Embed Size (px)
Citation preview

THE JOURNAL OF COMPARATIVE NEUROLOGY 364~254-266 (1996)
Glutamatergic Inputs From the Pedunculopontine Nucleus to Midbrain
Dopaminergic Neurons in Primates: Phaseolus uuZgaris-Leucoagglutinin
Anterograde Labeling Combined With Postembedding Glutamate and GABA
Immunohistochemistry
ALI CHARARA, YOLAND SMITH, AND ANDRE PARENT Centre de Recherche en Neurobiologie, HBpital de 1'Enfant-Jesus and Universite Laval,
Quebec, QC, G1J 124, Canada
ABSTRACT To verify the possibility that the pedunculopontine nucleus is a source of glutamatergic
terminals in contact with midbrain dopaminergic neurons in the squirrel monkey, we used the anterograde transport of Phaseolus uulgaris-leucoagglutinin in combination with preembed- ding immunohistochemistry for tyrosine hydroxylase and for calbindin D-28k and postembed- ding immunocytochemistry for glutamate and for y-aminobutyric acid. Following tracer injections in the pedunculopontine nucleus, numerous anterogradely labeled fibers emerged from the injection sites to innervate densely the pars compacta of the substantia nigra and ventral tegmental area. The major type of labeled fibers were thin with multiple collaterals and varicosities that established intimate contacts with midbrain dopaminergic neurons. At the electron microscopic level, the anterogradely labeled boutons were medium sized (maximum diameter between 0.9 pm and 2.5 pm) and contained numerous round vesicles and mitochon- dria. Postembedding immunocytochemistry revealed that 40-60% of anterogradely labeled terminals were enriched in glutamate and formed asymmetric synapses with dendritic shafts of substantia nigra and ventral tegmental area neurons. In triple-immunostained sections, some of the postsynaptic targets to these terminals were found to be dopaminergic. In addition, 30-40% of the anterogradely labeled terminals in both regions displayed immunoreactivity for y-aminobutyric acid and, in some cases, formed symmetric synapses with dendritic shafts.
In conclusion, our results provide the first ultrastructural evidence for the existence of synaptic contacts between glutamate-enriched terminals from the pedunculopontine nucleus and midbrain dopaminergic neurons in primates. Our results also show that the pedunculopon- tine nucleus is a potential source of y-aminobutyric acid input to this region. These findings suggest that the pedunculopontine nucleus may play an important role in the modulation of the activity of midbrain dopaminergic cells by releasing glutamate or y-aminobutyric acid as neurotransmitter. I 19% Wiley-Liss, Inc.
Indexing terms: basal ganglia, substantia nigra, ventral tegmental area, squirrel monkey, tract-tracing methods
It has been shown in Dharmacoloeical and electroohvsi- " 1 "
ological studies that glutamate plays a crucial role in the generation Of burst firing in midbrain dopaminergic neu- rOnS of the substantia nigra (SN) and of the ventral tegmental area (VTA; Grace and Bunney, 1984; Johnson et
Accepted July 1995, Address reprint requests to Andre Parent, Ph.D., Centre de Recherche en
Neurobinlogie, HBpital de 1'Enfant-Jesus, 1401, 18ieme Rue, QuBbec, QC, G1J 124, Canada.
O 1996 WILEY-LISS, INC.

PEDUNCULOPONTINE INPUTS TO SUBSTANTIA NIGRA 255
al., 1992; Overton and Clark, 1992). The pattern of dis- charge of midbrain dopaminergic neurons is directly corre- lated with the release of dopamine from terminals (Suaud- Chagny et al., 1992) that ultimately affects sensorimotor, cognitive, and emotional behaviors. Because the bursting activity is not observed in vitro (Grace and Onn, 1989), it is likely that glutamate is released by afferent inputs to the SN and the VTA. In accordance with these data, the results of the companion paper (Smith et al., 1996) have shown that a large proportion of glutamate-enriched terminals establishes synaptic contacts with dopaminergic neurons in the substantia nigra pars compacta (SNc) and in the VTAof squirrel monkey. The potential sources of these afferents are reported to arise from the cerebral cortex and from the subthalamic nucleus (STH). Indeed, a direct pathway from the frontal cortex to the SN and the VTA has been described (Sakai, 1988; Sesack et al., 1989; Sesack and Pickel, 1992), and electrical stimulation of the medial prefrontal cortex and anterior cingulate cortex induced burst firing in a small proportion of midbrain dopaminergic neurons (Gariano and Groves, 1988). Although the STH preferentially innervates substantia nigra pars reticulata (SNr) neurons (Kita and Kitai, 1987; Smith et al., 1990), stimulation of this nucleus induced burst firing in the dopaminergic cells in the rat SN (Hammond et al., 1978; Smith and Grace, 1992), whereas lesions of this nucleus decreased the baseline firing activity of midbrain dopaminergic cells (Smith and Grace, 1992). However, inputs from the cerebral cortex and STH are not sufficiently widespread to provide the large number of glutamatergic terminals in contact with midbrain dopamin- ergic neurons in monkey (Smith et al., 1996).
These observations raise the possibility that additional sources, namely the pedunculopontine nucleus (PPN), may also contribute to glutamatergic innervation of SNc and VTA neurons. In this regard, anatomical and immunohisto- chemical studies have shown that the PPN contains choline acetyltransferase ( C U T ) - and glutamate-immunoreactive cells (Clements and Grant, 1990; Lavoie and Parent, 1994a), both of which participate to the pedunculonigral projection in primates (Lavoie and Parent, 1994~) . Moreover, stimula- tion of the PPN evoked glutamate-mediated excitation of dopaminergic neurons in the rat SN (Scarnati et al., 1984, 1986) that was abolished by excitatory amino acid receptor antagonists but not by cholinergic antagonists (Scarnati et al., 1986; Di Loreto et al., 1992). Thus far, the existence of direct synapses between glutamatergic terminals from the PPN and midbrain dopaminergic neurons has never been demonstrated. Therefore, we decided to carry out the present study in order to verify the possibility that the PPN is a source of glutamatergic afferents to dopaminergic cells in the SNc and the VTA of squirrel monkeys. To reach this objective, the anterograde transport of Phaseolus vulgaris- leucoagglutinin (PHA-L) was combined with preembedding immunohistochemistry for tyrosine hydroxylase (TH) and calbindin D-28k (CaBP) and postembedding immunohisto- chemistry for glutamate and 7-aminobutyric acid (GABA). As mentioned in the companion paper (Smith et al., 1996), CaBP serves as a specific marker for dopaminergic neurons in the VTA and in the dorsal tier of the SNc (Rogers, 1992; Gaspar et al., 1993). Preliminary results of this study have been published in abstract form (Charara et al., 1993; Smith et al., 1994, 1995).
MATERIALS AND METHODS Three adult male squirrel monkeys (Saimirz sciureus)
weighing 950-1,050 g were used in the present study. They were anesthetized with a mixture of ketamine hydrochlo- ride (Ketaset; 70 mg/kg, i.m) and xylazine (10 mgikg, i.m) and received bilateral iontophoretic injections of the antero- grade tracer PHA-L (Vector Labs, Burlingame, CA; 2.5% solution in phosphate buffer; 0.01 M, pH 8.0). The depth of anesthesia was determined by monitoring heart rate, respi- ration rate, and muscle tone as well as corneal and toe- pinch reflexes. The surgical and anesthesia procedures were performed according to the guidelines of the Canadian Council on Animal Care. The injections were made accord- ing to stereotaxic coordinates chosen from the atlas of Emmers and Akert (1963). The solution was loaded into glass micropipettes with a tip diameter of 30-50 pm and was delivered iontophoretically at two different locations along the anteroposterior extent of the PPN with a 7-10 pA positive current in a 7 seconds on17 seconds off cycle over a period of 20 minutes (Gerfen and Sawchenko, 1984). The pipette was introduced through an oblique approach in order to avoid the contamination of structures overlying the PPN. During the first 2 days after the surgery, the animals received injections of analgesic (buprenorphine; 0.01 mgikg, i.m.) twice daily.
