Embed Size (px)
Citation preview

II mmunoglobulins are highlysoluble serum glycoproteinsinvolved in the defensemechanism of the immunesystem. IgGs are the major
components of the five differentclasses of immunoglobulins (namelyIgA, IgD, IgE, IgG, and IgM;Figure 1 provides a schematicrepresentation) (1). The serumconcentration of IgG is up to 13.5 mg/mL (Figure 1). The fourisotypes of IgGs are classified asIgG1, IgG2, IgG3 and IgG4 (Figure2). These IgG isotypes havedifferent numbers and arrangementsof interchain disulfide bonds (2). InIgG1 the disulfide bond linking thelight and heavy chains goes to thehinge region (Figure 2), whereas inthe IgG2, IgG3, and IgG4 subclassesthe disulfide bond linking the lightand heavy chains goes to thejunction between the variable andconstant regions.
Among the four isotypes, IgG1molecules exhibit predominanteffector functions (Table 1) (3). TheIgG2, IgG3, and IgG4 isotypescontain very minimal effectorfunctions. The N-linkedoligosaccharides present in theconstant CH2 domain of the Fchave greater influence on theeffector functions of IgG1 molecules(Figure 3 shows domain structure).These N-linked oligosaccharides ofIgGs are highly heterogeneous (alsoreferred to as microheterogeneity)and vary considerably with theexpression system (4).
Variations in conditions ofproduction systems greatly influencethe heterogeneity of IgGoligosaccharides (5). Thus,glycosylation of rIgGs varies from lot
to lot and among different cell-culture conditions. These variationsoften affect the biological activities oftherapeutic antibodies. This mightlead to potency changes in drugsubstances and drug products.Because of this, regulatory agenciesaround the world are very vigilantabout glycosylation variations oftherapeutic antibodies. This articleexamines the issues related toglycosylation variations with particularemphasis on expression systems andtheir impact on the effector functionsof therapeutic antibodies.
HUMAN IgG GLYCOSYLATION
N-Linked glycans of human IgGhave been extensively characterized(6). These glycans are mainlycomplex biantennary structures with
Glycosylation Variations with Expression Systemsand Their Impact on Biological Activity of Therapeutic ImmunoglobulinsT. Shantha Raju
PRODUCT FOCUS: ANTIBODIES
PROCESS FOCUS: CELL CULTURE,RECOVERY, ANALYSIS, RESEARCH
WHO SHOULD READ: SCIENTISTS,DIRECTORS, VPS, REGULATORS
KEYWORDS: IgG, GLYCOPROTEINS,OLIGOSACCHARIDES
LEVEL: BASIC TO ADVANCED
FFiigguurree 11:: Classes of immunoglobulins and their concentrations in serum
44 BioProcess International APRIL 2003

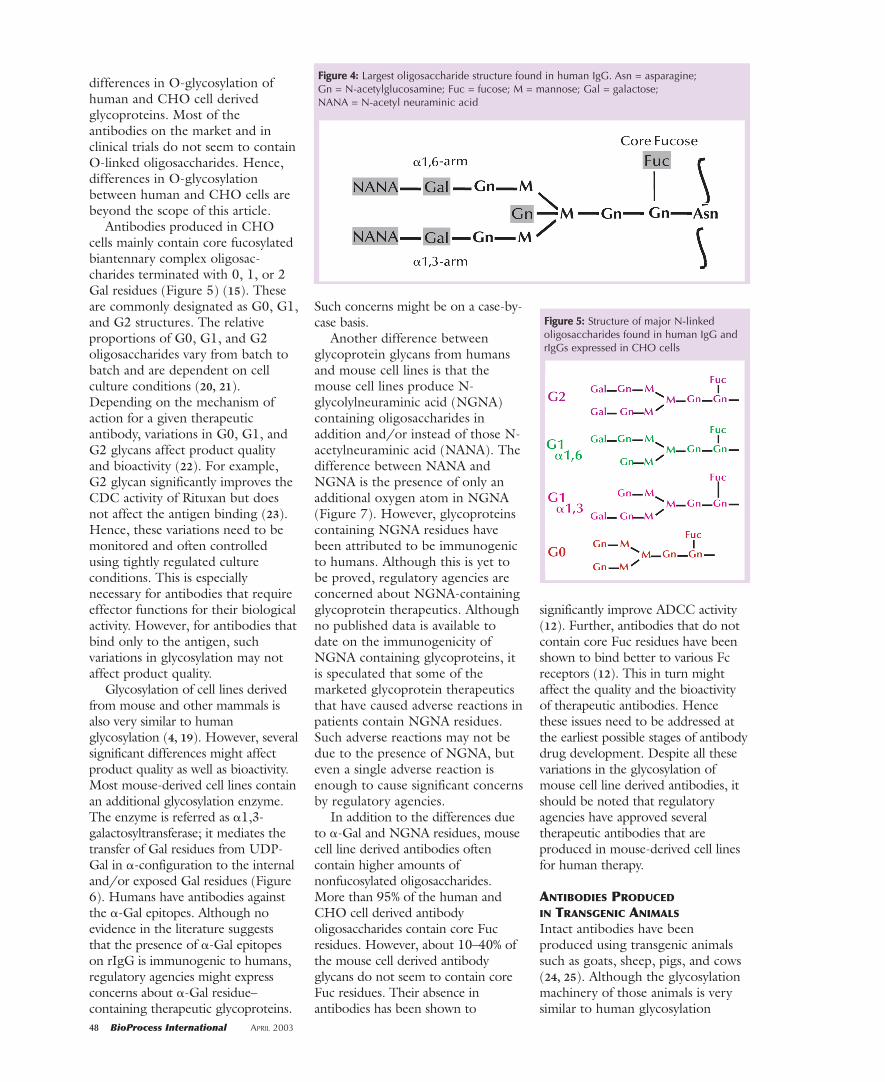
