Embed Size (px)
Citation preview

, . 183: 359–368 (1997)
GLOMERULAR PROLIFERATING CELL KINETICSIN ACUTE POST-STREPTOCOCCALGLOMERULONEPHRITIS (APSGN)
1*, 2, 2, 3, 3, 3, 3, 1, 4 4
1Department of Medicine, Sendai Hospital National Defence Agency, Sendai, Japan2Department of Public Health, National Defence Medical College, Saitama, Japan3Department of Medicine, National Defence Medical College, Saitama, Japan4Department of Nephrology, Sendai Shakaihoken Hospital, Sendai, Japan
SUMMARY
To investigate the time sequence of glomerular cell proliferation in acute human glomerulonephritis, renal biopsy tissues wereexamined from 15 acute post-streptococcal glomerulonephritis (APSGN) patients (who were biopsied 1–31 days after onset), using animmunoperoxidase technique with monoclonal antibodies against proliferating cell nuclear antigen (PCNA) and various cell surfacemarkers. Few, if any, PCNA+ cells were observed in normal glomeruli, but many cells were positive for PCNA in the acute phase ofAPSGN. Glomerular PCNA+ cells were observed either within glomerular tufts, or lining Bowman’s capsule (parietal epithelial cells);the number of positive cells tended to decrease exponentially as the disease duration increased (r="0·91, P<0·0001). PCNA+ cellswithin glomerular tufts were further identified by double immunostaining. PCNA was not found in PMN or T cells, but a smallproportion of macrophages were PCNA+. Most of the remaining PCNA+ cells were resident glomerular cells; the proportion ofPCNA+ endothelial cells (CD31+) was over 80 per cent in the early phase, but as the disease continued the proportion of mesangial cells(á-smooth muscle actin+) increased to about half of the total PCNA+ cells within the tuft. These data indicate that the hypercellularglomeruli in APSGN are due not only to immune cell infiltration, but also to resident glomerular cell proliferation, probably induced bylocally produced growth factors. ? 1997 John Wiley & Sons, Ltd.
J. Pathol. 183: 359–368, 1997.No. of Figures: 6. No. of Tables: 2. No. of References: 34.
KEY WORDS—acute post-streptococcal glomerulonephritis; glomerular cell proliferation; PCNA; immune cell infiltration
INTRODUCTION
Glomerular cell proliferation is a hallmark of varioustypes of human and experimental glomerulonephritis(GN). The proliferation may be related to the develop-ment of glomerulosclerosis,1–3 which is characterized bythe glomerular accumulation of extracellular matrix andis a common feature of end-stage glomerular injury.Analysis of cell proliferation is therefore of great import-ance in understanding the mechanisms of glomerularinjury. The phenomenon of proliferating cells in exper-imental GN has been extensively studied,4–8 but fewdata about the time course of glomerular cell prolifer-ation in human GN are available. One reason is that inmost human cases of GN, the precise point of diseaseonset is difficult to establish.Acute post-streptococcal glomerulonephritis
(APSGN) is presumed to be induced by the glomerulardeposition of nephritogenic streptococcal antigen–antibody complexes,9 and it is one of the few types ofhuman GN whose onset and time course are obvious. Inan earlier study,10 we demonstrated the glomerularaccumulation of prominent neutrophils and macro-
phages (Mö) and a small but significant number ofCD4+ T cells, and showed a significant positive linearcorrelation between the numbers of infiltrating CD4+ Tcells and Mö in APSGN. We therefore postulated that itis not only humoral immunity, such as type III hyper-sensitivity, but also cell-mediated immunity, especiallymechanisms such as delayed type hypersensitivity(DTH), which may be involved in the glomerular mani-festations of this disease. The induction of capillaryendothelial proliferation in a classical cutaneousDTH reaction has been reported.11 Little is known,however, about the contribution, if any, of endogenousglomerular cell proliferation to the hypercellularity,12 orabout which cell types are involved in the proliferationin APSGN. In the present study, we utilized variousmonoclonal antibodies and immunohistochemicaldouble-staining techniques to examine the time courseof the number and the phenotypes of glomerularproliferating cells in renal biopsy tissues from APSGNpatients.
MATERIALS AND METHODS
PatientsRenal biopsies collected from 15 patients with
APSGN were used in this study. All patients had
*Correspondence to: Takashi Oda, MD, Department of Medicine,Sendai Hospital National Defence Agency, 1-1 Minamimetate,Miyagino-ku, Sendai, Miyagi 983, Japan.
CCC 0022–3417/97/110359–10 $17.50 Received 14 October 1996? 1997 John Wiley & Sons, Ltd. Accepted 29 May 1997
overt symptoms of APSGN, such as facial edema,hypertension, and hematuria. To rule out progressiverenal diseases with acute nephritic syndrome, such asIgA nephropathy with crescents, lupus nephritis, orrapidly progressive GN, percutaneous needle biopsieswere performed for diagnostic purposes over a periodof about 7 years (1979–1986) in the Department ofMedicine, National Defence Medical College (Satama,Japan). The profiles of the 15 patients are shownin Table I. A diagnosis of APSGN was made on thebasis of serological and bacteriological evidence ofacute streptococcal infection before the onset ofnephritis, as well as from the characteristic histologicalfeatures of the renal tissue by light microscopy, immuno-fluorescence, and electron microscopy. The durationof the disease before renal biopsy ranged from 1 to 31days. Normal renal tissues from 10 kidneys thathad been removed for localized tumours served ascontrols.
Monoclonal and polyclonal antibodies
Details of the six monoclonal antibodies (MAbs) andone polyclonal antibody (all purchased from DAKO,Japan, Co. Ltd.) used in this study are shown in Table
II. PC-10 (proliferating cell nuclear antigen: PCNA/cyclin) served as a marker for cell proliferation. PG-M1(CD68), UCHL-1 (CD45RO), and 4KB5 (CD45RA)MAbs served as markers for Mö, memory T cells, andnaive T cells, respectively. JC/70A (CD31) MAb andanti-Factor VIII (VWF) polyclonal antibody served asmarkers for endothelial cells, and 1A4 (anti-á-smoothmuscle actin: á-SMA) MAb was used as a marker forphenotypically changed mesangial cells.13 Among these,PC-10, PG-M1, and 1A4 are ‘Enhanced Polymer One-Step Staining’ (EPOS, recently available from DAKO,Copenhagen, Denmark) reagents that are formed byMAb and horseradish peroxidase coupled to an inertpolymer backbone.