After a survival period of 10 days, the animals were deeply anesthetized with an overdose of pentobarbital and were perfused with 500 ml of a cold Ringer’s solution followed by 1,500 ml of a fixative containing 2% paraformal- dehyde and 1% glutaraldehyde in phosphate buffer (PB; 0.1 M), pH 7.4, and, finally, with 1,000 ml of PB (0.1 M), pH 7.4. Following perfusion, the brains were sectioned stereotaxi- cally in transverse blocks and were placed in cold phosphate- buffered saline (PBS; 0.1 M), pH 7.4. The blocks were then cut with a vibrating microtome in 60-km-thick sections in the transverse plane. The sections were collected in PBS and were treated for 20 minutes with sodium borohydride (1% in PBS). They were then washed several times in PBS and were processed to reveal PHA-L and TH or CaBP for light and electron microscopy.
Immunohistochemical localization of PHA-L and TH/CaBP
Light microscopy. A series of sections throughout the rostrocaudal extent of PPN and S N was prepared for the visualization of PHA-L in the light microscope. Sections were first incubated overnight with a goat anti-PHA-L antiserum (Vector Labs) diluted 1:2,000 in PBS containing 0.1% Triton X-100 and 2% normal rabbit serum (NRS). After this primary incubation, the sections were rinsed in PBS and were incubated with biotinylated rabbit anti-goat immunoglobulin (IgG; Vector Labs) diluted 1:200 in PBS containing 0.1% Triton X-100 and 2% NRS for 90 minutes. This was followed by washings in PBS and then an incuba- tion in the avidin-biotin-peroxidase complex (ABC; Vector Labs, 1:100 in PBS) for 90 minutes according to the method of Hsu et al. (1981). All of the incubations were carried out at room temperature. After a 30 minutes rinse in PBS and Tris buffer (0.05 M), pH 7.6, the bound peroxidase was revealed by placing the sections in a solution containing 3,3’-diaminobenzidine tetrahydrochloride (DAB; Sigma, St. Louis, MO; 0.05%) and hydrogen peroxide (H202, 0.005%) in Tris buffer for 10-15 minutes. The reaction was stopped by repeated washes in PBS.

256 A. CHARARA ET AL.
A series of PHA-L-immunostained sections throughout the SN was processed for TH or CaBP immunohistochemis- try. Sections were incubated overnight with mouse antisera directed against TH (INCSTAR; 1:1,000) or CaBP (Sigma; 1:2,500) diluted in PBS containing 0.1% Triton X-100 and 2% normal horse serum (NHS). They were then washed in PBS and incubated for 90 minutes with biotinylated horse anti-mouse IgG (Vector Labs) diluted 1:200 in PBS contain- ing 0.1% Triton X-100 and 2% NHS. After washing in PBS, the sections were incubated in ABC (1:lOO in PBS) for 90 minutes. Ail of the incubations were carried out at room temperature. The sections were washed in PBS and Tris buffer and were then incubated in a solution of Tris containing DAB (0.025%), nickel (Ni)-ammonium sulfate (Fisher; 0.35%), and HzOz (0.003%). The reaction was stopped by repeated washings in PBS. All of the sections were then mounted on gelatin-coated slides and dehy- drated, and a coverslip was applied with Permount.
Electron microscopy. A series of sections throughout the rostrocaudal extent of SN was processed to localize PHA-L and THiCaBP for electron microscopy. The sections were placed in a cryoprotectant solution (PB, 0.05 M, pH 7.4) containing 25% sucrose and 10% glycerol for 20-30 minutes. After sinking, they were frozen at -80°C for 20 minutes. They were then thawed and washed many times in PBS before being processed for the visualization of PHA-L and THiCaBP according to the protocol described above, except that Triton X-100 was not included in the solutions, and the incubations with primary antisera were carried out at 4°C for 48 hours. The DAB was used to reveal PHA-L, whereas TH and CaBP were localized with benzi- dine dihydrochloride (BDHC; Sigma) according to the proto- col introduced by Levey et al. (1986). Briefly, the sections were washed twice (for 20 minutes) in PBS and five times (for 30 minutes) in PB (0.01 M), pH 6.8, before being preincubated in a solution of BDHC for 10 minutes. The reaction was then initiated by adding hydrogen peroxide to a final concentration of 0.005% in fresh BDHC solution. The reaction was stopped by repeated washings in PB (0.01 M), pH 6.8, and the sections were processed for electron microscopy. They were postfixed in osmium tetroxide (1% in PB 0.01 M, pH 6.8) for 20 minutes. This was followed by washings in PB and dehydration in a graded series of alcohol and propylene oxide. Uranyl acetate was added to the 70% ethanol over 30 minutes to improve the contrast in the electron microscope. The sections were then embedded in resin (Durcupan; ACM, Fluka) on microscope slides, put in the oven, and cured for 48 hours at 60°C.
After detailed examination in the light microscope, re- gions in the SN and the VTA containing anterogradely labeled terminals that seemed to be apposed on TH- and CaBP-immunoreactive elements were drawn, photographed, cut out from the slides, and glued on the top of resin blocks with cyanoacrylate glue. Ultrathin sections were then cut on an ultramicrotome and were collected on pioloform- coated, single-slot nickel grids. They were then processed for postembedding glutamate and GABA immunohistochem- istry according to the protocol described in the companion paper (Smith et al., 1996). In brief, serial ultrathin sections were incubated overnight in a rabbit antisera raised against glutamate (Arne1 Products, New York, NY; 1:10,000) or GABA (1:10,000). The specificity of these antisera is de- scribed in detail elsewhere (Hodgson et al., 1985; Somogyi and Hodgson, 1985; Somogyi et al., 1985; Hepler et al., 1988). This was followed by an incubation in a secondary
antibody conjugated to colloidal gold (Biocell, Cardiff, United Kingdom; 1:25) for 90 minutes. Finally, the sections were stained with lead citrate (Reynolds, 1963) and were exam- ined with an electron microscope (Phillips EM 300).
Control experiments The control experiments that were carried out on the
present material are described in detail in the companion paper (Smith et al., 1996).
Analysis of the material In the sections prepared for light microscopy, the location
and extent of PHA-L injection sites in the PPN were determined and photographed. The anterograde labeling throughout the rostrocaudal extent of the SN and the VTA was examined and is schematically represented in Figure 1.
In the material prepared for electron microscopy, a total of five blocks containing PHA-L-labeled terminals apposed to the surface of dopaminergic elements were selected from the SNc, and four were taken in the VTA. All of these blocks except one were taken from the same animal. This animal was chosen because of the larger density of PHA-L-labeled terminals at midbrain level. The ultrathin sections were scanned for the presence of PHA-L-immunoreactive bou- tons. Once they were found, the terminals were examined in two to four serial sections to verify the type of synaptic specialization associated with them. The postsynaptic tar- gets contacted by these boutons were determined, and the number of those targets that displayed TH or CaBP immu- noreactivity was recorded.
