Embed Size (px)
Citation preview

Growth and Morphogenesis of Globular and Older Embryos of Capsella in CultureAuthor(s): V. Raghavan and John G. TorreySource: American Journal of Botany, Vol. 50, No. 6, Part 1 (Jul., 1963), pp. 540-551Published by: Botanical Society of AmericaStable URL: http://www.jstor.org/stable/2440029 .Accessed: 19/08/2011 13:23
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
Botanical Society of America is collaborating with JSTOR to digitize, preserve and extend access to AmericanJournal of Botany.
http://www.jstor.org

GROWTH AND MORPHOGENESIS OF GLOBULAR AND OLDER EMBRYOS OF CAPSELLA IN CULTURE1 2
V. RAGHAVAN AND JOHN G. TORREY
The Biological Laboratories, Harvard University, Cambridge 38, Massachusetts
A B S T R A C T
RAGHAVAN, V., and J. G. TORREY. (Harvard U., Cambridge, Mass.) Growth and morphogenesis of globular and older embryos of Capsella in culture. Amer. Jour. Bot. 50(6): 540-551. Illus. 1963.-Early heart-shaped and older embryos of Capsella bursa-pastoris (>80,u long), when cul- tured aseptically in a semi-solid medium containing mineral salts, vitamins, and 2% sucrose, de- veloped into small plantlets. When grown in a 12-hr light/12-hr dark cycle (light-grown cultures), formation of the primary root system was suppressed in embryos <1000u long, while embryos cultured in continuous dark routinely initiated a primary root system and several lateral roots. In embryos > 1000l long, the inhibition of formation of the root system in light did not occur. Growth of the shoot apex of intermediate and torpedo-shaped embryos was slightly accelerated in light. There was a correlation between the rate of growth of the embryos and their initial length. Better growth in length was observed in light-grown than in dark-grown cultures. Supplementing the basal medium with indoleacetic acid, kinetin, and adenine sulfate permitted successful develop- ment of globular embryos (<80M long) in vitro. Although a high sucrose concentration or a high salt concentration in the medium partially replaced the effect of the growth factors in inducing development of the globular embryos, no absolute requirement for a high osmotic concentration in the medium was found. The results favor caution in interpreting the inability of small embryos to grow in terms of the osmotic conditions of the culture milieu.
THE FACTORS that control the progressive and orderly development of embryos throughout their ontogeny have attracted the attention of experi- mental embryologists for some time (Needham, 1942; Brachet, 1950; Wardlaw, 1955). There is now substantial indirect evidence that growth and differentiation of plant embryos may result from an induction by gradients of nutritional substances and hormonal factors in the immediate environ- ment of the developing embryos (Rijven, 1952; Wardlaw, 1955). In the early stages of embryog- eny, embryos develop at the expense of the surrounding endosperm tissue, and as far as the evidence goes, such embryos are highly hetero- trophic in character. Attempts to culture very small plant embryos, especially at the morpho- logical stages designated as "globular" and "heart- shaped," in chemically defined nutrient media outside the plant tissues have met with little success (Van Overbeek, Conklin and Blakeslee, 1942; Rijven, 1952; Norstog, 1961; Swamy, 1961). It is only after the embryos have developed within the ovular tissues to the beginning of cotyle- donary development and the attainment of bi- lateral symmetry that they become sufficiently independent and autotrophic to lend themselves to culture in vitro even in complex nutrient media. While the nutritional requirements of young em- bryos are complex, and their responses often highly variable, progressively older embryos show less rigorous requirements and more definitive responses to added nutrients in the medium.
1 Received for publication November 8, 1962. 2 This work was supported by a grant (G-18684) from
the National Science Foundation.
The genus Capsella has been a favorite object for both descriptive and experimental embryol- ogy since the work of Hanstein (1870). Later, Famintzin (1879), Schaffner (1906), and Soueges (1914, 1919) made more thorough studies of the embryogenesis of C. bursa-pastoris. The first pub- lished report of in vitro culture of the embryos of this species was by Rijven (1952). A modifica- tion of Rijven's classification and terminology concerning embryo stages will be used in this paper (Table 1). Rijven found that growth in culture of the embryos excised from the ovules during the course of development depended upon the presence in the medium of amino acids which apparently regulated the metabolic processes in- volved in growth. In addition, Rijven (1952, 1955, 1956) observed that the addition of glutamine produced a stimulation of growth which was even greater than that obtained in the presence of casein hydrolysate or a complex mixture of amino acids of the composition of edestin. Nevertheless, Rijven did not succeed in cultivating embryos less than 100lu in mean initial length. Embryos at the heart-shaped or torpedo-shaped stages (140- 700,u long) responded consistently to the addition of nutrients only when grown in hanging-drop culture in a medium of high osmotic pressure obtained by the addition of 12-18% sucrose. More recently, using Rijven's experimental procedure, Veen (1961, 1962) studied the effects of substances like gibberellic acid and kinetin on the growth of Capsella embryos in culture. Embryos of this species have also been used for cyto-histochemical analysis of embryogenesis (Pollock and Jensen, 1961, 1962).
540

July, 1963] RAGHAVAN AND TORREY-CAPSELLA EMBRYOS 541
Rijven's (1952) work leaves the impression that the osmotic value of the nutrient medium is an important factor in the successful development of small embryos of Capsella in culture, the high osmotic pressure preventing the appearance of signs of precocious germination associated with cell enlargement and cell elongation. The same conclusion was reached by several other investi- gators (Ziebur and Brink, 1951; Rietsema, Satina and Blakeslee, 1953; Mauney, 1961; Norstog, 1961; Swamy, 1961). We have found that re- producible normal growth and differentiation can be achieved in immature embryos of Capsella when they are grown in vitro in a semi-solid medium containing 2% sucrose; thus the osmotic value of the culture medium may be of relatively little importance in the morphogenesis of the cultured embryos. The results of these studies are de- scribed in this paper. In addition, this report pre- sents experiments indicating the importance of growth factors in the culture of globular embryos. These results have significance in support of the view that the transition from heterotrophy to autotrophy during embryogenesis depends upon the availability of specific growth factors to the embryo.
MATERIAL AND METHODS-Embryos of Capsella bursa-pastoris Medic. were obtained from plants grown in the greenhouse in a mixture of equal parts of sand, perlite and leaf mold under a supplementary incandescent illumination of ca. 200 ft-c during the daytime. In the spring and summer months, preliminary experiments were done with embryos obtained from plants in nature, with essentially the same results.
