Embed Size (px)
Citation preview

iMedPub Journalshttp://www.imedpub.com
Translational BiomedicineISSN 2172-0479
2015Vol. 6 No. 2:17
1
Research Article
DOI: 10.21767/2172-0479.100017
© Under License of Creative Commons Attribution 3.0 License | This article is available in: www.transbiomedicine.com
Christopher T Turner1, Maria Fuller2, John J Hopwood3, Peter J Meikle4 andDoug A Brooks5
1 RegenerativeMedicine,MawsonInsti-tute,UniversityofSouthAustralia,Ad-elaide,Australia
2 GeneticsandMolecularPathology,SAPathologyatWomen’sandChildren’sHospital,NorthAdelaide,Australia
3 LysosomalDiseasesResearchUnit,SouthAustralianHealthandMedicalResearchInstitute,Adelaide,Australia
4 BakerIDIHeartandDiabetesInstitute,Melbourne,Australia
5 MechanismsinCellBiologyandDiseasesResearchGroup,SchoolofPharmacyandMedicalScience,DivisionofHealthSciences,UniversityofSouthAustralia,Adelaide,Australia
Corresponding author: DougABrooks
MechanismsinCellBiologyandDiseasesResearchGroup,SchoolofPharmacyandMedicalScience,DivisionofHealthSciences,UniversityofSouthAustralia,AdelaideSA5001,Australia.
Tel: +61-8-83021229
Citation:TurnerCT,FullerM,HopwoodJJ, etal.GlycogenExocytosisfromCulturedPompeSkinFibroblasts.TranslBiomed.2015,6:2.
BackgroundPompediseaseiscausedbyadeficiencyinthelysosomalenzymeα-glucosidase(GAA)andresultsintheprogressiveaccumulationof glycogen in autophagosome-autolysosome vesicles [1] andmultivesicularbodies [2] inaffectedcells.There iscurrentlynoeffective cure for this lysosomal storage disorder, but enzymereplacement therapy (ERT) has been clinically approved. ERThasbeenshowntoreducetheamountofglycogeninanumberofaffectedtissues, includingheartandskeletalmuscle[3],andthere is evidence of prolonged patient survival [4]. However,ithasbeenreported tobe ineffective insomepatients [3]andappears tohave limitedcapacity toaccess theglycogen storedin some major sites of pathology, including type II skeletalmuscle[1,5].ThisinabilityofGAAtoaccesstheglycogenstoredin certain autophagosomes-autolysosomes [6] necessitates the
development of novel or adjunct therapeutic options tomoreeffectivelytreatthedisease.
ThehallmarkofPompediseaseistheaccumulationofglycogenin autolysosomes, but a glucose tetrasaccharide (Glc4) hasbeenobserved inboth theurineandbloodofPompepatients[7]. This product of glycogen breakdown is presumed to arisefromdigestionby circulatingamylases [8] and tobe the resultof glycogen being released into circulation from affected cells.
Glycogen Exocytosis from Cultured Pompe Skin Fibroblasts
AbstractObjective: Pompe disease is a progressive form ofmuscular dystrophy causedby a deficiency in the lysosomal enzyme α-glucosidase (GAA), and leads tothe accumulation of glycogen in affected cells. Enzyme replacement therapy isapproved to treat infantile-onset Pompe disease, but this is not completelyeffective,necessitatingthedevelopmentofnewtherapeuticstrategies.Exocytosisinvolvesthefusionof intracellularvesicleswiththecellsurfaceandthereleaseofvesicularcontent,andisamechanismthatcouldbeusedinPompediseasetoremove stored glycogen fromaffected cells. The exocytosis of storagematerialfromPompepatientcellsintocirculationcouldresultinglycogendegradationbyotheramylases(i.e.notGAA)andthiscouldbedevelopedinthefutureasaneworadjuncttherapeuticstrategy.
Methods: AsensitivemassspectrometryassaywasusedtoquantifyglycogenincellextractsandtheculturemediafromconfluentPompeskinfibroblasts.
Results: Fourpercentofvesicularglycogenwasexocytosedafter2hoursinculture.Thisnaturalprocessofglycogenexocytosiswasenhancedinsub-confluentPompecells,whichreleased>80%ofglycogenafter2hoursinculture.
Conclusion: Under appropriate conditions exocytosis can release most of thestored glycogen in Pompe skin fibroblasts, identifying a potential target fortherapeuticintervention.
Keywords: Pompe; Lysosomal storagedisorder;Glycogen; Exocytosis; Substratedegradation
Received: June16,2015; Accepted: August13,2015; Published: August17,2015

2
ARCHIVOS DE MEDICINAISSN 1698-9465
2015Vol. 6 No. 2:17
Translational BiomedicineISSN 2172-0479
This article is available in: www.transbiomedicine.com
One possible explanation for this extracellular glycogen is celldeath,andthereissomeevidenceforapoptosisintheadvancedstagesofPompedisease[9].However,theamountofcirculatingtetrasaccharideinPompepatientsappearstobesimilarinbothearly- and late-stage disease [10], suggesting that cell death isatbestonlyapartialexplanationforthisglycogenreleasefromPompecells.Analternativeexplanationisthereleaseofstoredglycogen frommuscle and other cells by exocytosis, a processthat involves the fusion of intracellular vesicles with the cellsurfaceandthereleaseofvesicularcontent.
There is evidence that storage material can be released fromlysosomal storage disorder cells by exocytosis. In kidney cellsderived from metachromatic leukodystrophy patients, sulphatidestoragematerialisreleasedintotheculturemedium[11].Exocytosiswas induced in Niemenn Pick type C [12], multiple sulphatasedeficiency, mucopolysaccharidosis (MPS) type-IIIA and neuronalceroidlipofuscinosescells[13],andresultedinareducedamountofintracellularstoragematerial.Recently,theinductionofexocytosisinculturedPompecellswasshowntocauseareductioninintracellularglycogen storage [13-15], suggesting that glycogen exocytosis ispossible. Lysosomal exocytosis was induced by overexpressingtranscription factor EB, which activated lysosomal Ca2+ channelMCOLN1 and increased the intracellular Ca2+ concentration. Themodulationofexocytosis,byspecificmediaandculturingconditions[16,17] as well as a number of drugs/compounds targeting theexocytic machinery [18,19], could also be utilized to modulateglycogen release. Here, a mass spectrometry based glycogenquantification assay [20] was used to accurately define glycogenexocytosis from Pompe skin fibroblasts under culture conditionsthat induceCa2+-dependent exocytosis, confirming theworkofMedinaetal.[13],andCa2+-independentexocytosis,identifyingalternative targets with the potential to release a higherproportionofvesicularglycogen.Wealsodefinedtheamountofexocytosisandglycogenreleasedrelativetoothervesicularcargoand compared this process in fibroblasts from control, Pompedisease and MPS type I, a lysosomal storage disorder whichaccumulates different storage substrates. This led to the novelfindingthatalthoughincreasedexocytosisinPompecellsledtoelevatedglycogenrelease,itwaspartiallyimpaired.
Materials and MethodsCell cultureDe-identified Pompe and MPS I skin fibroblasts were isolatedfrom skinbiopsies referred to theNationalReferral Laboratoryfor Lysosomal, Peroxisomal and Related Genetic Disorders(Women’s and Children’s Hospital, Adelaide, Australia) andused in accordance with Women’s and Children’s HumanResearch Ethics Committee approval 668/4/2009. Unaffectedskinfibroblastswerederivedfromskinbiopsiesfromapparentlyhealthyvolunteers.Eachoftheculturedfibroblastlinesusedhadasimilar rateofgrowthandwas limited to less thanninesub-cultures.
Cytoplasmic glycogen depletionTodeplete theskinfibroblastculturesofcytoplasmicglycogen,cellswereculturedinFBS-andglucose-freeculturemedium.For
depletion,cellswerethenwashedtwicewith12mLofPBS,pH7.2 (Sigma,St.Louis,USA),andthencultured inFBS-freeBasalEaglemedium(Sigma,St.Louis,USA)for24hoursat37°C.Afterdiscardingthisculturemedium,thecellmonolayerwaswashedwith 12 mL of PBS (three times) and cultured in glucose-freeDulbecco’smodifiedEagle’smedium(Sigma,St. Louis,USA) for24hoursat37°C.
Trypan blue cell viability measurementToevaluatetheviabilityofculturedcellsatharvest,a20μLaliquotofcellsuspensionwasmixedwithanequalvolumeof0.1%(v/v)trypanblueand incubated for5minsat20°C, transferred toahaemocytometer and examined at 100X magnification. Non-viablecellswerestainedblueduetouptakeoftrypanblueintothecell.Cultureviabilitywasevaluatedasthepercentageoftotalcellsthatdidnotstainblue.Datawerenotcollectedfromcontrolfibroblastcultureswith<90%trypanblueexclusion.
