Embed Size (px)
Citation preview

Available online at www.sciencedirect.com
www.elsevier.com/locate/tcm
T R E N D S I N C A R D I O V A S C U L A R M E D I C I N E ] ( 2 0 1 3 ) ] ] ] – ] ] ]
1050-1738/$ - see frohttp://dx.doi.org/10
$Disclosure: J.D.Vfailure. G.B. and J.C
nCorresponding autE-mail address:
Review article
GPCR biased ligands as novel heart failure therapeutics$
Jonathan D. Violina,n, David G. Soergela, Guido Boerrigterb, John C. Burnett Jr.b, andMichael W. Larka
aTrevena Inc, 1018 West 8th Ave, Suite A, King of Prussia, PA 19406bCardiorenal Research Laboratory, Mayo Clinic and Foundation, Rochester, MN
a r t i c l e i n f o
Article history:
Received 21 December 2012
Received in revised form
22 January 2013
Accepted 23 January 2013
nt matter & 2013 Elsevie.1016/j.tcm.2013.01.002
., D.G.S., and M.W.L. are.B. have performed sponhor. Tel.: þ[email protected]
A b s t r a c t
G protein-coupled receptors have been successfully targeted by numerous therapeutics
including drugs that have transformed the management of cardiovascular disease.
However, many GPCRs, when activated or blocked by drugs, elicit both beneficial and
adverse pharmacology. Recent work has demonstrated that in some cases, the salutary and
deleterious signals linked to a specific GPCR can be selectively targeted by ‘‘biased ligands’’
that entrain subsets of a receptor’s normal pharmacology. This review briefly summarizes
the advances and current state of the biased ligand field, focusing on an example: biased
ligands targeting the angiotensin II type 1 receptor. These compounds exhibit unique
pharmacology, distinct from classic agonists or antagonists, and one such molecule is now
in clinical development for the treatment of acute heart failure.
& 2013 Elsevier Inc. All rights reserved.
1. Introduction
Heart failure, the impairment of ventricular filling or ejection,
is characterized by insufficient cardiac output to meet meta-
bolic demand (Hunt et al., 2009). Although the etiology is
highly variable, this imbalance, together with the body’s
compensatory responses, represents a derangement of the
interplay of the cardiac, renal, and vascular systems. In the
absence of successful intervention, inadequate systolic or
diastolic cardiac function yields poor systemic blood flow,
leading to compensatory neurohormone release, vasocon-
striction, and fluid retention. This can support end organ
perfusion but increases work required by the heart to main-
tain already compromised cardiac output. In chronic
heart failure, disease progression is generally slow as the
r Inc. All rights reserved.
employees of Trevena Incsored research on behalfx231; fax: þ1-610-354-88
(J.D. Violin).
compensatory renal and vascular responses stabilize cardio-
vascular function. Perturbation of this compensated state,
such as by myocardial infarction, intercurrent illness, or
change in fluid or salt intake, can lead to acute decompen-
sated heart failure and rapid exacerbation of clinical status.
The hormonal systems regulating the interplay of the
cardiac, renal, and vascular systems rely heavily on G
protein-coupled receptors (GPCRs), a superfamily of cell sur-
face receptors responsible for cellular responses to myriad
extracellular stimuli. Numerous GPCR-targeted therapies
have been developed to support cardiac, renal, and vascular
function; this review will focus on recent insights that
suggest that recently discovered GPCR-mediated pathways
may afford new pharmacological benefits by targeting recep-
tors with a novel class of GPCR-targeted drug.
, which is developing TRV120027 for the treatment of acute heartof Trevena Inc.50.

T R E N D S I N C A R D I O V A S C U L A R M E D I C I N E ] ( 2 0 1 3 ) ] ] ] – ] ] ]2
2. GPCRs revisited: more than G proteincoupling
Two distinct GPCR systems play a central role in treating
chronic heart failure and are the targets of mainstay therapy:
the angiotensin receptors (particularly the type I isoform
AT1R) and the b-adrenergic receptors (both the b-1 and b-2
isoforms b1AR and b2AR). Angiotensin receptor blockers
(ARBs), such as losartan and valsartan, improve hemody-
namics in heart failure patients in three ways: (1) by directly
blocking angiotensin II’s vasoconstrictor and fibrotic effects;
(2) by inhibiting aldosterone synthesis and therefore sodium
retention by the kidney; and (3) enhancing renal perfusion
through effects on the afferent and efferent arteriole of the
glomerulus (Awan and Mason, 1996). Angiotensin-converting
enzyme (ACE) inhibitors achieve effects similar to ARBs by
inhibiting angiotensin II generation from angiotensin I via
ACE. However, ACE inhibitors do not fully block AT1R effects
as alternative pathways, such as chymase, can generate
angiotensin II even when ACE is fully inhibited (Hollenberg
et al., 1998; Weber, 1999). b Blockers such as carvedilol and
metoprolol are sympatholytic, and work largely by blocking
b1AR-mediated cardiac chronotropy and inotropy, thereby
reducing cardiac workload and metabolic demand (McBride
and White, 2005; Ruffolo et al., 1998). In conjunction with
loop diuretics to offload fluid, successful targeting of the
AT1R and BARs has led to markedly improved prognosis
associated with chronic heart failure (Jafri, 2004; Pitt, 2002).
Fig. 1 – G proteins and b-arrestins are key determinants of rece
leads to G protein coupling and changes in second messenger
phosphorylated by GRKs, the GPCR kinases (3), leading to b-arr
phosphorylated receptor, desensitizing the receptor by reducing
signaling enzymes such as MEK, leading to G protein-independe
receptor internalization to endosomes (5). For some receptors su
stimulate signal transduction from the endosome. From the en
degraded, resulting in receptor downregulation.
