Embed Size (px)
Citation preview

MEIT{YL GROI.IP METABOLISM IN SHEEP
A Thesis
sutrnitted in fulfilment of the requirements
for admission to the degree of
DOCTOR OF PHII,OSOPHY
of the University of Adelaide
by
Gang-Ping )tue
(An assisLant lecturer of Biochemístry
in frrejiang Agricultural University, China)
Department of Ani¡nal Sciences
t'laite Agricultural Research InsLitute
Ttre University of Melaide
South Australia
WAITE INSTITUTË2-1- 4.81
LIBRARY
Novernber, 1986
A¡'cuúta rcl:;l'B'l

]-
TABLE OF CONTENTS
TABLE OF CONTENTS
LIST OF FIGURES
LIST OF TABLES
DECI-ARATION
ACKNOI^ILEDGEMENTS
PUBLICATIONS
NOÌ"ÍENCULATT]RE AITD ABBREVIATIONS
SI]MMARY
CFIAPTER 1. GENRAL INTRODUCTION
CI]APTER 2. LITMATURE REVIETN
2.t Introduction
2.2 The diet as a source of methyl groups
2.3 Biosynthesis of the methyl group of methionine
2.4. ULlLization of methyl groups via AdoMet-dependent
transmethylation
2.4.1 Biosynthesis of AdoMet
2. 4 . 2. Q:anti tatively predominan t Adollet-dependenL
transmethylation reactions
2.4.2.t Methylation of guanidinoacetate
2.4.2.2 Methylation of phospLratidylethanolamine
2.4.2.3 M,ethylation of glycine
2.5. C-atabolism of the main methyl compounds
2.5.L Methionine catabolism
Page No.
iviii
X
xii
xiii
xiv
xvi
xviii
L
4
4
5
9
I4
T4
15
t7
18
22
24
24

l_t
Page No.
2.5.2 AdoMet breakdown via 5'-methylthioadenosine format.ion 29
2.5.3 Ctroline oxidation 32
2.6. Regulatory aspects of methyl group metabolism 34
2.6.t. Cellular levels of substrates and I(m values of
competing enzymes 35
2.6.L.\ Methionine levels and Km values of
methionine-utilizing enzymes 35
2.6.L.2 AdoMet levels and Km values of methyltransferases 37
2.6.I.3 Homocysteine levels and Km values of
homocysteine-utilizing enzymes 39
2.6.2 Modulation of transmethylation reactions by the ratio
of AdoMet to AdoHcy 4L
2.6.3 Effects of availability of pre.formed methyl compounds 42
2.6.4 Effects of insulin 48
2.7 Concluding remarks on literature review 49
CHAruM 3. I"ÍATM.IAI-S AND GENMAL METHODS
3.1. Ibterials
3.1.1 Animals
3.L.2 Radiochemicals
3.1.3 Biochemicals
3.L.4 Chrromatographic materials
3.1.5 Enzymes
3.2. General methods
3.2.t. Preparation and purificat.ion of radiochemicals
3.2.I.t Preparation of fmethyl-3H]betaine
3.2.L.2 Purification of [methyl-3H]choline
50
50
50
50
50
51
51
51
51
51
52

3.2.1.3 Purification of [methyl-3H]AdoMet
3.2.2. Preparations and assays of enzymes
3.2.2.t 5-MethylH4F-homocysteine methyltransferase
3.2.2.2 Betaine-homocysteine methyltransferase
3.2.2.3 Phospholipid methyltransferase
3.2.2.4 Guanidinoacetate methyJ-transferase
3.2.2.5 Glycine methyltransferase
3.2.2.6 Chroline oxidase
3.2.2.7 Cystathionine ß-synthase
3.2.2.8 Y -Cystathionase
3.2.2.9 Methylmalonyl-C.oA nmtase
3.2.3. Metabolite assays
3.2.3.t AdoMet and AdoHcy
3.2.3.2 Choline
3.2.3.3 Creatine
3.2.3.4 Creatinine
3.2.3.5 Cysteine
3.2.3.6 Formiminoglutamic acid
3.2.4 Determination of protein
CHAPTM. 4. CO,IPARATIVE STUDIES ON BIOSYNTT{ESIS AI{D UTILIZATION OF TI{E
MSIÏTTL GROUP OF METT{IONINE IN SI{EEP AI{D RAT TISSUES
4.L Introduction
4.2. Experimental proeedures
4.2.I Animals
4.2.2 Enzyme preparations and assays
t].t
Page No.
53
53
53
55
56
57
58
59
60
61
62
63
63
64
66
66
66
67
67
68
68
70
70
7t
71.4.2.3 Measurement of methionine recycling in vitro

lv
Page No.
4.2.4 Other assays 73
4.3. Results 73
4.3.1 C-ornparison of the specific activities of enzyrnes in
sheep and rat tissues 73
4.3.2 Cornparison of methionine recycling in sheep and rat
tissues
4.3.3 Effects of metabolites on methionine recycling in
liver slices
4.4 Discussion
CHAITIER 5. DEVH¡PMENTAL CHANGES IN TI{E ACTIVITIES OF ENZYMES REI-ATED
TO ME'IT{YL GROUP ME'TABOLISM IN SHEEP TISSUES
5.1 Introduction
5.2. ExPerimental Procedures
5.2.1 Animals
5.2.2 Ûlzyme preparations and assays
5.3. Results
5.3.1 Develognental changes of choline oxidase
5.3.2 Developxnental changes of homocysteine-utilizing
enzymes
5.3.3 Developxnental changes of MoMet-dependent
methyltransferases
5.4 Discussion
88
88
88
88
89
89
89
9I
91
97
CÉIAPTER 6. REGUTATION OF ME.THYL GROI.TP ME.TABOLISM IN TAGTATING EI47ES
77
80
80
6.1 Introduction
6.2. Experimental Procedures
104
to4
105

v
Page No.
6.2.t Animals 105
6.2.2 Determination of milk choline, creaLine
and creatinine 105
6.2.3 Enzyme preparations and assays 106
6.3. Resulrs 106
6.3.1 The contents of choline, creatine and creatinine in
e,wet s mitk 106
6.3.2 Effects of lactatj-on on the specific activities of
methyltransferases and cystathionine ß-synLhase 108
6.4 Discussion 108
CHAPTM. 7. DISTURBAI{CE OF METHYL GROI.IP METABOLISM
IN AI-I,OXAI{-DIABETIC SHEEP 113
7.1 Introduction 113
7.2. Experimental procedures 114
7 '2'L Animars 1,L4
7.2.2 Enzyme prepa.rations and assays 115
7.2.3 Determi-nation of cysteine in urine 115
7'3' Resurts 715
7.3.t Effects of alloxan-diabetes on the specific
activities of homocysteine methyltransferases 115
7.3.2 Effects of alloxan-diabetes on the specific
activities of AdoMet-dependent methyltransferases 115
7.3.3 Effects of alloxan-diabetes on the specific activities
of cystathionine B-synthase and Y-cystathionase 118
7.3.4 Urinary excreLion of cyst(e)ine before and after
alloxan treatment 118

vt.
Page No.
1187.4 Discussion
G{APTM, 8. PERTURBATION OF MET{IONINE ME'IABOLISM IN SHEEP I{ITIT
NITROUS-OXIDE-INDUCED INACTIVATION OF COBAIÁI'IIN
8.1 IntroducLion
8.2. Experimental procedures
8.2.L Animals
8.2.2 kepa.ration of tissues
8.2.3 &rzyme preparations and assays
8.2.4 Metabolite assays
8.3. Results
8.3.1 Effects of N2O on the specific activities of vitamin
Bl2-requiring enzyrnes and betaine-homocys teine
methyltransferase
8.3.2 Effects of N2O on AdoMet. and methionj-ne
concentrations
8.3.3 Effects of N2O on urinary excretion of
formiminoglutamic acid and homocystine
8.4 Discussion
cHApTm 9. QUAITTTTATTVE EVALUATTON AND REGLLATTON OF
S-ADENOSYLMEMIONINE-DEPNDNT TRANSME'TIMATION
IN SHEEP TISSUES
9.7 Introduction
9.2. Experimental procedures
9.2.L Animals
9.2.2 Measurement of Adotlcy and AdoMet levels in tissues
t23
L23
L24
t24
t24
t25
L25
t25
L25
127
r29
t29
1_35
135
L36
136
t37

Page No.
9.2.3 Measurement of transmethylation rate in vivo L3l
9.2.4 preparations and assays of methyltransferases 138
9.3. Results L3g
9.3.1 Tissue distribution of AdoHcy and Adol4et in sheep 139
9.3.2 Efficacy of POA to block AdoHcy hydrolysis 139
9.3.3 In vivo rate of transmethylation in tissues L42
9.3.4 Effects of simultaneous adrninistration of methionine
with POA on AdoMet. and AdoHcy levels 146
9.3.5 In vitro irùribition of methyltransferases from sheep
liver by AdoHcy L48
9.3.6 Kinetic constants of methyltransferases from sheep
v].a
148
150
Iiver
9.4 Discussion
CHAPTM, 10. GENERAL DISCUSSION
BIBLIOGRAPHY
160
L66

2
4
5
7
L
7
5.3
5.4
5.5
5.6
5.7
9.t
9.2
9.3
9.4
V].I].
LIST OF FIGURES
Page No.
Major rnetabolic pathways of methyl groups '6
Time course of methionine recycling in tissue slices 78
The specific activity of choline oxidase in sheep Iiver during
developxnent 90
the specific activity of betaine-homocysteine methyltransferase
in sheep tissues during develognent 92
The specific activity of 5-methylH4F-homocysteine
methyltransferase in sheep tissues during development. 93
The specific activity of cystathionine ß-synthase in sheep
tissues during developxnent 94
The specific activity of phospholipid methyrtransferase in sheep
liver during developxnent 95
The specific activity of glycine methyltransferase in sheep
tissues during developxnent 96
The specific actirrity of guanidinoacetate methyltransferase in
sheep tissues during developxnent. 98
Inhibition of tissue AdoHcy hydrolysis by POA 1,4I
Time course of AdoHcy and AdoMet. accunmration in tissues after
the injection of POA L43
Effects of sinmltaneous administration of methionine with poA on
AdoHcy and AdoMet levels in sheep tissues I47
Lineweaver-Burk plots for determination of kinetic parameters of
guanidinoacetaLe methyltransferase from sheep liver I52
Lineweaver-Burk plots for determination of kinetic parameters of
glycine methyltransferase from sheep liver 153
5.2
9.5

ax
9
Page No.
6 Lineweaver-Burk plot. for determination of kinetic parameters of
phospholipid methyltransferase from sheep liver L54

4.L
4.2
4.3
4.4
4.5
6.t
6.2
6.3
7.t
7.2
7.3
8.1
8.2
8.3
LIST OF TABLES
Ttre specific activities of homocysteine methyltransferases
in sheep and rat tissues
The specific activities of AdoMet-dependent methyltransferases
and choline oxidase in sheep and rat tissues
C.omparison of methionine recycling ín the tissue slices of
sheep and rats
Effects of metabolites on methionine recycling in liver slices
Ratios of the specific activities of hepatic enzymes of
rats to sheep
The contents of choline, creatine and creatinine in ewe's milk
The specific activities of AdoMet-dependent methyltransferases
in the tissues of lactating and non-Iactating ewes
TLre specific activities of homocysteine-utilizing enzymes in
the liver of lactating and non-lactating ewes
The specific activities of homocysteine methyltransferases in
normal and alloxan-diabetic sheep liver
The specific activities of AdoMet-dependent methyltransferases
in norrnal and alloxan-diabet.ic sheep tissues
The specific activities of cystathionine S-synthase and
Y-cystathionase in normal and alloxan-diabetic sheep liver
The effect of N20 on the specific activities of vitamin
Bl2-requiring enzymes in sheep tissues
The effect of N2O on AdoMet concentrations in sheep tissues
Urinary excreLion of formiminoglutamic acid and homocystine in
sheep before and after N2O exposure
x
Page No.
l4
76
79
81
83
L07
109
LL6
LL7
719
L26
L28
110
130

9.L
9.2
9.3
9.4
Distribr:tion of AdoHcy and AdoMet in sheep tissues
Tentative estimation of transmethylation rates and AdoMet
turnover times in sheep and rat tissues
In vitro irùribition of methyltransferases from sheep liver
by AdoHc,y
Some kinetic constants of methyltransferases from sheep liver
x1
Page No.
L40
744
749
151

xii
DECIARATION
I hereby declare that this thesis does not contain any material
previously sutxnitted for the award of any other degree or diploma in any
university and thaL, to the best of my lcrowledge and bel-ief , the thesis
contains no material previously published or written by another person,
except v¡here due reference is made in Ehe text
I consent to this thesis being made available for photocopying and
loan if accepted for the award of the degree.
Gang- Xue

xt_11-
ACKNOI,ÍLEDGEI\4ENTIS
I acknowledge with deepest gratitude the invaluable advice,
insighLs and encouragement given throughout the course of this
work by my supervisor, Dr. A. M. Snoswell, Reader in Animal Sciences
(Biochemistry).
I or¿e a great. debt to my supen¡isor, Prof . B. P. Setchell for
his supporL, advice and hetp duríng various phases of this work.
I am extremely grateful to l,lr. R. c. Fishlock for help in
collection of sample, some technical assistance and critical reading of
the manuscript of this thesis.
The help of Dr. J. C. L. Mamo and l\tr. M. J. Snoswell in the
induction of diabetic sheep and collection of samples ís greatly
appreciaLed, and Dr. B. S. Robinson is also thanked for cheerful
companionship.
Many thanks go to Prof. D. J. D. Nicholas for his encouragement and
help.
I would like to Lhank Dr. I^1. B. Runciman for his advice on the
experiment of NZO exposure and the provision of the experimental
facilities and Mr. D. Boehm for help in analysis of amino acids.
I am indebted to },lr. P. E. Geytenbeek for his help in providing
experimental sheep at various ages and to I'tr. R. P. Fels and Mr- A. B.
Sanderson for maintenance and slaughter of the sheep.
Special thanks go to my wife, Xiaoming, wtro gave me encouragementt
love and undersLanding, and to my parents for their support and
inspiration.
Ttre financial support of tlniversity of Adelaide Postgraduate
Research Scholarship is gratefully acknowledged.

XIV
PUBLICATIONS
Part of the work presented in this thesis has been published:
Research papers
)fue, G. P. and Snoswell, A. M. (1935) Cornparative studies on the
methionine synthesis in sheep and rat tissues. C,ornp. Biochem.
Physiol. 808, 489-494.
Xue, G. p. and Snoswell, A. M. (1985) Disturbance of methyl group
metabolism in alloxan-diabetic sheep. Biochem. Int. 10, 897-905.
Xue, G. P. and
metabolism
Snoswell, A. M. (1985)
in lactating ewes.
Regulation of methyl grouP
Bioche¡n. Int. 11, 381-385.
Xue, G. P. and Snoswell, A. 1"1. (fg8$) Developxnental ctnnges in the
activities of enzymes related to methyl group metabolism in sheep
tissues. Cornp. Biochem. Physiol. 838' tL5-L20.
){¡e, G. P., Snoswell, A. l"l. and Runciman, I,l. B. (fOa6¡ Perturbation of
methionine metabolism in sheep with nitrous-oxide-induced
inactivation of cobalamin. Biochem. Int - 12, 6L-69.
)fue, G. P. and Snoswell, A. M. (1986) Quantitative evaluation and
regulation of S-adenosylmethionine-dependent transmethylation in
sheep tissues. Comp. Biochem- Physiol. in press.

XV
Abstracts
Snoswell, A. M. and Xue, G. P. (1985) The effects of alloxan-diabetes
on transmethylat.ion and transsulfuration reactj-ons in sheep
tissues. Proc. lr¡rst. Biochem. Soc. 17, L02.
Snoswe1l, A. M. and X:e, G. P. (1985) CIeanges in enzymes related to
methyl group metaboU-sm in preruminant versus ruminant lambs.
XIII International Congress of Nutrition (Brighton, U.K.), 50.
)fue , G. P. and Snoswell, A. M. (fggS) Effects of nitrous-oxide-induced
vitamin B12 deficiency on methionine and one-carbon metabolism in
sheep. Proc. NuLr. Soc. Aust. 10, 168.
Snoswell, A. M. and Xue, G. P. (fOaA¡ Rates and regulation of
transmethylation via S-adenosylmethionine in sheep tissues.
koc. drst. Biochem. Soc. 18, 92.

XVI
NOMM{CIATIJRE AND ABBREVIATIONS
The reconrnendations of the NomenclaLure Conrnittee of the
International Llnion of Biochemistry (1978) on the nornenclature and
classification of enzyrlìes have been followed as far as possible.
Acetylsero tonin methyltras f erase
S-Adenosylhomocys teine hydrolase
S-Adenosylmethionine cyclotrans f erase
S -Adenosylme thionine decarboxylas e
Arginine-glycine amidinotransf erase
Asparagine amino trans f erase
Betaine aldehyde dehydrogenase
Betaine-homocys teine methylLrans ferase
Catechol methyltransf erase
Choline dehydrogenase
Choline oxidase
r-Cystathionase
CysLathionine ß-synthr,ase
DNA methyltransferase
Glutamine aminotransferase
Glycine methyltransferase
Guanidinoaceta te methyltrans f eras e
His tamine methyltransf erase
His tidine aminotransferase
Hj-stone methyltransferase
I-eucine aminotransferase
Methionine adenosyltransferase
EC 2.t.I.4
EC 3.3.1.1
EC 2.5.L.4
EC 4.1.1.50
Eß 2.t.4.L
EC 2.6.t.L4
EC 1.2.1.8
EC 2.1.1.5
EC 2.7.t.6
EC 1.1 .99.L
EC 1.L.3.L7
EC 4.4.1.1
EC 4.2.t.22
EC 2.t.L.31
EC 2.6.1.15
EC 2.t.I.20
EC 2.t.1-.2
EC 2.1 .1 .8
EC 2.6.1.38
EC 2.L.t.42
EC 2.6.1.6
EC 2.5.1 .6

5, lO-Methylenetetrahydrofolate reductase
5-Methyltetrahydrofolate-homocys teine methyltrans ferase
Fhenyle thanolamine me thy I tran s f eras e
Pho sptr,a tidy Ie thano lamine me thy I tran s f era s e
Protein-carboxyl methyl trans f erase
Protein- lys ine methyltrans f erase
IRNA methyltransferases
XVI1
EC 1.1.99.15
Eß 2.1.1.13
EC 2. L.I.2B
EC 2.L.T.L7
EC 2.7.L.24
FC 2.1.I.43
EC 2.L.t.29-36
those
7L5,
Ttre abbreviations for chemicals, syrnbols and units follow
approved by Biochimica et Biophysica Acta [Biochim Biophys. Acta
L-23, (fOaZ¡;, except the following listed abbreviations:
AdoMet.
AdoHcy
DTI
H+F
5-MethyIH4F
MIA
POA
PtdCho
PtdEtn
PtdMeEtn
PtdMe2Etn
SEM
Trc
U.V.
S-adenosylmethionine
S-adenosylhomocys teine
dithiothreitol
tetrahydrofolate
5-me thy I te trahydrof olate
5' -methylthioadenosine
periodate-oxidized adenosine
phosphatidylcholine
pho s pha t idy Ie thano lamine
pho spha t idy Imonomethyle thano lamine
pho sphat idy Idime thylethanolamine
standard error of mean
thin layer chromatography
ultraviolet.

XVIII
SLll,ll',1ARY
Methyl group metabolism was investigated in sheep to elucidate
the mechanisms of metabolic adaptat.ion to a very low intake of methyl
nutrients in the post-ruminant staLe, due to the almosL complete
degradation of dietary choline by rumen microbes, the lack of dietary
creaLine and the relatively low content of methionine in microbial
protein.
The tissue capacities for the biosynthesis and utilization of the
methyl group of methionine were compared between adult sheep and rats.
The study reveals that the important features of the enzymatic patterns
of methionine synthesis in adult sheep were the small capacity of
betaine-homocysteine methyltransferase accompanied with the
great capacity of 5-methyltetrahydrofolate (5-methylH4F)-homocysteine
methyltransferase, in comparison to those of rats. TÏre specific
activit.ies of choline oxidase and glycine methyltransferase, viLrich are
enzymes involved in the catabolism of methyl groups, were 27 and 284
times lower in sheep liver than in rat liverr respectively. The
enzymatic capacities for the synthesis of creatine and choline in sheep
liver were lower than those of rat liver per unit of t,issue weight, but
the differences between the two species were relatively small. In vitro
overall rates of the reactions involved in methionine recycling in sheep
liver and kidney r^¡ere also lower than those in lhe corresponding tissues
of rats. The slow rate of methionine recycling in sheep liver
may be ascribed mainly to the very low levels of glycine
methyltrans f erase and betaine-homocys teine methyltrans f erase .
A study on the developnental changes in lhe specific activities of
these enzymes showed that the specific activities of choline oxidase and

xlx
betaine-homocysteine meLhyltransferase in sheep liver decreased from the
pre-ruminant to ruminant state aS a result of changes in the
availability of dietary choline. The rate of the synthesis of the
methyl group of methionine from 5-methylH4F in sheep liver may be
influenced by the reaction of homocysteine methylation from betaine, as
indicated by an inverse relationship between Ehe specific
activities of 5-methytH4F-homocysteine methyltransferase and
betaine-homocysteine methyltransferase during develoçxnent. Tkre specific
activity of hepatic glycine methyltransferase in newborn lambs increased
markedly from an extre¡nely low level in the fetuses ancl thereafter
decreased dramatically with age. This is in contrast to the
develognental pattern of the enzyme as observed in rals and rabbits,
vihere lhe specific activiLy of the enzyme continuously increases with
age until the adult level is reached. A marked developxnental increase
in the specific acLivity of phospholipid methyltransferase \¡ras observed
in Iambs a! the early postnatal age, but there was little change in
the specific activity of the enzyme from Èhe pre-ruminant to ruminant
state. No significant changes in the specific activiÈies of
guanidinoacetate methyltransferase and cystathionine ß-synLhase hlere
obsen¡ed in sheep liver during develoçxnent.
There are some unique patterns of the tissue distribution of these
enzymes in sheep. The specific activities of betaine-homocysteine,
glycine and guanidinoacetate mebhyltransferases in adult sheep Pancreas
were nn:ch higher than those in the liver. Develognental changes of the
specific activities of these methyltransferases are not parallel to
those of the hepatic enzymes. The specific activities of these
pancreatic methylÈransferases increased dramatically with age until Ehe
adult levels were reached.

xx
The requirement for methyl groups is higher during lactation.
l-actating e\^/es secreted approximately 10 milli-equivalents of methyl
groups in the forms of choline, creatine, creatinine and carnitine into
milk daily, which at least equals the reported values for the daily
output of methionine from the gut of non-lactating e\^Ies. This exlra
demand for methyl groups may be met by the elevated capacity for
the de novo synthesis of the methyl group of methionine via the
5-meLhylH4F-homocysteine methyltransferase reaction and a reduced
expenditure of the methyl group of S-adenosylmethionine (¡¿ol'tet) for
the nonessential methylation reaction caLaLyzed by glycine
methyltransferase. The level of hepatic phospholipid methyltransferase
\^/as elevated in lactating e\ÁJes. No significant dif ferences were
observed in the specific activities of hepatic guanidinoacetate
methyltransferase, betaine-homocysteine methyhransferase and
cystathionine ß-synthase between lactating and non-lactating e\Áles.
Severe alloxan-diabetes led Lo the reduced activities of
phospholipid methyltransferase and betaine-homocysteine
methyltransferase in sheep liver with no significant decrease of
5-methylH4F-homocysteine methyltransferase. An elevated rate of
methionine catabolism tikely occurred in severely diabetic sheep, as
indicated by a 65-fold j-ncrease in the specific activity of hepatic
glycine methyltransferase and a 4-fold rise in the specific activity
of y-cystathionase with the elevated uri-nary excretion of cyst(e)ine.
A block of methionine synthesis from the 5-methylH4F-homocysteine
methyltransferase reaction in sheep by exposure to nilrous oxide caused
marked decreases in the levels of plasma methionine and hepat.ic
AdoMet. There hTas no stimulaLion in the activity of betaine-homocysteine
methyltransferase to remove the surplus of homocysleine, vùren

XXI
5-methylH4F-homocysteine methyltransferase was strongly inactivated in
sheep tissues by nitrous oxide. Homocystinuria thus occurred in
nitrous oxide-exposed sheep. However, these nitrous oxide-induced
alterations are not often observed in non-ruminanL species as reported
by other investigators.
The overall raEe df Rdot'tet-dependent transmethylation was evaluated
in sheep tissues by measuring the init.ial rate of the accumulation of
S-adenosylhomocysteine (AdoHcy) plus AdoMet after blocking hydrolysis of
AdoHcy with a potent irùribitor, periodate-oxidized adenosine. A mininn:m
estimate of the Èotal requirernent of AdoMet for transmethylation in
50-kg sheep \.¡as at least 18 nmoles per day, virich is 2-4 times the
reported value for the daily output. of methionine from the gut. Sheep
liver possessed approximately 75 7. of Lhe total-body capacity of AdoMet
utilization for transmethylation. Methionine loading markedly elevaLed
the hepatic and pancreatic AdoMet levels and stimulated the rate of the
transfer of methyl groups from AdoMet in these tissues. Changes in
AdoMet and AdoHcy levels after methionine loading in other tissues of
sheep were relatively snnll, if they occurred at aII. The I(m values of
phospholipid methyltransferase and glycine methyltransferase from sheep
Iiver for AdoÞlet were, in the neighbourhood of its physiological
concentration, .bJt the Km of guanidinoacetate methyltransferase for
AdoMet was well below the physiological level. These meÈhyltransferases
from sheep liver were subject to strong irùribition by AdoHcy and a
certain degree of the irùribition occurred even at the physiological
concentrations.
From these findings, it is suggested that sheep obtain a
substantial amount of methyl groups from the de novo synthesis of methyl
groups and rely more heavily on the 5-methylH.F-homocysteine

XXII
methyltransferase reaction for the supply of methionine methyl than
most, if not aII, non-ruminant species. The extremely low enzymatic
capacities for the oxidat.ion of choline and the expenditure of the
methyl group of AdoMet for the nonessential methylation reaction,
glycine methylation, in adult sheep serf/e as a means for the
conservation of labile methyl groups. This greatly reduces the
requirement for labile methyl groups in this species. Thus, a low rate
of methyl group catabolism coupled r¿ith a high capacity of the de novo
slmthesis of methyl groups ensures that ruminant sheep are able to
survive successfully on a very low intake of methyl nutrients.

wÀìi! l|'l:jî¡-iljTIlrÈíÌ AR'l
f2
íT
ET
a
T
I L
CHAPTER 1. GENERAL INTRODUCTION
Methyl group metabolism is closely associated with methionine
metabolism, for not only does methionine act as a methyl donor for many
important transmethylation reactions via AdoMet, but also the reactions
involved in methionine methyl synthesis represent the routes for either
lhe recycling or the de novo synthesis of methyl grouPs. One becomes
increasingly aware thaL the amount of methionine required for
transmethylation reacLions is apparently in excess of its dietary intake
and ít is necessary to conserve homocysteine for the regeneration of
methionine (uu¿¿ and Poole, Lg75; l"h:dd et al., 1980). Methyl groups
required for homocysteine methylation are mainly derived either from one
of the methyl grouPs of betainer a product of choline oxidation, or from
5-methyltl4F, which is derived from one-carbon sources' The latter route
is a pathway f.or the de novo synthesis of methyl groups.
It is generally considered that predominant transmethylat'ion
reactions in the utilization of AdoMet in manrnalian tissues are those
involved in the synthesis of creatine, choline and sarcosine (Uuaa et
aL., 1980; Dawson et a1., 1981; Mitchell and Benevenga, L976;Ogawa and
Fujioka, 1982b). It is known that methionine, choline and creatíne
are predominant forms of preformed methyl nutrients in normal diets for
humans and non-ruminant animals and constitute the major body-pool of
methyl groups in manrnals (Ur:¿¿ and Poole, L975; Zeisel' 1981; Crim et
aL., L976). The utilizalion of methyl groups via AdoMet-dependent
transmethylation is thus balanced either by dietary intakes or by the
de novo synthesis of methyl grouPs from one-carbon sources.
In most manmals, dietary methyl compounds provide nust of the body
requirement for methyl groups. For example, in young adult humans on

2
normal diets, 70-80 7. of methyl groups utilized via AdoMeL-dependent
transmethylation are derived from dietary Iabile methyl contpounds,
methionine and choline (fq¡¿¿ and Poole, 1975; Itudd et al., 1930). l']nlike
most monogastrics, sheep derive a relat.ively small amount of methyl
groups frorn the diet, owing to the almost cornplete degradation of
dietary choline by rumen microbes (neitt et al. , L979; Dawson et al.,
1931) and the lack of creatine in planLs or microorganisms (t^ialker,
tgTg). Therefore, choline and creatine in sheep tissues must originaLe
entirely from their synthesis at the expense of the methyl group of
methionine. As estimated by l',ludd (1930), creatine synthesis in hurnans
consumes about 70 7" of toLa1 AdoMet formed under' nornnl dietary
condit.ions. Dawson et al. (1981) reported that the amount of methionine
methyl required for choline synthesis in adult sheep aPPears to be ntzlny
times greater than methionine supply from the gut. Thus, the
requirement for the methyl group of methionine must be quite large in
sheep. It has been deduced from nutritional experiments thaÈ methionine
is the firsr-limiting amino acid in sheep (ctr,alupa, L972; Schelling et
al., L973; Barry et a1., t973). Such a Poor nutritional condition of
methyl grouPs as sheep encounter would certainly lead to severe
pathological lesions and even death in many non-ruminant species, as
choline deficiency produces such problems in rnâny species (Kuksis and
Mookerjea , Lg78; Griffith et al. '
t97t).
The question is thus raised: how can sheep survive successfully
on such a low intake of methyl groups? Although some regulatory
mechanisms for the control of methyl grouP metabolism under various
nutritional conditions of methyl groups in hu¡nans and some non-ruminant
n¡anmals have been elucidated (these are presented in Literature Review),
regulatory aspects of meLhyl grouP melabolism in sheep are virtually

3
unkro\,.n. Presumably, the regulatory mechanisms observed in other
manrnalian systems are also likely to occur in sheep. Therefore, it is
Iikely ttrat the patterns of the flux of methyl conrpounds through the
various metabolic pathways of methyl groups in sheep may differ from
those in most non-ruminant species.
In order to elucidate the main features and regulation of methyl
group metabolism in sheep, the investigations presented in the thesis
have been organized to:
1) reveal the mechanisms of metabolic adaptation of sheep to the
low intake of methyl nutrienLs;
2) elucidate how diverse pathways of methyl group metabolism are
regulated under the influence of changes of nutritional and
physiological conditions to achieve methyl balances; and
3) gain quantitative lcrowledge of methyl group metabolism in this
species.

4
CFTAPTER 2. LITMATURE REVIE\,I
2.1 Introduction
A large ¡r-unber of biological cornpounds contain methyl groups, lùrich
are usually attached to nitrogen, sulfur, oxygen and carbon atoms'
Methyl groups in these biological compounds are not metabolically
equivalent, so that methyl compounds nay bre classified into two groups:
a metabolically labile group vilrich c¿n functiori biologically as methyl
donors; and a metabolically stable group. The former Sroup includes
AdoMet, meLhionine, 5-methylH4F, and one of the methyl groups of choline
or betaine. The latter group includes numerous cornpounds vùrich can be
synthesized from the methyl group of AdoMet via transmethylation- The
labile methyl groups may be recycled through the metabolic pathways of
methyl groups, but they are eventually either utilized by
transmethylation for the synthesis of stable methyl cornpounds or
directly catabolized.
In maÍïnals, labile methyl cornpounds can be obtained from diet
mainly in the forms of methionine and choline. However, mammalian cells
are also able to synthesize labile methyl groups in the form of
5-methylH4F from one-carbon sources. Among methyl cornpounds, methionine
is the most important and plays a central role in methyl group
metabolism. After activation to AdoMet, it acts as a methyl donor to a
wide variety of acceptors. Methionine may, in turn, be regenerated by
homocysteine methylation in vùrich either betaine or 5-methylH4F serves
as a methyl donor. Homocysteine may be further catabolized via the
transsul-furation pathway and its potency for methionine synthesis is
thus lost. Methionine nny also undergo transamination , through the

5
pathrday of v¡hich methionine methyl is directly oxidized. The overall
metabolic pathways are presented in Fi.g. 2't '
This review will discribe nutrition, biosynthesis, utilization,
catabolism and regulation of a group of methyl cornpounds v¡l'rich are
either metabolically irnportant in methyl metabolism or qrranti tatively
predominant in marrnalian cells, as all the biological reactions involved
in methyl group metabolism are too extensive to be covered in the length
of thesis review. The pathways, as shown in Fig. 2.I, of quantitaLive
irnportance in methyl group metabolism and their nutritional regulation
\^¡ill be stressed. Information on the regulation of the various
metabolic pathways of methyl compounds will be interlinked and presented
in the last section of the review. The review covers lcrowledge of the
subject obtained from ma¡rnnals available in literature up to January of
tg84, vùren this study \47as conmenced, and highlights the aspects related
to the scoPe of the study in this thesisr âs described in General
Introduction.
2.2. Tlr- Diet as a Source of Methyl Groups
Dietary sources of methyl conpounds for most of manmals are
predominantly present in the forms of methionine, choline and creatine.
The former two conrpounds are particularly important as their methyl
groups can be utilized as methyl donors for Lhe synthesis of other
methyl compounds. However, dietary supplemenL of creatine will reduce
its own synthesis at the expense of labile methyl groups (Walker , 1979).
In neonatal nutrition, carnitine may also be considered as one of major
methyl compounds in the diet, sj-nce milk contains the relatively high
content of carnitine (Snoswell and Linzell, L975) '
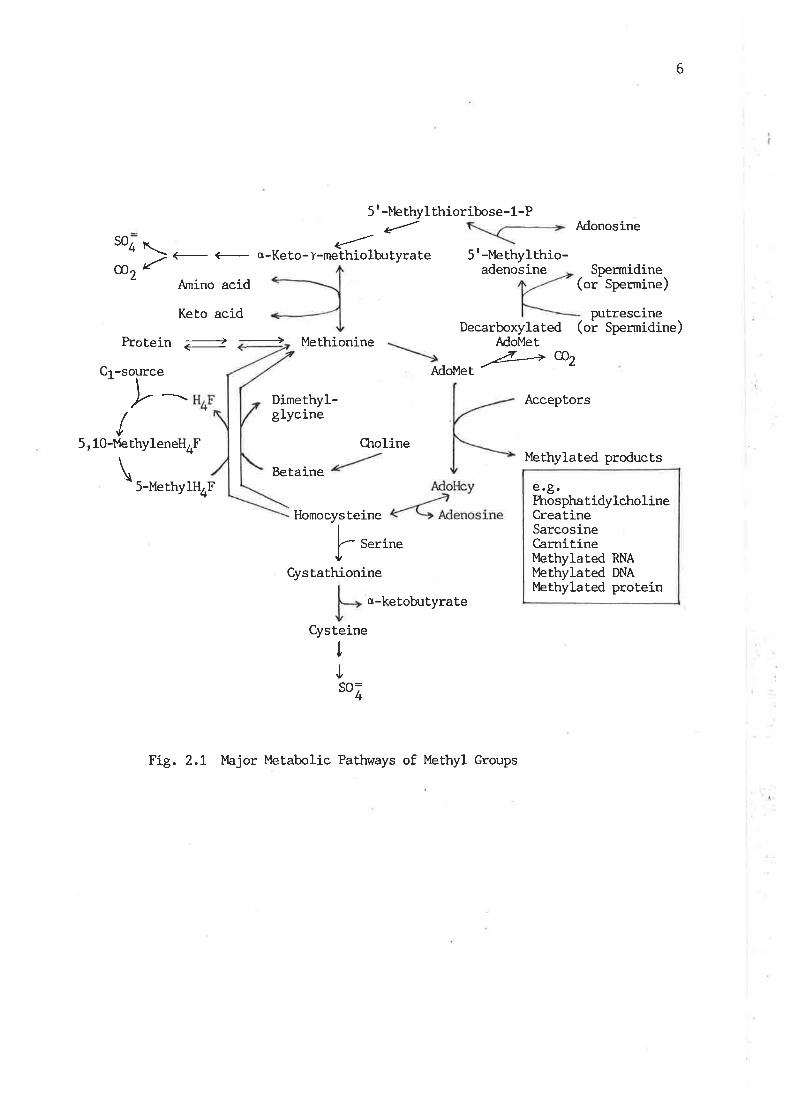
6
5'-t'tethyl¿
"'c -Ke to- Y-methiolbu tyrate
Anino acid
Keto acid
Methionine
Dimethyl-glycine
thioribose-1-P
5'-Methylthio-adenosine
Decarboxylated (Adol4et
AdoMet
AdonosíneSO
co
4\t-
2 Spermidine(or Spermine)
puÈrescineor Spermidine)
Protein <----)
/v5,10-MethyleneH4F
\5-MethylH4F
Betaine
Homocysteine
Choline
a-ketobuÈyrate
/Z----'+ CÐ2
Acceptors
Methylated products
e.g.PhosphatidylcholineCreatíneSarcosineCarnitineMethylated RNAI4,ethylated DNAlbthylated proLein
Cr -source
þ s.'ir'"CysEathionine
teine
Fig. 2.1 }bjor Metabolic Pathways of Methyl Groups
cys
4
IJSO

7
AIl manmals have an absolute requirement for methionine, because it
not only acts as a methyl donor afterconversion to AdoMet, but also is
an essential amino acid for protein synthesis as the homocysteine moiety
of methionine can not be synthesized in ma¡rmalian cells. Methionine
intake in adult humans on equilibration diets has been reporLed to be
8-11 nrnoles per day (Orr and I,latt, 1959; Ifudd and Poole, L975). In
ruminant animals, exogenous methionine is not directly derived from
dietary protei-n, but largely from microbial protein passing from the
rumen. The methionine contents of microbial protein are generally lo¡¡er
than those of tissue protein (Ørskov, L982). The output of methionine
from the gut in adult sheep is about. 4.5 mmoles per day from the study
of l,lolff et. al. (tglZ) and ranges from 5 Lo L2 nrnoles per day as
reported by Lindsay GggZ).
Dietary requirement for choline is influenced by the content of
methionine in the diet. Unlike methionine, all parts of the choline
molecule can be slmthesized in manrnalian cells and the limiting factor
for choline slmthesis is likely to be the labile methyl source.
Supplements of excess methionine in the diet certainly reduce choline
requirements. The choline requirements for non-ruminant species such as
rats, mice, guinea pigs, hamsters, dogs and pigs have been found to be
at least 0.1 7" of the total dietary intake of dry matter (National
Research Council, t974, !978, 1979). However, most of conmercial diets
for rats contain about 0.2 7" choline. In hunans, a conservative
estimation of total choline intake is about 300 mg per day, with an egg
and liver connoisseur ingesting more than three times this amount
(Zeisel, 1981). Experimental choline deficiency, vùrich leads to severe
pathological lesions and even death, has been produced in rnany
non-ruminant animals (Criffith et aL.,L97l; Kuksis and Mookerjea, 1978).