core fucose and often terminatedwith sialic acid residues. Figure 4shows the largest N-linkedoligosaccharide structure found inhuman IgG. This structure is presentas a very minor component. Themajority of oligosaccharides found inhuman IgG are truncated versions ofthe structure shown in Figure 4 dueto the microheterogeneity ofglycans. The microheterogeneity ofhuman IgG glycans is mainly due tothe presence and/or the absence ofeither core fucose, bisectingGlcNAc, terminal sialic acid, exposedGal, and often the exposed GlcNAcresidues (in either the �1,6- or �1,3-branch; Figures 4 and 5 show thestructures of human IgG glycans).Unlike other serum glycoproteins inwhich sialic acid residues are mainly�2,3-linked to penultimate Galresidues of N-glycans, in human IgGthe sialic acid residues are reportedto be mainly �2,6-linked to thepenultimate Gal residues of N-glycans (7).
The microheterogeneity ofhuman IgG glycans varies with ageand gender and is often indicative ofdisease status (8). For example, IgGs
from rheumatoid arthritis patientscontain more nongalactosylatedoligosaccharides (G0) than do thosefrom nonarthritic humans (9). Themicroheterogeneity of human IgGglycans reportedly affects biologicalfunctions such as complement-dependent cytotoxicity (CDC),antibody-dependent cytotoxicity(ADCC), binding to various Fcreceptors, and binding to C1qprotein (10). Further,nongalactosylated antibodies such asG0 glycoforms have been reportedto bind to mannose binding proteinpresent in serum and activate thealternative complement cascade(11). Also, terminal Gal, bisectingGlcNAc, and core fucose residuesaffect the effector functions oftherapeutic antibodies (12, 13).Hence, producing properlyglycosylated antibodies fortherapeutic purposes seems to bevery important.
RECOMBINANT IgG GLYCOSYLATION
About 11 rIgGs produced byrecombinant DNA technology arecurrently being marketed as humantherapeutics to treat life-threatening
diseases. Table 2 lists different IgGsand their indications. Additionally,more than 200 rIgGs are in variousphases of human clinical trials fordevelopment as human therapeutics.Biotechnology companies are usingdifferent methods of production tomake rIgG molecules. Most of thecurrently marketed rIgGs are beingproduced using in vitro cell culturemethods.
Because the demand forrecombinant antibodies far exceedsthe current in vitro cell cultureproduction capacity, alternativemethods to producing antibodies arebeing explored. These alternativemethods include producing theantibodies using transgenicmammals, plants, or avian eggs andalso amplifying the cell lines toimprove their yields. Severalantibodies have been successfullyproduced in transgenic goats, sheep,cows, and also in plants such as corn,alfalfa, and tobacco. Additionally,rIgGs have been successfullyproduced in avian eggs. For in vitrocell culture production, some celllines have been amplified and/or cellculture conditions manipulated toproduce up to 4–6g of antibody perliter of culture medium.
