Embed Size (px)
Citation preview

395
Biochimica et Biophysica Acta, 447 (1976) 395--405 © Elsevier/North-Holland Biomedical Press
BBA 98734
GIANT RNA IN INSECTS I. DIFFERENTIAL CHANGES DURING CALLIPHORA DEVELOPMENT AND THE ROLE OF ECDYSONE
ELI SHAAYA
Laboratory of Insect Physiology, Terra Santa, The Hebrew University of Jerusalem (Israel)
(Received April 13th, 1976)
Summary
The pattern of newly synthesized RNA in the epidermis cells of Calliphora during the larval life to the early pupal stage was analysed by electrophoretic separation of RNA extracts on polyacrylamide gels. During larval life, mainly rRNA was synthesized. The elevation of ecdysone concentration during the synthesis of the new larval cuticle resulted primarily in the stimulation of rRNA synthesis. In the transition period from larva to pupa, the principal RNA synthesized was rapidly labelled giant DNA-like RNA, with an average life span of 30 min. Prior to the change in RNA metabolism at this development stage, the synthesis of all classes of RNA was depressed. The initial increase in hnRNA synthesis occurs at the onset of the wandering stage, shortly after the appear- ance of a small peak of ecdysone and about two days prior to the main peak of hormone secretion and pupariation.
During induced pupariation by ecdysterone, 1 h after hormone treatment, more label was observed in giant hnRNA followed by elevation of rRNA syn- thesis. Processing and maturation of rRNA also seemed to be stimulated.
Introduction
Molting and metamorphosis are unique events in the development of insects [1,2]. The invariable requisite for these processes is tissue stimulation by the molting hormone ecdysone [3]. Thus, insect epidermis cells have been used in many laboratories for the study of the molecular action of ecdysone [4--6] and as a system for the study of the molecular aspect of development [7]. How- ever, the real understanding of the mechanism by which ecdysone exerts the diverse physiological and biochemical responses during insect life is essentially
Abbreviations: hnRNA, heterodisperse nuclear RNA; hRNA, heterodisperse RNA.

396
unknown. Taking advantage of improved methods for extraction and separa- tion of RNAs, the present s tudy was undertaken to compare the pattern of newly synthesized RNA in the epidermis cells during the larval life of Calli- phora to the early pupal stage, with the emphasis on two processes initiated by ecdysone. One, during larval molt , a process which is controlled by both ecdy- sone and juvenile hormone; and the other one, during larval pupal transition, which takes place in the absence of juvenile hormone. In addition, detailed studies on ecdysterone 's effect on epidermis RNA synthesis during induced metamorphosis by the hormone were also conducted.
Materials and Methods
Calliphora erythrocephala larvae were raised on pork at 23 + 0.5°C and at a relative humidi ty of about 60%. Under our experimental conditions, the larva which hatches from the egg undergoes two larval molts at the age of 2 and 3 days, and it becomes fully grown when about 6 days old. Then, it leaves its food and crawls around actively for two days before it becomes a white pupa. At the white pupa stage, various internal changes characteristic of early meta- morphosis have already commenced.
In order to s tudy the effect of ecdysterone on RNA metabolism, 0.05 #g ec- dysterone and 10 #Ci [5-3H]uridine (28 Ci/mmol, Schwarz/Mann) dissolved in 4 pl Ringer solution (0.15 M NaC1, 3 mM KC1, 2 mM CaC12 • 2 H20 buffered with I mM Na2HPO4 to pH 7.3) were injected into abdomina of Calliphora. Control animals received [aH]uridine in Ringer solution wi thout ecdysterone. Each abdomen was preinjected with 7 pl/Ringer solution 3 h before t reatment to stabilize the rate of uridine incorporation [8]. For induced pupariation by ecdysterone, ligated larvae before the period of ecdysone secretion were used [9].