Lactate dehydrogenase cell viability measurementA 300 μL aliquot of glucose-free DMEM from cultured cellswas mixed with 200 μL of lactate dehydrogenase (LDH) assaysubstrate,200μLofLDHcofactorand200μLofLDHdyesolution,and incubated for 30mins at 20°C in the dark, in accordancewiththeTOX-7kitinstructions.Eachreactionwasstoppedbytheadditionof90μLof1NHClandanalysedspectrophotometricallyat both 690 nm (background signal) and 490 nm.A sample ofglucose-free DMEM was included as a negative control. Forthepositive control, 10mLof LDHassay lysis solution (diluted1:10 in glucose-free DMEM) was added to a flask of cells torelease cellular LDH. All assays, including the positive control,wereperformedintriplicate.TheamountofLDHintheculturemediumwascorrectedfortotalcellproteinandexpressedasthepercentageofLDHreleasedperculture.Datawerenotcollectedfromcontrolfibroblastcultureswith>5μg/mgoftotalcellproteinLDHrelease.
Cell surface immune-fluorescenceSkin fibroblasts were seeded onto sterile coverslips in 6-wellplates at approximately 1×104 cells/mL (eachwell containing 2mLofcompleteculturemedium).Cellswereculturedtoeither20%to50%confluence(3.4to8.5×104cells/well)orconfluence(1.7×105cells/well).Eachwellwaswashedthreetimeswith4mLofPBS for5minutesat4°Conaplateshaker.OnehundredμLof mouse α-Lysosomal-associated membrane protein (LAMP)-1monoclonalantibodycloneBB6wasthenaddedtoeachwell(dilutedto2.2μg/mLincompleteculturemedium)andincubatedfor 1hour at 4°C.Wellswere thenwashed threetimeswith4mLofPBSfor5minutesat4°Conaplateshaker.Eachwellwasthenaspiratedand100μLofFluorophore-488conjugatedgoatα-mouseantibody(Invitrogen,Carlsbad,USA;dilutedto1:1000in complete culturemedium)was added and incubated in thedarkfor1hourat4°C.Allwellswerewashedthreetimeswith3mLofPBSfor5minutesat4°Conaplateshaker,inthedark.Tostainthenucleus,50µLofProlongGoldnuclearstain(Invitrogen,Carlsbad,USA),containingDAPI,wasaddedtoeachcoverslip.Thecoverslipwastheninvertedontoamicroscopeslide.Coverslips/
3
ARCHIVOS DE MEDICINAISSN 1698-9465
2015Vol. 6 No. 2:17
Translational BiomedicineISSN 2172-0479
© Under License of Creative Commons Attribution 3.0 License
cellswerestoredinthedarkat4°CuntilexaminedonaLeicaSP5spectral scanning confocalmicroscope (LeicaMicrosystems PtyLtd.,NorthRyde,Australia)at100Xmagnification.FluorescenceintensityperunitareawasdeterminedusingAnalySISsoftware(SoftImagingSystemGmbH,Munster,Germany).
Intracellular immune-fluorescenceTovisualisetheintracellularlocationofLAMP-1inskinfibroblasts,cellswerepermeabilisedand stainedby immune-fluorescence.To fix and permeabilise the cells, 1 mL of methanol/acetone(1:1)wasaddedtoeachwell(coverslip)andthenincubatedfor10minutesat–20○C.Eachwellwasthenaspiratedandair-driedfor20minutesat20○C.Topreventnon-specificantibodybinding,1mL of PBS containing 5% (w/v) bovine serum albumin (BSA;Sigma,St.Louis,USA)wasaddedtoeachwellandincubatedfor1hourat20○Conaplateshaker.Eachwellwasaspiratedand100μLofmonoclonalLAMP-1antibody(dilutedto2.2μg/mLin5%(w/v)BSAinPBS)wasaddedandincubatedfor1hourat20○C.Eachwellwaswashedthreetimeswith4mLofPBSfor5minutesat 4○C on a plate shaker. One hundred μL of Fluorophore-488conjugatedgoatα-mousesecondaryantibody(dilutedto1:200in5%(w/v)BSA inPBS)wasaddedtoeachwelland incubatedinthedarkfor1hourat20○C.Allwellswerewashedthreetimeswith4mLofPBSfor5minutesat20○Conaplateshaker,inthedark.To stain thenucleus,50µLofProlongGoldnuclear stainwas added to each coverslip; the coverslip was then invertedontoamicroscopeslide.Coverslips/cellswerestoredinthedarkat 20○CuntilexaminedonaLeicaSP5spectralscanningconfocalmicroscopeat100Xmagnification.
Glycogen quantificationThemassspectrometrybasedglycogenassayhasbeendescribedpreviously for the quantification of glycogen in mouse tissueextracts [20].Here, theassaywasapplied to thequantificationofglycogeninculturemedium(100µLanalysedpersample)andskinfibroblastextracts(0.1µgcellproteinmadeupto10µLinwateranalysedpersample).
β-Hexosaminidase activityTheβ-Hexosaminidasefluorometricassaywasusedasdescribedpreviously[21].
Evaluation of cell division in skin fibroblastsApproximately 1×106 cells were resuspended in 220 μL ofpropidiumiodidesolution,containing0.5%(v/v)ofTriton-X100(Sigma,St.Louis,USA),250µg/mLpropidiumiodide(Sigma,St.Louis,USA)and250µg/mLofRNaseI(Sigma,St.Louis,USA)inPBS,andincubatedinthedarkfor30minutesat20○C.Differentstages of the cell cycle were distinguished by the relativeproportion of propidium iodide incorporated into the nucleusof each cell as described previously by Givan [22]. To removeexcesspropidiumiodidesolution,cellswerewashedwith1mLofPBSfor1minuteonaplateshakerbeforefinalre-suspensionin1mLofPBS.CellsweretheninjectedintoaFACScaliburflowcytometer(BDBiosciences,FranklinLakes,USA)andtheamountof propidium iodide intercalated into each cell was evaluated
usingCellQuestsoftware (BDBiosciences,FranklinLakes,USA);20,000cellswerecountedforeachculture.
Statistical analysisDifferences between two independent groups of data with anormaldistributionweredeterminedbythestudentT-test,andthesignificancedefinedbyaPvalueof<0.05.Tocomparethreeormoregroupsofdataanalysisofvariance(ANOVA)wasutilised,with P-values adjusted by the Holm's Stepdown Bonferroniprocedure.
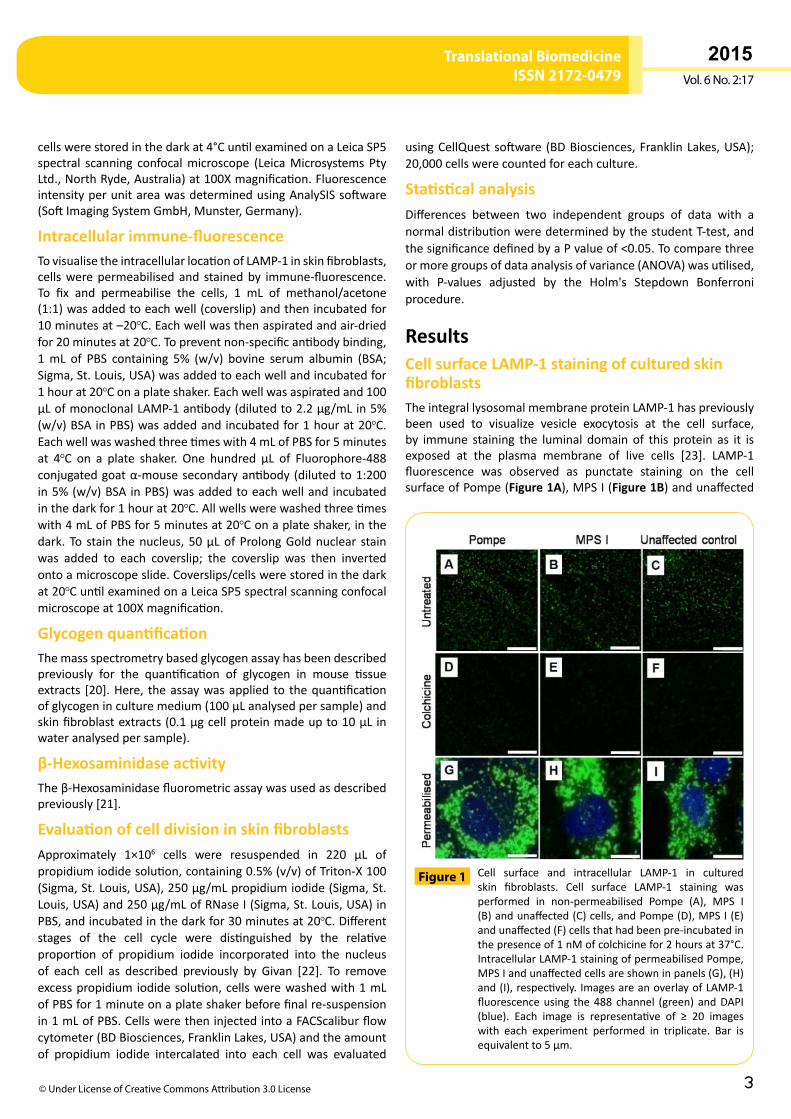
ResultsCell surface LAMP-1 staining of cultured skin fibroblastsTheintegrallysosomalmembraneproteinLAMP-1haspreviouslybeen used to visualize vesicle exocytosis at the cell surface,by immune staining the luminal domain of this protein as it isexposed at the plasma membrane of live cells [23]. LAMP-1fluorescence was observed as punctate staining on the cellsurfaceofPompe(Figure 1A),MPSI(Figure 1B)andunaffected
Cell surface and intracellular LAMP-1 in culturedskin fibroblasts. Cell surface LAMP-1 staining wasperformed in non-permeabilised Pompe (A), MPS I(B)andunaffected(C)cells,andPompe(D),MPSI(E)andunaffected(F)cellsthathadbeenpre-incubatedinthepresenceof1nMofcolchicinefor2hoursat37°C.IntracellularLAMP-1stainingofpermeabilisedPompe,MPSIandunaffectedcellsareshowninpanels(G),(H)and(I),respectively.ImagesareanoverlayofLAMP-1fluorescenceusing the488 channel (green)andDAPI(blue). Each image is representative of ≥ 20 imageswith each experiment performed in triplicate. Bar isequivalentto5µm.