In the sections processed for glutamate or GABA, the tissue was overlaid with gold particles. On visual inspec- tion, the gold particles appeared to be heterogeneously distributed over the tissue. Some elements were enriched in gold particles, whereas others were almost completely devoid of labeling in glutamate- and GABA-immunostained material. Quantitative measurements were carried out to substantiate the differences in intensity of labeling. The cross-sectional area of the boutons was measured with the aid of the Ultimage software (Graftek, France) and a Macintosh computer. All gold particles (including those over mitochondria) were counted over each profile. The results obtained for these measurements are illustrated and described in detail in Figure 3 of the companion paper (Smith et al., 1996). Because the same material was used in both studies, the immunoreactivity or nonimmunoreactiv- ity of anterogradely labeled terminals was determined by comparing the density of gold particles overlying the PHA- L-containing boutons to the density overlying neighboring terminals of the same category that did not contain the anterograde tracer. The density of gold particles associated with anterogradely labeled boutons was also compared to the density overlying putative structures with low levels of glutamate immunoreactivity in the same tissue. For in- stance, in the glutamate-immunostained material, the den- sity of gold particles overlying PHA-L-labeled boutons was compared to that associated with glutamate-enriched type I terminals (Smith et al., 1996), because they share similar ultrastructural features. The boutons forming symmetric synapses and the glial cells were used as negative controls (see Smith et al., 1996). In light of the quantitative data presented in Figure 3 of the companion paper (Smith et al., 1996), we considered a PHA-L-containing bouton enriched in glutamate if the density of gold particles associ-

v TA I
Fig. 1. Phaseolus uulgaris-leucoagglutinin (PHA-Ll injection sites in the pedunculopontine nucleus !PPN; A,Bl in two cases, and the resulting anterograde labeling in the substantia nigra pars compacta (SNcl and the ventral tegmental area !VTA C-Fl following the injection shown in A. In C-F, the anteroposterior levels derived from the atlas of Emmers and Akert (1963) are indicated in the lower right
corner of each drawing. CP, cerebral peduncle; ML, medial lemniscus; MLF, mesencephalic longitudinal fasciculus; RRA, retrorubral area; SNr, substantia nigra pars rcticulata; SP, superior peduncle; STH, subthalamic nucleus; 111, oculomotor nerve root fibers; IV, trochlear nerve root fibers. Scale bar = 1 mm in A !also applies to B-Fl.

258 A. CHARARA ET AL.
ated with it was at least 50/pm2. In the GABA-immuno- stained tissue, this was the reverse; i.e., glutamate-enriched type I terminals and glial cells served as negative controls, whereas terminals forming symmetric synapses were the positive elements.
RESULTS Location of PHA-L injection sites in the PPN In one of the three animals that received PHA-L injec-
tions in the PPN, the injection sites on both sides were small and were located slightly rostra1 to the PPN. This animal was not retained for the purpose of the present study because of the lack of labeling in the SNc and the VTA. In the two other animals, the injection sites were confined to the same regions of the PPN on both sides of the brain. The locations of these injection sites were deter- mined in relationship to the pars compacta (PPNc) and to the pars dissipata (PPNd) of the PPN (Olszewski and Baxter, 1954; Lavoie and Parent, 1994a). In our material, the extent of these two regions could be determined, because the PPN neurons displayed a slight background staining. The injection sites in the second animal covered the lateral part of the PPNc and PPNd but encroach- ed slightly on the pontine tegmentum (Fig. 1A). In the third animal, the injection sites were located in the central portion of the PPN that included the PPNc and the ventral part of the PPNd (Fig. 1B) without encroachment of adjacent structures. The injection sites consisted of a densely stained core surrounded by a lightly labeled periph- eral zone composed of perikarya and fibers that appeared to be completely filled with PHA-L. These four injections resulted in profuse anterograde labeling in the SNc and VTA.
Anterograde labeling in the SNc and VTA In the four cases retained for the purpose of the present
study, the overall pattern of anterograde labeling in the SNc and the VTA was the same, but the number of PHA-L-labeled fibers was much higher following the injec- tions in the third animal. Numerous PHA-L-labeled fibers emerged from the injection sites to innervate various subcortical regions in a pattern similar to that described recently by Lavoie and Parent (1994b) in the same species. The pattern of anterograde labeling in the SNc and the VTA that resulted from the PPN injections in the third animal is represented schematically in Figure 1C-F. A large number of PHA-L-labeled fibers and swelling-like terminal boutons were found along the rostrocaudal extent of the SNc. The density of anterogradely labeled elements appeared some- what greater in the medial aspect of the SNc (Fig. 1D,E). This pattern of arborization was especially well defined at the level where the SNc merged with the VTA (Fig. ID). The innervation was denser in the dorsaI than in the ventral part of SNc (Fig. 1C-El. Scattered PHA-L-labeled fibers and swelling-like boutons also occurred in the SNr (Fig. 1C-E).
The distribution of PHA-L-immunoreactive fibers and varicosities was uniform along the rostrocaudal extent of the VTA (Fig. lD,E), except that some PHA-L-labeled varicose fibers formed dense plexuses in the ventral part of the VTA near the oculomotor nerve root fibers (Fig. 2A,B). A substantial number of PHA-L-containing fibers also occurred more caudally at level of the retrorubral area (Fig. 1F). These fibers were in continuity with those labeled in
the SNc and the VTA. At different levels throughout the rostrocaudal extent of the ventral midbrain, long, thin, and varicose fibers crossed the midline toward the SNc and the VTA on the contralateral side.
Most of the anterogradely labeled fibers that were seen throughout the rostrocaudal extent of the SNc and the VTA were thin, had short collaterals (Fig. 2D,E), and possessed multiple round varicosities of different sizes separated one from another by intervaricose segments of variable length (Fig. 2D-F). Another type of PHA-L-labeled fibers that occurred less frequently was much thicker and possessed multiple large-sized varicosities (Fig. 2C).
In single labeled sections, the PHA-L-immunoreactive fibers and varicosities were often apposed on the surface of perikarya and dendrites in the SNc and the VTA (Fig. 2G). In double-labeled sections for PHA-L and THICaBP, the PHA-L-labeled elements were easily distinguishable from the THiCaBP-immunoreactive structures by the color of the reaction product associated with them. The dark blue reaction product of Ni-DAB was associated with perikarya and dendrites in the VTA and the SNc, whereas the brown DAB reaction product was localized exclusively in antero- gradely labeled fibers and terminals. At different levels of midbrain, close associations between DAB-labeled varicosi- ties and Ni-DAB-containing dendrites and perikarya were commonly seen.
Electron microscopic observations At the electron microscopic level, the PHA-L-labeled
terminals were characterized by the presence of the elec- tron dense, amorphous DAB reaction product (Figs. 3-5), whereas the TH- and CaBP-immunoreactive elements, which were stained with BDHC, contained electron dense crystals that were distributed randomly in the labeled perikarya and dendritic shafts (Fig. 5A-D). In the gluta- mate- or GABA-immunostained sections, the tissue was overlaid with gold particles that were associated preferen- tially with some neuronal structures (Figs. 3-5).
The PHA-L immunoreactivity occurred in myelinated axons and terminal boutons in the SNc and VTA. The PHA-L-labeled terminals displayed the same ultrastruc- tural features in both regions. Their maximum diameter ranged from 0.9 pm to 2.5 pm, and they contained a large number of round synaptic vesicles as well as mitochondria (Figs. 3-5). In the 34 cases where the synaptic specializa- tion could be seen, it was of the asymmetric type (30 cases) with a few exceptions, which formed symmetric synapses (4 cases). The major postsynaptic targets of these boutons were dendritic shafts of different sizes (maximum diameter ranged from 1-3 pm; Figs. 3-5). In only three cases, the labeled terminals formed synapses with perikarya.
In the SNc, a total of 81 PHA-L-immunoreactive boutons were tested for glutamate and/or GABA immunostaining (Table 1). Forty percent of the terminals tested for gluta- mate immunoreactivity (n = 33) were considered positive, because the density of gold particles associated with these boutons was not significantly different from that overlying neighboring terminals of the same category (Figs. 3, 5). In contrast, the density of gold particles associated with these terminals was three to five times higher than that associ- ated with glial cells in the same sections. The boutons that displayed a low level of immunoreactivity for glutamate were found to be immunoreactive for GABA in adjacent sections (Fig. 3A,B). In the seven cases where the synaptic

Fig. 2. Photomicrographs showing various features of antero- gradely labeled fibers and varicosities a t midbrain level after PHA-L injections in PPN. A: Low-power view of rich plexuses of PHA-L- imniunoreactive varicose fibers in the VTA. The plexus indicated by an arrow is shown a t higher magnification in B. C: An example of a rare type of PHA-L-labeled fibers in the SNc. This type of fiber is much thicker and possesses larger varicosities than the most common type of
anterogradely labeled axons shown in D-F. Note the short collaterals emerging from the main axon in D and E. Note also that the fiber shown in F possesses varicosities of different sizes. G: A neuronal perikaryon in the SNc apposed by PHA-L-immunoreactive varicosities. Scale bars = 300 pm in A, 30 Fm in B, 10 pm in C (also applies to E,G), 10 Fm in D (also applies to F).