For RNA extraction, epidermis from 5 abdomina was dissected and briefly rinsed in an ice-cold Ringer solution and immediately frozen on solid CO2. The frozen tissue was homogenized in a glass homogenizer (10 strokes) in 2-ml pro- nase solution pH 7.4 (2 pg/ml pronase, 30 mM Tris, 0.1 M NaC1, 1 mM Na2- EDTA, 1% sodium lauryl sulfate and 10/~g/ml polyvinyl sulphate) which had been pre-incubated at 37°C for 30 min. The homogenate was incubated at 37°C for 1 h. The homogenate was shaken with an equal volume of phenol saturated with Tris buffer pH 7.4 (30 mM Tris, 0.1 M NaC1, 1 mM Na2EDTA and 10/~g/ml polyvinyl sulfate) at ice temperature, centrifuged and the aque- ous layer was collected. The phenol layer was re-extracted with 0.5 vols. of the original buffer and the aqueous phases combined and re-extracted twice with cold phenol, followed by two chloroform extractions. The RNA solution was first brought to 0.2 M NaC1 and then precipitated twice from the aqueous phase with 2 vols. ethanol and stored at --20°C overnight. The epidermis of one abdomen yielded about 50 pg RNA. The concentrat ion of RNA in solution was estimated spectrophotometrical ly. The specific activity of radioactive RNA was measured as previously described [8].
For gel electrophoresis, gels of 2.2% polyacrylamide (10 × 0.6 cm) were pre- pared according to the method of Loening [10], with the exception that a ratio of bisacrylamide (Bis): total monomer of 0.021 was used to increase the pore

397
size of the gel. This modification allows the simultaneous resolutions of 18 S, 28 S rRNA, 38 S pre-rRNA and hRNA molecules of a molecular weight up to 20 • 106 [11]. For further details, see Shaaya and Clever [8].
Results
The changes in the wet weight of Calliphora epidermis compared with that of the whole animal during the larval life (Fig. la) show that in the first 5.5 days of larval life, the weight of the whole larva and the epidermis increase steadily. After this period and until pupariation, when the larva stops feeding, the weight of the epidermis remains fairly unchanged, while the total weight of the larva decreases gradually. The amount of bulk RNA per epidermis increases parallel to the increase in the weight of the epidermis, the maximum being 130 pg RNA per epidermis in 5.5~lay old larva (Fig. lb). However, in the older lar- va (Figure lb), the amount of ribonuleic acid steadily decreases and 55 pg RNA per epidermis was measured in the 6.5 day-old larva. This amount re- maines nearly unchanged for the rest of the larval life. In the epidermal cells, the study of the [3H]uridine incorporation into RNA, after a 1-h labelling period, revealed dramatic changes in the specific radioactivity of the RNA at various larval stages (Fig. lb). During the molting process, from the second to the third larval instar, a maximum incorporation of 800 cpm/pg RNA was measured during the synthesis of the new larval cuticle and prior to apolysis, the loosening of the old cuticle. The incorporation of uridine de- creased sharply to 220 cpm/pg RNA shortly before and after ecdysis, the shed- ding of the old cuticle. In 5--6 day-old larvae, the incorporation of uridine was very low, about 60 cpm/pg RNA. For the rest of the larval period, the epider- mis showed a gradual increase of [3H]uridine incorporation into RNA with a maximum of 840 cpm/#g, shortly before pupariation.
As can be seen from Fig. lb, the changes in the specific radioactivity of the RNA obtained in the various developmental stages cannot be attributed mainly
1 l ° r , , i , , , , ~ , 30
. a g e o f l a r v e ( d a y s ) ~ a g e o f l a r v e ( d a y s ) --,'-
F i e I . (a) Changes in the w e t w e i g h t o f to ta l larva (e o) and ep idermis ce l l s (o o) during larval life. (b) Changes in the a m o u n t o f to ta l bulk R N A / c p i d e r m i s ( o ) and [3H]urJd ine i n c o r p o r a t i o n in to R N A (e) during larval l i fe . Each larva was injected with 10 #Ci [3H]ur idine and i n c o r p o r a t i o n a l l o w e d for 60 m i n b e f o r e the animals w e r e killed. Five animals were used for each e x p e r i m e n t . Each p o i n t is based on three determinat ions . M, mol t ; WS, wandering stage; A0 apolysis; E° ecdysis; P, pupa,

398
to the changes in the amount of bulk RNA in the cells. Nor is the low incorpo- ration of [3H]uridine in the 5--6 day-old larva a consequence of pronounced changes in nucleotide uptake into the epidermis cells during this period, as only a slight increase in the rate of precursor uptake was measured in the epidermis cells of 7.5 day-old larva, compared to that of the 5--6 day-old larva {Figure not shown.) However, the gradual decrease in the bulk RNA in the epidermis cells during this period (Fig. l b ) shows clearly that very little RNA is synthe- sized to replace the normally degrading rRNA of the cells. Thus, the low incor- poration of uridine is a consequence of the reduced rate of RNA synthesis.