Although the integrity ofpolypeptide chains seems to belargely unchanged in the variousexpression systems and differentculture conditions, significantchanges in glycosylation have beennoticed. Glycosylation varies withcell line and animal species.Glycosylation of antibodies alsovaries with culture conditions (14).Hence, glycosylation of antibodiesexpressed under different cell
APRIL 2003 BioProcess International 45
FFiigguurree 22:: Schematic representation of human IgG isotypes
Table 1: Effector functions of human IgGisotypes
IgG1 IgG2 IgG4
Complement ++ + –
Fc�RI +++ – +Fc�RII +++ + +Fc�RIII ++ – –
Note: Receptors for Fc�RI, Fc�RII, and Fc�RIIIlocated on human monocytes, macrophages, andpolymorphonuclear cells

culture conditions will vary frombatch to batch. Because it variesfrom species to species, theglycosylation of antibodies producedin transgenic animals or plants or inavian eggs will be significantlydifferent from the glycosylation ofantibodies produced using in vitrocell culture processes (4–6, 15).Depending on the mechanism ofaction for each rIgG, these
variations will affect product qualityas well as product potency.
Regulatory agencies are payingclose attention to glycosylationvariations and their impact onproduct quality. It is thereforeadvisable to address these issues atthe early phases of development. Itis also recommended that theimpact of glycosylation on aparticular antibody product needs tobe addressed at the proof-of-
concept stage. Specific differences inglycosylation for different animalspecies and under varied cell cultureconditions are detailed below.
CHANGES DUE TO VARIATIONS
IN CELL CULTURE CONDITIONS
Many marketed recombinantantibodies are being produced eitherusing Chinese hamster ovary (CHO)cells or mouse-derived cell lines suchas NS0s or hybridomas as host cells.The glycosylation machinery of CHOcells is almost similar to the humanglycosylation machinery (16) – withseveral minor differences. Notableamong these differences is thatnormal CHO cells do not contain N-acetylglucosaminyltransferase-III(GnTIII), an enzyme that mediatesthe transfer of bisecting GlcNAc(Figure 4) to complex N-glycans(17). However, the amount ofbisecting GlcNAc-containingoligosaccharides in human IgG is<10% (4). Hence the absence ofbisecting GlcNAc-containingoligosaccharides in antibodiesproduced using CHO cells as hostsmay not significantly affect productquality and bioactivity.
Another difference betweenCHO cell derived IgG and humanIgG is in the nature of sialic acidlinkages. It has been reported thathuman IgG contain �2,6-linkedsialic acid residues whereas theCHO cell derived antibodiescontain �2,3-linked sialic acidresidues (18). However, the amountof sialylated oligosaccharides in bothhuman and CHO cell derivedantibodies is very small (<5% ofglycans contain sialic acid residues)(4, 19). So this difference in sialicacid linkages may not affect productquality. Moreover, the impact ofsialic acid residues on the antibodyfunction is not yet fully understood.Further, many of the human serumglycoproteins such as Fetuin and�1-acid glycoprotein contain �2,3-linked sialic acid residues. Becauseof this, regulatory agencies may notbe very concerned about the linkagedifference in sialic acid residuesbetween human and CHO cellderived antibodies. In addition tothese variations, there are several
46 BioProcess International APRIL 2003
FFiigguurree 33:: Domain structure of IgG molecule
TTaabbllee 22:: FDA-approved recombinant monoclonal antibodies in the market to treat human diseases
Type Product Marketer Approved
Murine Orthoclone OKT3 Johnson & Johnson June 1986allograft rejection
Chimeric ReoPro Lilly December 1994PTCA adjunct
Chimeric Rituxan Genentech November 1997non-Hodgkin’s lymphoma
Chimeric Simulect Novartis May 1998organ rejection prophylaxis
Chimeric Remicade Johnson & Johnson August 1998rheumatoid arthritis,Crohn’s disease
CDR-grafted Zenapax Roche December 1997organ rejection prophylaxis
CDR-grafted Synagis Medimmune June 1998respiratory syncytial virus (RSV)
CDR-grafted Herceptin Genentech September 1998metastatic breast cancer
CDR-grafted Mylotarg American Home May 2000acute myeloid leukemia Products
CDR-grafted Campath Millennium July 2001chronic lymphocytic leukemia
Human Humira Abbott Laboratories December 2002disease modifying antirheumatic drugs (DMARDs)
48 BioProcess International APRIL 2003
differences in O-glycosylation ofhuman and CHO cell derivedglycoproteins. Most of theantibodies on the market and inclinical trials do not seem to containO-linked oligosaccharides. Hence,differences in O-glycosylationbetween human and CHO cells arebeyond the scope of this article.
