Embed Size (px)
Citation preview

BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofit publishers, academic institutions, research libraries, andresearch funders in the common goal of maximizing access to critical research.
Genotype and soil nutrient environment influence aspen litter chemistry and in-streamdecompositionAuthor(s): Carri J. LeRoyStuart C. Wooley and Richard L. LindrothSource: Freshwater Science, 31(4):1244-1253. 2012.Published By: The Society for Freshwater ScienceDOI: http://dx.doi.org/10.1899/12-029.1URL: http://www.bioone.org/doi/full/10.1899/12-029.1
BioOne (www.bioone.org) is a nonprofit, online aggregation of core research in the biological, ecological, and environmentalsciences. BioOne provides a sustainable online platform for over 170 journals and books published by nonprofit societies,associations, museums, institutions, and presses.
Your use of this PDF, the BioOne Web site, and all posted and associated content indicates your acceptance of BioOne’s Terms ofUse, available at www.bioone.org/page/terms_of_use.
Usage of BioOne content is strictly limited to personal, educational, and non-commercial use. Commercial inquiries or rights andpermissions requests should be directed to the individual publisher as copyright holder.

Genotype and soil nutrient environment influence aspen litterchemistry and in-stream decomposition
Carri J. LeRoy1
Environmental Studies Program, Evergreen State College, Olympia, Washington 98505 USA
Stuart C. Wooley2AND Richard L. Lindroth3
Department of Entomology, University of Wisconsin Madison, Madison, Wisconsin 53706 USA
Abstract. A growing body of genes-to-ecosystems research has documented the ecosystem-levelconsequences of intraspecific variation in plants caused in large part by variation in chemical composition.Understanding how this genetic variation in trees might interact with elevated nutrients resulting fromincreases in anthropogenic deposition may give us insight into how future riparian forests might influenceadjacent streams through leaf-litter deposition. We examined the effects of tree genotype, soil nutrientenvironment, and their interaction on aspen (Populus tremuloides) litter chemistry and aquaticdecomposition. We used litter collected from 5 aspen genotypes grown in a common garden under lowand high nutrient availability and monitored decomposition over 112 d in a woodland stream. Genotype,environment, and genotype 3 environment interactions influenced litter chemistry and decompositiondynamics. Genotype and environment both strongly influenced initial litter chemistry, with significantgenotype 3 environment interactions for bound condensed tannins and C:N. Consistent with effects onlitter chemistry, decomposition rates were significantly affected by genotype, environment, and genotype3 environment interactions. These results suggest that future changes in intraspecific genetic variation oftree species and deposition of nutrients because of shifts in climate, land use, and related factors mayinfluence decomposition processes in terrestrial systems and in the aquatic systems with which they arecoupled via material transfer.
Key words: aquatic decomposition, aspen, genotypic variation, genotype 3 environment, G 3 E,nutrients, litter chemistry, Populus tremuloides.
A long history of research on leaf-litter inputs tostreams (Petersen and Cummins 1974, Webster andBenfield 1986, Tank et al. 2010, and many others)documents that plant traits, such as leaf toughness,nutrients (N and P), structural compounds (lignin,cellulose), and secondary compounds (e.g., tannins),explain significant variation in aquatic leaf-litterdecomposition rates. Variation in plant chemistrycan be interspecific or intraspecific (LeRoy et al. 2006,2007, Lecerf and Chauvet 2008) and can be altered byenvironmental conditions. Thus, under elevated nu-trient conditions caused by anthropogenic nutrientdeposition these important litter traits may change.
Meta-analysis has shown no overriding influence of Nadditions on decomposition (Knorr et al. 2005), butelevated N generally decreases C:N ratios in leaf litter.This shift can accelerate terrestrial decomposition forlow-lignin litters (Norby 1998, Hobbie 2000) butdecelerate decomposition for high-lignin litters be-cause of suppression of ligninolytic enzymes (Fog1988, Magill and Aber 1998, Carreiro et al. 2000). Inaddition, genetic variation in populations of ripariantrees is being reduced (Ledig 1992) by massive habitatlosses in riparian zones. The interaction of decreasedgenetic variation and increased nutrients may alterlitter quality and subsequent decomposition rates inforests (Madritch et al. 2006) and adjacent streamenvironments. Exploration of these important geno-type 3 environment interactions may further ourunderstanding of the ecosystem-level consequencesof genetic variation and the propensity of geneticgradients to interact with environmental gradients.
