Embed Size (px)
Citation preview

www.elsevier.com/locate/hydromet
Hydrometallurgy 71 (2003) 301–309
Geomicrobiology of the Tinto River, a model of interest for
biohydrometallurgy
E. Gonzalez-Torila,*, F. Gomezb, N. Rodrıguezb, D. Fernandez-Remolarb,J. Zuluagac, I. Marına, R. Amilsa,b
aCentro de Biologıa Molecular, UAM-CSIC, Cantoblanco, Madrid 28049, SpainbCentro de Astrobiologıa, INTA-CSIC, Torrejon de Ardoz 28850, Spain
cDepartamento de Quımica Fısica Aplicada, UAM, Cantoblanco, Madrid 28049, Spain
Abstract
The Tinto River (Huelva, southwestern Spain) is an extreme environment with a constant acidic pH (mean 2.3), a high
concentration of heavy metals and a remarkable level of microbial diversity (bacteria, archaea, photosynthetic and heterotrophic
protists, yeast and filamentous fungi). The extreme conditions found in the river are the direct consequence of the active
metabolism of chemolithotrophic microorganisms thriving in the rich polymetallic sulfides present in high concentrations in the
Iberian Pyritic Belt. Primary production in the river is driven mainly by oxygenic photosynthesis (protists and cyanobacteria),
although an important part is also due to the activity of chemolithotrophic prokaryotes. Conventional and molecular ecology
techniques were used to study the microbial ecology of the Tinto system. The results of both methods agreed. Although sulfur
metabolism plays an important role in the system, iron seems to be the key element in this habitat. Iron is not only an important
substrate for the rich population of iron oxidizing prokaryotes, but also an electron acceptor for anaerobic respiration in the
anoxic parts of the river. It is also responsible for the maintenance of a constant acidic pH (probably critical for biodiversity) and
for radiation protection. Laminar iron stromatolitic formations can be found along the river. These structures are related to
massive laminated bioinduced iron formations found at different elevations above the current river. The isotopic dating of these
formations leads to the conclusion that the Tinto River corresponds to a natural system and not to an industrial, contaminated
site. A geomicrobiological model of this habitat encompassing most of the geological, physical, chemical and biological
variables is presented and its biohydrometallurgical implications discussed.
D 2003 Elsevier B.V. All rights reserved.
Keywords: Geomicrobiology; Chemolithotrophy; Tinto River; Iron; Acidic pH
1. Introduction
Extremophilic microorganisms capable of develop-
ing in extreme environments have recently attracted
considerable attention because of the challenge that
their discovery has posed to our current notions of the
0304-386X/$ - see front matter D 2003 Elsevier B.V. All rights reserved.
doi:10.1016/S0304-386X(03)00169-5
* Corresponding author.
limits of life and its possible origin, but, most impor-
tantly, because of their biotechnological potential. Our
research group has characterized the microbial life of
an acidic habitat, the Tinto River, a 100-km long
acidic river containing a high concentration of heavy
metals in solution, which originates at Pena de Hierro,
at the core of the Rıo Tinto mining district, and flows
into the Atlantic Ocean at Huelva. The river gives its
name to an important mining site, which has been in

E. Gonzalez-Toril et al. / Hydrometallurgy 71 (2003) 301–309302
operation for more than 5000 years. In the past, the
extreme conditions of the river were considered the
consequence of intense mining activity in the area.
Conventional microbial ecology studies showed
that both sulfur and iron oxidizing bacteria are present
at rather high concentrations along the river (Gonza-
lez-Toril et al., 1999; Lopez-Archilla et al., 1993,
2001). In spite of the low pH values and the
corresponding high concentration of heavy metals,
an unexpected level of eukaryotic diversity has been
described in the system (Lopez-Archilla et al., 2001).
Classical microbial analysis has a strong bias as a
result of the absolute requirement of isolation of
microorganisms in selective media prior to their
characterization. The introduction of molecular biol-
ogy techniques, mainly in situ hybridization using
fluorescent probes (FISH) and denaturating gradient
gel electrophoresis (DGGE) enable the resolution of
PCR amplified rDNA using primers with different
phylogenetic specificity, has produced an authentic
revolution in microbial ecology, in general, and in the
study of extreme environments, in particular. The
combination of classical isolation with molecular
ecology techniques has advanced a geomicrobiolog-
ical model of the Tinto system, which has important
implications in biohydrometallurgy.