Antibodies produced in CHOcells mainly contain core fucosylatedbiantennary complex oligosac-charides terminated with 0, 1, or 2Gal residues (Figure 5) (15). Theseare commonly designated as G0, G1,and G2 structures. The relativeproportions of G0, G1, and G2oligosaccharides vary from batch tobatch and are dependent on cellculture conditions (20, 21).Depending on the mechanism ofaction for a given therapeuticantibody, variations in G0, G1, andG2 glycans affect product qualityand bioactivity (22). For example,G2 glycan significantly improves theCDC activity of Rituxan but doesnot affect the antigen binding (23).Hence, these variations need to bemonitored and often controlledusing tightly regulated cultureconditions. This is especiallynecessary for antibodies that requireeffector functions for their biologicalactivity. However, for antibodies thatbind only to the antigen, suchvariations in glycosylation may notaffect product quality.
Glycosylation of cell lines derivedfrom mouse and other mammals isalso very similar to humanglycosylation (4, 19). However, severalsignificant differences might affectproduct quality as well as bioactivity.Most mouse-derived cell lines containan additional glycosylation enzyme.The enzyme is referred as �1,3-galactosyltransferase; it mediates thetransfer of Gal residues from UDP-Gal in �-configuration to the internaland/or exposed Gal residues (Figure6). Humans have antibodies againstthe �-Gal epitopes. Although noevidence in the literature suggeststhat the presence of �-Gal epitopeson rIgG is immunogenic to humans,regulatory agencies might expressconcerns about �-Gal residue–containing therapeutic glycoproteins.
Such concerns might be on a case-by-case basis.
Another difference betweenglycoprotein glycans from humansand mouse cell lines is that themouse cell lines produce N-glycolylneuraminic acid (NGNA)containing oligosaccharides inaddition and/or instead of those N-acetylneuraminic acid (NANA). Thedifference between NANA andNGNA is the presence of only anadditional oxygen atom in NGNA(Figure 7). However, glycoproteinscontaining NGNA residues havebeen attributed to be immunogenicto humans. Although this is yet tobe proved, regulatory agencies areconcerned about NGNA-containingglycoprotein therapeutics. Althoughno published data is available todate on the immunogenicity ofNGNA containing glycoproteins, itis speculated that some of themarketed glycoprotein therapeuticsthat have caused adverse reactions inpatients contain NGNA residues.Such adverse reactions may not bedue to the presence of NGNA, buteven a single adverse reaction isenough to cause significant concernsby regulatory agencies.
In addition to the differences dueto �-Gal and NGNA residues, mousecell line derived antibodies oftencontain higher amounts ofnonfucosylated oligosaccharides.More than 95% of the human andCHO cell derived antibodyoligosaccharides contain core Fucresidues. However, about 10–40% ofthe mouse cell derived antibodyglycans do not seem to contain coreFuc residues. Their absence inantibodies has been shown to
significantly improve ADCC activity(12). Further, antibodies that do notcontain core Fuc residues have beenshown to bind better to various Fcreceptors (12). This in turn mightaffect the quality and the bioactivityof therapeutic antibodies. Hencethese issues need to be addressed atthe earliest possible stages of antibodydrug development. Despite all thesevariations in the glycosylation ofmouse cell line derived antibodies, itshould be noted that regulatoryagencies have approved severaltherapeutic antibodies that areproduced in mouse-derived cell linesfor human therapy.