Dissection of the embryos for culture was car- ried out aseptically in a transfer room previously illuminated by UV-sterilamps. Capsules in the desired stages of development were surface-ster- ilized for 4 min in a 5% filtered solution of "Pitt- chlor" (manufactured by Columbia-Southern Chemicals Co., Pittsburgh, Pa.) and washed re- peatedly in sterile water. Further operations were carried out under a dissecting microscope at a magnification of 90X, keeping the tissues in a depression slide containing a few drops of the liquid medium. The outer wall of the capsule was removed by a cut in the region of the placenta; the halves were pulled apart with forceps to ex- pose the ovules. Torpedo-shaped and younger embryos were confined to 1 longitudinal half of the ovule and were clearly visible through the chalaza either because of their green color (inter- mediate and torpedo-shaped), or because of the transparent vesicle of their suspensors (globular and heart-shaped). With the help of a sharp, mounted blade, the ovule was split longitudinally to isolate the embryo-containing half. Further manipulations to separate the embryos from the surrounding ovular tissues were performed with # 11 surgeon's blades (Clay-Adams). Early glob- ular embryos also could be removed with no
apparent injury. A small incision in the ovule, followed by slight pressure with a blunt needle, was enough to free the embryos in advanced stages of development from the ovules; these embryos, without any suspensor cells, floated freely in the medium. Unless otherwise stated, embryos in the globular and heart-shaped stages were cultured with the cells of the suspensor intact. The excised embryos were transferred by micropipettes to standard 10-cm Petri dishes con- tairiing 25 ml of the solid medium and/or sus- pended in a small drop of the liquid medium on a cover-slip over a cavity slide, and sealed with a melted 1:1 paraffin-petrolatum mixture (hanging drop cultures). Usually 6-8 embryos of the de- sired stages were cultured in a Petri dish or in individual hanging drops. All cultures of globular embryos were in hanging drops; older stages were cultured routinely on agar medium. Each experi- ment was done 3-4 times, with 6-8 embryos of a stage per treatment. The pooled data from these experiments assured the inclusion of a sufficient number of embryos of the same or closely similar size to allow statistical analysis. The Petri dishes were sealed with masking tape to prevent desic- cation of the cultures. Since the number of very small embryos that could be conveniently isolated and handled at a time was limited, in experiments involving many different concentrations of a test substance, there was an interval of several days between setting up the initial and final concen- tration series.
The basal medium used in this study consisted of macronutrient salts, trace elements, and vita- mins in proportions suggested by Dr. H. Castle, Yale University (personal communication, 1961). The composition of this basal medium is as follows:
I. Macronutrient salts (mg/liter): 480 Ca- (N03)2.4H20; 63 MgSO4*7H20; 63 KN03; 42 KCl; 60 KH2PO4.
II. Micronutrient salts (mg/liter): 0.56 H3B03; 0.36 MnCl2*4H20; 0.42 ZnCl2; 0.27 CuCl2. 2H20; 1.55 (NH4)6Mo7024.4H20; 3.08 fer- ric tartrate.
III. Vitamins (mg/liter): 0.1 thiamin hydro- chloride; 0.1 pyridoxin hydrochloride; 0.5 niacin.
IV. Sucrose 20 g/liter; specially purified Difco agar (where applicable) 9 g/liter.
V. Glass-distilled water to 1 liter. All the components of the medium were mixed
together and sterilized for 15 min at 15 lb/in2. A number of growth substances were added sepa- rately or in combination to the basal medium in attempts to induce growth in globular embryos. These supplements were mixed with the basal medium and autoclaved. The final pH of the medium was 4.9-5.0.
The cultures were kept in a culture room at 2541 C and given 12 hr illumination daily by a

542 AMERICAN JOURNAL OF BOTANY [Vol. 50
TABLE 1. Stages in the embryogenesis of Capsella. Length was measured from the base of the suspensor to the extreme tip of the embryo, including the cotyledons. (Classification modified from Rijven, 1952)
Developmental stage Length, microns
Early globular ca. 20-60 Late globular 61-80 Early heart-shaped 81-150 Late heart-shaped 151-250 Intermediate 251-450 Torpedo-shaped 451-700 Walking-stick-shaped 701-1000 Inverted-U-shaped 1001-1700 Mature embryo >1701
combination of cool-white fluorescent tubes and incandescent lamps (light-grown cultures) giving ca. 50 ft-c at the level of the cultures. Parallel sets of cultures were maintained in an incubator at 25 ? 1 C which was kept completely dark ex- cept for brief periods of opening for examination (dark-grown cultures). Transfers to fresh media were made at approximately 4-week intervals and the experiments were routinely discontinued after 12 weeks.
Unless otherwise specified, growth measure- ments were made at the end of 7 days with a dissecting microscope equipped with an ocular micrometer. Although the embryos were still growing after this period, further growth was less regular and unreliable for quantitative evaluation. Growth was expressed as the percentage increase over the initial length in the longitudinal axis of the embryo excluding the length of the suspensor. Since the initial length of the embryos varied, embryos were divided on the basis of their mor- phological stage of development into definite stage classes (Table 1). For clarity of presentation of results, data on embryos of approximately the same initial length were pooled to determine the average rate of growth.
The morphological changes occurring in the cultured embryos were followed by periodic ob- servations of the cultures under a dissecting micro- scope. For microscopic examination, samples were fixed in formalin-acetic acid-alcohol. They were dehydrated in n-butyl alcohol series and embedded in Tissuemat. Sections cut at 1l0, were stained in phloxine and fast green.
No systematic attempts were made to rear to maturity seedlings derived from the cultured em- bryos. In a few trials made, when the growing embryos were transferred to fresh media, many of them produced plantlets.
Statistical methods follow Snedecor (1956). RESULTS-Early in this work it became appar-
ent that the responses in culture of embryos differing in length, even over a narrow range, varied widely. To evaluate the effects of compo- sition of the medium on the growth of embryos of known developmental stages, a detailed morpho-
logical examination was made of embryos varying in length from 50 to 1700,u. With the help of camera lucida drawings of such isolated embryos, a classification of the developmental stages, based on Rijven (1952), was made (Table 1). Early globular embryos were generally 16-32 cells in number and ca. 40,u in diameter (Fig. 1). A transition from radial to bilateral symmetry was apparent in the early heart-shaped embryos (Fig. 2), which showed for the first time a visible demarcation between the presumptive shoot and root regions. Embryos in the intermediate and torpedo stages had basically the same morphology (Fig. 3, 4). The axis was short, the cotyledons were well developed, and internally they showed procambial tissue differentiation. In embryos >250,4 long, the cotyledons comprised over 3 of their length. The "walking-stick-shaped" and the "inverted-U-shaped" embryos were further differ- entiated by the elongation of the hypocotyl and the organization of the meristematic regions of the root and shoot (Fig. 5).