8
There are no reports concerning choline requirement. for sheep and other
ruminant, animals. However, choline deficiency has been reported in
pre-ruminant calves fed an artificial milk replacer devoid of choline
(Joknrson et al., I95L), but, has never been documented for sheep.
In contrast to choline nutrition of non-ruminant animalsr ân adult
sheep recieves no more Lhan 20-25 mg choline from the digesta passing
into the abomasum, owing to the almost complete degradation of dietary
choline by rumen microbes (Neitt et. al.,1979; Dawson et al., 1981). As
pointed out by Neill et, al.(t979), this is some f.ifty t.imes less, on a
body-weight. basis, than the minimum intake required to avoid
pathological lesions in a species sensit.ive to a low choline intake.
It should be noted that, in the diet of many herbivora, there may
exist a significant amount of betaine, a labile methyl compound.
However, the fate of betaine in sheep rumen is the Same as that of
choline (tlitchell, et al. , 1979).
Creatine is not. an essential nutrient, for manrnalsr âs it can be
synthesized in vivo as long as labile methyl groups are provided.
Creatine is abundantly present i-n most of vertebrate tissues, br:t has
not been found in plants or mj-croorganisms (I,{alker, L979). Therefore,
carnivora and onnivora ingest a considerable amount. of creatine from the
diet. For example, a moderate amount of creatine intake in humans is
near 1 g per day, though this varies widely (tykken et al.r 1980).
However, there is no creatine in the diet of herbivora. Thus, the amount
of creatine lost has to be replaced entirely by its synthesis at the
expense of labile methyl groups.
I¿bile methyl group requirement is higher in growing, pregnant and
lactating aninnls, since extra methyl groups are needed for the
synthesis of new Lissues or for milk produclion. In most of manrnals,
such extra dennnds for methyl groups may be met simply by relatively

9
higher dietary intake. However, this is not the case for post-rt¡ninant
sheep, as sheep derive no significant amounts of choline or creatine
from the diet even though amounts of the diet ingested is relatively
high. Howeverr Do sÈudies have been conducted to exami-ne regulatory
mechanisms of methyl group metabolism in these physiological states.
2.3. Biosynthesis of the Methyl Group of Methionine
Tkre concept that manrnals can synthesize the methyl moiety of
methionine by methylation of homocysteine arose from early nutrit.ional
experiments. \^lhite and Beactr (L937) first observed that, in certain
condiLions, homocysteine could support the growÈh of rats when
methionine \Àras lacking in the diet. A later surge of investigations into
methyl sources for homocysteine methylation leads to the present
knowledge that there are three homocysteine methylation reactions
utilizing naturally occurri-ng methyl donors in manrnals, but only tr¿o of
these reactions yield the net slmthesis of methionine (Finkelstein,
1970) r âs the third enzyme only transfers methyl groups from AdoMet to
homocysteine (Shapiro and Yphantis, 1959). Ttre direct methyl donors in
the two synthetic reactj-ons are beÈaine and 5-methylFlrF.
Du Vigneaud et al.(1939) first demonstrated that betaine or choline
could support the growth of rats vihen dietary methionine was replaced by
homocysteine. In vitro studies on transmethylation from the methyl group
of betaine or choline to homocysteine was first studied in liver
preparations by Borsook and Dubnoff. (L947). But choline is not, a direct
methyl donor and has Èo be converted to betaine by oxidation of its
alcohol group before the methyl transfer can occur (tntr:ntz, 1950) . The
methylation reaction is represented as follows:
Homocysteine * Betaine --+ Methionine + Dimethylglycíne

10
The reaction is catalized by betaine-homocysteine methyltransferase. In
horse and raL liver, there are two separable enzymes catalyzing this
reaction (Xtee et al., 1961). Awad et al. (1983) has demonstrated that
the methyl transfer in this enzyme reaction occurs directly from one
substrate to the other.
Betaine-homocysLeine methyltransferase was found to exist in all
species of mammals examined, but is primarily confined to the liver
(Ericson, 1960). The specific activity of the enzyme in rat kidney,
pancreas, spleen, adrenal, tesLes and adipose tissue is less than 1 7" of
that of the hepatic enzyme and was not detected in other tissues
investigated (Finkelstein eL aI., t97L). However, the specific activity
of the enzyme in human kidney is slightly higher than that in the liver
(¡ru¿¿ et al., t969r 7970). Itre specific actiwity of the enz)¡me decreases
slightly with age in rat liver (ninkelstein et aI., !97L) and is about
four-fold lower in human fetal liver than in the neonatal U-ver, but
there are no further changes with age (Sturman and Gaull , L978).
The de novo synthesis of methionine methyl was inferred in t94L,
vfrren du Vigneaud et al. (1941) found that' in rats given
deuterium-containing waLer, deuterium was incorporat.ed into the methyl
groups of choline. Requirement of folate and vitamin 812 in the de novo
synthesis of methionine methyl was discovered in 1949, vùten a nunber of
investigaLors found thât these two compounds could replace the preformed
methyl groups in a homocysteine diet for growth (reviewed by }4artin,
L975). Thereafter, many investigations revealed that the methyl sources
for the de novo synthesis of methionine methyl originaLe from a variety
of one-carbon units, such as fornnte, formaldehyde, the a-carbon of
glycine, the B-carbon of serj-ne and the formimino group of histidine
(reviewed by Greenberg, !963; Ifudd and C,antoni, 1964). These one-carbon
LI
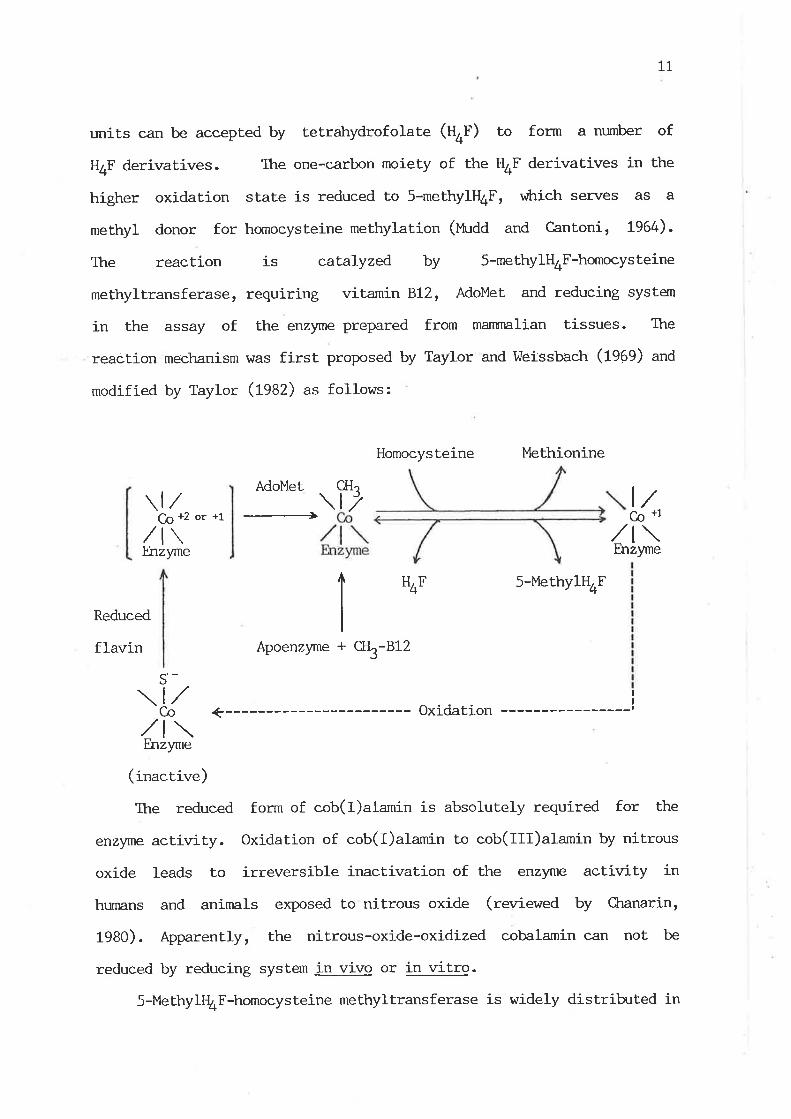
units can bre accepted by tetrahydrofolate (U+f) to form a nurnber of
H4F derivatives. Ttre one-carbon moiety of the H4F derivatives in the
higher oxidation state is reduced to 5-methyltl4F, vfiich serves as a
methyl donor for homocysteine methylation (Uudd and Cantoni, 1964).
The reaction is catalyzed by 5-methy1H4F-homocysteine
methyltransferase, requiring vitamin B12, AdoMet and reducing system
in the assay of the enzyme prepared from manrnalian tissues. The
reaction mechanism was first proposed by Taylor and l^leissbach (l9eg) an¿
modif ied by Taylor (ßAZ) as follows:
Homocysteine Methionine
Co +2 or +1/t\Enzyrne
CH.
\17
+
Adol'{et
--+
ltco *1,/l\
Enzyme
H+F 5-MethyIHOF
Reduced
flavin Apoenzyne ct]3-812
II
,/l\Fnzyme
(inactive)
The reduced form of cob(I)alamin j-s absolutely required for the
enzyme activity. Oxidation of cob(I)alamin to cob(Ill)alamin by nitrous
oxide leads to irreversible inactivation of the enzyme activity in
humans and animals exposed to nitrous oxide (rewiewed by Chanarin,
1980). Apparently, the nj-trous-oxide-oxidized cobalamin can not be
reduced by reducing system in vivo or in vitro.
5-MethylQ*F-homocysteine methylLransferase is widely distrib:ted in
s'-
Co

t2
marrnalian tissues (reviewed by Taylor, 1982). The enzyme activity in
rat tissues is the highest in the kidney, wtrere the specific activity is
about 4 times that of the hepatic enzyme (ninkelstein et al., l97t),
and the specific act.ivity of the enzyrne in human kidney is also higher
than that in the liver (¡,tu¿¿ er aI., 1969). The disrribution of
5-methylH4F-homocysteine methyltransferase has also been investigated in
various bovine tissues (l4angum et al., 1972). In contrast to rats and
humans, bowine pancreas possesses the highest. specific activity of the
enzyme, followed by brain, Iiver, adrenal, heart, kidney, spinal cord,
spleen and thyroid. The level of the enzyme for methionine synthesis in
bovine liver as reported by lhngum et aI. (t972) appears to be about 6
times higher than that of rat liver observed by Finkelstein et aI.
(1971). Ttre activiy of 5-methylH4F-homocysteine methyltransferase in
sheep has been exami-ned only in the liver (Gawthorne and Smith, 1974)
and the enzyme level of sheep liver appears to be similar to that of rat
liver as reported by Finkelstein et al- (L97L). But the enzyme assay
system in the study of Gawthorne and Smith (tgl+) for the purpose of
invest.igation of the effect of vitamin 812 deficiency on the enzyme
activity did not include vitamin 812, the addition of vùrlch would
greatly affect the enzyme activity.. Thus, it may indicate that the
enzyme level in sheep liver might be higher than that of rat liver.
The specific activity of 5-methylt{4F-homocystej-ne methyltransferase
declines markedly with age in rat liver (ninkelstein et al., 1971) and
in human liver during early life (¡,hrdd et aI. , t970; Sturnnn and Gaull,
1978; Kalnitsky et. al. , L982).
Relative eontributions of the two homocysteine methyltransferases
to methionine methyl synthesis has not been well established. From the
early studies of du Vigneaud and his colleâgues, it h/as shown

13
thât the utilizat,ion of preformed methyl SrouPS is predominant in
methionine synthesís in rats fed dieLs adequate in choline, folate,
vitamin 812, serine and glycine and that the de novo synthesis of methyl
groups is cornpãratively small (du Vigneaud and Rache1e, 1965). This
indicates that betaine-homocysteine methyltransferase is more inrportant
than 5-methylH4F-homocysteine methyltransferase in methionine synthesis
in rats. The irnportance of betaine-homocysteine methyltransferase in
maintaining tissue concentrations of methionine is also emphasized in
Lhe recent studies of Finkelstein et aI. (tgAZa,U). They found that
the actiwity of the hepa.tic enzyme is increased in rats fed a
methionine-free diet, cornpared with a diet conLaining a normal
methionine content, and ttnt fhe depletion of choline in rat diets le-ads
to a marked decrease of the hepatic levels of methionine and AdoMet. The
important contribution of 5-methylH4F-homocysteine methyltransferase to
methionine synthesis in raLs, particularly in non-hepatic tissues, was
not recognized until L977, as Finkelstein et al. (t97I) found that
5-methylH.F-homocysteine methyltransferase is nmch more rridely
distrih-rted in rat, Lissues than betaine-homocysteine methyltransferase
and the activity of the former enzyme increases under conditions vihere
there v¡as a need for methionine synthesis.
Predominance of one route of methionine synthesis over the other
may differ between species. Studies on patients v¡ho are defective in
5-methylH4F-dependent methylation of homocysteine revealed that these
patients have difficutty in maintaining nornnl plamsa and tissue
concentrations of methionine and extribit hornocystinuria (reviewed by
Finkelstein, t975; lfudd and levy, 1983). The conclusion r,¡hich emerges
from'these observaLions is that the route of the de novo synthesis of
methyl groups for methionine biosynthesis is of particular importance in

t4
humans. As C,antoni (L977) pointed out: "in this respect, adult humans
are different from growing rats , in v¡hich homocysteine methylation
occurs chiefly through the action of betaine-homocysteine
methyltransferase.t'
Tissue distribution of the two homocysLeine methyltransferases and
relative importance of the two enzymes in methionine slmthesis have not
been studied in sheep. Based on the facts that dietary choline is
almost. unavailable for sheep (Ueitt et aI-, 1979) and the tissue betaine
conLent is primarily dependent on dietary choline (I^long and Thompson,
t972; Finkelstein et a1., 1982b), the de novo synthesis of methyl groups
might be the predominant pathway in methionine slmthesis in sheep and
more inportant than in most of non-ruminant species.
2.4. ULTLizat.ion of Methyl Groups via AdoMet-dependent Transmethylation
2.4.t Biosyrthesis of AdoMet
AdoMet was first discovered by Cantoni in 1952, v¡?ro showed that.
this compound was synthesized from methionine and ATP. The reaction is
cataLyzed by methionine adenosyltransferase and represented as follows:
L-Methionine + ATP AdoMet + PPi + Pi
Methionine adenosyltransferase has been found in all nn¡r¡nalian
t.issues examined (¡tu¿¿ et al. , 1965; I-ombardini et al. , L973; Radcliffe
and Egan, L974; Hoffman and Kunz, L977; Eloranta, t977; Okadå et a1.,
1981) and it has now been dernonstrated that there are three isozynes
r^¡ith different kinetic properties in manmals (see Section 2.6.t.t).

15
Reports on the tissue levels of the enz)¡me are dif f icult to
sunrnarize because much of bhe data had been obtained before the three
isozymes r4¡ere lqrown. In general, the level of methionine
adenosyltransferase is the highest in manmalian liver. The specific
activities of the enzyme in non-hepatic tissues of rats is many times
less than that in the liver. The report of Radcliffe and Egan (1974) on
the enzyme levels in various tissues of sheep, goats, cat.tle and rats is
not in accord with those reporled by other irnzestigators. They. found
that the specifj-c activity of the enzyme in the liver of sheep, goats
and rats is lower tt¡,an ttlat in some other tissues, such as spleen,
kidney cortex and duodenum. The reason for such a difference is not
apparent.
The specific activity of methionine adenosyltransferase in
manrnalian tissues may change with age. It has been shown that the
enzyme activity is lower in the fetal liver of hu¡nans, rats, mi-ce and
rabbits than in the adult liver (Gautl et aI., 1972; Sheid and Bilik,
1968; Hancock, 1966). However, the specific activity of the hepatic
enzyme of . rats during postnatal developrnent uras reported to decrease
r,rith age (Finkelstein, f967), vùrile Eloranta (L977) showed that the
specific activity of the enzyme in rat liver remained fairLy unchanged
throughout pos tnatal developrnent.
2. 4 . 2. Qqan titatively Predominant AdoMet-dependent Transmethylation
React.ions
A subsLantial part of the utilization of methyl groups is involved
in the transfer of methyl groups from AdoMet, wtrich is a primary methyl
donor. The numerous transmethylation reactions have been revíewed by

16
Irtudd and Cantoni (1964) and Cantoni (1975). Tnterest in
transmethylation tns been heightened in recent yeârs by obsen¡ations
dcmonstratÍ-ng or suggesting ûrat tte methylation of RNAr'DNA and protein is
involved in the modulation of the biological functions of these
macrornolecules, such as the role of RNA methylation on protein synthesis
(revieved by Salvatore et a1., 1980), DUa methylation on gene expression
(reviewed by Doerfler, 1-983) and cell differentiation (reviewed by Jones
and Taylor, 1980), and protein carboxylmethylation on chernotaxis and
neurosecretion (reviewed by O'Dea et al-, 1981). However, amounts of
the methyl group of AdoMet required for the modification of nucleic
acids and protein are generally believed to be cornpa.ratively snrall in
total utilizat.ion of methyl groupsr âs indicated by the studies of Kerr
(L972) and }tudd and his colleagues (1975, 1980). Kerr (L972) reported
that the ratios of the specific activity of glycine methyltransferase to
LRNA methyltransferase is 2300 and 3500 in the nuclei and cytosol
preparation of rat lÍ-ver, respectively, even thougþ IRNA
methyltransferase used in the assay r^¡as more highly purified than
glycine methyltransferase- l,,ludd and his colleagues (1975, 1980)
concluded from thei-r studies thaL most of the methyl groups from AdoMet
are utilized for the synthesis of creatine and sarcosine in humans.
Ogawa and Fujioka (fggZU) pointed out that glycine methyltransferase
is the most active among methyltransferases in rat liver. Dawson et aI.
(1931) reported that a large amount of methionine methyl is required for
choline synthesis in adult sheep.
In the scope of the thesis, this section will only discuss the
methylation reactions vñich are quantitatively important in the
utilization of methyl groups.

L7
2.4.2.1 l4ethylation of Guanidinoacetate
Itre methyl group of methionine was found to be the precursor of the
methyl nroiety of creatine in 1940 both in wivo (du Vigneaud et aI. 11940)
and in witro (Borsook and Dubnoff, 1940). cantoni and vignos (rgs¿)
first demonstrated tt¡at the inrnediate methyl donor is AdoMet, using a
partially purified enz)lne from pig liver. The enzyme named
guanidinoacetate methyltransferase transfers the methyl group from
AdoMet. to guanidinoacetate to form creatine. Guanidinoacetate is in turn
synthesized from arginine and glycine (Borsook and Dubnoff , t94t), r,øtrich
is catalized by arginine-glycine amidinotransferase. The overall
reactions of creatine s¡mthesis are shown as follows:
Arginine Ornithine Ado}4et AdoHcy
Glycine Guanidinoacetate Creatine
Gr.¡anidinoacetate methyltransferase is widely distrib:ted in
manrnalian tissues (reviewed by l,tralker, L979; Yanokura and Tsukada,
7982). In general, manrnalian liver contains the highest level of
guanidinoacetate methyltransferase actiwity and is the nrain site for
the last step of the synthesis of creaLine, vfiich is subsequently
transported by blood to other tissues, mainly nmscle. The activity of
the methyltransferase is extremely low in norihepa.tic tissues except
kidney, pancreas and testes vùrich rnay possess a - moderate activity.However, hunran pancreas has a cornparable level of the enzyrrìe to that of
the liver (van Pilstrn et aL., L972).
An irnportant aspect of creatine slmthesis in methyl group metabolism

18
is that the formation of creatine represents a constant drain on the
organism's supply of methionine, as creatine does not yield its methyl
group for transfer and is lost. in urine as creatinine and creatine.
Creatine is one of major methyl compounds in manmalian tissues. For
example, a 70-kg rnan contains approximately 120 g of creatine plus
creatine-P in nmscle and nerr/e tissues ( Walker, L979). The mean value
of urinary excretion of creatinine plus creatine in adult humans is
about L9 nnples per day in males and 13 nmoles in fernales in normal
conditions ( ¡t¡¿¿ and Poole, 1975). This amount of loss from the body
pool of creatine has'to be replenishêd by endogenous biosynthesis
and / or dietary creatÍ-ne. Urinary excretion of creatine plus creatinine
in adulL sheep with 50-kg body weight is about 10 nrnoles per day
(Henderson et a1., 1983), vrtrich exceeds the daily output of methionine
frorn the gut in this speeies as reported by I,JoIff et al. (L972) and
Lindsay (L982). The methyl group required for creatine synthesis alone
in humans also exceeds methionine supplied from a normal diet and
accounts for about. 70 7" of the total utilizat.ion of AdoMet. under normal
dietary conditions (l'h:dd, 1980). Thus, the demand for labile methyl
groups in hunnns and presumably in most other manrnals is determined
to a large extenL by the requirements for creatine synthesis.
2.4.2.2 l'4ethylation of Phosphatidylethanolamine (ptdetn)
The transfer of methyl groups from methionine to choline was
discovered by du Vigneaud and his colleagues (fg¿f). Bremer et aI.
(fg0O) first demonstrated that choline is synthesized in
phosflratidylcholine (pt¿Oro) form by the stepr,rlse methylation of PtdEtn
with the intermediate formation of phosphatidylmonomethylethanolamine

L9
(ptdVentn) and phosphat.idyldimethylethanolamine (PtdMe2Etn) and rhat
AdoMet serves as the methyl donor for all three methyl æ+6 in ttÞ ctDlirÞ
molecule. The overall reactions are represented as follows:
AdoMet AdoHcy AdoMet AdoHcy AdoMet AdoHcy
PtdEtn PtdMeEtn PtdMe2Etn PtdCkro
The enzyme vihich catalyzes the overall reactions is generally
called phospholipid methyltransferase (or PtdEtn methyltransferase).
The conversion of PtdEtn to PtdMeEtn is proven to be the rate-limiting
step (Bremer and Greenberg, L961.). The question has arisen as to
r,¡trether the conversion of PtdEtn to Ptdcho is catalyzed by one or two
enzymes in nnnmals. Studies using the solubilized methyltransferase from
rat liver microsomes índicate that a single enzyme catalyzes the
stepwise methylation of PtdEtn to form PtdCkro (nehbinder and Greenberg,
1965; Tanaka et. al., L979; Schneider and Vance, L979). Beginning in
1978, several studies have suggested the existence of two
methyltransferases in mammalian tissues (Hirata and Axelrod, L978;
Hirata et al., L978; Crews et a1., 1980; Sastry et a1., 1981). These
two methyltransferases are distinguished with respect to pH optirmrm,
magnesium requirement and Km values for AdoMet. The first enzyme
(methyltransferase I) catalyzes the methylation of PtdEtn to form
Ptdl'{eEtn, and the second enzyme (methyltransferase II ) catalyzes the
incorporation of two methyl groups into PtdMeEtn to form Ptdcho.
However, this has been challenged by Audubert and Vance (1983), lùro have
shown that all three meLhylation reactions in microsomal preparations of
rat liver have essentially the same Km value for AdoMet and optimal pH,

20
using an equation r^¡ith vihich calculations of the rate of the formation
of Ptdl4eEtn, PtdMe2Etn and Ptdcho take into account the subsequent
conversion of Ptd},leEtn and PtdMe2Etn to PtdCLro. I4ato and Alernany (fgA:)
conrnented on this argument in great detail and their conclusion is that
the existence of the two methyltransferases in manrnals is undecided yet
and further unequivocal evidence has to be obLained. However, there
is convincing genetic evidence' for at least two distinct
methyltransferases in certain fungi (Scarborougþ and Nyc, 7967.) and
yeast (Yamashita et al. , L982).
Phospholipid methyltransferase is largely confined to the
endoplasmic reticulum and exists in a variety of manmalian tissues, but
is only quantitatively i-rnportant in liver (Bremer and Greenberg, 796t;
Skurdal and CornaLzer, L975; Hirata et aI., L978; Blusztajn et al. tL979;
Welsch et al., Ig81; Tanaka et aI., 1979; Vance andde Kruijff, 1980).
Ttre specific activity of phosphotipid methyltransferase in rat
Iiver increases markedly with the early postnatal age and thereafter
slowly drops to the adult value (Hoffman et al., L979; Pelech et aI.,
1983). It has been explained by Pelech et al. (1983) that the increase
of the enzyme level at the early postnatal age conLributes to the
changes in the molecular species of PtdCkro in neonatal rat liver, as the
species of PtdCho synthesized via the transmethylation pathway contain
more polyunsaturated fatby acids than those formed via an alternative
route, CDP-choline pathway.
Methylation of phospholipid has recently been suggested to play an
important role in many biological processes related to membrane
structure and funclion, such as changes in local membrane fluidiLy and
modulating the transmission of certaj-n biological signals across plasma
me¡nbrane (reviewed by tlirata and Axelrod, 1980; Hirata, t982; MaLo and

2L
Alemany, 1983).
Ptdcho is synthesized mainly by two rJutes, the methylation pathway
and the CDP-choline pathway. The methylation pathway is not a
predorninant route for PtdCkro slmthesis and amount.s to about 20-40 % of
the total synthesis of Ptd(ho in rat liver with most of the rest derived
from the CDP-choline pathway (Sundler and Åk ""o.,, 1975). The
contrib¡tion of the methylation pathway to Ptdcho synthesis in
non-hepatic tj-ssues is much less significant.
An inrportant aspect. of PtdEtn methylation in relation to methyl
group metabolism is that the synthesis of choline molecule involves a
transfer of thrree methyl groups from AdoMet , but only one of them in
the choline molecule can be recovered as a labile methyl group during
choline catabolism. Therefore, choline synthesis represents a constant
drain on the labite methyl group pool. Dawson et al. (1981) reported
that choline s¡rnthesis in adult sheep appears to be about 17 nunoles per
day and this would require 51 nrnoles of the methyl group of methionine,
which is 5-10 times the daily output of methionine from the gut in this
species (Woff et al., 1972; Lindsay, L982). Assessment of choline
synthesis in humans nny be indirectly derived from the study <-rf
Ifudd et aI. (1980), vùro have shown that the amount of sarcosine formed
from the oxidation of the endogenously synthesized choline plus that
from the reaction of glycine methylation in a young adult patient with
sarcosine dehydrogenase deficiency on a nonnal diet is 2.0-2.4 nrnoles
per day. Hence the quantity of choline synthesized in humans would be
less ttlan that va1ue. Such a difference in choline synthesis between
humans and sheep rdth a similar body size nny be part.ly explained by the
virtual unavailability of dietary choline for sheep. Clearly, PtdEtn
methylation is quantitatively significanL in the utilizaLion of labile

22
methyl groups, particularly in sheep
2.4.2.3 Methylation of Glycine
The methyl transfer from AdoMet to glycine \^tas discovered by
Bltrnenstein and tlilliams (1960) and is one of the two pathways in
sarcosj-ne forrnation. The other pathway of sarcosine synthesis
is via choline catabolism and will be discussed in Section 2.5.3.
Ttre enzyrne named glycine methyltransferase catalyzes the following
reaction:
Glycine + AdoMet ---) Sarcosine + AdoHcy
Relatively litt1e attention has been paid to glycine
methyltransferase since its discovery in 1960 and no review on the
subject has appeared to date. Glycine methyltransferase is largely
localized in cytæI, brt ar appreciable quantity of the enzyme also exists
in nuelei (Kerr, 1972). The enzyme acLivity has been detected in the
Iiver of rats, mice, hamsters, guinea pigs, rabbits, dogs, pigs, calves,
Iambs and humans (Blumenstein and l^lilliams, 1960, t963; Kerr, t972;
Heady and Kerr, L975; Liau et al., L979). Rabbit liver contains the
highest level of the enzyme activity among all species examined. The
enzyme activity in lamb liver r^ras reported to be low, but no actual
value was given (Blunenstein and liilliams, L960; Kerr, 1972). Glycine
methyltransferase has also been found in the kidney and pancreas of rats
and rabbits (Kerr, L972), the spleen, 1*g, heart and intestine of rats
(Yanokura and Tsukada, L982) and the breast of humans (Guerinot and

23
Bohuon, 1977). The specific activity of glycine methyltransferase in the
tissues of rats and rabbits is the highest in the liver, followed by
pancreas and kidney (Kerr, t972). The enzyme activity in rat lung,
heart and intèstine is extremely low (Yanokr:ra and Tsukada, t982).
It has been noted that significant levels of glycine
methyltransferase activity hTere found only in those organs which
contain significant levels of methionine adenosyltransferase
(Kerr, L972; Yanokrra and Tsukada, 7982). The enzyrne level in the
early fetal liver of rabbits and rats is very low and increases
narkedly with the prenatal and postnatal age (Heady and Kerr, t975;
Liau et aL., t979).
It has been reported that develognental changes in the glycine
methyltransferase level show an inverse relationship to LRNA
methyltransferase in rabbit liver (Heady and Kerr, 7975) and frog liver
(Heady, L979). The authors postulated that glycine methyltransferase can
modulate the activity of IRNA methyltransferase both by compet.ition for
Adollet and by the generation of the irùribitory product, AdoHcy. The
evidence supporting this hypothesis also emerges from studies of the two
enzyrne activities in rat tumor cells (Heady and Kerr, t975; Liau et. a1.,
L979i Yanolura and Tsukada, 1982). In contrast to these observations,
Guerinot and Bohuon (tgZZ) trave shown that there is no such inverse
relationship between the activities of the two enzymes j-n human
breast tumor tissues. Ttre content of glycine methyltransferase in rabbit
Iiver is very high and accounts for 0.9-3 7. of the soluble protein of
the liver (Heady and Kerr, L973). Ogawa and Fujioka (1982b) conrnented on
that the activity of glycine methyltransferase in rat liver is the
highest of lcrown methyltransferases. l"lLrdd et. al. (fgAO) observed that
increments of methionine added to a normal diet produced a rise in

24
sarcosine excretion equivalent to about 70 7" of the supplemental
intake in patients with sarcosine dehydrogenase deficiency. They
suggested that this increment in sarcosine excrebion is largely formed
by the methylation of glycíne. These observations suggest that the
methylation of glycine is one of the major reactions in the utilization
of labile methyl groups.
2.5. Catabolism of the Main Methyl Cornpounds
2.5.7 Methionine Catabolism
Catabolism of methionine can occur via the pathways of
transsulfuration and transamination. Ttre transsulfuration pathway of
methionine catabolism is through the fornration and cleavage of
cystathionine after the conversion of methionine to homocysteine via
AdoMet-dependent transmethylation. Consequently, the potency of
methionine regeneration from homocysteine is lost.
Binkley (1951) first. demonstrated tknt theformation ofcystathionine
in mamnnlian tissues from homocysteine and serinq¡ with rat liver
preparations free of the enzyme responsible for the cleavage of
cystathionine. The reaction is catalyzed by cystathionine B-synthase .
The enzynratic cleavage of cystathionine in manrnals \¡ras discovered by
Binkley et. al. (L942) v/ith the identification of cysteine as one
of the products and the other product rÁ/as identified as
cr-ketobr¡tyrate by Carrol et aI. (L949). The enzyme named
l-cystathionase catalyzes the reaction of the cleavage of
cystathionine. The overall transsulfuration reactions are

25
represented as follows:
NH4
Homocysteine * serine Cystathionine Cysteine + a-ketob-rtYrate
Both reactions require pyridoxal phosphate as a coenzpe.
The distribrt.ion of cystathionine 6-synthase and '¡-cystathionase
has been examined in the various tissues of rats and some tissues of
rnany other species of manrnals by l"hrdd et aI. (fS0S¡ and other
investigators ( na¿ctiffe and Egan, L974; Sturman and Gaull, L978;
Zlotkin and Anderson, 1982). These two enzyrnes have been found to occur
in all examined tissues of adult mannrals except heart and muscle v¡here
Y-cystathionase was not detectable (Mudd et al., 1965; Radcliffe and
Egan, tg74). In general, the activities of cysLathionine ß-slmthase and
y-cystathionase are high in marnnralian liver, Pancreas and kidney.
However, hunnn adrenal and sheep duodenum appear to contain a
high activity of y-cystathionase (Zlotkin and Anderson, t982;
Radcliffe and Egan, t974). The brain, adipose tissue and rnucosa
of the small intestine of rats contain a moderate level of
cystathionine 3-synthase, but a low level of Y-cystathionase'
The activities of these two enzymes are very low, if present at all, in
other tissues of rats.
The levels of cystathionine g-synthase and Y-cystathionase
increase with age in rat liver (Finkelstein, t967; Heinonen, L973),
hunran liver (Sturman and GauII, !978; Zlotkin and Anderson, t982;
+HzoH2o

26
Kalnitsky et al., lg82) and rat brain (volpe and aster, t972;
Heinonen, Lg73). The developxnental increase of cystathionine
6-synttr,ase has also been reported in the brain of humans and
monkeys (Gaull et aI. , t972; Sturman et. aI., L97Oi Rassin et aI.,
1981). However, in monkey liver, an increase in the specific activity of
y-cystathionase \¡Jrith age is accompanied with a decrease of
cystathionine ß -synthase activity (Sturman and Gaull, t978). In
contrast to hunans, monkeys and rats, the specific activity of
y-cystathionase declines markedly with age in sheep liver (nadcliffe
and Egan, Lg74). There is no data i-n literature concerning the
develo¡xnental change of cystathionine ß-synthase in sheep.
In contrast to the transsulfuration pathway wtrich has been well
established, the transamination pathway of methionine catabolism has not
been fully elucidated to date. The transamination reaction of
L-methionine in manrnalian tissues was first shown by Rowsell (tgStt
1956a,b), vùro demonstrated that both pyruvate and cr-ketogluLarale accept
amino groups from L-methionine in rat liver. However, in the
subsequent two decades, little progress had-been reported in this area.
Establishrnent of the transamination pathway is ascribed Lo
Benevenga and his colleagues. Case and Benevenga (tgl;, 1977) showed
that methionine can be extensively cataboLi-zed by a pathway
independent of AdoMet formation and that formaldehyde and formate \^Iere
two intermediates in the oxidation of methionine methyl by this
pathway. Further investigations identified methanethiol and
3-methylthiopropionate as j-ntermediates in this pathway (Steele and
Benevenga, Lg78, Lg19). The present understanding of the
transamination pathway has been sunrnarized by Benevenga and ngan (l-983)

27
as follows:
c -Ket o-Y -me thiolbutyrate
cD2
3-Me thyl thiopropiona te
Methionine
Methanethiol
a-Keto acid
a-Amino acid
3 carbon compounds ?
/\Fornnldehyde Hzs
Formate
@2
The ociÉtsne of the transamination pathway has been shown in the liver
of rats (C,ase and Benevenga, L976), pigs (Benevenga and Haas, L979),
monkeys (Steele and Benevenga, 1978), sheep (Benevenga and Egan, 1983;
Benevenga et al., 1983), and some other tissues of rats (¡titctrelt. and
Benevenga, 7978). A nurnber of aminotransferases have been shovn to be
able to catalyze methionine transamination in in vitro studies and they
are glutamine aminotransferase (Cooper and Meister, L972, !974),
asparagine aminotransferase (C-ooper, t977), histidine aminotransferase
soa

28
(Noguchi et al., L976) and leucine aminotransferase (fteaa et aI.,
1976). However, the aminotransferase(s) responsible for the formatíon of
a-keto-v-methiolbutyrate from methionine in vivo re¡nain unlanown. It has
been suggested that the in vivo actíon of glutamine aminotransferase rnay
caEaLyze the corn¡ersion of a-keto-T-rnethiolb:tyrate to methionine,
rather than the transamination of methionine (Cooper, 1983). The
reåson for this is that the nn:ch higher levels of glutamine in tissues,
coupled rrith the virtually irreversible nature of the reactiof¡. with
glutamine, should ensure that the reaction is directed toward glutamine
utilization. C-ooper (1983) also suggested that without such a mechanism
an excessive loss of essential methionine might occur. It has recently
been shown that a-keto-y-methiolbutyrate is not only derived directly
from methionine by transamination, but also formed from
5t-methylthioadenosine, a product of meLhionine breakdown via AdoMet
(see Section 2.5.2).
The transamination pathway has been demonstrated to operate at
physiological eoncentrations of methionine in rats (C-ase and Benevenga,
1977; EngsErorn and BenevengarlgSl), although the affinity of the
aminotransferases for methionine for its transaminat.ion is very low (see
Section 2.6.t.L). Q:antiLative inpætace of tte transamination paLhway in
methionine catabolism has been shown in rats (Case and Benevenga, t977)
and in sheep (Benevenga and Egan, 1983). However, the
transsulfuration pathway is considered Lo be the major route of
methionine catabolism (reviewed by Greenberg, t975; l'4udd and Levy, L978;
Finkelstein, t975; ¡'fudd and levy, 1983). Further quant-rtative
evaluation is needed to establish the relative contributions of these
two pathways Lo methionine catabolism under physiological conditions.
Tl:re pathway of transamination would become very important, vttten the

29
transsulfurati-on paLhway is inrpaired. The recognition of the Benevenga
pathway leads to possible ne$¡ approaches in the treatment of
homocystinuria due to cystathionine B-synthase deficiency. It has
been shown by Smolin et aI. (fgAf) that supplements of betaine are
beneficial in avoiding the detrimental effects of high homocysteine
concentrations by increasing the rate of homocysteine conversion Lo
methionine, vitrich might in turn increase the rate of methionine
catabolism via tte transamination pathway. However, the existencé of lhe
transamination pathway and its quantiLative significance in methionine
catabolism in humans rernain to be established.
2.5.2 AdoMet Breakdown via 5'-Methylthioadenosine (Uf¿,) Fornration
The enzymatic fornation of MTA from Adol"fet in manrnalian tissues by
three pathways has been reviewed by Zappi-a et. aI.(1980), t^Iilliams-Ashnran
et aI. (fgAZ) and Schlenk (1983) and is sunmarized as follows:
AdoMet
c02 Uridine (in tml¡,)
Decarboxylated AdoMet
R:trescine Homoserine
(or Spermidine)
Spermidine
(or Spermine)
3- ( 3 -Amino- 3-carboxypropyl )
Uridine
l',lTA

30
Among these, the pathway for the polyamine synthesis represents
the quantitatively most important route for l'fiA forrnation
(Wittiams-Ashman et aI., 1982). In this pa.thway, AdoMet. is first
decarboxylated, catalyzed by AdoMet decarboxylase, and then
donates its aminopropyl group for the synthesis of polyamines with the
equimolar formation of I'!TA. The enzymatic cleavage of AdoMet into
MTA and homoserine lactone, catalyzed by Adol4et cyclotransferase,
was found in manrnalian liver (Swiatek et al., 1973; I^lilson et
aL., L979; T,appia et al ., L979), ht the function of this reaction is
unloown. ¡fIA is also a by-prodrrct of the enzymatic Lransfer
of a 3-amino-3-carboxyl-propyl group from AdoMel to uridine in tRNA
(Williams-Ashman et aI., t982). Ttre authors pointed out that this
reaction contribrtes only negligibly to the total production of
l.{IA in vivo.
To the present lceowledge, the catabolism of MIA in manrnalian
ti-ssues mainly goes through a route by wtrich MTA is first converted to
5-methylthioribose-1-phosphate (t'finp) and then cr-keto-l-methiolbutyrate,
v¡hich enters the transamination pathway of meLhionine catâbolism
(Schlenk, 1983). Backlund and Smith (1931) first demonstrated
that carbons from the ribose portion, the intact methyl grouP
and the sulfur of MIA are aII incorporated into methionine by
soluble extracts of rat liver. Backlund et aI. (fggZ) further
showed that a final step of transamination to a-keto-Y-methiolbutyrate
from glutamine or asparagine leads to methionine. The authors proposed
that the pathway appears to be a significant salvage pathway for
methionine synthesis in manrnals. The sirnplified presentation of the
pathway to illustrate the salvage of methionine from MTA is shown in the
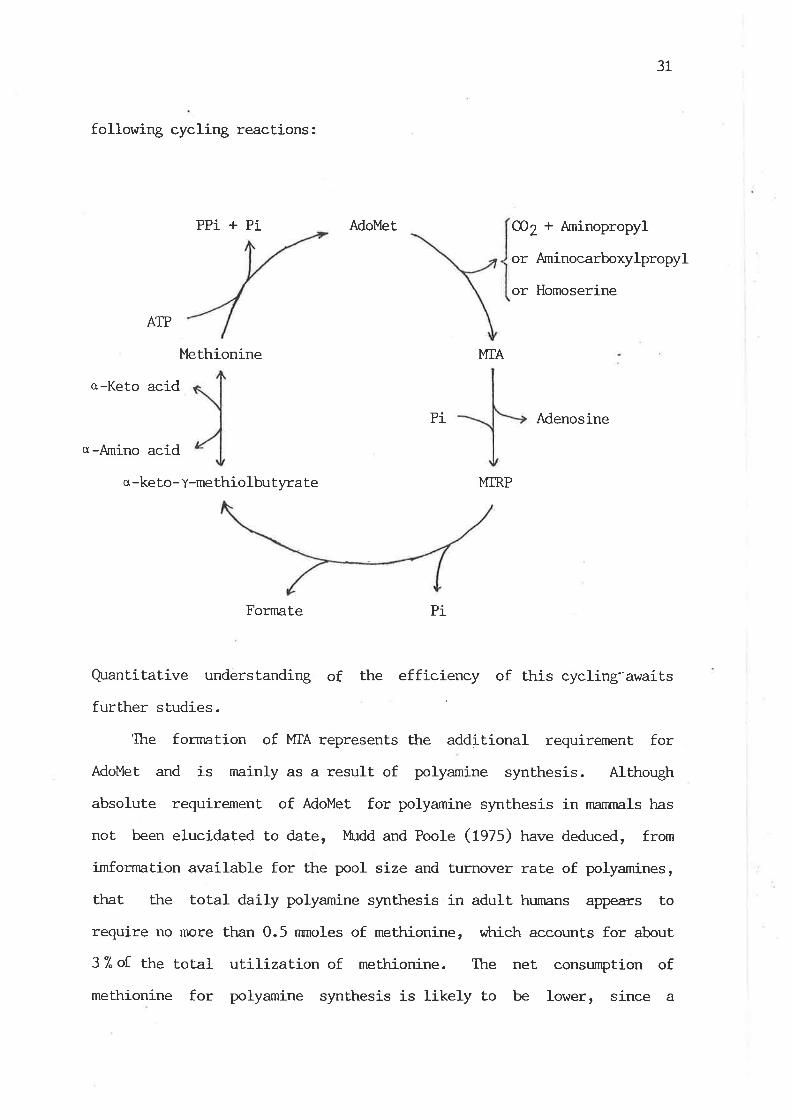
following cycling re¿ctions :
PPi + Pi
ATP
Methionine
AdoMet
31
CÐZ+ Aminopropyl
or Aminocarboxylpropyl
or Homoserine
MTA
o-Keto acid
Pi Adenosine
q -Arnino acid
o -keto- t-me thio lbutyra te MTRP
Fornate Pi
Quantitative understanding of the efficiency of this cycling-'awaits
further studies
The formation of MIA represents the additional requirement for
AdoMet and is mainly as a result. of polyamine synthesis. Although
absolute requirement of AdoMet for polyamine synthesis in nranrnals has
not been elucidated to date, lfudd and Poole (1975) have deduced, from
imfornation available for the pool size and turnover rate of polyamines,
that the total daily polyamine synthesis in adult hunnns appears to
require no more ttnn 0.5 nrnoles of methionine, vihich accounts for about
37"of the total utilization of methionine. The net consumption of
methionine for polyamine synthesis is likely to be lower, since a

32
portion of MTA, the by-product of polyamine synthesis, can be converted
back to methionine. Presumably, the daily requirement of methionine for
polyamine synthesis in sheep is similar to hunnns, as humans and sheep
have a similar body size.
2.5.3 Ckroline Oxidation
The catabolic pathway of choline has been sunmarized by MuCd and
c,anroni (1964) and illusrrated by }tudd and Poole (rszs; as follows:
Ckroline
Betaine
Homocysteine
I,lethionine
Dimethylglycine
Sarcosine ttHclìott
Glyeine
Througþ this pathway, choline is first oxidized to betaine and then
corn¡erted to dimethylglycine with recovery of one methyl group as
methionine methyl for each choline molecule. The oxidative demetþlation
of dimethylglycine occurs stepwise with sarcosine and finally glycine as
products. The carbon atorns of these methyl groups removed in the course
of the demethylation can enter the metabolic pathway of one-carbon

33
fragments. (tqcCitvery and Goldstein, 1983).
The oxidation of choline to betaine is catalyzed by the enzyme
system called choline oxidase r,ihich consists of two enzylnes, choline
dehydrogenase and betaine aldehyde dehydrogenase. Choline dehydrogenase
calalyzes the oxÍdation of choline to betaine aldehyde vùrich is further
oxidized to betaine, catalyzed by betaine aldetryde detrydrogenase.
Ctroline dehydrogenase is present in tle particulate fraction of cells,
mainly in mitochondrí-a, vùtereas betaine aldehyde dehydrogenase appears
to be present both in the particulate and soluble fractions (Wi[iams,
L952arb; tlatefi and Stiggall, 1976).
The activity of choline oxidase has been examined in several
manrnalian tissues of monogastric species (Bernheim and Bernheim, 1933,
1938; Kensler and langemann, L954; Sidransky and Farber, 1960; Gr:Lra and
I{egrnann, 1963). In general, the activity of choline oxidase is high in
liver and kidney and very low, if present at all, in other tissues.
Sidransky and Farber (1960) found that the activity of hepatic choline
oxidase is the highest in rats, followed by mice, dogs, hamsters,
rabbits, monkeys, guinea pigs and humans. This is in close agreement
with the observations of Haubrich and Gerber (1981) on the activity of
hepatic choline dehydrogenase among these species. Arì apparent
correlat.ion between hepatic choline oxidase activity of a species and
the tendency to develop choline-deficient. fatty liver has been suggested
by Handler (tgt+g) and tlandler and Bernheim (fg+g). The activity of
choline oxidase has also been examined in calf liver (Hopper and
Jotrrson, 1956), vitrich contains a nn:ch lower activity than rat liver,
but has not been reported in sheep tissues. However, studies of
tlaubrich and Gerber (1981) on choline dehydrogenase showed that the
enzyrne activity in sheep liver and kidney ís nn:ch lower than that in

34
the corresponding tissues of rats. This nny indicate that the level
of choline oxidase is low in sheep tissues to conserve this valuable
methyl cornpound.
The specific activity of choline oxidase has been shown to increase
nnrkedly with age in rat liver (Zeisel and l^Jurtman, 1981). A similar
postnatal increase in the specific activity of choline dehydrogenase has
been obsen¡ed in rat liver (Weinnotd and Sanders, L973) and mouse liver
and kidney (fAuUrich and Gerber, 1981)
An irnportant feature of choline catabolism in relation to methyl
group metabolism is the loss of two out of the three methyl groups
transfered from AdoMet in the course of choline synthesisr âs
dimethylglycine is successively dernethylated with no recovery of those
methyl groups. Apparently, the consen¡ation of choline by reducing its
oxidation in a condition v¡tren the dietary supply of choline is
inadequate is beneficial to the economy of methyl group uLilization.
2.6. Regulatory Aspects of Methyl Group Metabolism
The prirnary means of controlling the flow of nutrients through
their metabolic pathways is the regulation of the rates of enzymatic
reacLions. Like other nutrient metabolism, the rates of the enzymatic
reactíons involved in methyl group metabolism have been found to be
subject to alteration by three basic mechanisms: 1) changes in tissue
concentrations of enzymes, 2) modifícat.ion of enzyme to an active or
inactive form, and 3) changes in concentrations of substrates,
cofactors, aclivators or intribitors. In the scope of this thesis, the
enrphasis of the review of the regulation of methyl group metabolism is
placed only upon the control of the rates of enzymatic reactions by

35
ligand concentrations and of enzyme activities by the availability of
methyl cornpounds and by the hormone, insulin.
2.6.I. C-ellular Levels of Substrates and Km Values of Conrpeting &rzymes
The relative rates of flux of a metabolite through several
metabolic pathways are largely controlled by its cellular concentration
relative to the Km values of competing enzymes for the substrate and the
relative tissue capacities of the compet.ing enzymes measured under
conditions similar to the physiological status. As data for the
cornparison of tissue capacities of the competing enzymes are not
available, in this section lhe physiological concentrations of
substrates and Km values of the enzymes related to methyl group
metabolism are to be sunrnarized and compared to reveal one aspect of the
potential regulatory mechanisms.
2.6.1.L l4,ethionine l-evels and Km Values of Methionine-utilizing Enzymes
Ttre mean values of physiological concentrations of methionine are
50-130 nnrol/g wet tissue in rat tissues (Herbert et a1., 1966) and
50-140 nmol/g in rat liver (Herbert et al ., t966; Daniel and l^laisman,
L969a; Spector et a1., 1980; Finkelstein et aL., L982b). The mean values
of the concentrations of plasma methionine are 20-30 nmol/ml in humans
in the fasting state (¡'laclean et al., 1983; Boers et al., 1983), in
sheep (Strath and Shelford, 1978; Radcliffe and Egan, 1978), in goats
(Mepham and Linzell, 1966) and in fasting pigs (Chavez and Bay1ey,
7976), and 50-100 nmol/ml in rats (Spector et aI., 1980; Herbert et al.,
t966; Khader and Rao, L982) and in dogs (Longenecker and Hause, 1959).