The significant clinical impact of drugs targeting the AT1R
and BARs and the proliferation of tools available to study
these receptors have positioned them at the forefront of basic
GPCR research. While initially envisaged to operate as simple
switches engaging intracellular responses by virtue of a
ligand–receptor–G protein complex (De Lean et al., 1980),
more recent work has demonstrated that the AT1R and BARs
do not signal via linear signal transduction pathways, but
rather engage networks of responses that in some cases are
independent of G protein coupling. In addition to G proteins,
both the AT1R and BAR when activated are phosphorylated
by GPCR kinases (GRKs) and bind b-arrestins, scaffold pro-
teins that translocate from the cytoplasm to bind activated
and phosphorylated receptor (Hunyady and Catt, 2006; Noor
et al., 2011; Shukla et al., 2011). For both receptors, b-arrestins
are multipurposed: (1) they inhibit receptor coupling to G
proteins, stimulating receptor desensitization; (2) they couple
receptors to endocytic machinery, stimulating receptor inter-
nalization to endocytic vesicles; (3) they engage G protein-
independent signaling pathways leading to cellular responses
distinct from those elicited by classical G protein pathways
(DeWire et al., 2007). Further work has shown that these
pathways are shared widely across the GPCR family; since
almost every GPCR couples to at least one class of G protein
and at least one isoform of b-arrestin, it is now thought that
the divergence of intracellular GPCR signals is a generic
phenomenon.
This mechanistic understanding took on practical impor-
tance with the realization that agonists and antagonists need
ptor pharmacology. Inactive receptor (1) engaged by agonist
signaling (2). Following this, agonist-occupied receptor is
estin recruitment to bind the agonist-occupied and
further G protein coupling (4). b-Arrestin scaffolds both
nt signal transduction, and endocytic machinery to promote
ch as the AT1R, b-arrestin remains bound and continues to
dosome, receptor is either recycled to the cell surface or

T R E N D S I N C A R D I O V A S C U L A R M E D I C I N E ] ( 2 0 1 3 ) ] ] ] – ] ] ] 3
not activate or block all of a receptor’s functions; ‘‘biased
ligands,’’ also known as ‘‘functionally selective’’ agonists, can
selectively engage a subset of a receptor’s normal signaling
repertoire (DeWire and Violin, 2011; Kenakin, 2012; Violin and
Lefkowitz, 2007), likely by stabilizing different receptor con-
formational states than are stabilized by endogenous
‘‘unbiased’’ ligands (Kenakin, 2007; Swaminath et al., 2004,
2005; Watson et al., 2000). Although bias can, in principle, be
found between any two receptor-dependent signals, G pro-
teins and b-arrestins are perfectly positioned, as ubiquitous
receptor-interacting proteins, to serve as signaling nodes to
determine the pharmacology of biased ligands (Fig. 1).
Fig. 2 – Molecular, cellular, and tissue effects of b-arrestin-
biased AT1R ligands in vivo or ex vivo. Open boxes list
biochemical signals described in response to b-arrestin-
biased AT1R ligands; filled boxes list cellular responses to
the same ligands. References are as follows: (1) Violin et al.,
2010; (2) Rajagopal et al., 2006; (3) Hunton et al., 2005; (4)
Xiao et al., 2010; (5) DeWire et al., 2008; (6) Aplin et al.,
2007a,b; (7) Kim et al., 2012; (8) Ahn et al., 2009.
3. A case study in ligand bias: the AT1R
The AT1R has been a particularly rich source of discoveries
describing the existence of biased ligands, the differentiation
of biased ligands compared to unbiased ligands, and the
utility of biased ligands for uniquely targeting unmet medical
need. Once considered to signal primarily by Gaq coupling to
the phospholipase C signal transduction pathway, it is now
clear that the AT1R engages G protein-independent signals as
well. The AT1R GRK/b-arrestin axis has been characterized in
a number of in vitro and in vivo settings, and has been shown
to engage a wide range of cellular responses. In addition,
several other pathways have been described that may be
independent of both G protein and b-arrestin/GRKs. These
pathways are reviewed elsewhere (DeWire et al., 2007;
Hunyady and Catt, 2006; Shukla et al., 2011) and select
pathways of interest are illustrated in Fig. 2.
The GRK/b-arrestin axis of AT1R signaling has taken on
particular significance with the discovery of biased ligands
that selectively engage this pathway, beginning with angio-
tensin II analogue 1Sar 4Ile 8Ile-AngII (SII) (Holloway et al.,
2002). This peptide was first shown to stimulate ERK activa-
tion, AT1R phosphorylation, and AT1R internalization, akin to
angiotensin II; however, unlike angiotensin II, SII failed to
stimulate any phosphoinositide turnover. Additional work
showed that SII was in fact a b-arrestin-biased ligand, as it
stimulates recruitment of b-arrestin2 to the AT1R, again
without engaging G protein-linked signals (Wei et al., 2003).
This work also showed that SII-stimulated ERK activation was
b-arrestin dependent, proving that b-arrestin-dependent sig-
nals can also be independent of G proteins. This finding,
along with the continued b-arrestin recruitment and signal-
ing to a mutant AT1R incapable of engaging G proteins,
demonstrated that b-arrestin is both necessary and sufficient
for some AT1R signals, confirming the status of b-arrestin
signaling as distinct from and parallel to G protein signaling.
This launched a number of investigations into b-arrestin
AT1R signaling engaged by SII. These studies showed that
SII stimulates anti-apoptotic signals, chemotaxis, cell growth
and proliferation, and cardiac contractility (Ahn et al., 2009;
Aplin et al., 2007a,b; DeWire et al., 2008; Hunton et al., 2005;
Rajagopal et al., 2006).
It is unclear what impact the reduction of b-arrestin-
dependent effects at the AT1R may have on the clinical
pharmacology of ARBs and ACE inhibitors. These agents
block the effects of the renin–angiotensin system (RAS),
which is activated in response to inadequate cardiac output.