Immunohistochemical and enzyme histochemical staining
For immunohistochemical staining, we used boththe previously described classical indirect immuno-peroxidase method14 and the modified direct immuno-peroxidase (EPOS) method.15,16 Briefly, sections (3 ìmthick) of formalin-fixed, paraffin-embedded tissue werecollected on silane-coated (to improve adhesion) glassslides, deparaffinized in xylene, and rehydrated throughgraded ethanols and distilled water. After blocking
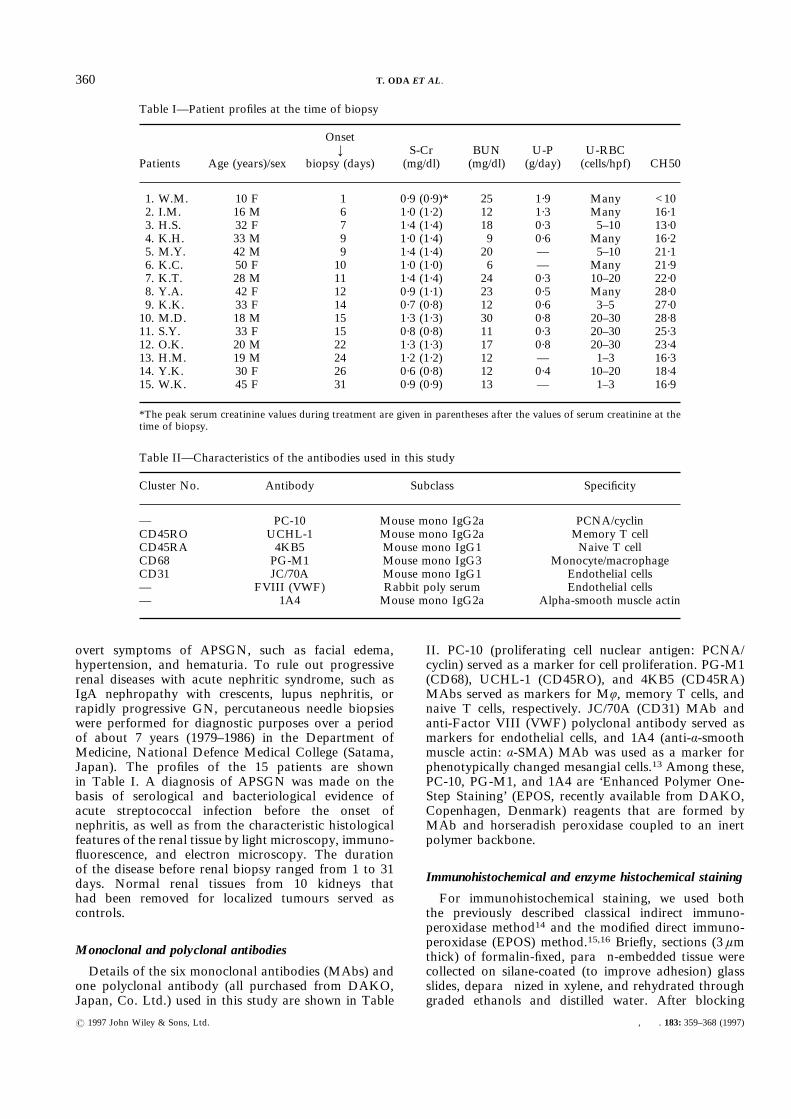
Table I—Patient profiles at the time of biopsy
Patients Age (years)/sex
Onset`
biopsy (days)S-Cr(mg/dl)
BUN(mg/dl)
U-P(g/day)
U-RBC(cells/hpf) CH50
1. W.M. 10 F 1 0·9 (0·9)* 25 1·9 Many <102. I.M. 16 M 6 1·0 (1·2) 12 1·3 Many 16·13. H.S. 32 F 7 1·4 (1·4) 18 0·3 5–10 13·04. K.H. 33 M 9 1·0 (1·4) 9 0·6 Many 16·25. M.Y. 42 M 9 1·4 (1·4) 20 — 5–10 21·16. K.C. 50 F 10 1·0 (1·0) 6 — Many 21·97. K.T. 28 M 11 1·4 (1·4) 24 0·3 10–20 22·08. Y.A. 42 F 12 0·9 (1·1) 23 0·5 Many 28·09. K.K. 33 F 14 0·7 (0·8) 12 0·6 3–5 27·010. M.D. 18 M 15 1·3 (1·3) 30 0·8 20–30 28·811. S.Y. 33 F 15 0·8 (0·8) 11 0·3 20–30 25·312. O.K. 20 M 22 1·3 (1·3) 17 0·8 20–30 23·413. H.M. 19 M 24 1·2 (1·2) 12 — 1–3 16·314. Y.K. 30 F 26 0·6 (0·8) 12 0·4 10–20 18·415. W.K. 45 F 31 0·9 (0·9) 13 — 1–3 16·9
*The peak serum creatinine values during treatment are given in parentheses after the values of serum creatinine at thetime of biopsy.
Table II—Characteristics of the antibodies used in this study
Cluster No. Antibody Subclass Specificity
— PC-10 Mouse mono IgG2a PCNA/cyclinCD45RO UCHL-1 Mouse mono IgG2a Memory T cellCD45RA 4KB5 Mouse mono IgG1 Naive T cellCD68 PG-M1 Mouse mono IgG3 Monocyte/macrophageCD31 JC/70A Mouse mono IgG1 Endothelial cells— FVIII (VWF) Rabbit poly serum Endothelial cells— 1A4 Mouse mono IgG2a Alpha-smooth muscle actin
360 T. ODA ET AL.
? 1997 John Wiley & Sons, Ltd. , . 183: 359–368 (1997)

endogenous peroxidase activity by incubating for 10 minin 3 per cent hydrogen peroxide (H2O2), the sectionswere allowed to react with EPOS reagents for 1 h atroom temperature. Colour development was performedin a solution of 0·02 per cent 3,3*-diaminobenzidine(DAB) and 0·005 per cent H2O2 in 50 mmol/l Tris (pH7·6), and sections were counterstained with methylgreen. We did not use any pretreatment, such as micro-wave irradiation, which is reported to improve thesensitivity of PCNA staining.Double immunohistochemical labelling was per-
formed by repeating the EPOS staining (in PCNA andMö or á-SMA), or by carrying out EPOS stainingand indirect immunoalkaline phosphatase staining (inPCNA and CD31) or indirect immunoperoxidase stain-ing (in PCNA and T cell) sequentially. The labelling isnuclear in PCNA staining, while it is cytoplasmic or cellsurface in cell-specific markers; therefore cross-reactivityis not a problem. In EPOS–EPOS staining or EPOS–immunoperoxidase staining, the first EPOS was devel-oped with DAB enhanced with NiCl2 (black) and thesecond EPOS or immunoperoxidase was developed with3-amino-9-ethylcarbazole (AEC) (red) without counter-stain. In EPOS–immunoalkaline phosphatase staining,the first EPOS staining was performed as describedabove and developed with DAB (brown). This wasfollowed by an indirect immunoalkaline phosphatasestaining procedure, which consisted of three washes inphosphate-buffered saline (PBS) after which the sectionswere allowed to react with appropriate dilutions ofthe primary antibodies for 2 h. Thereafter, they wereincubated for 1 h with alkaline phosphatase-conjugatedsecond-step antibodies, i.e., rabbit anti-mouse IgG orswine anti-rabbit IgG (DAKO, Copenhagen, Denmark),according to the source of the primary antibodies. Thecolour development of the alkaline phosphatase reactionwas carried out using the technique described by Cordellet al.17 When naphthol AS-bi-phosphate and Fast BlueBB salt were used as a substrate, the reaction productswere blue.In addition to immunohistochemical techniques, we
also used an enzyme histochemical technique, chloro-esterase staining, to detect PMN; the procedure has beendescribed by Li et al.18 The cytoplasm of PMN stainsblue when naphthol AS-D chloroacetate and Fast BlueBB salt are used as a substrate. Double staining forPCNA and PMN was performed by carrying out EPOSstaining and chloroesterase staining sequentially.