36
The recent kinetic studies of methionine adenosyltransferase in
mannnals have revealed that the enzyme has three isozynes with widely
different Km values for methionine. X* (SO.S) values of the three
isozymes in raL liver for methionine are 200-130 ffl, 14-30 pM and 6 tfi'respectively (liau et al., L979; Ktrnz et al., 1980; Hoffman and Kunz,
1980; Okada et al., 1981). The lowest-lh isozyrne was found to be the
sole form of methionine adenosyltransferase in non-hepatic tissues of
rats (Hoffman and Kunz, 1980; Okada et a1., 1981). TTre ðominant
isozyme in rat liver is a high-Km form (Líau et aI., 1979). The
Km values for methionine for the three isozymes of methionine
adenosyltransferase in human liver appear to be similar to those in rat
liver ( liau et aL., L919 ). A single species of methionine
adenosyltransferase with a low Ifu has also been demonstrated in human
erythrocytes (Oden and Clarke, 1983) and in human spleen, thynms
and brain ( Liau et al. , t979 ), although Tallan (L979) reported
at least two isozymes present in hr¡nan erythrocytes, cultured
fibroblasts and cultured lymphocytes, based on chromatographic
separation.
The Km values of Ehe L-form and K-form isozymes of glutamine
aminotransferase for methionine transamination are about 2 ml4 and 4 nù4
in rat liver and kidney, respectively (Cooper and l"leister, t972, 1974).
Although the Km values of other aminotransferases v¡trich were fourd to be
also active toward melhionine have not been determined for methioni-ne,
their I(m values for other amino acids are of an order of mM (fte¿a et
al., L976; Cooper, L977). Presunably, their Km values for methionine
are likely to be similar to those for other amino acids.
By cornparison of Km values for methionine between methionine
adenosyltransferase and the aminotransferases in relation to the

37
cellular levels of methioni-ne, apparently, methionine flux goes
preferentially through the transmethylation-transsulfuration route under
physiological conditions. Such a physiological arrangernent would ensure
methionine utilization both as a methyl donor and as the cysteine
precursor and minimize the unnecessary J-oss of methionine via the
Lransamination pathway. However, v¡hen an elevated level of methionine
occurs in a condit.ion of excess methionine, the aminotransferases
responsible for methionine transamination and the high-Km isozynle of
methionine adenosyltransferase in ma¡rmaliam liver would both become
functional to remove excess methionine.
2.6.I.2 AdoMet l,evels and Km Values of MethylLransferases
Physiological concentrations of AdoMet in most of rat tissues are
20-100 nnrol/g of wet tissue (Baldessarini, L966; Salvatore et al., L97L;
Eloranta, L977; Hoffman et al., L979; Ckriang and Cantoni, f979). The
hepatic level of AdoMet j-n rats is 50-100 nmol/g (Baldessari_ni, L966;
Salvatore eÈ al. , L97L; Hoffman, 1975; Eloranta et al. , L976; Elóranta,
t977; Hoffrnan et al. , L979; Ctriang and C,antoni, L979; Poirier et al.,L982; Finkelstein et al., L982b). A similar range of tissue AdoMet
levels has also been reported in mice (Hoffman, \975; Hoffman, 1980;
Schatz et a1., 1981a; Helland and Ueland, 1983), in fruit bats (van der
Idesthuyzen and MeLz, 1983) and in rabbits (Salvatore et aI. , 19lL).
Arnong numerous transmethylation reactions, guanidinoacetate,
glycine and phospholipid methyltransferases are quantitatively
important enzymes in the utilization of AdoMet. The Km value
of guanidinoacetate methyltransferase for Adol,{et is 6.7 FM
in rat liver (Qgawa et. a1., 1983) and 49 pM in pig liver ( Im er

38
a1., 1979b). Ttre Km value of glycine methyltransferase for Ado}4et is
100 FM in rabbir liver ( Heady and Kerr, 1973) and 30 pM (SO.S) in rat
liver (Ogawa and Fujioka, t982a). Ttre Kinetic constants of phospholipid
methyltransferase are difficult to sunmarize¡ owing to involvemenÈ of tte
stepnrise methylation of PtdEtn in Ptd0ro slmthesis. It has been
reported that the lfu value for AdoMet in rat liver is about 1 pM for
the first step methylation and 70 ¡:M for the last two methylation
reactions (Sastry et aI., 1981). However, Audubert. and Vance (1983) have
shown that. the Km values for the three methylation reactions in raL
liver range from .60 to 100 pM for AdoMet at a pH optinnrm of t}.25t
using an equation !úth vùrich calulations of the rates of the
fornntion of PtdMeEtn, PtdMe2Etn and Ptdcho take into account
the subsequent conversion of PtdMeEtn and Ptd}4e2Etn to Ptdcho.
Audubert and Vance (1983) further showed that, at pH6.6, the
Km.' values of phospholipid methyltransferase for AdoMet for the three
methylation reactions are 3-7 P", about lO-fold lower than those
obtained at pH 70.25. It seems likely that the I(m values of phospholipid
methyltransferase at physiological pH rnay be well below tissue lévels of
AdoMet. The Km values of most other AdoMet-dependent methyltransferases
for AdoMet appear to be below tissue levels of Adoì'l,et, such as 10 pM for
phenylethanolamine methyltransferase from rabbit adrenal (Deguchi and
Barchas, t97L) and for histamine methyltransferase from guinea pig brain
(Ttrithapandna and C.otur, 1978 ) , t4 ¡:M for catechol methyltransferase
from rat liver and acetylserotonin methyltransferase from bovine
pineal (Deguchi and Barchas , L97t), L2-t3 lrM for histone
methyltransferase from rat brain (Drerra et al. , L977), 3 pM for
protein-lysine methyltranferase frorn rat liver (Oliva et aI., 1980),
2-3 pM for protein-carboxyl methyltransferase from human and rat

39
erythrocytes (KÍm, 7974), !.3 ¡rM for DNA methyltransferase from rat
liver (Cox et aI., t977), and 0.3-3 pM for LRNA methyltransferases from
rat liver (Ctict and Leboy, L977; Glick et aI. , 1978). The analysis of
the Km values of various methyltransferases in marnmalian tissues
reveals that most methyltransferases are essentially saturated with
pkrysiological concentrations of AdoMet. Variations in tissue levels of
AdoMet trnder certain conditions rnay greatly influence the rate of
glycine methyltransferase. Relatively low Km values of the
methyltransferases for the methylation of DNA, RNA and protein indicate
that the rate of the methylation reactions of these macromolecules nlzly
not be controlled by the concentrations of Adolfet in most tissues. Ttris
appears to be a sensible physiological arrangement, as the methylation
of these macromolecules is most irnportant in biological processes of
life and lhe amount of AdoMet required is also small.
2.6.t.3 HomocysLeine l-evels and Km Values of
Homocys teine-utilizing EnzYmes
Homocysteine in manrnalian tissues is formed by the hydrolysis of
Adogcy. The reaction is reversible, catalyzedby AdoHcy hydrolase, and
the Lhernrodynamics favor the synthesis of AdoHcy. No data existed on
the levels of homocysteine in tissues, except human plasma, under
physiological conditions before 1984. There are several reports
concluding that the homocysteine content in marnnalian tissues is below
the detection Limit (Finkelstein et al. , I97L; Reed et al. ' 1980; Fahey
et al. , L98t; Dudman and l{ilcken, t982). However, sma1l amounts of
homocysteine have been demonstrated in human plasnn, being 1-18 nmol/ml
in the mixed disulfide form (Saetre and Rabenstein, 1978; Boers et al,

40
1983) and about 1.5 nrnol/ml in the protein-bound form (Kang et aI.,
LgTg). By a more sensitive method, Ueland et al. (1984) r¿ere able to
dernonstrate that total hornocysteine levels in the liver, kidney,
brain, heart, lt¡ng and spleen of mice and rats are in the range of
0.5-6 nnrol/g of wet tissue.
Three additional enzymes are quantitatively iruportant in the
utilizalion of homocysteine besides AdoHcy hydrolase for the reaction in
the synthetic direction. These enzymes are betaine-homocysteine
methyltransferase, 5-methylHOF-homocysteine methyltransferase
and cystathionine 6-synthase. Ttre Km Value of betaine-homocysteine
methyltransferase for homocysteine is L2 [rM it rat liver (Finketstein et
al., Lg12) and 120 FM ir human liver (stiua et al. , 1982). The I(m value
of 5-methylH4F-homocysteine methyltransferase for homocysteine is 60 PM
in rat liver (Finkelstein, 191L) and about. 100 FM in pig kidney (Burke
et al., L971,). The Km value of cystaLhionine B-synthase for homocysteine
is 1-15 mM in rat Iiver (sunmarized by Finkelstein, L979) and 0.59 mM in
htrnan liver (Kraus et al., L978). The kinetic studies of these three
enzymes clearly show that changes in t.issue levels of homocysteirre \.¡ill
greatly influence the rates of these enzynatic reactionsr âs the Km
values of all three enzyrnes are very nn:ch greater than tissue
homocysteine levels. In particular, the Km of cystathionine B-synthase
for hornocysteine is more than one order of magnitude greater than those
of homocysteine methyltransferases in rat liver. As pointed out by
Finkelstein (L979), homocysteine methyltransferases \^¡iII be at a
kinetic advantage relative to cystathionine ß-synthase' under
physiological conditions. Such a mechanism would favor the
consen¡ation of the homocysteine moiety of methionine at low tissue
Ievels of homocysteine and increase the fraction of homocysteine

4L
passing through the transsulfuration route at relatively high levels of
homocysteine as a consequence of excess methionine conditions, even
w1thout elevation of the concentraLion of cystathionine g-synthase.
However, the homocysteine moiety of methionine is eventually removed via
the transsulfuration route, as tteroute fæ tte conversion of homocysteine
to cysteine j-s unidirectional in vivo (Finkelstein, 1979).
2.6.2 Modulation of Transmethylation Reactions by the'Ratio
of AdoMet to AdoHcy
AdoHcy, one of the products of all transmethylation reactions
involving AdoMet, acts as a compet.itive intribitor of mosL, if not all,
of these reactions (Cantoni et aI. , 1979; Schatz et al., 1981a,b;
Kredich and Hershfield, 1980). The affinity of most methylLransferases
for AdoHcy is greater than for AdoMet (Cantoni et aI., t979). Tissue
concentrations of AdoHcy are generally nmch lower than those of AdoMet
(Eloranta, 1977; Hoffman et al; , t979; Helland and Ueland, 1983), bul
are high enough to cause some degree of irùribition of most of these
transmethylation reactions (Cantoni et al-. , 1g7g). Susceptibility
to such irùribition varies greatly among individual methyltransferases
(Kerr, L972; Cantoni et aI., t979; Kredich and Hershfield, 1980). It
has been demonstrated that intracellular ratj-os of AdoMet to AdoHcy
modulate AdoMet-dependent transmethylation reactions and play a
key role in the control of the utilization of AdoMet. (Cantoni et a1.,
L979; Hoffnran et. aI ., 1979; Cantoni and Clriang, 1980; Schatz et aI.,
1981arb). Such a regulatory mechanism provides the most acute form of
enz)rue regulation, with almost inrnediate nranifestations. Any factors
causing changes in tissue AdoHcy or AdoMet levels could exert an instant

42
control on the rate of transmethylation.
2.6.3 Effects of Availability of Preformed Methyl Compounds
Dietary methionine has been shown to have the most pronotnced
effect on the metabolic rates of methyl groupsr âs methionine plays
a central role in methyl group metabolism. Excessive methionine
administration to manrnals markedly elevates tissue concentrations of
methionine (Oaniel and I^laisman, t969a; Harper et al. , t97O; Strath and
Shelford, t978; Zeisel and lfurtman, 1979; Boers et aI. , 1983).
A pronounced elevation of Adol,4et levels, after methionine loading, has
been shor.¡n in rat liver (Schlenk, 1965; Baldessarini, 1966; Eloranta,
L977; Hoffman et a1., 1980). However, the elevation of AdoMet levels
is nn:ch srnaller in the non-hepatic tissues of rats (Baldessarini, L966;
Carl el al., !978; Zeisel et aI., L979), although methionine loading
produces a similar range of increase of methionine levels alnong rat
tissues (Daniel and l.]aisman , 7969a; Zeisel et aI., L979). These
obsen¡ations indicate that the capacíty for tte conversi¿¡ ofmethionine to
Adol4et in the non-hepatic tissues of rats is limited, vùrile the capacity
in the liver is nnrch greater. Presumably, this is because rat liver
possesses the high-Km isoz)¡me of methíonine adenosyltransferase vùrich
becomes functional at the elevated methionine concenLration. A marked
increase of hepatic AdoMet level, after a single dose of excessive
methionine administration, without the alteration of methionine
adenosyltransferase activity (Eloranta, t977) is tte evidence to support
the above suggestion. Iong-term administration of excess methionine
increases the activity of methionine adenosyltransferase in rat liver
(Ogawa and Fujioka, 7982b; Fau et a1., 1981), although an early study by

43
Sanchez and Swendseid (1969) showed no changes in the enzyme activity in
rats fed a diet containing 4 7" methionine. Feeding a higþ protein diet
also produces an increase of the enzyme act.ivity in rat liver
(Finkelstein, L967). Increases of AdoMet level and methionine
adenosyltransferase activity indicate that methionine flux through
the transmethylaLion-transsulfuration route could be increased under the
methionine excess conditions.
The further removal of excess AdoMet. produced after methionine
loading appeårs to be attributed mainly to glycine methyltransferase.
lfudd et aI. (fggO) have shown that an increment of methionine added to a
normal diet produces a rise in sarcosine excretion equivalent to about
70 7" of the supplemental intake in humans and it is likely that. this
incremental sarcosine synthesis is Iargely due to glycine methylation.
Benevenga and Flarper (fgZO) have demonstrated that. glycine
supplementation of the high methionine diet erùnnces the rate of the
conversion of methionine methyl to CO2 in rats, indicating that the
glycine methyltransferase reaction is a potential route to re¡nove excess
methionine methyl. A direct demonstration on the induction of glycine
methyltransferase in rat liver, afLer excess methionine feeding, has
been reported by Ogawa and Fujioka (1982b). In contrast Lo glycine
methyltransferase, the activity of guanidinoacetate methyltransferase in
rat liver is not affected by excess methionine feeding (Ogawa and
Fujioka, 1982b). This is in good agreement with the observations of lfudd
et al. (1SAO¡, vfrro found that creatinine excretion is not altered by
ctr,anges of methionine intake. There hras no report concerning effects of
excess methionine on phospholipid methyltransferase activity before
L984, but the reduction of methionine availability appears to decrease
the choline content in sheep liver (Smittr et al., L974), vùrich may

44
indicate the reduced rate of choline synthesis.
Investigation into the effects of methionine availability on
transsulfuration reactions in manrnals is cre of tte irçortart objects in the
study of methionine metabolism. It has been well estabf-lished that the
availability of dietary methionine greatly influences the rate of the
conversion of homocysteine to cysteine (Finkelstein, 7970, 1979).
Protein supplementation of a low protein diet produces an increase in
the activities of cysLathionine B-synthase and Y-cystathionase in rat
liver (Finkelstein, t967). Daniel and l^laisman (fg6gU) and Fau et aI.
(1981) reported thât excess methionine feeding increases only the
activity of Y-cystathionase, but not cystathionine ß-synthase, in rat
Iiver. Daniel and ln/aisman (tO0SarU) have also shown that the
cystathionine level is increased more than 1O-fold in rat liver after
injection with high doses of methionine or feeding a diet containing 3 7"
methionine. The resulLs of the latter two groups seern to suggest the
potential capacity of cystathionine ß-synthase is great enough to ensure
the removal of excess homocysteine produced from methionine even w1thout
an increase of the enzyme activity. Ihe high Km of cystathionine
ß-synthase for homocysteine provides such an explanation. However,
Finkelstein et al. (1975) has shown that Adol.,l,et acts as an activator
of cystathionine ß-synthase from rat liver ín in vitro experiments.
Therefore, the enzyrne activity in rat liver is certainly increased in
vivo under the condition of methionine excessr âs excess methionine
greatly elevates the hepatic AdoMet. Ievel. Further evidence to support
the idea that cystathionine ß-synthase in rat liver is a regulatory
enzyrne ernerges from the study of Finkelstein and Mudd (1967), vùro
observed that the cystathionine 8-synthase activiLy Í-s depressed by
cysteine supplement and the depression can be restored by

4s
administration of additional methionine'
Establishment of the regulation of methionine biosynthesis in
manrnals by the availability of methionine is mainly attribr-lted to
Finkelstein and his colleagues. Finkelstein et al. (fgZf) have
shown that the increase of dietary methionine contents decreases the
activity of hepatic 5-methylQ,F-homocysteine methyltransferase in rats.
Ttre supplernentat.ion of methionine to the rnethionine-deficient' diet
decreases the activity of betaine-homocysteine methyltransferase, but
an excess methionine addition to a 0.3 "/" m'ethionine diet increases the
enzyme activity in rat liver (Finkelstein et al.r.1982a). The authors
suggested that the betaine-homocysteine methyltransferase reaction may
function both to maintain tissue concentrations of methionine villen
the infake of this amino acid is 'limited and to remove homocysteine v¡hen
methionine intake is excessive. In vitro experiments have shown that
methionine irùribils both the reactions of betaine-homocysteine
methyltransferase (Finkelstein et aL. rt972) and 5-methylH4F-homocysteine
methyltransferase (Burke et al., L97t). Furthennorer methionine also
inciirectly affects the rate of its own slmLhesis through the action of
AdoMet, âs AdoMet inhribits the' reactions of betaine-homocysteine
methyltransferase (Finkelstein et al., L972) and 5r10-methyleneH4F
reductase in manrnalian t.isSues ( Kutzbach arxi Stokstad, t97I;
Matthews and Daubner, L982). These findings have clearly demonstrated
that. the rate of methionine slmthesis in manrnalian tissues is well
regulated by the availability of methionine througþ the action of both
the homocysteine methylation reactions.
cllanges in methionine levels could also influence the rate of its
oxidation via the transaminaLion pathway. Ttre rate of the oxidation of
methionine methyl to Cß2 increases markedly \^Iith methionine

46
concentrations in raLs both in vivo (Benevenga and Harper, t97O; C'ase
et al. , Lg76) and in vitro (Benevenga, !974; C-ase and Benevenga, 1976) '
C,ase and Benevenga (Lg76) have shov¡n that this increased rate of
methionine methyl oxidation i-n rats is mainly via the transamination
pathway, suggesting that it is one of the pathways in the modulation of
tissue methionine levels. However, the enzymatic response of this
pathway to changes in methionine intake has not been..studied to date,
probably owing to úncertainty in the recognition of the specific enzymes
responsible for this pathway in vivo at present'
Ckroline plays a significant role in the regulation of methyl group
metabolism, as dietary choline not only affects its o\¡rn synthesis
at the expense of the methyl group of AdoMet, but also provides methyl
groups for methionine synthesis. Skurdal and Cornatzer (fgZS) have
shown that the intraperitoneal injection of choline markedly decreases
prospholipid methyltransferase in rat liver microsomes' C'onversely,
choline deficiency in rats increases the hepatic phospholipid
methyltransferase activity by aL least 50 7" and irùribits the
CDP-choline pathway for PtdCtro synthesis (t-omUardi et aL', L969;
TLrompson et al. , 1969; Schneider and Vance, 1978). However, the
cornpensation by the increased rate of choline synthesis in raLs is
limited, as a marked decrease in the hepatic choline content occurs in
chol-ine deficiency (ttaines andRose, l97O; Thompson et al', 1969)'
Sctrneider and Vance (1978) noted that a 30-40 7" reduction in the
mibochondrial oxidation of choline to betaine takes place in response to
choline deficiency. The hepatic betaine content is drarnatically
reduced to a very low level in rats fed a choline-deficient diet (Wong
and Thornpson, L972; Aruidson and Asp¡ 1982; Finkelstein et a1', 1982b)'
Finkelstein et aI. (1983) further demonstrated ttìat excess choline

47
supplementation produces a marked elevation of the betaine content and
tt6t betaine-homocysteine methyltransferase activity in rat liver is
greatly increased by elevation of the tissue content of betaine' It
v¡as reported that the high level of dietary choline irhibits
5-methylH¿*F-homocysteine methyltransferase act.ivity in the liver of rats
fed on diets containing 0.5-0.7 7. methionine plus cysteine at a ratio
of 1:1 (Wgwira and Beames, L982), suggesting ttrat the de novo synthesis
of methyl groups is also controlled by the preformed methyl nutrient,
choline. In conclusion, choline supplernentation increases methionine
synthesis via the betaine-homocysteine methyllransferase reaction and
reduces the arnount of methionine'utilized for choline synthesis. BoLh
of these effects result in a reduction of dietary requirement of
methionine for its methyl group and explain the decrease in the de novo
synthesis of methyl grouPs from one-carbon sources'
Methyl group metabolism nEy be greatly influenced by dietary
creatine, as creatine synthesis in manrnals drains on a large proportion
of labile methyl groups from AdoMet. It has been well established that
creatine intribit.s iLs own synthesis both in vivo and in vitro (reviewed
by l{alker, Lg79). The feedback regulation of creatine does not seem to
be exerted on the guanidinoacetate methyltransferase reacti-on, but on
the amidinotransferase reaction for guanidinoacetate synthesis (I^lalker,
tglg). Effects of dietary creatine on other reactions of the pathways of
methyl group metabolism have not been investigated. A creatine-deficient
diet would greatly increase the requirement of the methyl Sroup of
methionine for creatine synthesis and consequently might increasq the
rate of methylneogenesis from one-carbon sources or decrease the rate of
methyl group catabolism via the glycine methylation pathway.

48
2.6.4 Effects of Insulin
l4any enzymes related to methyl group metabolism have been found to
be subject to regulation by some hormones (Finkelstein, 1970; Hoffnran et
al., 1981; Ilalker, t979-, C-astaño et al., 1980). However, there is a
particular reason to consider the enzymatic regulation of methyl group
metabolism by insulin, as insulin is ìrnown to regulate the rates of
protein synthesis and degradation and hence influences the utilization
of amino acids. Ttre pathways of methyl group metabolism consti-tute part
of the metaboliÈm of several amíno acids'
Finkelstein (1970) has shown that alloxan treatment, r'¡trich Ieads to
insulin deficiency in animals, produces marked increases in the
activities of methionine adenosyltransferase, cystathionine ß-synthase
and betaine-homocysteine methyltranferase in rat liver, b.rt the activity
of 5-methylH4F-homocysteine methyltransferase decreases after alloxan
treatment. Hoffman et al. (fg8f) reported that the activity of
phospholipid methyltransferase is decreased in alloxan-diabetic rat
liver. This is in agreement with a reduced level of hepatic Ptdcho
observed in diabetic rats ( T\rak¡Iov et aI., 1979). Alteration of
guanidinoacetate methyltransferase actiwity has been observed in
alloxan-diabetic rat liver, but not in alloxan-diabetic sheep liver
(Henderson et al., 1983). Ttre administration of insulin to normal rats
tr,as no effect on the activity of hepatic guanidinoacetate
methyltransferase (Carlson and van Pilsurn, t973). It has been shown that
insulin stinmlates the uptake of blood creatine by nnrscle (tüalker, L979)
and thus may decrease the end-product repression of arginine-glycine
amidinotransferase in other tissues. A reduced rate of creatine
synthesis in alloxan-diabetic sheep has been suggested by Henderson et

49
aI.(1S3). There is no report cn either the effect of insulin on glyeine
methyltranferase or study of this enzyme activity in diabetic animals.
These tinaings indicate that insulin deficiency leads to a
increased rale of methionine catabolism via the transsulfuration
pathway, however the utilizaLíon of methionine methyl for creatine
synthesis in sheep and choline slmthesis in rats is reduced. Tttus, the
elucidation of the regulation of methyl group metabolism in insulin
deficiency has to await further studies.
2.1 C-oncluding Remarks on Literature Review
Information presented in the literature review makes it clear that
post-ruminant sheep have a considerably lower availability of methyl
nutrients than most. non-ruminant species. However, most non-ruminant
species show some difficulty in achieving the metabolic balance of
methyl groups under the conditions of poor methyl nutrition similar to
those that sheep encounter. Ttre ewidence for regulatory aspects of
methyl group metabolism obtained from hurnans and non-ruminant aninals
suggest that an increase in the availability of methyl nutrients
enhances the rate of methyl group catabolism and reduces the rale of
methylneogenesis. Conversely, an inadequate supply of methyl groups
would result in an increased rate of methylneogenesis and a reduced rate
of methyl catabolism. Presumably, these might be the primary
mechanisms for sheep to sunrive on a low intake of methyl nutrients and
the capacity of operating these mechanisms might be even greater in
sheep than in most non-ruminant species. These considerations virere
proposed as a working hypothesis for the study on methyl group
metabolism in sheep.

50
CXIAPTER 3. MATM.IAI-S AND GENMAL }4ETHODS
3.1. l'bterials
3.1.1 Animals
Merino sheep were used througþout the course of this study. Aget
sex, maintenance and physical conditions of the aninrals are indicated in
the following specific experiments. Female l^Iistar rats aL the age of
3-6 months, weighing about 2OO g, were used in this sLudy. ltre animals
v¡ere housed in a windowless, weII ventilated room, fed a pelleted rat
diet (ftrarlicks, Adelaide,
access to water.
South turstralia) ad libitum and had ready
3.L.2 Radiochemicals
The following radiochemicals vùere purchased from A¡nersham
International plc. (Buckinghamshire, England): [nrethyl-3H]choline
(Za Cilnnrol), S-[14C]methyltetrahydrofolate (58.3 mCi/nrnol),
L-[merhyl-3u] methionine (l-S cilnrnol), L-[35S]methionine (1230 Cilnnrol),
S-adenosyl-L-[raethyl-3H]methionine (SOO mCi/nnrol) and [3H]acetic
arùrydride (500 Cilmol) .
3.1.3 Biochemicals
Biochemicals hrere obtained as follows: AdoMet (HSO4- ) from
Boehringer Manr¡heim GmbH, In. Gerurany; L-cystathionine from Koch-Light
LaboraLories Ltd, C-olnbrook, Br:cks., F:rgland and all other biochemicals

51
from Sigma Ctremical C-o., St. Louisr Mo.r U.S.A.. AlI reagents were of
the highest purity corTmercially available.
3.I.4 CLrromatographic Materials
Ac 50t^t-X8 (1OO-ZOO mesh, H + ), AG 50In-X4 (1OO-ZOO mesh, H+),
Dowex 1-X8 (100-200 mesh, Cl-_) and C-eltex-p (U+) were obtained
from Bio-RAD laboratories, Richmond, Calif . ' U.S.A.. Thin--layer
chrornatography (TfC) plates of cellulose (0.1 nm thiclcress, 20 x 20 cm)
and Silica gel 60 (O.Z *p thiclaress , 20 x 20 crn) were obtained from
Merck, Darmstadt, W. Germany.
3.1.5 Fnzymes
Ckroline oxidase (tZ.t+ units/mg protein) was obtained from Sigma
Ckremical Co.. Partially purified choline acetyltransferase was
prepared from sheep brain caudate nuclei by the method of Ryan and
lbClure (L979) to the step of Cl"I-Sephadex C-50 colu¡rn chromatography
purificat.ion.
3.2. General Methods
3.2.t Preparation and Purification of Radiochemicals
3.1.1.1 Preparation of [tt ttyt-3H]getaine
[u.ttyt-3tt]Betaine viras prepared from [methyl-3H]chorine'
reaction mixture in a fínal volume of 1 ml contained 20 P¡nol
The
of

52
porassium phosphate (pH 7.8), 20 ¡rmol of Ttis-HCf (pH 7.8), 5 Fnrol of
¡,tgSO4: 1.5 pmol of NAD, 100 ¡rci of [methyt-3H]cf,oline (500 mCi/nnrol) and
2.5 mg of choline oxidase (tZ.+ units/mg protein). Ttre incubation was
carried out at 30oC for 5 hr. Linder these conditions the conversion of
choline to betaine r^ras 95-98 7". Ttre re¿ct.ion v¡as terminated by heating
in boiling water for 3 min. Ttre denatured enzyme protein was rernoved by
centrifugation. The supernatant was applied to a colunn, 0.6 x 3'5 cm,
of AG 501,ü-Xg (ti*, 100-200 mesh) to separate betaine from choline and
betaine aldetryde, according to the method of tlilken (1970). Ihe lithium
form of AG 5OI^¡-X8 hras prepared frorn the hydrogen form by stirríng with
1 ¡,f LiOH. Betaine was eluted with 6 mI of water, vilrile choline and
betaine aldehyde were retained on the resin. Further purification of
[methyl-3g]betaine was carried out by paper chronatography. The eluate
obtained from the colunn was lyophilized, and the residue was dissolved
in a small amount of water and applied to l,lkratman 3 nrn chromatography
paper. The paper was developed by ascending ctrromatography for 15 hr
with the solvent system of isopropanol/acetic acid/water (fO:f ß, v/v).
The positions of betaine and choline on the paPer were
identified by exposure of authentic betaine and choline, rùrich had been
co-chromatographed, to 12 vapour. R¡ values for betaine and choline are
0.43 and 0.57, respectively. Radioactive betaine was eluted from the
paper with water, stored in sealed vials at -80oC and used for
betaine-homocys teine methyltrans f erase assay within one month'
3.2.1.2 Rrrif ication of [u"tt yt-3tt]Groline
Iløttryt-3g]ckroline r^las
described by l,ùilken (1970).
purified by paper chromatography as
The radioactive choline was eluted from

53
the paper hrith water, stored in sealed vials under a nitrogen gas ptr,ase
at -80"C and used for choline oxidase assay within one week.
3.2.t.3 Purification of [¡l.tt yt'3H]Ado]aet
lUettyt-3H]edo¡,let r^ras purified by TLC on cellulose coated plates
with a solvent system of n-b:tanol/acet.ic acid/water (60:L5225, v/v) as
described by Hoffnnn (1975). The chronratography was carried out at 5"C
for 5 hr. The radioactive.AdoMet on the plate was visualized under U.V.
Iigh! ( U.V.-fignt exposure time as short. as possible) and eluted from
the cellulose powder with cold 1 mM HCl. The purified [methyl-3U]ndoUet
had a radiochemical purity of approximately 98 "/"t as determined by
chronntography on a AG 5OIi-X4 colunn (NH+* form) as described in
guanidinoacetate methyltransferase assay (see Section 3.2.2.4). The
purified [methyl-3H]edo¡,tet in 1 nM HCI solution was stored at -80oC and
used for assays of AdoMet-dependent methyltransferases within 2 weeks.
3.2.2. Preparations and Assays of F-nzymes
For enzyrne preparations, homogenization was performed at OoC in an
ice bath and all other procedures \^/ere carried out at 0-4"C.
3 .2 . 2 .t 5-llethytH4F-homocys teine Methyltrans f erase
Preparation and assay of 5-methylH4F-homocysteine methyltrasferase
were based on the method of }fudd et al. (1970). AII tissues except
nmscle, heart and lung were homogenized in a Potter-Elvehjem tissue
homogenizer equipped with a motor-driven teflon pestle hrith 2-3 vol of

54
0.1 M potassium phosflrate b.rffer (pH 7.5) containing 1 mM reduced
glutathione. Ifuscle, he-art. and lung were homogenized with Kinematica
Polybron honrogenÍ-zer (ttt 10, L-rzern, Switzerland). The homogenates \,/ere
centrifuged aE 8000 x g for 10 min and then at 501000 x g for 2 hr. The
clear supernatants rnrere dialyzed three times against the above
b¡ffer for 3 hr. The dialyzed extracts were stored at -15oC and used
for the enz)fiìe assay next day
Ttre incubation mixLure for the measurement of the enzyme activity
contained 0.1 M potassium phosphate buffer (pH 7.4), 150 pM AdoMet,
50 FM cyano-812, 225 mM 2-mercaptoethanol, 250 pM L-homocysteine
[freshly prepa.red from homocysteine thiolactone hydrochloride by
incubation with 5 N NaOH, as described by D:erre and Miller (fgOO)],
150 f,,1"1 5-[14C]meLhytH4F (0.1-0.2 ¡:Ci) and the enzyme in a final volume
of 0.2 ml. The incubation was performed at 37"C trnder a nitrogen gas
phase for 40 min.
nitrogen.
The reaction was stopped by rapid freezing in liquid
The radioactive methionine formed t/as separated from radioactive
5-methylH.F according to the method of l,/eissbach et aI. (1963). The
frozen reaction mixture was thawed by the addition of 0.8 mI of cold
water, then i¡rnediately applied onto a 0.6 x 2 cm colunn of Dowex 1-X8
(Cl l 100-200 mesh) and the radioactive methionine lr7as eluted
with water in a total volume of 2.5 ml.
A portion of the eluate was mixed lrith 9 vol of Tfiton X-lOO/toluene
based scintillation fluid containing 7 g 2r5-diphenyloxazole and
0.3 g lr4-dL]2-(4-methyl-5-phenyloxazolyl]-benzene per litre of the
mixture of toluene and Triton X-100 (22L, v/v). The radioactivity was
determined in a Packard Tticarb Liquid Scintillation counter, l4cdel
3375. Counting efficiency v¡as determined by the channels ratio method of

55
quench correction.
IJnder the assay conditions, the rate of methionine fornntion in
sheep liver sarnples was linear with the enzyme protein up to 1 nrg and
\^/ith incubation time rsithin 60 min. The incubation with the boiled
enzyme (or without enzyme) was taken as a b1ank,value.
3.2.2.2 Betaine-homocysteine Methyltransferase
Betaine-homocysteine methyltransferase was prepared and assayed
essentially as described by Finkelstein'and lfudd (L967). Tissues were
homogenized in 3-4 vol of 40 mM potassium phosphate b-uffer (pH 7.5).
The homogenates were centrifuged at 181000 x g for 15 min and the
supernatant fractions vJere used for the enzyme assay.
Ttre incubation mixture contained 50 mM potassium phosphate
buffer (pH 7.4), 10 nM freshly prepared L-homocysteine, 2 nM
[methyl-3H]betaine (0.5-2 pCi) and the enzyme fraction in a total volume
of 0.2 mI. The L-homocysteine was prepared as follows z t5.4 mg
L-homocysLeine thiolactone hydrochloride was dissolved in 0.2 ml of 2 N
KOH, allowed to stand at room temperature for 5 min and then neutralized
as described by bfudd et aI. (fg6S). The incubation for the assay of the
enzyne activity was carried out at 37oC for 20 min . The reaction was
terminated by rapid freezing in liquid nitrogen. 0.4 ml of cold water
was added into the reaction mixture, before it vras applied onto a
0.6 x 3 cm colunn of Dowex 1-X8 (OH -, 100-200 mesh) to separate the
products, methionine and dimethylglycine, from betaine. Ttre colunn was
washed vdth 6 ml of water to re¡nove betaine. Then, methionine and
dimethylglycine were eluted with 4 ml of 1.5 M HCl. The radioactivity
in the acid eluates.was determined as described in Section 3.2.2.L. A

56
blank value obtained by incubation with the boiled enzyme was routinely
subtracted. Under the assay conditions, the product format.ion in sheep
liver sanrples vùas proportional to the enzyme content up to at least 1.5
mg protein and to incubation time within 30 min.
3.2.2.3 Phospholipid Methyltransferase
Assay of phospholipid methyltransferase activity was by a modified
method of Audubert et aI. (fgA+) by measuring the rate of Lhe transfer
of [3U]methyl groups from Imethyl-3H]ldoMet to phospholipid in
microsomes. Microsomes \,¡ere prepared from liver as follows. Tissues
were homogenized in 4 vol of 0.25 M sucrose containing 10 mM Tris-HCI
(pH 1.4) with a Potter-Elvehjem homogenizer for 10 strokes. The
homogenates l^¡ere centrifuged at 10,000 x g for 20 min and the resulting
supernatant fluids were centrifuged at 105,000 x g for 60 min toseparate the microsome and cytosol fractions. The microsomes were
resuspended in 0.25 M sucrose containing 10 mM Tris-HCI (pg 7.a) and
0.5 nrM dithiothreitol (Off) and used as a source of phospholipid
methyltransferase.
The incubation mixture contained 100 mM Tris-glycine
(pH 9.2), 1 mM cysLeÍ-ne, 10 mM MgCl2: 0.1 rnM EDTA , 0.15 mM
[methyl-3tt]Rdot',tet (0.5 pci) and the microsomes in a final volume of
0.1 ml. The endogenous PtdEtn in the microsomal preparations was used
as one of the substrates in the assay. The incubation was performed at
37oC for 15 min and the reaction was stopped by the addition of 3 'mI of
chloroform,/methanol/Hcl (2:LzO.O2, v/v). Radioactive phospholipid formed
was then extracted as described by Hirata et al. (1978). 1 ml of
chloroform ptr,ase containing the radioactive phospholipid was transferred

57
to a miniscintillation vial. Æter the solvent \,rras evaporated to
dryness at 8OoC, 4 mI of Triton X-1O0/toluene based scintillation fluid
v¡as added and the radioactivity was measured in a Iiquid scintillation
counter as described in Section 3.2.2.L. Ttre incubation r,rith boiled
microsomes r¡Jas taken as a blank value. under t-tre assay conditions, the?
rate qf ['H]methyl groups i-ncorporated into phospholipid in the samples
of sheep hepa.tic microsomes was linear with microsonnl protein at least
up to 0.2 mg and with incubation time within 20 min.
The [3H]methylated phospholipids were identified by TLC on Silica
GeI 60 plates as described by Castaño et aI. (1980). The [3tt]methylated
phospholipid produgls accounted for more than 937" of total radioactivity
recovered from the TIC plate, of vùrich approxinrately 85 7" of the
radioactivity was associated hrith PtdGto.
3.2.2.4 Guanidinoacetate Methyltransferase
The cytosol fractions prepared as described in Section 3.2.2.3 were
dialyzed 3 times against the buffer containing 50 mM Tris-HCf (ptf 7.5) t
0.2 nM III| and 0.1 mM EDTA for 18-20 hr. The dialyzed cytosol was used
as the source of guanidinoacetate methyltransferase.
Tlne measure¡nent of guanidinoacetate methyltransferase activity rnlas
based on the method of Im et aI. (tglga). The incubation mixture
contained 50 nM Tris-HCI (pH7.5), 5 mM 2-mercaptoethanol, 2 mM
guanidinoacetate, 0.2 nM [methyl-3H]AdoMet (O.f ¡:Ci) and the enzyme in a
final volume of 0.1 mI. After the mixture \¡ras incubated at 37oC for 15
min, the reaction was terminated by the addition of 100 ¡r1 of 10 7"
trichloroacetic acid. The resulting mixture rÁlas adjusted to pH 7.6 by
the addition of aOO ¡r1 of. 0.2 M unbuffered Tris solution and then poured

58
onto a colunn (0.6 x 2 cm) of AG 5OI4I-X4 (tmf, form, 100-200 mesh). The
colunn vras washed 3 times with 0.65-mI portions of
water to elute radioactive creatine formed. The radioactivity in the
eluates \.rras measured as described in Section 3.2.2.L. The incubation
with the boiled enzyme was taken as a blank value. Ünder the assay
conditions, the rate of creatine formation in sheep liver samples r{as
linear with the enzyme content up to 0.7 ,ttg protein and with incubation
time r¿ithin 40 min.
3.2.2.5 Glycine Methyltransferase
The activity of glycine methyltransferase was measured in the
dialyzed cytosol fractí-on, v¡irich \^Ias prepared as described in
Section 3.2.2.4, by a modification of the method of Heady and Kerr
(L973). The incubation mixture contained 50 rn'M Tris-HCl (pH 8.8), 20
nM glycine, 7 mM DTT, 0.2 nM [mehtyl-3H]adouet (O.S-f pci) and the
enzyrne fraction in a final volume of 0.1 mI. The incubation \^las
performed at 37"C for 15 min. The reaction was stopped by the addition
of 100 pI of tO 7" trichloroacetic acid. The separation of the
radioactive product from unreacted radioactíve AdoMet was Lhe same as
for the assay of guanidinoacetate methyltransferase (see
Section 3.2.2.4). The radioactivity in the eluates was determined as
described in Section 3.2.2.L. The incubation with the boiled enzyme or
without the substrate, glycine, was taken as a blank. llnder the assay
conditions, the rate of sarcosine formation in sheep liver sarnples \¡ras
linear with the enzyme content at least up to 0.8 ng protein and with
incubation time w.ithin 20 min.