This pathophysiological increased RAS activity elevates
angiotensin II levels, which in turn activates the AT1R,
leading to vasoconstriction, increased contractility, hyper-
trophy, and fibrosis of the heart, and increased sodium and
fluid retention at the kidney. Blocking these effects with ACE
inhibitors or ARBs improves mortality and morbidity in
chronic heart failure. Importantly, ARBs directly block both
G protein coupling and b-arrestin coupling of the AT1R (Violin
et al., 2010), and ACE inhibitors indirectly achieve the same
effect; although the net effect of this blockade is clearly
beneficial in chronic heart failure, it remains possible that
selective blockade by a biased AT1R ligand may deliver a
superior intervention in RAS-elevated states. In particular,
the direct stimulation of SII on cardiac contractility raised the
possibility that b-arrestin-biased AT1R ligands could support
cardiac function in acute decompensated heart failure via a
completely novel mechanism. SII is very weakly potent but
provided a lead for an optimization campaign that discovered
TRV120027, a 30-fold more potent b-arrestin-biased AT1R
ligand that has been studied in vitro and in vivo in rats and
dogs, and is now in Phase II clinical studies for the treatment
of acute heart failure (AHF). The preclinical data suggest that
TRV120027 has the potential to support heart, vasculature,
and kidney function in acute heart failure, interacting bene-
ficially with furosemide to alleviate symptoms and improve
long-term outcomes following hospitalization.
In the heart, TRV120027 acts as a mild inotrope, modestly
increasing the slope of the end-systolic pressure–volume

T R E N D S I N C A R D I O V A S C U L A R M E D I C I N E ] ( 2 0 1 3 ) ] ] ] – ] ] ]4
relationship (ESPVR) in normal mice and rats, and moderately
increasing cardiac output in normal and heart failure dogs
(Boerrigter et al., 2011; Violin et al., 2010). This was consistent
with the AT1R- and b-arrestin2-dependent inotropy seen in
isolated mouse cardiomyocytes and with a related b-arrestin-
biased ligand in mice (Kim et al., 2012; Rajagopal et al., 2006),
and contrasted with ARBs, which do not increase isolated
cardiomyocyte contractility, do not increase the slope of
ESPVR (Kim et al., 2012; Violin et al., 2010) and have not
shown increased cardiac output in dogs (Chan et al., 1992).
This suggests that in AHF patients, in whom RAS activity is
frequently elevated both on admission and by diuretic ther-
apy (Johnson et al., 2002), angiotensin II may contribute to
inotropic tone; whereas an ARB could block this inotropic
support in some patients, TRV120027 may preserve or even
increase cardiac contractility. Indeed, a study of the hemody-
namic effects of losartan showed that while cardiac output
increased with chronic therapy, acute treatment decreased
cardiac output (Crozier et al., 1995) which together with the
potential for sustained hypotension arising from the long
half-lives of ARBs and ACE inhibitors has largely prevented
their study and use in treating acute heart failure (Silvers
et al., 2007; Swedberg et al., 1992).
The mechanism responsible for the inotropic effect of b-
arrestin-biased AT1R ligands is unclear. SII increased frac-
tional shortening of isolated cardiomyocytes from wild-type
mice but not AT1R, b-arrestin2, or GRK6-knockout mice
(Rajagopal et al., 2006). Likewise, TRV120027 increased frac-
tional shortening of cardiomyoctes from wild-type but not
from b-arrestin2-knockout mice and the effect on wild-type
cardiomyocytes was blocked by pretreatment with the ARB
valsartan (Violin et al., 2010). Neither SII nor TRV120027
generated any detectable G protein-mediated effect in a
range of cell types, including calcium mobilization in cardi-
ovascular cells (Hunton et al., 2005; Kim et al., 2009; Rajagopal
et al., 2006). These findings demonstrate the necessity of the
AT1R GRK/b-arrestin axis for TRV120027 inotropy, and distin-
guish such ligands from classic inotropes such as dobut-
amine and milrinone which increase contractility by
mobilizing intracellular calcium, but do not elucidate the
signaling mechanism linking b-arrestins to contractile func-
tion. The wide array of kinase and phosphatase pathways
entrained by AT1R coupling to b-arrestins (Kendall et al.,
2011; Xiao et al., 2010) suggests the possible involvement of
sarcomeric protein phosphorylation, an established mecha-
nism to modify cardiomyocyte contractility (Solaro and
Kobayashi, 2011).
In addition to effects on contractility, emerging evidence
suggests that b-arrestins can exert anti-apoptotic signals via
the AT1R in the heart and that b-arrestin-biased ligands like
TRV120027 may protect against apoptosis and cell loss
generated by the increased metabolic stress of the decom-
pensated heart. Isolated hearts from WT mice subjected to
left ventricular mechanical stretch respond by activating
prosurvival signals to limit apoptosis; this is evident by
decreased ERK and Akt signaling, and increased apoptosis,
in stretched hearts from AT1R, b-arrestin2, or GRK-knockout
mice (Rakesh et al., 2010). Mechanistically, this response is
mediated by ligand-independent stretch activation of the
AT1R; both mechanical and osmotic stretch engaged
b-arrestin recruitment and b-arrestin-dependent prosur-
vival signaling in vitro (Rakesh et al., 2010). This b-arrestin
pathway is blocked by the ARB losartan, leading to increased
apoptosis following cardiac stretch. Consistent with this, a
b-arrestin-biased ligand enhanced Akt activation and
reduced apoptosis in a cardiac ischemia–reperfusion model
in WT but not b-arrestin2-knockout mice; in contrast the
ARB losartan had no effect in WT mice (Kim et al., 2012).
Together, these findings suggest that despite their estab-
lished benefits, ARBs may adversely affect beneficial com-
pensatory responses in the heart, and that b-arrestin-biased
ligands may be able to supplement a compensatory AT1R
prosurvival pathway in response to pressure overload,
potentially ameliorating myocyte loss and declining cardiac
performance.
In addition to contractility and apoptosis, the AT1R is
linked to adverse cardiac remodeling in heart failure. The
effects of biased ligands on cardiac remodeling have not been
tested directly, but ARBs directly inhibit pro-fibrotic signals.