Quantitation
Although the immunoreactivity of PCNA differedconsiderably in intensity between cells, we judged onlypositive or negative in comparison with backgroundstaining. The total number of PCNA+ cells was countedin each glomerulus and expressed in terms of the numberof positive cells per glomerular cross-section (cells/gcs).In double immunostaining, we calculated the absolutenumber of each double positive glomerular cells/gcs, andthe percentage of each double positive cells in totalPCNA+ cells within the tuft. The number of glomeruliin the biopsies varied from 3 to 27 (average 11).
Statistical analysis
Data are expressed as mean&SD. Student’s t-test wasused to evaluate differences between means. Differenceswere considered significant if P¡0·05.
RESULTS
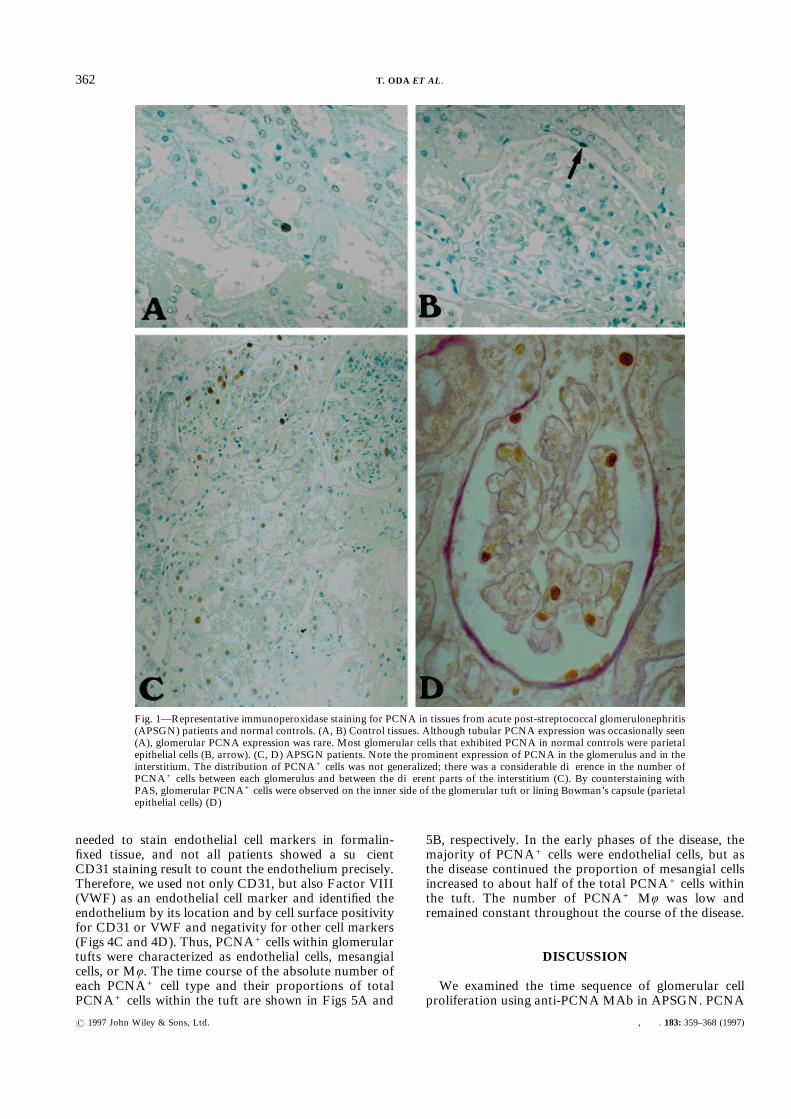
Immunohistochemically, few or no PCNA+ cells werefound in the glomeruli of normal controls, althoughtubular PCNA+ cells were occasionally seen (Fig. 1A).Most glomerular cells that exhibited PCNA in thenormal control were parietal epithelial cells (Fig. 1B).On the other hand, many cells in the glomeruli ofAPSGN patients were positive for PCNA in the earlyphases of the disease (Fig. 1C). The distribution ofglomerular PCNA+ cells was not generalized; glomerulidiffered greatly with respect to the number of PCNA+
cells present. The distribution of tubular PCNA+ cellswas similar, i.e., tubular expression of PCNA wasfocally seen near glomeruli that had many PCNA+ cells,but occasional cells were positive for PCNA in thetubules of other areas. By counterstaining with PAS,glomerular PCNA+ cells were observed either on theinner side of the glomerular tuft or on the lining ofBowman’s capsule (parietal epithelial cells). (Fig. 1D).No visceral epithelial cell was positive for PCNA. Thenumber of labelled cells decreased exponentially as thedisease duration increased (r="0·91, P<0·0001) (Fig.2). Because