportion of the cells are still in S phase and express histoneH3 mRNA. Moreover, a second peak of histone H3 mRNAexpression occurs at 23 h after hydroxyurea release (Fig. IB,Lane j), which corresponds to the S phase of the second cellcycle.
Expression of p53 and Histone H3 RNA in Normal and SV40-transformed Fibroblasts. We next ask whether the expressionof p53 RNA is altered in SV40-transformed established humanfibroblast cell lines. The human diploid fibroblasts strains (WI-38, IMR-90, and GM1142) and their SV40-transformed counterparts (VA13-2RA, SVIMR-90, and SVGM1142) were examined and the level of p53 mRNA expression was comparedto the levels of histone H3 mRNA expression. Because theexpression of histone H3 is limited to the S phase of the cellcycle (18, 19), this approach allowed us to evaluate the dependence of the levels of expression of a growth-regulated gene (p53)on the growth fraction of each matched set of cells analyzed.
Fig. 2, left, shows a Northern blot of RNA from normalIMR-90 and GM1142 cell strains and their SV40-transformed
ab cd
a b
-H3
Fig. 2. Left, expression of p53 and H3 in exponentially growing IMR-90 andGM1142 fibroblasts and their SV40-transformed cell lines. Blot hybridization ofRNA was made as described in Fig. \.A and B. The filter was first hybridized tothe H3 probe (pFo 422) and subsequently to the p53 probe (php53c-l). The H3mRNA size is 0.5 kilobase; the p53 mRNA size is 3.0 kilobases. The sources ofthe RNAs in the various lanes are as follows: Lane a. IMR-90; Lane b, SVIMR-90; Lane c, GM 1142; Lane d, SVGM1142. Right, expression of p53 and H3 inexponentially growing WI-38 and VA13-2RA. Blot hybridization of RNA was asdescribed in Fig. 2, left. Lane a. WI-38: Lane b, VA13-2RA.
established cell lines SVIMR-90 and SVGM 1142, hybridizedfirst with histone H3 and next with the p53 probes. We measured the levels of hybridization by soft laser densitometry ofthe autoradiograph of this blot and found very little, if any,difference in the levels of p53 RNA expression relative tohistone H3 expression in these normal and SV40-transformedcells. Fig. 2, right, is a composite picture of a Northern blot ofRNA from the normal cell strain WI-38 and its SV40-trans-formed derivative VA13-2RA. It is rather obvious that there isa modest difference in the intensity of the H3 band betweenWI38 and VA13-2RA cells; whereas, the difference in intensityof the p53 band is much more pronounced. We found that thereis at most a 2-fold difference for H3, but more than a 10-fold
difference for p53. Therefore, the increase in the level of p53mRNA expression in VA13-2RA in comparison to the parentalcell strain WI38 is not merely a reflection of the difference inthe growth fraction.
Expression of p53 in Normal and Neoplastic Colon Mucosa.We next decided to extend the same type of analysis to spontaneously occurring human neoplasia. We have selected theadenocarcinoma of the colon because this neoplasm is rathercommon in Western countries and exhibits a pattern of growththat is a clear subversion of the regular pattern found in thenormal mucosa. The specimens from which total cellular RNAwas isolated were examined macroscopically and microscopically. The histológica! diagnosis and the location of the lesionsare listed in Table 1. The normal mucosa had a similar histologyin all cases. Tubulovillous adenoma was present in 4 cases andin 2 it was the seat of cancer origin (cases 1 and 7). Thecarcinoma was moderately differentiated in 8 cases, poorlydifferentiated in 2, well differentiated in one (Table 1). Asdescribed below, the degree of tumor differentiation had noapparent effect on the degree of expression of the tested genes.