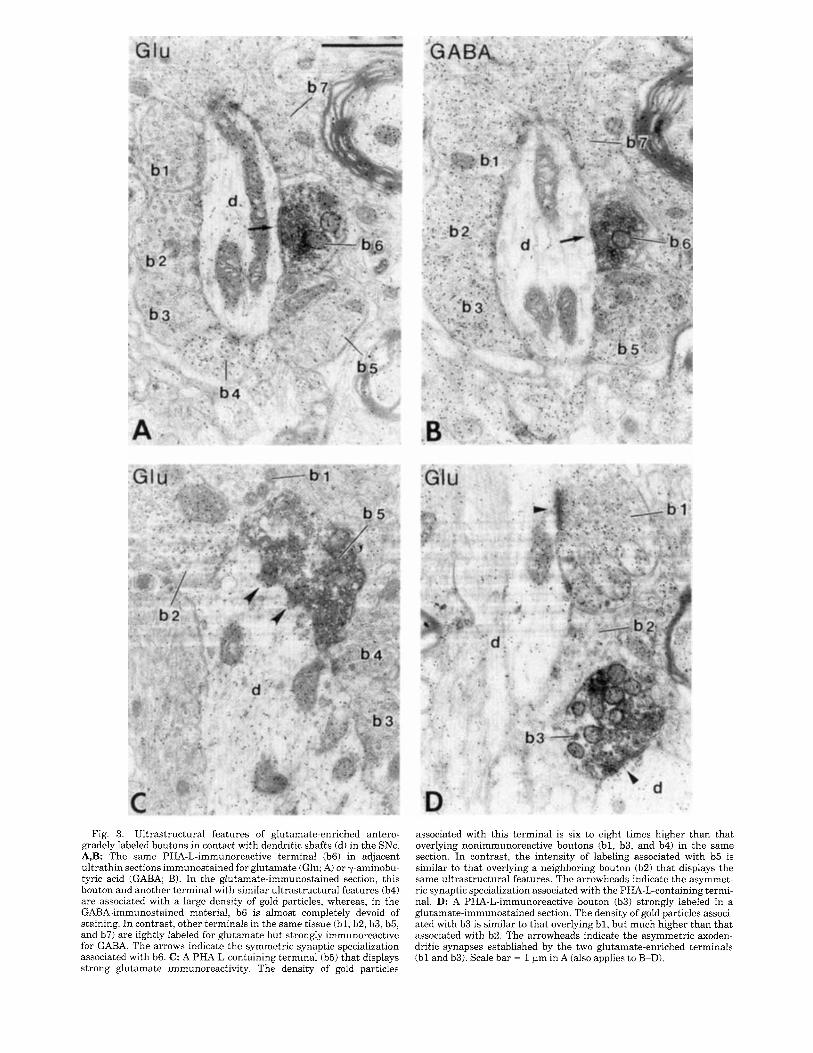
Fig. 3. Ultrastructural features of glutamate-enriched antero- gradely labeled boutons in contact with dendritic shafts (di in the SNc. A,B: The same PHA-L-immunoreactive terminal (b6) in adjacent ultrathin sections immunostained for glutamate (Glu; Ai or y-aminobu- tyric acid (GABA; B). In the glutamate-immunostained section, this bouton and another terminal with similar ultrastructural features (b4) are associated with a large density of gold particles, whereas, in the GABA-immunostained material, b6 is almost completely devoid of staining. In contrast, other terminals in the same tissue (bl , b2, b3, b5, and b7i are lightly labeled for glutamate but strongly immunoreactive for GABA. The arrows indicate the symmetric synaptic specialization associated with b6. C: A PHA-L-containing terminal (b5) that displays strong glutamate immunoreactivity. The density of gold particles
associated with this terminal is six to eight times higher than that overlying nonimmunoreactive boutons (bl , b3, and b4) in the same section. In contrast, the intensity of labeling associated with b5 is similar to that overlying a neighboring bouton (b2) that displays the same ultrastructural features. The arrowheads indicate the asymmet- ric synaptic specialization associated with the PHA-L-containing termi- nal. D: A PHA-L-immunoreactive bouton (b3) strongly labeled in a glutamate-immunostained section. The density of gold particles associ- ated with b3 is similar to that overlying bl, but much higher than that associated with b2. The arrowheads indicate the asymmetric axoden- dritic synapses established by the two glutamate-enriched terminals (b l and b3i. Scale bar = 1 pm in A (also applies to B-D).

PEDUNCULOPONTINE INPUTS TO SUBSTANTIA NIGRA 261
Fig. 4. Ultrastructural features of PHA-L-labeled boutons that strongly immunoreactive for both GABA and glutamate. C,D: Two display GABA immunoreactivity in the VTA. A,B: The same PHA-L- PHA-L-containing boutons ( b l ) that display immunoreactivity for immunoreactive terminal (b3) in a GABA (A) and a glutamate- GABA. In C, note the low density of gold particles associated with b2 immunostained (B) section. This bouton is strongly immunoreactive and b3 in comparison to that overlying b l . In D, b2 also displays GABA for GABA but is lightly labeled for glutamate. In contrast, a neighbor- immunoreactivity. The arrows in A and C indicate symmetric synaptic ing terminal (b2) is nonimmunoreactive for GABA but is strongly specializations. Scale bar = 0.5 pm in A (also applies to B-D). stained for glutamate. Another bouton in the same tissue ( b l ) is
specialization could be seen, six PHA-L-immunoreactive terminals that displayed a strong immunoreactivity for glutamate formed asymmetric synapses (Figs. 3C,D, 5B,C). In only one case, the synapse associated with such a terminal was of the symmetric type (Fig. 3A,B). Three of the 13 elements contacted by glutamate-enriched PHA-L- immunoreactive boutons displayed TH (two cases) or CaBP (one case) immunoreactivity (Fig. 5 ) .
Among the 44 PHA-L-positive terminals that were tested for GABA immunoreactivity, 28% were found to be positive (Fig. 4) . In the GABA-immunostained sections, the density of gold particles associated with these boutons was not significantly different from that overlying other terminals
in the neuropil that formed symmetric synapses (Fig. 4A-C). In the glutamate-immunostained sections, these boutons were almost completely devoid of gold particles (Fig. 4A,B). In the two cases where we could see the synaptic specialization associated with these terminals, it was of the symmetric type (Fig. 4C). However, the ultra- structural features of these boutons were not different from those enriched in glutamate (compare Figs. 3 and 5 to Fig. 4). None of the postsynaptic targets contacted by the PHA-L-positive boutons that displayed GABA immunoreac- tivity were found to be immunoreactive for TH or for CaBP (Fig. 4). Among the four terminals that were tested for glutamate and GABA, three displayed a strong immunore-

Fig. 5. Ultrastructural features and neurotransmitter content of PHA-L-labeled houtons forming asymmetric synaptic contacts (arrow- heads) with tyrosine hydroxylase (TH)-immunoreactive and calbindin D-28k (CaBPj-immunoreactive elements in the SNc (A,B) and VTA (C,D). In B-D, the sections have been processed for glutamate (Glu; B,C) or GABA immunostaining (D). A: A TH-immunoreactive dendrite (d/TH) that receives a synaptic contact from a PHA-L-immunoreactive bouton (b l ) in the SNc. Note the numerous crystals of benzidine dihydrochloride (BDHC) in the TH-positive dendrite. B: A PHA-L- containing bouton (b2) forming an asymmetric synapse with a TH- immunoreactive dendrite (dlTH) in the SNc. This bouton (b2) displays
strong glutamate immunoreactivity. Note the difference in the inten- sity of glutamate-immunostaining between b l (low immunoreactivityj and b2. C: An anterogradely labeled bouton (b l ) in contact with a CaBP-immunoreactive dendrite (d/CaBPj in the VTA. The PHA-L- positive bouton is associated with a large density of gold particles in comparison to another terminal in the neuropil (b2). D: A PHA-L- containing bouton (h2) in contact with a TH-positive dendrite (dlTH) in the VTA. This bouton is not immunoreactive for GABA. Note the larger density of gold particles associated with b l and b3 in comparison to b2. Scale bar = 1 Frn in A (also applies to B-D).