there was no correlation between diseaseduration before renal biopsy and disease severity ofAPSGN patients as assessed by peak serum creatinine(Table I, in parentheses), it seems likely that the timecourse of PCNA+ cell numbers was not caused bydifferences in disease severity.Intra-tuft PCNA+ cells were further identified by
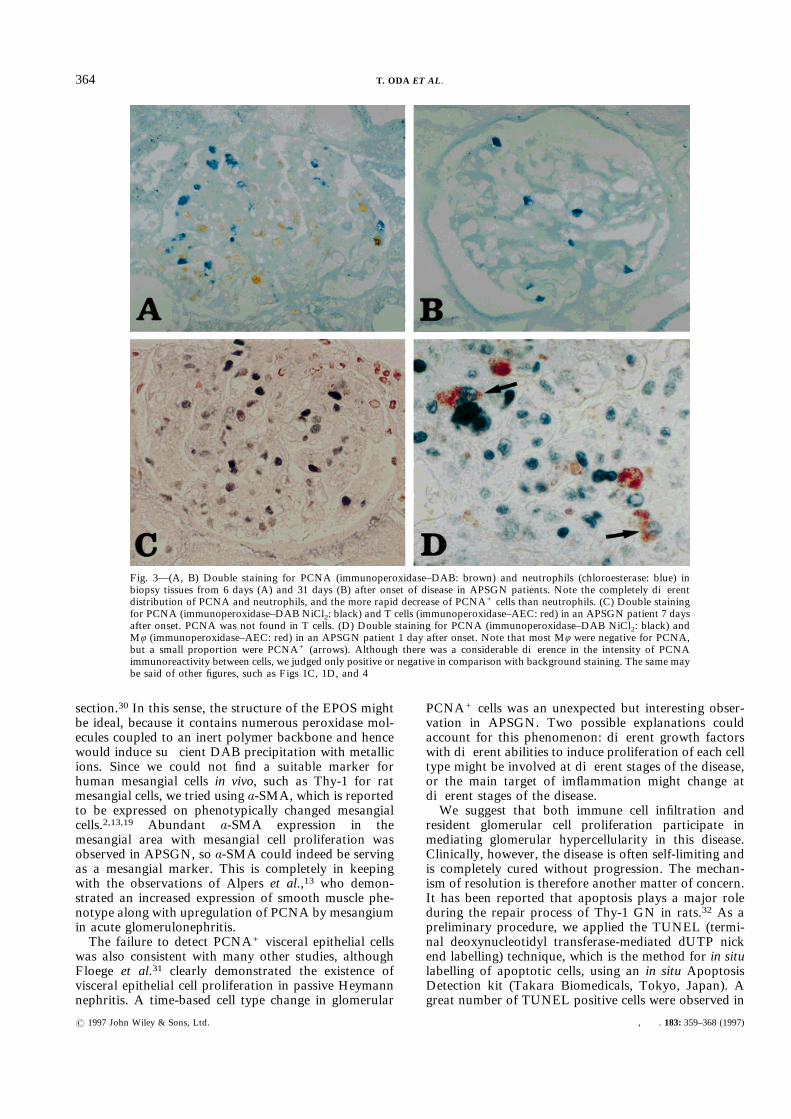
various combinations of double staining. As noted ear-lier,10 glomerular non-resident infiltrating cells consistedof neutrophils, Mö, and a few T cells. PCNA was notfound in neutrophils (Figs 3A and 3B) or in T cells (mostof the T cells showed a CD45RO+ CD45RA" pheno-type) (Fig. 3C), but a proportion of Mö were PCNA+
(Fig. 3D). Since PCNA+ Mö were few, most PCNA+
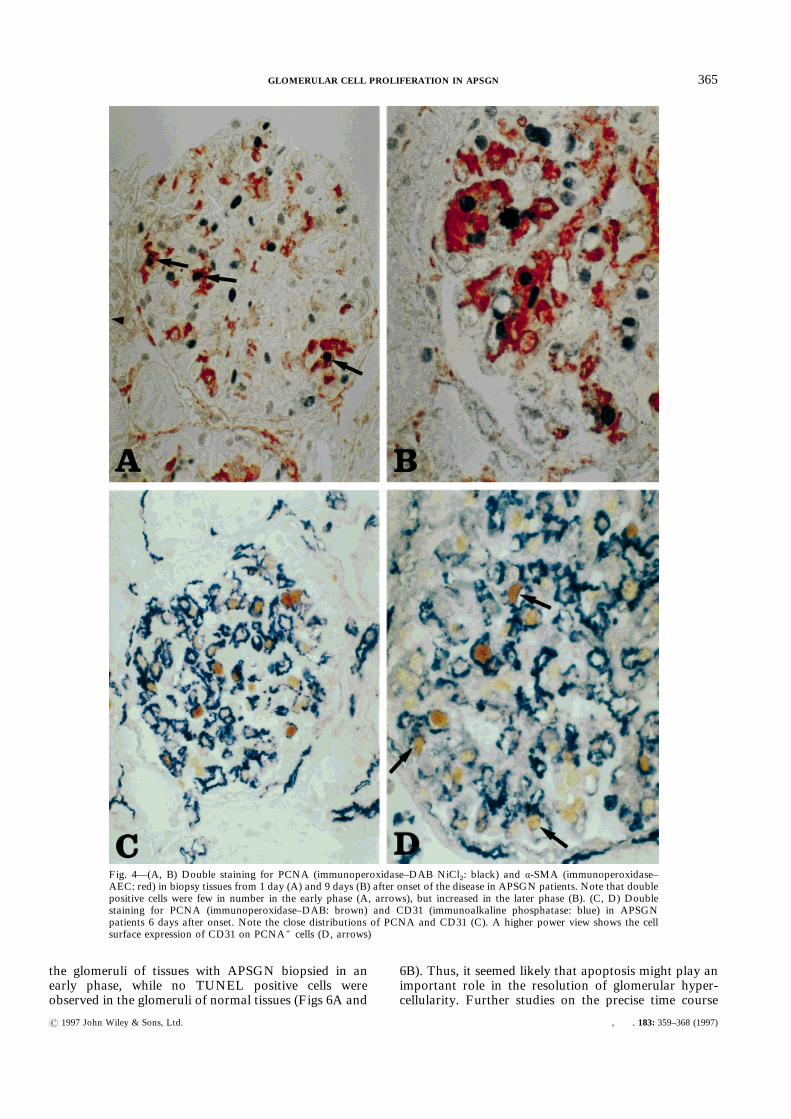
cells were revealed to be resident glomerular cells.PCNA+ resident glomerular cells were identified usingá-SMA and CD31 (endothelial cells). It has beenreported that á-SMA is expressed on the phenotypicallychanged mesangial cells that are going to prolifer-ate.2,19,20 Although glomerular á-SMA was scarce in thenormal controls, all APSGN patients strongly expressedá-SMA in the mesangial areas of glomeruli. Thus,á-SMA could be used as a mesangial marker. In earlierphases of the disease, only small proportions of PCNA+
cells were observed in á-SMA+ areas (Fig. 4A, arrow).However, as the disease progressed, the proportion ofPCNA+ cells in á-SMA+ areas increased to nearly halfof the total PCNA+ cells within the tuft (Fig. 4B). SincePCNA+ infiltrating cells were rare, intra-tuft á-SMA"
PCNA+ cells were suspected of being endothelial cells,but exact identification of the endothelial cells was verydifficult because extended enzymatic digestion was
361GLOMERULAR CELL PROLIFERATION IN APSGN
? 1997 John Wiley & Sons, Ltd. , . 183: 359–368 (1997)
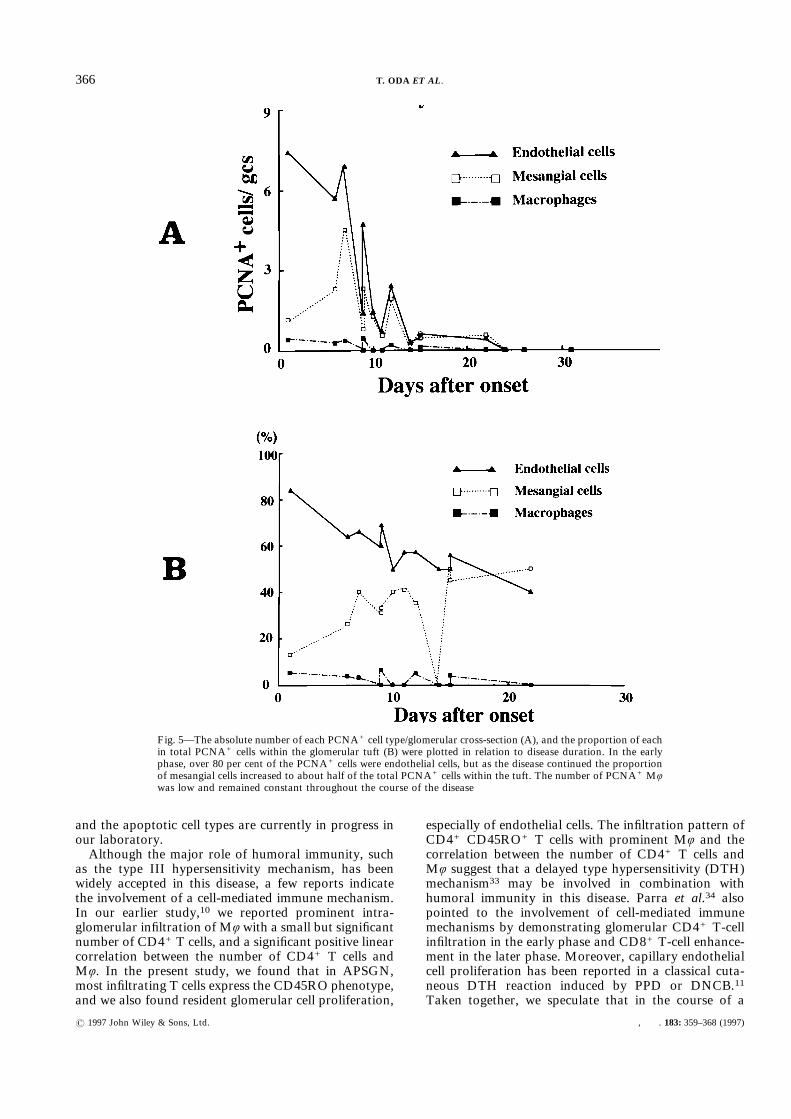