Freshwater Science jnbs-31-04-20.3d 7/10/12 08:10:49 1244 Cust # 12-029R1
1 E-mail address: [email protected] Present address: Department of Biological Sciences,
California State University Stanislaus, Turlock, California95382 USA. E-mail: [email protected]
3 E-mail address: [email protected]
Freshwater Science, 2012, 31(4):1244–1253’ 2012 by The Society for Freshwater ScienceDOI: 10.1899/12-029.1Published online: 16 October 2012
1244

Research into the effects of plant genetic variationon community dynamics and ecosystem processes hasdemonstrated the strong organizing power of genes atthe ecosystem scale (see reviews by Whitham et al.2003, 2006, 2008, Hughes et al. 2008, Bailey et al. 2009).In studies with a variety of model plant species(including riparian cottonwood, aspen, and birch),litter chemistry varied significantly among genotypeswithin species, and this variation in litter chemistryhad extended effects on ecosystem processes, suchas terrestrial decomposition and nutrient cycling(Schweitzer et al. 2004, 2005, Madritch et al. 2006,2009, Silfver et al. 2007). These influences of plantgenetics on ecosystem processes are equally strongwhen examined in nearby aquatic environmentsdependent on allochthonous inputs of plant material(LeRoy et al. 2006, 2007, Whitham et al. 2006, Lecerfand Chauvet 2008, Bailey et al. 2009), although theinfluences may diminish across spatial and temporalscales according to the Genetic Diffusion Hypothesis(LeRoy et al. 2006). Nevertheless, the influence ofwithin-species genetic variation is often on par withthe influence of variation among species. For example,variation in decomposition of litter from withinPopulus species can be as great as variation acrossmajor plant families (LeRoy et al. 2007).
In addition to genetic variation, resource (e.g.,nutrients, light) availability is a major determinantof plant chemical composition within and amongspecies (Coley et al. 1985, Cunningham et al. 1999).Direct manipulation of soil nutrient availability viafertilizer supplementation can strongly influence thechemical phenotypes of plants (Kyto et al. 1996,Lindroth et al. 2002) and associated ecosystemprocesses (Madritch et al. 2006).
Previous work with our experimental system(trembling aspen, Populus tremuloides Michx.) hasdemonstrated strong genotype, environment, andgenotype 3 environment effects on litter chemistry.Trees grown in low-nutrient environments havelower foliar quality (lower N, higher condensedtannins), resulting in lower defoliation rates, reducedherbivore growth rates (Osier and Lindroth 2001,Donaldson and Lindroth 2007), and reduced terres-trial decomposition (Madritch et al. 2006) compared totrees grown in high-nutrient environments. Wepredict that low foliar N and high foliar condensedtannin concentrations may similarly affect coloniza-tion by aquatic bacteria, fungi, and shreddinginvertebrates, thereby decelerating decomposition indetrital food webs in streams. In one other recentstudy of microbial diversity on decomposing leaves(but not decomposition itself), genotype, environ-ment, and genotype 3 environment effects influenced
fungal richness on leaves, but environmental effectson other microbial measures were typically strongerthan genotypic effects (Marks et al. 2009).
We experimentally manipulated soil nutrient envi-ronment for 5 aspen genotypes to evaluate howgenotype, environment, and genotype 3 environmenteffects on litter chemical composition influence ratesof allochthonous litter decomposition in an aquaticecosystem. We hypothesized that: 1) aspen genotypicvariation would influence litter chemistry, leading todifferential rates of in-stream decomposition, 2) soilnutrient (N, P, and K) amendment would increaselitter quality and accelerate in-stream decomposition,3) the nutrient treatment would differentially affectgenotypes leading to significant genotype 3 environ-ment interactions for both litter chemistry and in-stream decomposition, and 4) the nutrient environ-ment effect would influence these variables through-out the decomposition process, whereas the genotypeeffect would become weaker through time as sug-gested by the Genetic Diffusion Hypothesis (LeRoyet al. 2006).