Figure 1
4
ARCHIVOS DE MEDICINAISSN 1698-9465
2015Vol. 6 No. 2:17
Translational BiomedicineISSN 2172-0479
This article is available in: www.transbiomedicine.com
cells (Figure 1C), with no apparent difference in fluorescenceintensity and cell surface distribution among different celltypes. Colchicine, a cytoskeletal destabiliser that inhibitsexocytosis [23], causeda>70%reduction incell surfaceLAMP-1fluorescence intensity onPompe,MPS I andunaffected cellswhencomparedtountreatedcells(P<0.005;Figure 1D-F).Therewasnodifference incell surfaceLAMP-1fluorescence intensityfor Pompe,MPS I andunaffected cells treatedwith colchicine,whiletheabsenceofDAPInuclearstainingindicatedthatthecellshad not been permeabilised. Permeabilised cells displayed anextensivenetworkoflargerLAMP-1-positivevesicularstructures(Figure 1G-I),andDAPInuclearstaining.
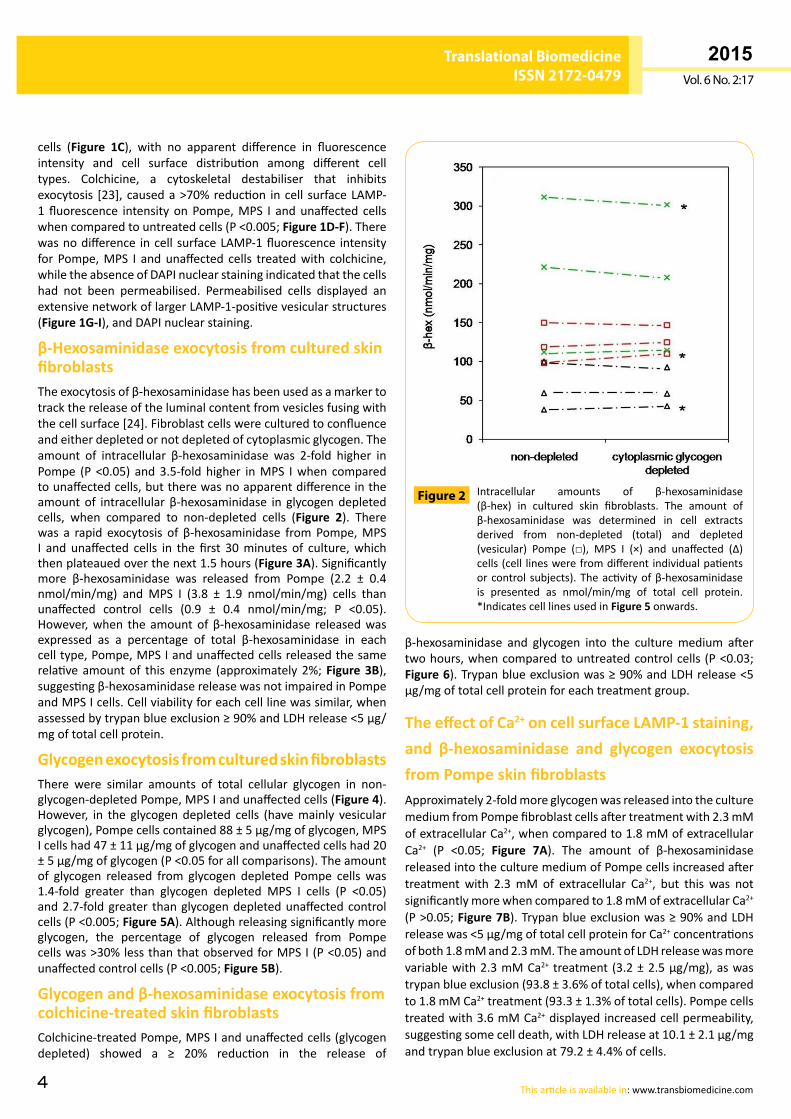
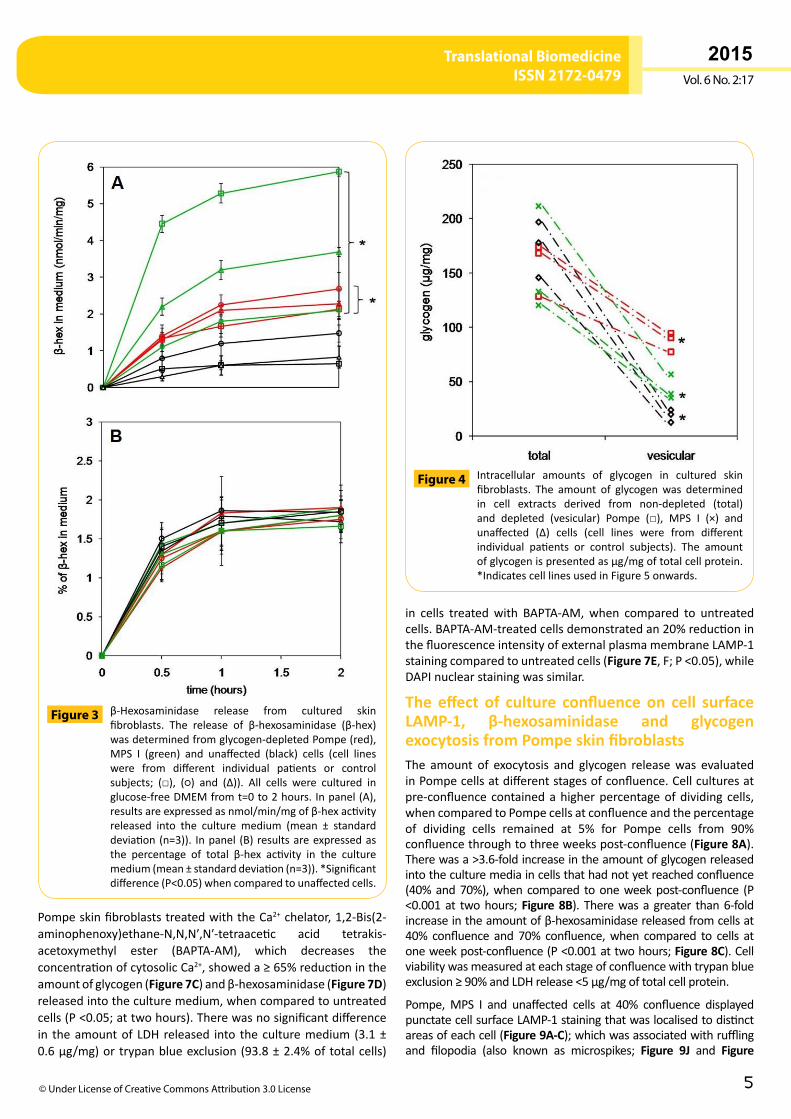
β-Hexosaminidase exocytosis from cultured skin fibroblastsTheexocytosisofβ-hexosaminidasehasbeenusedasamarkertotrackthereleaseoftheluminalcontentfromvesiclesfusingwiththecellsurface[24].Fibroblastcellswereculturedtoconfluenceandeitherdepletedornotdepletedofcytoplasmicglycogen.Theamount of intracellular β-hexosaminidase was 2-fold higher inPompe (P <0.05) and3.5-foldhigher inMPS Iwhen comparedtounaffectedcells,buttherewasnoapparentdifferenceintheamountof intracellularβ-hexosaminidase in glycogendepletedcells, when compared to non-depleted cells (Figure 2). Therewas a rapid exocytosis of β-hexosaminidase fromPompe,MPSI and unaffected cells in the first 30minutes of culture,whichthenplateauedoverthenext1.5hours(Figure 3A).Significantlymore β-hexosaminidase was released from Pompe (2.2 ± 0.4nmol/min/mg) and MPS I (3.8 ± 1.9 nmol/min/mg) cells thanunaffected control cells (0.9 ± 0.4 nmol/min/mg; P <0.05).However,when theamountofβ-hexosaminidase releasedwasexpressed as a percentage of total β-hexosaminidase in eachcelltype,Pompe,MPSIandunaffectedcellsreleasedthesamerelativeamountof this enzyme (approximately2%;Figure 3B),suggestingβ-hexosaminidasereleasewasnotimpairedinPompeandMPSIcells.Cellviabilityforeachcelllinewassimilar,whenassessedbytrypanblueexclusion≥90%andLDHrelease<5µg/mgoftotalcellprotein.