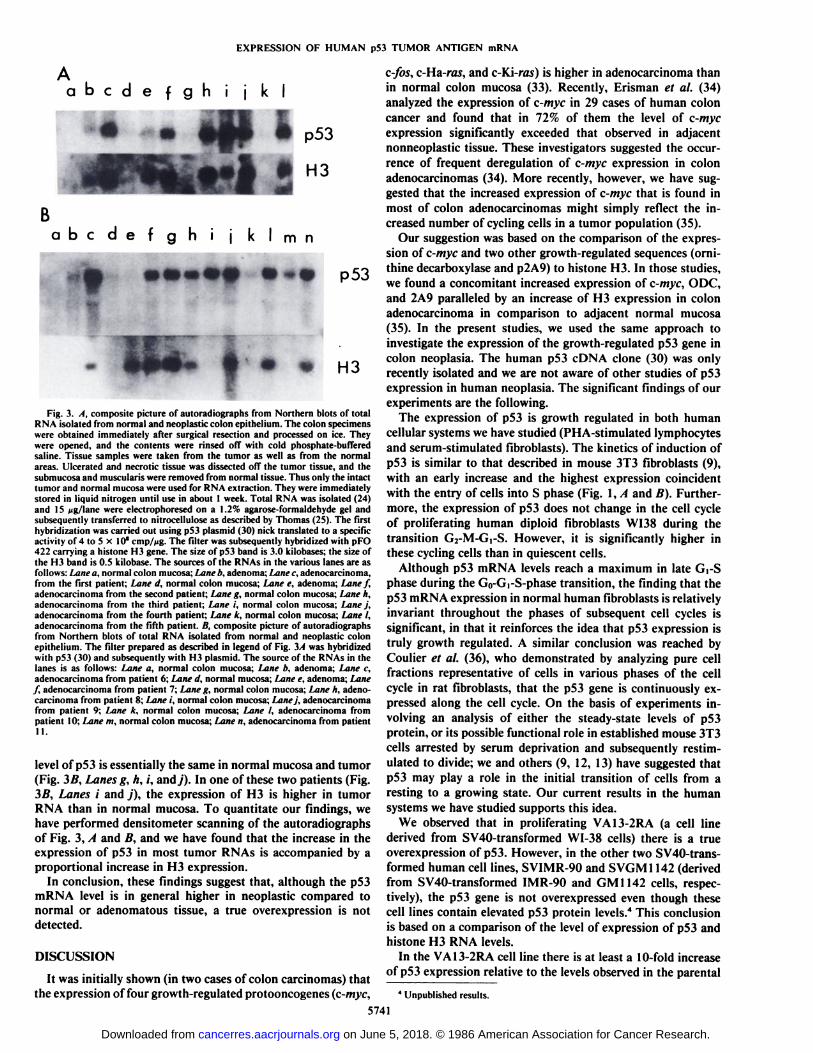
Fig. 3/1 is a composite picture of autoradiograms of a filterhybridized first to p53 and subsequently to histone H3. Theincreased expression of p53 found in most cases in tumor RNA(Fig. 3/1, Lanes c,f, h, and /) is paralleled by an increase in themRNA level of histone H3. In Fig. 3/1, Lanes i and j, corresponding to normal and neoplastic tissue of patient 4 theexpression of p53 is essentially equal, although in the tumorRNA there is a higher H3 expression.
Fig. 3Äshows the same type of analysis extended to sixadditional patients. Once again, the filter was hybridized to p53and next to histone H3. It is evident that the increased expression of p53 found in most cases in tumor RNAs (Fig. 3A, Lanesc,f, I, and n) is paralleled by an increase in the mRNA level ofhistone H3. It is noteworthy that at variance with the prevalentpattern of p53 expression, there are two patients in whom the
Table 1 Location and histological diagnosis of colonie neoplasms
Case1°23456T891011LocationCecumAscending
colonSigmoid
colonAscendingcolonSigmoidcolonAscendingcolonCecumCecumSigmoid
colonAscendingcolonSigmoid
colonHistológica!
diagnosisModerately
differentiatedadenocarcinoma.tubulovillousadenomaWell
differentiated adenocarcinoma, tubulovillousadenomaPoorly
differentiatedadenocarcinomaModeratelydifferentiatedadenocarcinomaModeratelydifferentiatedadenocarcinomaModeratelydifferentiatedadenocarcinoma.tubulovillous
adenomaModeratelydifferentiatedadenocarcinoma.tubulovillous
adenomaModeratelydifferentiatedadenocarcinomaModeratelydifferentiatedadenocarcinomaModeratelydifferentiatedadenocarcinomaPoorly
differentiated adenocarcinoma' In cases 1 and 7 the carcinomas arose in the tubulovillous adenomas.