Methods
Study design
To differentiate the effects of genotypic andenvironmental factors on phenotypic traits thatprobably influence aquatic decomposition, we grew5 aspen genotypes in a common garden under 2 soilnutrient regimes. The 5 genotypes used in our studywere collected from naturally occurring aspen clonesin southern Wisconsin, USA: Dan1, Dan2 (DaneCounty), Sau3 (Sauk County), Wau1 (WausharaCounty), and PI12 (Pine Island Wildlife Area, Colum-bia County). These genotypes are distinct (confirmedby microsatellite analysis of 16 loci, C. T. Cole,Department of Biology, University of Minnesota–Morris, MN and RLL, unpublished data) and werepropagated by micropropagation techniques (Sellmeret al. 1989, Donaldson and Lindroth 2004), plantedindividually into 40-L pots containing a 60:40 sand:siltloam mixture (see Madritch et al. 2006 for additionaldetails), and grown outdoors. Half of the trees (n = 5for each genotype) were allocated to the high-nutrienttreatment and supplemented with fertilizer (Osmo-cote 18:6:12 NPK 8–9-mo slow-release fertilizer,4.5 g/L soil, applied once/y). Remaining treesreceived 0.5 g/L fertilizer (low-nutrient treatment).Roots had minimal soil volume in which to forage fornutrients because the trees were grown in pots.Fertilizer was applied at rates determined empirically(Hemming and Lindroth 1999) to facilitate slow andfast growth rates and to produce foliage with a range
Freshwater Science jnbs-31-04-20.3d 7/10/12 08:10:50 1245 Cust # 12-029R1
2012] IN-STREAM GENOTYPE–ENVIRONMENT INTERACTIONS 1245

of chemical composition similar to that of trees grownnaturally (Donaldson and Lindroth 2007) and under Ndeposition conditions found in aspen forests (Kochyand Wilson 2001). We collected litter in autumn 2005after the trees had completed 2 seasons of growthunder the nutrient treatments. Prior to leaffall, wewrapped each tree with 1-cm mesh to capture leaves asthey abscised. Whole-tree leaf collections continueduntil the end of leaffall in October 2005. We removedleaves regularly from the mesh bags, placed them inpaper bags, and air-dried them.
Litter decomposition took place in Otter Creek (lat43u239N, long 89u479W), in Baxter’s Hollow NatureConservancy Preserve (central Sauk County, Wiscon-sin, USA). Otter Creek is a small (3 m wide onaverage), 2nd-order stream that drains a watershed of,2100 ha. Common woody riparian vegetation at thelocation includes aspen (P. tremuloides), oaks (Quercusrubra L., Quercus alba L.), yellow birch (Betulaalleghaniensis Britton), green ash (Fraxinus pennsylva-nica Marsh.), shagbark hickory (Carya ovata (Mill.) K.Koch), sugar maple (Acer saccharum Marsh.), andwhite pine (Pinus strobus L.). We did not measurewater-chemistry variables during this study. Werecognize that this constraint limits our ability togeneralize our findings to similar systems.
Litter decomposition
We mixed litter collections for each tree thoroughly(keeping litter from replicate trees separate) andpacked ,4 g of air-dried leaf litter from each replicategenotype into litter bags constructed from 6.4-mmdiamond-shaped mesh (TricalH netting, Aquatic Eco-Systems, Apopka, Florida). We allocated litter fromeach of the 5 replicate trees per genotype 3 environ-ment combination to 5 litter bags to provide forsequential retrieval of litter over 5 collection dates(5 genotypes 3 2 nutrient treatments 3 5 trees 3 5collection periods = 250 litter bags). We attached litterbags to randomly chosen 3.1-m lengths of steel rebarwith color-coded cable ties and placed them on thestream bed at a depth of 0.6 to 0.7 m. We placed therebars in the stream in an area of active litterdeposition, ,3 m 3 6 m, to minimize differences indecomposition among litter bags caused by streamflow, leaf deposition, or other environmental factors.On each collection date, we cut litter bags (1/replicatetree) from the rebar and returned them to thelaboratory for processing. We collected litter bags after7, 15, 29, 57, and 112 d in the stream. Twenty-three bagswere not available for analysis because they were lostin the stream or destroyed by ice, leaving a total of227 litter bags for collection and analysis.