2. Materials and methods
2.1. Sampling and analysis of physico-chemical
parameters
Samples were collected in triplicate from different
sampling stations along the river. Total content of
metals was measured by atomic absorption spectro-
photometry and X-ray fluorescence reflection. Sulfate
concentrations were determined by a turbidimetric
method (Keith, 1996a) and ferrous iron by a colori-
metric method (Keith, 1996b). Conductivity, pH,
oxygen and redox potential values were measured in
situ using specific electrodes.
2.2. Isolation and phenotypic characterization of
microorganisms
The isolation and characterization of chemolitho-
trophic and heterotrophic bacteria, archaea, photosyn-
thetic and heterotrophic protists, yeast and filamentous
fungi were performed as previously described (Lopez-
Archilla et al., 2001).
2.3. Fluorescence in situ hybridization
The basic protocol described by Amann (1995)
with some modifications required for its adaptation to
acidic environments has been used throughout the
work. One milliliter of each sample was denaturated
with 0.1 ml of formaldehyde solution (37%) and kept
for 4 h at 4 jC. Samples were filtered through GTTP
Millipore filter (0.22 Am) and washed with 10 ml of
minimal Mackintosh medium (Mackintosh, 1978) to
eliminate excess of formaldehyde and heavy metals,
followed by a 10-ml wash with PBS buffer (130 mM
NaCl, 10 mM sodium phosphate pH 7.2) and air
dried. The following fluorescent probes were used:
EUB338 (GCTGCCTCCCGTAGGAGT) for bacteria,
Non338 (ACTCCTACGGGAGGCAGC) as a nega-
tive control, GAM42a (GCCTTCCCACATCGTTT)
for g-proteobacteria, Bet42a (GCCTTCCCACTTCG-
TTT) for h-proteobacteria, Thio1 (GCGCTTTCTGG
GGTCTGC) for Acidithiobacil lus , ALF968
(GGTAAGGTTCTGCGCGTT) for a-proteobacteria,
NTR712 (CGCCTTCG CCACCGGCCTTCC) for
members of the Nitrospira group and FER656
(CGTTTA ACCTCAC CCGATC) for Ferroplasma.
2.4. Denaturating gradient gel electrophoresis
One liter of water from selected sampling stations
was filtered through a 0.22-Am Millipore filter and
washed with Mackintosh minimal media at pH 1.5.
Filters were kept at 4 jC until used. A piece of filter
membrane was treated with 2 ml of SET buffer (25%
saccharose, 50 mM Tris pH 8, 2 mM EDTA) over-
night at � 20 jC. After thawing, the sample 120 Al ofSDS (25%) and 200 Al of pronase (2 mg/ml) were
added and the mixture incubated at 4 jC for 30 min.
Each sample was extracted with 6 ml of PCI (phenol/
chloroform/isoamyl alcohol, 24:24:1) and centrifuged
at low speed to separate the phases. A volume of 0.5
ml of sodium acetate 2 M, pH 5.2 was added to the
water phase and the extraction with 6 ml of PCI
repeated until complete elimination of the interphase.
The nucleic acids were precipitated by addition of 2.5
volumes of ice-cold ethanol (96%) at � 20 jC for 3 h.

E. Gonzalez-Toril et al. / Hydrometallurgy 71 (2003) 301–309 303
The pellet was washed with 1 ml of ice-cold ethanol
(70%), centrifuged at low speed, vacuum-dried and
resuspended in water (MQ). To facilitate a comparison
with the results obtained with FISH, RNA rather than
DNA amplification is convenient. To do so, 50 Al ofeach sample was treated with RNAse-free DNAseI
prior to its amplification with M-ML V reverse tran-
scriptase. The protocols for rDNA amplification and
DGGE were those described by Muyzer et al. (1996).
Each band was amplified and sequenced. Sequences
were analyzed with the BLAST program using the
NCBI data bank and the ARB program using the
Technical University of Munich data bank. The fol-
lowing primers were used for specific amplification:
GM5-GC (CCTAC GGGAGGCAGCAG) and
907RM (CCGTCAATTCMTTTGAGTTT) for
bacterial rDNA, ARC344F-GC (ACGGGGYGCA-
GGCGCGA) and ARC907R (GTGCTCCCGC
GCCAATTCCT) for archaeal rDNA and CYA781R
(GACTACTGGGGTATCTAATCCCWATT) and
CYA781R (GACTACTGGGGTATCTAAYCCCW-
TT) for cyanobacterial and chloroplast rDNA.