This effect is likely through blockade of G protein signaling as
Gaq activation generates fibrosis and can induce heart failure
in transgenic mice (Fan et al., 2005; Wettschureck et al., 2001).
Consistent with this, transgenic mice expressing constitu-
tively active but b-arrestin-uncoupled AT1R display increased
cardiac fibrosis (Billet et al., 2007), and mice expressing a Gaq-
uncoupled AT1R develop less fibrosis than WT AT1R trans-
genic mice (Zhai et al., 2005). Thus it is likely that in the
setting of chronic therapy, b-arrestin-biased ligands like
TRV120027 would, like ARBs, reduce fibrosis and ameliorate
adverse cardiac remodeling.
At the level of vasculature, TRV120027 exerts all the
mechanistic benefits of ARBs. TRV120027 antagonizes angio-
tensin II-mediated hypertensive responses in normal rats
(Violin et al., 2010), and decreases both MAP and PCWP in
both normal dogs and tachypaced heart failure dogs
(Boerrigter et al., 2011). However, unlike ARBs and ACE
inhibitors which have long half-lives, TRV120027 is short-
lived, with a half-life of approximately 2 min in rats and dogs,
and its hemodynamic effects are rapidly reversible (Boerrigter
et al., 2011; Violin et al., 2010). This provides an important
safety feature for TRV120027, as sustained hypotension could
impede development of novel therapeutics for AHF. The
effect of TRV120027 on MAP is preserved in furosemide-
treated heart failure dogs, and the effect of TRV120027 on
pulmonary capillary wedge pressure is additive with the
effect of furosemide (Boerrigter et al., 2012). Since pulmonary
capillary wedge pressure is associated with dyspnea
(Teerlink, 2003), this suggests that TRV120027 will be benefi-
cial when co-administered with loop diuretics such as furo-
semide, which are first-line therapy for AHF.
TRV120027 also acts similarly to ARBs in the kidney. In
normal dogs, TRV120027 decreased both systemic and renal
vascular resistance, leading to increased renal blood flow
(Boerrigter et al., 2011). In these animals TRV120027 also
increased glomerular filtration rate (GFR), urine volume, and
urine sodium and potassium. These findings all echo the
effects of the ARB losartan in the same model (Chan et al.,
1992). As in normal dogs, TRV120027 increased renal blood
flow and urinary sodium and potassium excretion in heart
failure dogs.
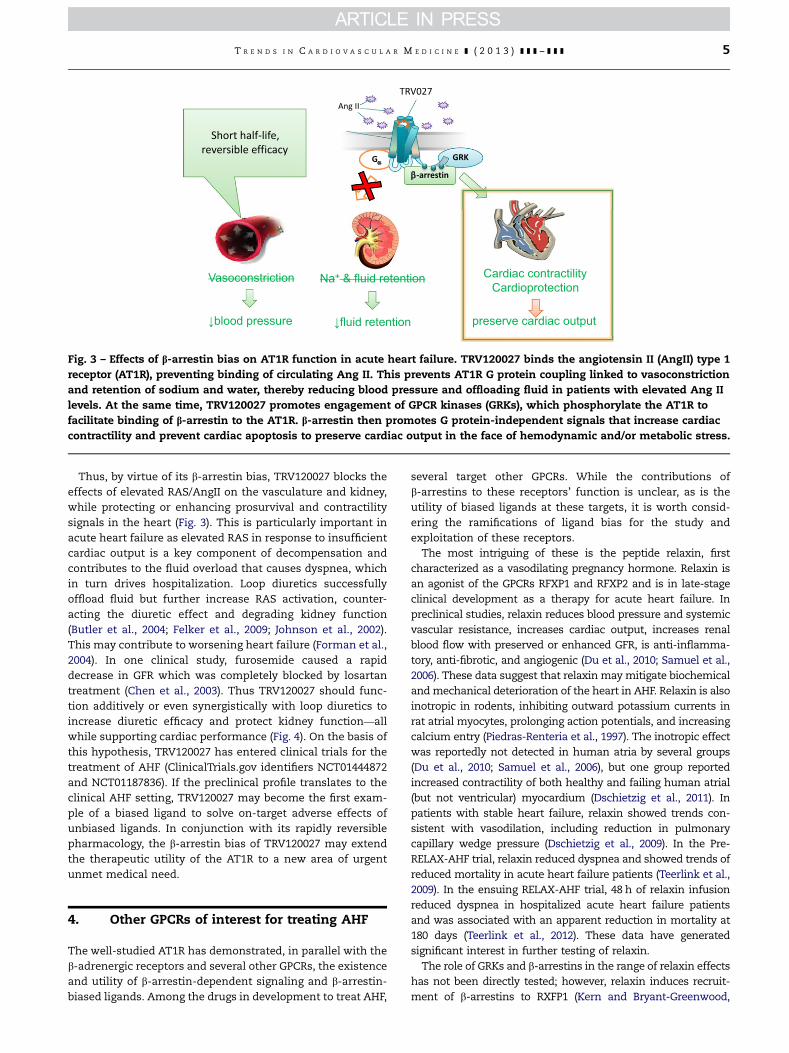
Fig. 3 – Effects of b-arrestin bias on AT1R function in acute heart failure. TRV120027 binds the angiotensin II (AngII) type 1
receptor (AT1R), preventing binding of circulating Ang II. This prevents AT1R G protein coupling linked to vasoconstriction
and retention of sodium and water, thereby reducing blood pressure and offloading fluid in patients with elevated Ang II
levels. At the same time, TRV120027 promotes engagement of GPCR kinases (GRKs), which phosphorylate the AT1R to
facilitate binding of b-arrestin to the AT1R. b-arrestin then promotes G protein-independent signals that increase cardiac
contractility and prevent cardiac apoptosis to preserve cardiac output in the face of hemodynamic and/or metabolic stress.