needed to stain endothelial cell markers in formalin-fixed tissue, and not all patients showed a sufficientCD31 staining result to count the endothelium precisely.Therefore, we used not only CD31, but also Factor VIII(VWF) as an endothelial cell marker and identified theendothelium by its location and by cell surface positivityfor CD31 or VWF and negativity for other cell markers(Figs 4C and 4D). Thus, PCNA+ cells within glomerulartufts were characterized as endothelial cells, mesangialcells, or Mö. The time course of the absolute number ofeach PCNA+ cell type and their proportions of totalPCNA+ cells within the tuft are shown in Figs 5A and
5B, respectively. In the early phases of the disease, themajority of PCNA+ cells were endothelial cells, but asthe disease continued the proportion of mesangial cellsincreased to about half of the total PCNA+ cells withinthe tuft. The number of PCNA+ Mö was low andremained constant throughout the course of the disease.
DISCUSSION
We examined the time sequence of glomerular cellproliferation using anti-PCNAMAb in APSGN. PCNA
Fig. 1—Representative immunoperoxidase staining for PCNA in tissues from acute post-streptococcal glomerulonephritis(APSGN) patients and normal controls. (A, B) Control tissues. Although tubular PCNA expression was occasionally seen(A), glomerular PCNA expression was rare. Most glomerular cells that exhibited PCNA in normal controls were parietalepithelial cells (B, arrow). (C, D) APSGN patients. Note the prominent expression of PCNA in the glomerulus and in theinterstitium. The distribution of PCNA+ cells was not generalized; there was a considerable difference in the number ofPCNA+ cells between each glomerulus and between the different parts of the interstitium (C). By counterstaining withPAS, glomerular PCNA+ cells were observed on the inner side of the glomerular tuft or lining Bowman’s capsule (parietalepithelial cells) (D)