In the laboratory, we rinsed sediment and inverte-brates from the leaves, which were then placed inpaper sacks and air-dried. Once dried, we groundleaves in a Wiley mill (US standard no. 40 meshscreen, 0.425 mm) and vacuum-dried the leaf powder.We subsampled dried, ground leaf material forchemical analysis. We also combusted leaf powderat 500uC for 3 h to measure ash-free dry mass (AFDM)of the remaining fraction for initial litter andsubsamples of litter from each collection date. Wecalculated decomposition rate constants (k) by re-gressing ln(AFDM) against time (Jenny et al. 1949).
Litter chemistry
We conducted chemical analyses on initial leaf litter(before stream deployment) and on decomposedleaves from each collection date. Examination oftemporal trends in litter chemical traits is importantbecause some compounds (e.g., soluble condensedtannins) can be leached from litter material, whereasothers (e.g., N and P) can accumulate in litter tissuesthrough microbial immobilization or become a largerproportion of litter mass remaining (e.g., fiber andlignin) because of differential loss of individualconstituents through time. We measured N, C, aciddetergent fiber (ADF), acid detergent lignin (ADL),soluble condensed tannins (SCT), and bound con-densed tannins (BCT) on all initial samples. We alsomeasured ADF, ADL, and BCT on each sample afterincubation in the stream. We did not measure P, andwe did not measure initial N levels for genotype Wau1 or for litter of any genotypes beyond day 7 of thedecomposition process. Most investigators who reporttannin concentrations report SCT values, but thesetannins leach rapidly from litter immersed in water,so we did not measure SCT after day 7. Instead, wemeasured BCT in initial samples and in all samplesthrough time.
We measured initial litter C and N by combustionin a Thermo Finnigan CNS Analyzer (ThermoFinnigan, San Jose, California) with atropine as theN standard. We measured ADF and ADL with amodified gravimetric method and an ANKOM 200fiber analyzer (AOAC 2000; ANKOM Technologies,Macedon, New York). We calculated %ADL based onthe initial dry mass of each sample. We extractedsoluble condensed tannins from leaf material with70% acetone:30% water (with 10 mM ascorbic acid).BCTs are not extractable because they remain boundto cell-wall constituents, but we solubilized BCTs byhydrolyzing the leaf tissue after SCT extraction. Weassayed both SCT and BCT with a modified butanol-HCl method (Porter et al. 1986) with purified aspen
Freshwater Science jnbs-31-04-20.3d 7/10/12 08:10:50 1246 Cust # 12-029R1
1246 C. J. LEROY ET AL. [Volume 31

tannin as the standard (Hagerman and Butler 1989).We quantified concentrations of SCT and BCT spec-trophotometrically (550 nm, SpectraMax-Plus 384;Molecular Devices, Sunnyvale, California).
Statistical analyses
We ran all statistical analyses in JMP (version 9.0.2;SAS Institute, Cary, North Carolina). We comparedinitial leaf-litter chemistry among genotypes, betweennutrient environments, and analyzed the genotype 3
environment interaction with 2-way analysis ofvariance (ANOVA). We selected genotypes to providea range of chemical compositions, so we denotedgenotype as a fixed factor in ANOVA. We constructeda correlation matrix to identify relationships amonglitter traits (Pearson’s correlation coefficient, r at a =
0.05). We reported litter chemistry values as % drymass, so we arcsin!(x)-transformed these valuesbefore statistical analysis. We used Tukey’s HonestlySignificant Difference (HSD) as a post hoc test toidentify differences among genotypes followingANOVA.