To investigate whether the changes in the incorporation of label into RNA also reflect a differential change in the synthesis of various species of RNA, we made the following analysis:
1. We analysed epidermis RNA from various stages during the process of molting from the second to the third instar. Gel electrophoresis profiles of RNA samples isolated after 1 h labelling period can be seen in Figs. 2a and 2b. The absorbance profiles of all the RNA samples analysed were the same. The label in epidermis RNA isolated from animals 6--8 h before apolysis, 1--2 h prior to ecdysis, was found mainly distributed among 38 S pre-rRNA, 28 S and 18 S rRNA. Thus, the increase in the specific radioactivity shortly before apo- lysis is due mainly to the elevation of the label in all species of rRNA, including pre-rRNA. The profile of epidermis RNA isolated from animals 10 h after ec- dysis was similar to that of animals of 1--2 h prior to ecdysis (Figure not shown).
2. We analysed RNA extracted from epidermis of third instar larvae at differ- ent ages till pupariation (Figs. 3,a-d). In the epidermis of 4-day old larvae (Fig. 3a), the shape of RNA profiles was similar to that in Fig. 2b. During the de- pression period of uridine incorporation into RNA in the 5--6 day-old larva, the synthesis of all species of RNAs was depressed (Fig. 3b). After this period, when the epidermis cells started again to incorporate a high amount of [3H]- uridine into RNA at day 6.5 of larval life till pupariation, the main portion of the label was found in the region of molecular weight higher than 40 S (Fig. 3e, 3d). It is clear that though the amount and the specific radioactivity of the RNA are nearly the same before apolysis to the third larval instar and prior to pupariation (Fig. lb ) , the various types of RNA synthesized in these two devel- opmental stages are different. Before and after apolysis, around 70% of the to- tal radioactivity in the gel was in rRNA, calculated from Figs. 2a, 2b and 3a, while from the onset of the wandering stage till pupariation, more than 70% of the label was in hnRNA of s value higher than 40 S, calculated from Figs. 3c and 3d.
The s tudy of the effect of eedysterone on RNA metabolism in the epidermis cells during the process of induced pupariation showed that ecdysterone admin- istered for 30 min followed by an [3H]uridine pulse of 15 min caused no in- crease in the specific radioactivity of the RNA compared to that of control ani- mals. In animals where eedysterone was administered for 60 and 90 min, the specific activity of the RNA was increased by 30 and 60% respectively. After 4 h, ecdysterone was found to have no effect. The profiles shown (Fig. 4a,b) were those obtained from gel elec~ophoresis of the same RNA of hormone- treated and the control animals as described above. These show that the in-
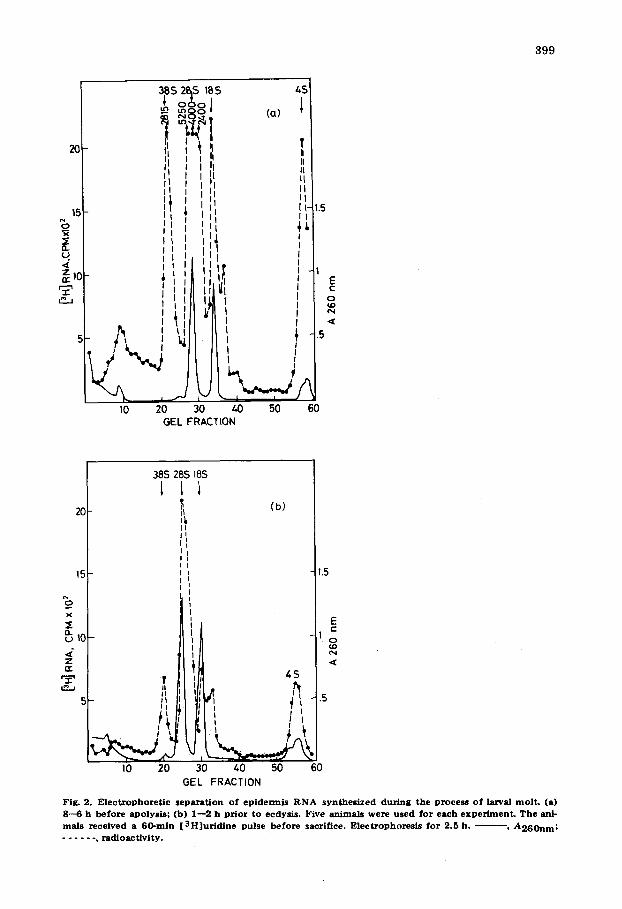
399
20
15
5 : a.