362 T. ODA ET AL.
? 1997 John Wiley & Sons, Ltd. , . 183: 359–368 (1997)
has been identified as an auxiliary protein of DNApolymerase delta; its synthesis reaches its maximumduring the S phase of the cell cycle.20 We could detectextremely rare glomerular PCNA+ cells in normal con-trols, and most glomerular cells that exhibited PCNA inthe normal controls were parietal epithelial cells. This issomewhat, but not entirely consistent with Pabst andSterzel’s4 report on autoradiographic analysis of normalrat kidney using [3H]thymidine; they found no labellingin podocytes, a large amount in Bowman’s capsule cells,some in endothelial cells, and little in mesangial cells. Itis not clear why our results differ, such as our detectionof extremely rare PCNA+ cells within the tufts innormal kidneys, but the discrepancy may rest upondifferences between the species examined (human vs.rats) and/or a difference in sensitivity between auto-radiographic analysis and immunohistochemistry withPCNA. It should be noted that some reports point to thesuperior sensitivity of PCNA immuhistochemistry forthe detection of cell proliferation, compared with auto-radiography with [3H]thymidine.21 However, there havebeen some reports indicating difficulties in using PCNAas a proliferation marker.22,23 Overexpression of PCNA,probably because of its long half-life and because of itsinvolvement in DNA repair as well as DNA synthesis,has been reported.22 In fact, we observed numerousPCNA+ cells, even in normal renal tissues, with pre-treatment such as microwave irradiation. In this study,therefore, we did not use any pretreatment. Moreover,we tried polyclonal antibody against Ki-67 antigen(rabbit anti-human Ki-67, recently available fromDAKO), which is applicable to paraffin sections withmicrowave irradiation, is reported to have a muchshorter half-life than PCNA, and correlates well withother markers of proliferation, such as S-phase factionmeasured by DNA flow cytometry.22 Although the
number of positive cells was generally smaller by Ki-67than by PCNA, similar results were obtained with Ki-67;many Ki-67+ cells were observed in the glomeruli ofearly-phase APSGN patients, while positive cells werescarce in normal patients or late-phase APSGN patients(data not shown).The precise mechanism whereby glomerular cells pro-
liferate in vivo has not been fully elucidated. However,PCNA mRNA stability or PCNA expression is generallyknown to be regulated by certain growth factors.24 Inthe present study, PCNA expression was greatly upregu-lated in APSGN but the distribution of PCNA+ cellswas irregular, suggesting that locally produced growthfactors, rather than circulating factors, acted on theglomerular cells and subsequent nephron tubular cells inAPSGN. In the glomerulus, not only infiltrating cellssuch as Mö6,25 or platelets,2,5 but also endothelial cells26or mesangial cells27–29 themselves have been suspected ofinvolvement in glomerular proliferation by secretingseveral growth factors. Moreover, with the EPOStechnique we could precisely identify the types of prolif-erating cells by double immunostaining. EPOS is arecently developed, highly sensitive direct immunoper-oxidase technique.15,16 In the present PCNA immuno-staining study, the EPOS method was more sensitivethan the biotin–streptavidin–peroxidase method (datanot shown) and was more suitable for double immuno-staining because the procedure is simple, rapid, and onestep. Furthermore, no detectable cross-reactivity wasobserved in these double staining results. In doublestaining when both enzymes were peroxidase, wedeveloped the first stain with DAB enhanced with NiCl2to avoid colour mixing of the peroxidases, in accordancewith previous reports showing that sufficient DAB pre-cipitation by metallic ions would block peroxidaseactivity and mask the antibodies previously added to the
Fig. 2—Log value of glomerular PCNA+ cells/glomerular cross-section (gcs) of each APSGNpatient was plotted with reference to duration of the disease before renal biopsy. There was asignificant negative linear correlation between the disease duration and the log value ofglomerular PCNA+ cell number (r="0·91, P<0·0001)
363GLOMERULAR CELL PROLIFERATION IN APSGN
? 1997 John Wiley & Sons, Ltd. , . 183: 359–368 (1997)
section.30 In this sense, the structure of the EPOS mightbe ideal, because it contains numerous peroxidase mol-ecules coupled to an inert polymer backbone and hencewould induce sufficient DAB precipitation with metallicions. Since we could not find a suitable marker forhuman mesangial cells in vivo, such as Thy-1 for ratmesangial cells, we tried using á-SMA, which is reportedto be expressed on phenotypically changed mesangialcells.2,13,19 Abundant á-SMA expression in themesangial area with mesangial cell proliferation wasobserved in APSGN, so á-SMA could indeed be servingas a mesangial marker. This is completely in keepingwith the observations of Alpers et al.,13 who demon-strated an increased expression of smooth muscle phe-notype along with upregulation of PCNA by mesangiumin acute glomerulonephritis.The failure to detect PCNA+ visceral epithelial cells
was also consistent with many other studies, althoughFloege et al.31 clearly demonstrated the existence ofvisceral epithelial cell proliferation in passive Heymannnephritis. A time-based cell type change in glomerular
PCNA+ cells was an unexpected but interesting obser-vation in APSGN. Two possible explanations couldaccount for this phenomenon: different growth factorswith different abilities to induce proliferation of each celltype might be involved at different stages of the disease,or the main target of imflammation might change atdifferent stages of the disease.We suggest that both immune cell infiltration and
resident glomerular cell proliferation participate inmediating glomerular hypercellularity in this disease.Clinically, however, the disease is often self-limiting andis completely cured without progression. The mechan-ism of resolution is therefore another matter of concern.It has been reported that apoptosis plays a major roleduring the repair process of Thy-1 GN in rats.32 As apreliminary procedure, we applied the TUNEL (termi-nal deoxynucleotidyl transferase-mediated dUTP nickend labelling) technique, which is the method for in situlabelling of apoptotic cells, using an in situ ApoptosisDetection kit (Takara Biomedicals, Tokyo, Japan). Agreat number of TUNEL positive cells were observed in
Fig. 3—(A, B) Double staining for PCNA (immunoperoxidase–DAB: brown) and neutrophils (chloroesterase: blue) inbiopsy tissues from 6 days (A) and 31 days (B) after onset of disease in APSGN patients. Note the completely differentdistribution of PCNA and neutrophils, and the more rapid decrease of PCNA+ cells than neutrophils. (C) Double stainingfor PCNA (immunoperoxidase–DAB NiCl2: black) and T cells (immunoperoxidase–AEC: red) in an APSGN patient 7 daysafter onset. PCNA was not found in T cells. (D) Double staining for PCNA (immunoperoxidase–DAB NiCl2: black) andMö (immunoperoxidase–AEC: red) in an APSGN patient 1 day after onset. Note that most Mö were negative for PCNA,but a small proportion were PCNA+ (arrows). Although there was a considerable difference in the intensity of PCNAimmunoreactivity between cells, we judged only positive or negative in comparison with background staining. The same maybe said of other figures, such as Figs 1C, 1D, and 4