Experiments with heart-shaped and larger em- bryos-Isolated embryos of Capsella > 80,u in length (early heart-shaped; Fig. 2) grew readily in the basal medium described above and differ- entiated regularly into shoot, root and leaf pri- mordia, although the process was relatively slow. Since reference will be made to the pattern of development of the embryos in the basal medium, a brief description of the sequence will be given. The growth and organization of meristematic regions in the early and late heart-shaped embryos cultured in a semi-solid medium appeared alike. In Fig. 6-9 the external morphological forms of embryos initially 81 A in length are shown at various times during 2-3 months in culture. The first evidence of growth in the embryos was an increase in diameter due to rapid proliferation of cells during the first week. This was followed by the development of the distal half of the embryo into 2 cotyledonary lobes, at an angle of 450 to the main axis. After 3 weeks in culture, a root apex was initiated at the proximal end. The sus- pensor cells were sloughed off from the main body of the embryo shortly afterwards. Further devel- opment of the embryo was marked by the initia- tion and increase in size of the shoot apex which was visible externallv as a mound of tissue be- tween the cotyledons (Fig. 6-8, 13-15). The shoot meristem showed little internal organization and lacked the characteristic demarcation into cyto- histological layers or zones (Fig. 14, 15). The apex developed slowly, producing the first pair of leaves in 4-6 weeks (Fig. 16). By this time the embryos had grown to about 3 mm. Almost si- multaneously in dark-grown cultures, the root became active, elongating rapidly to form a pri- mary root several millimeters long and, subse- quently, a few laterals (Fig. 9). Elongation of the primary root showed the highest rate observed for any organ of the cultured embryo, averaging
July, 1963] RtAGHAVAN AND TORREY-CAPSELLA EMBRYOS 543
between 200 and 300,u daily. Development of the primary root was suppressed in embryos grown in light. This inability of embryos cultured in light to initiate a primary root system persisted throughout the period of the experiment. During further development the embryos produced addi- tional leaves rapidly. The leaves were 6-8 mm in length, and were thick and linear and had none of the features of the leaves of the adult plant. Shoot length at this time was less than 1 cm, and the plantlets were characterized by an almost complete absence of internodal elongation (Fig. 9). No evident correlations were obtained between
shoot development and the conditions of illumi- nation in the heart-shaped embryos.
These attempts to grow heart-shaped embryos in a simple medium demonstrate that immature embryos are strikingly similar to the mature em- bryos in their organogenesis. All stages of embryo development through the torpedo stage were nor- mal in embryos cultured in vitro. Thereafter, cultured embryos developed without the char- acteristic curvature apparent in the walking-stick- shaped and inverted-U-shaped embryos which de- veloped within the restrictions imposed by the ovule. No significant deleterious effect was caused
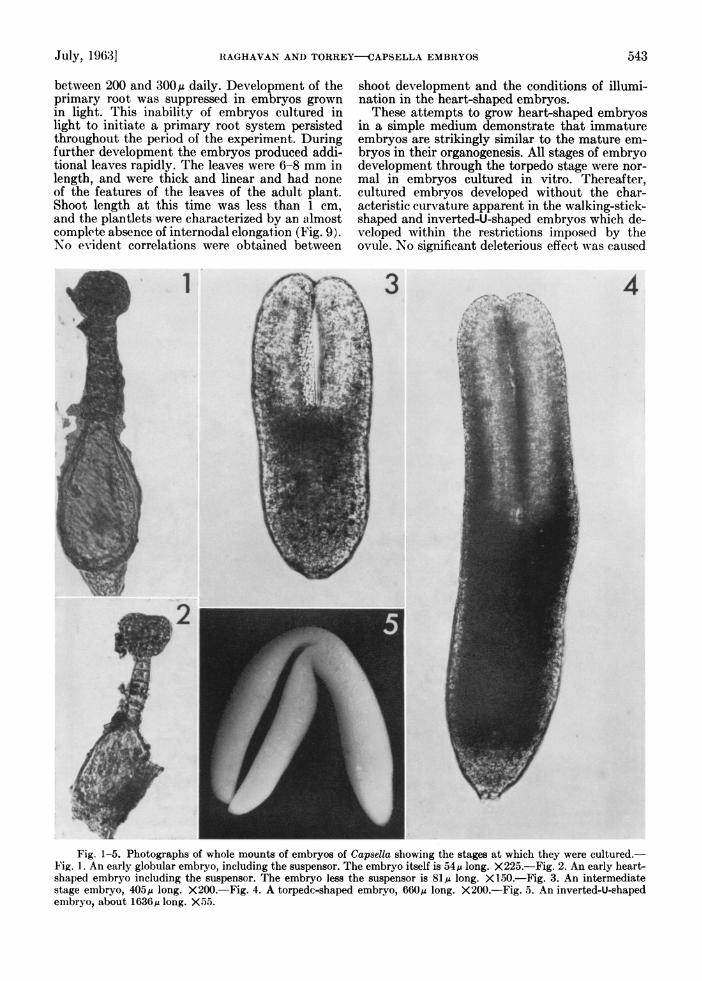
Fig. 1-5. Photographs of whole mounts of embryos of Capsella showinig the stages at which they were cultured.- Fig. 1. An early globular embryo, including the suspensor. The embryo itself is 54/.A long. X225.-Fig. 2. An early heart- shaped embryo including the suspensor. The embryo less the suspensor is 811A long. X 150.-Fig. 3. An intermediate stage embryo, 405MA long. X200 -Fig. 4. A torpedc-shaped embryo, 660jA long. X200.-Fig. 5. An inverted-U-shaped embryo, about 1636A long. X5.
544 AMERICAN JOURNAL OF BOTANY [Vol. 50
-~~~~7_
_v_ _ _ . .~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~.. . .. ........