59
3.2.2.6 Choline Oxidase
The assay of choline oxidase was modified frorn a combination of the
methods of Zeisel and l^Jurtman (1981) and l{ilken (1970). Tissues r¡lere
homogenized in 10 vol of 0.1 l''l potassium phosphaLe buffer (pH 7.a) for
10 strokes with a Potter-Elvehjem homogentzer. The homogenates were
centrifuged at 121000 x g for 15 min. The pellets were washed once
\^rith 10 vo1 of the same buffer as above and then suspended in 5 wol of
0.2 M potassium phosphate (pH 7.a) with a homogenizer. of this
suspension, 1OO ¡rl was incubated w-ith 0.2 ¡-rno1 of NAD, 1 pmol of LlgSO4
and 1 ¡LmoI of [methyt-3tt]choline (1 pci) in a final volume of 0.2 mI.
The incubation r,ras performed at 31oC fot 15 min under an atmosphere of
pure oxygen. The reaction was stopped by the addition of 0.8 ml of
ice-cold water and 1 ml of chloroform,/methanol (1:1, v/v). After being
shaken vigorously for 5 min, the two phases v¡ere separated by
centrifugation. 0.9 mI of the aqueous phase was withdrawn and put under
an air stream to renove methanol and then poured onto a colu¡rur
(O.O x 3.5 crn) of AG 50I^l-X8 (li*, 100-200 mesh). The reaction product,
betaine, rnras eluted r^rith water to obtain a final volume of 3 mI of the
eluaLe, vùrile choline and betaine aldehyde were retained on the resin.
Ttre radioactivity in the eluates was measured as described in
Section 3.2.2.L. The incubation with the boiled enzyrne was taken as a
blank va]ue. The rate of betaine formation in sheep liver sarnples \{as
linear for 20 min under the conditions described.
To identify the radioactive cornpounds in the eluate obtained from
the AG 50t^l colunnr, 2 mL of the eluate of rat. liver sample was dried in a
rotatory evaporator at 35oC and reduced pressure. The residue was
dissolved in 100 ¡rI of water and 20 ¡rmoles of carrier betaine and

60
chloine were added. Of this solution, 60 pI was applied to a silica
ge1 60 plate (ZO x 20 cm, 0.2 nm thick). The plate vras developed by
ascending chromatography for 1.5 hr with a solvent system of
methanol,/4O mM potassium phosphate buffer, pH 7.5, (1:1, v/v). The
positions of betaine, choline, phosphorycholine and acetylcholine on the
plate \^/ere visualized by 12 vapour. V values for betaine,
phosphorycholine, acetylcholine and choline were 0.53, 0.30, 0.26 and
O.22, respectively. The bands of betaine, choline, the
choline-containing compounds and the rest of the silica gel on the plate
were scraped off and eluted vrith 1 N HCl. The radioactivity was
measured as described in Section 3.2.2.t. The radioactivity in betaine
accounted for approxinaLeLy 97 % of the total radioactiwity recovered
from the plate.
3.2.2.7 Cystathionine B -Synthase
For preparat.ion of cystathionine 6-synthase, tissues were
homogenized in 4 vol of 1.15 7" KCI for 20 strokes with a Potter-Elvejhem
homogenizer. The homogenates were allowed to stand for at least 20 min
and then centrifuged aL I2r000 x g for 30 min. The clear supernatant
was brought to 40 7" saturaLion of a¡rmonium sulphate by Lhe slow addition
of the saturated solution (pH 7.5) with stirring. After standing foro60 min aL 0 C, the precipitate was collected by centrifugation at.
181000 x g for 20 min and dissolved in 0.1 M potassium phosphate buffer
(pH 7.5). After the solution was dialyzed overnight against the same
buffer, it was used as a source of cystathionine ß -synthase.
The activity of cystathionine ß-synthase $ras measured essentially
according to the method of Kashiwamata and Greenberg (fgZO). The

61
incubation mixture contained 0.1MTris-HClbuffer (pH 8.3), 80 mM
L-serine, 0.12 mM pyridoxal-P, 0.5 nM fuSO4, 1ffi mM freshly prepared
Dl-homocysteine and the enzyrne in a final volume of 0.5 mI. For the
preparation of Dl-homocysteine, 1 nmole of Dl--homocysteine thiolactone
hydrochloride v¡as dissolved in 0.36 mI of 5 N NaOH, and after standing
at room temperature for 5 min, 1.56 ml of waterr 780 pI of 0-66 M
Ttis-HCL (ptt 8.3) and 600 ¡:1 of 0.88 M HCI were added in sequence \^rith
stirring. The enzyme reacLion started by the addition of the
solutions of Dl-homocysteine and serj-ne after pre-incubation for 10 min
aL 37oC *ithout these substrates. After incubation at 37oC fot 80 min,
the reaction \,ùas stopped by the addition of 50 ¡:I of 50 7"
trichloroacetic acid. An incubation blank was obtained with the boiled
enzyme.
The measurement of cystathionine formed was the same as described
by Kashiwarnata and Greenberg (fgZO). Llnder these conditions, the rate of
cysÈathionine formation in sheep liver sarnples was linear \^/ith the
enzyme content up to 3 mg prolein and with incubation time for 100 min.
3.2.2.8 y -Cystathionase
For assay of .¡-cystathionase actiwity, the enzyme was prepared as
follows: Tissues ulere homogenized in 2 vol of 0.01 M potassium
phosphate buffer (pH 7.5) containing 0.15 M KCl, 0.1 mM pyridoxal-P and
0.5 mM EDIA. The homogenates were centrifuged at 10,000 x g for 10 min.
Tlre supernatant was dialyzed 3 times against the txrffer containing 50 mM
potassium phosphate buffer (pH 7.5), 0.5 mM EDIA and 0.05 mM pyridoxal-P
for 16-20 hr. Insoluble material formed during dialysis was removed by
centrifugation. The resulting supernatant was used as a source of the

62
enzyme.
The incubation medium used for the determination of the activity
of y-cystathionase v/as the same as described by Greenberg (1962),
except that 5 mM L-cystathionine v¡as used as subslrate instead of 20 mM
Dl-cystathionine. The reaction mixture in a,total volume of 1 ml was
incubated aL 37oC for 15 min. The reaction was stopped by the addition
of 160 ¡r1 of 30 7. perchloric acid. The precipitated protein was rernoved
by centrifugation. An incubation blank was obtained with the -boiled
enzyrne. The cysteine formed was estimated by the acid ninhrydrin method
of Gaiton¿e (fg6Z). The cysteine forn¡ation in sheep liver samples was
proportional to the enzyme content up to 8 mg protein and to incubation
time for 30 min.
3.2.2.9 Methylmalonyl-CoA ltfutase
Methylmalonyl-C,oA nn:Lase \¡Ias prepared in the dark. Livers were
frozen at -80oC for at least 2 hr before they were homogenized in 10 vol
of 0.25 M sucrose containing 10mMTris-HCl (pH 7.5)' 0.2 nM EDIA
and 0.5 mM reduced glutathione with a Kinematica Polytron homogenizer
(m 10, I-uzern, Switzerland) at full speed for 1 min and
then sonicated for 1 min (tab sonicator set at 100 !ü). The homogenates
hrere stored at -80oC and used for the enzyme assay. The nmtase
activity r^ras assayed in the dark based on the method of Kolhouse and
Allen (L977). The incubation mixture j-n a final volume of 0.1 mI
containing 100 mM Tris-HCf (pH 7.5iù, 5 ¡re/ml antimycin A , 2 mM reduced
glutathione, 40 mM NaCl and 2 nM DL-2-lmethyl-14C]methylmalonyl-CoA
(0.1 pci) was used for the assay of the holo-nmbase activity. I,lhen the
total nn-rtase activity was determined, 100 F¡'l coenzyme 812 was included.

63
After the enzyme was preincubated with or without. coenzyme 812 at 30oC
for 10 mi-n, the reaction vtras initiated by the addition of
methylmalonyl-CoA and stopped after 10 min at 30oC \,rith 250 ¡rI of 0.45 M
perchloric acid. After 3 min at 100"C, the precipitated protein was
removed by centrifugation and 250-¡tI portions of the supernatants
hrere added to 50 pl of 2 M perchloric acid followed by 500 pI of 4 7"
potassium permanganate. After 10 min at 1O0oC, 100 ¡rI of propionic acid,,
hTas added and continued to heat for 5 min, then cooled in ice a¡d
centrifuged. A portion of the supernatant was added to a glass
scintillation vial, dried at 90oC (30-45 min). The radioactivity was
measured as described in Section 3.2.2.L. A blank value was obtained
from the incubation with the boiled enzyme. IJnder the assay
conditions, the rate of the reaction was linear r¿ith the enzyme content
at least up to L20 ¡E protein and with incubation time for 12 min.
3.2.3. Metabolite Assays
3.2.3.t AdoMet and AdoHcy
Tj-ssues were homogenized ín 2-6 vol of 10 7" cold trichloroacetic
acid witha Kinematica Polytron homogenizer at 0"C. The homogenates were
centrifuged at 10,000 x g and 4oC for 15 min. The supernatants v¡ere
extracted 3 times with an equal volume of ether to rernove most. of the
trichloroacetic acid and then filtered through a l,thatman No. 1 filterpaper. Generally, 4-6 ml of the resulting solution was diluted to 10 ml
\^rith 1 nM HCl and then applied to a C,ellex-P (tt * form) colunn
(t x Z cm). The chromatography was carried out. at 5"C according to the
procedure of Eloranta et al. (tglí). AdoHcy and AdoMet. were eluted in

64
5-ml fractions \^rith 50 mM- and 0.5 M HCI, respectively. The measurerrent
of Adotby and Adoltet in the eluates were the same as described by
Eloranta et aI. (1976). The recoveries of these two adenosyl compounds
througþ these procedures, es determined by the addition of 300 nmoles of
conrnercial Adollcy and AdoMet into samples before tissue homogenization,
were abouL 90 7" for AdoHcy and 88 % for Adolufet.
The specificity of the method for different tissues was checked by,
Trc of AdoHcy and AdoMet eluates on cellulose plate r^rith a solvent
system of n-b:tanol/acetic acid/water (60:L5225, v/v)r âs described by
Hoffman (1975). The method was found to be acceptable for all tissues
of sheep, except sheep kidney in vùrich AdoMet was contaninated by a
considerable amount of unlcrown U.V.-absorbing material.' Therefore, in
the kidney samples, AdoMet in the eluates obtained from the C-elIex-P
column was further purified by TI,C-
3.2.3.2 Choline
Free choline and lipid choline containing fractions of milk lrere
prepared as follows. MiIk was ulLrafiltered through a membrane cone
(centriflo cF 25; Amicon, U.s.A.; rnol. wt. ) 251000 excluded) by
centrifugation at 1000 x g and 4oC for 20 min to yield ultrafiltrates
containing free choline. Lipid choline was extracted from milk by the
method of Folch et al. (1957) and hydroLyzed to free choline vdth 6 M
HCI at l1OoC for 48 hr in sealed test tubes. The hydrolysate was taken
to dryness to remove HCl. The residue was redissolved in 10 mM
phoSphr,ate b-rffer (ptt 7.S). The soluti-on was filtered througþ l,lhatman
No. 1 filter paper and the filtrate was used for the determination of
choline.

65
Chroline was determined by a radioenzynratic method, by vrtrich choline
\¡ras corn¡erted to [3U]acetylcholine by choline acetyltransferase.
[3H]acetyl-C,oA was prepared from free CoA and f3H]acetic anhydride,
according to the method of Stadtman (t957), by Þb. R. C. Fishlock. The
incubation medium for the determination of choline were modified from
that described by Hebb et aI. (1975). The reaction mixture in a final
volr¡ne of 50 pI contained 2.5 ¡:moles of sodium phosphate buffer
(pH7.8)r 50nmoles of EDTA, 25pg ofbovine serumalbuminr l0nmoles of
eserine salicylate, 20 nmoles of [3H]acetyl-CoA (approximately 1001000
dEn/nnrol), sample or standard choline (O-Z nmoles), and 10 pl of the
choline aceLyltransferase solution (specific activityi 0.1 ¡:mo1/rnin/ml).
The incubation $¡as performed at 37oC for 60 min. The reaction \¡ras
stopped by the addition of 550 ¡:I of 100 mM sodium phosphate buffer
(pH 7.8) and 300 pl of 2-heptanone containing sodium tetraphenylboron
(ZS ng/nt). Af ter being shaken for 10 min, the two phases \Àrere separated
by centrifugation. By this method of extraction, the radioactive
acetylcholine formed was extracted into the 2-heptanone phase, vùrile
other radioactive compounds, such as [3tt]acetyl-CoA anC
2_['HJacetylcarnitine stayed in the aqueous phase. 150 ¡rl of the
2-heptanone phase was mixed with 3.7 mI of Triton X-1OO/toluene based
scintillat.ion fluid and the radioactivity was determined as described in
Section 3.2.2.L. A blank value was obtained from the incubation
without choline and sample. The formation of acetylcholine was linear
with the amount of choline at. Ieast up to 2 nmoles under the conditions
described. Recovery of choline by this method measured by the addition
of 1 nmole of choline into samples !{as approximately 95 7".

66
3.2.3.3 Creat.ine
For the preparation of protein-free extracts used in the estimation
of creatj-ne in milk, protein was precipitated by the addition of 2 vol
of 0.15 M Ba(OH)2 solution and 2 vol of 0.15 M ZnSO4 solution, according
to the method of Kim et al. (fgA:). Creatine was determined by
a-naphthol diacetyl reaction, according to the method of l^long (1971).
3.2.3.4 Creatinine
. Extraction of creatinine from rnilk was as follows. Protein \¡/as
precipitated by the addition of 8 vol of tungstic acid reagent as
described by Giorgio (I97a). After centrifugation, the supernatant was
filtered through lJhatman No. 42 paper.
One mI of the filtrate or standard creatinine solut.ion was mixed
!ùith 0.1 mI of saturated oxalic acid and 0.2 mI of 10 7" FuIIer's earth
suspension (shaken before use). After being shaken for 10 min, the
mixture was cenLrifuged at 3000 rpxn for 10 min. The supernatant. \das
cornpletely aspirated. Creatinine adsorbed in the Fuller's earth was
eluted by the addition of 0.9 mI of alkaline picrate solution consisting
of 8 mM picric acid and 156 mM NaOH and measured as decri-bed by Flaeckel
(fggf). The reaction was linear with amounts of creatinine from 5 to
40 nnroles under the conditions described.
3.2.3.5 Cysteine
To remove pigment and some interferring compounds in urine in the
estÍmation of cysteine, 1-4 mI urine was diluted to 6 ml with distilled

67
Lrater, adjusted to pH 4-4.5 with HCI to remove CO2 and then
pa.ssed rhrough a colunn (0.8 x 6 cm) of AG 50tn-x8 (H+, 100-200mesh) ar a
rate of 0.5 ml,/min. The colunn was washed rrith 25 ml oL CD2-free water.
The anino acids were eluted wiLh 25 mr of 1 N HCL. The eruate v/as
evaporated to dryness in vacuo. The residue v.7as dissolved in water and
the solution was adjusted to pH 8-8.5 with NaOH. Reduction of cystine
to cysteine by [IlT and the measurement of cysteine were performed
according to the method of Gaitonde (1967). The cysteine analyzed
includes both cysteine and cystine excreLed in urine. Recovery of
cysteine added to urine through this procedure was about 97 7".
3.2.3.6 Formiminoglutamic Acid
Formiminoglutamic acid in urine \¡/as determined by a
spectrophotometric method, as described by (Lranarin and BenneLL (1962).
3.2.4. Determination of Prot.ein
kotein concentrations of enzyme'extracts \{ere measured þ tte Biuret
method of lt.zhaki and GiIl (1964), after precipitation of protein with
10 7" trichloroacetic acid to remove interfering substances. Bovine
serum albumin was used as standard protein.

68
CTIAPTER 4. COI"ÍPARATIVE STUDIES ON BIOSYNTHESIS AND UTILIZATION OF THE
MEIT{YL GROI.JP OF METHIONINE IN SHEEP A}ID RAT TISSUES
4.L Introduction
The important metabolic function of methionine in biological
transmethylation and its central role in methyl group metabolism
are weII recognized. It appears that the amount of methionine required
for various transmethylaLion reactions is far in excess of the
dietary intake (¡tr:¿¿ and Poole , t975; Itudd et al., 1980) and that it
is necessary to conserve the homocysLeine moiety of methionine by the
methylation of homocysteine to replenish methionine. This conservative
metabolism of methionine involves a series of reactions (a cycle)
referred to as methionine recycling, in vùrich the four-carbon moiety of
methionine is successively converted to AdoMet, AdoHcy and homocysleine
(Fig. 2.1). Methyl groups for homocysteine methylation may be derived
either from one of the methyl groups of betaine, a product. of choline
oxidation, or from the one-carbon pool, in which tetrahydrofolate serves
as a carrier for one-carbon units.
It has been shown that most of methionine methyl is utilized for
the synthesis of creatj-ne and choline in manrnals (ttaa and Poole, L975;
Dawson et aI., 1981). These two compounds are also the major forms of
methyl nutrients in diet.s besides methionine. Dietary creatine and
choline can greatly reduce their own synthesis and thus spare the methyl
group of methionine (l,lalker, L979; Skurdal and Cornatzer, I975;
Schneider and Vance, L918). herefore, the requirernent of the methyl
group of methionine would be largely determined by dietary contents of
choline and creatine. There is another methylation reaction cataLyzed

69
by glycine methyltransferase, vùrich is of quantitative irnportance in
the utilization of methionine methyl (¡t¿¿ et aI., 1980; Mitchell and
Benevenga, t976), although the reaction product., sarcosine, has no
physiological function. However, this enzyme has been proposed to play
a significant role in modulation of methionine levelsr or more exactly
AdoÙlet. levels (Ogawa and Fujioka' 1982b).
There is a unique feaLure in methyl group nutrition in sheep.
This species derives a relatively small amount of methyl nutrients from
the diet, owing to almost cornplete degradation of dietary choline (Neitt
et. al., t979; Dawson et a1., 1931) and the nonexistence of creatine in
plants and microbes (l,lalker, f979). Thus, the source of choline and
creatine in sheep would rely ent.irely on their synthesis at the exPense
of the methyl group of methionine. However, microbial protein, a nrajor
source of protein for ruminant nutrition, contains a relatively low
content of methionine, compared to that. of tissue protein (Ørskov,
L1SZ). Nutritional experiments showed that methionine is the first
timiting amino acid in sheep (chalupa, t972; schelling et al., 1973;
Storm and @rskov, 1984). In addit.ion, choline, after its oxidation to
betaine, is an important methyl source for methionine methyl synthesis
j-n most nrannnals. However, the tissue contents of betaine are dependent
largely on dietary choline (finkelstein et aL., t982b; Arvidson and Asp,
Lg82). Such a difference in methyl group nutrition beLween sheep and
most non-ruminant species would indicate that there may be some
unique enzymatic patterns in the biosynthesis and utilization of
methionine methyl in sheeP.
Extensive studies concerning the tissue distrib-ltions of
betaine-homocys teine , 5-methylH4F-homocys teine , phospholipid ,
guanidinoacetate and glycíne methyltransferases as well as choline

70
oxidase have been reported for rats (ninkel-stein et al., L97t; Bremer
and Greenb"rg, L96I; Yanolqrra and Tsukada, L982; Guha and l^legmann, 1963).
The activities of many of these enzymes in rats have been found to be
influenced by dietary methyl nutrients (see Section 2.6.3). However, no
information on tissue distributions of these enzymes in sheep is
available in the literature, except a few reports concerning
the presence of some of these enzymes in sheep liver. Thus, the
metabolic features of methionine methyl for adaptation to the poor
methyl group nutrition in sheep is poorly understood
In an attempt to elucidate the enzymatic patterns of methionine
methyl metabolism in sheep on adaptation to poor nutrit.ional conditions
in respect to methyl groups, the activities of the above mentioned
enzymes were examined in sheep tissues in cornparison with those in rats.
Particular attention viras paid to tissue distributions of two
homocysteine methyltransferasesr âs it has been shown that significant
amotrnts of 5-methylH4F-homocysteine methyltransferase occur in most
tissues of r:ats (finkelstein et aI., L97t). To assess the overall rate
of the slmthesis and utilizaLion of methionine methyl, the rates of in
vitro methionine recycling in the liver and kidney of sheep and rats
r^rere measured and cornpared. The physiological significance of the
results is discussed in respect to the metabolic adaptation of sheep to
poor metl-ryl nutrition.
4.2. Experimental Procedures
4.2.I Animals
Adult Merino ewes were maintained on pasture and adult female

7t
Southl{istar rats were fed a pelleted rat diet (Crarticts, Adelaide
Australia) ad libitum.
4.2.2 Enzyme Preparations and Assays
The preparations and assays 9f betaine-homocysteine,
5-methylH4F-homocysteine, glycine, gi:anidinoacetate and phospholipid
methyltransferases and choline oxidase IÀIere described in Chapter 3 .
4.2.3 Measurement of Methionine Recycling in vitro
Methionine recycling in viLro was determined according to the
method of Barak et aI. (tggZ) by measuring the radioactivities of AdoMet.
incorporated from the [methyl-9H]- and 35S-t.U.tIed meLhionine as the
index of the recycling, since the rqtio of [3H]- to [35s]-AdoMet
represents that of methionine.
Tissue slices, 0.5 to 1.0 nrn thick, were prepared from fed
animals. The slices weighing 1.5 g were placed in 9 mI of Krebs-Ringer
bicarbonate buffer ( pH 7.4 ) containing 6 mM glucose and 50 pM
radioactive L-methionine (O.ZS ¡:Ci fmethyl-3H]methionine and 0.25 pci
[35S]methionine per ml). The slices were incubated under 957" 02. 57" CÐ2
at 37oC with shaking. At the end of the incubalion, the slices
were washed 3 times in ice-cold saline and homogenized in 11 mI of 10 7"
trichloroacetic acid. The homogenate was Lhen centrifuged at 10,000 x g
and 4oC for 15 min. The supernatant was extracted 3 times with an
equal voltrne of ether to remove most of the trichloroacetic acid. The
supernatant, after being filtered through Wlratman No. 1 filter PaPerr
hras applied to a C-eIIex-P (g* form) colunn (f x 7 cm). The

72
chromatography r,iras carried out at 4"C according to the procedure of
Eloranta et aI. (1976). AdoMet was eluted in 5-mI fractions r¿ith 0.5 M
HCI. 1.5 mI of the eluate from the fraction containing the highest
concentration of AdoMet and 50 ¡rI of original incubation medium plus 1.5
mI of 0.5 M HCI were placed in separate scintillation vials.
Then, t2 nL of Triton X-t}O/ toluene based scintillation fluid was added
into the vials. The [3tt] an¿ [35s]contents of various sarnples vrere
assayed in a Rack Beta II Liquid Scintillation Counter, using the
method for double-isotope counting.
In order to determine the radiopurity of AdoMet eluted from Lhe
coltrnn, the fraction containing the highest. concentration of AdoMet
was lyophlLized, dissolved in a small quantity of water and applied
to cellulose coated plates (O.t mn thiclaness). Tbo solvent systems
vùere tested: System A, n-butanol/acetic acid/water (60/L5/25; v/v),
described by Hoffn'nn(1975); System B, 10 mM sodium dihydrogen
phosphate/2-propanoL (L0O/2; v/v) in vùrich Adol'fet was well separated
from some unlanown ninhydrin-posítive compounds usually present in the
AdoMet eluate. It was found that the rat.io of 3tt to 35S after TI,C with
either of the systems was ídentical to the ratio in the original eluate
from the colurnn, although in the samples from kidney AdoMet was
contaminated with unloown U.V.-absorbing materials.
Ttre index of methionine recycling was calculated by the following
equation:
ratio or [3H] to ¡35s1aaoMet in tissue
index of methionine recycling - 1 -
ratio of [3u] to [35s]mettrionine in
original incubation meditrn
As methionine recycling occurs in tissue with time, the [methyt-3U]

73
groups are repl-aced by unlabelled methyl groups, but the 35S t"U"t
remains unchanged. Thus, an index of methionine recycling with a value
of 0 indicates no recycling and progressively greater values denote
increasing degrees of the recycling.
4.2.4 Other Assays
The concentrations of protein and tissue levels of AdoMet r^/ere
determined as described in Chapter 3 .
4.3. Results
4.3 1 C-omparison of the Specific Activities of Erzymes
in Sheep and Rat Tissues
As shown in Table 4.1, 5-methylH4F-homocysteine melhyltransferase
\,{as widely distributed in all tissues of sheep and rats. The highest
specific activity of this enzyme was found in rat kidney, which is in
consistent with the observations of Finkelstein et aI. (L97L), vÌrile in
sheep the relatively high activity was present in the pancreas and
Iiver. By comparison, it was found that. the activity of
5-methylH4F-homocysteine methyltransferase in most tissues of sheep was
significantly higher than those of rats. Particularly, the enzyme
activity in sheep liver was 3.5-fold higher than that. of rat liver. Itis worth noticing that rmrscle, the largest tissue mass of the body, also
contained a significant level of the enzyme and that the enz¡rme level in
sheep nmscle was 2.5-fold higher than that of rats. Obviously, the
results indicate that the total body capacity of methionine synthesis
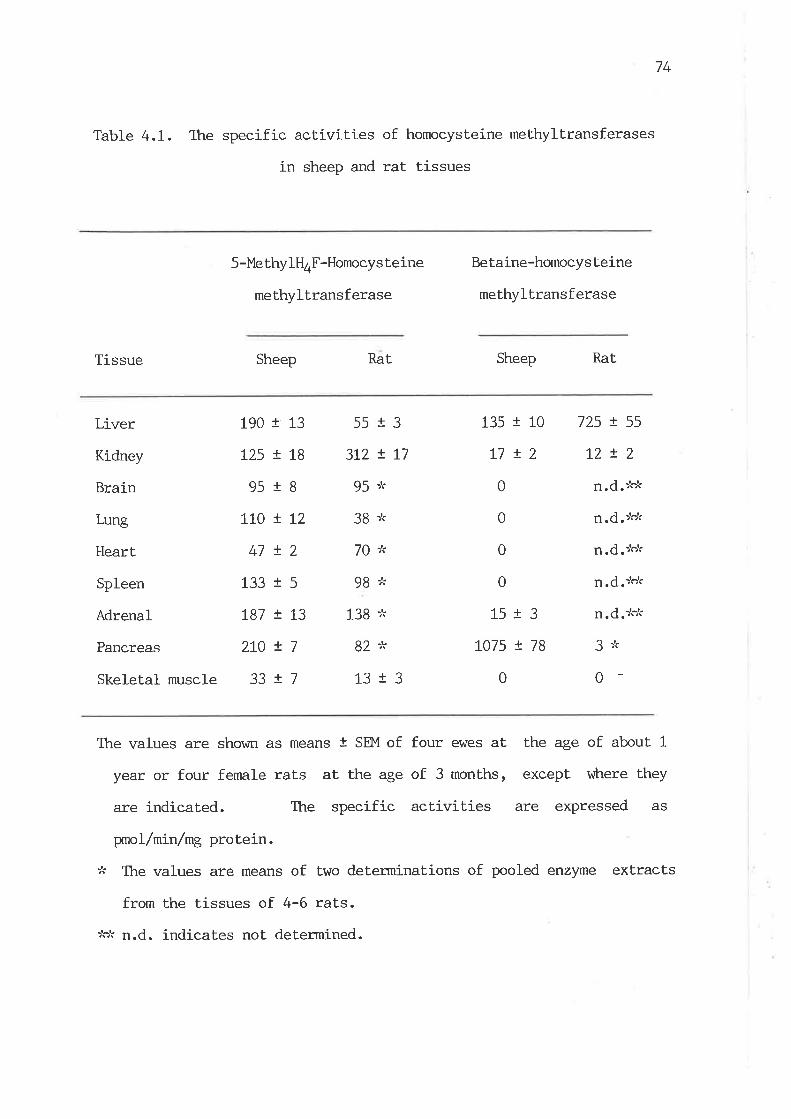
74
Table 4.1. The specific act.ivit.ies of homocysteine meLhyltransferases
in sheep and rat tissues
Tissue
5-Me thylH4F-Homocys teine
methyltransferase
Sheep Rat
Betaine-homocysteine
methyltransferase
Sheep Rat.
Liver
Kidney
Brain
Lung
Heart
Spleen
Adrenal
Pancreas
Skeletal muscle
135 t 10
L7 !20
0
0
0
15r3L075 ! 78
0
725 ! 55
t2!2¡.d.:'cJ<
¡.d.:'c)r
n.d.>'ok
¡.d.:'.r'r
¡.d.:'.r'c
3tk
0
190
L25
95
1_10
47
133
!87
2L0
33
r13
r18
tg
!12+)
+q
r13
!7+1
55
3t2
95
38
70
9B
138
82
13
l3L 1'7!Lt
+?
The values are shown as means t SEM of four ewes at Lhe age of about 1
year or four fenrale rats at the age of 3 months, except vrhere they
are indicated. The specific activities are expressed as
WoL/nLn/mg protein.
:k The values are means of two determinations of pooled enzyme extracts
from the tissues of 4-6 rats.
:'.rk ¡.d. indicates not determined.

75
by S-methylH4F-homocysteine methyltransferase per unit of body weight
in sheep is considerably greater than that in rats.
Table 4.1 also shows that Lhe distribution of betaine-homocysteine
methyltransferase in sheep is sLrict.ly confined to a few tissues as
observed in rats by Finkelstein et al. (t:glt). Surprisingly, the
highest. activity was found in sheep pancreas r¡Èrere the specific activity
vras 8-fold higher than that of sheep liver, vùrile in rats the
significant activity \,vas found only in the liver. The level of this
enzyme in sheep liver was 5.4 times lower than thaL in rat liver.
Although sheep pancreas contains the highest activity of the enzyme,
the absolute amount of methionine synthesized in this .tissue could not
be expected to be much because the relative weight of pancreas in sheep
is small, only about 4 7" of that of the liver. Therefore, the total
body capacity of betaine-homocysteine methyltransferase for methionine
synthesis in sheep is st.ill much smaller than that of rats on the basis
of body weight.
TabLe 4.2 shows that the tissue distribution of AdoMet-dependenl
methyltransferases and choline oxidase in sheep and the corresponding
enzymes in rat liver for comparison. All these enzymes vüere.present in
the liver, pancreas and kidney of sheep. It \,ùas found that the
specific activities of glycine methyltransferase and guanidinoacetate
methyltransferase in sheep pancreas vùere many times higher than Lhose of
the liver, vùrereas in rats the highest activities of these enzymes are
present in the liver (Kerr, L972; van Pilsum et aI., 7972; Yanokura and
Tsukada, 7982). Comparison of these four hepatic enzymes between sheep
and rats shows that the levels of these enzymes were much lower in sheep
liver than those in rat liver (taUle 4-2). In particular, the specific
activities of choline oxidase and glycine methylLransferase in sheep
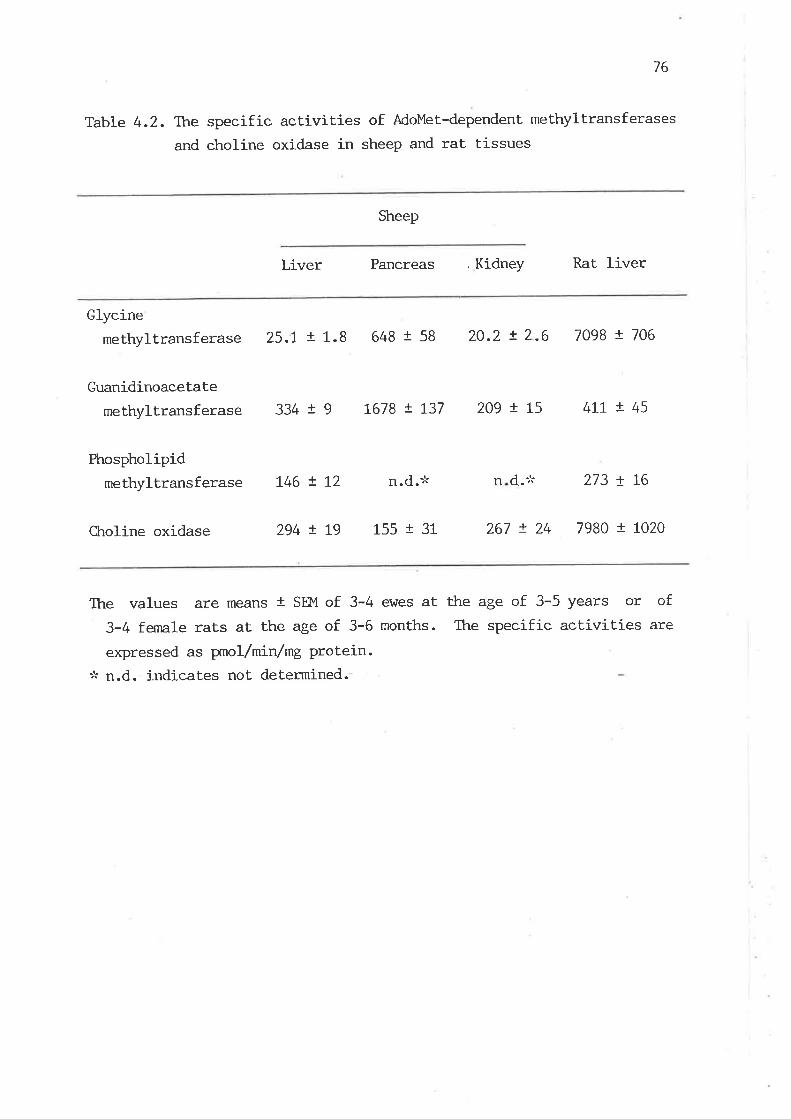
76
Table 4.2. Tþre specific activities of AdoMet-dependent methyltransferases
and choline oxidase in sheep and rat tissues
Sheep
Liver Pancreas , Kidney Rat liver
Glycinemethyltransferase 25.I ! t.8 648 t 58 20.2 ! 2.6 7098 t 706
Guanidinoacetate
methyltransferase
Phospholipidmethyltransferase
334 r 9 t678 ! t37 209 ! L5 4rL ! 45
L46 ! L2 n.d.,k n d.* 273 ! L6
Chroline oxidase 294 t t9 155 t 31 267 ! 24 7980 t 1020
The values are means t SEM of 3-4 ewes at the age of 3-5 years or of
3-4 femate rats at. the age of 3-6 months. The specific activities are
expressed as prnol/min/ng protein.:k n.d. indicates not determined.

77
liver hrere 27 Eimes and 284 times lower than those in rat liver,
respectively. The activities of these four enzymes in other tissues of
sheep were not examined, as the data obtained frorn other species of
animals show ttrat the Ievels of these enzymes are generally very low,
if present at aII, in other tissues (see Literature Review).
4.3.2 Comparison of Methionine Recycling in Sheep and Rat Tissues
In order to minimize the effect of the pool size variability
on tissue methionine specific radioactivity, v¡?rich affects the apparent
rate of methionine recycling, 50 ¡rM methionine was included in the
incubat.ion medium for the purpose of the cornparat.ive study. AdoMet
Ievels in the liver of sheep and rats were also measured and found to be
similar (Ag f 3 and 7I r 4 nmol/g wet, tissue in sheep liver and rat
liver, respectively). Therefore, the compari-son of the rates of
methionine recycling should reflec! the true difference between sheep
and rats.
Data in Fig. 4.1 show that methionine recycling in kidney -slices
steadily increased throughout a 60-min period. However, the curve
obtained in Iiver slices was linear only within 30 min, and then a
decreased rate was observed. Therefore, a 30-min period of incubation
was used for cornpa.rative studies.
Itre indices of methionine recycling measured in the tissues of
sheep and rats are shown in Table 4.3. The rates of methionine recycling
in both tissues of sheep were lower ttran the corresponding tissues of
rats, bJt the difference was only significant for hepatic tissues
(p < 0.01). It can be seen that the rates of the recycling in the
livers of both species were nmch faster than those in the kidneys.

78
o.4olcoooo.Ec.9-coE
oxo1t.g
o.5
o.3
o.2
O.1
20 40Time (min)
60
Fig. 4.1. Time course of meÈhionine recycling in tissue slices. Each
point represents the mean of two animals with duplicate assays. Tissue
slices $/ere prepared from l-year-old ewes or 3-month-old female rats.Symbols: Or rat liveri A I rat kidney; O, sheep liver; trsheep kidney.

79
Table 4.3. C,onrparison of methionine recycling in the tissue slices
of sheeP and rats
Tissue
Index of methionine recYcling
Sheep Rat
Ratio of the index
of rat to sheep
Liver 0.16 t o.o1 (4) 0.44 ! o.o2 (4)
Kidney O.os t 0.01 (3) 0.10 t 0.02 (3)
2.8
2.0
The values are means i SEM rrith the nurnber of animals,in parentheses.
Index of methionine recycling was measured after 130-min incubation.
Tissue slices \^/ere prepared from l-year-old ewes or 3-month-old female
rats.

80
4.3.3 Effects of Metabolites on Methionine Recycling ín Liver Slices
As seen in Table 4.4, the addition of betaine, a substrate of
betaine-homocysteine methyltransferase, gave a sna1l but significant
stinnllation on methionine recycling in the liver of both species, wìrile
the addition of formate and tetrahydrofolate v¡hich are precursors of
5-methylH4F, a substrate of S-methy1tl4F-homocysteine methyltransferase,
had no effect on the hepatic methionine recycling.
4.4 Discussion
The present study demonstrates that sheep liver possesses a nmch
lower level of betaine-homocysteine methyltransferase and a higher level
of 5-methyltl4F-homocysteine methyltransferase, compared with rat liver.
5-Methylt-l4F-homocysteine methyltransferase activity in most non-hepatic
tissues of sheep is also higher than that in the corresponding tissues
of rats. Clearly, sheep possess a relatively large capacity of
methionine methyl synlhesis from 5-methylH4F. This can be considered as
a compensatory mechanism for methionine synthesisr âs the body capacity
of betaine-homocysLeine methyltransferase in sheep is relatively low. In
view of the relative weight of tissues, the potential capacity of
5-methylH4F-homocysteine methyltransferase for methionine synthesis in
non-hepatic tissues of sheep is of significance. Particularly, in sheep
nn:scle the total ti-ssue capacity of methionine slmthesis by
5-methyltl4F-homocysteine methyltransferase was approximately 1.5-fold
that of the liver since the relative weight of the nn:scle is about
2O-fold that of the liver, although the enzyme activity in this tissue
was 13 times lower than that in the liver v¡tren the data are expressed

81
Table 4.4. F,f.fects of metabolites on methionine recycling in liver slices
Index of methionine recycling% of control
Additive Sheep Rat
None 100
Betaine (e .S rnt'l) 123 t e''. (4) 73L t 5:'.:'r (4)
Fornnte (Z.O rnU) an¿
tetrahydrofolate (0.1 nM) 110 r 4 (4) e813 (3)
Ttre results are expressed as percent index of methionine recycling,measured in liver slices after 3O-rnin incubation. The tissue slicesr^rere prepared from 1-year-old ev/es or 3-month-o1d fe¡nale rats. The
control values for sheep and rats are 0.16 t 0.01-and O.44 ! 0.02,respectively. The concentrations of metabolites added and the nurnber
of determinations for each condition are given in parentheses.
Significance was deterrninined for the effect. of additive as compared
to its respective control, using Studentrs paired t-test.:'r p < 0.05.:'c,'r p ( 0.01.
100

82
as nnrot/min/g wet tissue (f .: and 17.0 nnrol/min/g \^¡et tissue in the
muscle and liver, respectively). Comparison of the enzynratic patterns
of homocysteine methylation between the two species clearly shows that
sheep rely nn¡ch more heavily on 5-methylH4F-homocysteine
methyltransferase for methionine synthesis than rats.
The low activity of betaine-homocysteine'methyltransferase in sheep
seelns to be related to the very low activity of choline oxidase in
this species, since betaine in mannnalian tissues is mainly derived from
choline. The high ratios of these two enzyme activities of rats to sheep
(table 4.5) should be a good indication of the slow rate of choline
oxidation in sheep. The conservation of choline apPears to be an
irnportant. mechanism for adaptation of sheep to the Poor choline
nutrition. ,
By comparison of the activities of the three AdoMet-dependent
methyltransferases between sheep and rats (table 4-5), it was found that
the ratio for the specific activity of glycine methyltransferase in rat
Iiver to sheep liver was 284, vfrrile the ratios for guanidinoacetate
methyltransferase and phospholipid methyltransferase were 1.2 and t.9,
respectively. These findings suggest that glycine methyltransferase
activity in sheep liver may be reduced to an extremely low level in
order to conserve the methyl group of methionine for physiologically
significant methylaLion reactions, since the physiological significance
of sarcosine, the product of glycine methylation, is unknown so far and
it is generally considered that the function of this enzyne is solely to
dispose excess methionine (Qgawa and Fujioka, 1982b).
Despite no dietary creatine and negligible amounts of dietary
choline available to sheep, the levels of guanidinoacetate and
pLrospholipid methyltransfearses in sheep liver were still lower than

83
Table 4.5. Ratios of the specific activities of hepatic enzynres
of rats to sheeP
Êrzyme Ratio
Betaine-hornocys teir,re methyl Lrans f erase
5-MethylffuF-homocys teine methyltrans f erase
Glycine methyltrans f erase
Guanidinoacetate methyltransf erase
Phospholipid methyltrânsferase
CLroline oxidase
5.4
o.29
284
r.2
L.9
27
Ttre values fo-r the speeific activities of these enzymes are shown
in Table 4.L-4.2.

84
those in rat liver. Ttris nay reflect a general metabolic pattern that a
species of animals with a large body size tends to possess a slower
metabolic rate per unit of body weight than a small species of animals.
The ratio of basal metabolic rates of rats to sheep was found to be
about 3.5 on the basis of body weight (Uitctrett ' L962). However, the
differences in the activities of these two enzymes between sheep and
rats were found to be small. If the values for the rat.ios of the hepatic
enzyme activities of rats to sheep (laUte 4.5) are divided by a factor
of 3.5, Lhe capacities for creatine and choline synthesis in sheep liver
will be greater than those of rats on the basis of basal metabolic rate.
Yet the ratios of rats to sheep for other enzymes will still indicate
thaÈ the hepatic capacities of betaine-homocysteine methyltransferase,
glycine methylLransferase and choline oxidase in sheep are much smaller
than those in r"r", v¡hereas the hepatic capacity of
5-methylH4F-homocysteine methyltransferase in sheep will be even greater
than that of rats.
The relative overall rate of the reactions involved in the slmthesis
and utilization of methionine methyl can be assessed by means
of comparison of the rates of methionine recycling beLween the
two species. Ttre results demonstrate that the recycling rates
r,vere considerably lower in sheep liver than in rat liver. Ttris is in
close agreement with the total enzymatic capacity in meLhionine
recycling reactions in the liver of the two speciesr âs the total
capacities of predominant AdoMet-dependent methyltransferases and
homocysteine methyltransferases \ilere Lower in sheep liver than
rat liver. The results also show that the capacity for such
demethylation and remethylation reactions of methionine in liver is nmch
greater than that of kidney in both species. These observations

85
directly demonstrate that liver is a quantitat,ively irnportant organ in
the synthesis and utilization of methionine methyl. The data in
Table 4.3 do not allow quant.itative evaluation of the rate of methionine
recycling in tissues, but do indicale that conservative metabolism of
methionine occurs very actively in both species, as 30-min incubation
results in a significant portion of the hornocysteine moiety of
methionine reappearing in AdoMet molecules in tissues, particularly in
IiVer. l,fudd and his coworkers (1975, 1980) have shown that the average
homocysteine moiety in hunans Æn normal diets cycles between
methionine and homocysteine about 2 times before being converted to
cystathionine. Finkelstein and Þfartin (fgAe) have regently shown that
63 7. of homocysteine goes through the remethylation route in the liver
of rats fed a 0.3 7" methionine diet. 1lhese findings highlight that
homocysteine methylation is of quantitaLive irnportance in the supply of
the methyl group of methionine for AdoMet-dependent methylation
reactions in manrnals.
Ttre present study demonstrates that betaine could stimulate the
hepatic methionine recycling in both species, but formate plus
tetrahydrofolate, the precursors of 5-methylH4F, could not. The data
indicate both that the betaine enzyme is not saturated rfith endogenous
betaine in the tissue slices and that the enzyme plays an important role
in hepatic methionine synthesis. By calculation of the ratios of the
hepatic capacity of 5-methylH4F-homocysteine methyltransferase to
betaine-homocysteine methyltransferase, it was found tttat t'he rati-os
were !.2 in sheep (the specific activity expressed as nmol/min/g wet
tissue was 17.0 for 5-methylH4F-homocysteine methyltransferase vs. 13.8
for betaine-homocysteine methyltransferase in sheep liver) and
0.06 in rats (4.7 vs. 74.4 in rat liver). This suggests that

86
5-methylH4F-hornocysteine methyltransferase may make an equal
contrib-ltion to the hepatic methionine synthesis in sheep r^rith the
betaine enzyme if other conditions for these two enzymatic reacti-ons in
vivo are similar, r,¡hereas the methionine synthesis in rat liver is
attrih-rted nninly to betaine-homocysteine meLhyltransferase if dietary
choline is not limited. Finkelstein (L979) has shown that the affinity
of the betaine enzyme for homocysteine is relatively higher than that of
the 5-methylH4F enzyme in rat liver, although the Km values of these two
enzymes from sheep are unlmown. Nevertheless, methionine recycling
observed in kidney is virtually dependent on lhe action of
5-methylH4F-homocysteine methyltransferase, since Ëhe contents of
betaine-homocysteine methyltransferase are extremely low in the kidneys
of these two species.
Observations on the activiÈies of these enzymes in sheep pancreas
are striking. Unlike monogastrics, the levels of betaine-homocysteine
methyltransferase, glycine methyltransferase and guanidinoacetate
methyltransferase in sheep pancreas were nn:ch higher than those in the
Iiver. However, the significance of the high levels of these enzymes in
sheep pancreas is unlanown.
In conclusion, the cornparative studies between the two species
reveal ttrat the overall rate of the synthesis and utilization of
methionine per unit of tissue weight is lower in sheep than in rats. The
enzymatic capacity for methyl group catabolism in sheep is extremely
low, compared with that in rats. The relatively low capacity of
betaine-homocysteine methyltransferase in sheep is likely due to the
slow rate of oxidation of choline, v¡trich nny tle a result of adaptation
to the poor choline nutrition in this species. Ttre nmch higher capacity
of 5-methylH4F-homocysteine methyltransferase in sheep, as comPared \fith

87
rats, can compensate for the relatively snnll amount of methionine
methyl synthesized from the betaine-homocysteine methyltransferase
re¿ction. Thus, the de novo synthesis of methyl groups is a very
important source of labile methyl groups in sheep, r,ihich rnay be a
response of sheep to a relatively low intake of methyl nutrients.
The questÍ-on raised is vÈrether these metabolic features of sheep
represent only the characteristics of a species or are formed in
response to the nutritional conditions of methyl groups. Thusr-further
investigations are needed to elucidate the effect of nutritional changes
of methyl groups on these enzynntic patterns in sheep. These will be
presented in the next chapter.