5740
on June 5, 2018. © 1986 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
EXPRESSION OF HUMAN p53 TUMOR ANTIGEN mRNA
«
abcdefghijkl
Babcdefqhi ¡ kl m n
p53
H3
Fig. 3. A, composite picture of autoradiographs from Northern blots of totalRNA isolated from normal and neoplastic colon epithelium. The colon specimenswere obtained immediately after surgical resection and processed on ice. Theywere opened, and the contents were rinsed off with cold phosphate-bufferedsaline. Tissue samples were taken from the tumor as well as from the normalareas. Ulcerated and necrotic tissue was dissected off the tumor tissue, and thesubmucosa and muscularis were removed from normal tissue. Thus only the intacttumor and normal mucosa were used for RNA extraction. They were immediatelystored in liquid nitrogen until use in about 1 week. Total RNA was isolated (24)and IS tig/lane were electrophoresed on a 1.2% agarose-formaldehyde gel andsubsequently transferred to nitrocellulose as described by Thomas (25). The firsthybridization was carried out using pS3 plasmid (30) nick translated to a specificactivity of 4 to 5 x 10*cmp//ig. The filter was subsequently hybridized with pFO
422 carrying a histone H3 gene. The size of p53 band is 3.0 kilobases; the size ofthe H3 band is 0.5 kilobase. The sources of the RNAs in the various lanes are asfollows: I.itni' a, normal colon mucosa; Lane b, adenoma; I iinf c, adenocarcinoma,from the first patient; Lane d, normal colon mucosa; Lane e, adenoma; Lane f,adenocarcinoma from the second patient; Lane g, normal colon mucosa; Lane h,adenocarcinoma from the third patient; Lane i, normal colon mucosa; Lane j,adenocarcinoma from the fourth patient; Lane k, normal colon mucosa; Lane I,adenocarcinoma from the fifth patient. /i. composite picture of autoradiographsfrom Northern blots of total RNA isolated from normal and neoplastic colonepithelium. The filter prepared as described in legend of Fig. 3A was hybridizedwith p53 (30) and subsequently with H3 plasmid. The source of the RNAs in thelanes is as follows: Lane a, normal colon mucosa; Lane b, adenoma; Lane c,adenocarcinoma from patient 6; Lane d, normal mucosa; Lane e, adenoma; Lane/, adenocarcinoma from patient 7; Lane g, normal colon mucosa; Lane h, adenocarcinoma from patient 8; Lane i, normal colon mucosa; Lanej, adenocarcinomafrom patient 9; Lane k, normal colon mucosa; Lane I, adenocarcinoma frompatient 10; Lane m, normal colon mucosa; Lane n, adenocarcinoma from patientII.
level of p53 is essentially the same in normal mucosa and tumor(Fig. 3B, Lanes g, h, i, andy). In one of these two patients (Fig.3B, Lanes i and j), the expression of H3 is higher in tumorRNA than in normal mucosa. To quantitate our findings, wehave performed densitometer scanning of the autoradiographsof Fig. 3, A and B, and we have found that the increase in theexpression of p53 in most tumor RNAs is accompanied by aproportional increase in H3 expression.
In conclusion, these findings suggest that, although the p53mRNA level is in general higher in neoplastic compared tonormal or adenomatous tissue, a true overexpression is notdetected.
DISCUSSION
It was initially shown (in two cases of colon carcinomas) thatthe expression of four growth-regulated protooncogenes (c-myc,
c-fos, c-Ha-ras, and c-Ki-ras) is higher in adenocarcinoma than
in normal colon mucosa (33). Recently, Erisman et al. (34)analyzed the expression of c-myc in 29 cases of human coloncancer and found that in 72% of them the level of c-mycexpression significantly exceeded that observed in adjacentnonneoplastic tissue. These investigators suggested the occurrence of frequent deregulation of c-myc expression in colon
adenocarcinomas (34). More recently, however, we have suggested that the increased expression of c-myc that is found in
most of colon adenocarcinomas might simply reflect the increased number of cycling cells in a tumor population (35).
Our suggestion was based on the comparison of the expression of c-myc and two other growth-regulated sequences (orni-
thine decarboxylase and p2A9) to histone H3. In those studies,we found a concomitant increased expression of c-myc, ODC,and 2A9 paralleled by an increase of H3 expression in colonadenocarcinoma in comparison to adjacent normal mucosa(35). In the present studies, we used the same approach toinvestigate the expression of the growth-regulated p53 gene incolon neoplasia. The human p53 cDNA clone (30) was onlyrecently isolated and we are not aware of other studies of p53expression in human neoplasia. The significant findings of ourexperiments are the following.