3. Results
3.1. Microbial ecology of the Tinto River
The basin of the Tinto River covers an area of 1676
km2. The gentle slope of the river (0.6%) facilitates
the settlement of a dense microbial community on the
riverbed. The river is subject to a Mediterranean type
regime with an extremely variable flow depending on
the season. The highest flow values are reached
during winter (8.1 m3/s) and the lowest during the
summer (0.07 m3/s). Representative values of some of
Table 1
Quantitative ecology of prokaryotes at different sampling sites using FISH
Sampling site Bacteria a-prot h-prot g-p
Origin I 75 – 41 –
Origin II 63 3 – 54
Intersection 67 4 1 9
Anabel’s Garden 70 17 3 15
3.2 69 51 – 1
Berrocal 88 3 – 64
La Palma 43 26 – –
Abbreviations: a-prot =a-proteobacteria, h-prot =h-proteobacteria, g-prot
the physico-chemical parameters measured in differ-
ent sampling stations are shown in Table 1. Comple-
mentary information has been published recently
(Lopez-Archilla et al., 2001). The pH remains low
and rather constant year-round (mean 2.3), regardless
of the temperature or the river flow. This is the
consequence of the buffer effect produced by the
presence of high concentrations of ferric iron along
the river:
Fe3þ þ H2OZFeðOHÞ2þ þ Hþ
FeðOHÞ2þ þ H2OZFeðOHÞþ2 þ Hþ
FeðOHÞþ2 þ H2OZFeðOHÞ3 þ Hþ
When the river is diluted by tributaries or rain,
ferric hydroxide is formed and protons are released,
thus maintaining the acidic pH of the system. During
the summer, when extremely high temperatures are
reached, intensive evaporation is produced, but the pH
is maintained constant because the reverse reaction
operates: ferric hydroxide precipitates dissolve, con-
suming protons.
The Tinto River ecosystem is unique in that its
biological community is exclusively microbial. Mul-
ticellular eukaryotes have not been detected in any of
the sampling stations. In order to elucidate the degree
of biodiversity conventional microbiological methods
were used for the isolation and characterization of
the microorganisms present in the habitat. To date,
we have identified and characterized chemolithotro-
(in %)
rot Nitros Archaea pH Total Fe (g/l)
– – 2.9 0.8
31 1 1.8 19.8
27 – 2.7 9.6
21 5 1.9 2.3
27 – 1.9 1.8
2 – 2.4 3.1
14 1 2.2 0.9
= g-proteobacteria, Nitros =Nitrospira.

E. Gonzalez-Toril et al. / Hydrometallurgy 71 (2003) 301–309304
phic bacteria and archaea (responsible for the ex-
treme conditions of the system), heterotrophic bacte-
ria, heterotrophic and photosynthetic protists, yeasts
and filamentous fungi. A summary of the isolated
prokaryotic microorganisms is presented in Table 2.
The different microbial populations found in the
Tinto River can be grouped according to their
ecological role as primary producers, decomposers
and consumers.
3.1.1. Primary producers
Chemolithotrophic prokaryotes, mainly iron and
sulfur oxidizing bacteria, together with sulfur oxidiz-
ing archaea have been isolated from the Tinto eco-
system (Lopez-Archilla et al., 2001). The iron
oxidizing bacteria have been identified as different
strains of Acidithiobacillus ferrooxidans (formerly
Thiobacillus ferrooxidans) and Leptospirillum fer-
rooxidans based on their morphology, metabolism
and low-frequency restriction fragment analysis
(LFRFA) profiles (Gonzalez-Toril et al., 1999). In
addition to the members of the Acidithiobacillus
genus, other sulfur oxidizing bacteria unable to oxi-
dize iron have been characterized. Chemolithoauto-
trophic archaea related to the Sulfolobales have been
also isolated in the Tinto. Algae accounted for the
Table 2
Comparative prokaryotic ecology of the Tinto River using different
techniques
Isolation DGGE FISH
Acidithiobacillus
ferrooxidans
(different LFRFAsa)
Acidithiobacillus
ferrooxidans
(different sequences)
+ + +
Acidithiobacillus
thiooxidans
Acidithiobacillus
thiooxidans
?