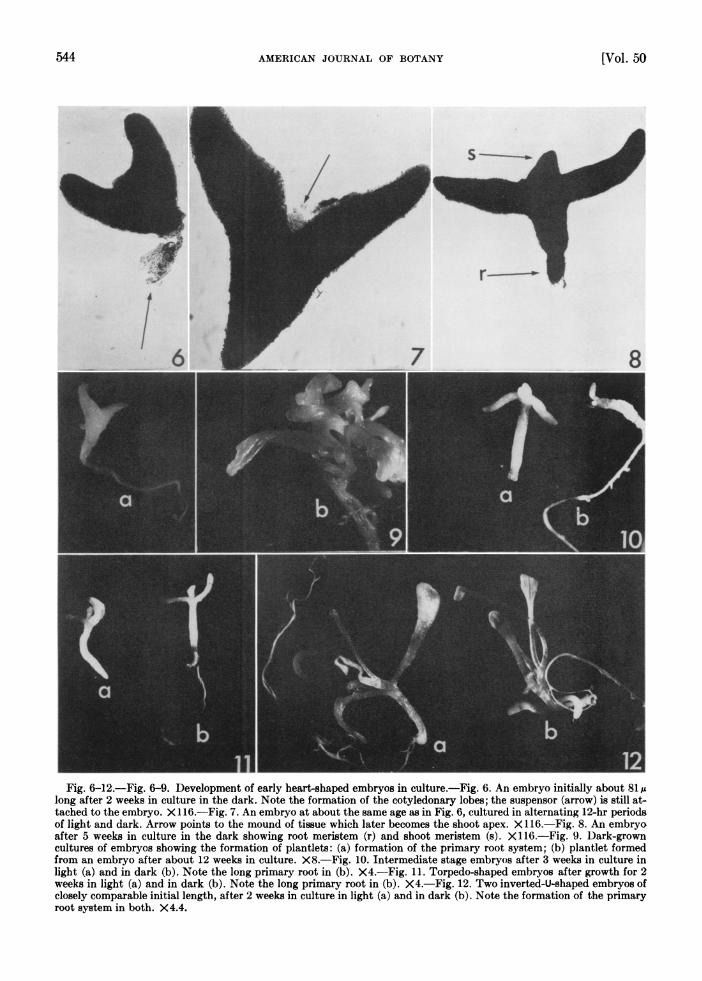
Fig 6 12 Fig. 6-9. Development of early heart-shaped embryos in culture-Fig. 6. An embryo initially about.81.
::
:~~~~~~~~~~~~~~~~~. :::.:. : ... . .. :.:. .. ...........j:::: of....... l a.... ... g.. 8. A e r
after . w in cams) .. ..u.... in. ter. 9
light (a)1andFin dar(b. Neeotmenth long priarly rootsine (mb).sX4ulur.-Fig. 116TrpdosApe embryos intafter growth for
tahe toteeby. X 16-Fig 7. An emry ataottesm gsii. 6_ cutrd in alentg 12heid
aftr ek ncluei h aksoigro eriste .r and sho meise S_) X11. ig 9_ Dr-rw
wek nlight (a) and in dark (b). Note the long primary root in (b). X 4.-Fig. 121. Twreo ivre--shaped embryos atrgohfof clsl oprbeiiillnt,atr2weeks i utr in light (a) and in dark (b). Note the fogPia-
oo n() .Fi.1.Toivrmationshaof teprmaryoso
ro syse in both X4.4.I

July, 1963] RAGHAVAN AND TORREY-CAPSELLA EMBRYOS 545
TABLE 2. Growth of Capsella embryos of different ages in light and dark. Growth data represent increase in length as per cent of initial length dt standard error after 7 days in vitro
Dark cultures Light cultures Developmental stage -- --
Initial length, IU Percent growth Initial length, IA Percent growth
Early heart-shaped 88.0 81.9 + 5.9 88.0 90.4 + 11.3
Late heart-shaped 180.0 130.8 4 9.9 208.0 184.3 + 18.0
Intermediate 333.0 113.1 ? 9.2 277.5 166.4 4+ 25.1 375.0 89.9 ? 8.8 347.0 173.9 ? 10.2 444.0 100.0 ? 10.4
Torpedo-shaped 569.0 134.8 + 11.0 569.0 198.1 ? 12.2 655.0 107.2 ? 8.7 652.0 171.2 ? 9.7
Walking-stick-shaped 777.0 142.7 + 12.4 832.5 111.6 4 5.0 874.0 125.3 ? 9.1 888.0 114.0 ? 9.7
943.5 150.1 4 7.8
Inverted-U-shaped 1401.0 132.3 ? 2.4 1540.0 137.8 ? 4.1 1628.0 128.0 ? 5.9 1679.0 122.2 4 6.3
by lack of high cencentrations of sucrose in the medium, as claimed by Rijven (1952).3
The pattern of organogenesis in the interme- diate and torpedo-shaped embryos (Fig. 3, 4) was more or less similar to that of the heart-shaped embryos. Growth in length of the embryos was initiated within 48 hr after transfer to the medium. This growth represented the over-all growth of the embryos and not merely the growth due to elongation of the hypocotyl. Thus, the initial responses of the embryos, when grown in a semi- solid medium, were profoundly different from the precocious germination described by Rijven (19,52). While growth of the primary root was suppressed in light-grown cultures (Fig. 10, 11), a considerable number of embryos in light de- veloped a shoot system a few days ahead of the dark cultures.
Other experiments in which embryos of the walking-stick-shaped, inverted-U-shaped and ma- ture stages were cultured in the basal medium were also of interest with respect to the factors affecting the initiation and growth of the shoot and root primordia. Walking-stick embryos responded in the same way as torpedo-shaped embryos with re- spect to their morphogenesis in light and dark. On the other hand, when embryos about 1000,u long (Fig. 5) were planted under both light and dark conditions, the responses were uniform. They showed prolific growth of the primary and lateral roots in 1 week in culture and an elaborate shoot system composed of several linear leaves within 2 weeks (Fig. 12).
3 In a personal communication Dr. H. Castle (Yale University) informs us that he also succeeded in growing normal plants from Capsella embryos isolated at the 16-32-cell stage, and grown in the medium described in this paper (see also Castle, quoted in Naylor, 1952).
Data on the growth of embryos of different ages after 7 days in culture are summarized in Table 2. The results show clearly the greater growth in length of the embryos in light than in dark: in this respect our observations are not in agreement with those of Rijven (1952) who found light inhibitory or without effect on the growth of the embryos. In agreement with Rijven, we ob- served a correlation between the rate of growth of the embryos and their initial length at excision. Thus, younger embryos did not grow to the same proportional extent as the older ones, suggesting a nutritional limitation. This effect was consistent in embryos up to ca. 700, long in light, while in dark-grown cultures and in longer embryos no clear relationship was apparent.