In studies based on bulk litter, unique estimatesof k cannot be made for each replicate for use insubsequent analyses. Instead, all replicates are used tocalculate 1 k-value per treatment. However, wepacked leaf litter from each replicate genotype 3
environment treatment combination separately intoindividually labeled litter bags. This experimentaldesign enabled us to calculate 5 independent k-valuesper treatment. We used a 2-way ANOVA withgenotype, environment, and genotype 3 environmenteffects to analyze these estimates of k. All exponentialregressions (with slopes k/d) were significant at p ,
0.05. We used simple linear regression to assessrelationships between k and all litter chemistryvariables because multiple estimates of k werecalculated per treatment.
We examined patterns in litter chemical constitu-ents and mass remaining through time with 2-wayanalysis of covariance (ANCOVA) models to deter-mine the effects of genotype, environment, collectiondate (covariate), and all possible interactions (includ-ing the genotype 3 environment interaction). Wereported all values as percentages (% chemicalconstituents and % mass remaining), so we arc-sin!(x)-transformed them before analysis. We usedANOVAs with random effects (genotype, environ-ment, and genotype 3 environment) to assesschanges in mass remaining and litter chemistry(ADF, ADL, BCT) at each harvesting date to estimatethe proportion of variation explained by each effect inrelation to the residual variation through time.
Results
Initial litter chemistry
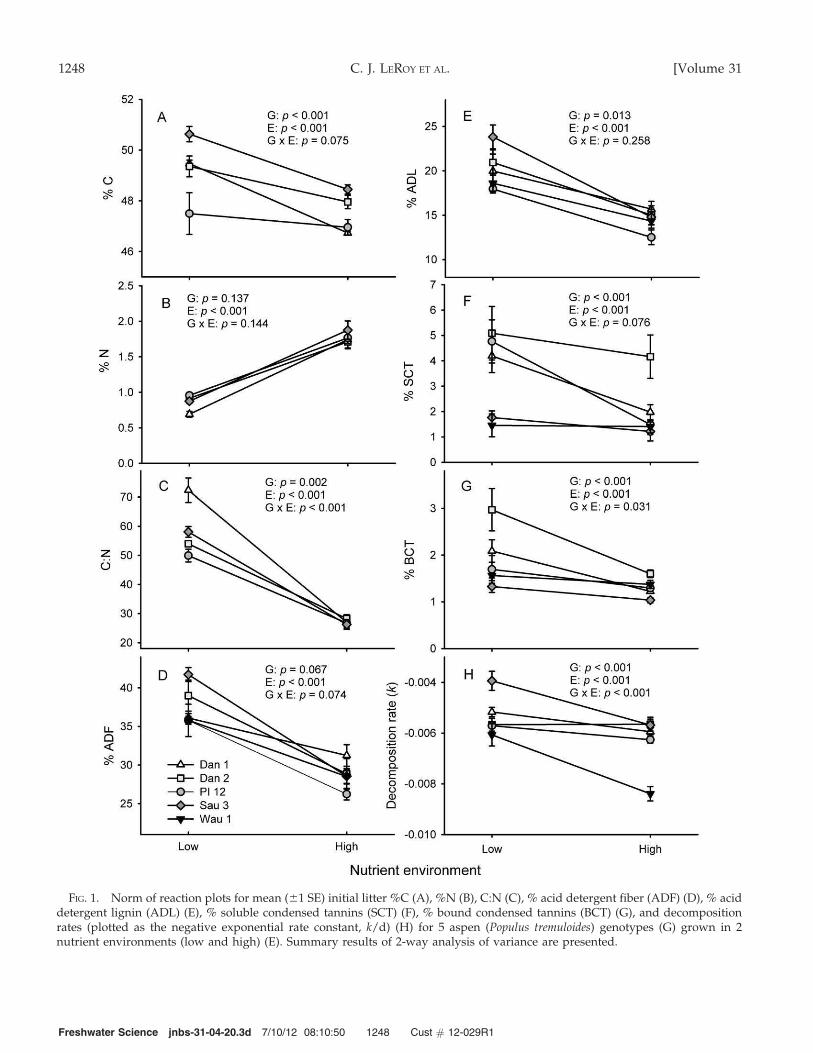
Initial concentrations of litter chemical componentsvaried among aspen genotypes, nutrient environ-ments, or both (genotype 3 environment interaction;Fig. 1A–H). Leaf-litter C, C:N, ADL, SCT, and BCTcontent differed among aspen genotypes, whereasADF differed only marginally among genotypes (p =
0.067). N did not differ among genotypes (p = 0.137).In contrast, nutrient environment influenced all litterchemical components (p , 0.001). Litter C wassomewhat lower for high- than for low-nutrient-environment trees (Fig. 1A), but N was nearly 23
higher in leaves from the high- than the low-nutrientenvironment (Fig. 1B), causing litter C:N ratios to besignificantly lower for trees grown in the high- than inthe low-nutrient environment. Litter in the low-nutrient environment showed greater variability inC:N than did litter in the high-nutrient environment,resulting in a significant genotype 3 environmentinteraction (Fig. 1C). ADF and ADL content weresignificantly higher in litter from the low- than thehigh-nutrient environment and differences wereconsistent across genotypes (no genotype 3 environ-ment interactions; Fig. 1D, E, respectively). SCT andBCT content tended to be higher in the low- than inthe high-nutrient environment (Fig. 1F, G, respective-ly), but genotypes responded differently to nutrientenvironment (marginally significant and significantgenotype 3 environment interactions for SCT andBCT, respectively). These litter variables were allnegatively correlated with N, and content of moststructural and phenolic compounds were positivelycorrelated with one another (Table 1).