Leptospirillum
ferrooxidans
Leptospirillum
ferrooxidans
+ + +
? Ferroplasma
acidiphilum
+
Acidiphilium sp. Acidiphilim
cryptum
++
Different
Bacillus species
– ?
? Acidimicrobium
ferrooxidans
?
sulfate reducing
? ? related bacteria
? cyanobacteria ?
a LFRFAs: low-frequency restriction fragment analysis.
greatest proportion of biomass (over 65%) in the
system. Together with the chemolithoautotrophic bac-
teria, algae constitute the primary producers of the
system. Members of the Chlorophyta (Klebsormi-
dium, Zignema, Chlamydomonas), Euglenophyta (Eu-
uglena), Bacillariophyta (Diatoms) and Rhodophita
(Galdieria) phyla have been observed and some of
them isolated (Lopez-Archilla et al., 2001).
3.1.2. Decomposers
A high number of heterotrophic bacteria were
isolated from enrichment cultures (Lopez-Archilla et
al., 2001), but many of them did not grow after the
second or third transfer, probably because some com-
ponents of the original inoculum were diluted out,
affecting their growth. Some of the isolates corre-
sponded to the genus Acidiphilium. Members of this
genus have been shown to be frequently associated
with chemolithoautotrophic bacteria, especially iron
oxidizers. Many bacterial isolates were Gram-positive
bacilli, aerobic spore formers of the genus Bacillus.
Within the decomposers, fungi were very abundant
and exhibited great diversity, including yeasts and
filamentous forms. A high percentage of the hypho-
mycetes isolates were able to grow in the Tinto River
conditions. Some of the yeast species isolated can also
be found in other less extreme aquatic environments.
But the isolated Dematiaceae seem to be specific to
the extreme conditions of the habitat, since they are
rarely present in neutral fresh waters (Lopez-Archilla
et al., 2001).
3.1.3. Consumers
Among the eukaryotes, heterotrophic protists con-
stitute the major consumer group in the Tinto ecosys-
tem. Different flagellates (phylum Zoomastigina),
amoebae of the class Lobosea (phylum Rhizopoda),
some representatives of the class Heliozoa (phylum
Actinopoda) and ciliates (Phylum Ciliophora) have
been observed mainly associated to the biofilms
(Lopez-Archilla et al., 2001).
3.2. Bioinduced iron formations in the Tinto ecosystem
Most of the biomass is located on the riverbed in
the form of dense, compact biofilms, mainly com-
posed of filamentous algae and fungi in which bacte-
ria are trapped. Also, heterotrophic protists can be

Fig. 1. Fluorescent in situ hybridization of a sample from the origin
of the Tinto River using (a) DAPI, universal stain, and (b) a specific
probe for Nitrospira (NTR712).
E. Gonzalez-Toril et al. / Hydrometallurgy 71 (2003) 301–309 305
found associated to the biofilms. Significant mineral
precipitation is observed on the surface of the bio-
films, forming iron stromatolites, which grow season-
ally following the exposure of the biofilms to the
water. As mentioned, the system has been altered by
mining activities during the last 5000 years, but
evidence of its antiquity, older than 0.3 My, has been
found in massive laminated iron beds in three iron
rock levels occupying different elevations above the
present river. One of them corresponds to an extensive
formation, more than 35 m above the river and over
10 m thick, organized into two different lithostrati-
graphic units, one with paraconglomerate and massive
facies and the other with laminar-stromatolitic, mas-
sive microbiolitic, as well as some plant fossils. If the
morphology of the outcrops, the strata thickness and
continuity are taken into account, this formation could
be interpreted as an acid lake generated by the
metabolic activity of chemolithotrophic microorgan-
isms. The observation of the actual river iron laminae
allows us to understand the origin of these facies and
their environmental significance.
3.3. Molecular ecology of the Tinto River
As mentioned in the introduction, the use of con-
ventional microbiological techniques has important
limitations for a complete enumeration of the microbial
diversity present in any given system and, most impor-
tant, for the quantification of their populations. The
introduction of PCR based technologies prompted the
accumulation of rDNA sequences from many different
microorganisms, thus allowing the development of
complementary molecular ecology techniques like
FISH and DGGE. One of the goals of this work was
to use in situ hybridization techniques complemented
with DGGE for identification and quantification of
microorganisms present in different sampling stations
along the river and to compare the results with those
obtained using conventional methodologies.