Experiments with globular embryos-Globular embryos (<80,g long) cultured in the basal med- ium, either in semi-solid cultures or in hanging drops, did not show any signs of cell division and growth even after prolonged periods in culture. In considering the factors controlling the growth in vitro of these embryos, it became increasingly apparent that the problem was chiefly a nutri- tional one, the embryos probably being limited in their potentialities for cell division and growth by the lack of one or more specific growth factors in the medium. Indirect support for this view came from observations gathered during attempts to culture globular embryos. Anticipating the fact that the tiny embryos would be particularly sensi- tive to heavy metal contaminants in the medium, an experiment was run using macronutrient salts specially purified to reduce the total heavy metal content of the medium to less than 10 ppb. Em- bryos >80 , cultured in the purified medium sur- passed the growth obtained in medium using rea- gent grade salts, but in no cases were cell divisions

546 AMERICAN JOURNAL OF BOTANY [Vol. 50
TABLE 3. Growth of globular embryos in different media. Growth values are expressed as the percentage increase after 10 days in culture in the length of the embryo over the initial length i standard error
Initial length of the embryos, microns Composition of media
27.0 40.5 54.0 67.5
Basal medium alone 0(7)a 0(9) 75.0(1/24)b,c 35.0 i 5.0(4/18)0
Basal medium with IAA: 0.0001 mg/liter 0(3) 0(1) 0(3) 0(3) 0.001 mg/liter 0(1) 0(5) 0(8) 40.0(1/6)c 0.01 mg/liter - 0(6) 25.0 : 0(2/8)c 40.0(1/3)0 0.10 mg/liter 0(1) 33.3(1/4)C 25.0 i 0(2/8)0 25.0 4 5.0(4/5)c 1.0 mg/liter 0(3) 0(5) 0(4)
10.0 mg/liter - 0(2) 0(1) 0(5)
Basal medium + 0.10 mg/liter IAA and kinetin: 0.0001 mg/liter - 0(6) 32.1 ? 7.1(7/10)c 40.0 i 0(4/5)0
0.001 mg/liter 0(1) 33.3(1/6)0 23.0 i 1.2(5/8)d 45.0 i 5.0(4/4)d 0.01 mg/liter 0(6) - 25.0(1/5)d 0.10 mg/liter 0(3) - 0(4) 1.0 mg/liter 0(5) - 0 (1)
10.0 mg/liter - 0(6)
Basal medium + 0.10 mg/liter IAA, 0.001 mg/liter kinetin and adenine sulfate:
0.0001 mg/liter 50.0(1/3)0 33.3 i 0(2/3)! 68.8 ? 15.9(4/6)0 60.0(1/1)d 0.001 mg/liter 0(4) 77.7 4 29.4(3/4)d 154.2 ?46.2(13/14)0 287.5 i 16.4(8/9)0 0.01 mg/liter 0(3) 0(1) 50.0 i 0(2/5)d 0.10 mg/liter 0(3) 0(2) 1.0 mg/liter 0(3) 0(1) 0(1)
10.0 mg/liter 0(3) 0(2) 0(1) Basal medium with 12.0% sucrose 33.0(1/2)0 85.0 i 16.3(10/10)e 171.4 i 18.4(14/14)e
Basal medium with 18.0% sucrose - 0(2) 99.5 i 23.2(10/10)e 150.0 i 33.6(8/8)e Basal medium with l0X major salts 0(1) 0(3) 60.7 A 13.2(7/8)0 111.4 i 13.7(7/8)e Basal medium with 0.10 mg/liter IAA,
0.001 mg/liter kinetin and 0.001 mg/liter adenine sulfate, and 12.0% sucrose - 0(3) 75.0 i 22.4(5/5)e 83.3 i 15.8(6/6)0
Basal medium with 0.10 mg/liter IAA, 0.001 mg/liter kinetin and 0.001 mg/liter adenine sulfate and IOX major salts 0(1) 31.6 ? 8.7(6/7)d 105.0 4 3.7(4/4)d,e
a The figures in parenthesis indicate the number of embryos cultured. b The pairs of figures in parenthesis indicate the number of embryos which showed any measurable increase in size
and the number of embryos cultured. Thus, (1/24) means that out of 24 embryos cultured, only 1 had grown to the extent shown on the left of this figure in the table.
e Growth due to increase in volume of the embryos without evident cell divisions. These embryos did not show any appreciable growth or morphogenesis even after prolonged periods in culture.
d Embryos which develop by both cell enlargement and cell division. Those showing cell division predominantly assumed the shape of a sphere. Often growth was terminated at this stage and no differentiation of cotyledons was observed.
e Cell division was rapid during the first 1-2 weeks in culture. Many embryos in this group proceeded to form the cotyledonarv lobes and the root and shoot meristems. For description see text.
and growth observed in globular embryos. It was also conceivable that certain of the organic supple- ments in the medium such as the vitamins were below the level required for optimum growth of the globular embryos. However, even in media with 5 X or ] OX vitamins, attempts to grow these embryos were unsuccessful. In additional experi-
ments, high or low pH of the medium, high oxygen tension, and high (30 C) or low (20 and 10 C) temperatures were without beneficial effect.
In further attempts to achieve growth of the globular embryos, various mixtures containing the growth factors indoleacetic acid (IAA), kinetin, and adenine sulfate were tested as components of

July, 1963] RAGHAVAN AND TORREY-CAPSELLA EMBRYOS 547
the basal medium, because their profound influ- ence on organ initiation had been observed in heart-shaped and older embryos. The results of these experiments are summarized in Table 3. Because of the difficulty in handling globular embryos, cultures described in this section were made in hanging drops and kept in the incubator in the dark at 25 C.
When used alone, IAA was generally toxic to the embryos, although in a few cases increases in length of up to 30 ,I were noted during the first week in culture. On addition of kinetin several instances of limited growth in size of embryos accompanied by cell division were recorded at
0.0001-0.001 mg/liter kinetin (Veen, 1962). The consistency of the promotive effect of kinetin, although admittedly small, made us look for other substances which, in combination with IAA and kinetin, would sustain cell division and growth in the embryos. As is apparent in Table 3, the addi- tion of low concentration of adenine to the medium had a dramatic effect in inducing continued growth of the embryos. The results show that a combina- tion of IAA, kinetin, and adenine sulfate added as supplements to the basal medium provides the best medium for continued cell division and growth of embryos initially in the range 50-80,u in length.
/ *.~~ ~ ~ ~ ~~~~~~~~~~~~~~~~~~~ . .........