The expression of p53 is growth regulated in both humancellular systems we have studied (PHA-stimulated lymphocytesand serum-stimulated fibroblasts). The kinetics of induction ofp53 is similar to that described in mouse 3T3 fibroblasts (9),with an early increase and the highest expression coincidentwith the entry of cells into S phase (Fig. I, A and B). Furthermore, the expression of p53 does not change in the cell cycleof proliferating human diploid fibroblasts WI38 during thetransition G2-M-Gi-S. However, it is significantly higher inthese cycling cells than in quiescent cells.
Although p53 mRNA levels reach a maximum in late d-Sphase during the G0-Gi-S-phase transition, the finding that the
p53 mRNA expression in normal human fibroblasts is relativelyinvariant throughout the phases of subsequent cell cycles issignificant, in that it reinforces the idea that p53 expression istruly growth regulated. A similar conclusion was reached byCoulier et al. (36), who demonstrated by analyzing pure cellfractions representative of cells in various phases of the cellcycle in rat fibroblasts, that the p53 gene is continuously expressed along the cell cycle. On the basis of experiments involving an analysis of either the steady-state levels of p53protein, or its possible functional role in established mouse 3T3cells arrested by serum deprivation and subsequently restim-ulated to divide; we and others (9, 12, 13) have suggested thatp53 may play a role in the initial transition of cells from aresting to a growing state. Our current results in the humansystems we have studied supports this idea.
We observed that in proliferating VA13-2RA (a cell linederived from SV40-transformed WI-38 cells) there is a trueoverexpression of p53. However, in the other two SV40-trans-formed human cell lines, SVIMR-90 and SVGM1142 (derivedfrom SV40-transformed IMR-90 and GM1142 cells, respectively), the p53 gene is not overexpressed even though thesecell lines contain elevated p53 protein levels.4 This conclusion
is based on a comparison of the level of expression of p53 andhistone H3 RNA levels.
In the VA13-2RA cell line there is at least a 10-fold increase
of p53 expression relative to the levels observed in the parental
4 Unpublished results.
5741
on June 5, 2018. © 1986 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

EXPRESSION OF HUMAN p53 TUMOR ANTIGEN mRNA
WI38 cell strain; whereas the expression of histone H3 is atmost 2-fold higher (Fig. 2, right).
Northern blot analysis on mRNA isolated from a variety ofhuman tumor cell lines shows that they all contain p53 mRNA;but that p53 protein levels in these cell lines varies considerably,yet this is not reflected at the mRNA level so that the amountof p53 protein in these cell lines must be posttranscriptionallyregulated (37). Thus, the functional significance of the over-expression of p53 mRNA in VA13-2RA cells as it relates totransformation and immortalization of these cells is unclear. Itis however, noteworthy to mention that the clear-cut over-expression of p53 RNA in VA13-2RA is not associated with adetectable rearrangement or gene amplification of p53 locus(data not shown).
In the large majority of colon adenocarcinomas we haveanalyzed, the expression of p53 is increased in comparison tonormal or adenomatous mucosa. This increased expression islikely to reflect the fact that a higher number of cycling cells isfound in most colon tumorous tissue than in the normal oradenomatous mucosa. This is strongly suggested by the proportional increased expression of histone H3. The comparisonof c-myc, ornithine decarboxylase, 2A9 (35), and p53 (ourcurrent study) RNA expression relative to the expression ofhistone H3 in adjacent normal and neoplastic tissue alerts oneto the fact that the growth fraction of cells in a given populationmust be taken into account when evaluating the "overexpres-sion" of a given oncogene, especially when the gene under studyis expressed in a growth-regulated fashion.
ACKNOWLEDGMENTS
We wish to thank Renato Baserga for his guidance and criticalreading of the manuscript. We thank Drs. D. Givol and G. Stein forproviding us with p53 and H3 probes, respectively.
ADDENDUM
While this work was in progress, Reed et al. (Proc. Nati. Acad. Sci.USA, 83: 3982, 1986) reported similar kinetics of p53 mRNA expression in PHA-stimulated human peripheral blood lymphocytes.
REFERENCES
1. Lane, D. P., and Crawford, L. V. T-antigen is bound to a host protein inSV40 transformed cells. Nature (Lond.), 278: 261-263, 1979.
2. Linzer, D. I. H., and Levine, A. J. Characterization of a 54k dalton cellularSV40 tumor antigen present in SV40 transformed cells and uninfectedembryonal carcinoma cells. Cell, 17: 43-52, 1979.
3. Rotter. V.. Boss, M. A., and Baltimore, D. Increased concentration of anapparently identical cellular protein in cells transformed by either Abelsonmurine leukemia virus or other transforming agents. J. Viro!., 38: 336-346,1981.