=z,0
385, 2~S 185 45
°°o I ~ 8 o (a)
i I , , ll'
I
!
%.,....d I0 20 30 40 50 60
GEL FRACTION
'1 I II I I
I I t 1.5 I 1,
E (::
o M} N
2C
15
x
~, ~0
Z (:l:
5
38S 28S 18S
II
] I
(b)
1.5 i
.5
10 20 30 40 50 60 GEL FRACTION
Fig. 2. E l e c t r o p h o r e t i c s epa ra t i on o f ep ide rmis R N A syn thes i zed dur ing the process o f larval mo l t . (a) 8--6 h be fo re apolysis ; (b) 1- -2 h p r io r to ecdysis. Five an imals were used for each e x p e ~ e n t . T h e ani- mals rece ived a 60-ra in [ 3 H ] u r i d i n e pulse be fo re sacrifice. E lec t rophores i s fo r 2.5 h. ~ - , A 2 6 0 n m ; . . . . . . , r ad ioac t iv i ty .

4 0 0
2 0
15
x
(,9
' ~ ' 5
38 2B
1
/! ,^:
1o 20 30 ~0 GEL FRACTION
18 I ~(a)
1.0
o,1
.5 "~
50 60
2 2BS IBS (b)
1 1
~ _ (c) I I I I ~
I0 20 30 ~0 50 6( GEL FRACTION
2j,s
11 I i i ~10l ) ~ V
2 Io 20 30 40
GEL FRACTION
185 J (d)
1
E c
(.D
I .s
I
50 60
Fig . 3. E l e c t r o p h o r e t i c s e p a r a t i o n o f e p i d e r m i s R N A s y n t h e s i z e d d u r i n g t h e t h i r d l a r v a l i n s t a r t i l l s h o r t l y b e f o r e p u p a r i a t i o n . (a ) 4 - d a y - o l d l a r v a ; ( b ) 5 - d a y - o l d l a r v a ; (e ) 6 . 5 d a y - o l d l a r v a ; ( d ) 1 0 - - 8 h p r i o r t o
p u p a r i a t i o n , T h e a n i m a l s r e c e i v e d a 6 0 - m i n [ 3 H ] u r i d i n e p u l s e b e f o r e t h e y w e r e k i l l e d . F i v e a n i m a l s w e r e
used f o r e a c h e x p e r i m e n t . Electrophoresis w a s f o r 4 h . - - , A 2 6 0 n m ; . . . . . . , r a d i o a c t i v i t y .
crease in the specific activity of the RNA after 60 min of hormone treatment represented mainly an increase in the incorporation of label in RNA of s value higher than 40 S, while the amount of label incorporated in the 38 S pre-rRNA remained the same as that of the control animals. Also, the profile of RNA af- ter 9Omin of hormone t reatment showed an increase in label of RNA of s value higher than 40 S compared to the control. RNA synthesized in the epidermis 30 min and 4 h after hormone treatment showed a profile identical to that of the control (Results not shown).

4 0 1
25~1 (a)
0 tO 20 30 40 50 60 GEL FRACTION
25
~ 20
~E
z
5
0 0
3S.s (b) l
10 20 30 40 50 60 GEL FRACTION
Fig. 4. E l e c t r o p h o r e t i c s e p a r a t i o n o f R N A e x t r a c t e d f r o m e c d y s t e r o n e - t r e a t e d (o) a n d c o n t r o l a b d o m e n s ($) . A b d o m e n s were f i rs t i n j e c t e d w i t h e e d y s t e r o n e ( 0 . 0 5 #g) o r R i n g e r s o l u t i o n for : (a) 6 0 r a in ; (b) 9 0 r a i n ; f o l l o w e d b y a 15 vain [ 3 H ] u r i d i n e pulse b e f o r e sacr i f ice o f t h e a b d o m e n s . V a l u e s o f s r e f e r to t he a b s o r b a n c e p e a k s a t 2 6 0 rim. O t h e r de ta i l s as in Fig. 3.