PEDUNCULOPONTINE INPUTS TO SUBSTANTIA NIGRA 263
TABLE 1. Anterogradely Labelled Terminals Tested for Glutamate and/or GABA at Midbrain Level
Number of terminals tested (% of irnmunoreactive) for Total number
Brain of terminals regions Glutamate GABA GlutamateitiABA' tested
SKC 33 140'4 1 44 128'V 1 4 175'h G I . ' / G J 81
W A 33 160'4 1 21 140'4 J 125'4 GI,'/(; 1 58 125'4 GI, / G i
125% GL iG 150% (;L iG'1
1C1. , high GI,LI-IR: GL , low GI,U-IR; G ' , GABA-positive; G , GABA-negative; SNc, suhstantia ni~rra pars cornpacta; VTA, ventral tegrnental area.
activity for glutamate but were nonimmunoreactive for GABA, whereas the other terminal was immunoreactive for GABA but was lightly labeled for glutamate (see Table 1).
In the VTA, a total of 58 terminals were tested for glutamate and/or GABA immunoreactivity (Table 1). Sixty percent of the 33 terminals tested for glutamate were strongly immunoreactive. The ultrastructural features and the type of synaptic specialization associated with these terminals were not different from those visualized in the SNc (compare Fig. 3 to Fig. 5C). In four cases, the postsyn- aptic targets displayed TH (two cases) or CaBP (two cases) immunoreactivity (Fig. 5 ) . Among the 21 PHA-L-positive terminals that were tested for GABA immunoreactivity, 40% were found to be positive. These terminals and those in the SNc shared similar ultrastructural features (Fig. 4). In the four terminals that were tested for glutamate and GABA, two were immunoreactive for GABA but not for glutamate, one displayed a high level of glutamate immuno- reactivity but was nonimmunoreactive for GABA, and one was nonimmunoreactive for glutamate and GABA (see Table 1). None of the postsynaptic targets contacted by GABA-positive terminals displayed TH or CaBP immunore- activity.
DISCUSSION The major finding of the present study is that a signifi-
cant proportion of PPN terminals in the ventral midbrain contain glutamate, and some of them establish asymmetric synapses with dopaminergic neurons in the monkey. In addition, some PPN terminals in the SNc and the VTA display GABA immunoreactivity. These results and their functional significance will be discussed in the light of previous anatomical and electrophysiological observations.
The PPN: A major source of afferents to the SNc and the VTA
The present study provides further evidence that the PPN sends direct projections to the midbrain dopaminergic neurons. This is consistent with the previous observations obtained by means of retrograde and anterograde tracing methods in rodents (Saper and Loewy, 1982; Jackson and Crossman, 1983; Sugimoto and Hattori, 1984; Woolf and Butcher, 1986; Gould at al., 1989) and cats (Nomura et al., 1980; Moon Edley and Graybiel, 1983) indicating that the PPN projections to the SN are more dense in the pars compacta than in the pars reticulata. Our observations also confirm recent findings obtained in the squirrel monkey by Lavoie and Parent (199413) using anterograde tracing meth- ods. In keeping with our data, these authors found that PPN efferent fibers arborize more profusely in the dorsal than in the ventral part of the SNc. In both studies, the
VTA was found to be uniformly innervated except for the rich plexuses of varicosities that were seen in our material. Furthermore, the intimate relationships between PPN fibers and SNc dopaminergic neurons that were previously described (Lavoie and Parent, 1994b) were also seen in the present study. Our findings showed the existence of differ- ent types of fibers labeled in the SNc after injection in the PPN. Although the majority consisted of thin and elongated fibers like those described by Lavoie and Parent (1994b), other thicker fibers with large varicosities were also seen in our material. The possibility that these two types of fibers arise from different populations of cells in the PPN remains to be established.
Glutamate in PPN neurons The results of our study demonstrate that a large propor-
tion of anterogradely labeled terminals in the SNc and the VTA display strong glutamate immunoreactivity, and some of them form asymmetric synaptic contacts predominantly with dendritic shafts of midbrain dopaminergic neurons. In line with the data reported in the companion paper demon- strating that VTA neurons receive a more substantial glutamatergic input than SNc cells (Smith et al., 1996), the proportion of PHA-L-positive terminals that displayed glu- tamate immunoreactivity was higher in the VTA than in the SNc. The glutamatergic innervation of midbrain dopa- minergic cells by PPN afferents has been previously sug- gested by multiple experiments. First, electrophysiological studies have reported that electrical stimulation of the PPN in the rat evokes glutamate-mediated excitation of SNc neurons (Scarnati et al., 1984, 1986, 1987; Di Loreto et al. 1992). Second, ultrastructural studies using the antero- grade degeneration method have shown that PPN termi- nals form asymmetric synaptic contacts with dendrites of SNc neurons in the rat (Scarnati et al., 1988; Tokuno et al., 1988). Third, immunohistochemical studies have shown that a large number of neurons in the PPN exhibit gluta- mate immunoreactivity in the rat (Clements and Grant, 1990) and monkey (Lavoie and Parent, 1994a). Fourth, a combination of retrograde tract-tracing methods with gluta- mate immunohistochemistry has revealed that several ret- rogradely labeled cells in the PPN display glutamate immu- noreactivity after injections in the SN of squirrel monkey (Lavoie and Parent, 1994~) . Taken together, these data support the existence of a glutamatergic projection from the PPN to the midbrain dopaminergic neurons. However, the demonstration of the existence of such a pathway requires the determination of whether PPN terminals in the SNc and the VTA are enriched with glutamate. Because this amino acid is involved in several metabolic pathways that are not related to synaptic transmission (for review, see Fonnum, 1984; Ottersen et al., 1992), the occurrence of glutamate immunoreactivity in cell bodies does not necessar- ily indicate that these neurons use glutamate as a neuro- transmitter. In this respect, our findings provide the first ultrastructural evidence that glutamate-enriched terminals from the PPN form synaptic contacts with dopaminergic neurons in the SNc and the VTA. In general, the ultrastruc- tural features displayed by the anterogradely labeled termi- nals resembled those of the type I glutamate-enriched boutons described in the companion paper (Smith et al., 1996). Although a substantial proportion of PHA-L-labeled terminals formed synapses with nonimmunoreactive struc- tures, this does not necessarily imply that PPN afferents contact predominantly nondopaminergic elements. The

264 A. CHARARA ET AL.
fact that BDHC, which was used to localize TH and CaBP immunoreactivity, does not penetrate the tissue as well a DAB may explain why such a high number of elements contacted by PHA-L-immunoreactive terminals were non- immunoreactive. Of course, the possibility that nondopamin- ergic SNc and VTA neurons (Van der Kooy et al., 1981; Domesick et al., 1983) receive glutamatergic afferents from the PPN must also be considered.