ANTIBODIES PRODUCED
IN TRANSGENIC ANIMALS
Intact antibodies have beenproduced using transgenic animalssuch as goats, sheep, pigs, and cows(24, 25). Although the glycosylationmachinery of those animals is verysimilar to human glycosylation
FFiigguurree 44:: Largest oligosaccharide structure found in human IgG. Asn = asparagine; Gn = N-acetylglucosamine; Fuc = fucose; M = mannose; Gal = galactose; NANA = N-acetyl neuraminic acid
FFiigguurree 55:: Structure of major N-linkedoligosaccharides found in human IgG andrIgGs expressed in CHO cells

50 BioProcess International APRIL 2003
machinery, several significantdifferences do exist (4, 19, 26). Theseare very similar to the differencesdescribed above for the antibodiesderived from mouse cell lines.Currently, several glycoproteinsincluding antibodies that wereproduced in transgenic animals areundergoing human clinical trials.Genzyme Transgenics reported asuccessful completion of pivotalphase III trials of anti-thrombin-IIIthat was produced in transgenic goatmilk (27). However, the companydid not follow it through by filing abiological license application tomarket the product. Despite acertain amount of skepticism in theindustry, still, about the success oftransgenic technology, there areseveral significant benefits ofpursuing it to produce humantherapeutics. Notable among thesebenefits are potential savings in costof goods and services (COGS) andproduction of unlimited amounts ofprotein therapeutics using easilyavailable resources. Additionally, itshould be noted that no significantchanges have been observed in thepolypeptide chains of proteintherapeutics produced usingtransgenic animals other than thedifferences in glycosylation (28).
In addition, efforts are under wayto produce animals lacking the �-
galactosyltransferase gene. AlreadyVanhoue et al. reported that theintracellular expression in pig cellsof anti-�1,3galactosyltransferasesingle-chain FV antibodies reducesGal �1,3Gal expression and inhibitscytotoxicity mediated by anti-Galxenoantibodies (29). This type oftechnology development shouldadvance the field significantly. Suchadvanced transgenic technology isnot only useful to produce humantherapeutics; it is also useful forhuman tissue grafting and organtransplantation. Hence it is veryimportant to continue the researchin this area to advance thebiotechnology field.
ANTIBODIES PRODUCED
IN AVIAN EGGS
Therapeutic antibodies to treathuman diseases have been successfullyproduced in avian eggs (30).However, to date no publishedglycosylation analysis data areavailable in the literature about therecombinant antibodies produced inavian eggs. Some information isavailable in the literature about theglycosylation of IgG purified fromchicken serum (4, 19, 30, 31). Werecently reported that chicken IgGcontains a highly heterogeneousmixture of complex biantennaryoligosaccharides as well as highmannose structures (4). Figure 8shows a comparative analysis ofchicken IgG neutral glycans with
human IgG neutral glycans. Majorglycan structures of chicken IgG werefound to be Glc3Man9GlcNAc2 anda complex biantennary structurecontaining core Fuc, bisectingGlcNAc and terminated with Galresidues (see Figure 9 for structuresof high mannose glycans and Figure4 for the complex biantennarystructure). This observation suggeststhat the chicken IgG contains bothfully processed glycans (complexglycans) as well as unprocessedglycans (high mannose structures) inalmost equal amounts. Theglycosylation of rIgG produced inavian eggs might be very similar tothat observed in chicken IgG.
It should be noted here that nopotential immunogenic epitopessuch as �-Gal and NGNA residueswere observed in chicken IgG. Allthe sialylated oligosaccharides ofchicken IgG contained only NANA;no NGNA-containing oligosac-charides were seen. It has beenreported that the IgGs containinghigh mannose type oligosaccharideswere cleared faster than thosecontaining complex type oligosac-charides (32–34). Because chickenIgG contains significant amounts ofhigh mannose type oligosaccharides(~50% glycans), the pharmacokineticproperties of avian egg derivedrIgGs might be different from thoseof the rIgGs containing complextype oligosaccharides. This will bethe case if the avian-egg–derivedrIgGs contain glycans similar to thechicken IgG glycans. This mightpose some hurdles during clinicaltrials as well as during productapproval to market a drug. Hence,research studies should be continuedto understand the physicochemicaland biological properties of rIgGsproduced in avian eggs.
ANTIBODIES PRODUCED
IN TRANSGENIC PLANTS
Both glycosylated andnonglycosylated antibodies havebeen successfully produced intransgenic plants such as corn,alfalfa, and tobacco for therapeuticpurposes (35–41). Plant glycosylationis somewhat different from humanglycosylation, but the initial steps in
FFiigguurree 66:: Structure of G2 oligosaccharide containing �-Gal residues at the nonreducingtermini
FFiigguurree 77:: Structure of N-acetylneuraminicacid (NANA) and N-glycolylneuraminicacid (NGNA)
FFiigguurree 88:: MALDI-TOF-MS analysis ofneutral glycans of human and chicken IgGmolecules

APRIL 2003 BioProcess International 51
the biosynthetic pathway are verysimilar (38). The core N-glycanstructure that constitutes the twoGlcNAc residues and the three Manresidues is similar in both plants andanimals including humans (Figure10). Even the two GlcNAc residuesthat are �1,2-linked to both �1,3-Man and �1,6-Man are the same inplants and animals (Figure 4).Although plants add Gal residues tothe GlcNAc residues in theantennae, the Gal residues are �1,3-
linked in plants whereas they are�1,4-linked in animals (42). Inaddition many plants add a Xylresidues to the �1,4-Man residues atO-2 position in �-configuration.Further, plants add �1,3-Fuc to theGlcNAc residue, which in turn linksto the Asn residue of the polypeptidechain. In animal glycans, the coreFuc residue is �1,6-linked to thevery same GlcNAc (Figure 10).