T R E N D S I N C A R D I O V A S C U L A R M E D I C I N E ] ( 2 0 1 3 ) ] ] ] – ] ] ] 5
Thus, by virtue of its b-arrestin bias, TRV120027 blocks the
effects of elevated RAS/AngII on the vasculature and kidney,
while protecting or enhancing prosurvival and contractility
signals in the heart (Fig. 3). This is particularly important in
acute heart failure as elevated RAS in response to insufficient
cardiac output is a key component of decompensation and
contributes to the fluid overload that causes dyspnea, which
in turn drives hospitalization. Loop diuretics successfully
offload fluid but further increase RAS activation, counter-
acting the diuretic effect and degrading kidney function
(Butler et al., 2004; Felker et al., 2009; Johnson et al., 2002).
This may contribute to worsening heart failure (Forman et al.,
2004). In one clinical study, furosemide caused a rapid
decrease in GFR which was completely blocked by losartan
treatment (Chen et al., 2003). Thus TRV120027 should func-
tion additively or even synergistically with loop diuretics to
increase diuretic efficacy and protect kidney function—all
while supporting cardiac performance (Fig. 4). On the basis of
this hypothesis, TRV120027 has entered clinical trials for the
treatment of AHF (ClinicalTrials.gov identifiers NCT01444872
and NCT01187836). If the preclinical profile translates to the
clinical AHF setting, TRV120027 may become the first exam-
ple of a biased ligand to solve on-target adverse effects of
unbiased ligands. In conjunction with its rapidly reversible
pharmacology, the b-arrestin bias of TRV120027 may extend
the therapeutic utility of the AT1R to a new area of urgent
unmet medical need.
4. Other GPCRs of interest for treating AHF
The well-studied AT1R has demonstrated, in parallel with the
b-adrenergic receptors and several other GPCRs, the existence
and utility of b-arrestin-dependent signaling and b-arrestin-
biased ligands. Among the drugs in development to treat AHF,
several target other GPCRs. While the contributions of
b-arrestins to these receptors’ function is unclear, as is the
utility of biased ligands at these targets, it is worth consid-
ering the ramifications of ligand bias for the study and
exploitation of these receptors.
The most intriguing of these is the peptide relaxin, first
characterized as a vasodilating pregnancy hormone. Relaxin is
an agonist of the GPCRs RFXP1 and RFXP2 and is in late-stage
clinical development as a therapy for acute heart failure. In
preclinical studies, relaxin reduces blood pressure and systemic
vascular resistance, increases cardiac output, increases renal
blood flow with preserved or enhanced GFR, is anti-inflamma-
tory, anti-fibrotic, and angiogenic (Du et al., 2010; Samuel et al.,
2006). These data suggest that relaxin may mitigate biochemical
and mechanical deterioration of the heart in AHF. Relaxin is also
inotropic in rodents, inhibiting outward potassium currents in
rat atrial myocytes, prolonging action potentials, and increasing
calcium entry (Piedras-Renteria et al., 1997). The inotropic effect
was reportedly not detected in human atria by several groups
(Du et al., 2010; Samuel et al., 2006), but one group reported
increased contractility of both healthy and failing human atrial
(but not ventricular) myocardium (Dschietzig et al., 2011). In
patients with stable heart failure, relaxin showed trends con-
sistent with vasodilation, including reduction in pulmonary
capillary wedge pressure (Dschietzig et al., 2009). In the Pre-
RELAX-AHF trial, relaxin reduced dyspnea and showed trends of
reduced mortality in acute heart failure patients (Teerlink et al.,
2009). In the ensuing RELAX-AHF trial, 48 h of relaxin infusion
reduced dyspnea in hospitalized acute heart failure patients
and was associated with an apparent reduction in mortality at
180 days (Teerlink et al., 2012). These data have generated
significant interest in further testing of relaxin.
The role of GRKs and b-arrestins in the range of relaxin effects
has not been directly tested; however, relaxin induces recruit-
ment of b-arrestins to RXFP1 (Kern and Bryant-Greenwood,

Fig. 4 – Clinical hypothesis for how a b-arrestin-biased ligand such as TRV120027 could intercede in RAS pathophysiology to
improve outcomes in AHF. (A) symptomatic HF patients presenting to the hospital are given loop diuretics such as
furosemide to offload fluid. This relieves shortness of breath (dyspnea) but also stimulates the renin–angiotensin system
(RAS), which is already activated by chronic pathology underlying acute heart failure. RAS activation increases circulating
angiotensin II (Ang II) levels, stimulating the AT1R to vasoconstrict and retain sodium and fluid, counteracting the effects of
furosemide on dyspnea and impairing renal function. (B) TRV120027 (‘‘027’’) interrupts the negative feedback of furosemide
actions while promoting cardiac performance, which may facilitate fluid offloading to shorten hospital stay and improving
outcomes.
T R E N D S I N C A R D I O V A S C U L A R M E D I C I N E ] ( 2 0 1 3 ) ] ] ] – ] ] ]6
2009), and b-arrestins participate in a pre-assembled signaling
complex containing several key signal transduction enzymes
(Halls and Cooper, 2010). Interestingly, this pre-assembled com-
plex is sensitive to extraordinarily low levels of relaxin, in stark
contrast to the high relaxin levels that are required for canonical
signaling form RXFP1. This differential sensitivity to drug
concentration may contribute to a puzzling feature of relaxin:
in preclinical and clinical studies relaxin pharmacology displays
biphasic ‘‘bell-shaped’’ receptor binding and dose–response
relationships, eliciting different effects at markedly different
doses (Du et al., 2010; Teichman et al., 2009). Thus, although
there is no clear hypothesis for how a biased ligand might differ
pharmacologically from relaxin, it is possible that selective
signaling could begin to precisely target the differential phar-
macology underpinning the complicated dose–response behav-
ior of relaxin.