4. DeLeo, A. B., Gilbert. J, Appella, E., DuBois, G. C, Law, L. W., and Old,L. J. Detection of a transformation related antigen in chemically inducedsarcomas and other transformed cells of the mouse. Proc. Nati. Acad. Sci.USA. 76: 2420-2424. 1979.
5. Crawford, L. V., Pirn. D. C, Gurney, E. G., Goodfellow. P.. and Taylor-Papadimitrous, J. Detection of a common feature in several human tumorcell lines—a 53,000-dalton protein. Proc. Nati. Acad. Sci. USA, 78: 41-45,1982.
6. Eliyahu. D., Raz, A., Gruss, P., Givol, D.. and Oren, M. Participation of p53cellular tumor antigen in transformation of normal embryonic cells. Nature(Lond.), 312: 646-649. 1984.
7. Parada, L. F.. Land. H., Weinberg, R. A., Wolf. D.. and Rotter, V. Cooperation between gene coding p53 tumor antigen and ras in cellular transformation. Nature (Lond.), 312:649-651, 1984.
8. Jenkins. J. R.. Rudge, K., and Currie, G. A. Cellular immortalization by acDNA clone encoding the transformation associated phosphoprotein p53.Nature (Lond.). 312: 651-659, 1984.
9. Reich, N. C., and Levine, A. J. Growth regulation of cellular antigen p53 innontransformed cells. Nature (Lond.), 308: 199-201, 1984.
10. Mercer, W. E., and Baserga, R. Expression of the p53 protein during the cellcycle of human peripheral blood lymphocytes. Exp. Cell Res., 100: 31-46,1985.
11. Milner, J., and Milner, S. SV40-53k antigen: a possible role of 53k in normalcells. Virology, 112: 785-788, 1981.
12. Mercer, W. E., Nelson, D., DeLeo, A. B., Old, L. J., and Baserga, R.Microinjection of monoclonal antibodies to protein p53 inhibits seruminduced DNA synthesis in 3T3 cells. Proc. Nati. Acad. Sci. USA, 79:6309-6312, 1982.
13. Mercer, W., Avignolo, C., and Baserga, R. Role of the p53 protein in cellproliferation as studied by microinjection of monoclonal antibodies. Mol.Cell. Biol., 4: 276-284. 1984.
14. Kaczmarek, L., Oren, M., and Baserga, R. Co-operation between the p53protein tumor antigen and platelet-poor plasma in the induction of cellularDNA synthesis. Exp. Cell Res., 162: 268-272, 1986.
15. Nowell, P. C. The clouai evolution of tumor cell populations. Science (Wash.DC), / 74: 23-30, 1976.
16. Klein, G. The role of gene dosage and genetic transposition in carcinogenesis.Nature (Lond.). 294: 313-318, 1981.
17. Lipkin, M. Phase 1 and phase 2 proliferative lesions of colonie epithelialcells in diseases leading to colonie cancer. Cancer (Phila.), 34: 878-888.1974.
18. Plumb, M., Stein, J., and Stein, G. Coordinate regulation of multiple histonemRNAs during the cell cycle in HeLa cells. Nucleic Acids Res., //: 2301-2410, 1983.
19. Hirschhorn, R. R., Marashi, F., Baserga, R., Stein, J., and Stein, G. Expression of histone genes in a G, specific temperature sensitive mutant of thecell-cycle. Biochemistry, 23: 3731-3736, 1984.
20. Baserga, R. The cell-cycle. N. Engl. J. Med., 304:453-459, 1984.21. Maizel, A. L., Mehta, S. R., Häuft,S., Franzini, D., Lachman, L. B., and
Ford, R. J. Human T lymphocyte/monocyte interaction in response to lectin:kinetics of entry into the S-phase. J. Innmmol., 127: 1058-1064, 1981.
22. Bombik. B. M., and Baserga, R. Increased RNA synthesis in nuclear mono-layers of WI38 cells stimulated to proliferate. Proc. Nati. Acad. Sci. USA,71: 2038-2042. 1974.
23. Hirschhorn, R. R., Aller, P., Yuan, Z-A., Gibson, C. W., and Baserga, R.Cell-cycle specific cDNAs from mammalian 1984 cells temperature sensitivefor growth. Proc. Nati. Acad. Sci. USA, 81: 6004-6008, 1984.