Fig. 1 shows an in situ hybridization performed
with a fluorescent probe specific for members of the
Nitrospira group (in which members of the genus
Leptospirillum are included) compared with DAPI
(universal) stain. The use of probes with different
specificity to hybridize samples collected along the
river generated data on the population of prokaryotic
microorganisms present in the Tinto system. Table 1
shows quantitative FISH results obtained with differ-
ent probes at different sampling sites along the river.
Complementary information was obtained from the
sequence of the different rDNAs amplified by PCR
using selected primers with different specificity and
separated by DGGE. Fig. 2 shows the rDNA bands
resolved after amplification using bacterial and ar-
chaeal universal primers of RNA samples obtained
from different sampling stations.
The diverse bands can be excised, amplified,
sequenced and compared using different programs
(BLAST, ARB) with the rDNA sequence data banks.
Any RNA donor whose homology is greater than 96%
was considered a member of the same species as the
reference sequence. Both techniques, FISH and
DGGE, are complementary in the sense that DGGE
allows possible candidates to be identified and spe-
cific fluorescent probes for in situ hybridization to be
designed. Table 2 shows a summary of comparative
results obtained using conventional and molecular
ecology techniques. It should be noted that there is
substantial agreement among all of them.

Fig. 2. DGGE of amplified rDNA fromRNA extracted from different
river locations : (1) origin (II), (2) origin (I), (3) intersection and (4)
berrocal, using primers with different specificity: Archaea and
Bacteria.
E. Gonzalez-Toril et al. / Hydrometallurgy 71 (2003) 301–309306
Interestingly enough, using DGGE we were able to
amplify rDNAs which according to their sequence
clustered together with sulfate reducing bacteria. Also,
using specific primers for cyanobacteria and chloro-
plast, we amplified several bands whose sequence
clustered with cyanobacteria, strongly suggesting their
presence in the Tinto ecosystem. We are currently
trying to isolate both types of bacteria from enrich-
ment cultures.
4. Discussion
The objective of this work was to compare the
results obtained using conventional microbial techni-
ques with those derived from molecular ecology
techniques, and to generate a functional model of
the Tinto River system. The particular geology and
climatology of the region favor the creation of the
Tinto River’s special environment, which provide the
base on which the biological communities establish
themselves and proliferate. The river rises in the
middle of the Iberian Pyritic Belt, one of the world’s
richest complex polymetallic sulfide deposits. The
abundance of sulfide minerals, mainly pyrite and
chalcopyrite, provides the necessary substrate for the
development of chemolithotrophic bacteria, responsi-
ble for the extreme conditions of the habitat. The high
water table, which has been a serious hindrance to the
exploitation of the mines in the past, maintains the
river flow during the extremely hot summers, in the
absence of rain and with a high rate of evaporation.
The Tinto River originates at Pena de Hierro from
very close tributaries, which have quite different
microbial content, probably related to the character-
istics of the mineral substrates that feed them. ‘‘Origin
I’’ is less acidic (mean 2.9) and has a rather low
content in iron (mean 0.8 g/l) when compared to
‘‘Origin II’’, which has a higher concentration of
oxidized iron (mean 19.8 g/l) and is more acidic
(mean pH 1.8). In the first case, there is a high
concentration of unidentified bacteria belonging to
the h-proteobacteria, which might have a strong sulfur
oxidizing capability, because of the high content of
sulfate in the water and its pH. ‘‘Origin II’’ holds a
high concentration of different strains of At. ferroox-
idans and L. ferrooxidans. This site seems to be the
output of a large underground reactor, which may be
in contact with an acidic lake formed in an abandoned
open mine pit (Pena de Hierro). This acidic lake has a
diameter of about 500 m and is 50 m deep. Prelim-
inary results indicate that this lake is anoxic. Because
of its difficult accessibility, a minisubmarine has been
designed to explore the characteristics of this interest-
ing opaque habitat in collaboration with the Centro de
Astrobiologıa. This lake is a good indicator of the
water table level in the area, allowing the problems
that the water table caused to past and present mining
activities to be explained, and raises serious questions
about the flooding of open pits as a safe environmen-
tal practice in sulfidic mine closing projects.