Other GPCRs have been pursued as potential therapies for
acute heart failure, including the A1 adenosine, V2 vaso-
pressin, urotensin CRF2, and endothelin receptors
(Gheorghiade et al., 2005; Rademaker et al., 2011; Tamargo
et al., 2010). Each of these receptors engages b-arrestins, but
the contribution of the GRK/b-arrestin axis to therapeutic
hypotheses for these receptors is unclear. However, as the
field learns more about these targets, it will be important to
consider opportunities for ligand bias to deliver more precise
desired pharmacology. Broadly, biased ligands offer the
potential to (1) increase or extend efficacy in the form of
non-desensitizing or non-internalizing G protein-biased
ligands; (2) solve on-target adverse events by selectively
engaging therapeutic signal pathways; and (3) add novel
pharmacological benefits to a class of established unbiased
ligands. Alternatively, in some cases, unbiased agonists or
antagonists may be the optimal pharmacological therapy,
and ligand bias will be important as a means to help under-
stand variable pharmacology during lead optimization of
drug discovery.
5. Conclusion
The concept of ligand bias has evolved rapidly in the last
decade. Once deemed a theoretical consideration or an
idiosyncratic phenomenon of no clear importance, biased
ligands are now well-characterized at a range of receptors,
differentiated from unbiased ligands both in vitro and in vivo,
and, increasingly, viewed as viable therapeutic targets in the
effort to deliver safer and more efficacious pharmacotherapy.
In particular, non-canonical signal transduction pathways,
and biased ligands to target or avoid these pathways, should
be especially valuable to efforts targeting different pharma-
cology at distinct organ systems, as in acute heart failure.
r e f e r e n c e s
Ahn S, Kim J, Hara MR, Ren XR, Lefkowitz RJ. b-Arrestin-2mediates anti-apoptotic signaling through regulation of BADphosphorylation. Journal of Biological Chemistry 2009;284:8855–65.

T R E N D S I N C A R D I O V A S C U L A R M E D I C I N E ] ( 2 0 1 3 ) ] ] ] – ] ] ] 7
Aplin M, Christensen GL, Schneider M, Heydorn A, Gammeltoft S,Kjolbye AL, et al. The angiotensin type 1 receptor activatesextracellular signal-regulated kinases 1 and 2 by G protein-dependent and -independent pathways in cardiac myocytesand langendorff-perfused hearts. Basic & Clinical Pharmacol-ogy & Toxicology 2007a;100:289–95.
Aplin M, Christensen GL, Schneider M, Heydorn A, Gammeltoft S,Kjolbye AL, et al. Differential extracellular signal-regulatedkinases 1 and 2 activation by the angiotensin type 1 receptorsupports distinct phenotypes of cardiac myocytes. Basic &Clinical Pharmacology & Toxicology 2007b;100:296–301.
Awan NA, Mason DT. Direct selective blockade of the vascularangiotensin II receptors in therapy for hypertension andsevere congestive heart failure. American Heart Journal1996;131:177–85.
Billet S, Bardin S, Verp S, Baudrie V, Michaud A, Conchon S, et al.Gain-of-function mutant of angiotensin II receptor, type 1A,causes hypertension and cardiovascular fibrosis in mice.Journal of Clinical Investigations 2007;117:1914–25.
Boerrigter G, Lark MW, Whalen EJ, Soergel DG, Violin JD, Burnett Jr JC.Cardiorenal actions of TRV120027, a novel ss-arrestin-biasedligand at the angiotensin II type I receptor, in healthy and heartfailure canines: a novel therapeutic strategy for acute heartfailure. Circulation Heart Failure 2011;4:770–8.
Boerrigter G, Soergel DG, Violin JD, Lark MW, Burnett Jr JC.TRV120027, a novel beta-arrestin biased ligand at the angio-tensin II type I receptor, unloads the heart and maintainsrenal function when added to furosemide in experimentalheart failure. Circulation Heart Failure 2012.
Butler J, Forman DE, Abraham WT, Gottlieb SS, Loh E, Massie BM,et al. Relationship between heart failure treatment anddevelopment of worsening renal function among hospitalizedpatients. American Heart Journal 2004;147:331–8.
Chan DP, Sandok EK, Aarhus LL, Heublein DM, Burnett Jr JC.Renal-specific actions of angiotensin II receptor antagonismin the anesthetized dog. American Journal of Hypertension1992;5:354–60.
Chen HH, Redfield MM, Nordstrom LJ, Cataliotti A, Burnett Jr JC.Angiotensin II AT1 receptor antagonism prevents detrimentalrenal actions of acute diuretic therapy in human heart failure.American Journal of Physiology. Renal Physiology 2003;284:F1115–1119.
Crozier I, Ikram H, Awan N, Cleland J, Stephen N, Dickstein K,et al. Losartan in heart failure. Hemodynamic effects andtolerability. Losartan Hemodynamic Study Group. Circulation1995;91:691–7.
De Lean A, Stadel JM, Lefkowitz RJ. A ternary complex modelexplains the agonist-specific binding properties of the adeny-late cyclase-coupled beta-adrenergic receptor. Journal of Bio-logical Chemistry 1980;255:7108–17.
DeWire SM, Ahn S, Lefkowitz RJ, Shenoy SK. Beta-arrestinsand cell signaling. Annual Review of Physiology 2007;69:483–510.
DeWire SM, Kim J, Whalen EJ, Ahn S, Chen M, Lefkowitz RJ. Beta-arrestin-mediated signaling regulates protein synthesis. Jour-nal of Biological Chemistry 2008;283:10611–20.
DeWire SM, Violin JD. Biased ligands for better cardiovasculardrugs: dissecting G-protein-coupled receptor pharmacology.Circulation Research 2011;109:205–16.
Dschietzig T, Alexiou K, Kinkel HT, Baumann G, Matschke K,Stangl K. The positive inotropic effect of relaxin-2 in humanatrial myocardium is preserved in end-stage heart failure: roleof G(i)-phosphoinositide-3 kinase signaling. Journal of CardiacFailure 2011;17:158–66.
Dschietzig T, Teichman S, Unemori E, et al. Intravenous recombi-nant human relaxin in compensated heart failure: a safety,tolerability, and pharmacodynamic trial. Journal of CardiacFailure 2009;15:182–90.