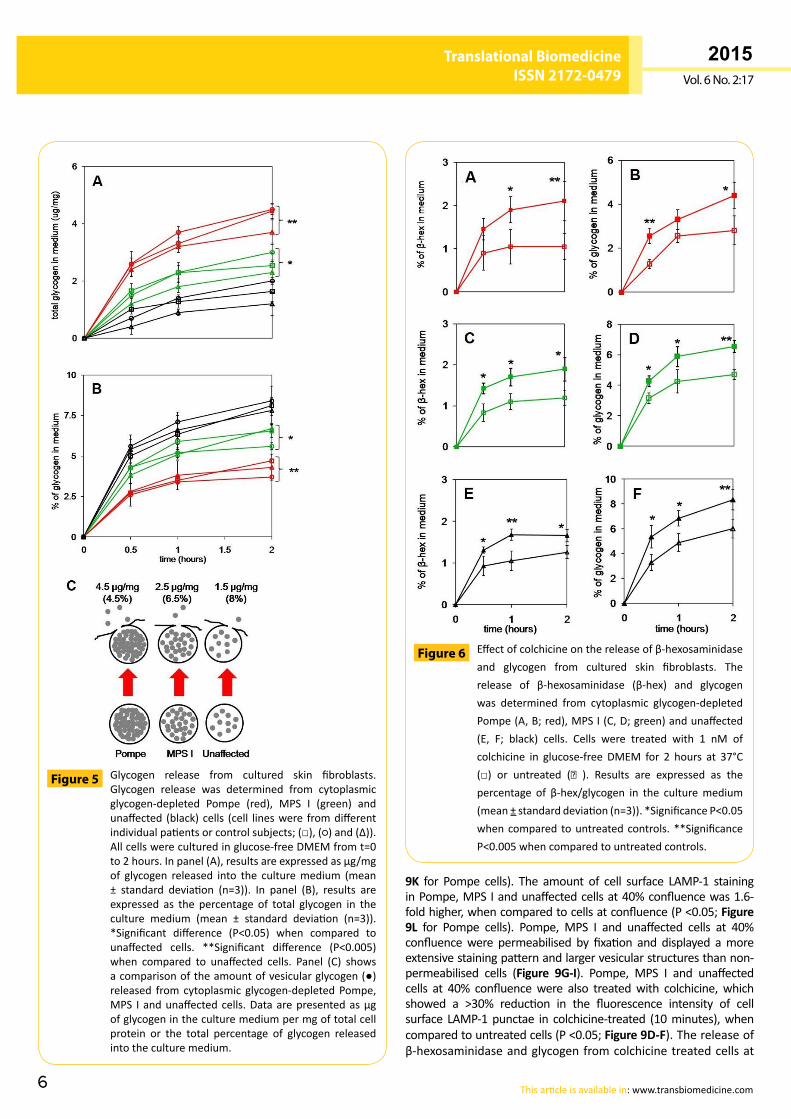
Glycogen exocytosis from cultured skin fibroblastsThere were similar amounts of total cellular glycogen in non-glycogen-depletedPompe,MPSIandunaffectedcells(Figure 4).However, in the glycogendepleted cells (havemainly vesicularglycogen),Pompecellscontained88±5µg/mgofglycogen,MPSIcellshad47±11µg/mgofglycogenandunaffectedcellshad20±5µg/mgofglycogen(P<0.05forallcomparisons).Theamountof glycogen released from glycogen depleted Pompe cells was1.4-fold greater than glycogen depleted MPS I cells (P <0.05)and2.7-foldgreater thanglycogendepletedunaffectedcontrolcells(P<0.005;Figure 5A).Althoughreleasingsignificantlymoreglycogen, the percentage of glycogen released from Pompecellswas>30%lessthanthatobservedforMPSI(P<0.05)andunaffectedcontrolcells(P<0.005;Figure 5B).
Glycogen and β-hexosaminidase exocytosis from colchicine-treated skin fibroblastsColchicine-treatedPompe,MPSIandunaffectedcells(glycogendepleted) showed a ≥ 20% reduction in the release of
β-hexosaminidase and glycogen into the culturemedium aftertwohours,whencomparedtountreatedcontrolcells (P<0.03;Figure 6).Trypanblueexclusionwas≥90%andLDHrelease<5µg/mgoftotalcellproteinforeachtreatmentgroup.
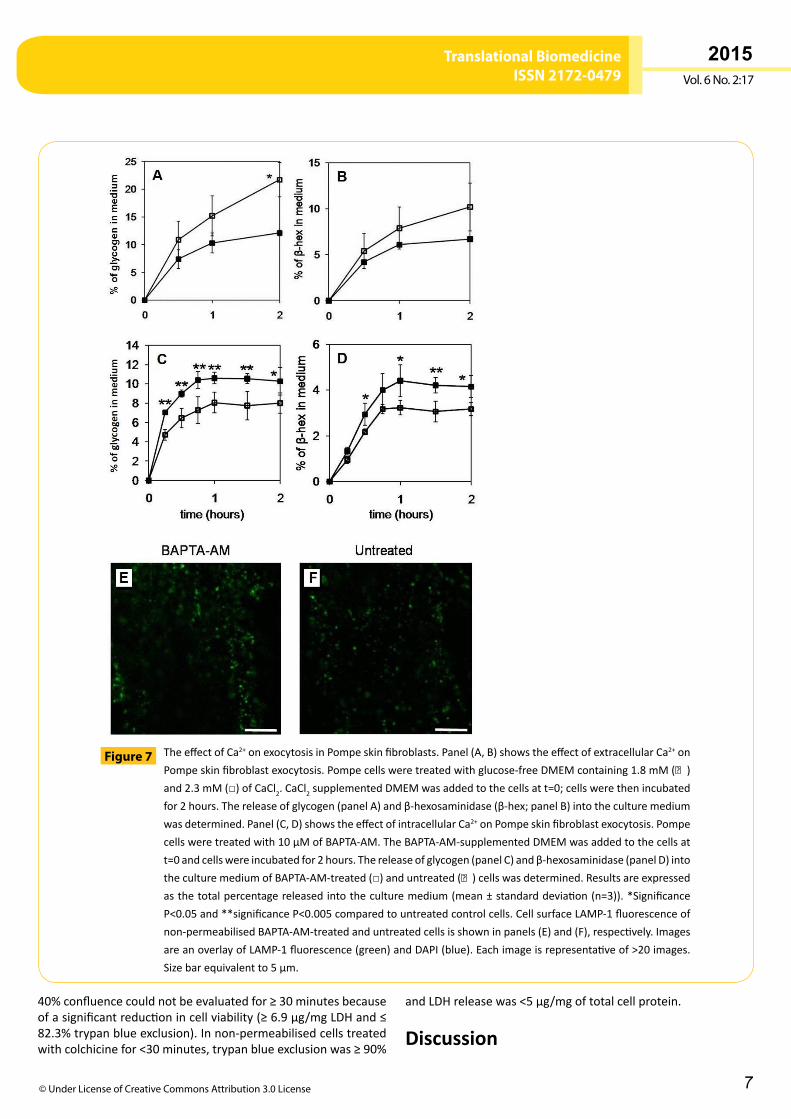
The effect of Ca2+ on cell surface LAMP-1 staining, and β-hexosaminidase and glycogen exocytosis from Pompe skin fibroblastsApproximately2-foldmoreglycogenwasreleasedintotheculturemediumfromPompefibroblastcellsaftertreatmentwith2.3mMofextracellularCa2+,whencomparedto1.8mMofextracellularCa2+ (P <0.05; Figure 7A). The amount of β-hexosaminidasereleasedintotheculturemediumofPompecellsincreasedaftertreatment with 2.3mM of extracellular Ca2+, but this was notsignificantlymorewhencomparedto1.8mMofextracellularCa2+ (P>0.05;Figure 7B).Trypanblueexclusionwas≥90%andLDHreleasewas<5µg/mgoftotalcellproteinforCa2+concentrationsofboth1.8mMand2.3mM.TheamountofLDHreleasewasmorevariablewith2.3mMCa2+ treatment(3.2±2.5µg/mg),aswastrypanblueexclusion(93.8±3.6%oftotalcells),whencomparedto1.8mMCa2+treatment(93.3±1.3%oftotalcells).Pompecellstreatedwith3.6mMCa2+displayedincreasedcellpermeability,suggestingsomecelldeath,withLDHreleaseat10.1±2.1µg/mgandtrypanblueexclusionat79.2±4.4%ofcells.