To investigate further the effect of ecdysterone on rRNA synthesis, we in- creased the uridine pulse from 15--45 min to permit rRNA maturation. Acryl- amide gel analysis {Fig. 5) of the RNA were in good agreement with the report. ed data in Fig. 4. Thus, the label in RNA migrated slower than 40 S, increased after 45 min of hormone treatment, and rRNA synthesis was elevated only 1 h after hormone treatment. This shows that ecdysterone may first act by increas- ing the synthesis of high molecular hnRNA, and shortly after may effect the synthesis of rRNA. This is also evident from calculating, at various times of hormone treatment, the percentage change of control in the amount of label measured in the region where RNA migrated slower than 40 S, and in the re- gions of the different species of rRNA (Fig. 6). Ecdysterone administered for 90 min followed by 15 min [3H]uridine pulse before the animals were killed caused a maximum of 56% stimulation of hRNA of s value higher than 40 S without effect on rRNA, mainly 38 S pre-rRNA. A 35% stimulation of rltNA synthesis occurred only 120 min after hormone treatment (Fig. 6a). In other

402
25
~2o
,-if, It
I/A
I I I I I
10 20 30 40 50 60 GEL FRACTION
? i 38.S 28S 18.S fl , , 1 1 1
40 -,~ :', (b) $
II I II
36 -" I I I I I o
3 o i : ~, I
'I "_o25
x
2(3 n I I
z A ,o
~ ~,oL i I ~ I:l I
I I I I L ,
10 20 30 40 50 60 GEL FRAC lION
Fig. 5. E l e c t r o p h o t e t i c s epa ra t ion of R N A e x t r a c t e d f r o m e c d y s t e r o n e - t r e a t e d (o) and co n t ro l (o) abdo- mens . A b d o m e n s were first in jec ted wi th e e d y s t e r o n e or R inger so lu t ion for (a) 45 rain, (b) 6 0 rain; fol- l owed by a 45-ra in [ 3 H ] u r i d l n e pulse be fo re the an imals were killed. O t h e r detai ls as in Fig. 3.
similar experiments with a longer [3H]uridine pulse of 45 min to permit rRNA maturation, the hormone was also found to stimulate first hnRNA and only later to stimulate rRNA (Fig. 6b). The effect of the hormone was diminished 4 h after the treatment. This is to be expected, since ecdysone was found to be totally metabolized 5 h after administration [12].
In continuous labelling, in which the hormone and isotopically labelled pre- cursor were injected simultaneously, 2 h before the animals were killed (Fig. 7), the specific activity of the total RNA of the treated animals increased by 107%

403
._J
8 ~,o bJ
~: 2o "r
o -10
MIN
(a)
I I I jl I
0 45 2~0 Mi~-
(b)
Fig. 6 . P e r c e n t a g e c h a n g e o f c o n t r o l in the a m o u n t o f l abe l m e a s u r e d in h n R N A h i g h e r t h a n 4 0 S (e ) a n d t o t a l r R N A (o) a t v a r i o u s t i m e s o f e c d y s t e r o n e a d m i n i s t r a t i o n . (a) Absc i ssa : m i n a f t e r e c d y s t e r o n e a d m i n - i s t r a t i o n f o l l o w e d b y 15 r a i n [ 3 H ] u r i d i n e pu l se b e f o r e the an ima l s we re ki l led. The va lues were c a l c u l a t e d f r o m the gel p ro f i l e s s h o w n in Fig. 4 a n d o t h e r s n o t s h o w n . (b) Absc i ssa : r a i n a f t e r e c d y s t e r o n e admin i s - t r a t i o n f o l l o w e d b y 4 5 r a in [ 3 H ] u r i d i n e pu l se b e f o r e the an ima l s w e r e sac r i f i ced . T h e va lues we re ca lcu - l a t e d fo r the gel p ro f i l e s in Fig . 5 a n d o t h e r s n o t s h o w n .