GABA in PPN neurons Our findings also show that a significant number of
PHA-L-labeled boutons displayed GABA immunoreactivity and established contact with dendritic shafts in the SNc and the VTA. To our knowledge, these results provide the first evidence that the PPN is a source of GABAergic afferents to the SNc and the VTA in primates. The results obtained in previous immunohistochemical studies are controversial with regard to the existence of GABAergic cells in the PPN. Although some investigators detected only a few glutamic acid decarboxylase (GAD)-immunoreactive cells in the rat PPN (Mugnaini and Oertel, 1985), other experiments in the same species revealed the existence of a significant number of GAD-immunoreactive neurons in this region (Jones, 1990). Another finding that suggests the existence of GABAergic cells in the PPN is the fact that this nucleus contains a small number of parvalbumin-immuno- reactive cells in monkeys (Cste, Fortin and Parent, unpub- lished observations). Although the occurrence of GABA in these neurons has not been shown, parvalbumin is often associated with GABA-containing neurons in other brain areas (Celio, 1990). These observations indicate that a small proportion of PPN neurons projecting to the mid- brain may use GABA as neurotransmitter in monkeys. The fact that we did not find any PHA-L/GABA-positive bou- tons in contact with TH- or CaBP-positive neurons in the triple-immunostained tissue can be explained by the techni- cal limitations described above for the PHA-L/glutamate- positive terminals.
Possible colocalization of acetylcholine and glutamate in PPN neurons
The PPN is known to contain cells immunoreactive for ChAT (Armstrong et al., 1983; Mesulam et al., 1984; Jones and Beaudet, 1987; Tag0 et al., 1989; Lavoie and Parent, 1994a), calbindin D-28k (C6te and Parent, 1992; Guela et al., 19931, and various neuropeptides (see Vincent et al., 1986). Although some studies have denied the existence of a cholinergic projection from the PPN to the SN in the rat (Sugimoto and Hattori, 1984; Rye et al., 1987; Lee et al., 1988), other experiments carried out by means of retro- grade tract-tracing methods combined with ChAT immuno- cytochemistry have demonstrated that a proportion of cholinergic cells in the PPN project to the SN in different species (Woolf and Butcher, 1986; Beninato and Spencer, 1987; Clarke et al., 1987; Gould et al., 1989; Lavoie and Parent, 1994~). These observations are supported by the existence of ChAT-immunoreactive terminals in the sub- stantia nigra of primates (Mesulam et al., 1992) and nonprimates (Gould and Butcher, 1986; Henderson and Greenfield, 1987; Martinez-Murillo et al., 1989a,b; Bolam et al., 1991). Moreover, CUT-positive terminals have been found to form asymmetric synapses with TH-immunoreac- tive dendrites and somata in the SNc of the ferret (Bolam et al., 1991). The ChAT-immunoreactive terminals displayed ultrastructural features similar to those of the antero-
gradely labeled boutons seen in the present study. These data combined with data supporting the coexistence of glutamate and ChAT at the level of single neuronal peri- karya in the PPN (Lavoie and Parent, 1994a) open the possibility that single PPN terminals in contact with mid- brain dopaminergic neurons contain both glutamate and acetylcholine.
Functional considerations and potential clinical relevance
The dopaminergic nigrostriatal and mesolimbic path- ways are implicated in the control of voluntary movements, cognition, and emotional behavior. It is well established that extrinsic afferents, especially those using excitatory amino acids, play a major role in the generation of burst firing in midbrain dopaminergic neurons in the rat (for references, see Smith et al., 1996). The fact that glutamate- enriched terminals from the PPN form synaptic contacts with SNc and VTA neurons suggests that the PPN may be involved in the control of the firing pattern of midbrain dopaminergic neurons in primates. However, the mecha- nism by which the PPN influences the activity of midbrain dopaminergic cells has not been elucidated. Excitatory amino acids are believed to act at midbrain level on N-methyl-D-aspartate (NMDA), a-amino-3-hydroxy-5- methyl-4-isoxazoleproprionate (AMPA), and metabotropic glutamate receptors (Seutin et al., 1990; Petralia and Wenthold, 1992; Wang and French, 1993; Testa et al., 1994). Recent data obtained by means of iontophoretic application of excitatory amino acids and their antagonists combined with stimulation of the PPN have shown that non-NMDA receptors are involved in the excitatory effect generated in nigrostriatal dopaminergic neurons after PPN activation (Di Loreto et al., 1992).
The companion to this paper (Smith et al., 1996) dis- cusses in detail a series of findings indicating that glutama- tergic afferents may be involved in the degeneration of midbrain dopaminergic neurons in Parkinson’s disease. Although the exact mechanism by which glutamatergic afferents may become cytotoxic for dopaminergic neurons remains to be established, it is likely that activation of NMDA receptors is involved in the degenerative process. It is well established that the channel associated with NMDA receptor is highly permeable to Ca2+ (for review, see Nakanishi, 1992). An overstimulation of these receptors by glutamatergic afferents may therefore induce a rapid rise in the intracellular concentration of Ca2+, which ultimately becomes lethal and leads to neuronal death. The glutamater- gic afferents to SNc neurons that arise from the cerebral cortex and from the subthalamic nucleus activate NMDA receptors, as mentioned above, whereas those arising from the PPN generate their excitatory influences via the activa- tion of non-NMDA receptors. Therefore, a dysfunction in cortical or subthalamic afferents, but not necessarily in afferents arising from the PPN, may become excitotoxic for midbrain dopaminergic neurons. However, because NMDA and non-NMDA receptors closely interact to control the activity of midbrain dopaminergic neurons (Mereu et al., 1991), the possibility that PPN afferents participate in the degenerative process must also be considered.
ACKNOWLEDGMENTS The authors thank Carole Emond, Lisette Bertrand, and
Jean-FranGois Pare for technical assistance as well as

PEDUNCULOPONTINE INPUTS TO SUBSTANTIA NIGRA 265
Isabelle Deaudelin for the art work. We also thank Dr. Peter Somogyi for the generous gift of the GABA antise- rum. This research was supported by grants from the Medical Research Council of Canada to A.P. and from the Canadian Foundation of Parkinson's Disease to Y.S. A.C. holds a studentship from the FCAR.
LITERATURE CITED Armstrong, D.M., C.B. Saper, A.I. Levey, B.H. Wainer, and R.D. Terry
11983) Distribution of cholinergic neurons in rat brain demonstrated by the immunocytochemical localization of choline acetyltransferase. J. Comp. Neurol. 216.53-68.
Beninato. M., and R.F. Spencer 11987) A cholinergic projection to the rat substantia nigra from the pedunculopontine tegmental nucleus. Brain Res. 412:169-174.
Bolarn. J.P., C.M. Francis, and 2. Henderson (1991) Cholinergic input to dopaminergic neurons in the substantia nigra: A double immunohisto- chemical study. Neuroscience 41r483-494.
Celio. M.R. (1990) Calbindin D-28k and parvalbumin in the rat nervous system. Neuroscience 35,375-475.
Charara, A,, Y. Smith, and A. Parent (1993) Glutamatergic inputs from the pedunculopontine nucleus to the midbrain dopaminergic neurons in primates: A light and electron microscopic study. Sac. Neurosci. Abstr. 191437.
Clarke. P.B., D.W. Hommer, A. Pert, and L R . Skirboll(1987) Innervation of the substantia nigra neurons by cholinergic afferents from the peduncu- lopontine nucleus in the rat: Neuroanatomical and electrophysiological evidence. Neuroscience 23:1011-1019.
Clements, J .R., and S. Grant ( 1990) Glutamate-like immunoreactivity in neurons of the laterodorsal tegmental and pedunculopontine nuclei in the rat. Neurosci. Lett. 120r70-73.
CBte. P.Y.. and A. Parent (19921 Calbindin D-28k and choline acetyltransfer- ase are expressed by different neuronal populations in pedunculopontine nucleus but not in nucleus basalis in the squirrel monkey. Brain Res. 593;245-252.