Fig. 13 . An embryo after 3 weeks in cultureinthedark. The:position:oftheshoote . . by a w T
ape is no viil in, the...plane....of..sectioning... X6.-Fig. 14. A lih-rw cutr of comarbl ag. The shoo men
* ~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~. . .. . .. .. ...: ....... s
*~~~~~~~~~~~~~~~~~~~~~~~~~~~~~ .. . ......... ..... ..."
. . . ....... .. ... ... ... ........... . ... .. .. .. ..... . ..... . ... .. . .
:. ~ ~ ~ ~ ~ ~~~~~~~~~~~~~~ .. .. ..:.
/ ~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~...: .........:_..:::.......j
stem (arrow) is prominent. X260.-Fig. 15. Section of an embryo after 5 weeks in the dark: (r) root; (s) shoot. Note the lack of cytohistological zonation pattern in the apical meristem. X 130.-Fig. 16. A light-grown embryo 5 weeks old showing the formation of the first pair of leaves. X 130.
548 AMERICAN JOURNAL OF BOTANY [Vol. 50
; A ~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~-~ffi _ 19d;
tT~ ~ ~ ~~~~~2
... .. ........ _ _
. . ;sbS S _E _ I i E .r:2i
:~~~~. g i . - g ?. .
........... l . ....... _ . _ I . . f L~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~...........
...... i - _ . _ I * ~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~.....
* .! Rs E I | ~~~~~~~~~~- T
....... .....
*'.'t.',-::'ts s X_ ~~~~~~~~~~~~~~~~~~~~~m4 .'
'X'. . '4_ l?BS#. _ * , a _ e 3;,,~~~~~~~~~~~~~~~~~~~~~S"p9

July, 1963] RAGHAVAN AND TORREY-CAI SELLA EMBRYOS 549
The only other supplements to the basal med- ium which supported growth of embryos of com- parable length were either high concentrations (12% and 18%) sticrose or IOX the concentration of major salts. Thus, the effect of a delicate bal- ance between IAA, kinetin, and adenine sulfate can be substituted for, in part, by a high sucrose concentration or an increased salt level. However, even these media were inadequate to induce cellu- lar divisions and growth of still smaller embryos.
In Fig. 17, 18 is illustrated the development of an early globtular embryo, initially 54,u long (cf. lF'ig. 1), in the basal medium supplemented with IAA, kinetin and adenine at 0.1, 0.001 and 0.001 mg/liter respectively. Duiring the first 7-10 days in culture, the embryo increased in size as a sphere by irregular cell div-isions and cell enlarge- ment (Fig. 17). The lack of an organized division se(luence was especially evident in the outer layers of cells, some of which protruded from the body of the embryo. After the embryo had grown to a few hundred microns in length, bilateral differen- tiation was observed with the, formation of the cotyledons. Fturther growth of the embryo was similar to that described earlier for heart-shaped embryos. Occasionally, abnormal structures, rem- iniscent of no specific stages in the embryogenesis of the species, resulted from continued division at and following the sphere-like stage (Fig. 19, 20). Most embryos <54 p long at isolation did not grow in this medium, while a few which showed signs of growth did not develop beyond the period of early cell divisions (lig. 21). The pattern of development observed in the globuilar embryos when cutltured in high sucrose or high salt media was similar to that described above.
DISCUSSION-One important modification of the basal meditum made by previous workers in at- tempts to cullture very small plant embryos has been the adjustment of its osmotic value. Im- proved development of immature embryos follow- ing such modification led them to the concluision that a medium isotonic with the ovular sap is a sine qua non for successful culture of very small embryos. In their work on immatuire barley em- bryos, Ziebur and Brink (1,951) and Norstog (1961) secured suitably high osmotic pressures by the uise of 12.0-12.5% suicrose and succeeded in growing embryos of ()0-3'OOA initial length. Iso- lated embryos of Datura strammSium of different ages required different minimal suicrose concentra- tions for development, the optimum for preheart- shaped embryos being 8-12% (Rietsema et al., 1953). Rijven (1952) routinely used 12% and 18% sucrose to grow torpedo-shaped and heart-shaped
embryos of Capsella. Embryos of cotton (Mauney, 1961) and Citrus (Swamy, 1961) have also been shown to be sensitive to the osmotic value of the culture medium. On the other hand, we have found that in a semi-solid medium containing only mineral salts, vitamins, and 2% sucrose, embryos of Capsella as small as 81 , regularly undergo cellular divisions and differentiation without evi- dent dependence on a high osmotic value in the medium.
In view of the implications from our findings, it is necessary to re-examine the above-mentioned reports about the importance of the osmotic bal- ance of the medium in the culture of small em- bryos. Our findings provide new information in 2 respects: First, the nutritional requirements of heart-shaped and older embryos of Capsella are not as complex as reported by Rijven (1952). For example, no requirement was found for an exog- enous amino acid supply. Second, the inhibition in growth of embryos in a hypotonic medium ob- served by Rijven need not be ascribed to the physical state of the culture medium. The growth of heart-shaped embryos into small plantlets in a simple medium indicates that for these embryos the nutritional requirements are suirprisingly simple. In such embryos development is sustained by amino acids and growth factors available within the embryos themselves, or the embryos can syn- thesize them from the mineral salts and sucrose provided in the medium.
Rijven (1952) attempted to explain the ob- served effects of the osmotic value of the culture medium in terms of the control of growth that occurs in the embryos; in his view high osmotic pressure prevents cell elongation leading to germi- nation. The pattern of development in vitro of early heart-shaped embryos described in this paper is not germination in Rijven's sense, since it does not involve "incipient elongation" of the hypocotyl. Furthermore, the sequence. of develop- ment of such embryos is similar to that of the ma- ture ones cultured in vitro. Thus, like mature em- bryos, even the smallest cultured embryos rou- tinely initiated a root, a shoot apex, and later, several pairs of leaves, in that order.