The ‘‘Intersection’’ sampling site refers to the area
in which both streams (Origin I and II) join with an
almost neutral tributary (pH between 3.9 and 4.5). The
microbial content of the ‘‘Intersection’’ site and its
physico-chemical conditions correspond to the differ-
ent contributions of these streams to the river. Another
interesting observation can be made at the sampling
station known as ‘‘Anabel’s Garden’’. This site was
considered in the beggining of the work as the origin
of the Tinto system, although careful Global Position-
ing System (GPS) measurements have proven that it is
located at a lower altitude than the ‘‘Origin’’. ‘‘Ana-
bel’s Garden’’ is the origin of a tributary that crosses
the modern Rıo Tinto mine operation and was used by
the British company as a source of acidic waters for

E. Gonzalez-Toril et al. / Hydrometallurgy 71 (2003) 301–309 307
direct copper extraction from the river using cemen-
tation procedures. This tributary has a broader spec-
trum of microbial diversity, not only at the prokaryotic
level (Table 2), but also at the eukaryotic one, exhib-
iting important blooms of algae, mainly Diatoms and
Euglenas, especially during spring and summer.
This stream gives rise to the ‘‘3.2’’ site, which
corresponds to a reservoir of acidic waters used for
cementation. This reservoir was originally 12 m deep.
Today, it is half full of sediment. Interestingly enough,
the microbial composition of this site is rather differ-
ent from the ‘‘Anabel’s Garden’’ site which feeds it.
There is an important increase in cell density, and in
relation to prokaryotes the number of bacteria that
give a positive signal with the a-proteobacteria probe
is rather high, whereas the concentration of At. fer-
rooxidans seems rather low. An interesting feature of
this reservoir is that from several centimeters below
the surface to its bottom it is completely anoxic,
which might explain the difference in bacterial pop-
ulation observed in its waters.
The ‘‘Berrocal’’ site is representative of the middle
course of the river. This site gives reproducible results
in which the proportion of At. ferrooxidans is rather
high when compared to other sampling sites along the
river. This is the part of the river in which important
fields of iron stromatolites can be found. Also, the
flow of the river in this section is rather slow,
facilitating the formation of biofilms and their
corresponding mineral precipitation, and produces in
many places (especially in the meanders) deep basins,
which are basically anoxic. Finally, the sampling
station of ‘‘La Palma’’, near the mouth of the river,
shows that more than fifty per cent of the biota at this
site is eukaryotic. Large numbers of filamentous algae
thrive on the surface of the rocks where dark ferric
iron waters do not impede sunlight penetration. There
are some important and reproducible differences be-
tween ‘‘La Palma’’ and ‘‘Berrocal’’ in terms of their
prokaryotic populations, which are probably related to
the physico-chemical differences between both sites,
mainly a lower content of heavy metals, especially
iron, in the lowest part of the river.
DGGE techniques amplified two sets of signatures
from RNA samples from several sites which clustered
with bacteria with interesting properties. One of these
corresponds unequivocally to cyanobacterial sequen-
ces. This observation is interesting because up to now
no one has been able to isolate a cyanobacteria that
can grow at acidic pH. So far, we have been unable to
identify the possible candidates and to isolate them.
One of the bands has a sequence 93% homologous
with Stanieria cyanosphaera, others cluster with un-
identified cyanobacteria. The other signature clusters
with sulfate reducing bacteria. For years, our group
has been interested in the isolation of acidic sulfate
reducing bacteria to explain the recycling of reduced
sulfur substrates for use by strict chemolithotrophic
bacteria along the main basin of the Tinto River.
Knowledge of the area in which the signal has been
collected and the sequence of its rDNA should help in
the design of tools to isolate these bacteria which have
important biotechnological applications (Sen and
Johnson, 1999).
As mentioned, both sulfur and iron have a funda-
mental role in this fluvial ecosystem. Of these, much
emphasis has been given in the literature to the sulfur
cycle bacteria, probably because from a biochemical
point of view much more energy can be obtained from
its reduced forms. Interestingly enough, iron bacteria
have recently acquired a more predominant role in
environmental microbiology since the discovery of
the importance of iron in bioleaching operations
(Brierley, 2001), its use as reducing power for anaer-
obic photosynthesis (Ehrenreich and Widdel, 1994)
and its use as substrate for anaerobic respiration
(Straub et al., 1996). We believe that the Tinto River
is a system controlled by iron (Fig. 3). Iron is not only
a source of energy for iron oxidizing prokaryotes (At.
ferrooxidans, L. ferrooxidans and Ferroplasma acid-
iphilum), but also can be used as an electron acceptor,
both in aerobic (At. thiooxidans and Acidiphilium sp.)
and anaerobic conditions (At. ferroxidans). In addi-
tion, ferric iron is responsible for the maintenance of a
constant pH in the ecosystem, a property that must be
important to the maintenance of a high level of
eukaryotic diversity found in the system. Finally, acid
ferric iron solutions provide good UV irradiation
protection.