Both �1,2-Xyl and �1,3-Fucresidues are reported to produce
IgE responses in humans. However,at this point, consequences of thepresence of �1,3-Gal residues islargely unknown. Because of thepresence of these potentialimmunogenic epitopes in transgenicplant derived antibodies (also calledplantibodies), nonglycosylatedantibodies have been produced intransgenic plants (35). Thenonglycosylated plantibodies aregenerated using site-specificmutations. Such plantibodies do not
FFiigguurree 99:: Structure of high-mannose type oligosaccharides found in chicken IgG

52 BioProcess International APRIL 2003
contain the conserved glycosylationsite in the Fc. Several glycosylatedand nonglycosylated plantibodieshave entered human clinical trials,and their results should be availablesoon.
Efforts are under way to obtainplantibodies with human typeglycosylation. A research group inHolland has produced plants thatgenerate plantibodies without �-Xyland �1,3-Fuc residues (36, 41–43).This was achieved using geneknockout technology. If thisresearch effort continues to showimprovements at the productionscale, it will be advantageous to thebiotechnology industry to producehuman therapeutics using transgenicplants. They could produce largeamounts of economically viablehuman protein therapeutics.
LONG-TERM PLANNING
Literature is replete with the role ofN-linked glycans on antibodyfunction. Considering the fact that somany therapeutic antibodies are indevelopment to treat human diseases,regulatory agencies and researchersare making additional efforts tounderstand the impact of glycans onantibody functions. Hence it isnecessary for biotechnologycompanies to allocate necessaryresources for studying the role ofglycans on their drug candidate/s.
Although many antibodies indevelopment and on the market arestructurally similar, their biophysicaland biochemical properties are quitedifferent, as is the impact of glycanson their functions. As long as theantibody drug is a full-lengthmolecule containing glycosylation inthe Fc, it is necessary to understandthe impact of glycans as much as
possible and at the earliest possiblestages. From the literature it is veryclear that antibody glycosylation isspecies-specific and varies from cellline to cell line. Within a particularcell line, antibody glycosylation variesfrom batch to batch and also withscale-up. So it is necessary to developproper analytical tools to detect thoseminor variations— because antibody-based drugs are here to stay.
REFERENCES1 Spiegelberg, HL; Dainer, PM. Fc
Receptors for IgG, IgM and IgE on HumanLeukaemic Lymphocytes. Clin. Exp.Immunol. 1979, 35(2): 286–295.
2 Jefferis, R. Structure-FunctionRelationships in Human Immunoglobulins.Neth. J. Med. 1991, 39(3–4): 188–198.
3 Jefferis, R. The Glycosylation ofAntibody Molecules: Functional Significance.Glycoconjugate J. 1993, 10(5): 358–361.
4 Raju, TS; et al. Species-SpecificVariation in Glycosylation of IgG: Evidencefor the Species-Specific Sialylation andBranch-Specific Galactosylation andImportance for Engineering RecombinantGlycoprotein Therapeutics. Glycobiology2000, 10(5): 477–486.
5 Parekh, RB; et al. N-Glycosylation andIn Vitro Enzymatic Activity of HumanRecombinant Tissue Plasminogen ActivatorExpressed in Chinese Hamster Ovary Cellsand a Murine Cell Line. Biochemistry 1989,28(19): 7670–7679.
6 Mizuochi T; et al. Structural andNumerical Variations of the CarbohydrateMoiety of Immunoglobulin G. J. Immunol.1982, 129(5): 2016–2020.
7 Jassal, R; et al. Sialylation of HumanIgG-Fc Carbohydrate By Transfected RatAlpha2,6-Sialyltransferase. Biochem. Biophys.Res. Commun. 2001, 286(2): 243–249.