8B
CHAFTER 5. DEVEI-OPMNVIAL CHANGES IN THE ACTIVITIES OF ENZYMES RHATED
TO METTIYL GROIJP ME'IABOLISM IN SHEEP TISSUES
5.1 Introduction
The data presented in (trapler 4 show that sheep appear to possess
a greater enzymatic capacity for the de novo synthesis of methyl groups
and a lower capacity for methyl group catabolism, as compared with rats.
It seems likely that these features of methyl group metabolism might be
related to a low intake of methyl nutrients in adult sheep, mainly due
to the almost complete degradation of dietary choline by rumen microbes
(neitt eL aI., 1979; Dawson et al., 1981) and the lack of creatine in
plants and microorganisms (Walker, L9l9). However, milk is rich in
these methyl nutrj-ents (Zeisel, L98!; Macy and Kelly, 1961) and they are
thus available for pre-ruminant lambs. Therefore, it would appear that
there is a vital ctrange in methyl group nutritionr âs sheep plogress
from a pre-nminant to a ruminant state. Such a change might lead to
some alterations in the enzynatic patterns of methyl group metabolism.
In the present investigation, the develognental changes of a number
of enzymes related to methyl group metabolism were studied in sheep to
reveal the mechanisms of adaptation to nutritional changes of methyl
groups from the pre-nrminant to the mminant state.
5.2. Experimental Procedures
5.2.I Animals
Merino sheep of various ages vtere used. Each group of lambs

89
contained both sexes. The animals were slaughtered at the following
ages: ferus (last 2L-25 days of gestation), 1-day-old (Z+-SZl'v
postpartum), lO-day-old, 4O-day-old, 90-day-o1d and 3- to 5-year-old.
The 4Q-day-old group of Iambs had suckled only ewets milk without access
to any grass or solid food in order to keep the lambs in the
pre-ruminant state. gO-day-o1d lambs had been weaned for about 5 weeks
and had well-developed rumen systerns. ltre weaned and mature sheep were
nnintained on pasture.
5.2.2 Enzyme Preparations and Assays
The preparations and assays of betaine-homocysteine
methyltransferase, 5-methylH4F-homocysteine methyltransferase, glycine
methyltransferase, guanidinoacetate methyltransferase and phospholipid
methyltransferase, and choline oxidase and cystathionine ß-synthase
are described in Chapter 3 .
5.3. Results
5.3.1 Developrnental CLnnges of Oroline Oxidase
Fig. 5.1 shows that the level of choline oxidase was low in
prenatal and adult sheep liver. lhe specific activity of the enzyme \^ras
postnatally increased in pre-ruminant lambs at least until 40 days of
ager b¡t a subsequent decrease in the enzyme actiwity was observed v¡hen
the animals reached the ruminant state.
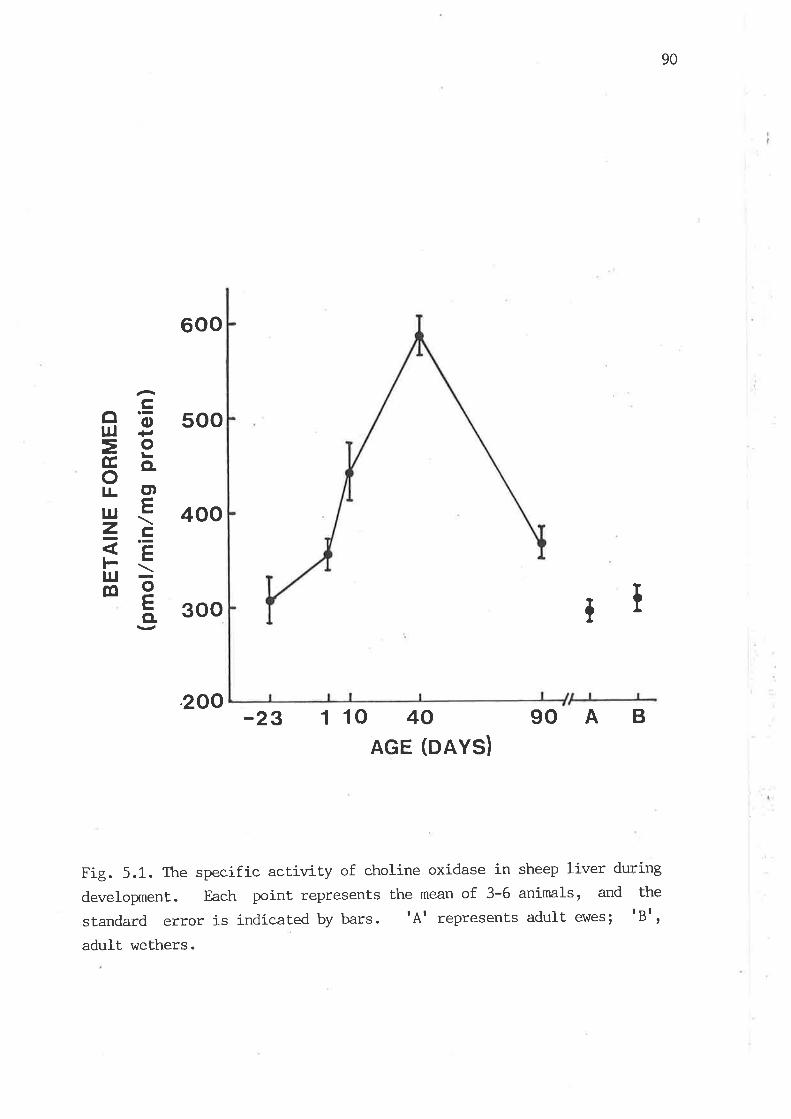
90
?ffþ=95õ.]L ct)cru.t-zè.-
FÈg
600
500
400
300 TI
200 90AB-23 1 10 40AGE (DAYS)
Fig. 5.1. The specific activity of eholine oxidase in sheep liver during
developrnent. Each point represent.s the mean of 3-6 animals, and the
standard error is indicated by bars. tAt represents adult e\47es; tBt,
adult wethers.

9l
5.3.2 Developmental Ctranges of Homocysteine-utilizing enzynes
Fig. 5.2 shows that the specific activity of pancreatic
betaine-homocysteine methyltransferase increased with age and reached
the adult level at the age of 90 days. The specific activity of the
hepatic enzyme was also found to increase with age in pre-ruminant
lambs, but a marked decrease in the hepatic enzyme activity in
post.-mminant larnbs and adults was observed. It can be seen in Fig. 5.3
that the developxnental changes in the specific activity of hepatic
5-methylH4F-homocysteine methyltransferase show a pattern opposite to
the ckr,anges of hepatic betaine-homocysteine methylLransferase, virile the
pancreatic enzyme level did not show significant changes during
developnent. No significant developnrental changes in the cystathionine
8-synthase level was observed in sheep liver and pancreas (fig. 5.4).
5.3.3 Developrnental Changes of AdoMet-dependent Methyltransferases
As shown in Fig. 5.5, the hepat.ic phospholipid methyltransferase
level in fetuses and l-day-old larnbs was much lower than that in adults.
The specific activity of the enzyme in lO-day-old lambs was increased
4-fold, as compared with that in l-day-old lambs. Thereafter a slight.
decrease in the enzyme acLivity during maturation vras seen. The
dramatic developmental changes in the specific activity of glycine
methyltransferase are shown in Fig. 5.6. The enzyme level was very low
in both prenatal and adult sheep liver. The specific activity of the
hepatic enzyme in l-day-old lambs increased 55-fold as compared with
that of fetuses and was 13-fold higher than that of adults. Thereafter
a rapid decrease in the enzyme level with age followed. However, the
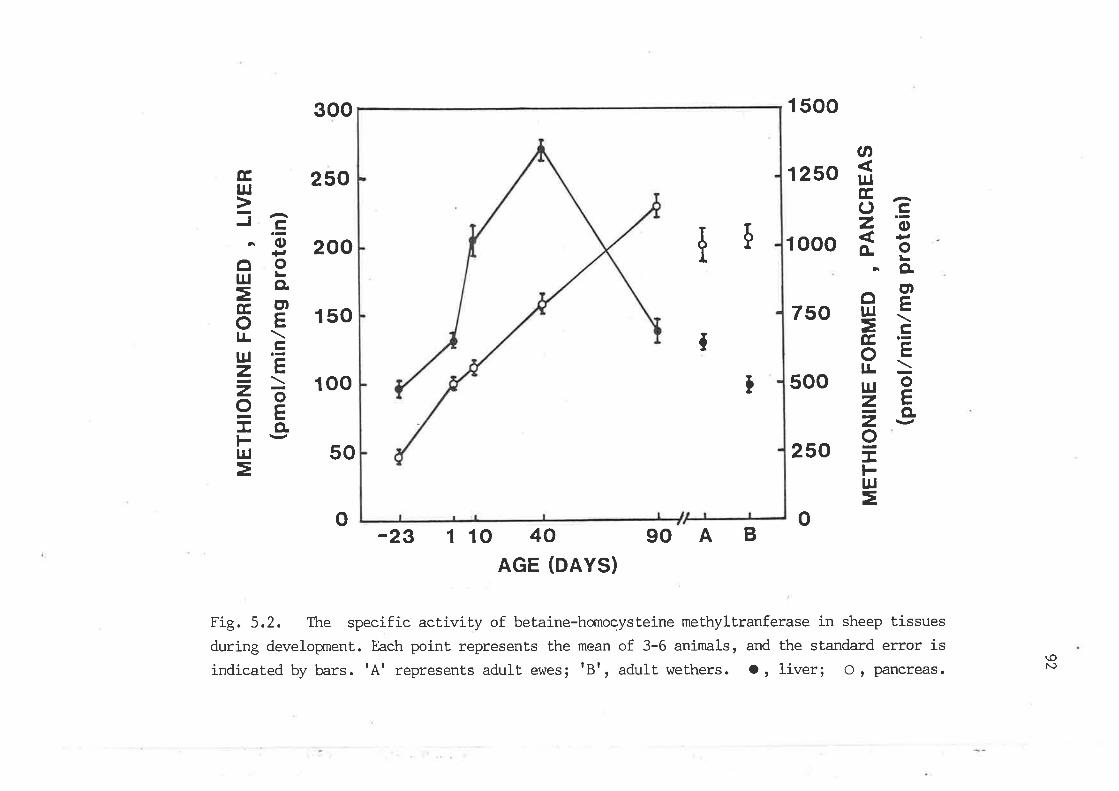
t
Þ
T
I
300 1 500
250 1 250
200 1 000
150 750
100 500
50 250
o-23 1 10 40
AGE (DAYS)
gOAB
Fig. 5.2. The specific activily of betaine-homocysteine methyltranferase in sheep tissuesduring developrnent. Each point represents the mean of 3-6 animals, and the standard error isindicated by bars. tAt represents adult evres; tBt, adult wethers. o , Iiveri o r pancreas.
.n
]UÊ,^ocz'õiE
-CLogI¡J .L
=è8Sr¡J 9zts-A É4\/o.Lt-ul
=
Ê,UJ
-^Je
' 'õl+,oor¡J =
=;otsr!>lu '=z.E<-AoË:ECLl- \/l¡J
=
o
\oN)

93
oG,ËsÉ,oPä
ct)l¡J ç-b1\
=, e,
9El-ou¡tr=õ.
300
250
200
150
100
50
-23 1 10 40AGE (DAYS)
gOAB
Fig. 5.3. The specific activity of 5-methylH4F-homocysteine
methyltransferase in sheep tissues during developxnent'. Each point
represents the mean of 3-6 animals, and the standard error is indicated
by bars. tAt represenLs adult ehles; tBt, adult wethers. o, Iiver;O, pancreas.
5I
tt
o

94
Þ40
oUJ
=ã6E 30tLO¡-ucLz ct)z E 20o>-f-+trF<sõ-a E 10
o
I
t t
o-23 1 10 40
AGE (DAYS)90AB
Fig. 5.4. The specific activity of cystaLhionine ß -synthase in sheep
t.issues during developnent. Each point represents the mean of 3-6animals, and the standard error is indicated by bars. tAt represenLsadult e\,ües; tBt, adult wethers. o, liver ; o, pancreas.
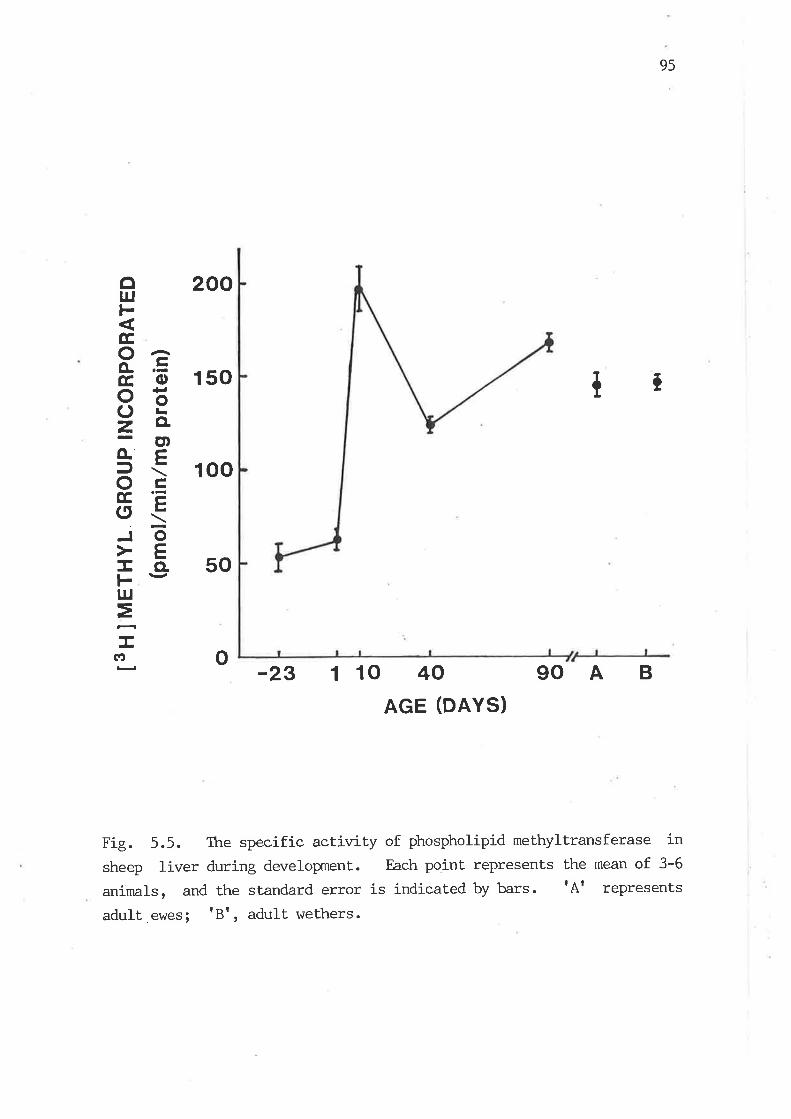
o
ol¡Jl-Ê,Oão. .=Ê,ooã2ä
ct)o-ÊJ\oÈG.E(5<Jõ:EcL.\/Ft¡J
==Ic¡
95
20o
150 I7
100
50
-23 1 10 40AGE (DAYS)
90A B
Fig. 5.5. The specific activity of phospholipid methyltransferase insheep liver during developnent.. Each point represenLs the mean of 3-6
anímals, and the standard error is indicated by bars. tAt represents
adult,ewes; tBt, adult wethers.
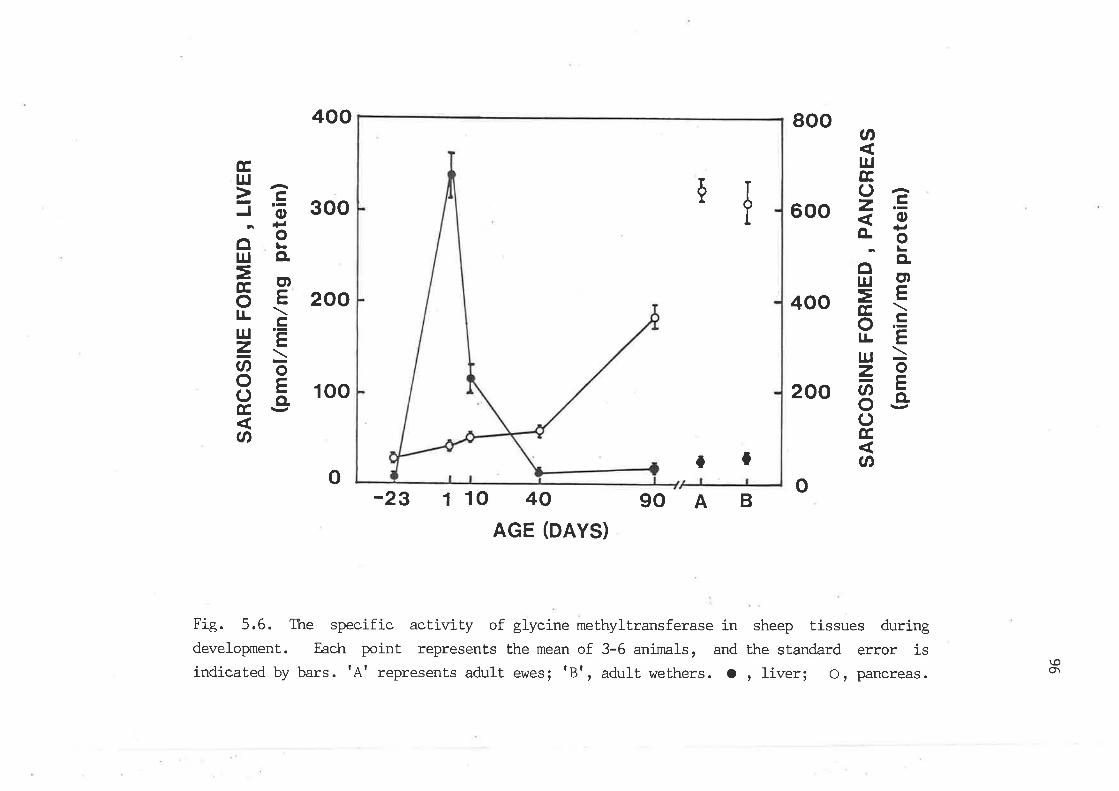
Þ
tt
400 800
300 600
20o 400
100 200
o-23 1 10 40
AGE (DAYS)90AB
Fig. 5.6. The specific activity of glycine methyltransferase in sheep tissues duringdeveloprnent. Each point represents the mean of 3-6 animals, and the standard error isindicated by bars. tAt represents adult e\des; tBt, adult wethers. o , liver; o, pancreas.
Øl¡JEOã-DEI
f.3¡L
ffr'=tsÉ,>o.=l!tt¡l >=O
8ëoÉ,
Ø
Ê,
5EJO
¡ùlo9I¡J Ét
ËeotslrèUJ 'E-, t-É\
8Eoã?l' v-
Ø
o
\oOt

97
specific activity of pancreatic glycine methyltransferase \'\7as found to
increase with age. The developxnental increase in the specific
activity of guanidinoacetate methyltransferase \,ùas also found in sheep
pancreas (Fig. 5.7), v¡trile no changes in the hepatíc enzyme level were
observed.
5.4 Discussion
The present investigation demonstrates postnatal increases in the
Ievels of choline oxj-dase and betaine'homocysteine methyltransferase in
pre-ruminant lamb liver and subsequent decreases v¡hen the lambs reached
the ruminant state. However, the magnitude of increase in the specific
activity of choline oxidase with age in pre-ruminant sheep \'Ùas much
smaller than that in rats, vùtere the enzyme activity after reaching the
highest ì evel at- tle age of 40 days \^/as maintained throughout further
developrnent (Zeizel and I,üurtman,1981). Increase in the specific activity
of betaine-homocysteine methyltransferase with age in pre-ruminant lamb
Iiver showed a different pattern from the developnental changes of the
enzyme activity observed in rats, it v¡hich the enzyme level slightly
d.ecreases with age (Finkelstein et aI., t97L). This difference may be
due to the fact that the hepatic betaine'homocysteine methyltransferase
activity in fetuses and newborn lambs was at a nmch lower level than
that in rals and was subsequently inducqd by the ar¡ailability of milk
choline. The low levels of choline oxidase and betaine-homocysteine
methyltransferase in adult sheep liver can be considered as a
consequence of adaptation to the low availabilily of dietary choline'
These results suggest that the availability of choline regulates the
activity of choline oxidase as well as betaine-homocysteine
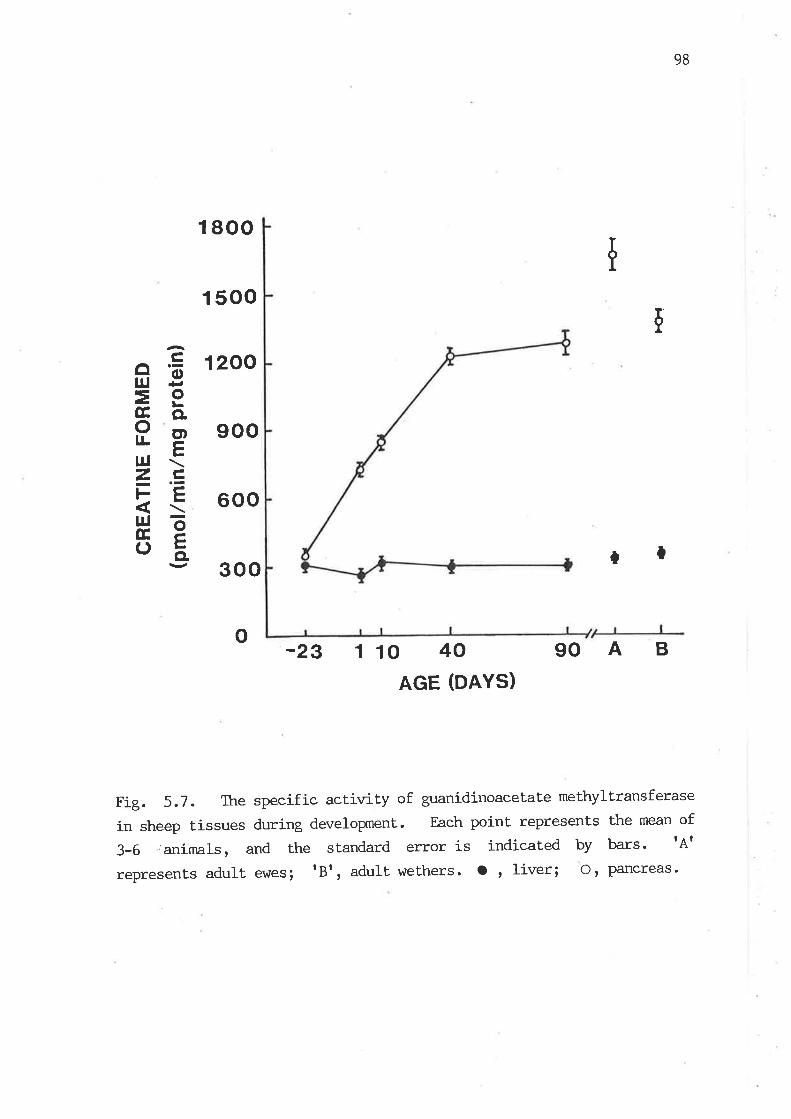
98
1 800
I 500
I 200
900
600
300 rl
-23 I 10 40AGE (DAYS)
gOAB
Fig. 5.1. The specific activity of guanidinoacetate methyltransferase
in sheep tissues during developxnent. Each point represents t'he mean of
3-6 'animals, and the standard error is indicated by bars' tAt
represents adult e\^JeS; tBt, adult wethers. O , Iiver; Or Pancreas.
I
le
ffB=9Ê, õ.Pel¡r <Zc,l'- Et¡¡ =É,=oã
o

99
methyltransferase in sheep. It is worth notice that lowering the raLe of
choline oxidation in post-ruminant sheep would conserve methyl grouPs
in the body, since only one of the methyl grouPs in the choline molecule
after its oxidation can reenter the labile methyl group Poolr vilrile the
synthesis of choline involves a transfer of three methyl groups from
AdoMet.
The decrease of the specific activity of 5-methylH4F-homocysteine
methyltransferase with age in pre-ruminant lamb liver presents a similar
pattern to the developrnental change of the enzyme activity observed in
human liver (Uuaa et a1., t97O; Sturman and Gaull, L978; Kalnitsky et
al., tg82) and rat liver (n'inkelstein et a1., t97t). But. following
this decrease, the specific act.ivity of the hepatic enzyme in
post-ruminant sheep was markedly increased, vilrereas in adult humans and
rats there is no such rise observed. A close examination of the
developmental pat.terns of the two homocysteine methyltransferases
reveals that changes in the level of hepatic 5-methylH4F-homocysteine
methyltransferase during develoçxnent shows an inverse relationship with
those of hepatic betaine-homocysteine methyltransferase in sheep. As
these two methyltransferases play a similar role in methionine synthesis
and the need of methionine for numerous biological methylation reactions
is far in excess of its dietary intake (Uu¿¿ and Poole, 1975), lowering
the level of hepatic betaine-homocysteine methyltransferase in ruminant
sheep would cause a crucial problem in naintaining the physiological
Ievel of methionine, if the activity of 5-methylH4F-homocysteine
methyltransferase were not accordingly increased. These findings
suggest that the' elevatj-on of the actiwity of hepatic
5-methylH4F-homocysteine methyltransferase in sheep serves as a means to
compensate the low capacity of betaine-homocysteine methyltransferase in

100
methionine synthesis in order to maintain the physiological level of
methionine. Tttus, the de novo synthesis of methyl groups becomes a
predominant route in the synthesis of the methyl group of methionine in
sheep under the condition of poor choline nutrition.
The specific actiwity of cystathionine ß-synthase in sheep showed
no significant changes during developxnent. However, it was reported
that, in rats and humans, the enzyme level increases with age
(Finkelstein, t97O; Volpe and laster, t972; Sturman and GauII, 1978).
Radcliffe and Egan (1974) found that the specific activity of
v-cystathionase, which catalyzes the conversion of cystathionine to
cysteine, in 2-day-old lamb liver is about 30 times higher than that in
adult sheep. It is lcrown that the availability of dietary methionine is
Iower in adulL sheep than Lhat in milk-suckling larnbs, as ttte methionine
content of microbial protein for the post-ruminant animals is relatively
low (Storm and prskov, L984; Ørskov, 1982). Thus, if f-cystathionase is
an irdícatiræ szyre of tle rate of lulnq¿steire catabolism in sheep, it would be
reasonable Lo assume that in adult sheep more homocysteine is conserved
for methionine synthesis by lowering the catabolic rate of homocysteine.
The developrnental pat.tern of hepatic phospholipid methyltransferase
in sheep was similar to the observations of Pelech et al. (1983) in
rats. They suggested that a postnatal elevation in the phospholipid
methyltransferase level may account for a rapid increase in the
proportion of polyunsaturated speci-es of PtdQro in postnatal rat liver
as observed by Ogino et al. (fggO)r âs polpnsaturated species of
diglyceride are preferentially utilized for PtdEtn synthesis (Holub,
1978; Morj-moto and Kanoh, !978). But this does not seem to be the case
in sheep. Noble et aI. (tglt) reported that the proportion of
polyunsaturated species of PtdGro in sheep is reduced during an early

T' -'-
Irtv'o'il'¡
ir il'{5il'íLl1[Ll-ìf-rAtlY
postnatal period. However, they showed that the hepatic content of
ptdcho i-s markedly increased in lambs during the first postnatal week.
Although PtdCho can be synthesized via the CDP-choline pathway, v¡hich is
also enhanced during the postnatal period as observed in rats (Pelech et
aI., 1983), the contribution of the marked elevation of the activity of
phospholipid methyltransferase to the postnatal increase in the hepatic
PtdCLro level should be considered. However, there was little ctrange in
the specific activity of phospholipid methyltransferase from the
pre-ruminant Lo post-ruminant state, despite the change of choline
intake. Ttrese findings suggest that the metabolic balance of choline in
post-ruminant sheep under the poor choline nutrition is achieved by
means of a reduction of the capacity for choline oxidation rather than
the elevation of the capacity for choline synthesis.
The specific activity of guanidinoacetate methyltransferase in
sheep liver \,rIaS not subject. to changes with age: even though milk
generally contains a considerable amount of creatine, as reported for
human and bovine milk (t'{..y and KeIIy , 196I), vf,rile the diet of adult
sheep contains virtually no creatine. Hence, üds obsenration is qsístsrt
with the prevlous finding ttrat this enzyme in sheep liver is not a
rate-lirniting enzyme in creatine synthesis (Henderson et aI., 1983).
One of the important findings in this investigat.ion is the
observation of dramatic changes of the glycine methyltransferase level
in sheep liver during developxnent. The striking developxnental changes of
this enzyme have also been observed in rat livei (Liau et a1., 7919) and
rabbit liver (Ueady and Kerr, L975). The prenatal increase of the
hepatic glycine methyltransferase level in rats and rabbits during
developnent appears to be similar to that observed in sheep. However,
unlike rats and rabbits, vilrere the hepatic enzyme level continuously
1

toz
i-ncreases in postnatal age, the enzyme level in sheep was the highest in
the neonatal liver and thereafter decreased dramat.ically with age. A
d.ecrease of glycine methyltransferase activity would reduce the animalts
requirement. for labile methyl grouPs. Therefore, this dramatic
reduction of glycine methyllransferase activity in adult sheep could be
considered as one of the primary mechanisms for adaptat.ion to the low
intake of methyl nutrients. It is worth noLice thal the decrease of the
glycine methyltransferase level with postnatal age in sheep Iiver is
parallel to the postnatal change of V-cystathionase as reported by
Radcliffe and Egan (7914). The decreased activity of hepatic glycine
methyltransferase could provide another explanation for the reduced
Ievel of hepat.ic Y=cystathionase, as lowering the rate of glycine
methyltransferase reaction would produce a Iesser amount of homocysteine
from AdoMet. Likewise, the develoEnental increase of glycine
methyltransferase activity in rat liver (tiau et al., 1979) may be one
of the factors Ieading to the increase of the enzymatic capacity of
transsulfuration reactions with age in rats (Finkelstein, L967i
Heinonen, L913).
Developrnemtal changes of the levels of betaine-homocysteine,
glycine, guanidinoacetate methyltransferases in sheep pancreas are not
parallel to those of the hepatic enzymes. The specific activities of
these pancreatic enzymes increased dramatically with age until the adult
leyels were reached. Yet significance of the developrnental increases of
the levels of these enzymes in sheep pancreas'andvùrether they are a
unique feature of ruminant animals are unlctown.
Based on Lhe results of developxnental chnnges in the activities of
the enzymes involved in transmethylation reactions and choline
catabolism, it is suggested that adaptation of post-ruminant sheep to

103
the low availability of dietary preformed methyl nutrients is achieved
by a nnrkedly reduced rate of methyl catabolism and an elevated rate of
the de novo synthesis of methyl groups from one-carbon sources. However,
to gain a better understanding on the meclnnisms of such nutritional
adaptation in ruminant sheep, further studies on regulatory and
quantÍ-tative aspects of methyl group metabolism are desirable, vùrich are
presented in the following chapters.

L04
CHAPTER 6. REGUTATION OF ME'THYL GROUP ME'IABOLISM IN IACIATING EI{ES
6.1 Introduction
I-actation results in a nu¡nber of meLabolic changes, viLrich are
directed towards producing substrates for the production of milk
(Williamson, 1980). It has been reported, in some manrnals, that milk
contains a considerable amount of methyl cornpounds, such as choline,
creatine and carnitine (lbcy and Kelly, t967; Zeisel, 1981; Snoswell and
Linzell, 1975), vilrich are irnportant nutrients to neonates.
In most nra¡imals, the extra requirement for the production of these
methyl nutrients in milk may be met sirçly by a liigher dietary intake in
the Iactating state. However, sheep, vùrich are ruminant,s, have a very
srnall amounl of these methyl nutrients available from the diet, because
dietary choline is degraded by rumen microorganisms (lleill et al., L979)
and plant material is krrown to contain virtually no creatine (Walker,
L979) and low carnitine (tuitchell, L978). Therefore, Lhe secretion of
these cornpounds in milk is mainly dependent on their syn thesis in vivo
by transmethylation at the expense of the meLhyl groups from methionine.
However, methionine is lanown to be the first limiting amino acid in
sheep (Ckralupa., L972; Schelling et aI. , L973; Storm and Ørskov, 1984).
It has been shown that the content of methionine in microbial protein is
lower than that of milk protein (Øfoskov, 1982). Davis et al. (fgZg)
found ttnt, in lactating ewes, methionine has the highest extraction
ratio of all amino acids by nranrnary g1and. Thus, the extra dernand on
the utilization of methionine for transmethylation reactions, as weII as
for the synthesis of milk protein, would require the further adaptation
in methyl group metabolism for sheep to achieve methyl group balance

105
during lactation. Yet the mechanisms vilrich operate in the lactating
state are virtually unlcrown.
The lactating state was thus considered as an ideal
experimental model for elucidation of some regulatory mechanisms of
methyl group metabolism in sheep under a condition v¡hen there is a high
dernand for methyl groups. In this investigation, creatine,
creatinine, free choline and lipid choline in e\,ùer s milk \¡/ere
quantitated, as there are no data on the contenLs of these methyl
compounds ín ewe's milk available in literature. The activities of a
nurnber of enzyrnes involved in transmethylation and transsulfuration
reactions LTere also measured in the tissues of lactating and
non-lactating ewes.
6.2. Experimental Procedures
6.2.L Animals
Merino ewes at
lactating er4les with
the age of 3-5 years were maintained on pasture.
twin la¡nbs were slaughtered at 10 days after
parturition.
postpartum.
Milk hTas obtained from lactating ewes' at L5-25 days
6.2.2 Determination of MiIk Ctroline, Creatine and Creat.inine
The concentrations of free choline, lipid choline, creatine and
creatinine in milk were determined as described in Oeapter 3.

106
6.2.3 Enzyme Preparations and Assays
5-MethylH4F-homocysteine methyltransferase, betaine-homocysteine
methyltransferase, guanidinoacetate methyltransferase, glycine
methyltransferase and phospholipid methyltransferase and cystathionine
8-synthase uTere prepared and assayed as described in Ctrapter 3.
6.3. Results
6.3.1 The Content.s of Choline, Creatine and Creatinine in Ewe's Milk
The concentrations of lipid choline, free choline, creatine and
creatinine in ewets milk are presented in Table 6.1. Qroline in ewe's
milk was present mainly in lipid form. Total choline content of ev/e's
milk is similar to the reported values for that of hunan and bovine milk
(Jenness, t974; Þlacy and Kelly, 1961).
A considerable amount of creatine vJas also found to exist
in ewet s milk, v¡frile the content of creatinine, v¡trich has no
nutritive value, was relatively low. Similar values for
the contents of creatine and creatinine in bovine colostrum
were also observed ( 49t t 83 dnd 118 t 5 nmol/m1 for creatine and
creatinine, respectively, n = 10 ). The total contenL of creatine
plus creatinine of ewe's milk is similar to the reported value
for that of hunnn milk (Macy and Kelly, t96I), but the ratio of the
content of creaLine to creatinine in e'nlets milk appears to be higher
than that of human milk.

L07
Table 6.1. The eontents of choline, creatine and creatinine
in ev¡e's milk
C,ompound runol/ml
The values are means t sEM for the milk samples from 4
lactating ewes at L5-25 days postpartum'

108
6.3 2 Effects of Lactation on the Specific Activities of
Methyltransferases and Cystathionine g-Synthase
As shown in Table 6.2, there was a significant increase in the
activity of phospholipid methyltransferase in the liver of e\des in
the lactating state. However, the activiLy of guanidinoacetate
methyltransferase in the liver and pancreas of lactating ewes l^/as not
significantly changed, although there is an extra need for the
production of creatine and creatinine in the milk. Table 6.2 also shows
that the act.ivity of glycine methyltransferase h/as markedly decreased in
the liver and pancreas of lactating ewes. It was found that the
act.ivity of hepatic 5-methylH4F-homocysteine methyltransferase \'ras
increased by 35 7" in the lactating state, v¡hereas there \^lere no
significant changes in the activities of hepatic betaine-homocysteine
methyltransferase and cysLathionine ß-synthase (Table 6.3).
6.4 Discussion
Like other manrnals, evre's milk contained considerabl.e amounts of
methyl compounds. The values for total choline and the sum of creatine
plus creatinine in the milk were 1160 and 732 nmoles per ml,
respectively. Snoswell and Linzell (1975) reported that the carnitine
content of ewe's milk ts 872 t 67 nmoles per mI. Thus, the total value
of these methyl conrpounds was about 6800 nano-equivalenLs of methyl
groups per ml of milk, as choline and carnitine contain three methyl
groups each molecule. If the daily milk yield is 1.5 titre (Snoswell and
Linzell, Lg75), a ev/e will require about 10 milli-equivalents of methyl
groups for the secretion of these methyl cornpounds in milk per day- This

109
Table 6.2. The specific. activities of AdoMet-dependent methyltransferases
in the tissues of lactating and non-Iactating ewes
Phospholipid
methyltransferase
Guanidinoacetate
methyltransferase
Glycine
methyltransferase
Liver
Non-lactating
l-actating
Pancreas
Non-lactating
I-actating
146 ! L2 (4)
1es t 7 (4)
(p < 0.0s)
334 I e (3)
362 ! 32 (3)
(n. s .'t)
1678 ¡ 137 (3)
t47o ! 18e (3)
(n. s .'t)
25.L ! 1.S (4)
10.4 r 2.2 (4)
(p < 0.01)
648 r s8 (4)
44s ! 24 (4)
(p < 0.05)
The enzlne activities are expressed as pmol/min/nrg protein. Ttre values
are means + SÞ{ for the nurnber of animals shov¡n in parenthesis.
Differences between two groups \.vere analyzed by Studentrs L-test.
:k Not significant.

110
Table 6.3. The specific activities of homocysteine-utilizing enzymes in
the liver of lactating and non-l-actating ewes
5-Me thyIH4F-homocYs teine
methyltransferase
(fxnol/min/mg protein)
Betaine-homocysteine Cystathionine
methyltransferase g -slmthase
(prnol/min/mg protein) (nmol/min/mg
protein)
Non-lactating
Lactating
203 ! L2
274 ! t4
(p < 0.01)
13017
L25!8(n. s .:t)
15.5 t 1.0
19.2 ! t.l(n. s .''t)
The values are means t SEM of four animals. Differences between two groups
were analyzed by SLudentrs t-test.
:k Not significant.

LLT
amount at least. equals the daily output of methionine from the gut in
sheep in the non-lactating state (Wolff et aI. , L972; Lindsay, L982).
Subsequent work on the enzymatic capacities revealed that the
activity of phospholipid methyltransferase, v¡hich is responsible for
choline synLhesis, vras increased by 34 7" in the liver of lactating ewes.
Thus, an increase in choline slmthesis would compensate for the loss of
choline in milk production without depletion cf t-e body choline pool.
However, the activity of guanidinoacetate methyltransferase remained
unchanged, vilrich is not. in accordance with the increased production of
creatine for tte secretion of this compound in milk. The explanation for
this rnay be ttnt this enzyme does not appear to be rate-Iimiting in
creatine symthesis in sheep as observed prewiously (Henderson et aL.,
1983), although the elevated urinary excretion of creatinine accornpanied
with an increased activity of hepatic guanidinoacetate methyltransferase
was observed in the legume-fed rats (Santidrian et aI., 1981-).
A marked decrease in the activity of glycine methyltransferase
in the liver and pancreas of lactating ev¡es is of physiological
importance, since the formaLion of sarcosine by this enzymatic reaction
expends methyl groups from AdoMet and could be considered as a wasteful
pathway of methyl group metabolism, for no physiological function of
sarcosine is lcrown. Therefore, slowing down the rate of sarcosine
formation could spare the methyl group of AdoMet for physiologically
essential methylalion reactions, vftrich are erihanced in lactating ewes.
Another possible mechanism to maintain the labile methyl pool in
lactating e\¡/es may be the regulation of the tissue methionine level by
alteration of the relative raLes of homocysteine methylation and
transsulfuration. The observations on three homocysteine-utilizing
enz)nìes suggest that 5-methylH4F-homocysteine methyltransferase

1L2
appears to play this role. The elevation of the activity of
5-methylH4F-homocysteine methyltransferase in lactating ewes provides a
means to replenish methyl groups used for the synthesis of milk choline,
creatine and carnitine.
Therefore, it is sUggested that, in lactating ewes, an increase in
methionine synthesis via the methylation of homocysteine frorn
5-methylH4F and a decrease in Èhe rate of glycine meÈhylation are
essential mechanisms to ensure that the extra de¡rnnd for labile methyl
groups for the produetion of the methyl compounds in milk is met.
i
I
j
¡
I
L
I
I
I
t
I
I
,

11_3
CHAPTER 7: DISTURBANCE OF METIIYL GROUP ME.TABOLISM
IN ALI,OXAN-DIABE'TIC SHEEP
T.L Introduction
Insulin deficiency in uncontrolled diabetes results in net
total-body protein loss with manifestation of negat.ive nitrogen balance
(Naklrooda et al., 1980; Huszar et al. , 1982). The experiments of Forker
et al. (1951) demonstrated that diabetic dogs incorporated only
one-third to one-fifth as nmch 35s-methionine into muscle protein as
normal dogs. Mårlensson and Hermansson (fgA+) reported the increased
urinary excretion of cyst.(e)ine, taurine and total sulfur in ketotic
diabetes, indicating the enhanced rate of methionine catabolism. The
marked increase of the activity of hepatic methionine
adenosyltransferase in rats after alloxan administration (Finkelstein,
Lg67) is in line with the findings of lårtensson and Hermansson
(1984). Henderson et al. (1983) have shown that a dramatic increase of
carnitine synthesis with the decreased production and excretion of
creatine occurs in diabetic sheep. A reduced level of hepaLic
PtdClro has been shown in diabetic rats (Johnson and Cornatzer, L969;
Turakulov et al., L979). The act.ivity of phospholipid methyltransferase
is markedly decreased in alloxan-diabetic rat liver (Hoffman et al.,
1981) and diabetic rat myocardium (Ganguly et aI., 1984). This might
also occur in alloxan-diabetic sheep, but no such study has been
reported in this species to date.
As the amount of methyl groups required for carnitine synthesis is
relatively small, compared with that for the synthesis of choLine and

t1.4
creatine, the total ut.ilization of methyl groups, as indicated by these
three biosynthet.ic reactj-ons, would be decreased in diabetic sheep.
Thus, an increased rate of methionine catabolism with the decreased rate
of total methyl group utilizat.ion suggests that there must be some
alterations in other transmethylation reactions. The reactions
caLaLyzed by homocysteine methyltransferases and glycine
methyltransferase are likely to be candidaLes, as it has been
previously shown that these reactions could exert the control of
methyl group metabolism in sheep ( Chapter 4-6 ). It. \^/as thus
considered that the diabetic state could offer a good experimental
model for the elucidation of regulatory mechani-sms of methyl group
metabolism j-n sheep under a condition v¡hen methyl groups appear to be in
excess.
7 .2. Experimental Procedures
7.2.1 Animals
Approximately 3-year-old Merino wethers were divided into two
groups, maintained in metabolic crates and fed on lucerne-hay chaff
ad libitum. Diabetic animals \^Iere induced by inject.ing alloxan
(60 mg/kg body wt.) intravenously. The animals \^7ere slaughtered
approximately one week after alloxan treaLmenL, vltren the blood glucose
level r^ras more than 4-fold higher than the normal value (assayed
by R. C. Fishlock) and the level of urinary ketones r^/as bretween
moderate and high, as determined by Ketostix strips (Rmes Company,
Melbourne, Australia). 24-hr-urine samples r,ùere collected with toluene
as preservative.