compared to that of the control. On the other hand, analysis of this RNA showed that the amount of label o f the mature rRNA increased by 200%, com- pared to an only 90% increase in the 38 S pre-rRNA calculated from Fig. 7. This shows that the stimulation of rRNA after hormone treatment is due not only to an increase in the synthesis of the pre-rRNA, but also to an increase in
38 S 28S 11~S Q
.~1" u-> 0
40 u~
30
c~
o
1C
5
,; 2'0 3'o ,'o s'o 6o GEL FRACTION
Fig. 7. E l e c t r o p h o r e t i c seps . ra t ion o f R N A e x t r a c t e d f r o m e c d y s t e r o n e - t r a a t e d (o) a n d c o n t r o l a b d o m e n s (e ) . A b d o m e n s r ece ived s i m u l t a n e o u s l y e c d y s t e r o n e a n d [ 3 H ] u r i d i n e f o r 1 2 0 m i n b e f o r e the a n i m a l s we re ki l led. O t h e r de ta i l s w e r e as in Fig . 3. P e r c e n t a g e s t i m u l a t i o n of: t o t a l R N A , 107%; h R N A h i g h e r t h a n 4 0 S, 60%; 3 8 S p r e - r R N A , 90%; r R N A , 2 0 0 % .

404
the rate or efficiency of processing of the rRNA precursors. The increase of only 60% of radioactivity in hnRNA can be explained by the short life span of about 30 min of this RNA species [14].
Discussion
The data presented here show a pronounced variation in the specific radio- activity of epidermis RNA during the postembryonic development of Calli- phora and differential changes in the amount of the various RNA classes syn- thesized in the different developmental stages.
The observation that the increase in the specific radioactivity around apol- ysis is due mainly to ~he elevation of rRNA synthesis, while the increase during the transition from the larva to pupa is due to elevation in hnRNA, is of partic- ular interest as the two processes are known to be under the control of ecdys- one. The only difference is that ecdysone alone in the absence of juvenile hor- mone will lead to progress in development and pupariation, while for larval molt, juvenile hormone must be present as well. The stimulation of rRNA syn- thesis around apolysis is understandable, because increased growth implies an increase in the protein synthetic capacity of the cells, and therefore an in- creased formation of ribosomes. The finding that about 70% of the RNA syn- thesized during the larval-pupal transition is a DNA-like RNA, and represents molecules of molecular weight up to 20 • 106 with a life span of about 30 min [13,14] is of particular significance in regard to a possible precursor-product relationship between nuclear hRNA and mRNA [15--18]. However, it should be noted that this informational change does not necessarily mean an increase in the amount of cytoplasmic mRNA.
A study of ecdysone titer during the development of Calliphora and other in- sects [ 19,20] showed that at the process of larval-larval molt, a substantial in- crease in ecdysone titer was measured during the process of the new cuticle for- mation, shortly before apolysis. This points to a causal relationship between the increase in the ecdysone titer and the elevation of rRNA synthesis. Of par- ticular interest is the finding that the initial increase in hnRNA synthesis occurs at the onset of the wandering stage, about two days prior to the appearance of the main peak of ecdysone and pupariation. However, a small peak of the hor- mone could be measured shortly before the changes in RNA synthesis took place. It is likely that the observed depression in RNA synthesis followed by a substantial increase in hnRNA, is related to the reprogramming of the genome at this stage. Further work is needed in order to ascertain whether these events are controlled by minor changes in the concentrat ion of ecdysone in the epider- mis cells and/or, by the reduced level of juvenile hormone at this particular stage [21]. It is commonly accepted that juvenile hormone inhibits the mor- phogenetic changes associated with metamorphosis [22,23]. Therefore, it is possible that those genes determining pupal or imaginal characteristics will not be expressed' in the presence of ecdysone plus juvenile hormone, but only by ecdysone alone.