8 Routier, FH; et al. Quantitation of theOligosaccharides of Human Serum IgG fromPatients with Rheumatoid Arthritis: A CriticalEvaluation of Different Methods. J.Immunol. Methods 1998, 213(2): 113–130.
9 Tsuchiya, N; et al. Distribution ofGlycosylation Abnormality Among SerumIgG Subclasses from Patients withRheumatoid Arthritis. Clin. Immunol.Immunopathol. 1994, 70(1): 47–50.
10 Wright, A; Morrison, SL. Effect ofGlycosylation on Antibody Function:Implications for Genetic Engineering. TrendsBiotechnol. 1997, 15(1): 26–32.
11 Malhotra, R; et al. GlycosylationChanges of IgG Associated with RheumatoidArthritis Can Activate Complement Via theMannose-Binding Protein. Nat. Med. 1995,1(3): 237–243.
12 Shields, RL; et al. Lack of Fucose onHuman IgG1 N-Linked OligosaccharideImproves Binding to Human Fcgamma RIII
and Antibody-dependent Cellular Toxicity. J.Biol. Chem. 2002, 277(30): 26733–26740.
13 Uman,a P; et al. EngineeredGlycoforms of an Antineuroblastoma IgG1with Optimized Antibody-DependentCellular Cytotoxic Activity. Nat. Biotechnol.1999, 17(2): 176–180.
14 Raju, TS; et al. Glycoengineering ofTherapeutic Glycoproteins: In VitroGalactosylation and Sialylation ofGlycoproteins with Terminal N-Acetylglucosamine and Galactose Residues.Biochemistry 2001, 40(30): 8868–8876.
15 Routier, FH; et al. The GlycosylationPattern of Humanized IgGI Antibody (D1.3)Expressed in CHO Cells. Glycoconjugate J.1997, 14(2): 201–207.
16 Stanley, P; Raju, TS; Bhaumik, M.CHO Cells Provide Access to Novel N-Glycans and Developmentally RegulatedGlycosyltransferases. Glycobiology 1996, 6(7):695–699.
17 Campbell, C; Stanley, P. A DominantMutation to Ricin Resistance in ChineseHamster Ovary Cells Induces UDP-GlcNAc:Glycopeptide Beta-4-N-Acetylglucosaminyltransferase III Activity. J.Biol. Chem. 1984, 259(21): 13370–13378.
18 Shah, P; et al. Physiological Influenceson Recombinant IgG Glycosylation.Biochem. Soc. Trans. 1998, 26(2): S114.
19 Hamako, J; et al. Comparative Studiesof Asparagine-Linked Sugar Chains ofImmunoglobulin G from Eleven MammalianSpecies. Comp Biochem Physiol. B 1993,106(4): 949–954.
20 Umana, P; Jean, M; Bailey, JE.Tetracycline-Regulated Overexpression ofGlycosyltransferases in Chinese HamsterOvary Cells. Biotechnol. Bioeng. 1999, 65(5):542–549.
21 Mimura, Y, ; et al. Butyrate IncreasesProduction of Human Chimeric IgG inCHO-K1 Cells Whilst Maintaining Functionand Glycoform Profile. J. Immunol. Methods2001, 247(1–2): 205–216.
22 Mimura, Y, ; et al. The MolecularSpecificity of IgG-Fc Interactions with FcGamma Receptors. Adv. Exp. Med. Biol.2001, 495: 49–53.
23 Davies, J, ; et al. Expression of GnTIIIin a Recombinant Anti-CD20 CHOProduction Cell Line: Expression ofAntibodies with Altered Glycoforms Leads toan Increase in ADCC Through HigherAffinity for FC Gamma RIII. Biotechnol.Bioeng. 2001, 74(4): 288–294.
24 Echelard, Y. Recombinant ProteinProduction in Transgenic Animals. Curr.Opin. Biotechnol. 1996, 7(5): 536–540.
25 Young, MW; et al. Production ofRecombinant Antibodies in the Milk ofTransgenic Animals. Res. Immunol. 149(6):609–610.
26 Pollock, DP; et al. Transgenic Milk AsA Method for the Production ofRecombinant Antibodies. J. Immunol.Methods 1999, 231(1–2): 147–157.