115
7.2.2 Enzyme Preparations and Assays
The preparations and assays of betaine-homoeysteine
methyltransferase, 5-methylFI4F-homocysLeine methyltransferase, glycine
methyltransferase, guanidinoacetate methyltransferase, phospholipid
methyltransferase, cystathionine ß-synthase and v-cystathionase were
described in Chapter 3.
1.2.3 Determination of Cysteine in Urine
Urinary cysteine \^ras measured as described in Chapter 3.
7.3 Results
7.3.I Effects of Alloxan-diabetes on the Specific Activities
of Homocyst.eine Methyltransferases
Table 7.1 shows that the activity of hepat.ic betaine-homocysteine
methyltransferase was significantly reduced in severely diabetic sheep.
while there was slight., but not. significant, decrease in the activity of
hepatic 5-methylH4F-homocysteine methyltransferase. Thus Lhe rate of
hepatic methionine synthesis may be reduced in diabetic sheep.
7.3.2 Effects of Alloxan-diabetes on the Specific Activities of
AdoMe t -dependen t Methyl trans f eras es
As shown rn TabLe 7.2,, the activity of pancreatic guanidinoacetate
methyltransferase \,vas found to decrease 3-fold in diabetic sheep,

L16
Table 7.1. Iþe specific activities of homocysteine methyltransferases
in normal and alloxan-diabetic sheep liver
5 -Me thylH4F-homocY s teine
methyltransferase
Betaine-homocysteine
meLhyltransferase
Nornal 2L8 ! t8 98.2 ! 6.0
Diabetic 52.0 ! 7.5
(p < 0.01)
Each value represents rnean t SEI',I of 5-7 animals and is expressed as
¡xno/min/ mg protein. Differences between normal and diabet.ic animals
were analyzed by Studentrs L-test.
* Not significant.
176 t 24
(n.s.io)

It7
Table 7.2. Tlne specific activities of AdoMet-dependent methyltransferases
in normal and alloxan-diabetic sheep tissues
Glycine
methyltransferase
Guanidinoacetate
methyltransferase
Phospholipid
methyltransferase
Liver
NornnI
Diabetic
Pancreas
Normal
Diabetic
29.O ! 3.4
1872 ! 493
(p < 0.001)
619 r 85
729 ! 95
(n.s.*)
363 ! 26
308 I 34
(n.s.*)
1388 t 79
442 ! 98
(p < 0.01)
1481 6
81 !L3
(p < 0.01)
Each value represents mean I SEM of 3-7 aninnls and is expressed as
pnol/min/mg protein. Differences between normal and diabetic animals
were analyzed by Studentrs t-test.
:k Not signif icant.

11-8
v¡hereas the hepatic enzyme was noL significantly affected. The act.ivity
of phospholipid methyltransferase in severely diabetic sheep liver was
also significantly deereased. It can also be seen that a 65-fold
increase in the activity of hepatic glycine methyltransferase with
unchanged pancreat.ic enzyme occurred in severely diabetic sheep. These
results indicate that the three AdoMet-dependent methyltransferases are
all affected in diabetic sheep.
7.3.3 Effects of Alloxan-diabetes on the Specific ActiviLies of
Cystathionine ß-Synthase and Y-Cystathionase
Cystathionine ß-synthase and v-cystathionase are involved in the
homocysteine catabolic pathway. Table 7.3 shows that the activity of
y -cystaLhionase in diabetic sheep liver increased 4-fold, v¡hile no
significant change in the activity of cystathionine ß-synthase \das
observed.
7.3.4 Urinary Excret.ion of Cyst(e)ine before and after Alloxan Treatment
The values of cyst(e)ine (cysteine plus cystine) excretion in sheep
urine, expressed as ¡rmole /day per kg body wt., were 0.58 t 0.05 before
alloxan inject,ion and 0.96 t 0.05 (mean t SW) at the 6th and 8th day
after alloxan inject.ion. Thus, urinary excretion of cyst(e)ine j,n
diabet,ic sheep was increased by 66 7".
7.4 Discussion
The present investigation demonstrated a 3-fold decrease in the

L19
Table 7.3. The specific activities of cystathionine ß-synthase and
y-cystathionase in normal and alloxan-diabetic sheep liver
Cyslathionine ß-synthase r-Cystathionase
i
i
I
I
i
I
i
Nornral 17.5 t 1.5
16.3 I 0.7
(n. s .:"t)
0.44 r 0.04
1.65 I 0.11
(p < 0.01)
Diabetic
Each value represents mean t SEM of 3-6 animals and is expressed as
nnlc:.1-/ni:n/mg protein. Differences between normal and diabetic animals
were analyzedby Studentrs t-test.
:'i Not significant.

L20
activity of guanidinoacetate methyltransferase in diabetic sheep
pancreas, while the enzyme in the liver remained unchanged as reported
previously (Henderson et aI., 1983). However, it is difficult to assume
that a 3-fold decrease in the pancreatic enzyme activity could explain a
significant reduction in total-body creatine synthesis (Henderson et
al., 1-933): âs the tissueweight of pancreas is much less than that of
liver although the higher enzyme activity exists in the pancreas.
The reduced activity of phospholipid methyltransferase in diabetic
sheep liver is in agreement with the observation of Hoffman et al.
(fggf) and Cabrero et al. (fgge) in rats. The possible explanation is
that. the reduced act.ivity of the enzyme is a result of insulin
deficiency, as this reduction of the enzyme activity can be restored by
injection of insulin (Ganguly et al., 1984) and insulin has been shown
to stimulate the enzyme activity in vitro (fetty et aI., L984; Kiechle
et al., 1986). Decrease in the activiLy of phospholipid
methyltransferase indicates that. the rate of PtdCtro synthesis via the
methylation pathway is reduced in severely diabetic sheep liver. Thus,
the reduced activity of betaine-homocysteine methyltransferase in
diabetic sheep could be attributed to the decreased oxidation of
choline, owing to the reduction of choline synthesis in the diabetic
sheep, as the origin of choline in this species is mainly from in vivo
synthesis by the successive methylation of PtdEtn (Dawson et al., 1981;
NeiII eL al. , L979).
The most important finding from this investigation is a dramat.ic
increase in the activity of hepatic glycine methyltransferase in
severely diabetic sheep. As it. is known that most of AdoMet. formed in
mammalian tissues under normal condilions is ut.ili-zed for creatine and
choline slmLhesis (tUuaA and Poole, L975; Dawson et al., 1981), decrease

L2L
in the synthesis of these methyl group compounds in diabetic sheep would
markedly reduce the utilizati-on of AdoMet. Furthermore, it is known
that, in ketotic diabetes, tissue protein degradation is markedly
increased and amino acid utiltzaLion for protein synthesis is markedly
reduced (Huszar et. al., 1982; Jefferson eL aI., 1983; Felig et al.,
t977). Elevated levels of blood amino-N and urinary amino acid
excretion also occur in diabetes (Ivy et al., 1951). The increased
urinary excretion of cyst(e)ine in severely diabetic sheep observed here
agrees with the finding of Mårtensson and Hermansson (1984). They
explained the elevated urinary excretion of cyst(e)ine, taurine and
total sulfur in ketot.ic diabetes as a resull of the surplus of
sulfur-containing amino acids released from t.issue prot.ein mobilization.
An increase in the activity of hepatic methionine adenosyltransferase in
alloxan-treated rats (Finkelstein, L967; Cabrero et al., 1936) could
ensure the enhanced rate of methionine catabolism via the
Lransmethylation-transsulfurat.ion rouLe. Cabrero et al. (1986) recently
showed a marked increase in the levels of AdoMet and AdoHcy in
alloxan-diabetic raL hepatocytes. Their findings confirm the enhanced
rate of AdoMet synthesis and also imply the increased rate of the total
AdoMeÈ utilization in the diabetic animals. Therefore, it. is reasonable
to assume that. the dramat.ic increase in the activity of hepatic glycine
methyltransferase is a means of cataboLizing any surplus AdoMet in
severely diabet.ic sheep, owing to the reduced ut.ilizat.ion of AdoMet for
the synthesis of creatine and choline and the enhanced production of
AdoMet.
The observation of a 4-fold increase in the activity of
t-cystathionase in diabetic sheep liver provides a further explanation
for the elevated excretion of cyst(e)ine in urine. Although no change

t22
in the activity of cystathionine ß-synthase r^7as observed, there is some
evidence indicating that Y-cystathionase is more likely to be a key
enzyme in the homocysteine catabolic pathway in sheep. Radcliffe and
Egan (L974) observed that Y-cystathionase is subject to postnatal
changes in sheep, while cystathionine ß-synthase was not (Chapter 5).
Therefore, the rise in the Y-cystathionase activity reflects the
elevated caLabolic rate of homocysLeine, owing to increased homocysteine
production from methionine by means of the elevated activity of glycine
methyltransferase in diabeLic sheep.
Based on the results of the effect of diabetes on the activities of
the enzymes involved in transmethylation reactions and homocysteine
transsulfuration reactions, it appears that the diabetes-induced
alterations in methyl group metabolism in sheep are characterized by an
accelerated rate of methionine catabolism via the
transmethylation-transsulfuraLion route and a reduced rate of choline
and crealine synthesis. Most. importantly, the results highlight the
important. role of glycine methyltransferase in Lhe disposal of labile
methyl groups in sheep.

123
CHAPTm, 8. PERTURBATION OF METTIIONINE ME'IABOLISM IN SFIEEP I^rITFl
NITROUS-OXIDE-INDUCED INASTIVATION OF COBAUMIN
8.1 Introduction
Oxidation of cob(I)alamin to cob(Ill)alamin by the anaesthetic gas,
nitrous oxide (NZO), io humans and some species of experimental animals
leads to neurological changes and metabolic alterations v¡hich resemble a
state of vitamin 812 deficiency (Chanarin, 1980; Scott et aI.' 1981; van
der l^lesthuyzen eL a1., L982; Eells et al ., 1982). The primary mechanism
of NZO toxicity has been suggested to be due to the inhibition
of the cob(I)alamin-dependent enzymer 5-methyltl¿*F-homocysteine
methyltransferase (Chanarin, 1980), which is one of the two main
homocysteine methyltransferases responsible for methionine synthesis in
mannnals. It is further proposed that neurological pathogenesis in
N2O-exposed animals may be a result of methyJ- group deficiency due to
the irnpairment of methionine synthesis by 5-methylQ*F-homocysteine
methyltransferase (Scott et al., 1981). However, there is no general
agreement on the effects of N2O exposure on tissue methionine and AdoMet
levels in recenL studies in vùrich rats, mice and frrit bats were used as
experimental anirnals (Ckranarinr1981; llrnb et al., 1983; Bukin and Orlov,
L984; van der Westhuyzen and Metz, 1983). Thus, the mechanism of the
N2O-induced neuropa.thy still remains the subject of controversy.
In the present study, sheep were used as a suitable animal model to
investigate the effects of N2O-induced inactivation of cobalamin on
methionine metabolism, because the previous results (CIrapter 4) suggest
that. sheep, unlike most of other species, are heavily dependent on
5-methylH4F-homocysteine methyltransferase for methionine synthesis-

124
This is because of the unavailability of dietary choline, vùrich is a
methyl precursor for an alternative pathway of methionine synthesis by
betaine-homocysteine methyltranferase, due to the breakdown of dietary
choline in the rumen (neitt et aI ., 1979). The purPose of this study is
mainly focused on elucidation of Lhe importance of
5-methylH4F-homocysteine methyltransferase in ttÊ supply of labile methyl
groups in sheep, by using N2O to block the synthesis of the methyl group
of methionine from 5-methylH4F.
8.2 Experimental Procedures
8.2.I Aninrals
Merino wethers at tle age of 2-3 years were divided into two groups and
kept in metabolic crates individually and fed lucerne chaff ad libitum.
One group of sheep with plastic face masks v/ere connected to a gas
supply through tubing and exposed to a mixture of N2o/O2 (4;L, v/v) at
a rate of 7.5 litres/min for 2 hr per day for 8 days. The NZO
concentraLion in the mask was 36% on average, aS measured by mass
spectrometry (determined by Dr. A. M. Snoswell). In the control group,
the animals were exposed to room air.
8.2.2 Preparation of Tissues
Blood was obtained between l-2.00 and 13.00 hours frun tejugular rein
of sheep before and after N2O exposure for 8 days. Plasnn itlas
deproteinized by the addition of L/j voL of. L2 7. sulphosalicylic acid
within 30 min after sampling and stored at -80oC until assay. 24-hr

L25
urine sanples htere collected with HCl as preservative from sheep before
and after Nzo exposure for 7 days and stored at -15 o c untir assay.
s'heep hTere slaughtered at the end of N20 exposure on the 8th day.
Tissues \,rrere rsnoved as rapidly as possible, inrnediately dropped into
Iiquid nitrogen and stored at -80oC until assay.
8.2.3 Enzyme Preparations and Assays'
5-MethylH4F-homocysteine methyltransferase, betaine-homocysteine
methyltransferase and methylmalonyl-C.oA mutase \^/ere prepared and assayed
as described in Ckrapter 3, except that for assay of
5-methy1H4F-homocysteine methyltransferase the reduced glutathione vrras
omitted in tlehomogenizing buffer ad the supernatant obtained from 50,000
x g centrifugation was used without dialysis.
8.2.4 Metabolite Assays
The concentrations of AdoMet and formiminoglutamic acid were
measured as described in Grapter 3. Plasma methionine and urinary
homocystine concentrations were determined on a Technicon amino acid
anaLyzer ( by },lr. D. Boehm ).
8.3. Results
8.3.1 Effects of N2O on the Specific Activities of Vitamin 812-requiring
Eezyrnes and Betaine-homocysteine Methyltransferase
Table 8.1 shows that the activity of 5-methyrH4F-homocysteine

L26
Table 8.1. The effect of N2O on the specific activities of
vitamin Bl2-requiring enzymes in sheep tissues
Exposure Liver
5-Me thylH4F-homocys teine
methyltransferase
(prnol/min/mg protein)
Methylmalonyl-CoA
holo-nmtase
(nmol/min/mg protien)
LiverBrain Heart
Air t98 ! Lt+ 105 t L0 59 I 10 4.55 ! 0.27
3.76 ! 0.26
(n.s.''t)
Nzo 19r3(p < 0.001)
28!2 1014
(p < 0.01) (p < 0.01)
Sheep \^rere slaughtered after exposure to N2O fot 2 hr per day for 8
days. Each value represents themean I SEM of 4 animals. Differences
between animals breathing air and those exposed to N20 were analyzed
by Studentrs t-test.:k Not signif icant.

L27
methyltransferase in sheep tissues after intermittent exposure to NtO
for 8 days was reduced to IO7", 18% and 267" of the control values in the
Iiver, heart, and brain, respectively. In contrast, Lhe activity of the
holo-enzyme of methylmalonyl-CoA mutase, cre of Ûe vitamin Bl2-requiring
enzyrnes, \¡/as not. subject to a significant decrea"" (p < 0.1) after 8-day
exposure to N2O (taUle 8.1). The activity of the total mltase \47as
also measured by the addition of coenzyme B12 into the assay medium.
The total muLase activity was about 4 times higher than that of the
holo-nrutase and no significant difference of the enzyme act.ivity was
observed'beb¡em tle b,io groups ( 16.8 t 0.5 and 13.4 t 0.8 nmol/min/mg
protein in the control and N2O-exposed animalsr respectively). These
results demonsLrate that N2O exposure causes the strong irihibition
of 5-methylH4F-homocysteine methyltransferase but not methylnnlonyl-CoA
mutase in the period of N2O exposure studied.
In order to elucidate vilrether the inhibit.ion of
5-methylH4F-homocysteine methyltransf erase led to the stirrn-llation
of an alternative pathway for methionine slmthesis, the activity of
betaine-homocysteine melhyltransferase I^Ias also measured. It was found
that there was no significant change in this enzyme activity in sheep
liver after N2O exposure (tZq x 9 pxnol/min/mg protein in the control
group vs. 109 t 16 in N2O-exposed sheep)-
8.3.2 Effects of N2O on Adollet and Methionine Concenlrations
AdoMel level in the liver of sheep fell rnarkedly after 8 days of
N2O exposure (Table 8.2) and there was a slight but significant decrease
in its concentration in the heart. By contrast AdoMet level in the
brain was not changed. The concentrations of plasma methionine in sheep
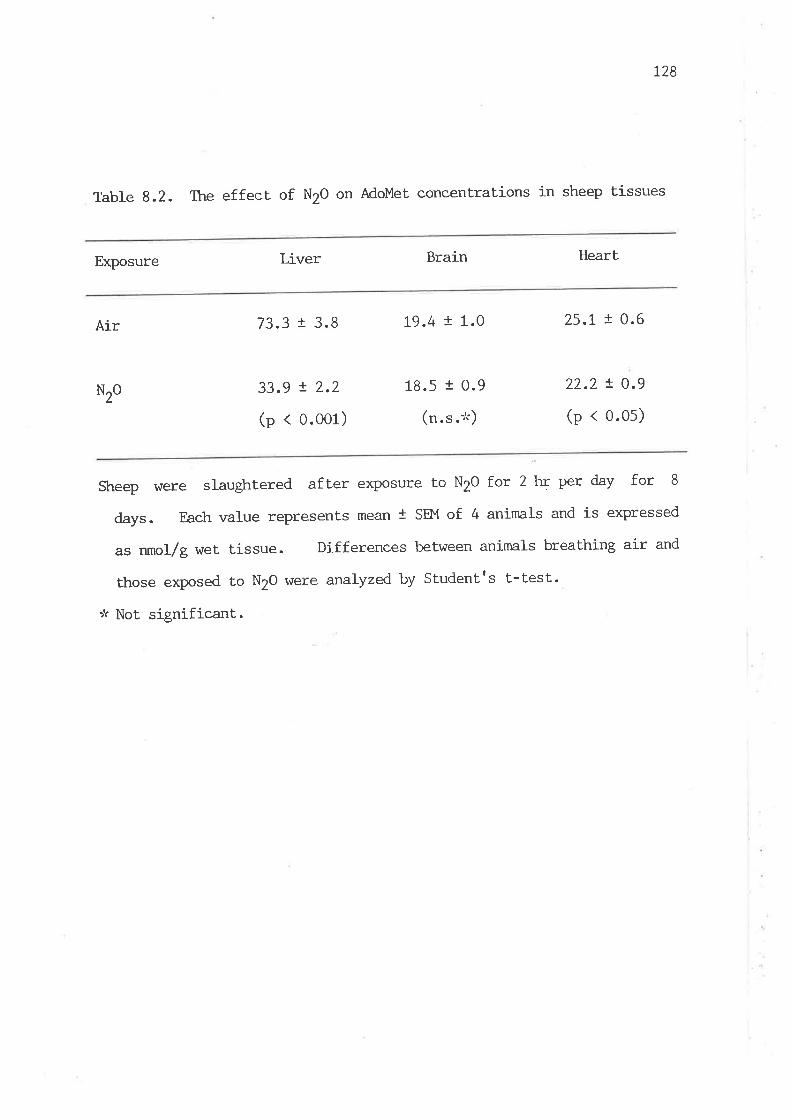
L28
Table 8.2. The effect of N2O on AdoMet concentrations in sheep tissues
Exposure LÍver Brain Heart
Air 73.3 t 3.8
33.9 ! 2.2
(p < 0.001)
19.4 t 1.0
18.5 1 0.9
(n. s .'t)
25.1 I 0.6
22.2 ! 0.9
(p < 0.0s)
N20
sheep were slaughtered after exPosure to N2O for 2 krr per day for 8
days. Each value represents mean t SEM of 4 anirnals and is expressed
as nmol/g wet tissue. Differences between animals breathing air and
those exposed to N2O were analyzed by Studentrs t-test.
-,'r Not significant.

L29
were found to @ L2.4 t 3.4 and 3.6 I 2.7 ¡Ìa before and after 8-day
exposure to N2O, respectively. Thus, the value in the N2O-exposed
animals was reduced to 30 % of- its initial level.
8.3.3 Effects of N2O on Urinary Excretion of Formiminoglutamic
Acid and HomocYstine
Data in Table 8.3 show that exposure to N2O for 7 days caused the
excretion of formiminogluLamic acid. Formiminoglutamic acid \'{as
virtually undetectable in the urine of sheep brefore N20 exposure and
became detectable in a considerable amount on the second day after
N2O exposure (o.z t 0.3 nnnol/ 24 hr). Homocystine \^7as slightly
detectable in the urine of two of the four sheep before N2O exposure.
Tkre elevated urinary excretion of homocystine occurred in all sheep
after 7 days of N2o exposure (table 8.3).
8.4 Discussion
Methionine synthesis via 5-methylH4F-homocysteine methyltransferase
reaction is highly active in sheep (Chapter lr), owing to the poor supply
of preformed methyl nutri-ents from the gut. However, in most other
species an alternaLive pathway of melhionine synthesis by homocysteine
methylation from betaine, vilrich is derived from choline, can produce a
substantial amount of methionine. As observed in rats, this reaction is
able to sustain normal growth vùren the animals fed a methionine-free
diet supplemented with homocysteine and choline or betaine (du Vigneaud,
fg52). Therefore, it is not surprising that N2O-exposed rats are able
to maintain normal methionine levels of the tissues (Chanarin, L98t;

130
Table 8.3. Urinary excretion of formiminoglutamic acid and homocystine
in sheep before and after N2O exposure
' Exposure of N2O
Urinary formiminoglutamic Urinary homocystine
acid
(mrnor/24 hr) ( pmol/24 hr)
Before n.d.t'. 0.2 r 0.1
After 1.3 r 0.3 5.1 I 1.9
Urine samples were collecLed from sheep before and after exposure to N20
f.or 2 hr per day for 7 days. Ttre values are mean t SEM of 4 animals.
:k Not detectable.

73L
hmb et al., 1933), âs long as a certain amount of methionine and
adequate choline are present in the diet. Hence, to observe the effects
of N2O on Lissue methionine and AdoMet levels in raLs, the amount of
dietary methionine and choline has to be carefully controlled.
As was expected, a marked fall in plasma methionine and hepatic
AdoMet levels \47as readily demonstrable in sheep af ter tissue
5-methylH*F-homocysteine methyltransferase had been severely inactivated
by N2O. Studies on changes of plasma methionine level in humans exposed
to NrO during surgical operation are inconclusive (Skacel eL al., 1983;
Nunn et aI., 1986). Van der l,{esthuyzen eL af.(19S5) reported that tissue
methionine levels are decreased in fruit bats after prolonged continuous
exposure to N2O for L7 weeks. However, they found no effects of N20 on
tissue AdoMet levels of fruil bats after the long-term exPosure to N20
(van der l,,Testhuyzen and Metz, 1983; Gibson and van der I'Jesthuyzen,
1934). Bukin and Orlov (1934) also found no change of the hepalic
AdoMet level in N2O-exPosed mice.
Although plasma methionine and hepatic AdoMet levels in N2O-exposed
sheep \,vere reduced to one-third and half of the control values,
respect.ively, the concentrati-on of AdoMet showed only a slight
decrease in the heart and no change in the brain. There is no
clear explanation for these observations, since there is no knowledge of
any alternative pathways for methionine synthesis present in these
tissues so far. However, unlike liver, the I(m value of methionine
adenosyltransferase for methionine in non-hepat.ic tissues was reported
to be well below the physiological levels of this amino acid in rats and
humans (Hoffman and Kunz, 1980; Okada et al., 1981-; Carl et al., L978;
Oden and Clarke, 1983; Liau et a1., t979). Furthennore, the rate of
AdoMet utilization for biological methylation in brain and heart must be

t32
nuch less than that in liver, since the activities of phospholipid
methyltransferase and guanidinoacetate methyltransferase, vf,rich are
lcnov¡n to expend most of Adol'{et (Mudd and Poole, L975; Dawson et al',
1981) , are very low in these Lissues of nanrnals (Bre¡ner and Greeberg'
L96L; van Pilsum et al., 1972; Yanokura and Tsukada, L982).
InN2Q-exposedsheeprhomocystinuriawasobserved'bltitdoes
not occur in rats even v¡tren 5-methylH4F-homocysteine methyltransferase
was almost cornpletely inactivated (Deacon et al.r 1980)' The reason for
this can be ascribed to highly active betaine-homocysteine
methyltransferase present in rat liver, in v¡hich the enzyme level is
5 times higher than that in sheep liver (Chapter 4 ). In addition, the
activity of betaine-homocysteine methyltransferase is increased in rats
after NZO exPosure (f.umU et aL', 1-983) ' In contrast' Lhe enzyme
activity remained unchanged in sheep. Therefore, the elevated activity
of betaine-homocysteine methyltransferase in N2o-exposed rats relnoves
the extra amount of homocysteine v¡hich would othen¡ise be removed by
5-rnethylH4F-homocysteine methyltransferase in normal conditions'
However, in the case of sheep, owing to the much lower activity of
betaine-homocysteine methyltransferase and a much lesser amount of
betaine derived from choline (Caapter 4), the enzyme activity is too
Iow to prevent the accumulation of homocysteine in the N2o-exposed
animals and consequently homocystinuria occurs'
ItappearsthâtNzodoesnothaveadirecteffectupon
methylmalonyl-C-oA nmtase in rats (Deacon et al., L978), although the
prolongedexposuretoNzoeventuallyleadstoadecreaseinthe
holo-nmtase activity due to decreases in tissue cobalamin levels (Kondo
et aI., 1981). In contrast, elevated urinary excretion of methylmalonic
acid occurs in nan after 24-hr exposure to N2o (nask et al., 1983)'

133
Methylmalonyl-CoA nn-rLase is particularly irnportant in ruminant. animals
as a key gluconeogenic enzyme in the synthesis of glucose from
propionate (Smitn et aI., 7967). The present results show that the
holo-nn-ltase vùas not signif icantly decreased in sheep af ter 8-day
exposure to N20. It is possible that the holo-nmtase activity in sheep
nny eventually decrease, if N2O exposure continues for a longer period
until tissue cobalamin concentrations decline markedly. The present
finding suggests that the oxidized form of cobalamin is effective for
the mutase activity in sheep. This renders N20 a rnore specific tool to
explore the effects of cobalamin-inactivation on methionine metabolism.
Urinary excretion of formiminoglutamic acid is recognized as
a symptom of the disturbance of one-carbon metabolism in vitamin
Bl2-deficient animals (Silverman and Pitney, 1958; Smith et al., L974)
and in N2O-exposed rats (Deacon et al., 1983). It is clear thatt
ir¡tribition of 5-methylH4F-homocysteine mlhyltransferase by N2O leads to
impaired utilization of formimino residues derived from histidine in
sheep. Presumably, this is due to the reduced levels of tissue
non-methyl folate, as obsen¡ed in rats ( Eells et al ., L982; Flansen and
Billings, 1985; t/ilson and Horne, 1986), although the reduction of
tissue non-methyl folate can not be satisfactorily explained by
methylfolate trap hypothesis (¡ells et al., L982; llmb et al., 1981).
Nevertheless, urinary excretion of formiminoglutamic acid was readily
demonstrable on the second day of NZO exposure, indicating the
occurrence of severe inhibition of 5-methylQrF-homocysteine
meLhyltransferase by N2O exposure.
The results presented herein suggest thnt. sheep are more sensitive
to N20 exposure in respect of perturbation of methionine metabolism than
some other species. These findings further document that the

t34
methylation of homocysteine from 5-methylH4F is of upxnost importance in
methionine synthesis and is an indispensable methyl source for
nnintaining physiological levels of methionine and Adol4et in sheep.

135
CHAPTER 9. QUANIITATIVE EVALUATION AND REGULATION OF
S-ADENOSYLI.4ETHIONINE-DEPENDENI TRANSME'TTM-ATION IN SHEEP TI S SUES
9.t InLroduction
AdoMet, vùrich is derived from methionine, is known to be the
principal methyl donor for various biological methylation reactions,
vùrich are vital in diverse physiological and biochemical processes.
However, there is a relative lack of information on general aspects of
transmethylation such as absolute rates of transmethylation and AdoMet
turnover, toLal requirement of methyl groups and factors regulating
AdoMet levels in most of ma¡nrnalian tissues.
A quantitative estimation of total requirement of AdoMet for
transmethylation has been attempted only in humans by l'fudd and his
colleagues (fgZS, 1980) by measurement of urinary excretion of methyl
compounds vitrich are derived from the methyl group of AdoMet. Their
excellent studies provide the first convincing evidence that. the amount
of methionine used for transmethylation is far in excess of its dietary
intake. Ttrerefore, a significant portion of methionine has to be
reslmthesized by homocysteine methylation, vil.rich is an indispensable
source of body meLhionine in manrnals. A minimal estimate for the
conservative metabolism of methionine in young adult humans on normal
diets is that about 50 7" of homocysteine is converted to methioníne
(Mudd and Poole, L975; Itudd et al., 1980). In particular, methionine is
a first-limiting amino acid in ruminant sheep with a poor supply of
preformed methyl nutrients (Chalupa, 19721 Storm and Ørskov, !984; NeiII
et aI., 1979). The extent of homocysteine methylation in ruminant sheep
might be even greater. Although the data presented in Qeapter 4-5 have

t36
shown that sheep possess conservative features of methyl group
metabolism, it appears that the quantitative evaluaLion of AdoMet
utilization for biological methylation in sheep tissues and the
esLimation of the total-body requirement of methyl groups from AdoMet
are highly desirable to the quantitat.ive understanding of methionine and
AdoMeL metabolism in this species.
As little is known to date about regulatory aspecLs of the
metabolism of AdoMet in sheep tissues, the relationship bet'ween
methionine and AdoMet levels in various tissues of sheep is not fully
understood at present. For example, it \,vas previously observed
(Chapter 8) that. AdoMet level is markedly diminished in the liver of
sheep with N2O-induced methionine deficiency, vhile there is no decrease
of AdoMet level in the brain. Such a difference in response of tissue
AdoMet. Ievel to changes of methionine availability in sheep has remained
unexplained.
In this investigation, the rates of transmethylat.ion and AdoMet
turnover in various Lissues of sheep were quantitaLively evaluat'ed and
a tentative estimation of the total requirement of methyl groups in
this species \^/as determined. The kinetic properties of major
meLhyltransferases and the effects of methionine loading on tissue
AdoMet levels and the rate of transfer of methyl groups from AdoMet were
explored to elucidate the regulatory aspects of AdoMet metabolism'
9.2. Experimental Procedures
9.2.L Animals
Tho to three-year-old Merino e\^/es weighing about 50 kg \dere

r37
mainLained on pasture and 3-month-old female l,/istar rals weighing about
200 g were fed ad libitum a pelleted rat diet (Charlicks, Adelaide,
South Australia).
9.2.2 Measurement of AdoHcy and AdoMet levels in Tissues
AdoHcy and AdoMet concentrations were measured as described in
Chapter 3.
9.2.3 Measurement of Transmethylation RaLe in vivo
The measurement of the rate of transmethylation was based on the
method of Hoffman (1980), which involved blocking hydrolysis of AdoHcy,
a product of transmethylat.ion reactions, with an extremely potent
inLribitor of AdoHcy hydrolase, periodate-oxidized adenosine (pOA). POA
was prepared essentially according to the method of Hoffman (1930).
Adenosine was oxidized by periodate at room temperature for l- hr and the
suspension' was chilled on ice for about 2 hrr. The precipitate of POA
\,{as collected by centrifugat.ion and washed 4 times by suspension in,
and centrifugation from, cold water (7 ml lþO/g adenosine). The washed
POA was kept. on íce and used on the day of preparation.
Sheep and rats were injected with a freshly prepared POA solution
in 0.9 7" (w/v) NaCl intravenously and intraperitoneally, respect,ively.
In the group of sheep in which transmethylation rates virere measured 2 hr
after treaLment r^rith POA, another dose of POA was given l- hr after first
injection. In studies of the effect of methionine loading on AdoMet
level and the rate of methyl transfer from AdoMet., a dose of 85 mg
methionine per kg body weight was simultaneously injected with POA.

138
Animals \^rere rapidly killed at given times after treatment with POA.
Tissues were quickly excised, dropped into liquid N2 and stored at -800C
for determination of tissue AdoHcy and AdoMet levels.
Transmethylation rate was estimated from the initial rate of the
sum of MoHcy plus AdoMet accunmlated, i.e. the incremenLs after
treatment with POA over the normal levels in control animals. The rate
is expressed as nmoles/min/g wet tissue.
9.2.4 Preparations and Assays of Methyltransferases
The methods for the preparation and assay of guanidinoacetate
methyltransferase, glycine methyltransferase and phospholipid
methyltransferase were basically the same as described in Chapter 3.
Some modifications were made for the preparation of partially purified
guanidinoacetate and glycine methyllransferases. The supernatant
obtained after the 105,000 x g centrifugation of the homogenate of
sheep liver was brought to pH 5.0 with 1 N acetic acid.
After centrifugation at 10,000 x g for 1-0 min, the pH 5.0 supernatant
was inrnediately neutralized with 2 N KOH. Solid (ma)2SOa (20 g,/100 mI)
was added slowly to the neutralized supernatant with gentle
stirring. After 30 min, the solution was centrifuged at 101000 x g
for 30 min to rernove precipitated protein. The resulting
supernatant was again treated with (NH4)2SOa O e/t}O mI). After
standing for a further 30 min, the solution was centrifuged as above to
obtain the precipitated protein containing guanidinoacetate and glycine
methyltransferases. The precipitated enzymes \tere dissolved in 50 mM
potassium phosphate hrffer (pH 7.4) containing 1 mM lt.IT and 1 mM EyfA
and dialyzed 3 times against the same buffer for t6-20 hr and used as

139
enz)lne sources. In order to eliminate the effect of pH -m tle kinetic
constants, Tris-HCl buffer at pH 7.5 was used in the assays of the
three methyltransferases to obtain their kinetic parameters in a
condition sjmilar to the physiological environment. Km values \^/ere
determined by double reciprocal plot and Ki values were calculated from
the apparent Km values of the double reciprocal plot in the presence of
the inhibitor.
9.3. Results
9.3.1 Tissue Distribution of AdoHcy and AdoÙIet in Sheep
Table 9.t shows the tissue distrib.ltion of AdoHcy and AdoMet in
sheep. The liver and pancreas contained relatively high levels of
AdoHcy and AdoMet cornpared to other Lissues. The concentrations of
AdoHcy were much lower than those of Adol1et in all the tissues examined.
In the heart, brain, skeletal nmscle and v¡l'role blood, the concentrations
of AdoHcy were too low to be accurately deLermined.
9.3.2 Efficacy of POA to Block AdoHcy Hydrolysis
POA has been shown to be an extremely potent irhibitor of AdoHcy
hydrolase r¿ith the Ki of 2.4 nM, (patel-thornbre and Borchardt, 1985).
Fig. 9.1 demonstrates the efficacy of this inhibitor in preventing the
in vivo hydrolysis of AdoHcy in sheep and rat tissues. The maxinmm
inhibition seems to be reached at 10 Fmof/kg body wt. in sheep tissues
and at 30 ¡Lmof/þ in rat liver. To ensure the cornplete inhibition,
50 ¡mol/ke of POA was chosen for the measurement of the overall rates of

140
Table 9.1. Distribrtion of AdoHcy and AdoMet in sheep Lissues
Tissue AdoHcy
(nmol/g wet tissue)
AdoMet
(nmol/e wet tissue)
Liver
Pancreas
Kidney
Spleen
Heart
I-ung
Brain
Skeletal muscle
l.lhrole blood
Adipose
Skin
L4.2 ! L.3
14.6 ! 3.7
9.2 ! t.3
2.8 ! O.2
<2
3.9 t 0.4
<2
<2,an.o.,an.Cl.
2.6 ! 0.5
66.6 t
36.8 I
28.7 !
24.5 !
23.2 !
23.6 !
19.8 t
L2.6 !
5.7
2.8
' t4.4 !
3.9
3.8
1.4 b
2.2
t.4
0.9
0.4
0.5
c
c
1.0
The values are means t SEM of three animals except vürere
indicated.a Not detectable.
b Determined after further purification by TI-C-
c The mean value of two animals
they are

L4L
o=tnU,
O+tIooìiSoEtr
500
400
300
200
100
,/
o
10010 20 30 40 50
Dosage (^¡mot/kg)
Fig. 9.1. Inhibition of tissue AdoHcy hydrolysis by POA. Sheep and raLs
\,{ere injected \,'rith varying concentrations of POA intravenously and
intraperitoneally, respectively. Each point represents the mean value
of. 2-4 animals killed at 30 min after the injection. o , sheep liver;
l, sheep pancreasi a r sheep kidneyi o r rat liver.
o

L42
transmethylation.
9.3.3 In vivo Rate of Transme thylation in Tissues
As shown in Fig. 9.2, tissue AdoHcy concentrations increased
markedly with time after the injection of POA. However, the rates of
AdoHcy accumulation hlere maximal at first 15 min in sheep liver,
pancreas and kidney after the intravenous injection of POA. In rat
Iiver, the rate was maximalbeU,æsr 15 ¿rd I min after the intraperitoneal
injection. Increases of AdoMet concentraLions with time after the
treatmenL were also obserùed, but to a nmch lesser degree cornpared with
the AdoHcy accunn:lation. Or,ring to a dramatic rise of AdoHcy level with
a nmch lesser increase of Adol'fet level, the ratios of AdoMet to AdoHcy
in these tissues fell markedly within 15 min and continued to decrease
slightly thereafter.
The concentrations of AdoMet and AdoHcy in vñrole blood of sheep
\,ùere examined during tle co¡:se of ttteasuremellt of tle transmethylation rate.
It was found, that there \4¡ere no changes in the levels of both compounds
up to 30 min after injection of POA. However, there \^las a slight rise
in AdoHcy and Adol4et levels 2 hr after POA administration and the mean
values of AdoHcy rose Lo 3.0 nmol/g of r,rhole blood from an undetectable
Ievel and AdoMet from 5.7 to 8.4 nmol/g of vileole blood. These results
indicate that ceIl membranes are relatively impermeable to AdoHcy and
AdoMet under the conditions investigated.
The overall rates of transmethylation shown in Table 9.2 vÍere
estimated from the initial rates of accumulation of tissue AdoHcy plus
Adol'4et. It is considered that the rates of accumulation of the sum of
AdoFIcy plus AdoMet should provide a better estimation of the true rate

L43
800
600
400
200
o
300
200
100
30 6() 90 120
Time (min)
Fig. 9.2. Time course of AdoHcy and AdoMet accumulation in tissues afterthe injection of POA. Sheep and rats were injected with 50 pmof/kg ofPOA intravenously and intraperitoneally, respectively. In the group
of sheep v¡here transmethylation rates lvere measured at 2 trr after POA
treatmenL, anoLher dose of POA \,{as given at t hr after the
first injection. Each point represents the mean value of 2-4 animals.
o, sheepliver; I, sheeppancreasi ar sheepkidney; orraL liver.
o3ant,
>rroeroۓ
oEc
oaaDaD
y!Of= (¡,6ì9 crt
õEc
o
4
3
2
1
o
()
oT'
o
o
=o!
oIo
CE
o
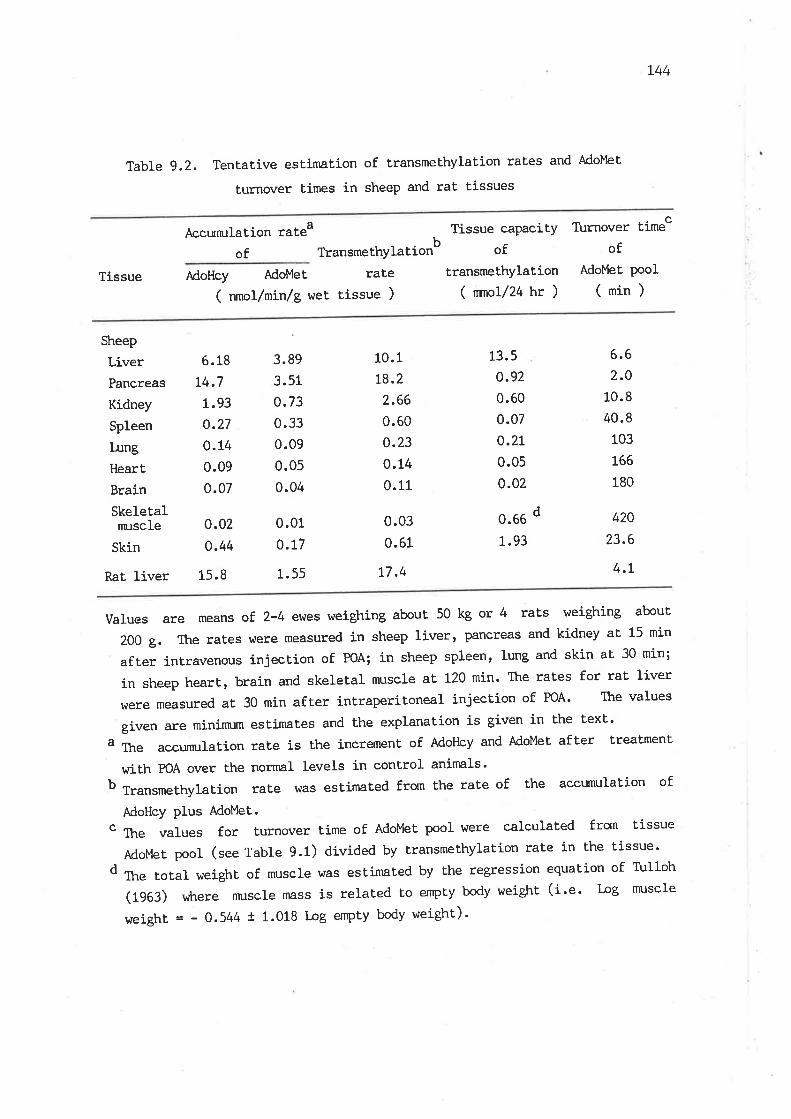
L44
Table 9.2. TentaLive estimation of transmethylation rales and Ado}'let
turnover times in sheep and rat tíssues
Tissue
Accunn:Iation rateaof Ttansmethylationb
Tissue capacityof
transmeEhylation( nmol/24 hr )
Turnover timec
ofAdoMet pool
(min)AdoHcy AdoMet rate
( nmol/min /8 wet tissue )
Sheep
LiverPancreas
Kidney
Spleen
I¡ngHeart
Brain
Skeletalnn:scIe
Skin
Rat liver
6.18
t4.71.93
o.27
o.t40.09
0.07
0.02
0.44
15.8
10.1
t8.22.66
0.60
o.23
0.14
0.11
0.03
0.61
t7.4
13.5
o.92
0.60
0.07
o.2t0.05
0.02
0.66
L.93
3.89
3.51
0.730.33
0.09
0.05
0.04
0.01
0.17
1.55
d
6.62.O
10.8
40.8
103
166
180
420
23.6
4.t
varues are means of. 2-4 ewes weighing about 50 kg or 4 rats weighing about
200 g. The rates were measured in sheep liver, Pancreas and kidney at' 15 min
after intravenous injection of PoA; in sheep spleen, Iung and skin aE 30 min;
in sheep heart, brain and skeletal nnrscle aE t2Q min' The rates for rat liver
vrere measured aE 30 min afEer intraperitoneal injection of PoA' The values
givenareminirn:mestimatesandtheexplanationisgiveninthetex!.
" Th" accunn:lation rate is the increment of AdoHcy and AdoMet after treatment
vrith POA over the normal levels in control animals'b TransmethylaLion rate was estimated from the raLe of the accunn'llation of
AdoHcy plus AdoMet.c The values for turnover time of AdoMet pool were calculated from tissue
AdoMe! pool (see Table 9.1) divided by Eransmethylation rate in the tissue'd Th" total weight of muscle was estimated by Ehe regression equation of Tulloh
(1963) where rnrscle mass is related to empty body weight (i.e. log muscle
weight = - 0.544 t 1.018 log empty body weight)'