Our findings show that growth and differen- tiation in the cultured embryos are regulated, not by the osmotic condition of the culture me- dium, but by specific constituents of the medium which may act by their control of differential cell division and cell enlargement during development. Results of experiments on the culture of globular embryos (<80,u long) tend to confirm conclusions concerning the general importance of growth fac-
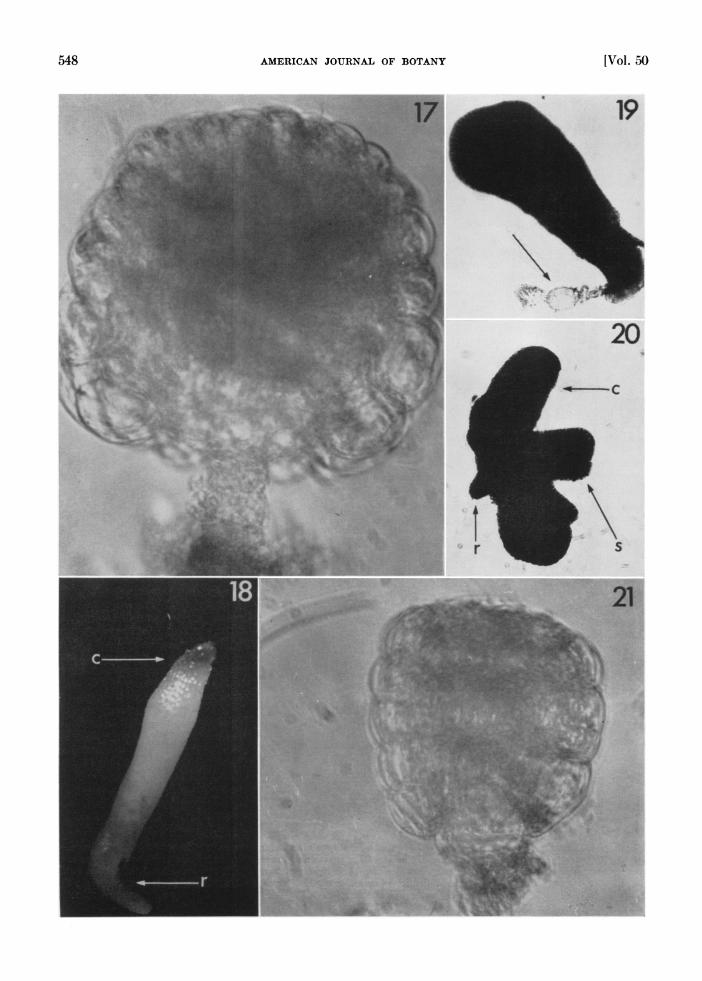
Fig. 17-21. Cultures of globular embryos.-Fig. 17. Formation of a spherical mass from an embryo initially 54,u long, after 10 days in culture. X260.-Fig. 18. D)ifferentiation of the cotyledons (c) and root meristem (r) after 6 weeks in culture. The cotyledons are folded together aiid not visible separately. X57.-Fig. 19. Abnormal growth of a globular embryo in vitro. The suspensor (arrow) is still attached to the embryo. D)evelopment has proceeded without apparent differentiation into the cotyledons. X 130.-Fig. 20. Another type of abnormtal growth in globular embryo in vitro. Note the wing-like cotyledons (c), root (r) and shoot (s) regions. X130.-Fig. 21. An early globular embryo of 40.5,p initial length, after 12 weeks in cuiltuire. I)evelopment terminated after a few early cell (livisions. X260.

550 AMERICAN JOURNAL OF BOTANY LVol. 50
tors in stimulating embryonic development. In other studies, we have found that beneficial effects on the development of heart-shaped and older em- bryos are exerted by varying concentrations of IAA, adenine sulfate and kinetin provided sepa- rately. No such effects were observed in the devel- opment of globular embryos when these sub- stances were supplied individually in the medium. Such observations on the different sensitivities of embryos of different sizes led to the idea that chemical factors, hormone-like in their action, were involved in the inhibition or stimulation of cell division and cell enlargement in embryos of different developmental stages. Clearly, these con- trolling systems must be amenable to experi- mental manipulation through changing the com- position of the growth factor components of the medium. For globular embryos, a balance among all 3 factors-IAA, adenine and kinetin-was achieved which permitted development to proceed. Presumably, a different balance of these and per- haps other factors is necessary for normal devel- opment of isolated embryos of still younger stages. It is perhaps too simple to visualize kinetin as a cell division factor (Miller, 1961) and IAA as controlling cell enlargement; rather the interaction of these factors on the young embryos in isolation provides the necessary chemical enlvironment for the developing embryo to proceed in its normal development drawing upon its chemical environ- ment.
An especially interesting observation from ex- periments with globular embryos is that the bene- ficial effect of added growth factors can be par- tially replaced by growing the embryos in a me- dium with 12-18% sucrose or with an increased level of the macronutrient salts. Thus, globular embryos show a reduced requirement for exo- genous growth factors like IAA, adenine and kine- tin in the presence of high concentrations of sucrose or major salts. Some support for this line of thinking is provided by the works of Van Overbeek et al. (1942) and Rietsema et al. (1953) on the in vitro culture of embryos of Datura stramonium. Van Overbeek et al. found that heart- shaped embryos (150-500,u long) of this species could be successfully grown in a medium con- taining mineral salts, 1% dextrose and a mixture of physiologically active substances (glycine, thi- amin, ascorbic acid, nicotinic acid, pyridoxin HC1, adenine, succinic acid and pantothenic acid) sup- plemented with non-autoclaved coconut milk. In later experiments (Van Overbeek, 1942; Van Overbeek, Siu and Haagen-Smit, 1944), beneficial effects of the same magnitude were obtained using a purified embryo factor preparation from coconut milk in a dilution of 1:19,000. On the other hand, Rietsema et al. have reported that Datura embryos could be grown in a mineral salt medium con- taining 400 mg/liter casein hydrolysate and an optimum sucrose concentration of 8-12%. Since coconut milk is known to contain "auxins" and substances which specifically stimulate cell divi-
sion (probably kinins) in the less completely identitied "active fraction" (Pollard, Shantz and Steward, 1961), this represents a parallel case where the requirement for growth factors has been satisfied by the high sucrose concentratioin.
At this stage of investigation, it is not certain whether the high concentration of sucrose in the absence of added growth factors exerts an osmotic effect, or whether it influences the metabolism of the cultured tissues in some other unknown way. Although the embryos develop better in a medium with growth substances plus 2% sucrose than do those cultured in high sucrose or high salt medium, our present evidence does not eliminate a role for sucrose in the in vitro development of these em- bryos as balancing the osmotic medium of the sap. It is probable that the activity or one component of the balanced hormonal control system is in turn controlled by the high sucrose or salt concentra- tion, perhaps through osmotic processes prevent- ing cell elongation, which determines the optimum balance between cell division and cell enlargement (cf. Adamson, 1962). It is further possible that culture of larger embryos on agar rather than in liquid affects in some way the osmotic contribu- tion of the nutrient medium.