364 T. ODA ET AL.
? 1997 John Wiley & Sons, Ltd. , . 183: 359–368 (1997)
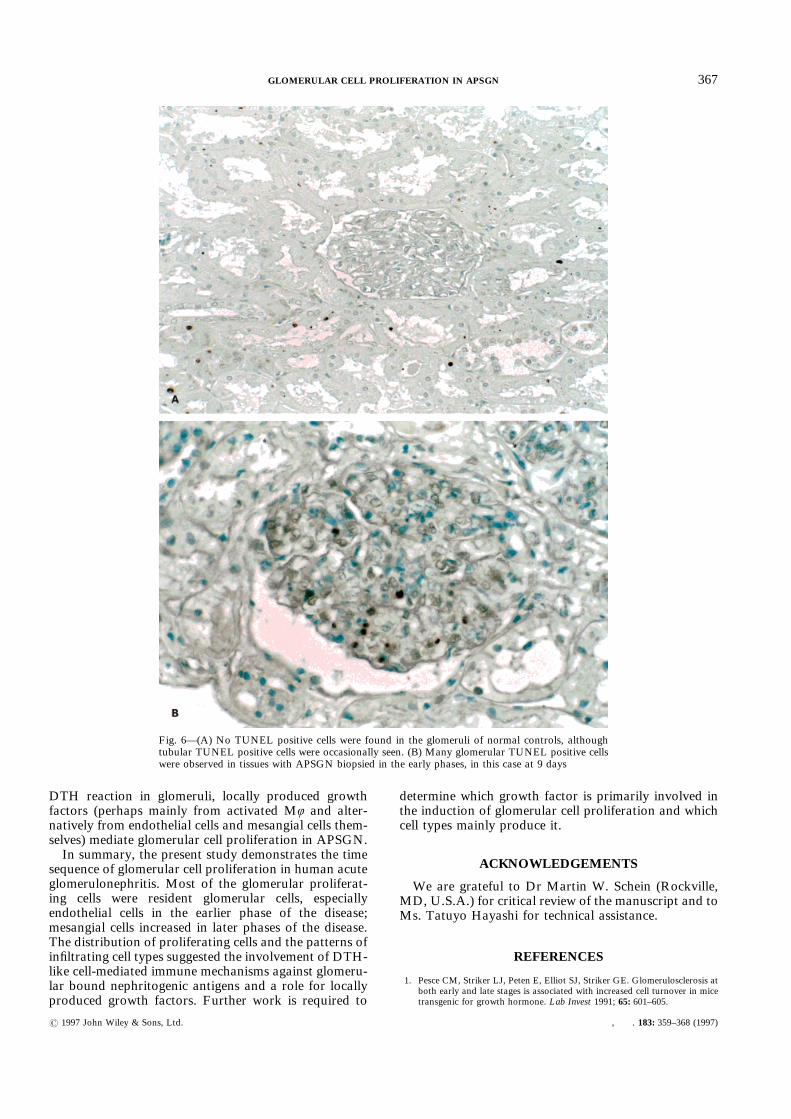
the glomeruli of tissues with APSGN biopsied in anearly phase, while no TUNEL positive cells wereobserved in the glomeruli of normal tissues (Figs 6A and
6B). Thus, it seemed likely that apoptosis might play animportant role in the resolution of glomerular hyper-cellularity. Further studies on the precise time course
Fig. 4—(A, B) Double staining for PCNA (immunoperoxidase–DAB NiCl2: black) and á-SMA (immunoperoxidase–AEC: red) in biopsy tissues from 1 day (A) and 9 days (B) after onset of the disease in APSGN patients. Note that doublepositive cells were few in number in the early phase (A, arrows), but increased in the later phase (B). (C, D) Doublestaining for PCNA (immunoperoxidase–DAB: brown) and CD31 (immunoalkaline phosphatase: blue) in APSGNpatients 6 days after onset. Note the close distributions of PCNA and CD31 (C). A higher power view shows the cellsurface expression of CD31 on PCNA+ cells (D, arrows)
365GLOMERULAR CELL PROLIFERATION IN APSGN
? 1997 John Wiley & Sons, Ltd. , . 183: 359–368 (1997)
and the apoptotic cell types are currently in progress inour laboratory.Although the major role of humoral immunity, such
as the type III hypersensitivity mechanism, has beenwidely accepted in this disease, a few reports indicatethe involvement of a cell-mediated immune mechanism.In our earlier study,10 we reported prominent intra-glomerular infiltration of Mö with a small but significantnumber of CD4+ T cells, and a significant positive linearcorrelation between the number of CD4+ T cells andMö. In the present study, we found that in APSGN,most infiltrating T cells express the CD45RO phenotype,and we also found resident glomerular cell proliferation,
especially of endothelial cells. The infiltration pattern ofCD4+ CD45RO+ T cells with prominent Mö and thecorrelation between the number of CD4+ T cells andMö suggest that a delayed type hypersensitivity (DTH)mechanism33 may be involved in combination withhumoral immunity in this disease. Parra et al.34 alsopointed to the involvement of cell-mediated immunemechanisms by demonstrating glomerular CD4+ T-cellinfiltration in the early phase and CD8+ T-cell enhance-ment in the later phase. Moreover, capillary endothelialcell proliferation has been reported in a classical cuta-neous DTH reaction induced by PPD or DNCB.11Taken together, we speculate that in the course of a
Fig. 5—The absolute number of each PCNA+ cell type/glomerular cross-section (A), and the proportion of eachin total PCNA+ cells within the glomerular tuft (B) were plotted in relation to disease duration. In the earlyphase, over 80 per cent of the PCNA+ cells were endothelial cells, but as the disease continued the proportionof mesangial cells increased to about half of the total PCNA+ cells within the tuft. The number of PCNA+ Möwas low and remained constant throughout the course of the disease
366 T. ODA ET AL.
? 1997 John Wiley & Sons, Ltd. , . 183: 359–368 (1997)
DTH reaction in glomeruli, locally produced growthfactors (perhaps mainly from activated Mö and alter-natively from endothelial cells and mesangial cells them-selves) mediate glomerular cell proliferation in APSGN.In summary, the present study demonstrates the time
sequence of glomerular cell proliferation in human acuteglomerulonephritis. Most of the glomerular proliferat-ing cells were resident glomerular cells, especiallyendothelial cells in the earlier phase of the disease;mesangial cells increased in later phases of the disease.The distribution of proliferating cells and the patterns ofinfiltrating cell types suggested the involvement of DTH-like cell-mediated immune mechanisms against glomeru-lar bound nephritogenic antigens and a role for locallyproduced growth factors. Further work is required to
determine which growth factor is primarily involved inthe induction of glomerular cell proliferation and whichcell types mainly produce it.
ACKNOWLEDGEMENTS
We are grateful to Dr Martin W. Schein (Rockville,MD, U.S.A.) for critical review of the manuscript and toMs. Tatuyo Hayashi for technical assistance.
REFERENCES
1. Pesce CM, Striker LJ, Peten E, Elliot SJ, Striker GE. Glomerulosclerosis atboth early and late stages is associated with increased cell turnover in micetransgenic for growth hormone. Lab Invest 1991; 65: 601–605.
Fig. 6—(A) No TUNEL positive cells were found in the glomeruli of normal controls, althoughtubular TUNEL positive cells were occasionally seen. (B) Many glomerular TUNEL positive cellswere observed in tissues with APSGN biopsied in the early phases, in this case at 9 days
367GLOMERULAR CELL PROLIFERATION IN APSGN
? 1997 John Wiley & Sons, Ltd. , . 183: 359–368 (1997)

2. Floege J, Burns MW, Alpers CE, et al. Glomerular cell proliferation andPDGF expression precede glomerulosclerosis in the remnant kidney model.Kidney Int 1992; 41: 297–309.
3. Floege J, Eng E, Young BA, Couser WG, Johnson RJ. Heparin suppressesmesangial cell proliferation and matrix expansion in experimental mesan-gioproliferative glomerulonephritis. Kidney Int 1993; 43: 369–380.
4. Pabst R, Sterzel RB. Cell renewal of glomerular cell types in normal rats.An autoradiographic study. Kidney Int 1983; 24: 626–631.
5. Bhan AK, Collins AB, Schneeberger EE, McCluskey RT. A cell mediatedreaction against glomerular-bound immune complexes. J Exp Med 1979;150: 1410–1420.