14)
of transmethylation vùrich normally occurs in tissue rather than the rate
of AdoHcy accunruIation alone as used by Hoffman (fgAO). This is because
AdoHcy is lcrown to act as a cornpetitive inhibitor of many
methyltransferases and the rate of the AdoMet accumulation represents
that port.ion of the total transmethylation rate due to the inhibitory
effect of the elevated AdoHcy level, in turn produced by blocking
hydrolysis of AdoHcy. In contrast to rat liver, the rate of AdoMet
accu¡rnrlaLj-on in sheep tissues in the experimental conditions is
particularly significant.. In the tissues with very low rates of
transmethylation, the values were obtained at a time v¡hen measurable
amounts of AdoHey had accumulated.
A comparison of transmethylation rates and the turnover times of
AdoMet in tissues is given in Table 9.2, sheep liver had the great.est.
capacity of transmethylation among the t.issues of this species. However,
the pancreas possessed the highest. rate of transmethylation, although
the total capacity of transmethylation in this tissue was only
one-fifteenth of that in the liver due to the small nnss of the former
tissue. It is worth noting that sheep skin had a relat.ively high rate
of transmethylation cornpared with the rate in the brain, heart, spleen,
lung and skeletal nmscle. The transmethylation rate in sheep adipose
tissue vfas also measured and no measurable amount of AdoHcy
accumulation h/as found in the period of time studied. It can also be
seen frorn Table 9.2 that the transmethylation rate in rat liver was at
least 1.7 times higher than that. in sheep liver.
On the assurnption that AdoMet catabolism via transmelhylation
accounts for most of AdoMet utilization j-n manrnalian tissues (¡'tu¿¿ and
Poole, 1975; Itudd et al., 1980; Eloranta and Kajander, L984), it can be
calculated that the minimum values of turnover times of tissue AdoMet

t46
pools, based on the estimated rate of transmethylation, range from 2 min
in sheep pancreas to 7 hr in the muscle (table 9.2). It is worth
mentioning that the turnover tjme of AdoMet in sheep liver was similar
to that in human liver as estimated by }tudd et aI. (1980), using a
totally different experimental approach.
9.3.4 Effects of Sinmltaneous Administration of Methionine with POA
on AdoMet and AdoHcy levels
Since AdoHcy is a competitive inhibitor of nnny methyltransferases
with respect to AdoMet and changes in the ratio of AdoMet to AdoHcy
affect the transmethylation rate (CLriang and C,antoni, 1979; Schatz et
aI., 1-981a,b), methionine loading may elevate tissue AdoMet levels and
consequently enhance the rate of methyl transfer from AdoMet by
increasing Lhe ratio of AdoMet to AdoHcy. Fig. 9.3 shows that
sinmltaneous administration of methionine r¿ith POA elevated AdoMet
Ievels markedly in sheep liver and pancreas and part.ially reversed the
ratio of AdoMet to AdoHcy in these tissues (the ratios in the liver and
pancreas áre 1.0 and 0.29 in the POA-treated group, respectively, vs.
2.2 and 0.78 in the POA plus methionine-treated group). It can also
been seen that the AdoHcy level in the pancreas was more than double
with simultaneous administration of methionine and was significantly
increased in the liver. Methionine loading also significantly increased
Adolulet level, but not AdoHcy level, in sheep kidney, spleen and skin, as
compared to the values with administration of POA alone. However, io
sheep brain, heart, lung and skeletal nmscle, neither AdoMet levels, nor
Adotlcy levels r^/ere affected by methionine loading. These results
indicate that the rates of transfer of methyl groups from AdoMet in

800
a))8 600
.r{
o!tr +rooEB4oo
ò0
rloE
-9 200
600
400
200
OJ
ØØ.-l+)
¡Jo3ö0
JoE
+Jo2.o.d
Liver Pancreas Kidney Spleen Skin Brain Heart lung }fuscle
Fig. 9.3. Effects of simultaneous administration of methionine with POA on AdoHcy and AdoMet
len¡els in sheep tissues. In treated groups, sheep were given a dose of 50 ¡:mol/kg of POA r¿ith
or without methionine (85 mg/kg) intravenously and killed at 30 min after the injection. TTre
data are presented as the means t SÞ1 of three aninrals. Significance was determined for the
effects of POA plus methionine administration as compared to the POA-treated group in each
tíssue by Studentfs t-test. :k p ( 0.05, :'.:k p ( 0.01,'Å-:'ck p < 0.001.
50
40
30
20
10
00
0
60
40
20
_0
Ps.!
1-] No treatment: PoA
Ø POA + methionine

148
aresheep liver and pâncreas under the experimental conditions
stinulated by methionine loading, but not in other tissues.
9.3.5 In vitro Inhibition of Methyltransferases from Sheep Liver
by AdoHcy
To estimate the degree of the irùribition of transmethylation by
AdoHcy, guanidinoacetate, phospholipid and glycine methyltransfe¡ases,
vùrich are quantitatively predominant in AdoMet utilization (¡tl¿¿ et aI.,
1980; Dawson et aI., 1981; Mitchell and Benevenga, t976; Ogawa and
Fujioka, L982), \,ùere studied in vitro. Under the experimental
conditions used to estimate the transmethylation rate, both hepatic
AdoHcy and AdoMet levels reached about 100 nmol/g wet. tissue \^rith a
ratio of AdoMet to AdoHcy of t.2 at 15 min after the injection of POA
and the normal levels of hepat.ic AdoHcy and AdoMet were 14 and 67 nnoL/g
wet tissue, respectively. Therefore, 100 ¡rM AdoMet and 10-100 pM AdoHcy
\^/ere used in the enzyme assay systems. As shown in Table 9.3, t'hese
methyltransferases were all subject to a great degree of inhibition by
AdoHcy. Phospholipid methyltransferase was the most sensitive to the
intribition by AdoHcy and the activity was reduced by 74 7" aL 100 pM
Adollcy. A considerable degree of inhribition also occurred in the
physiological range of hepatic Adot{cy concentrations-
9.3.6 Kinetic Constants of Methyltransferases from Sheep Liver
In order to gain a better understanding on the physiological
regulation of transmethylation rate in sheep by the conrnon substrate and
product, it is necessary t.o ianow the Km values for AdoMet and Ki values
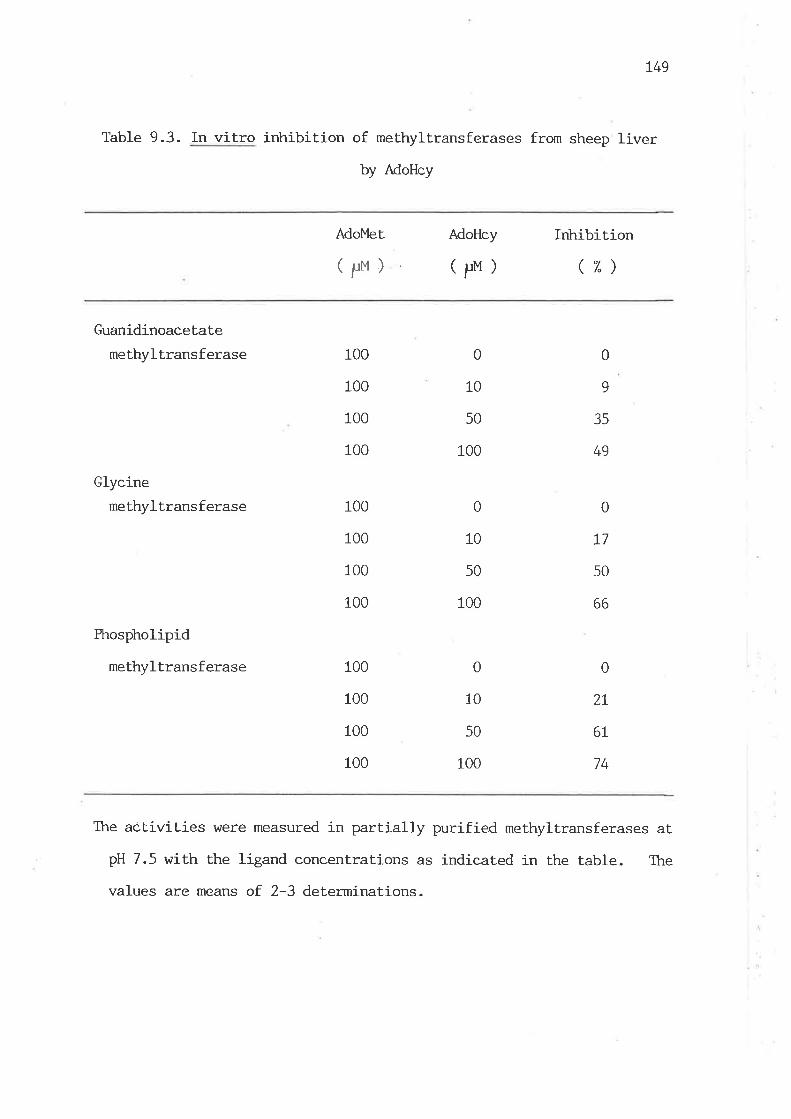
L49
Table 9.3. In vitro inhibition of methyltransferases frorn sheep liver
by AdoHcy
AdoMet AdoHcy
(Pr)Inhibition
(7")
Guanidinoacetate
methyltransferase
Glycine
methyltransferase
Phospholipid
methyltransferase
0
10
100
100
100
100
0
9
100
100
100
100
100
100
100
100
50
100
50
100
0
10
50
100
49
0
T7
50
66
6L
74
35
0
10
0
2L
The act.ivities !üere measured in partially purified methyltransferases at
pH 7.5 with the ligand concentrations as indicat.ed in the table. The
values are means of. 2-3 determinat.ions.

150
for AdoHcy for methyltransferases. The apparent Km of phospholipid
methyltransferase for AdoMet was delermined by measuring the overall
rate of the incorporaLion of methyl groups from AdoMet into phospholipid
with a testing range of AdoMet concentrations of 5-200 ¡:M. Reports on
the number of the methyltransferases involved in the s¡mthesis of PtdGro
from PtdEtn in rat liver are controversial (Tanaka et aI., L979; Sastry
et aI., 1981; Audubert and Vance, 1983). The possible existence of more
than one methyltransferase for stepwise methylation of PtdEtn in the
slmthesis of PtdGro in sheep liver was not investigated.
Table 9.4 sunnnarizes the kinetic parameters of three
AdoMet-dependent methyltransferases from sheep liver as shown in
Fig. 9.4-9.6. It can be seen that guanidinoacetate methyltransferase
had the lowest Km value for AdoMet, being 1-0 t.imes lower than those for
the other two methyltransferases. Ki values for AdoHcy for the three
methyltransferases were in the neighbourhood of physiological levels of
AdoHcy in lhe liver. Thus, it seems likely that a certain degree of
inhibition of transmethylation nay occur under physiological conditions.
The Km of guanidinoacetate methyltransferase in sheep liver for
guanidinoacetate is similar to the reported value in pig liver (f* et
aL., L979b), br¡t is considerably lower thran that in rat liver (Ogawa et
aL., 1983). The Km of glycine methyltransferase in sheep liver for
glycine was very high, about 100 times that for AdoMet. A similar hi-gh
Km of this enzyme for glycine vùas also observed in rabbit liver (Heady
and Kerr, t973).
9.4 Discussion
The present irnrestigation quantitatively demonstrated a wide range
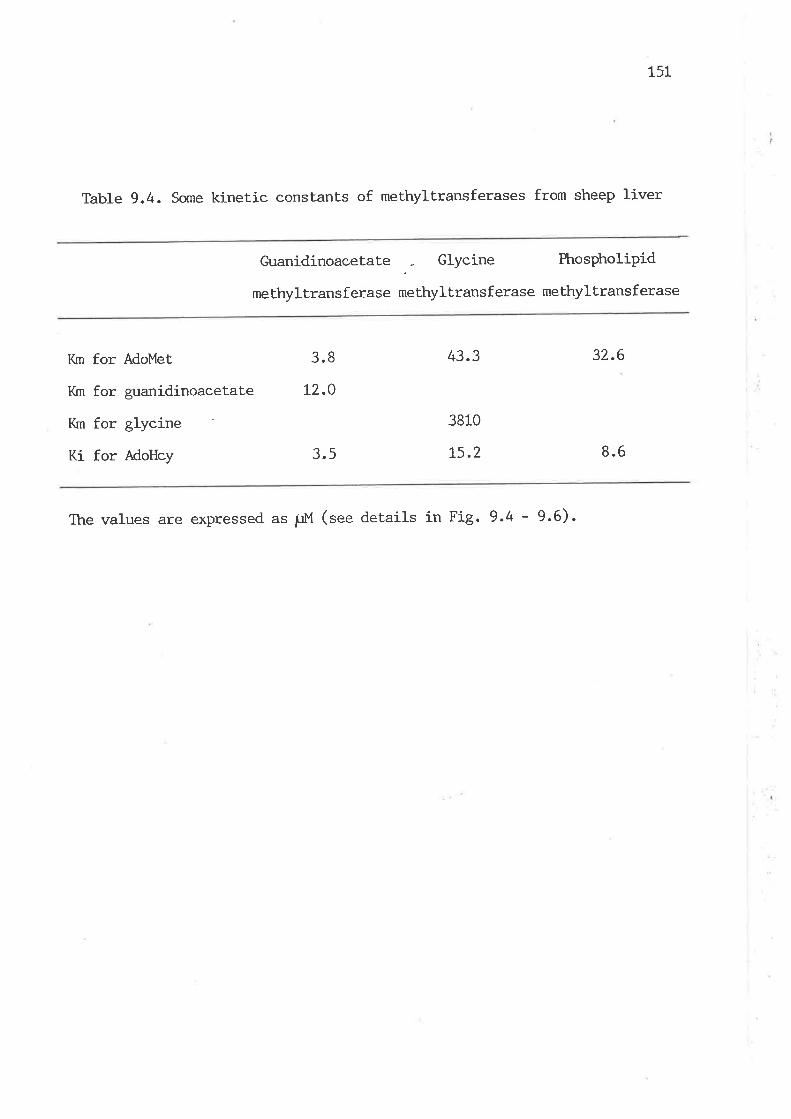
151
Table 9.4. Some kinetic constants of methyltransferases from sheep liver
Guanidinoacetate Glycine Phospholipid
me bhyl trans f erase me thyl trans f erase me thyl trans f erase
Ifu for AdoMet
I(m for guanidinoacetate
Km for glycine
Ki for AdoHcy
3.8
L2.O
3.5
43.3
3810
L5.2
32.6
8.6
The values are expressed as pM (see deLails in Fig. 9.4 - 9.6).
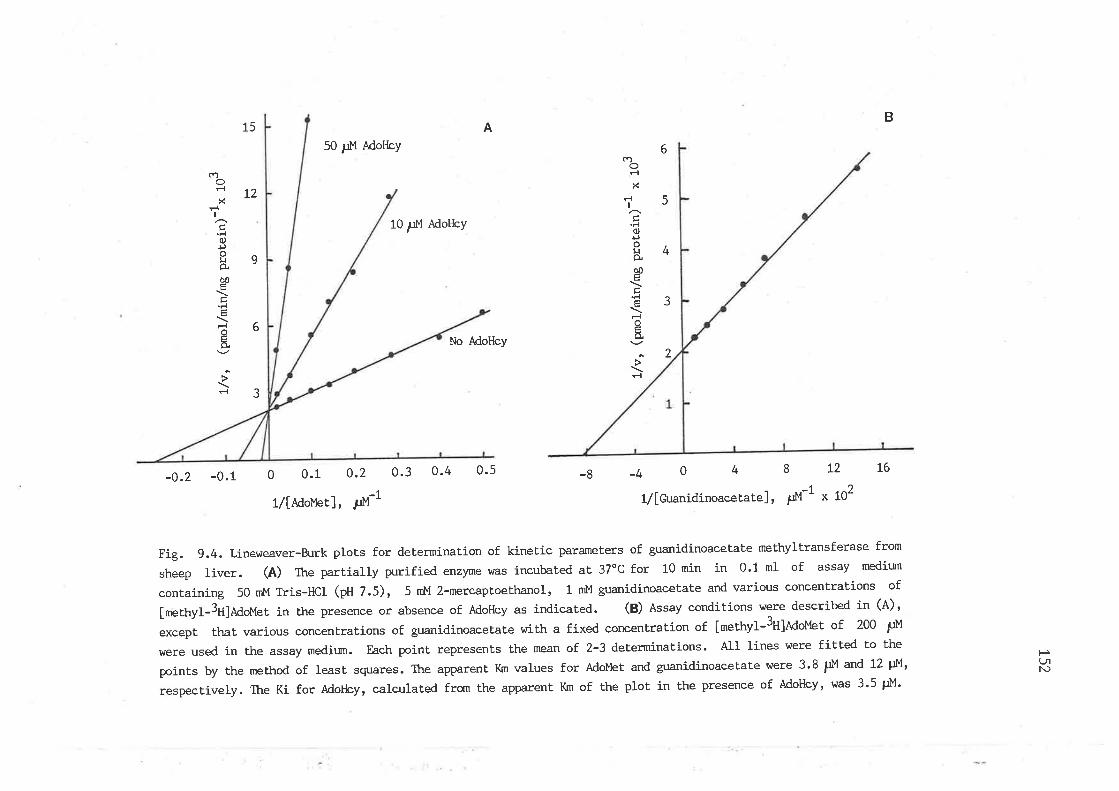
B15
t2
A
6
5
4
3
2
9
6
3
cnOFI
XF{
I
'F{o¡Joþâò0Eç
'F{È
F{
oõ.
FI
50 pM Adot{cy
10 ¡:M MoHcY
cîot-.1
xr'l
I
ç'Fl0,ÐoÞq00EÉ.r{
Jgã.
É.1
No Adotlcy
-0.2 -0.1 0 0.1 0.2 0.3 0.4 0.5 -40481/[GuanidinoacetaÈe], È,M-1
L2 t6-8
f/l¿douet], ¡rM-1
Fig. 9.4. Liner¿eaver-Burk ploEs for determination of kinetic parameters of guanidinoacetate methyrtransferase from
sheep liver. (A) The partially purified enzyme was incubated at 37oC for 10 min in 0.1 ml of assay mediurn
containing 50 nM Tris-HCl (pH 7.5), 5 mM 2-mercaptoethanol, 1 mM guanidinoacetate and various concentraÈions of
[mettryf-3H]MoMet in the presence or absence of AdoHcy as indicated. (B) Assay conditions were described in (A),
except that various concentrations of guanidinoacetate w'ith a fixed concentration of [methyl-3tt]ldot"tet of 200 PM
were used in the assay medir-un. Each point represenEs the mean of 2-3 determinalions. All rines were fitted Èo the
points by the method of least squares. The apparent Km values for AdoMet and guanidinoacetate were 3.8 ¡rM and 72 tN,
respectively. The Ki for Adotlcy, calculated from the apparent Km of the plot in the presence of AdoHcy, was 3'5 pM'
x ß2
P(-¡lN)

35
30
25
20
15
10
A
€t*e-a$'
NoFl
xFlt
É.F{
o+Jo!o.ò0E
..{
F{
oã.
Fl
100 pM Adotlcy
20 pM AdoFby
No AdoHcy
c\oFl
xr{
I
'F{o+JotJÊô0É,
tr'F{É,
F{I¡l
-{
75
t2
9
6
35
-0.25 0 0.25 0.5 0.75 ! 1.25-2-L0123456
l/[AdoMet], ¡rlt-1 * to21/[Gtycine], d-1
Fig. 9.5. Lineweaver-Burk plots for determination of kinetic parameters of glycine methyltransferase from sheep
liver. (Ð The partially purified enzyme hras incubated at 37oC for 20 min in 0.1 ml of assay mediun contaíning
50 ml4 Tris-HCf (pH 7.5)r 20 mM glycine, 1 mM DTI and various concentrations of [methyl-3H]AdoMet in the presence or
absence of AdoHcy as indicated. (B) Assay conditions were the same as described in (A), excePt that various
concentrations of glycine w-ith a fixed concentration of [methyl-3H]AdoMet of 300 pM were used in the assay medium'
Each poÍnt represents the mean of.2-3 determinations. All lines were fitted to the points by the method of least
squares. The apparent Iç¡ values for AdoMet and glycine were 43 pM and 3.81 mM, respectively' Ttre Ki for MoHcy,
calculaLed from the aPParent Km of this plot in the presence of AdoHcy, was 15'2 p!1'
HtJr(Ð
B
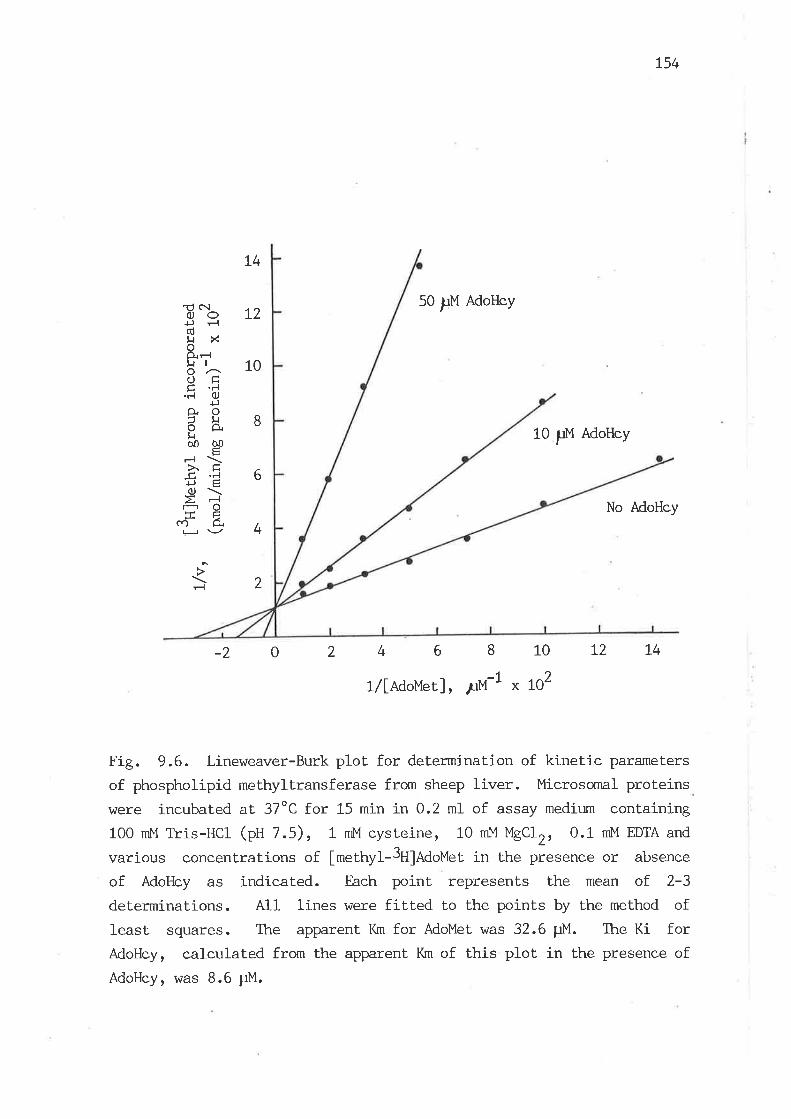
'U c\¡OOL¿+J rl(ú$rXoô,Ft
8'^ 10otrÉ .rl.r.l O
+Jo.o¿ ä 8lrò0 00
ËFl\.xÉ6'É bc,) \-E-l oFE
"'ì- ia 'lJ\-/+
L4
2
50 FM AdoHey
L54
No AdoHey
10 FM AdoHcy
rl
-20 24 6810t214
t/[Rdo¡,tet], ¡ùf-l x tO2
Fig. 9.6. Lineweaver-Burk plot for determinat.ion of kinet.ic parameters
of phospholipid methyltransferase from sheep liver. Microsomal proteins
\^rere incubated at 37oC for 15 min in 0.2 mI of assay medium containing100 mM Tris-HCl (pH 7.5), 1 mM cysteine, 10 mM UgClZ, 0.1 mM EDTA and
various concentrations of fmethyt-3H]n¿oUet. in Lhe presence or absence
of AdoHcy as indicated. Each point represents the mean of 2-3
determinations. All lines were fitted to the points by the method ofIeast. squares. The apparent Km for AdoMet was 32.6 ttM. The Ki forAdoHcy, calculated from the appa.rent Km of this plot in the presence ofAdoHcy, was 8.6 p1"1.

155
of transmethylation rates existing in sheep tissues- Among these, the
pancreas had the highest rate of transmethylat.ion, virich is consistent
with the distrib:tion of quantitatively inrportant methyltransferses in
sheep tissues (Ctrapter 4). Presumably, most of AdoMet utilization in
this tissue may be used for the synthesis of creatine and sarcosine from
glycine, since, unlike most of other species, the activities of
guanidinoacetate and glycine methyltransferases in sheep pancreas are
very nmch higher than those j-n the liver (C:rapter 4) ' In relation to
the quantitat.ive utilization of AdoMet, sheep liver was the predominant
tissue, vilrich accounted for approximately 757"of- tetotal-body capacity
for transmethylation. Particular attention has been paid to the
transmethylation rate in sheep skin, as Pisulewski and Butt.ery (fgaS)
proposed that the rate of conversion of methionine to cysteine in sheep
skin might be even faster than that. in the liver. The results from the
present investigation indicate that this may not be the case. However,
sheep skin possessed a'moderate rate of transmethylation, being 20 times
faster than thåt in the muscle, and its tissue capacity of
transmethylation is thus quantitatively significant.
It should be emphasized that the rates of tissue transmethylation
determined in this study are still an approximate estimation, even
though AdoMet accumulation was taken account of in the calculation of
transmethylation rate to correcl the decreased rate of transmethylation
drre to the inhribitory effect of AdoHcy on methyltransferases as shown in
Table 9.3. However, a certain degree of feedback ir¡tribition of AdoMet
on it.s own synthesis is likely to have occurred in the tissues with
elevated AdoMet levels. Another less likely factor v¡hich might
introduce so{ne error into the est.imation of transmethylation rate is the
unproven asstnnption of cornplete inkribition of AdoHcy hydrolase by KA'

r-56
although the dosage of POA used in the present study is at least twenty
thousand times higher than the Ki value for PoA for AdoHcy hydrolase
from calf liverr âs reported by Pateì--Thornbre and Borchardt (1985).
These above mentioned factors may have led to an underestimated rate of
transmethYlation-
One of the nrain purposes in this investigation was to obtain a
quant.itative assessment of body AdoMet requirement, v¡?rich is very
important. for quantitative understanding of methionine and AdoMet
metabolism. A tentative estimate of tte total-body requirement of AdoMet
for biological methylation, as calculated from the values of all the
measured tissue capacities of transmethylation, is at least 18 nnnoles
per day for a female sheep \^rith 5o-k8 body weight. In comparison with
humans, the estimated daily requirement of AdoMet for transmelhylation
in adult fe¡nale sheep is similar to that for adult young men on normal
dieLs, but higher than that for adult young \^Tomenr as estimated by
l,4udd and his colleagues (tgzs, 1980). It is interesting to note that
the proportion of the methyl group of AdoMet used for creatine syrthesis
in sheep \^ras no more than 55 7", vùrich is considerably less than that in
hurnans, âS a 5o-kg sheep excretes about LO nrnoles of creatine plus
creatinine daily (Henderson et al., 1933). Presumably, the amount of
AdoÙlel required for choline slmthesis in sheep may be greater than tlr¿t
in ht¡nans, since dietary choline is virtually unavailable for sheep due
to the degradation of dietary choline by rumen microbes (¡leill et al',
7g7g). The relatively high requirement of melhionine for choline
synthesi-s in a rulrninant species has been reported by F¡nrEnuel and
Kennelly (fgg+), vùro observed that approxinntely 28 % of methionine is
utilized for choline synthesis in goats, presumably, the situation of
choline nutrition in this ruminant species is sjmilar to that in sheep.

L57
However, the reported value for choline synthesis in sheep (Dawson et
a1., 1981) is 5-10 times that of methionine uptake from the gut
and seens likely to be overestimated, as compared with the value for
humans, goats and the data from the present investigation.
The present results also give a quantitative view on the
importance of in vivo synthesis of methionine, as the estimated value of
total-body requirement of AdoMet for methylat.ion in sheep i's 2-4 Limes
higher than the daily output of methionine from the gul as reported by
I,rTolff et al . (L972) and Lindsay (ßgZ). Therefore, it is clear that
more than half the amount of methionine required for maintaining
normal physiological funct.ion of sheep has to be synthesized via
methionine recycling, mainly from the 5-methylH4F'homocysteine
methyltransferase reaction (Chapter 4). Such a need for methionine
synthesis is even greater in lactating ewes (Cfrapter e). Furthermore,
it. has been demonstrated that a significant portion of methionine is
catabolized via the transamination pathway in sheep (Benevenga and
Egan, 1983). In adult sheep in metabolic st.eady sLates, a certain amount
of methionine is also required for polyamine synthesis and wool growth
vilrich represents irreversible loss of meLhionine from its body pool.
Obviously, the requiremenL for methionine in adult sheep musL be
considerably greater than the value required for transmethylation.
Thus, it further proves that the de novo synthesis of methyl groups
provides a major source of labile methyl groups in adult sheep.
Based on the Km values of the three methyltransferases and the
effect of methionine loading on tissue AdoMet levels, it can be deduced
Lhat ûr vivo rates of transmethylation in sheep pancreas and liver are
regulated by AdoMet and methionine, as the Km values for AdoMet. for
phospholipid and glycine methyltransferases r,æne in tìre neighbourhood of

158
physiological concentraLions, wtrich were affected by the avaíIability
of methionine, in these Lissues. This finding provides an explanation
for the reduced level of hepatic choline and the increased hepat.ic ratio
of PtdEtn to PtdCtro in vitamin Bl2-deficient sheep (Smittr et al., L974;
Iough et al., L982), owÍ-ng Lo a reduced amount of methionine synthesized
from the reaction of vitamin Bl2-dependent homocysteine methylation.
However, Adol'let levels in other tissues are not or much less regulated
by methionine. An explanation for this may lie 1n the fact that there
exist three isozymes of methionine adenosyltransferase with quite
different Km values for methionine as found in rat and human liver
(Hoffman and Kunz, 1980; Liau et a1., L979). One of the isozymes has a
high Km, vùrich is many times higher than the physiological level of
methionine in hunan and rat liver, vùrile in non-hepatic tissues
there is only one form of the enzyme for AdoMet synthesis with a
low I(m for methionine, vùrich is very much lower than its physiological
levels (Liau et al., t979; Carl et aI., 1978; Hoffman and Kunz, 1980;
Oden and Clarke, 1983).
It is possible that three isozymes of methionine
adenosyltransferase might exist in sheep liver as well as the pancreas,
as these two tissues showed a marked response of AdoMet levels to
methionine loading. The unresponsiveness of AdoMe! level to methionine
loading in the Lissues with a slow rate of transmethylation suggests
that the Km of methionine adenosyltransferase for methionine in these
tissues nay be well below its physiological level.
One of the interesting findings in the present investigation is
that there exists a positive relationship between the responsive
capacity of AdoMet level Lo excess methionine and the turnover rale of
AdoMet in sheep tissues. Such coordination is of importance in AdoNlet

t59
metabolism to ensure that the extra AdoMet synthesized from excess
methionine can be rapidly removed via transmethylation reactions. It. is
clear that AdoMet levels in the tissues v¡ith slow rates of
transmethylation and with insensitivity or relative insensitivity to
methionine loading are not or nnrch less affected by changes of
methionine availability. This nny explain the previous observations on
the different response of tissue AdoMet level to N2O-induced methionine
deficiency in sheep (Ctnpter 8).

160
CHAPTER 10. GM{RAL DISCUSSION
The results presenLed in this thesis clearly elucidate the main
mechanisms of adaptation of sheep to a low intake of methyl nutrients in
the ruminant sLate. The relatively low enzymatic capacities of choline
oxidation and AdoMet utilization for Lhe methylation of glycine in adull
sheep, aS compared with those in rats, would greatly reduce the
requirement for labile methyl groups. The develognental changes in
the specific activities of choline oxidase apPears to be closely
related to choline intake in sheep. It is tikely that the low level of
choline oxidase in post-ruminant sheep is a reponse to the virtual
unavailability of dietary choline. This is in accordance with the
findings of Schneider and Vance (1978) in regard to the regulation of
choline oxidase activity by dietary choline in rat liver. However, the
capacity for such nutrilional adaptation in rats is limited, as choline
deficiency could produce a marked reduction in the hepatic choline
content (Haines and Rose, L970; TLrompson et al., L969) and pathological
lesions (".g. f.aLly liver and haemorrhagic kidney) in rats (Kuksis and
Mookerjea, L978; Griffith et. al., t97L).
The extremely low level of glycine methyltransferase in adult sheep
liver is of significance in the conservaLion of the methyl group of
AdoMet, as the reaction product, sarcosine, has no physiological
function and this reaction is one of quantitaLively predominant
methylation reactions utilizing Adol4et, as observed in humans (ptu¿¿ et
aI., 1980) and in rats (t'titcfrell and Benevenga, 1976; ogawa and Fujioka,
t982b). Glycine methyltransferase activity could be further reduced
in sheep in the lactating state, v¡here there is an increased
labile methyl requirement for the synthesis of choline, creatine

r6t
and carnitine. Conversely, the enzyme activity could be dramatically
induced in severely diabetic sheep liver, vfrrere there l^7as a
surplus of AdoMet, owing to the decreased utilization of AdoMet for the
synthesis of choline and creatine and Lhe increased amount of methionine
flux through the transmethylation-transsulfuration rouLe. An increase
in the activity of glycine methyltransferase in alloxan-diabetic sheep
liver is nmch more pronounced than that observed in alloxan-diabetic rat
liver (unpublished data from Professor C. Inlagner, personal
connnrnication). E¡¿en the increase of glycine methyltransferase activiLy
in the liver of rats fed excess methionine is relatively small (Ogawa
and Fujioka, t982b; Tsukada et. aI., 1985), as compared to the dramatic
change of the enzyme activity in severely diabetic sheep liver. These
findings indicate that the capacity of glycine methyltransferase in
sheep liver under normal conditions is very small and a dramatic
increase of the enzyme activity has to be achieved in order to dispose
excess labile methyl groups.
It was found that there existed a direct relationship between
glycine methyltransferase and Y-cysLathj-onase in sheep liver,
as observed from the develo¡xnental changes in the specific
activities of these two enzymes and the effects of alloxan-diabetes on
the enzyme activities. The significance of this relationship is to
provide a means of disposal of excess methionine. There is a general
agreement in the literature that the glycine methyltransferase reaction
could be markedly stimulated by excess methionine (tntudd et a1., 1980;
Ogawa and Fujioka, I982b; Tsukada et al., 1985), vùtereas the activity of
phospholipid methyltransferase is not affecLed by excess methionine
(Tsukada et al., 1935) and reports on the effects of excess methionine
on the aclivity of guanidinoacetaLe mettryltransferase in rat liver are

162
conLroversial (Ogawa and Fujioka, L982b; Tsukada et aI., 1985).
Nevertheless, the rate of the in vivo synthesis of creatine is not
ínf luenced by excess methionine (l^/alker , 1979; lfudd et aL., 1980).
Therefore, glycine methyltransferase appears to be Lhe only enzyme v¡irich
could play a significant role in the disposal of surplus AdoMet in
rnanrnalian tissues.
Despite the virtual unavailability of dietary choline (Ueill et
al., L979) and the lack of exogenous creatine from the diet in
post.-ruminant sheep, the specific activities of the enzymes responsible
for choline and creaLine synthesis do not appear to increase from the
pre-ruminant to post-ruminanL state. An explanation for the
unresponsj-veness of guanidinoacetate methyltransferase to changes of
dietary crealine in sheep could be that the feedback control of creatine
synthesis is exerted aL Lhe amidinotransferase reaction rather than the
methyltransferase reaction, as observed in other species (l,Jalker, L979).
The increased enzymatic capacity for Ptdcho synthesis via the
transmethylation pathway in the liver of rats fed a choline-deficient
diet has been clearly demonstrated (t¡mbardi et a1., L969; Thompson et.
al., t969; Schneider and Vance, L918). However, there is no such
response of phospholipid methyltransferase in sheep liver to nutritional
changes of choline from the pre-ruminant to posL-ruminant state. These
findings suggest that the reduct.ion of choline oxidat,ion rather than
increase of its synthesis is the mechanism for the adaptation of
ruminant sheep to poor choline nutriLion. However, phospholipid
methyltransferase activity increased in sheep in the lactaLing status,
vùrere there is an exLra denrand for the synthesis of choline for the
secretion of milk, suggesting that this extra need for choline can not
be mct. by a furlher reduction of choline oxidation.

1,63
An estimation of total Adol4et requirement for biological
transmethylat.ion reaclions in sheep revealed that this requirement \^7as
at least 2-4 times the reported values for the daily uptake of
methionine from the gut (wotff eL al., L972; Lindsay, 1982). Most of
the methyl group of AdoMet. must be utilized for the synthesis of
creatine and choline in adult sheep, as the enzymatic capacity for the
methylation of glycine \tlas very small. Sheep liver is a predominant
tissue in AdoMet utilization and possessed approximaLeLy 757" of the
total-body capacity of AdoMet-dependenl Lransmethylat.ion. The major
source of the methyl group of methionine appears to originate from
methyl donor compounds in homocysteine methylation reactions. The
capacity of betaine-homocysteine methyltransferase for methionine
synthesis in sheep is relatively small, owing to a slow rate of choline
oxidation. Most of the methyl group of methionine is synthesized
via the 5-methylH4F-homocysteine methyltransferase react,ion in sheep,
as the body capacity of this enzyme was much greater than that of
betaine-homocysteine methyltransferase in this species.
A block of methionine methyl synthesis from 5-methylH4F in sheep
by N2O exposure led to a marked decrease in the concentrations of plasma
methionine and hepatic AdoMet and caused homocystinuria. These
alterations are often not observed in non-ruminant species with NZO
exposure (O:ranarin , L98L; Imb et al., 1983; Bukin and Orlov, 1984; van
der I,/esthuyzen and MeLz, 1983; Deacon et aI. , 1-980) r âs the
betaine-homocysteine meLhyltransferase reacLion in these species can
produce a substantial amount of methionine if dietary choline is noL
Iimited. The elevation of hepatic betaine-homocysteine methyltransferase
has been observed in N2O-exposed rats (Lumb et al., 1983), bul not in
N2O-exposed sheep. It has long been recognized that betaine or choline

164
could supporL the growth of rals fed a homocysteine diet free of
methionine (du Vigneaud et al., 1-939). These fíndings suggest
that sheep reLy more heavily on Lhe 5-methylH4F-homocysteine
methyltransferase reaction for labile methyl grouPs than most, if not
all, non-ruminant species.
It. can be deduced from the kinetic study of methylt.ransferases
that a decrease of hepatic AdoMet level in NtO-exposed sheep might
further lead to a reduced rate of choline synthesis, buL would not
affect the rate of creatine synthesis, as the Km of phospholipid
methyltransferase fiun sheep liver for AdoMet r,'as in the neighbourhood of
it.s physiological concentration, whereas the Km of guanidinoacetate
methyltransferase for AdoMet was one order of magnitude lower than the
nornnl tissue level of AdoMet. The relatively high Km of glycine
methyltransferase in sheep liver for AdoMet indicates that the in vivo
rate of this enzynratic reacLion could be directly regulated by changes
in the tissue levels of AdoMet.
The increase of the capacity of 5-methylH4F-homocysteine
methyltransferase can be considered as cre of the primary mechanisms for
adaptation of sheep to the lor¿ intake of methyl nutrients in the
post-ruminant. staLe. Changes in the specific activity of the hepatic
enzyme during developnent show an inverse relationship with those of
hepatic betaine-homocysteine methyltransferase, v¡trich i-s regulat.ed by
the availability of choline intake in sheep. Further increase in the
capacity of 5-methylH4F-homocysteine methyltransferase could occur in
sheep liver when there is a high demand for labile methyl groups, such
as during lactation. It should be noLed that the potential capacity of
an animal for the generation of the methyl group of 5-methylHOF is
relatively larger âS this is related to the metabolisrn of one-carbon

165
units, sources of vûrich (".g. serine, glycine, histidine and formate)
are abundant in the body.
The overall results suggest that a highly reduced raLe of methyl
group catabolism and a markedly elevated capacity of the de novo
synthesis of methyl groups are primary mechanisms for adaptation of
ruminant sheep to a very low intake of methyl nutrients. The capacity
of such metabolic adaptation to nutritional changes of methyl groups in
sheep appears to be much greater than that of raLs, v¡hich explains the
occurrence of pathogenesis in rats under the poor nutritional conditions
of methyl groups similar to those vùrich ruminant sheep normally
encounter.