Du XJ, Bathgate RA, Samuel CS, Dart AM, Summers RJ. Cardio-vascular effects of relaxin: from basic science to clinicaltherapy. National Reviews Cardiology 2010;7:48–58.
Fan G, Jiang YP, Lu Z, Martin DW, Kelly DJ, Zuckerman JM, et al. Atransgenic mouse model of heart failure using inducibleGalpha q. Journal of Biological Chemistry 2005;280:40337–46.
Felker GM, O’Connor CM, Braunwald E. Loop diuretics in acutedecompensated heart failure: necessary? Evil? A necessaryevil? Circulation Heart Failure 2009;2:56–62.
Forman DE, Butler J, Wang Y, Abraham WT, O’Connor CM,Gottlieb SS, et al. Incidence, predictors at admission, andimpact of worsening renal function among patients hospital-ized with heart failure. Journal of American College of Car-diology 2004;43:61–7.
Gheorghiade M, Teerlink JR, Mebazaa A. Pharmacology of newagents for acute heart failure syndromes. American Journal ofCardiology 2005;96:68G–73G.
Halls ML, Cooper DM. Sub-picomolar relaxin signalling by a pre-assembled RXFP1, AKAP79, AC2, beta-arrestin 2, PDE4D3complex. EMBO Journal 2010;29:2772–87.
Hollenberg NK, Fisher ND, Price DA. Pathways for angiotensin IIgeneration in intact human tissue: evidence from compara-tive pharmacological interruption of the renin system. Hyper-tension 1998;32:387–92.
Holloway AC, Qian H, Pipolo L, Ziogas J, Miura S, Karnik S, et al.Side-chain substitutions within angiotensin II reveal differentrequirements for signaling, internalization, and phosphoryla-tion of type 1A angiotensin receptors. Molecular Pharmacol-ogy 2002;61:768–77.
Hunt SA, Abraham WT, Chin MH, Feldman AM, Francis GS,Ganiats TG, et al. 2009 Focused update incorporated into theACC/AHA 2005 Guidelines for the Diagnosis and Managementof Heart Failure in Adults A Report of the American College ofCardiology Foundation/American Heart Association TaskForce on Practice Guidelines Developed in Collaboration Withthe International Society for Heart and Lung Transplantation.Journal of American College of Cardiology 2009;53:e1–90.
Hunton DL, Barnes WG, Kim J, Ren XR, Violin JD, Reiter E, et al.Beta-arrestin 2-dependent angiotensin II type 1A receptor-mediated pathway of chemotaxis. Molecular Pharmacology2005;67:1229–36.
Hunyady L, Catt KJ. Pleiotropic AT1 receptor signaling pathwaysmediating physiological and pathogenic actions of angioten-sin II. Molecular Endocrinology 2006;20:953–70.
Jafri SM. The effects of beta blockers on morbidity and mortalityin heart failure. Heart Failure Reviews 2004;9:115–21.
Johnson W, Omland T, Hall C, Lucas C, Myking OL, Collins C, et al.Neurohormonal activation rapidly decreases after intravenoustherapy with diuretics and vasodilators for class IV heartfailure. Journal of American College of Cardiology 2002;39:1623–9.
Kenakin T. Functional selectivity through protean and biasedagonism: who steers the ship? Molecular Pharmacology2007;72:1393–401.
Kenakin T. New concepts in pharmacological efficacy at 7TMreceptors. British Journal of Pharmacology 2012; 168(3): 554–75.
Kendall RT, Strungs EG, Rachidi SM, Lee MH, El-Shewy HM,Luttrell DK, et al. The beta-arrestin pathway-selective type1A angiotensin receptor (AT1A) agonist [Sar1,Ile4,Ile8]angio-tensin II regulates a robust G protein-independent signalingnetwork. Journal of Biological Chemistry 2011;286:19880–91.
Kern A, Bryant-Greenwood GD. Characterization of relaxin recep-tor (RXFP1) desensitization and internalization in primaryhuman decidual cells and RXFP1-transfected HEK293 cells.Endocrinology 2009;150:2419–28.
Kim J, Ahn S, Rajagopal K, Lefkowitz RJ. Independent beta-arrestin2 and Gq/protein kinase Czeta pathways for ERKstimulated by angiotensin type 1A receptors in vascular

T R E N D S I N C A R D I O V A S C U L A R M E D I C I N E ] ( 2 0 1 3 ) ] ] ] – ] ] ]8
smooth muscle cells converge on transactivation of theepidermal growth factor receptor. Journal of Biological Chem-istry 2009;284:11953–62.
Kim KS, Abraham D, Williams B, Violin JD, Mao L, Rockman HA.b-Arrestin-biased AT1R stimulation promotes cell survivalduring acute cardiac injury. American Journal of Physiology.Heart and Circulatory Physiology 2012;303:H1001–1010.
McBride BF, White CM. Critical differences among beta-adrenoreceptor antagonists in myocardial failure: debating theMERITof COMET. Journal of Clinical Pharmacology 2005;45:6–24.
Noor N, Patel CB, Rockman HA. Beta-arrestin: a signaling mole-cule and potential therapeutic target for heart failure. Journalof Molecular and Cellular Cardiology 2011;51:534–41.
Piedras-Renteria ES, Sherwood OD, Best PM. Effects of relaxin onrat atrial myocytes. II. Increased calcium influx derived fromaction potential prolongation. American Journal of Physiology1997;272:H1798–1803.
Pitt B. Clinical trials of angiotensin receptor blockers in heartfailure: what do we know and what will we learn? AmericanJournal of Hypertension 2002;15:22S–7S.
Rademaker MT, Charles CJ, Ellmers LJ, Lewis LK, Nicholls MG,Richards AM. Prolonged urocortin 2 administration in exper-imental heart failure: sustained hemodynamic, endocrine,and renal effects. Hypertension 2011;57:1136–44.