6. Sterzel RB, Pabst R. The temporal relationship between glomerular cellproliferation and monocyte infiltration in experimental glomerulonephritis.Virchows Arch B 1982; 38: 337–350.
7. Johnson RJ, Garcia RL, Pritzl P, Alpers CE. Platelets mediate glomerularcell proliferation in immune complex nephritis induced by anti-mesangialcell antibodies in the rat. Am J Pathol 1990; 136: 369–374.
8. Diamond JR, Ding G, Frye J, Diamond IP. Glomerular macrophages andthe mesangial proliferative response in the experimental nephrotic syn-drome. Am J Pathol 1992; 141: 887–894.
9. Yoshizawa N, Oshima S, Sagel I, Shimizu J, Treser G. Role of a strepto-coccal antigen in the pathogenesis of acute post streptococcal glomerulone-phritis. J Immunol 1992; 148: 3110–3116.
10. Yoshizawa N, Oda T, Oshikawa Y, et al. Cell-mediated immune response inacute poststreptococcal glomerulonephritis. Jpn J Nephrol 1994; 36: 322–330.
11. Polverini PJ, Cotran RS, Sholley MM. Endothelial proliferation in thedelayed hypersensitivity reaction. An autoradiographic study. J Immunol1977; 118: 529–532.
12. Shigematsu S, Shishido H, Kihara K, et al. Participation of monocytes intransient glomerular hypercellularity in poststreptococcal glomerulonephri-tis. Virchows Arch B 1973; 12: 367–370.
13. Alpers CE, Hudkins KL, Gown AM, Johnson RJ. Enhanced expression of‘muscle-specific’ actin in glomerulonephritis. Kidney Int 1992; 41: 1134–1142.
14. Takeuchi A, Yoshizawa N, Yamamoto M, et al. Basic fibroblast growthfactor promotes proliferation of rat glomerular visceral epithelial cells invitro. Am J Pathol 1992; 141: 107–116.
15. Bisgaad K, Linme A, Rolsted H, Pluzek K-Z. Polymeric conjugates forenhanced signal generation in enzyme linked immunoassays. XXIVthAnnual Meeting of the Scandinavian Society for Immunology, Aathus,Denmark, 1993 (Abstract).
16. Tsutsumi Y, Serizawa A, Kawai K. Enhanced polymer one step staining(EPOS) for proliferating cell nuclear antigen (PCNA) and Ki-67 antigen.Application to intra-operative frozen diagnosis. Pathol Int 1995; 45: 108–115.
17. Cordell JL, Falini B, Erber WN, et al. Immunoenzymatic labeling ofmonoclonal antibodies using immune complexes of alkaline phosphataseand monoclonal anti-alkaline phosphatase (APAAP complexes). J Histo-chem Cytochem 1984; 32: 219–229.
18. Li CY, Lam KW, Yam LT. Esterases in human leukocytes. J HistochemCytochem 1973; 21: 1–12.
19. Johnson RJ, Iida H, Alpers CE, et al. Expression of smooth muscle cellphenotype by rat mesangial cells in immune complex nephritis: á-smoothmuscle actin is a marker of mesangial cell proliferation. J Clin Invest 1991;87: 847–858.
20. Bravo R, Frank R, Blundell P, Bravo H. Cylcin/PCNA is the auxiliaryprotein of DNA polymerase-delta. Nature 1987; 326: 515–517.
21. Zeymer U, Fishbein MC, Forrester JS, Cercek B. Proliferating cell nuclearantigen immunohistochemistry in rat aorta after balloon denudation. Com-parison with thymidine and bromodeozyuridine labeling. Am J Pathol 1992;141: 685–690.
22. Carmen CW, Filipe MI. Update on proliferation-associated antibodiesapplicable to formalin-fixed paraffin-embedded tissue and their clinicalapplications. Histochem J 1993; 25: 843–853.
23. McCormick D, Hall PA. The complexities of proliferating cell nuclearantigen. Histopathology 1992; 21: 591–594.
24. Jaskulski D, Gatti C, Travali S, Calabretta B, Baserga R. Regulation of theproliferating cell nuclear antigen cyclin and thymidine kinase mRNA levelsby growth factors. J Biol Chem 1988; 263: 10175–10179.
25. Van-Diemen-Steenvoorde R, Lambers A, van der Waal A, et al. Macro-phages are responsible for mesangial cell injury and extracellular matrixexpansion in anti-Thy-1 nephritis in rats. J Am Soc Nephrol 1991; 2: 585A.
26. Castellot JJ, Hoover RL, Karnovsky MJ. Glomerular endothelial cellssecrete a heparin-like inhibitor and a peptide stimulator of mesangial cellproliferation. Am J Pathol 1986; 125: 493–500.
27. Doi T, Striker LJ, Elliot SJ, Conti FG, Striker GE. Insulin like growthfactor-1 is a progression factor for human mesangial cells. Am J Pathol1989; 134: 395–404.
28. Silver BJ, Jaffer FE, Abboud HE. Platelet-derived growth factor synthesis inmesangial cells: induction by multiple peptide mitogens. Proc Natl Acad SciUSA 1988; 86: 1056–1060.
29. Lovett DH, Szamel M, Ryan JL, Sterzel RB, Gemsa D, Resch K.Interleukin 1 and the glomerular mesangium I. Purification and characteri-zation of a mesangial cell-derived autogrowth factor. J Immunol 1986; 136:3700–3705.
30. Hsu SM, Soban E. Color modification of diaminobenzidine (DAB) precipi-tation by metallic ions and its application for double immunohistochemis-try. J Histochem Cytochem 1982; 30: 1079–1082.
31. Floege J, Johnson RJ, Alpers CE, et al. Visceral glomerular epithelial cellscan proliferate in vivo and synthesize plate-derived growth factor B-chain.Am J Pathol 1993; 142: 637–650.
32. Shimizu A, Kitanura H, Masuda Y, Ishizaki M, Sugisaki Y, Yamanaka N.Apoptosis in the repair process of experimental proliferative glomerulone-phritis. Kidney Int 1995; 47: 114–121.
33. Waldorf HA, Walsh LJ, Schechter NM, Murphy GF. Early cellular eventsin evolving cutaneous delayed hypersensitivity in humans. Am J Pathol1991; 138: 477–486.
34. Parra G, Platt JL, Falk PJ, Rodriguez-Iturbe B, Michael AF. Cell popula-tions and membrane attack complex in glomeruli of patients with post-streptococcal glomerulonephritis: identification using monoclonal anti-bodies by indirect immunofluorescence. Clin Immunol Immunopathol 1984;33: 324–332.
368 T. ODA ET AL.
? 1997 John Wiley & Sons, Ltd. , . 183: 359–368 (1997)