166
BIBLIOGRAPITT
Abe, T., Okada, G., Teraob, H. and Tsukaù, K. (fSaZ¡ J. Biochern. 91,
1081-1084.
Anridson, G.A.E. and Asp, N.-G. (L982) Ann. Nutr. Metab. 26, I2-t7.
Audubert, F. and Vance, D.E. (fgg:) J. BioI. Ckrem. 258, 10695-10701-.
Audubert, F., Pelech, S.L. and Vance, D.E. (1934) Biochim. Biophys.
Acta 792, 348-357.
Awad, I^/.M.Jr., Whitney, P.L., Skiba, W.E., Mangum, J.H. and Wells, M.S.
(1983) J. Biol. Cbrem. 258, t2790-72792.
Backlund, P.S.Jr. and Smith, R.A. (fggf) J. Biol. Crem. 256, 1533-1535.
Backlund, P.S.Jr., Chr,ang, C.P. and Smith, R.A. QSAZ) J. Biol. Ctrem.
257, 4t96-4202.
Baldessarini, R.J. and Kopin, I.J. (fSe:¡ Anal. Bioche¡n. 6, 289-292.
Baldessarini, R.J. (f900) Biochem. Pharmacol. 15, 74t-748.
Baldessarini, R.J. and Kopin, I.J. (1966) J. Neurochem. L3, 769-777.
Baldessarini, R.J., Stramentinoli, G. and Lipinski, J.F. (fgZA) fn
"Biochemical and Pharrnacological Roles of Adenosylmethionine and
the Central Nervous System" (Z,appta, V., Usdin, E. and Salvatore,
F., eds.), pp. L27-L40, Pergamon Press, Oxford.
Barak, A.J., Beckenhauer, H.C. and TLrma, D.J. (neZ¡ Anal. Biochem.
t27, 372-375.
Barry, T.N., Fennessy, P.F. and Duncan, S.J. (tglZ) N. Z. J. Agric.
Res. 16, 64-68.
Benevenga, N.J. and tlarper, A.E. (1-970) J. Nutr. 100, I2O5-L2L4.
Benevenga, N.J. (tglt+) J. Agric. Food Chrem. 22, 2-9.
Benevenga, N.J. and Haas, L.G. (1979) J. Ar¡im. Sci. 49 (Suppl. 1,),94.

t67
Benevenga, N.J. and Egan, A.R. (fgg:) Prog. Clin. BioI. Res. I25,
327-34L.
Benevenga, N.J., Radcliffe, B.C. and Egan, A.R. (fg8:) Aust. J. Biol.
Sci. 36, 475-485.
Benevenga, N.J. (1984) Adv. Nutr. Res. 6, 1-18.
Bernheim, F. and Bernheim, M.L.C. (fg::) Am. J. Physiol. IO4, 438-440
Bernheim, F. and Bernheim, M.L.C. (fg:A) Am. J. Physiol. LzI, 55-60.
Binkley, F., Anslow, W.P;Jr. and du Vigneaud, V. (L942) J. Biol. Chem.
743,559-560.
Binkley, F. (1951) J. Biol. Chem. L9L, 53t-534.
Bltrnenstein, J. and l,Jilliams, G.R. (1960) Biochem. Biopìeys. Res.
C-omnnrn. 3, 259-263.
Blunenstein, J. and hiilliams, G.R. (fgO:) Can. J. Biochem. Physiol . 4!,
20r-2L0.
Blusztajn, J.K., Tei.seL, S.H. and l,Jurtnran, R. J. (tglg) Brain Res. 179,
319-327.
Boers, G.H., Smals, 4.G., Trijbels, F.J., l-eermakers, A.I. and
Kloppenborg, P.tn7. (fgg:) J. Clin. Invesr. 72, L97!-1976.
Borsook, H. and Dubnoff, J.W. (fg+O) J. Biol. Chem. L32, 559-514.
Borsook, H. and Dubnoff , J.W. (tgt*t) J. BioI. Crem. 138, 389-403.
Borsook, H. and Dubnoff , J.W. (1947) J. Biol. Chem. L69, 247-258.
Bremer, J., Figard, P.H. and Greenberg, D.M. (fg0O) Biochim. Biophys.
Acta 43, 477-488.
Bremer, J. and Greenberg, D.M. (fg6f) Biochim. Biophys. Acta 461 205-216.
Bukin, Yu.V. and Orlov, E.N. (fga+) Vestn. Akad. l4ed. Nauk SSSR 5,
9-L4.
Bnrke, G.T., I'langum, J.H. and Brodie, J.D. (L97I) Biochemistry 10,
3079-3085.

168
cabrero, c., Merida, I. , orLiz, P. varela, I. and Mato, J.M. (rga0) "*
Biochem. Pharmacol. 35, 226L-2264.
C,antoni, G.L. (L952) J. Am. Chrem. Soc. 14, 2942-2943.
Cantoni, G.L. and Vignos, P.J. Jr. (fgS+) J. Biol. Chem. 209, 647-659-
Cantoni, G.L. (1975) Ann. Rev. Biochem. 44, 435-45L.
Cantoni, G.L. (t977) In "The Biochemistry of Adenosylmethionine"
(Salvatore, F., Borek, 8., Zappi'a, V., I'Jilliams-Ashman, H.G. and
Schlenk, F., eds.)r PP. 557-577, Coltlnbia University Press, New
York.
Cantoni, G.L. , Richards, H.H. and Chiang, P.K. (L979) In
"TransmeLhylalion" (Usdin, E., Borchardt, R.T. and Creveling,
C.R., eds.)r pp. L55-L64, Elsevier/North Holland, New York.
C,antoni, G.L. and Chiang, P.K. (fOaO¡ In "Natural Sulfur Compounds"
(Cavallini, D., Gaull, G.E. and Zappia, V., eds.), pp. 67-80,
Plenum Press, New York.
Carl, G. F., Benesh, F.C. and Hudson, J.L. (1978) Biol. Psychiat. L3,
66L-669.
C,arlson, M. and van Pilsum, J.F. (1973) Proc. Soc. Exp. Biol. Med.
L43, L256-L259.
Carrol, W.R., Stacy, G.W. and du Vigneaud, V. (tg+g) J. BioI. Chem.
180,375-382.
C,ase, G.L. and Benevenga, N.J. (t976) J. Nutr. 106, Ll2L-t736.
Case, G.L., Mitchell, 4.D., Harper, A.E. and Benevenga, N.J. (L976)
J. Nutr. 106, 735-746.
case, G.L. and Benevenga, N.J. (1977) J. Nutr. L07, L665-I676.
Castafio, J.G., Alemany, S., Nieto, A. and Mato, J.M. (fggO) J. BioI.
Chem. 255,9041,-9043.
Chalupa, I,l. (tglZ) Fcd. Proc., Fed. tun. Soc. Exp. Biol. 31 , LL52'LL64.

L69
Cllranarin, I. and Bennett,, M.C. (tgøZ) Br. Med. J. L, 27'29.
Chanarin, I. (1980) J. Clin. Pathol. 33, 909-916.
Chanarin, I. (1931) Lancet ii, 755.
Chavez, E.R. and Bayley, H.S. (tglí) Br. J. Nutr. 36, 189-198.
Chiang, P.K. and Cantoni, G.L. (t979) Biochem. Pharmacol. 28, L897'L902.
Cooper, A.J.L. and Meister, A. (tSlZ¡ Biochemistry LI, 66L-67L.
Cooper, A.J.L. and Meister, A. (1974) J. Biol. ClÌrem. 249, 2554-256t.
Cooper, A.J.L. (L977) J. Biol. Chem. 252, 2032-2038
Cooper, A.J.L. (1983) Ann. Rev. Biochem. 52, L87-222.
Cox, R. Prescott, C. and lrving, C.C. (L977) Biochim. Biophys. Acta
474, 493-499.
Crews, F.T., Hirata, F. and Axelrod, J. (fgAO) J. Neurochem. 34,
L49L-L498.
Crim, M.C., C,alloway, D.H. and Margen, S. (tglî) J. Nutr. 106, 371-331.
Daniel, R.G. and !üaisman, H.A. (fgega) J. Neurochem. t6, 787-795.
Daniel, R.G. and Waisman, H.A. (1969b) J. Nutr. 99, 299-306.
Davis, S.R., Bickerstaffe, R. and Hart, D.S. (fgZA) Aust. J. BioI. Sci.
3L, t23-L32.
Dawson, R.M.C., Grime, D.ld. and Lindsay, D.B. (fgAf) Biochem. J. 796,
499-504.
Deacon, R., Lumb, M., Perry, J., Chanarin, I., Minty, B., Halsey, M.J.
and Nunn, J.F. (1978) I-ancet ü, I023-L024.
Deacon, R., hmb, M., Perry, J., Ckranarin, I., Minty, 8., Flalsey, M.J.
and Nunn, J.F. (1980) Eur. J. Biochem. I04, 4L9-422.
Deacon, R., Perry, J., hmb, M. and Chanarin, I. (1983) Eur. J.
Biochem. 129, 627-628.
Deguchi, T. and Barchas, J. (L97L) J. BioI. Ckrem. 246, 3L75-3181.
Doerfler, I,rl. (1-983) Ann. Rev. Biochem. 52, 93-L24.

L70
Du Vigneaud, V., Ctmndler, J.P., Moyer, A.W. and Keppelr D.M. (fg:g)
J. Biol. Chem. 1.3L, 57-76.
Du Vigneaud, V., Ckrandler, J.P., fuh, M. and Brown, G.B. (fg+O) J.
Biol. Grem. L34, 787-788.
Du Vigneaud, V., bh, M., Chandler, J.P., Schenck, J.R. and Sínrnonds,
S. Jr. (f94L) J. BioI. Chrem. L4O, 625-64L.
Du Vigneaud, V. (tgSZ) A Trial of Research in Sulfur Chemistry and
Metabolism, Cornell University Press, Ithaca, New York.
Du Vigneaud, V. and Rachele, J.R. (fg6S) In "Transmethylation and
Methionine Biosynthesis" (Shapiro, S.K. and Schlenk, F., eds.), pp.
L-20, Univ. of Cfricago Press, Ckricago.
Dudman, N.P.B. and l{ilcken, D.E.L. (1982) Biochem. Med. 27, 244-253.
Duerre, J.A. and Miller, C.H. (fge6) Anal. Biochem. L7, 3LO-3L5.
Duerre, J.A. and l,rlalker, R.D. (L977) In "The Biochemistry of Adenosyl-
methionine" (Salvatore, F., Borek, E., Zappia, V., Williams-Ashman,
H.G. and Schlenk, F., eds.), pp. 43-57, Coltrnbia University Press,
New York.
Duerre, J.4., hlallwork, J.C., Quick, D.P. and Ford, K.M. (L977) J.
Biol. Ckrem. 252, 598L-5985.
Eells, J.T., Black, K.4., Makar, 4.8., Tedford, C.E. and Tephly, T.R.
(1OSZ¡ Arch. Biochem. Biophys. 2L9, 3t6-326.
Eloranta, T.O., Kajander, E.O. and Raina, A.M. (tglA) Biochem. J. 160,
287-294.
Eloranta, T.O. (1977) Biochem. J. L66, 52L-529.
Eloranta, T.O. and Kajander, E.0. (fgg+) Biochem. J. 224, L37-L44.
E¡nrnanuel, B. and Kennelly, J.J. (fgg+) J. Diary Sci. 67, I9L2-1918.
Engstrom, M.A. and Benevenga, N.J. (1981) Fed. Proc., Fed. Am. Soc.
Exp. Biol. 40, 84L.

flL
Ericson, L.E. (fg6O) Acta Chem. Scand. t4, 2LQ2-2LL2.
Fahey, R.C., Newlon, G.L., Dorian, R. and Kosower, E.M. (1981) Anal.
Biochem. LLL, 357-365.
Fau, D., Bois-Joyeux, 8., C,hanez, M., Delhonrne, B. and Peret, J. (fg8f)
Reprod. Nutr. Develop. 2L, 5L9'529.
Felig, P., Inlahren, J., Sherwin, R. and Palaiologos, G. (L917) Arch.
Intern. Med. L37, 507-513.
Finkelstein, J.D. (L967) Arch. Biochem. Biophys. 122, 583-590.
Finkelstein, J.D. and Mudd, S.H. (t961) J. Biol. Chem. 242, 873-880.
Finkelstein, J.D. (1970) In "sulfur in Nutrition" (t"tuth, 0.H. and
Oldfield, J.F., eds.)r PP. 46-60., Avi Press, I,{estport, Conn.
Finkelstein, J.D. (L97t) In "Inherited Disorders of Sulfur Metabolism"
(Carson, N.A.J. and Raine, D.N. , eds. ) , Pp. I-13, Ckrurchill
Livingstone, Edinburgh.
Finkelstein, J.D., Kyle, I^I.E. and Flarris, B.J. (tglt) Arch. Biochem.
Biophys. L46, 84'92.
Finkelstein, J.D., Harris, B.J. and Kyle, Il.E. (lglz) Arch. Biochem.
Biophys. 153, 320-324.
Finkelstein, J.D. (tglS) In "Metabolic Pathways", 3rd edn., VoI. VII
(Greenberg, D.M., ed.), pp. 547-597, Academic Press, New York.
Finkelstein, J.D., Kyle, W.E., Mart.in, J.J. and Pick, A.M. (1975)
Biochem. Biophys. Res. Conrnun. 66, 81-87.
Finkelstein, J.D. (L979) In "Transmethylation" (Usdin, E., Borchardt,
R.T. and Creveling, C.R., eds.), pp.49-58, Elsevier/North
Holland, New York.
Finkelstein, J.D., tlarris, 8.J., Martin, J.J. and Kyle, \.{.E. (tgïZa)
Biochem. Biophys. Res. Connnun. l-08, 344-348.

172
Finkelstein, J.D., Martin, J.J., Flarris, B.J. and Kyle, W.E- (1982b)
Arch. Biochem. Biophys. 2L8, L69-I73-
Finkelstein, J.D., Martin, J.J., Élarris, B.J. and Kyle, Inl.E. (fga:) J.
Nutr. LI3, 519-521.
Finkelstein, J.D. and Martin, J'J' (1-986) J' Biol' cltrern' 261, t582-L587'
Folch, J., Lees, M. and Sloane-Stanley, G.H. (tgSl) J. Biol. Chem.
226,497-509.
Forker, L.L., Chaikoff, I.L., EnLenman, C. and Tarver, H. (1951) J.
Biol. Chem. 188, 37-48.
Gaitonde, M.K. (L967) Biochem. J. L04, 627-633.
Ganguly, P.K., Rice, K.M., Panagia, V. and DhaIIa, N.S. (1984) Circ.
Res. 55, 504-5L2.
Gaull, G.E., Sturman, J.A. and Raiha, N.C.R. (L972) Pediatr. Res. 6,
538-547.
Gawthorne, J.M. and Smith, R.M. (1974) Biochem. J. L42, LL9-t26.
Gibson, J. and van der Ilesthuyzen, J. (1934) Internal. J. Vit. Nutr.
Res. 54, 329-332.
Giorgio, J.D. (t974) In "Clinical Chemislry: Principles and Technics",
2nd edn. (Henry, R.J., Cannon, D.C. and l{inkelman, J.W., eds.),
pp. 541-553, llarper and Row Publishers Inc., U.S.A..
Glick, J.M. and l.eboy, P.S. (L977) J. Biol. Ctrem. 252, 4790-4195.
Glick, J.M., Averyhart, V.M. and Leboy, P.S. (fgZA) Biochim. Biophys.
Acta 518, L58-L7L.
Greenberg, D.M. (1962) In "Methods in Enzymology" (Colowick, S.P. and
Kaplan, N.O., eds.), Vol. 5r pP. 936-942, Academic Press, New York-
Greenberg, D.M. (1963) Adv. Enzymol. 25, 395-43L.
Greenberg, D.M. (L975) rn "Metabolic Pathwaysrr, vol. vrr, 3rd edn.
(Greenberg, D.M., ed.), pp. 529-534, Academic Press, New York.

L73
Griffith, W.H., Nyc., J.F., tlarris, R.S., Flartroftr I'J.S. and Porta, E.A.
(L971,) rn "The vitamins'r, 2nd edn. (Sebrell, I,J.H. Jr. and [larris,
R.S., eds.), Vol. 3, pp. l-t54, Academic Press, New York.
Guerinot, F. and Bohuon, C. (L977) Eur. J. Cancer L3, 1'257-L259.
Guha, S. and Ilegmann, R. (1963) Nature (ton¿on) 200, t2I8-t2L9.
Haeckel, R. (fg8f) clin. Chem. 27, I79-L83.
tlaines, D.S.M. and Rose, C.I. (1970) Can. J. Biochem. 48, 885-892.
Hancock, R.L. (f900) Cancer Res. 26, 2425-2430.
Handler, P. (L949) Proc. Soc. Exp. Biol. Med. 70, 70-73.
Handler, P. and Bernheim, F. (L949) Proc. Soc. Exp. Biol. Med. 72,
569-57L.
Hansen, D.K. and Billings, R.E. (1985) Dev. Pharmacol. Ther. 8, 43-54.
Élarper, 4.E., Benevenga, N.J. and l,lohlhueter, R.M. (1970) Physiol.
Rev. 50, 428-558.
Hatefi, Y. and Stiggall, D.L. (L976) In "The Enzyme", 3rd edn. (Boyer,
P.D., ed.), Vol. XIIIr pp. 175-297, Academic Press, New York.
Haubrich, D.R. and Gerber, N.H.(fggf) Biochem. Pharmacol. 30, 2993-3000.
Heady, J.E. and Kerr, S.J. (L973) J. Biol. Chem. 248, 69-72.
Heady, J.E. and Kerr, S.J. (L975) C,ancer Res. 35, 640-643.
Heady, J.E. (L979) Develop. Biol. 68, 624-630.
Hebb, C., Mann, S.P. and Mead, J. (L975) Biochem. Pharmacol . 24,
1007-1011.
Heinonen, K. (tglS) Biochem. J. 136, 1011-1015.
Helland, S. and Ueland, P.M. (fgg:) Cancer Res. 43, 1847'1850.
Henderson, G.0., Xue, G.P. and Snoswell, A.M. (1983) Comp. Biochem.
Physiol. 768, 295-298.
Herbert, J.D., Coulson, R.A. and Hernandez, T. (fg66) Comp. Biochem.
Physiol. 17, 583-598.

Ll4
Hirata, F. and Axelrod, J. (1978) Proc. Natl. Acad. Sci. USA 75,
2348-2352.
Hirata, F., Viveros, 0.H., Diliberto, E.J. and Axelrod, J. (1978) Proc.
Natl. Acad. Sci. USA 75, t7L8-172L.
Hirata, F. and Axelrod, J. (1980) Science 209, L082-1090.
Hirata, F. (L982) In "Tkre Biochemistry of S-Adenosylmethionine and
Related Compounds" (Usdin, E., Borchardt, R.T. and Creveling,
C.R., eds.)r pp. L09-LL7, Macmillan, London.
Hof fman, D.R., Cornatzer, W.E. and Duerre, J.A. (L979) Can. J. Biochem.
57, 56-65.
Hof fman, D.R., Marion, D.W., Cornatzer, W.E. and Duerre, J.A. (1980)
J. Biol. C?rem. 255, t0822-I0827.
Hoffman, D.R., Haning, J.A. and Cornatzer, W.E. (fgAf) Proc. Soc. Exp.
Biol. Med. L67, L43-I46.
Hoffman, J. (tglS) Anal. Biochem. 68, 522-530.
Hoffnran, J.L. and Kunz, G.L. (L977) Biochem. Biophys. Res. Commun. 77,
t23L-L236.
Hoffman, J.L. (fgAO) Arch. Biochem. Biophys. 205, L32-I35.
Hoffman, J.L. and Kunz, G.L. (1980) Fed. Proc., Fed. Am. Soc. Exp.
Biol. 39, L690.
Holub, B.J. (fgZA) J. Biol. Ctre¡n. 253, 686-69I.
Hopper, J.H. and Johnson, B.C. (1956) Proc. Soc. Exp. Biol. Med. 9t,
497-499.
Huszar, G., Koivisto, V., Davis, E. and Felig, P. (1982) Metabolism.
3L, 188-191.
Ikeda, T., Konishi, Y. and lchihara, A. (L976) Biochim. Biophys. Acta
445,622-63I.
Im, Y.S., Cantoni, G.L. and Chiang, P.K.(tglga) AnaI. Biochem. 95, 87-88.

Ll5
Im, Y.S., Cltriang, P.K. and C,antoni, G.L. (1979b) J. Biol. Ckrem. 254,
11047-11050.
Itzhaki, R.F. and Gill, D.M. (L964) Anal. Biochem. 9' 40L-4t0.
Ivy, J.H., Svec, M. and Freeman' S. (fgSf) Am. J. Physiol - 167,
t82-L92.
Jefferson, L.S., Liao, W.S.L., Peavy, D.E., l'4iller, T-8., Appelr M.C.
and Taylor, J.M. (fgeg) J. Biol. Chem. 258, I369-L375.
Jenness, R. (1974) In "I-actation: A Comprehensive Treatise"(I-arson, B.L.
and SmiLh, V.R., eds.), Vol. 3, PP. 3-t07, Academic Press, NewYork.
Johnson, B.C., Mitchell, H.H. and Pinkos, J.A. (fgSf) J. Nutr. 43,
37-47.
Johnson, J.D. and cornalzer, I,J.E. (rg0g) Proc. soc. Exp. Biol. Med.
L3L,474-478.
Jones, P. and Taylor, S. (fgaO) CeIl 20, 85-93.
Kalnitsky, Ã., Rosenblatt, D. and Zlotkin, S. (nAZ) Pediatr. Res. 16,
628-63L.
Kang, S.-S., I,{ong, P.I^I.K. and Becker, N. (tglg) Pediatr. Res. 13,
tt4L-tL43.
Kashiwamata, S. and Greenberg, D.M. (1970) Biochim. Biophys. Acta 2L2,
488-500.
Kelly, K.L., Kiechle, F.L. and Jarrett' L. (1984) Proc. NatI. Acad.
Sci. USA 81, L089-L092.
Kensler, C.J. and langemann, H. (1954) Proc. Soc. Exp. Biol. Med. 85,
364-361.
Kerr, S.J. (L972) J. Biol. Chem. 241 , 4248'4252.
Khader, V. and Rao, S.V. (1932) Ann. Nutr. Metab. 26, 353'359.
Kiechle, F.L., l4alinski, H., Strandbergh, D.R. and Artiss, J-D. (1986)
Biochem. Biophys. Res. Commun. I37, L-7.

L76
Kim, G.S., Chevli, K.D. and Fitch, C.D. (1933) Experientia 39,
1_360-L362.
Kim, S. (t974) Arch. Biochem. Biophys. t6L, 652-657.
Klee, W.4., Richards, H.H. and Cantoni, G.L. (1961) Biochim. Biophys.
Acta 54,757-L64.
Kolhouse, J.F. and Allen, R.H. (1977) Proc. Nat.l. Acad. Sci. USA J4,
92r-925.
Kondo, H., Osborne, M.L., Kolhouse, J.F., Binder, M.J., Podell, E.R.,
Ut.ley, C.S., Abrams, R.S. and Allen, R.H. (1981) J. Clin. Invest.
67, L270-L283.
Kraus, J., Packnan, S., Fowler, B. and Rosenberg, L.E. (1978) J. Biol.
Ctrem. 2531 6523'6528.
Kredich, N.M. and Hershfield, M.S. (1980) Adv. Enzyme. Regul. 18,
181-191.
Kuksis, A. and Mookerjea, S. (1978) Nutr. Rev. 36, 2OL-207.
Kunz, G.L., Hoffman, J.L., Chia, C.-S. and Stremel, B. (fgAO) Arch.
Biochem. Biophys. 202, 565-572.
Kutzbach, C. and Stokstad, E.L.R. (tglt) Biochim. Biophys. Acta 250,
459-477.
Liau, M.C., Chang, C.F., Belanger, L. and Grenier, A. (tglg) Cancer
Res. 39, L62-L69.
Lindsay, D.B. (nAZ) Fed. Proc., Fed. Arn. Soc. Exp. Biol .4tr 2550-2554.
l,ombardi, 8., Pani, P., Schlunk, F. and Chen, S.-H. (fg6g) Iipids 4,
67-75.
Lombardini, J.B. and Talalay, P. (tglt) Adv. Erzyme. Regul. 9, 349'384.
I-ombardini, J.8., Chou, T.C. and Talalay, P. (1973) Biochem. J. 135'
43-47.

t77
longenecker, J.B. and Hause, N.L. (1959) Arch. Biochem. Biophys . 84,
46-59.
Iough, 4.K., Duncan, Id.R.H., EarI, C.R.A. and Coutts, L. (1982) Proc.
Nutr. Soc. 4t, L6^.
trmb, M., Perry, J., Deacon, R. and CLranarin, I. (1981) Am. J. Ctin.
Nutr. 34, 24L2-24t7.
Ixmb, M., Sharer, N., Deacon, R., Jennings, P., Rrrkiss, P., Perry, J.
and Clranarin, I. (fgg:) Biochim. Biophys. Acta 756, 354-359.
I-r:nd, P. (fgao) FEBS Lett. tL7, K86-92.
Lykken, G.I., Jacob, R.4., Munoz, J.M. and Sandstead, H.H. (fggO) Am.
J. Clin. Nutr. 33, 2614-2685.
Maclean, I,i.C. Jr., Graham, G.G., Placko, R.P. and I-åpez de Ronaña, G.
(1983) J. fturr. rL3, 779-785.
M..y, I.G. and Kelly, H.J. (fg6f) In "Milk: the Ùbnmary Gland and its
Secretion" (Kon, S.K. and Cowie, 4.T., eds.), VoI. 2¡ pp. 265-304,
Academic Press, Iondon.
l',langum, J.H., Steuart, B.W. and North, J.A. (tglZ) Arch. Biochem.
Biophys. 148, 63-69.
Mårtensson, J. and Hermansson, G. (fggA) Metabol-ism 33, 425-428.
lulartin, F. (rgZS) rn "Metabolic Pathways" (Greenberg, D.M., ed.), vol.
VII, pp. 457-503, Acadernic Press, New York.
Mato, J.M. and Alemany, S. (fg8:) Biochem. J. 2L3, L-t}.
Mato, J.M., Pajares, M.A. and Varela, I. (1984) TIBS 9, 47L-472.
Mat.thews, R.G. and Daubner, S.C. (tggZ) Adv. Enzyme. Regul. 20, L23-L3I.
l4cGi-lvery, R.W. and Goldstein, G.t^/. (1933) Biochernistry: A Functional
Approach, pp. 633-636, Saunders, W.B. Cornpany, Japan.
Mepham, T.B. and Linzell, J.L. (fq00) Biochem. J. 101' 76-83.
Mirchell, A.D. and Benevenga, N.J. (tglø) J. ltutr. 106, L7O2-17L3.

L78
Mitchell, A.D. and Benevenga, N.J. (1978) J. NuEr. 108, 67-78.
Mitchell, 4.D., CLrappell, A. and l(rrox, K.L. (L979) J. Anim. Sci. 49,
764-774.
Mitchell, H.H. (tS1Z¡ C-ornparat,ive Nutrition of Man and Domestic
Animals, Vol. I, Academic Press, New York.
Mir,chell, M.E. (tgle) Am. J. Clin. Nurr. 37, 293-306.
Morimoto, K. and Kanoh, H. (1978) J. BioI. CLrem. 253, 5056-5060.
l4udd, S.H. and Cantoni, G.L. (t964) In "C-omprehensive Biochemistry:
Group Transfer Reactions" (Florkin, M. and Stotz, E.H., eds.),
Vol. 15, pp. t-47, Elsevier Publishing Co., Arnsterdam.
Mrdd, S.H., Finkelstein, J.D., Irreverre, F. and l-asteî, L. (1965) J.
Biol. Clrem. 240, 4382-4392.
l"fudd, S.H., Levy H.L. and Abeles, R.H. (1969) Biochem. Biophys. Res.
C-onnrn:n. 35 , 72L-726.
Mrdd, S.H., Ievy, H.L. and Morrow, G. (1970) Biochem. Med. 4, L93-2L4.
l"lrdd, S.H. and Poole, J.R. (1975) Metabolism 24, 72L-735.
Mrdd, S.H. and levy, H.L. (1978) In "Ttre Metabolic Basis of Inherited
Disease" , 4tlrr edn. (Stanbury, J.8., t{yngaarden, J.B. and
Fredrickson, D.S., eds.), pp. 458-503, |deGraw-tlill, New York.
Mrdd, S.H. (1930) In "Sulfur in Biology", Ciba Found. Synrp. 72,
pp. 239-258, Excerpta Medica, New York.
Mrdd, S.H., Ebert, M.H. and Scriver, C.R. (1930) Metabolism 29, 707-720.
Mrdd, S.H., and Levy, H.L. (1983) In "Ttre Metabolic Basis of lriherited
Disease", 5th edn. (Stanbury, J.8., t$mgaarden, J.B., Fredrickson,
D.S. , Goldstein, J.L. and Brown, M.S. ; eds. ), PP. 522-559,
McGraw-Hill, New York.
D4nnr.z, J.A. (1950) J. Biol. Ckrem. L82, 489-499.
Nal<Lrooda, 4.F., t/eirC-N. and ]4arlissrE.B.(1980) Metabolism 29, L272-L277.

t79
National Research Council (L974) Nutrient Requirements of Dogs, National
Academy of Sciences, I^/ashington, D.C.
National Rese¿rch C-ouncil (1975) Nutrient Requirements of Sheep, 5th
edn., National Academy of Sciences, [.lashington, D.C.
National Research Council (1978) Nutrient. Requirements of Laboratory
Animals, 3rd rev. edn., National Academy of Sciences, washington,
D.C.
National Research Council (1919) Nutrient Requirements of Swine, 8th
rev. edn., National Academy of Sciences, I^Tashington, D.C.
Nayman, R., Thomson, M.E., Scriver, C.R. and CIow, C.L. (fgZg) Arn. J.
Clin. Nutr. 32, L279-L289.
Neill, 4.R., Grime, D.W., Snoswell, 4.M., NorthroP, 4.J., Lindsayr D.B.
and Dawson, R.M.C. (t979) Biochem. J. 180' 559-565.
Ngwira, T.N. and Beames, R.M. (L982) Nutr. Rep. Int- 25, 235-244-
Noble, R.C., Steele, I^1. and Moore, J.H. (tglt) Lipids 6, 926-929-
Noguchi, T., Okrno, E. and Kido, R. (ple) Biochem. J. 159, 607'6L3.
Nunn, J.F., Sharer, N.M., Bottiglieri, T. and Rossiter, J. (1986) Br.
J. Anaesth. 58, 1-10.
O'Dea, R.F. , Viveros, O.H. and Diliberto, E.J. Jr. (1981) Bioche¡n.
Pharmacol. 30, 1163-1168.
Oden, K.L. and Clarke, S. (fggg) Biochemistry 22, 2978-2986.
ogawa, H. and Fujioka, M. (tggZa) J. Biol. Cre¡n. 257, 3447-3/+52-
ogawa, H. and Fujioka, M. (1982b) Biochern. Biophys. Res. con.nmn. l-08,
227-232.
Ogawa, H., Ishiguro, Y. and Fujioh, M. (fgA:) Arch. Biochem. Biophys.
226, 265-275.
Qgino, H., Matsunmra, T., Satouchi, K. and saito, K. (rgao) Biochim.
Biophys. Acta 6L8' 43L-438.

180
Okada, G., Teraoka, H. and Tsukada, K. (fg8f) Bioche¡nisLry 20, 934-940.
oliva, A., Galletti, P. and Tappi-a, V. (1980) In "Natural Sulfur
C,ompounds: Novel Biochemical and Structural Aspect.s" (C,avallini,
D., Gaull, G.E. and Zappia, V., eds.), pp. 55-66, P1enum Press, New
York.
Orr, M.L. and Ilatt, B.K. (f0Sl¡ Amino Acid C-ontent of Foods, U.S. Dept..
of Agriculture Home Economics Research Report, No. 4, l^/ashington,
D.C., U.S. Government Printing Office.
Ørskov, E.R. (1982) Protein Nutrition in Ruminants, Academic Press,
I-ondon.
Patel-Thombre, U. and Borchardt, R.T. (1985) BiochemisLry 24, 1130-1136.
Pelech, S.L., Power, E. and Vance, D.E. (1983) Can. J. Bioche¡n. CeII
Biol. 6I,71-47-LL52.
Pisulewski, P.M. and Buttery, P.J. (fggS) Br. J. Nutr. 54, I2l-t29.
PoirÍ-er, L.4., Shivapurkar, N., Hyde, C.L. and Mikol, Y.B. (fgaZ) In
"The Biochemistry of S-Adenosylmethionine and Related Compounds"
(Usdin, E., Borchardt, R.T. and Creveling, C.R., eds.),
PP. 283-286, Ihcmillan, London.
Radcliffe, B.C. and Egan, A.R. (L974) Aust.. J. Biol. Sci. 27, 465-47I.
Radcliffe, B.C. and Egan, A.R. (L978) Aust. J. Biot. Sci. 3L, !05-LL4.
Rask, H., Olesen, 4.S., Mortensen, J.Z. and Freund, L.G. (fgA:) Scand.
J. tlaenratol. 31 , 45-48.
Rassin, D.K., Sturman, J.A. and Gaull, G.E. (1981) J. Neuroche¡n. 36,
\263-r27t.
Reed, D.J., Babson, J.R., Beatty, P.W., Brodie, 4.E., Ellis, W.l,l. and
Potter, D.lnl. (fgAO) AnaI. Biochem. L06, 55-62.
Rehbinder, D. and Greenberg, D.M. (fg0S) Arch. Biochem. Biophys. 109'
110-115.

181
Reis, P.J., Ttrnks, D.A. and Downes, A.M. (tglS) Aust. J. Biol. Sci.
26, 249-258.
Rowsell, E.V. (fgsf) Nature (r-on¿on) 168, 104-106.
Rowsell, E.V. (fgS6a) Biochem. J. 64, 246-252.
Rowsell, E.V. (1956b) Biochem. J. 64, 235-245.
Ryan, R.L. and McClure, W.O. (L919) Biochemistry 18, 5357-5365.
SaeLre, R. and Rabenstein, D.L. (fgZA) Anal. Biochem. 90, 684-692.
Salvatore, F., Utili; R. , Tappia, V. and Shapiro, S.K. (I97I) AnaI.
Biochem. 4L, t6-28.
Salvatore, F., Traboni, C., Colonna, A, Ciliberto, G., Paolella, G. and
Cimino, F. (1980) In "Natural Sulfur Cornpounds: Novel Biochemical
and Structural Aspects" (Cavallini, D., Gaull, G.E. and Zappi-a, V.,
eds.)r pp. 25-40, Plenum Press, New York.
Sanchez, A. and Swendseid, M.E. (1969) J. Nutr. 99, t45-L5I.
Santidrian, S., Sobrini, F.J., Bello, J. and larralde, J. (f98f¡ Enzyme
26,103-106.
Sastry, ts.V.R., Statharn, C.N., Axelrod, J. and Hirata, F. (fgaf) Arch.
Biochem. Biophys. 27L, 162-773.
Scarborough, G.A. and Nyc, J.F. (1967) J. Biol. Che¡n. 242, 238-242.
Schatz, R.4., I^/ilens, T.E. and Sellinger, O.Z. (fg8fa) J. Neurochem.
36, L739-L148.
Schatz, R.4., h7ilens, T.E. and Sellinger , O.Z. (fggfU) Biochem.
Bioflrys. Res. Comrmn. 98, L097-L|O7.
Schelling, G.T., Chandler, J.E. and Scott, G.C. (1973) J. Anim. Sci.
37, 1034-1039.
Schlenk, F. (1965) In "Transmethylation and Methionine Biosynthesis"
(Shapiro, S.K. and Schlenk, F., eds.), pp. 48-65., llniversity of
Gricago Press, Chicago.

L82
Schlenk, F. (1983) Adv. Enzynrol. 54, 195-265.
schneider, w.J. and vance, D.E. (rgza) Err. J. Biochem. 85, 181-187.
schneider, w.J. and vance, D.E. (L979) J. Biol. G:rem. 254, 3886-3891.
Scott, J.M., Dinn, J.J., trlilson, P., and Weir, D-G- (1981) lancet ii,
334-331.
Shapiro, S.K. and Yphantis, D.A. (1959) Biochim. Biophys. Acta 36,
24t-244.
Sheid, B. and Bilik, E. (1968) C,ancer Res. 28, 25L2-25L5.
Sidransky, H. and Farber, E. (1960) Arch. Biochem. Biophys . 87, t29-133.
silvernnn, M. and Pitney, A.J. (1958) J. BioI. Ctrem. 233, 1L79-a182.
Skacel, P.O., Hewlett, A.M. , Iewis, J.D. , Lumbr M. , Nunn, J.F. and
Cknnarin, I. (fgA:) Br. J. Flaenntol. 53, 189-200.
Skiba, W.E., Taylor, M.P., VJells M.S., Mangum, J.H. and Award, I{.M. Jr.
(1982) J. Biol. Ctrem. 251 , 14944-L4948.
Skurdal, D.N. and Cornatzer, W.E. (1975) Int. J. Biochem. 6' 579-583.
Srnith, R.M., Osborne-lJhite, L7.S. and Russell, G.R. (t967) Biochem. J.
L04, 44t-449.
Srnith, R.M., Osborne-l.r7hite, I,r/.S. and Gawthorne, J.M. (7974) Biochem. J.
142, L05-1L7.
Srnolin, L.4., Benevenga, N.J. and Berlow, S. (1981) J. Pediatr. 99,
467-472.
Snoswell, A.M¡ and Linzell, J.L. (tglS) J. Dairy Res. 42r 371-380.
Spector, R., Coakleyr G. and BIakeIy, R. (fggO) J. Neuroche¡n. 34,
t32-L37.
Sradtman, E.R. (1957) MeLhods Enzymol. 3, 931-94L.
Sreele, R.D. and Benevenga, N.J. (1973) J. Biol. Ctrem. 253, 7844-7850.
steele, R.D. and Benevenga, N.J. (1979) J. Biol. Chrem. 254, 8885-8890.
SLorm, E. and Brskov, E.R. (1984) Br. J. lfutr. 52, 6t3-620.

183
Strath, R.A. and Shelford, J.A. (fgZg) Can. J. Anim. Sci. 58, 479-494.
Sturman, J.4., Gaull, G.E. and Raiha, N.C.R. (1970) Science 169, 74-76.
Sturman, J.A. and Gaull, G.E. (fgZg) In "Advances in Polyamine Research"
(Ca¡np¡eff, R.A., Morris, D.R., Bartos, D., Daves, G.D. and Bartos,
F., eds.), Vol. 2, pp. 2L3-240, Raven Press, New York.
Sundler, R. and fu"""on, B. (1SZS¡ J. Biol. Cihem. 250, 3359-3367.
Swiatek, K.R., Simon, L.N. and Chao, K.-L. (L973) Biochemistry !2,
4670-4674.
Tallan, H.H. (f979) Biochem. Med. 2t, t29-L40.
Tanaka, Y., hi, O. and Akamatsu, Y. (tglg) Biochem. Biophys. Res.
C-ornrmn. 87, 1109-1115.
Taylor, R.T. and i,rleissbach, H. (fgeg) Arch. Biochem. Biophys. 729,
745-766.
Taylor, R.T. (fggZ) In "812" (oolphin, D., ed.), Vol. 2, pp.307-355,
JoLrr l{iley and Sons, New York.
Thithapandha, A. and C-ohrr, V.H. (fgZS) Biochem. Pharmacol . 27, 263-27L.
Thompson, InI., IkDonald, G. and Mookerjea, S. (1969) Biochim. Biophys.
Acta 776, 306-315.
Tsukada, K., Yamano, H., Ah, T. and Okadâ, G. (fggO) Biochem. Biophys.
Res . Connrmn. 95, LL6O-LL67 .
Tsukada, K., Abe, T., Kuwahata, T. and Mitsui, K. (1985) Life Sci. 37,
665-672.
T\rlloh, N.M. (1963) In "C,arc¿.ss C,onrposition and þpraisal of Meat
Animals" (Tribe, D.E., ed.), Paper No. 5, CSIRO, Australia.
T\rrakulov, Ya.lQ'r., SaaLov, T.S., Isaev, E.I. and Sadykov, S.S. (lglg)
Problemy Endokrinologgi (¡,losk) 25, 54-57.
Ueland, P.M., Helland, S., Broch, O.J. and Schanche, J.-S. (1984) J.
BioI. Chem. 259, 2360-2364.

184
Van der l,Jesthuyzen, J., Fernandes-Costa, F. and Metz, J. (IgB2) Life
Sci. 3t, 2O0L-20I0.
Van der l^lesthuyzen, J. and Met.z, J. (rga:) Br. J. Nutr. 50, 325-330.
Van der Westhuyzen, J., van Tonder, S.V., Gibson, J.E., Kilroe-Smith,
T.A. and Metz, J. (1995) Br. J. Nurr. 53, 657-662.
Van Pilsum, J.F., Stephens, G.C. and Taylor, D. (tglZ) Biochem. J.
t26, 325-345.
Vance, D.E. and de Kruijff, B. (1OAO¡ Nature (t-onaon) 2gg, 2ll-279.
Volpe, J.J. and I-asrer, L. (tglZ) Biol. Neonate 20, 385-403.
I^/alker, J.B. (tglg) Adv. Fnzynxrl. 50, t77-242.
I,rTeintrold, P.A. and Sanders, R. (tOlS¡ Life Sci. 13, 62l-629.
Ileissbach, H., Peterkofsky, 4., Redfield, B.G. and Dickerman, H. (rgo:)
J. Biol. Qrern. 238, 3318-3324.
I,lelsch, F., I{enger, w.c. and stedman, D.B. (rggr) pracenta 2, 2Lr-22L.
IiLrite, A. and Beach, E.F. (L937) J. Biol. Chem. LZZ, Zt9-226.
I^/ilken, D.R. (fSZO; Anal. Biochem. 36, 323-33I.
Wj lliams, J.N. Jr. (L952a) J. Biol. Crem. 194, L39-I42.
!ùilliams, J.N. Jr. (1952b) J. BioI. Chem. L95, 37-41.
I,Jilliams-Ashnran, H.G., seidenferd, J. and Galletti, p. (tggz) Biochem.
Pharmacol. 31, 217-288.
I,Jilliamson, D.H. (fgeO) FEBS I-erL . LL7, K93-105.
LTiIson, J., Corti,4., Flawkins, M., I,rlilliams-Ashman, H.G. and pegg, A.E.
(1979) Biochem. J. 180, 5L5-522.
I,/ilson, s.D. and Horne, D.w. (rga6) Arch. Biochem. Biophys. 244, 249-253.
I^lohlt, J.E., Kleyn, D.H., Vandernoot, G.W., Selfridge, D.J. and
Novorney, C.A. (fg8f) J. Dairy Sci. 64, 2Ll5-2L94.
I,lolff, J.E., Bergnan, E.N. and l^/illiams, H.H. (tslz¡ Am. J. physiol.
223, 438-446.

185
In/ong, E.R. and Thompson, [,,7. (1972) Biochim. Biophys. Acta 260, 259-27L.
Liong, T. (L97L) h"]. Biochem. 40, 78-28.
Yamashita, S., Oshinn, 4., Nikawa, J. and Hosaka, K. (tgAZ) Err. J.
Biochern. 128, 589-595.
Yanola;ra, M. and Tsukada, K. (fggZ) Biochem. Biophys. Res. Conrmrn.
to4, r464-L469.
Z.appía, V., Carteni-Farina, 1"1. and Porcelli, M. (L979) In
"Tbansmethylation" (Usdin, E., Borchardt, R.T. and Creveling,
C.R., eds.)r pp; 95-704, Elsevier/North HoIIand, New York.
Z,appia, V., Carteni--Farina, M., C-acciapuoti, G., Oliva, A. and
Gambacorta, A. (fggO) In "Natural Sulfur Compounds: Novel
Biochemical and Structural Aspects" (Cavallini, D., Gaull, G.E. and
Tappia, V., eds.)r pp. 133-148 , Plenum Press, New York.
Zeisel, S.H. and lfurtman, R.J. (L979) In "Transmethylation" (Usdin, E.,
Borchardt, R.T. and Creveling, C.R., eds.), pp.59-68,
Elsevier/North HoIIand, New York.
Zeisel, S.H. (1-981) Ann. Rev. Nutr. l, 95-I2I.
Zeisel, S.H. and lÌurtman R.J. (1981) Biochem. J. 198, 565-570.
Zlotkin, S.H. and Anderson, G.H. (1SAZ¡ Pediatr. Res. t6, 65-68.





