24. Frazier, M. L., Mars, W., Fiorine. L. D., Montagna, R. A., and Saunders,G. F. Efficience extraction of RNA from mammalian tissue. Mol. Cell.Biochem., 56: 113-122, 1983.
25. Thomas. P. S. Hybridization of denatured RNA and small DNA fragmentstransferred to nitrocellulose. Proc. Nati. Acad. Sci. USA, 77: 5201-5205,1980.
26. Maniatis, T., Jeffrey. A., and Kleid, G. D. Nucleotide sequence of the rightward operator of phage. Proc. Nati. Acad. Sci. USA, 72: 1184-1188, 1975.
27. Rigby, P. W. S., Dieckman, M., Rhodes, C., and Berg. P. Labeling deoxyri-bonucleic acid to high specific activity in vitro by nick-translation with DNApolymerase I. J. Mol. Biol., 113: 237-251, 1977.
28. Lai, E. C, Woo, S. L. C.. Dugaiczyk, A. K., and O'Malley, B. W. The
ovalbumin gene: alÃelescreated by mutations in the intervening sequences ofthe natural gene. Cell, 16: 201-211, 1979.
29. Wahl. G. M., Stern, M., and Stark. G. R. Efficient transfer of large DNAfragments from agarose gels to diarobenzyloxymethil-paper and rapid hybridization by using dextran sulfate. Proc. Nati. Acad. Sci. USA, 76: 3683-3687, 1979.
30. Zakut-Houri. R.. Bienz-Tadmor. B., Givol, D., and Oren, M. Human p53cellular tumor antigen: cDNA sequence and expression of COS cells. EMBOJ., 4: 1251-1255, 1985.
31. Torelli, G., Seller!, L., Donelli, A., Ferrari, S., Emilia, G., Venturelli, D.,Moretti, L., and Torelli, U. Activation of c-myc expression by PHA stimulation in normal human T-lymphocytes. Mol. Cell. Biol., S: 2874-2877,1985.
32. Kaczmarek, L., Calabretta, B., and Baserga, R. Expression of cell-cycledependent genes in phytohemagglutinin stimulated human lymphocytes.Proc. Nati. Acad. Sci. USA, 82: 5375-5379, 1985.
33. Slamon, D. J., de Kernion, J. B., Verma, I. M., and Cline, M. J. Expressionof cellular oncogenes in human malignancies. Science (Wash. DC), 224:250-262, 1984.
34. Erisman, M. D., Rothberg. P. G.. Diehl, R. E., Morse, C. C., Spandorfer, J.M., and Astrin, S. M. Deregulation of c-myc gene expression in human coloncarcinoma is not accompanied by amplification or rearrangement of the gene.Mol. Cell. Biol., 5: 1969-1976, 1985.
35. Calabretta. B.. Kaczmarek, L., Ming. P-M. L., Au, F., and Ming, S-C.Expression of c-myc and other cell-cycle dependent genes in human colonneoplasia. Cancer Res.. 45: 6000-6005. 1985.
36. Coulier, F., Imbert, J., Albert, J.. Jeunet, E., Lawrence. J-J., Crawford, L.,and Birg, F. Permanent expression of p53 in FR373 rat cells but cell cycle-dependent association with large T antigen in simian virus 40 transformants.EMBO J.. 4: 3413-3418, 1985.
37. Matlashewski, G., Lamb, P., Pirn, D., Peacock, J.. Crawford, L., and Ben-chimol, S. Isolation and characterization of a human p53 cDNA clone:expression of the human p53 gene. EMBO J., 3: 3257-3262.
5742
on June 5, 2018. © 1986 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

1986;46:5738-5742. Cancer Res Bruno Calabretta, Leszek Kaczmarek, Licia Selleri, et al. Antigen Messenger RNA in Normal and Neoplastic Cells
53,000 TumorrMGrowth-dependent Expression of Human
Updated version
http://cancerres.aacrjournals.org/content/46/11/5738
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/46/11/5738To request permission to re-use all or part of this article, use this link
on June 5, 2018. © 1986 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from





![DECISION - rtbuvic.com.au · [2016] FWCA 5738 DECISION [2016] FWCA 5738 2 [7] The Agreement is approved and, in accordance with s.54 of the Act, will operate from 23 August 2016](https://img.pdfslide.us/doc/110x75/5f8a85cbb1c7e85c8f719c02/decision-2016-fwca-5738-decision-2016-fwca-5738-2-7-the-agreement-is-approved.jpg)