We know that iron is an important limiting factor
for the growth of phytoplankton in the oceans
(Martin, 1990). We know that most cells, both
prokaryotes, but especially eukaryotes devote an
important amount of genetic information and energy
to scavenging iron. A possible advantage to the
photosynthetic protists that thrive in the extreme
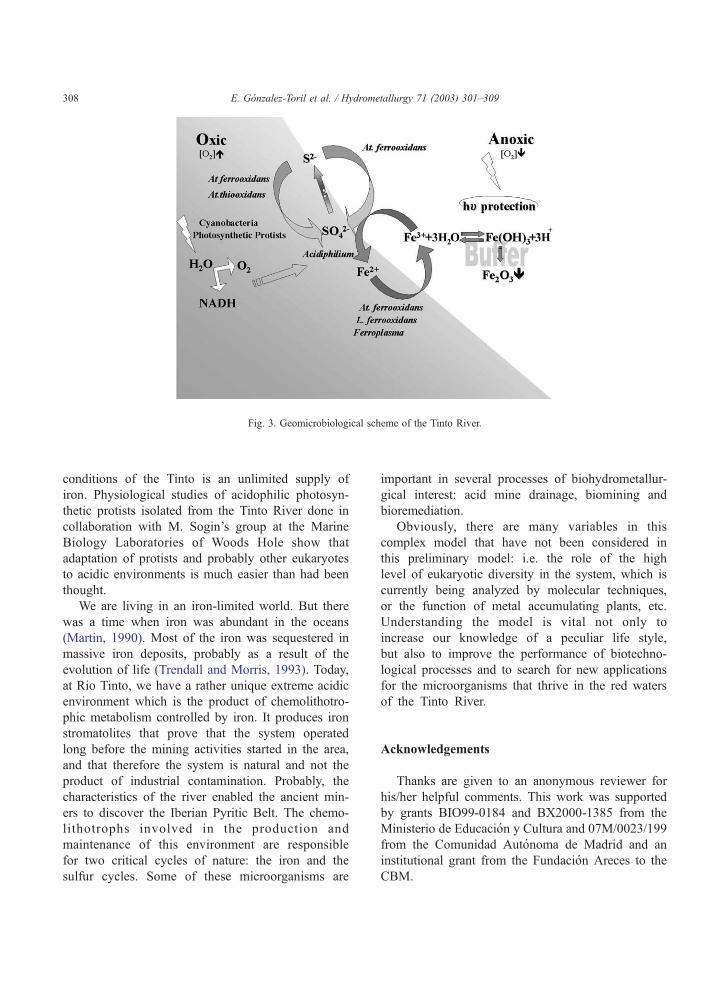
Fig. 3. Geomicrobiological scheme of the Tinto River.
E. Gonzalez-Toril et al. / Hydrometallurgy 71 (2003) 301–309308
conditions of the Tinto is an unlimited supply of
iron. Physiological studies of acidophilic photosyn-
thetic protists isolated from the Tinto River done in
collaboration with M. Sogin’s group at the Marine
Biology Laboratories of Woods Hole show that
adaptation of protists and probably other eukaryotes
to acidic environments is much easier than had been
thought.
We are living in an iron-limited world. But there
was a time when iron was abundant in the oceans
(Martin, 1990). Most of the iron was sequestered in
massive iron deposits, probably as a result of the
evolution of life (Trendall and Morris, 1993). Today,
at Rio Tinto, we have a rather unique extreme acidic
environment which is the product of chemolithotro-
phic metabolism controlled by iron. It produces iron
stromatolites that prove that the system operated
long before the mining activities started in the area,
and that therefore the system is natural and not the
product of industrial contamination. Probably, the
characteristics of the river enabled the ancient min-
ers to discover the Iberian Pyritic Belt. The chemo-
lithotrophs involved in the production and
maintenance of this environment are responsible
for two critical cycles of nature: the iron and the
sulfur cycles. Some of these microorganisms are
important in several processes of biohydrometallur-
gical interest: acid mine drainage, biomining and
bioremediation.