As it is well known, in Calliphora larva, the distraction of the ring gland, the source of ecdysone, by ligation prior to the appearance of the main ecdysone peak, results in the maintenance of the larval character of the posterior part of

405
the animal. The epidermis cells of this part continue to synthesize hnRNA with no pronounced change for at least 48 h after ligation. This points out that the presence of the ring gland is not necessary for the persistence of hnRNA syn- thesis. During induced pupariation in these abdomina by ecdysterone, there is an increase in the label measured in hnRNA within 1 h of hormone administra- tion fol lowed by an elevation of rRNA synthesis. The observation that poly- somes appear in sito shortly before pupariation after the main peak of ecdys- one secretion, and in ligated larvae within 3 h of ecdysone administration [24] fol lowed by a new production of the enzyme Dopa-decarboxylase [4] , suggests that in the presence of a high level of endo- or exogenous ecdysone, much more information is transported to the cytoplasm. Whether this is due to new synthe- sis of RNA or modification of the existing hnRNA, is now under intensive study.
As there is suggestive evidence that the molecular action of ecdysone is dic- tated not only by the variation in the hormone titer, but also by the type of ecdysone [25- -27] , it is likely that this is also the case in the various develop- mental stages of Calliphora.
References
1 Locke, M. (1964) in The Phys io logy o f Insec ta (Rockstein, M., ed.), Vol. 3, pp. 379--470, Academic Press, New York
2 Jenkin, P.M. (1966) Ann. Endocrinol. 27, 331--341 3 Gilbert, L.I. (1974) Recent Prog. Horm. Res. 30, 347--390 4 Sekeris, C.E., Karlson, P. and Congote, L.F. (1971) in The A c t i o n o f H o r m o n e s , Genes to Popula t ion
(Poa, P.P., ed.), pp. 7--19, Charles C. Thomas, Springfield :, 5 Krishnakumaran, A., Berry, S.J., Oberlander, H. and Schneiderman, H.A. (1967) J. Insect Physiol. 13,
1--57 6 Neufeld, G.J., Thomson, J.A. and Horn, D.S.H. (1968) J. Insect Physiol. 14, 789--804 7 Doane, W.W. (1973) in D e v e l o p m e n t a l S y s t e m s : Insec ts (Counce, S.J. and Waddington, C.H., eds~),
Vol. 2, pp. 291--466, Academic Press, New York 8 Shaaya, E. and Clever, U. (1972) Biochim. Biophys. Acta 272, 373--381 9 Karison, P. and Shaaya, E. (1964) J. Insect Physiol. 10, 797--804
10 Loening, V.E. (1967) Biochem. J. 102, 251--257 11 Shaaya, E. (1976) Anal. Biochem., 74 in the press 12 Shaaya, E. (1969) Z. Naturforsch. 24b, 718--721 13 Shaaya, E., Schnieders, B., Kunz, W. and Sekeris, C.E. (1971) Insect Biochem. 1, 113--121 14 Shaaya, E. (1976) Insect Biochem., in the press 15 Mendecki, J., Lee, S.Y. and Brawerman, G. (1972) Biochemistry 11 ,792 - -798 16 Molly, G.R., Spron, M.B., Kelly, D.E. and Perry, R.P. (1972) Biochemistry 11, 3256--3260 17 Imaizumi, I., Diggelmann, H. and Scherrer, K. (1973) Prnc. Natl.. Acad. Sci. U.S. 70, 1122--1126 18 Ruiz-Carillo, A., Beato, M., Schutz, G., Fiegeison, P. and Allfrey, V.G. (1973) Proc. Natl. Acad. Sci.
U.S. 70, 3641--3645 19 Shaaya, E. and Karlson, P. (1965) J. Insect Physiol. 11, 65--69 20 Shaaya, E. and Karlson, P. (1965) Develop. Biol. 11, 424--432 21 Shaaya, E. and Levinson, Z. (1966) Riv. Parassitol. 82, 211--215 22 Williams, C.M. (1961) Biol. Bull. 121, 572--585 23 Willis, J.H. (1969) J. Embryol . Exp. Morphol. 22, 27---44 24 Marmaras, V.J. and Sekeris, C.E. (1972) Exp. Cell Res. 75, 143--153 25 Clever, U., Clever, I., Storbeck, I. and Young, N. (1973) Dev. Biol. 31, 47--60 26 Oberlander, H. (1972) J. Insect Physiol. 18, 223--228 27 Courgeon, A.M. (1972) Nature 238, 250--251





