27 Yeung, PK. Technology Evaluation:
FFiigguurree 1100:: Comparison of the trimannosecore region structure in human and plantglycans

Transgenic Antithrombin III (rhAT-III),Genzyme Transgenics. Curr. Opin. Mol. Ther.2000, 2(3): 336–339.
28 Edmunds, T; et al. TransgenicallyProduced Human Antithrombin: Structuraland Functional Comparison to HumanPlasma-Derived Antithrombin. Blood 1998,91(12): 4561–4571.
29 Vanhove, B; et al. IntracellularExpression in Pig Cells of Anti-Alpha1,3galactosyltransferase Single-ChainFV Antibodies Reduces Gal alpha1,3GalExpression and Inhibits CytotoxicityMediated by Anti-Gal Xenoantibodies.Transplantation 1998, 66(11): 1477–1485.
30 Mohammed, SM; et al. Deposition ofGenetically Engineered Human Antibodiesinto the Egg Yolk of Hens.Immunotechnology 1998, 4(2): 115–125.
31 Morrison, SL; et al. Sequences inAntibody Molecules Important for Receptor-Mediated Transport into the Chicken EggYolk. Mol. Immunol. 2002, 38(8): 619–625.
32 Wright, A; et al. In Vivo Traffickingand Catabolism of IgG1 Antibodies with FcAssociated Carbohydrates of DifferingStructure. Glycobiology 2000, 10(12):1347–1355.
33 Rifai, A; et al. The N-GlycansDetermine the Differential Blood Clearanceand Hepatic Uptake of HumanImmunoglobulin (Ig)A1 and IgA2 Isotypes.J. Exp. Med. 2000, 191(12): 2171–2182.
34 Zuckier, LS; et al. Chimeric Human-Mouse IgG Antibodies with ShuffledConstant Region Exons Demonstrate ThatMultiple Domains Contribute to in VivoHalf-Life. Cancer Res. 1998, 58(17):3905–3908.
35 Ma, JK; et al. Assembly ofMonoclonal Antibodies with IgG1 and IgAHeavy Chain Domains in TransgenicTobacco Plants. Eur. J. Immunol. 1994,24(1): 131–138.
36 De, W; et al. Accumulation Pattern ofIgG antibodies and Fab Fragments inTransgenic Arabidopsis Thaliana Plants. PlantCell Physiol. 1998, 39(6): 639–646.
37 Ma, JK; et al. Characterization of aRecombinant Plant Monoclonal SecretoryAntibody and Preventive Immunotherapy inHumans. Nat. Med. 1998, 4(5): 601–606.
38 Cabanes, M; et al. N-Glycosylation ofa Mouse IgG Expressed in TransgenicTobacco Plants. Glycobiology 1999, 9(4):365–372.
39 Khoudi, H; et al. Production of aDiagnostic Monoclonal Antibody inPerennial Alfalfa Plants. Biotechnol. Bioeng.1999, 64(2): 135–143.
40 Sharp, JM; Doran, PM.Characterization of Monoclonal AntibodyFragments Produced By Plant Cells.Biotechnol. Bioeng. 2001, 73(5): 338–346.
41 Bouquin, T; et al.. Human Anti-
Rhesus D IgG1 Antibody Produced inTransgenic Plants. Transgenic Res. 2002,11(2): 115–122.
42 Bakker, H; et al. Galactose-ExtendedGlycans of Antibodies Produced ByTransgenic Plants. Proc. Natl. Acad. Sci. USA2001, 98(5): 2899–2904.
43 Edelbaum, O; et al. Expression ofActive Human interferon-Beta in TransgenicPlants. J. Interferon Res. 1992, 12(6):449–453. ��
T. Shantha Raju is a senior scientist andleader of the CMC (Chemistry,Manufacturing, and Controls) subteamfor an antibody development project atFibroGen, where he overseesdevelopment and CMC activities forIND and BLA submissions. FibroGenInc., 225 Gateway Blvd, South SanFrancisco, CA 94080; 650-866-6456;fax 650-866-7255; [email protected].








![Site-specific N-glycosylation regulates the GPS auto ... · EMR2- and CD97-mFc expression constructs have been described previously [8,25]. The chimeric and N-glycosylation site mutants](https://img.pdfslide.us/doc/110x75/5febcf30dec34c5d1c72ca55/site-specific-n-glycosylation-regulates-the-gps-auto-emr2-and-cd97-mfc-expression.jpg)




