Intracellular amounts of β-hexosaminidase(β-hex) in cultured skin fibroblasts. The amount ofβ-hexosaminidase was determined in cell extractsderived from non-depleted (total) and depleted(vesicular) Pompe (□), MPS I (×) and unaffected (∆)cells(cell lineswerefromdifferentindividualpatientsorcontrol subjects).Theactivityofβ-hexosaminidaseis presented as nmol/min/mg of total cell protein.*IndicatescelllinesusedinFigure 5onwards.
Figure 2
5
ARCHIVOS DE MEDICINAISSN 1698-9465
2015Vol. 6 No. 2:17
Translational BiomedicineISSN 2172-0479
© Under License of Creative Commons Attribution 3.0 License
PompeskinfibroblaststreatedwiththeCa2+chelator,1,2-Bis(2-aminophenoxy)ethane-N,N,N′,N′-tetraacetic acid tetrakis-acetoxymethyl ester (BAPTA-AM), which decreases theconcentrationofcytosolicCa2+,showeda≥65%reductionintheamountofglycogen(Figure 7C)andβ-hexosaminidase(Figure 7D)releasedintotheculturemedium,whencomparedtountreatedcells(P<0.05;attwohours).Therewasnosignificantdifferencein theamountof LDH released into theculturemedium (3.1±0.6µg/mg)ortrypanblueexclusion(93.8±2.4%oftotalcells)
in cells treatedwith BAPTA-AM,when compared to untreatedcells.BAPTA-AM-treatedcellsdemonstratedan20%reductioninthefluorescenceintensityofexternalplasmamembraneLAMP-1stainingcomparedtountreatedcells(Figure 7E,F;P<0.05),whileDAPInuclearstainingwassimilar.
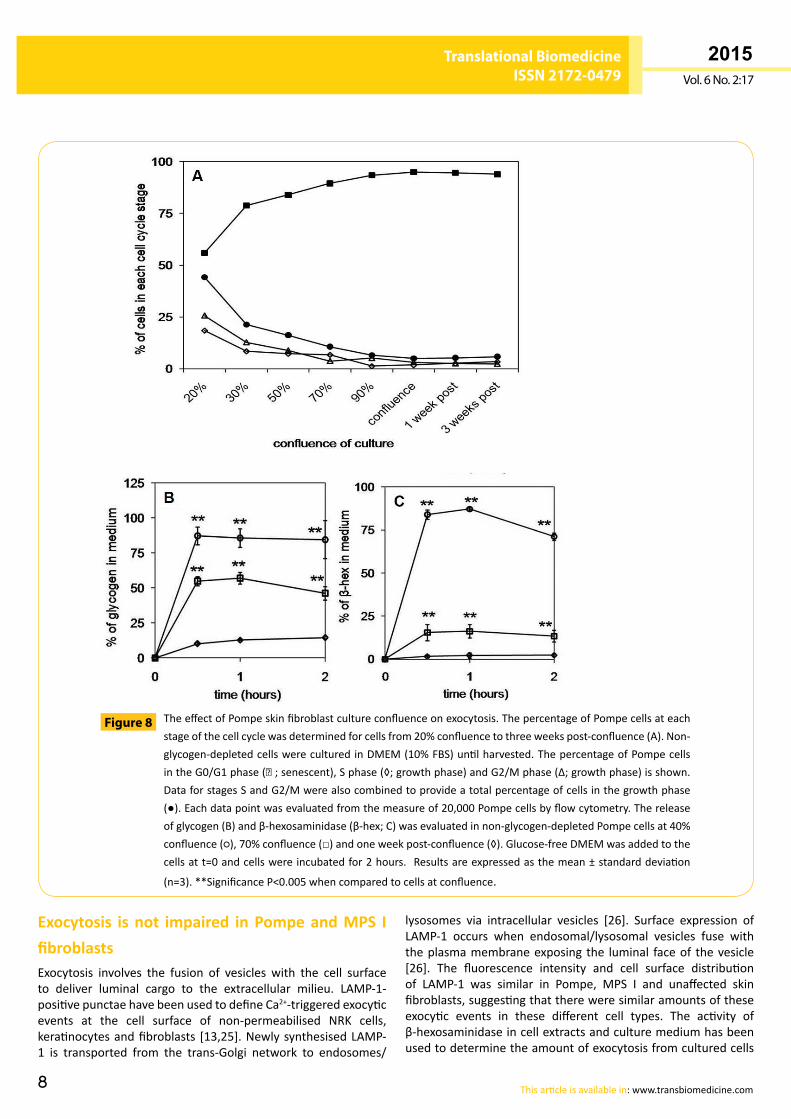
The effect of culture confluence on cell surface LAMP-1, β-hexosaminidase and glycogen exocytosis from Pompe skin fibroblastsThe amount of exocytosis and glycogen releasewas evaluatedinPompecellsatdifferentstagesofconfluence.Cellculturesatpre-confluencecontainedahigherpercentageofdividing cells,whencomparedtoPompecellsatconfluenceandthepercentageof dividing cells remained at 5% for Pompe cells from 90%confluencethroughtothreeweekspost-confluence(Figure 8A).Therewasa>3.6-foldincreaseintheamountofglycogenreleasedintotheculturemediaincellsthathadnotyetreachedconfluence(40%and70%),whencompared tooneweekpost-confluence (P<0.001at twohours;Figure 8B). Therewasagreater than6-foldincreaseintheamountofβ-hexosaminidasereleasedfromcellsat40% confluence and 70% confluence,when compared to cells atoneweekpost-confluence(P<0.001attwohours;Figure 8C).Cellviabilitywasmeasuredateachstageofconfluencewithtrypanblueexclusion≥90%andLDHrelease<5µg/mgoftotalcellprotein.
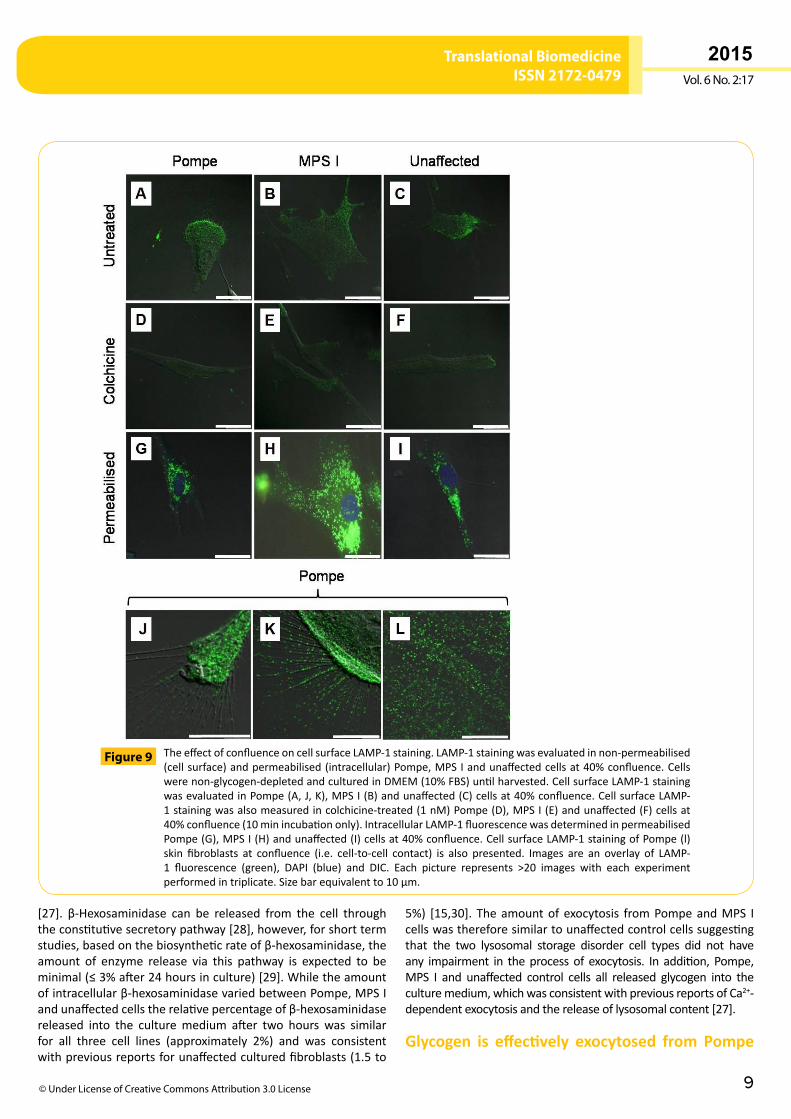
Pompe,MPS I and unaffected cells at 40% confluence displayedpunctatecellsurfaceLAMP-1stainingthatwaslocalisedtodistinctareasofeachcell(Figure 9A-C);whichwasassociatedwithrufflingand filopodia (also known as microspikes; Figure 9J and Figure
β-Hexosaminidase release from cultured skinfibroblasts. The release of β-hexosaminidase (β-hex)wasdeterminedfromglycogen-depletedPompe(red),MPS I (green) and unaffected (black) cells (cell lineswere from different individual patients or controlsubjects; (□), (○) and (∆)). All cells were cultured inglucose-freeDMEMfromt=0to2hours.Inpanel(A),resultsareexpressedasnmol/min/mgofβ-hexactivityreleased into the culture medium (mean ± standarddeviation(n=3)). Inpanel (B) resultsareexpressedasthe percentage of total β-hex activity in the culturemedium(mean±standarddeviation(n=3)).*Significantdifference(P<0.05)whencomparedtounaffectedcells.