Rajagopal K, Whalen EJ, Violin JD, Stiber JA, Rosenberg PB,Premont RT, et al. Beta-arrestin2-mediated inotropic effectsof the angiotensin II type 1A receptor in isolated cardiacmyocytes. Proceedings of the National Academy of Sciencesof the United States of America 2006;103:16284–9.
Rakesh K, Yoo B, Kim IM, Salazar N, Kim KS, Rockman HA.b-Arrestin-biased agonism of the angiotensin receptorinduced by mechanical stress. Science Signaling 2010;3:ra46.
Ruffolo Jr. RR, Feuerstein GZ, Ohlstein EH. Recent observationswith beta-adrenoceptor blockade. Beneficial effects in hyper-tension and heart failure. American Journal of Hypertension1998;11:9S–14S.
Samuel CS, Du XJ, Bathgate RA, Summers RJ. ‘Relaxin’ thestiffened heart and arteries: the therapeutic potential forrelaxin in the treatment of cardiovascular disease. Pharma-cology & Therapeutics 2006;112:529–52.
Shukla AK, Xiao K, Lefkowitz RJ. Emerging paradigms of beta-arrestin-dependent seven transmembrane receptor signaling.Trends in Biochemical Sciences 2011;36:457–69.
Silvers SM, Howell JM, Kosowsky JM, Rokos IC, Jagoda AS. Clinicalpolicy: critical issues in the evaluation and management ofadult patients presenting to the emergency department withacute heart failure syndromes. Annals of Emergency Medicine2007;49:627–69.
Solaro RJ, Kobayashi T. Protein phosphorylation and signal trans-duction in cardiac thin filaments. Journal of Biological Chem-istry 2011;286:9935–40.
Swaminath G, Deupi X, Lee TW, Zhu W, Thian FS, Kobilka TS,et al. Probing the beta2 adrenoceptor binding site withcatechol reveals differences in binding and activation byagonists and partial agonists. Journal of Biological Chemistry2005;280:22165–71.
Swaminath G, Xiang Y, Lee TW, Steenhuis J, Parnot C, Kobilka BK.Sequential binding of agonists to the beta2 adrenoceptor.Kinetic evidence for intermediate conformational states. Jour-nal of Biological Chemistry 2004;279:686–91.
Swedberg K, Held P, Kjekshus J, Rasmussen K, Ryden L, Wedel H.Effects of the early administration of enalapril on mortality inpatients with acute myocardial infarction. New England Jour-nal of Medicine 1992;327:678–84.
Tamargo J, Amoros I, Barana A, Caballero R, Delpon E. Newinvestigational drugs for the management of acute heartfailure syndromes. Current Medicinal Chemistry 2010;17:363–90.
Teerlink JR. Dyspnea as an end point in clinical trials of therapiesfor acute decompensated heart failure. American HeartJournal 2003;145:S26–33.
Teerlink JR, Cotter G, Davison BA, Felker GM, Filippatos G, Green-berg BH, et al. 2012. Serelaxin, recombinant human relaxin-2,for treatment of acute heart failure (RELAX-AHF): a rando-mised, placebo-controlled trial. Lancet.
Teerlink JR, Metra M, Felker GM, Ponikowski P, Voors AA, Weath-erly BD, et al. Relaxin for the treatment of patients with acuteheart failure (Pre-RELAX-AHF): a multicentre, randomised,placebo-controlled, parallel-group, dose-finding phase IIbstudy. Lancet 2009;373:1429–39.
Teichman SL, Unemori E, Dschietzig T, Conrad K, Voors AA,Teerlink JR, et al. Relaxin, a pleiotropic vasodilator for thetreatment of heart failure. Heart Failure Reviews 2009;14:321–9.
Violin JD, DeWire SM, Yamashita D, Rominger DH, Nguyen L,Schiller K, et al. Selectively engaging beta-arrestins at theangiotensin II type 1 receptor reduces blood pressure andincreases cardiac performance. Journal of Pharmacology andExperimental Therapeutics 2010;335:572–9.
Violin JD, Lefkowitz RJ. Beta-arrestin-biased ligands at seven-transmembrane receptors. Trends in Pharmacological Scien-ces 2007;28:416–22.
Watson C, Chen G, Irving P, Way J, Chen WJ, Kenakin T. The use ofstimulus-biased assay systems to detect agonist-specificreceptor active states: implications for the trafficking ofreceptor stimulus by agonists. Molecular Pharmacology2000;58:1230–8.
Weber MA. Interrupting the renin-angiotensin system: the role ofangiotensin-converting enzyme inhibitors and angiotensin IIreceptor antagonists in the treatment of hypertension. Amer-ican Journal of Hypertension 1999;12:189S–94S.
Wei H, Ahn S, Shenoy SK, Karnik SS, Hunyady L, Luttrell L, et al.Independent beta-arrestin 2 and G protein-mediated pathwaysfor angiotensin II activation of extracellular signal-regulatedkinases 1 and 2. Proceedings of the National Academy ofSciences of the United States of America 2003;100:10782–7.
Wettschureck N, Rutten H, Zywietz A, Gehring D, Wilkie TM,Chen J, et al. Absence of pressure overload induced myocar-dial hypertrophy after conditional inactivation of Galphaq/Galpha11 in cardiomyocytes. Nature Medicine 2001;7:1236–40.
Xiao K, Sun J, Kim J, Rajagopal S, Zhai B, Villen J, et al. Globalphosphorylation analysis of beta-arrestin-mediated signalingdownstream of a seven transmembrane receptor (7TMR).Proceedings of the National Academy of Sciences of theUnited States of America 2010;107:15299–304.
Zhai P, Yamamoto M, Galeotti J, Liu J, Masurekar M, Thaisz J, et al.Cardiac-specific overexpression of AT1 receptor mutant lack-ing G alpha q/G alpha i coupling causes hypertrophy andbradycardia in transgenic mice. Journal of Clinical Investiga-tions 2005;115:3045–56.





