Obviously, there are many variables in this
complex model that have not been considered in
this preliminary model: i.e. the role of the high
level of eukaryotic diversity in the system, which is
currently being analyzed by molecular techniques,
or the function of metal accumulating plants, etc.
Understanding the model is vital not only to
increase our knowledge of a peculiar life style,
but also to improve the performance of biotechno-
logical processes and to search for new applications
for the microorganisms that thrive in the red waters
of the Tinto River.
Acknowledgements
Thanks are given to an anonymous reviewer for
his/her helpful comments. This work was supported
by grants BIO99-0184 and BX2000-1385 from the
Ministerio de Educacion y Cultura and 07M/0023/199
from the Comunidad Autonoma de Madrid and an
institutional grant from the Fundacion Areces to the
CBM.

E. Gonzalez-Toril et al. / Hydrometallurgy 71 (2003) 301–309 309
References
Amann, R.I., 1995. In situ identification of microorganisms by
whole cell hybridization with rRNA-targeted nucleic acid
probes. In: Akkermans, A.D.L., Van Elsas, J.D., De Bruijn,
F.J. (Eds.), Molecular Microbial Manual. Kluwer Academic
Publishers, Dordrecht, pp. 1–15. 3.3.6.
Brierley, C.L., 2001. Bacterial succession in bioheap leaching. Hy-
drometallurgy 59, 249–255.
Ehrenreich, E., Widdel, F., 1994. Anaerobic oxidation of ferrous
iron purple bacteria, a new type of photosynthetic metabolism.
Appl. Environ. Microbiol. 60, 4517–4526.
Keith, L.H. (Ed.), 1996a. EPA’s Sampling and Analysis Methods,
2nd ed. CRC, Lewis Publishers, Boca Raton, p. 1036.
Keith, L.H. (Ed.), 1996b. EPA’s Sampling and Analysis Methods,
2nd ed. CRC, Lewis Publishers, Boca Raton, p. 1424.
Gonzalez-Toril, E., Gomez, F., Irazabal, N., Amils, R., Marın, I.,
1999. Comparative genomic characterization of iron oxidizing
bacteria isolated from the Tinto River. In: Amils, R., Ballester,
A. (Eds.), Biohydrometallurgy and the Environment towards the
Mining of the 21st century, vol. B. Elsevier, Amsterdam,
pp. 149–157.
Lopez-Archilla, A.I., Marın, I., Amils, R., 1993. Bioleaching and
interrelated acidophilic microorganisms from Rıo Tinto, Spain.
Geomicrobiol. J. 11, 223–233.
Lopez-Archilla, A.I., Marın, I., Amils, R., 2001. Microbial com-
munity composition and ecology of an acidic aquatic environ-
ment: the Tinto River, Spain. Microb. Ecol. 41, 20–35.
Mackintosh, M.E., 1978. Nitrogen fixation by Thiobacillus ferroox-
idans. J. Gen. Microbiol. 105, 215–218.
Martin, J.H., 1990. Glacial-interglacial CO2 change: the iron hy-
pothesis. Paleoceanography 5, 1–13.
Muyzer, G., Hottentrager, S., Teske, A., Wawer, C., 1996. Denatur-
ing gradient gel electrophoresis of PCR-amplified 16S rDNA.
A new molecular approach to analyse the genetic diversity of
mixed microbial communities. In: Akkermans, A.D.L., Van El-
sas, J.D., De Bruijn, F.J. (Eds.), Molecular Microbial Manual.
Kluwer Academic Publishers, Dordrecht, pp. 1–23. 3.4.4.
Sen, A.M., Johnson, B., 1999. Acidophilic sulfate-reducing bacte-
ria: candidates for bioremediation of acid mine drainage.
In: Amils, R., Ballester, A. (Eds.), Biohydrometallurgy and
the Environment Towards the Mining of the 21st Century, vol.
A. Elsevier, Amsterdam, pp. 709–718.
Straub, K.L., Benz, M., Schink, B., Widdel, F., 1996. Anaerobic,
nitrate-dependent microbial oxidation of ferrous iron. Appl. En-
viron. Microbiol. 62, 1458–1460.
Trendall, A.F., Morris, R.C., 1993. Iron-Formation: Facts and Prob-
lems. Elsevier, Amsterdam.





