Figure 3
Intracellular amounts of glycogen in cultured skinfibroblasts. The amount of glycogenwas determinedin cell extracts derived from non-depleted (total)and depleted (vesicular) Pompe (□), MPS I (×) andunaffected (∆) cells (cell lines were from differentindividual patients or control subjects). The amountofglycogenispresentedasµg/mgoftotalcellprotein.*IndicatescelllinesusedinFigure5onwards.
Figure 4
6
ARCHIVOS DE MEDICINAISSN 1698-9465
2015Vol. 6 No. 2:17
Translational BiomedicineISSN 2172-0479
This article is available in: www.transbiomedicine.com
9K for Pompe cells). The amount of cell surface LAMP-1 staininginPompe,MPSIandunaffectedcellsat40%confluencewas1.6-foldhigher,whencomparedtocellsatconfluence(P<0.05;Figure 9L for Pompe cells). Pompe, MPS I and unaffected cells at 40%confluencewere permeabilised by fixation and displayed amoreextensivestainingpatternandlargervesicularstructuresthannon-permeabilised cells (Figure 9G-I). Pompe, MPS I and unaffectedcells at 40% confluencewere also treatedwith colchicine,whichshowed a >30% reduction in the fluorescence intensity of cellsurface LAMP-1punctae in colchicine-treated (10minutes),whencomparedtountreatedcells(P<0.05;Figure 9D-F).Thereleaseofβ-hexosaminidaseandglycogenfromcolchicinetreatedcellsat
Glycogen release from cultured skin fibroblasts.Glycogen release was determined from cytoplasmicglycogen-depleted Pompe (red), MPS I (green) andunaffected (black)cells (cell lineswere fromdifferentindividualpatientsorcontrolsubjects;(□),(○)and(∆)).Allcellswereculturedinglucose-freeDMEMfromt=0to2hours.Inpanel(A),resultsareexpressedasμg/mgof glycogen released into the culturemedium (mean± standard deviation (n=3)). In panel (B), results areexpressed as the percentage of total glycogen in theculture medium (mean ± standard deviation (n=3)).*Significant difference (P<0.05) when compared tounaffected cells. **Significant difference (P<0.005)when compared to unaffected cells. Panel (C) showsacomparisonoftheamountofvesicularglycogen(●)released fromcytoplasmic glycogen-depletedPompe,MPS Iandunaffectedcells.Dataarepresentedasµgofglycogenintheculturemediumpermgoftotalcellprotein or the total percentage of glycogen releasedintotheculturemedium.
Figure 5
Effectofcolchicineonthereleaseofβ-hexosaminidaseand glycogen from cultured skin fibroblasts. Therelease of β-hexosaminidase (β-hex) and glycogenwas determined from cytoplasmic glycogen-depletedPompe(A,B;red),MPSI(C,D;green)andunaffected(E, F; black) cells. Cells were treated with 1 nM ofcolchicine in glucose-free DMEM for 2 hours at 37°C(□) or untreated (■ ). Results are expressed as thepercentage of β-hex/glycogen in the culturemedium(mean±standarddeviation(n=3)).*SignificanceP<0.05whencompared tountreatedcontrols.**SignificanceP<0.005whencomparedtountreatedcontrols.
Figure 6
7
ARCHIVOS DE MEDICINAISSN 1698-9465
2015Vol. 6 No. 2:17
Translational BiomedicineISSN 2172-0479
© Under License of Creative Commons Attribution 3.0 License
40%confluencecouldnotbeevaluatedfor≥30minutesbecauseofasignificantreductionincellviability(≥6.9µg/mgLDHand≤82.3%trypanblueexclusion).Innon-permeabilisedcellstreatedwithcolchicinefor<30minutes,trypanblueexclusionwas≥90%
andLDHreleasewas<5µg/mgoftotalcellprotein.
Discussion
TheeffectofCa2+onexocytosisinPompeskinfibroblasts.Panel(A,B)showstheeffectofextracellularCa2+onPompeskinfibroblastexocytosis.Pompecellsweretreatedwithglucose-freeDMEMcontaining1.8mM(■)and2.3mM(□)ofCaCl2.CaCl2supplementedDMEMwasaddedtothecellsatt=0;cellswerethenincubatedfor2hours.Thereleaseofglycogen(panelA)andβ-hexosaminidase(β-hex;panelB)intotheculturemediumwasdetermined.Panel(C,D)showstheeffectofintracellularCa2+onPompeskinfibroblastexocytosis.Pompecellsweretreatedwith10µMofBAPTA-AM.TheBAPTA-AM-supplementedDMEMwasaddedtothecellsatt=0andcellswereincubatedfor2hours.Thereleaseofglycogen(panelC)andβ-hexosaminidase(panelD)intotheculturemediumofBAPTA-AM-treated(□)anduntreated(■)cellswasdetermined.Resultsareexpressedasthetotalpercentagereleasedintotheculturemedium(mean±standarddeviation(n=3)).*SignificanceP<0.05and**significanceP<0.005comparedtountreatedcontrolcells.CellsurfaceLAMP-1fluorescenceofnon-permeabilisedBAPTA-AM-treatedanduntreatedcellsisshowninpanels(E)and(F),respectively.ImagesareanoverlayofLAMP-1fluorescence(green)andDAPI(blue).Eachimageisrepresentativeof>20images.Sizebarequivalentto5µm.
Figure 7
8
ARCHIVOS DE MEDICINAISSN 1698-9465
2015Vol. 6 No. 2:17
Translational BiomedicineISSN 2172-0479
This article is available in: www.transbiomedicine.com
TheeffectofPompeskinfibroblastcultureconfluenceonexocytosis.ThepercentageofPompecellsateachstageofthecellcyclewasdeterminedforcellsfrom20%confluencetothreeweekspost-confluence(A).Non-glycogen-depletedcellswereculturedinDMEM(10%FBS)untilharvested.ThepercentageofPompecellsintheG0/G1phase(■;senescent),Sphase(◊;growthphase)andG2/Mphase(Δ;growthphase)isshown.DataforstagesSandG2/Mwerealsocombinedtoprovideatotalpercentageofcellsinthegrowthphase(●).Eachdatapointwasevaluatedfromthemeasureof20,000Pompecellsbyflowcytometry.Thereleaseofglycogen(B)andβ-hexosaminidase(β-hex;C)wasevaluatedinnon-glycogen-depletedPompecellsat40%confluence(○),70%confluence(□)andoneweekpost-confluence(◊).Glucose-freeDMEMwasaddedtothecellsatt=0andcellswereincubatedfor2hours. Resultsareexpressedasthemean±standarddeviation
(n=3).**SignificanceP<0.005whencomparedtocellsatconfluence.
Figure 8
Exocytosis is not impaired in Pompe and MPS I fibroblastsExocytosis involves the fusion of vesicles with the cell surfaceto deliver luminal cargo to the extracellular milieu. LAMP-1-positivepunctaehavebeenusedtodefineCa2+-triggeredexocyticevents at the cell surface of non-permeabilised NRK cells,keratinocytesandfibroblasts [13,25].NewlysynthesisedLAMP-1 is transported from the trans-Golgi network to endosomes/
lysosomes via intracellular vesicles [26]. Surface expression ofLAMP-1 occurs when endosomal/lysosomal vesicles fuse withtheplasmamembraneexposing the luminal faceof thevesicle[26]. The fluorescence intensity and cell surface distributionof LAMP-1 was similar in Pompe, MPS I and unaffected skinfibroblasts,suggestingthatthereweresimilaramountsoftheseexocytic events in these different cell types. The activity ofβ-hexosaminidaseincellextractsandculturemediumhasbeenusedtodeterminetheamountofexocytosisfromculturedcells
9
ARCHIVOS DE MEDICINAISSN 1698-9465
2015Vol. 6 No. 2:17
Translational BiomedicineISSN 2172-0479
© Under License of Creative Commons Attribution 3.0 License
TheeffectofconfluenceoncellsurfaceLAMP-1staining.LAMP-1stainingwasevaluatedinnon-permeabilised(cellsurface)andpermeabilised(intracellular)Pompe,MPSIandunaffectedcellsat40%confluence.Cellswerenon-glycogen-depletedandculturedinDMEM(10%FBS)untilharvested.CellsurfaceLAMP-1stainingwasevaluatedinPompe(A,J,K),MPSI(B)andunaffected(C)cellsat40%confluence.CellsurfaceLAMP-1stainingwasalsomeasuredincolchicine-treated(1nM)Pompe(D),MPSI(E)andunaffected(F)cellsat40%confluence(10minincubationonly).IntracellularLAMP-1fluorescencewasdeterminedinpermeabilisedPompe(G),MPSI(H)andunaffected(I)cellsat40%confluence.CellsurfaceLAMP-1stainingofPompe(l)skin fibroblasts at confluence (i.e. cell-to-cell contact) is also presented. Images are an overlay of LAMP-1 fluorescence (green), DAPI (blue) and DIC. Each picture represents >20 images with each experimentperformedintriplicate.Sizebarequivalentto10µm.