Di Loreto. S., T. Florio, and E. Scarnati 119921 Evidence that non-NMDA receptors are involved in the excitatory pathway from the pedunculopon- tine reDon to nigrostriatal dopaminergic neurons. Exp. Brain Res. 89:79-86.
Domesick, V.B., L. Stinus, and P.A. Paskevich (1983) The cytology of dopdminerpc and nondopaminergic neurons in the substantia nigra and ventral tegnental area of the rat: A light- and electron-microscopic study. Neuroscience 8.743-765.
Emmers. R., and K. Akert (1963) A Stereotaxic Atlas of the Brain of the Squirrel Monkey (Saimiri sciureus). Madison, WI: University of Wiscon- sin Press.
Fonnum. F. (1984) Glutamate: A transmitter in mammalian brain. J. Neurochem. 4 2 - 1 1 .
Gariano, R.F., and P.M. Groves 11988) Burst firing induced in midbrain dopamine neurons by stimulation of the medial prefrontal and anterior cinhwlate cortices. Brain Res. 462: 194-198.
Gaspar, P.. C.W. Heizmann, and J.H. Kaas (19931 Calbindin D-28K in the dopaminergic mesocortical projection of a monkey (Aotus trzuzrgatusi. Brain Res. 603166-172.
Gerfen. C.K.. and P.1:. Sawchenko (1984) An anterograde neuroanatomical tracing method that shows the detailed morphology of neurons, their axons and terminals: Immunohistochemical localization of an axonally transported plant lectin, Phaseolus uulgaris-leucoagglutinin IPHA-L). Brain Res. 290r219-238.
Gould, E., and L.L. Butcher 119861 Cholinergic neurons in the rat substantia n i p x Neurosci. Lett. 63.315-319.
Gould, E., N . J . Woolf, and L.L Butcher 11989) Cholinergic projections to the substantia nigra from the pedunculopontine and laterodorsal tegmental nuclei. Neuroscience 28r611-623.
Grace, A.A., and B.S. Bunney (1984) The control of firing pattern in nigral dopamine neurons: Burst firing. J . Neurosci. 42877-2890.
Grace. A.A., and S.P. Onn (1989) Morphology and electrophysiological properties of immunocytochemically identified rat dopamine neurons recorded in vitro. J. Neurosci. 9:3463-3481.
Guela, C . , C.R. Scatz, and M.-M. Mesulam (1993) Differential localization of NADPH-diaphorase and calbindin D-28k within the cholinergic neurons of the basal forebrain, striatum and brainstem in the rat , monkey, baboon and human. Neuroscience 54:461476.
Hammond, C., J.M. Deniau, A. Rizk, and J. Feger (1978) Electrophysiologi- cal demonstration of an excitatory subthalamonigral pathway in the rat . Brain Res. 151:235-244.
Henderson, Z., and S.A. Greenfield (1987) Does the substantia nigra have a cholinergic innervation? Neurosci. Lett. 73:109-113.
Hepler, J.R., C.S. Toomim, K.D. McCarthy, F. Conti, G. Battaglia, A. Rustioni, and P. Petrusz (1988) Characterization of antisera to gluta- mate and aspartate. J . Histochem. Cytochem. 36.13-22.
Hodgson, A.J., B. Penke, A. Erdei, I.W. Chubb, and P. Somogyi (1985) Antisera to y-aminobutyric acid. I. Production and characterization using a new model system. J. Histochem. Cytochem. 33229-239.
Hsu, S.M., L. Raine, and H. Fanger (1981) Use of avidin-biotin-peroxidase complex (ABCl in immunoperoxidase techniques: A comparison between ABC and unlabelled antibody (PAP) procedures. J. Histochem. Cyto- chem. 21:557-580.
Jackson, A., and A.K. Crossman 11983) Nucleus tegmenti pedunculoponti- nus: Efferent connections with special reference to the basal ganglia, studied in the rat by anterograde and retrograde transport of horserad- ish peroxidase. Neuroscience 10:725-765.
Johnson, S.W., V. Seutin, and R.A. North 119921 Burst firing in dopamine neurons induced by N-methyl-D-aspartate: Role of electrogenic sodium pump. Science 258.665-667.
Jones, B.E. (1990) Relationship of GABAergc to cholinergic neurons within the laterodorsal and pedunculopontine tegmental nuclei. Soc. Neurosci. Abstr. 16:300.
Jones, B.E., and A. Beaudet (1987) Distribution of acetylcholine and catecholamine neurons in the cat brainstem: A choline acetyltransferase and tyrosine hydroxylase immunocytochemical study. J. Comp. Neurol. 261: 15-32.
Kita, H. , and S.T. Kitai (19871 Efferent projections of the subthalamic nucleus in the rat: Light and electron microscopic analysis with the PHA-L method. J. Comp. Neural. 260r435-452.
Lavoie, B., and A. Parent (1994a) The pedunculopontine nucleus in the squirrel monkey. Distribution of cholinergic and monoaminergic neu- rons in the mesopontine tegmentum with evidence for the presence of glutamate in cholinergic neurons. J. Comp. Neurol. 344:190-209.
Lavoie, B., and A. Parent (1994bi Pedunculopontine nucleus in the squirrel monkey: Projections to the basal ganglia as revealed by anterograde tract-tracing methods. J. Comp. Neurol. 344:210-231.
Lavoie, B., and A. Parent ( 1 9 9 4 ~ ) Pedunculopontine nucleus in the squirrel monkey: Cholinergic and glutamatergic projections to the substantia nigra. J . Comp. Neurol. 344.232-241.
Lee, H.J., D.B. Rye, A.E. Hallanger, A.I. Levey, and B.H. Wainer (19881 Cholinergic vs. noncholinergic efferents from the mesopontine tegmen- tum to the extrapyramidal motor system nuclei. J . Comp. Neurol. 275:469492.
Levey, A.I.. J .P. Bolam, D.B. Rye, A.E. Hallanger, R.M. Demuth, M.-M. Mesulam, and B.H. Wainer (1986) A light and electron microscopic procedure for sequential double antigen localization using diaminobenzi- dine and benzidine dihydrochloride. J. Histochem. Cytochem. 34:1449- 1457.
Martinez-Murillo, R., R.M. Villaba, M.I. Montero-Caballero. and J. Rodrigo (1989a) Cholinergic somata and terminals in the rat substantia nigra: An immunohistochemical study with optical and electron microscopic tech- niques. J . Comp. Neurol. 281397416 .
Martinez-Murillo. R., R.M. Villaba, and J. Rodrigo (1989bi Electron micro- scopic localization of' cholinergic terminals in the rat substantia nigra: An immunocytochemical study. Neurosci. Lett. 96:121-126.
Mereu, G., E. Costa, D.M. Armstrong, and S. Vicini 11991) Glutamate receptor subtypes mediate excitatory synaptic currents of dopamine neurons in midbrain slices. J. Neurosci. 11:1359-1366.
Mesulam, M.-M., E.J. Mufson, A.I. Levey, and B.H. Wainer (1984) Atlas of cholinergic neurons in the forebrain and upper brainstem of the macaque based on monoclonal choline acetyltransferase immunohisto- chemistry and acetylcholinesterasc histochemistry. Neuroscience 12:669- 686.
Mesulam, M.-M., D. Mash, L. Hersh, M. Bothwell, and C. Guela (1992) Cholinergic innervation of the human striatum, globus pallidus, subtha- lamic nucleus, substantia nigra, and red nucleus. J . Comp. Neurol. 323:252-268.
Moon Edley, S., and A.M. Graybiel (1983) The afferent and efferent connections of the feline nucleus tegmenti pedunculopontinus, pars compacta. J. Comp. Neurol. 21 7.187-215.