Similarly, the role of high salt concentration in substituting for the added growth factors is not understood. In their ability to dispense with exog- enous precursors when grown in a medium with high salt, globular embryos of Capsella resemble the crown gall tumor tissue recently described by Wood and Braun (1961). They found that the normal and partially altered tumor cell types of Vinca rosea lost their requirement for glutamine, asparagine, cytidylic and guanylic acids, and inositol and glutamine, respectively, when grown in White's medium with increased inorganic salt levels. In the case of embryos, the growth factors were not limiting, since no enhancement in growth occurred in a medium with 12% sucrose or lOX salts to which adenine sulfate, kinetin and IAA were added in the usual proportions.
Although the embryos of a number of angio- sperms have been successfully grown in vitro, we have little knowledge of the physiological mecha- nism of organ initiation in cultured embryos. In the present experiments, even the smallest em- bryos cultured were potentially capable of pro- ducing a shoot and/or a root system characteristic of the adult plant. Complete development of the shoot system is slightly accelerated in light. In isolated embryos <l10001 long, a primary root system developed only in cultures grown in the dark, while longer embryos produced root systems in both light and dark. To explain this variation in embryos differing only in a few hundred mi- crons, it is suggested that mature embryos produce the necessary stimulus for the growth of the primary root system characteristic of the adult plant when grown in light or dark conditions. In younger embryos if cultured in light, the stimulus is lost or not formed in adequate amount.

July, 1963] RAGHAVAN AND TORREY-CAPSELLA EMBRYOS 551
LITERATURE CITED
ADAMSON, D. 1962. Expansion and division in auxin- treated plant cells. Canad. Jour. Bot. 40: 719-744.
BRACHET, J. 1950. Chemical embryology. Interscience Publishers, Inc., New York.
FAMINTZIN, A. 1879. Embryologische Studien. Mem. Acad. Imp. Sci. St. Petersburg. 10.
HANSTEIN, J. 1870. Entwicklungsgeschichte der Keime der Monokotyle und Dikotyle. Bot. Abhandl. Bonn. 1: 1-112.
MAUNEY, J. R. 1961. The culture in vitro of immature cotton embryos. Bot. Gaz. 122: 205-209.
MILLER, C. 0. 1961. Kinetin and related compounds in plant growth. Ann. Rev. Plant Physiol. 12: 395-408.
NAYLOR, A. W. 1952. Physiology of reproduction in plants, p. 259-300. In George S. Avery, Jr., [ed.], Survey of biological progress, Vol. II. Academic Press, Inc., New York.
NEEDHAM, J. 1942. Biochemistry and morphogenesis. University Press, Cambridge.
NORSTOG, K. 1961. The growth and differentiation of cultured barley embryos. Amer. Jour. Bot. 48: 876- 884.
POLLARD, J. K., E. M. SHANTZ, AND F. C. STEWARD. 1961. Hexitols in coconut milk: their role in nurture of dividing cells. Plant Physiol. 36: 492-501.
POLLOCK, E. G., AND W. A. JENSEN. 1961. A cyto- chemical analysis of embryogenesis in higher plants. (Abstr.) Amer. Jour. Bot. 48: 530. _-, AND -. 1962. DNA synthesis and patterns of cell division in developing higher plant embryos. (Abstr.) Amer. Jour. Bot. 49: 656.
RIETSEMA, J., S. SATINA, AND A. F. BLAKESLEE. 1953. The effect of sucrose on the growth of Datura stramo- nium embryos in vitro. Amer. Jour. Bot. 40: 538-545.
RIJVEN, A. H. G. C. 1952. In vitro studies on the em- bryo of Capsella bursa-pastoris. Acta Bot. Neerland. 1: 157-200.
- -. 1955. Effects of glutamine, asparagine and other related compounds of the in vitro growth of embryos of Capsella bursa-pastoris. Proe. Koninkl. Ned. Akad. Wetensch. C58: 368-376.
. 1956. Glutamine and asparagine as nitrogen sources for the growth of plant embryos in vitro: A comparative study of 12 species. Austral. Jour. Biol. Sci. 9: 511-527.
SCHAFFNER, M. 1906. The embryology of the Shep- herd's Purse. Ohio Naturl. 7: 1-8.
SNEDECOR, G. W. 1956. Statistical methods. The Iowa State College Press, Ames.
SOUEGES, E. C. 1914. Nouvelle recherches sur le developpement 1'embryon chez les Cruciferes. Ann. Sci. Nat. Bot., IX, 19: 311-339.
. 1919. Les premieres divisions de l'oeuf et les differenciations du suspenseur chez le Capsella bursa- pastoris Moench. Ann. Sci. Nat. Bot., X, 1: 1-28.
SWAMY, N. S. R. 1961. Experimental studies on female reproductive structures of Citrus mnicrocarpa Bunge. Phytomorphology 11: 109-127.
VAN OVERBEEK, J. 1942. Hormonal control of embryo and seedling. Cold Spring Harb. Symp. Quant. Biol. 10: 126-134. -, M. E. CONKLIN, AND A. F. BLAKESLEE. 1942. Cultivation in vitro of small Datura embryos. Amer. Jour. Bot. 29: 472-477.
, R. SIuI, AND A. J. HAAGEN-SMIT. 1944. Factors affecting the growth of Datura embryos in vitro. Amer. Jour. Bot. 31: 219-224.
V'EEN, H. 1961. The effect of gibberellic acid on the embryo growth of Capsella bursa-pastoris. Proc. Koninkl. Ned. Akad. Wetensch. C64: 79-85.
- -. 1962. Preliminary report on effects of kinetin on embryonic growth in vitro of Capsella embryos. Acta Bot. Neerland. 11: 228-229.
WARDLAW, C. W. 1955. Embryogenesis in plants. John Wiley & Sons, Inc., New York.
WOOD, H. N., AND A. C. BRAUN. 1961. Studies on the regulation of certain essential biosynthetic systems in normal and crown-gall tumor cells. Proc. Natl. Acad. Sci. 47: 1907-1913.
ZIEBUR, N. K., AND R. A. BRINK. 1951. The stimula- tive effect of Hordeum endosperms on the growth of immature plant embryos in vitro. Amer. Jour. Bot. 38: 253-256.












![Molecular specification of germ layers in vertebrate embryos...ducers in frog [19–22], fish [23], chick [24] and mouse embryos [25]. Blocking FGF function in these embryos leads](https://img.pdfslide.us/doc/110x75/60faba4f90a41b60861d0402/molecular-specification-of-germ-layers-in-vertebrate-embryos-ducers-in-frog.jpg)