Figure 9
[27]. β-Hexosaminidase can be released from the cell throughtheconstitutivesecretorypathway[28],however,forshorttermstudies,basedonthebiosyntheticrateofβ-hexosaminidase,theamount of enzyme release via this pathway is expected to beminimal(≤3%after24hoursinculture)[29].Whiletheamountofintracellularβ-hexosaminidasevariedbetweenPompe,MPSIandunaffectedcellstherelativepercentageofβ-hexosaminidasereleased into the culturemedium after two hours was similarfor all three cell lines (approximately 2%) and was consistentwithpreviousreportsforunaffectedculturedfibroblasts(1.5to
5%) [15,30]. Theamountofexocytosis fromPompeandMPS Icellswasthereforesimilartounaffectedcontrolcellssuggestingthat the two lysosomal storage disorder cell types did not haveany impairment in theprocessof exocytosis. In addition, Pompe,MPS I and unaffected control cells all released glycogen into theculturemedium,whichwasconsistentwithpreviousreportsofCa2+-dependentexocytosisandthereleaseoflysosomalcontent[27].
Glycogen is effectively exocytosed from Pompe

10
ARCHIVOS DE MEDICINAISSN 1698-9465
2015Vol. 6 No. 2:17
Translational BiomedicineISSN 2172-0479
This article is available in: www.transbiomedicine.com
fibroblastsThe treatment of cultured cells with colchicine, a cytoskeletaldestabiliser, has been reported to inhibit exocytosis throughits action on microtubules and this is thought to impede thetrafficof vesicles to the cell surface [24]. Colchicine treatmentofPompe,MPSIandunaffectedcellsledtoa>30%reductionintheexposureofLAMP-1atthecellsurface,andalsothereleaseof β-hexosaminidase and glycogen, but with no change in cellviability. The release of β-hexosaminidase was similar to thatobservedinNRKcells,withcolchicinetreatmentleadingtoa15%reduction in the exocytosis of this enzyme [30]. Treatment offibroblastcellswithcolchicinewasabletoreducetheamountofglycogenreleasedfromPompe,MPSIandunaffectedcontrolcellssupportingthenotionthatthiswasbeingreleasedbyexocytosis.In cultured Pompe, MPS I and unaffected fibroblast cells,therewasa rapid increase in theamountofβ-hexosaminidasereleased into the culture medium for the first 30 minutes,followed by a plateau in the release. For exocytosis fromNRKcells [24], fibroblasts [12] and mast cells [14] there was also aplateauintheamountofβ-hexosaminidasereleasedaftertimeinculture.Extracellularβ-hexosaminidasecanbere-internalised intofibroblasts by endocytosis [29] and may therefore contribute tothis apparent plateauing effect. Importantly, glycogen release didnot plateau to the sameextent as β-hexosaminidase release andthismayindicatethatthereisnotaspecificuptakemechanismforglycogeninfibroblasts.
Ca2+ concentration affects glycogen exocytosis in Pompe fibroblastsTheconcentrationofCa2+ intheculturemediumhadasignificantimpact on the amount of glycogen exocytosis from Pompe cells.Increasing the concentration of Ca2+ in the culture medium hasbeen reported to facilitate the release of acid hydrolases fromcells [15], whereas decreasing the concentration of Ca2+ reducesthisexocyticrelease[31].Therewasa70%increaseinthereleaseof β-hexosaminidase from Pompe cells in response to a higherconcentration of Ca2+; but a 300% increase in β-hexosaminidasereleasehasbeenreportedinNRKcells[15].Similarly,therewasa65% reduction in the release of β-hexosaminidase from Pompecells in response to BAPTA-AM, whereas NRK cells displayed a>90% reduction [32]. Cultured fibroblasts may therefore be lessresponsivetoCa2+thanothercelltypes.Despitethesedifferencesinβ-hexosaminidaserelease,modulationofCa2+-dependentexocytosiswasabletoalterglycogenexocytosis,confirmingtheworkofMedinaetal.[13].
Cell confluence affects glycogen exocytosis in Pompe fibroblastsPompe cells undergoing cell division released >75% of the totalcell β-hexosaminidase and >80%of the glycogen into the culturemedium after only 2 hours of culture, which was >7-fold higherthanthatobservedforconfluentcells.Thishighamountofglycogenreleasewas surprising asPompe cells only contain approximately50%vesicularglycogen(singlepointintimemeasurement),withtheremainderbeinglocalisedtothecytoplasm.Apotentialexplanationfor thishighpercentageofglycogen releasemaybe thatduring
cell growth cytoplasmic glycogen is rapidly autophagocytosedto provide an energy source to drive these processes, therebydepleting the cytoplasmic stock of glycogen and providing apoolofvesicularglycogenthatissusceptibletoexocyticrelease.Increasedglycogenautophagyprovidesanenergysourceinlivercellsduringthehighenergydemandpost-natalperiod[33].
Up-regulation of glycogen exocytosis in sub-confluent Pompe fibroblasts correlates with in-creased cell migrationCell division and migration are elevated in cultures at pre-confluence[34]. Increasedexocytosishasbeenreportedduringcell division and occurs at the cleavage furrow, as cells divide[35].Exocytosisalsooccursduringcellmigrationandthisinvolvesmembrane localised to the leadingedgeof thecell [36].Therewasnoobvious increase in cell surface LAMP-1 staining at thecleavage furrowofdividingPompecells.Golgi-derivedvesicles,but not endosomal or lysosomal compartments, have beenlinkedtoincreasedexocytosisduringcelldivision[35],implicatingCa2+-independentexocytosis.Insub-confluent,migratingPompecells, there was an increase in cell surface LAMP-1 staining attheleadingedgeofthecell,inareasofrufflingandonfilopodia.Thesestructuresareinvolvedinthetransferofmembranefroman intracellular location to the cell surface, thereby enablingforward motion [36]. The elevated exocytosis associated withsub-confluentPompeskinfibroblasts,asmeasuredbyincreasedLAMP-1stainingandβ-hexosaminidaserelease,maythereforeberelatedtocellmigrationratherthancelldivisionperse.
Glycogen release from Pompe and MPS I fibroblasts is partially impairedWhen expressed as a percentage of total cellular glycogen foreachcell,Pompecellsreleasedthelowestproportionofvesicularglycogen compared toMPS I and unaffected cells (Figure 5C).However, theamountof exocytosis for eachof these cell lineswassimilar,asdeterminedbythepresenceofcellsurfaceLAMP-1stainingandthereleaseofβ-hexosaminidaseintotheculturemedium. This suggested that the reduced glycogen releasefromPompe(andtoalesserextentMPSI)cellswasnotduetoimpairedexocytosisperse.OnepossibleexplanationisthatCa2+-inducedexocytosis,whichisthelikelymechanismresponsibleforglycogenrelease,results incavicapture[37], thepartial releaseof vesicle content through an exocytic pore. An explanationfor this reduced glycogen exocytosismaybe that the glycogengranules in Pompe cellsmay be larger in diameter than thosein unaffected cells, and therefore limited in their ability to beexocytosed. Alternatively, the diameter of the exocytic pore inPompe cells may be restricted, when compared to unaffectedcells,whichwouldalsolimitglycogenrelease.Thesetwopossiblemechanisms for the reduced release of glycogen from Pompecellsarethefocusoffuturestudies.
MPS I fibroblasts accumulate glycogenMPS I cells were included in this study as these cellsaccumulatedifferentstoragesubstratestoPompecells,namelyglycosaminoglycans.However,MPSIcellswereshowntocontain

11
ARCHIVOS DE MEDICINAISSN 1698-9465
2015Vol. 6 No. 2:17
Translational BiomedicineISSN 2172-0479
© Under License of Creative Commons Attribution 3.0 License
moreglycogenthanunaffectedcells,indicatingelevatedvesicularglycogen stores. While the defect in GAA explains increasedglycogeninPompecells,adefectinα-L-iduronidasewouldnotbeexpectedtoresultinglycogenstorage.ItremainsunknownastowhyglycogenaccumulatesinMPSIcells,andfurtherstudiesarerequired.However,itcouldbespeculatedthatglycosaminoglycanstorage in MPS I cells either impairs the catalytic activity inendosome-lysosome compartments or limits lysosomal fusionandthereforedegradationinautolysosomes.
ConclusionIn this study, we showed no impairment of exocytosis inPompe cells as defined by cell surface LAMP-1 staining andβ-hexosaminidase release; and that glycogen was exocytosedfrom these cells. Importantly, glycogen release from Pompefibroblasts was up-regulated by certain culture conditions,presumablyduetospecificeffectsontheexocyticmachinery.Thisstudy provided proof of concept that glycogen exocytosismaybeusedtore-locatestoragematerialfromPompecellsintothe
extracellularmilieu.TheinductionofCa2+-independentexocytosisappearedtoreleasemoreglycogenfromPompecellsthanCa2+-dependent exocytosis. A greater understanding of the Ca2+-independentexocyticmechanismmayleadtotheidentificationofnewtherapeutictargets,whichmaybemorebeneficialthanthose identified for the inductionofCa2+-dependentexocytosis[13].The inductionofglycogenexocytosisopensthepossibilityforanalternativetherapeuticstrategybasedondegradationbycirculatingamylases.