Mugnaini, E.. and W.H. Oertel (1985) An atlas of the distribution of GABAerbic neurons and terminals in the rat CNS by GAD immunohisto-

266 A. CHARARA ET AL.
chemistry. In A. Bjorklund and T. Hokfelt (eds.): Handbook of Chemical Neuroanatomy, Vol. 4. Amsterdam: Elsevier, pp. 436-608.
Nakanishi, S. (1992) Molecular diversity of glutamate receptors and implica- tions for brain function. Science 258t597-603.
Nomura, S., N. Mizuno, and T. Sugimoto (1980) Direct projections from the pedunculopontine tegmental nucleus to the subthalamic nucleus in the cat. Brain Res. 196.223-227.
Olszewski, J., and D. Baxter (1954) Cytoarchitecture of The Human Brain Stem. New York: S. Karger.
Ottersen, O.P., N. Zhang, and F. Walberg (1992) Metabolic compartmenta- tion of glutamate and glutamine: Morphological evidence obtained by quantitative immunocytochemistry in rat cerebellum. Neuroscience 46.519-534.
Overton, P., and D. Clark (1992) Iontophoretically administered drugs acting at the N-methyl-D-aspartate receptor modulate burst firing in A9 dopamine neurons in the rat. Synapse I0:131-140.
Petralia, R.S., and R.J. Wenthold (1992) Light and electron immunocyto- chemical localization of AMPA-selective glutamate receptors in the rat brain. J. Comp. Neurol. 318.329-354.
Reynold, E.S. (1963) The use of lead citrate a t high pH as electron opaque stain in electron microscopy. 3. Cell. Biol. 17:208-212.
Rogers, J.H. (1992) Immunohistochemical markers in rat brain: Colocaliza- tion of calretinin and calbindin D-28k with tyrosine hydroxylase. Brain Res .58 7t203-2 10.
Rye, D.B., C.B. Saper, H.J. Lee, and B.H. Wainer (1987) Pedunculopontine tegmental nucleus of the rat: Cytoarchitecture, cytochemistry, and some extrapyramidal connections of the mesopontine tegmentum. J. Comp. Neurol. 259t483-528.
Sakai, S.T. (1988) Corticonigral projections from area 6 in the raccoon. Exp. Brain Res. 73:498-504.
Saper, C.B., and A.D. Loewy (1982) Projections of the pedunculopontine tegmental nucleus in the rat: Evidence for additional extrapyramidal circuitry. Brain Res. 252.367-372.
Scarnati, E., A. Proia, E. Campana, and C. Pacitti (1984) Pedunculopontine evoked excitation of substantia nigra neurons in the rat. Brain Res. 304t35 1-36 1.
Scarnati, E., A. Proia, E. Campana, and C. Pacitti (1986) A microiontopho- retic study of the nature of the putative synaptic neurotransmitter involved in the pedunculopontine-substantia nigra pars compacta path- way of the rat. Exp. Brain Res. 62.470-478.
Scarnati, E., A. Proia, S. Di Loreto, and C. Pacitti (1987) The reciprocal electrophysiological influence between the nucleus tegmenti pedunculo- pontinus and the substantia nigra in normal and decorticed rats. Brain Res. 423:116-124.
Scarnati, E., F. Hadju, C. Pacitti, and T. Tombol (1988) An electron microscope and Golgi study on the connection between the nucleus tegmenti pedunculopontinus and the pars compacta of the substantia nigra in the rat. J. Hirnforsch. 29.95-105.
Sesack, S.R., and V.M. Pickel (1992) Prefrontal cortical efferents in the rat synapse on unlabeled neuronal targets of catecholamine terminals in the nucleus accumbens septi and on dopamine neurons in the ventral tegmental area. J. Comp. Neurol. 320:145-160.
Sesack, S.R., A.Y. Deutch, R.H. Roth, and B.S. Bunney (1989) Topographical organization of the efferent projections of the medial prefrontal cortex in the rat: An anterograde tract-tracing study with Phuseolus uulguris- leucoagglutinin. J. Comp. Neurol. 290:213-242.
Seutin, V., P. Verbanck, L. Massote, and A. Dresse (1990) Evidence for the presence of N-methyl-D-aspartate receptors in the ventral tegmental area of the rat: An electrophysiological in vitro study. Brain Res. 514:147-150.
Smith, I.D., and A.A. Grace (1992) Role of the subthalamic nucleus in the regulation of nigral dopamine neuron activity. Synapse 12,287-303.
Smith, Y., L.-N. Hazrati, and A. Parent (1990) Efferent projections of the subthalamic nucleus in the squirrel monkey as studied by the PHA-L anterograde tracing method. J. Comp. Neurol. 294.306-323.
Smith, Y., A. Charara, and A. Parent (1994) Differential synaptic innerva- tion of dopaminergic neurones in the ventral tegmental area and the substantia nigra pars compacta by glutamate-containing terminals in squirrel monkeys. Movement Disorders 9:SllO.
Smith, Y., A. Charara, M. Paquet, and A. Parent (1995) Glutamatergic innervation of midbrain dopaminergx neurones in monkeys. Brain Res. Assoc. Abstr. I2t38.
Smith, Y., A. Charara, and A. Parent (1996) Synaptic innervation of midbrain dopaminergic neurones by glutamate-enriched terminals in the squirrel monkey. J. Comp. Neurol. (this issue).
Somogyi, P., and A.J. Hodgson (1985) Antisera to y-aminobutyric acid. Demonstration of GABA in Golgi-impregnated neurons and in conven- tional electron microscopic sections of cat striate cortex. J. Histochem. Cytochem. 33:249-257.
Somogyi, P., A.J. Hodgson, I.W. Chubb, B. Penke, and A. Erdei (1985) Antisera to y-aminobutyric acid. 11. Immunocytochemical application to the central nervous system. J. Histochem. Cytochem. 33,240-248.
Suaud-Chagny, M.F., K. Chergui, G. Chouvet, and F. Gonon (1992) Relation- ship between dopamine release in the rat nucleus accumbens and the discharge activity of dopaminergic neurons during local in vivo applica- tion of amino acids in the ventral tegmental area. Neuroscience49.63-72.
Sugimoto, T., and T. Hattori (1984) Organization and efferent projections of nucleus tegmenti pedunculopontinus pars compacta with special refer- ence to its cholinergic aspects. Neuroscience 11.931-946.
Tago, H., P.L. McGeer, E.G. McGeer, H. Akiyama, and L.B. Hersh (1989) Distribution of choline acetyltransferase immunopositive structures in the rat brainstem. Brain Res. 495t271-297.
Testa, C.M., D.G. Standaert, A.B. Young, and J.B. Penney (1994) Metabo- tropic glutamate receptor mRNA expression in the basal ganglia of the rat. J. Neurosci. 14t3005-3018.
Tokuno, H., K.V. Moriizumi, M. Kudo, and Y. Nakamura (1988) A morpho- logical evidence for monosynaptic projections from the nucleus tegmenti pedunculopontinus pars compacta (TPC) to the nigrostriatal projection neurons. Neurosci. Lett. 85: 1-4.
Van der Kooy, D., D.V. Coscina, and T. Hattori (1981) Is there a nondopamin- ergic nigrostriatal pathway? Neuroscience 6:345-357.
Vincent, S.R., K. Satoh, D.M. Armstrong, P. Panula, W. Vale, and H.C. Fibiger (1986) Neuropeptides and NADPH-diaphorase activity in the ascending cholinergic reticular system of the rat. Neuroscience 17.167-182.
Wang, T., and E.D. French (1993) Electrophysiological evidence for the existence of NMDA and non-NMDA receptors on rat ventral tegmental dopamine neurons. Synapse 13t270-277.
Woolf, N.J., and L.L. Butcher (1986) Cholinergic systems in the rat brain: 111. Projections from the pontomesencephalic tegmentum to the thala- mus, tectum, basal ganglia, and basal forebrain. Brain Res. Bull. 16.603-637.





