AcknowledgementsThe authors gratefully acknowledge funding from theMasonicFoundation. C. Turner was funded by the NHMRC Dora LushScholarship.
FundingThe authors gratefully acknowledge funding from theMasonicFoundation. C. Turner was funded by the NHMRC Dora LushScholarship.

12
ARCHIVOS DE MEDICINAISSN 1698-9465
2015Vol. 6 No. 2:17
Translational BiomedicineISSN 2172-0479
This article is available in: www.transbiomedicine.com
References1 Qureshi OS, Paramasivam A, Yu JC, Murrell-Lagnado RD (2007)
Regulation of P2X4 receptors by lysosomal targeting, glycanprotectionandexocytosis.JCellSci120:3838-3849.
2 CardoneM, Porto C, Tarallo A, VicinanzaM, Rossi B, et al. (2008)Abnormal mannose-6-phosphate receptor trafficking impairsrecombinantalpha-glucosidaseuptakeinPompediseasefibroblasts.Pathogenetics1(1):6.
3 KishnaniPS,BeckemeyerAA(2014)NewtherapeuticapproachesforPompedisease:enzymereplacement therapyandbeyond.PediatrEndocrinolRev.12Suppl1:114-124.
4 Sugo T, Tachimoto H, Chikatsu T, Murakami Y, Kikukawa Y, et al.(2006) Identificationofa lysophosphatidylserine receptoronmastcells.BiochemBiophysResCommun341:1078-1087.
5 Hawes ML, Kennedy W, O'Callaghan MW, Thurberg BL (2007)DifferentialmuscularglycogenclearanceafterenzymereplacementtherapyinamousemodelofPompedisease.MolGenetMetab91:343-351.
6 SesakiH,OgiharaS (1997)Protrusionof cell surface coupledwithsingle exocytotic events of secretion of the slime in Physarumplasmodia.JCellSci110:809-818.
7 RodríguezA,Martinez I, ChungA,BerlotCH,AndrewsNW (1999)cAMP regulates Ca2+-dependent exocytosis of lysosomes andlysosome-mediatedcellinvasionbytrypanosomes.JBiolChem274:16754-16759.
8 Kumlien J, Andrén-Sandberg A, Zopf D, Lundblad A (1989)Determination of a glucose-containing tetrasaccharide in urine ofpatientswithacutepancreatitis.IntJPancreatol4:139-147.
9 HesselinkRP,WagenmakersAJ,DrostMR,VanderVusseGJ(2003)LysosomaldysfunctioninmusclewithspecialreferencetoglycogenstoragediseasetypeII.BiochimBiophysActa1637:164-170.
10 An Y, Young SP, Kishnani PS, Millington DS, Amalfitano A, et al.(2005)Glucose tetrasaccharide as a biomarker formonitoring thetherapeutic response to enzyme replacement therapy for Pompedisease.MolGenetMetab85:247-254.
11 Klein D, BüssowH, Fewou SN, Gieselmann V (2005) Exocytosis ofstorage material in a lysosomal disorder. Biochem Biophys ResCommun327:663-667.
12 Chen FW, Li C, Ioannou YA (2010) Cyclodextrin induces calcium-dependentlysosomalexocytosis.PLoSOne.5:e15054.
13 Martinez I, Chakrabarti S, Hellevik T,Morehead J, Fowler K, et al.(2000)SynaptotagminVIIregulatesCa(2+)-dependentexocytosisoflysosomesinfibroblasts.JCellBiol.148:1141-1149.
14 Spampanato C, Feeney E, Li L, Cardone M, Lim JA et al. (2013)TranscriptionfactorEB(TFEB)isanewtherapeutictargetforPompedisease.EMBOMolMed5:691-706.
15 Martina JA, Diab H, Lishu L, Jeong-A L, Patange S, et al. (2014) Thenutrient-responsive transcription factor TFE3 promotes autophagy,lysosomalbiogenesis,andclearanceofcellulardebris.SciSignal21:7.
16 BargS,MacadoJD(2008)Compensatoryendocytosisinchromaffincells.ActaPhysiol192:195-201.
17 RodríguezA,Webster P,Ortego J, AndrewsNW (1997) Lysosomesbehave as Ca2+-regulated exocytic vesicles in fibroblasts andepithelialcells.JCellBiol137:93-104.
18 Jaiswal JK, Andrews NW, Simon SM (2002) Membrane proximallysosomesarethemajorvesiclesresponsibleforcalcium-dependentexocytosisinnonsecretorycells.JCellBiol159:625-635.
19 Medina DL, Fraldi A, Bouche V, Annunziata F,Mansueto G, et al.(2011)Transcriptionalactivationof lysosomalexocytosispromotescellularclearance.DevCell21:421-430.
20 FullerM,DuplockS,TurnerC,DaveyP,BrooksDA,etal.(2012)MassspectrometricquantificationofglycogentoassessprimarysubstrateaccumulationinthePompemouse.AnalBiochem.421:759-763.
21 LeabackDH,WalkerPG(1961)Studiesonglucosaminidase.4.ThefluorimetricassayofN-acetyl-beta-glucosaminidase.BiochemJ78:151-156.
22 GivanAL(2001)Flowcytometry:firstprinciples2ndedn.Wiley,John&Sons,Inc.NewYork.
23 PanCY,LeeH,ChenCL(2006)Lysophospholipidselevate[Ca2+]iandtrigger exocytosis in bovine chromaffin cells. Neuropharmacology.51:18-26.
24 Kuncl RW, Bilak MM, CraigSW, Adams R (2003) Exocytotic"constipation" is amechanism of tubulin/lysosomal interaction incolchicinemyopathy.ExpCellRes285:196-207.
25 LaPlante JM, SunM, Falardeau J, Dai D, Brown EM, et al. (2006)LysosomalexocytosisisimpairedinmucolipidosistypeIV.MolGenetMetab.89:339-348.
26 EskelinenEL,TanakaY,SaftigP(2003)Attheacidicedge:emergingfunctionsfor lysosomalmembraneproteins.TrendsCellBiol13:137-145.
27 SheaLRabenN(2009)Autophagyinskeletalmuscle:implicationsforPompedisease.IntJClinPharmacolTher.47Suppl1:S42-S47.
28 Rozaklis T, Ramsay SL,Whitfield PD, Ranieri E, Hopwood JJ, et al.(2002) Determination of Oligosaccharides in Pompe Disease byElectrospray IonizationTandemMassSpectrometry.ClinChem48:131-139.
29 TiwariN,WangCC,BrochettaC,KeG,VitaF,etal. (2008)VAMP-8segregatesmastcell-preformedmediatorexocytosis fromcytokinetraffickingpathways.Blood111:3665-3674.
30 RabenN,FukudaT,GilbertAL,deJongD,ThurbergBL,etal.(2005)Replacing acid alpha-glucosidase in Pompe disease: recombinantand transgenic enzymes are equipotent, but neither completelyclearsglycogenfromtypeIImusclefibers.MolTher11:48-56.
31 LiD,RopertN,KoulakoffA,GiaumeC,OheimM(2008)Lysosomesare the major vesicular compartment undergoing Ca2+-regulatedexocytosisfromcorticalastrocytes.JNeurosci28:7648-7658.
32 Ito N, Yokomizo T, Sasaki T, Kurosu H, Penninger J, et al. (2002)Requirementofphosphatidylinositol3-kinaseactivationandcalciuminfluxforleukotrieneB4-inducedenzymerelease.JBiolChem277:44898-44904.
33 Kotoulas OB, Kalamidas SA, Kondomerkos DJ (2004) Glycogenautophagy.MicroscResTech64:10-20.
34 Boucrot E, Kirchhausen T (2007) Endosomal recycling controlsplasmamembraneareaduringmitosis.ProcNatlAcadSciUSA104:7939-7944.
35 GossJW,ToomreDK(2008)Bothdaughtercellstrafficandexocytosemembraneatthecleavagefurrowduringmammaliancytokinesis.J Cell Biol181:1047-1054.
36 SagherianC,ThornerP,MahuranD(1994)Thepro-peptideofthepro

13
ARCHIVOS DE MEDICINAISSN 1698-9465
2015Vol. 6 No. 2:17
Translational BiomedicineISSN 2172-0479
© Under License of Creative Commons Attribution 3.0 License
beta-polypeptidechainofhumanbeta-hexosaminidaseisnecessaryforproperproteinfoldingandexitfromtheendoplasmicreticulum.BiochemBiophysResCommu204:135-141.
37 Jaiswal JK, Chakrabarti S, Andrews NW, Simon SM (2004)SynaptotagminVIIrestrictsfusionporeexpansionduringlysosomalexocytosis.PLoSBiol2:233.





