Litter decomposition
Values of k differed among genotypes of aspen (p ,
0.001; Fig. 1H), and high-nutrient litter tended todecompose faster than low-nutrient litter (p , 0.001).However, the response of k to nutrients differedamong genotypes (significant genotype 3 environ-ment interaction; p , 0.001). Specifically, the Wau1genotype decomposed fastest, whereas Sau3 andDan1 decomposed slowest. Overall, k-values wereup to 1.53 higher for high- than for low-nutrient litteracross all genotypes (Table 2). The genotype 3
environment interaction reflected the fact that k-values for genotypes Dan1, Dan2, and PI12 werenot influenced by the nutrient environment, whereask-values for genotypes Sau3 and Wau1 were signifi-cantly faster in high- than in low-nutrient environ-ments (Fig. 1H).
Freshwater Science jnbs-31-04-20.3d 7/10/12 08:10:50 1247 Cust # 12-029R1
2012] IN-STREAM GENOTYPE–ENVIRONMENT INTERACTIONS 1247
Freshwater Science jnbs-31-04-20.3d 7/10/12 08:10:50 1248 Cust # 12-029R1
FIG. 1. Norm of reaction plots for mean (61 SE) initial litter %C (A), %N (B), C:N (C), % acid detergent fiber (ADF) (D), % aciddetergent lignin (ADL) (E), % soluble condensed tannins (SCT) (F), % bound condensed tannins (BCT) (G), and decompositionrates (plotted as the negative exponential rate constant, k/d) (H) for 5 aspen (Populus tremuloides) genotypes (G) grown in 2nutrient environments (low and high) (E). Summary results of 2-way analysis of variance are presented.
1248 C. J. LEROY ET AL. [Volume 31
Correlations with decomposition
In general, we found weak linear relationshipsbetween k-values and initial leaf-litter chemistry andlitter chemistry measured through time. Values of kwere negatively related to initial concentrations ofADF (r = 0.346, p = 0.049), but not C, N, C:N, ADL,SCT, or BCT (p . 0.140). Values of k decreased as litterADL increased over time (R2
= 0.100, F(1,43) = 4.79,p = 0.034), but k was not linearly related to ADF (R2
=
0.000, F(1,43) = 0.001, p = 0.968) or BCT (R2= 0.029,
F(1,43) = 1.27, p = 0.265) over time. N, C, and SCT werenot measured through time.
Patterns through time
Examination of losses of leaf structural compounds(ADF and ADL), BCT, and overall leaf mass at eachharvesting date could elucidate patterns and provideinsight into the relative importance of genetic orenvironmental factors at various times in the decom-position process. A full factorial ANCOVA revealedthat genotype, environment, and time significantlyinfluenced all 3 chemical constituents and litter massremaining (Table 3). The genotype 3 environmentinteraction affected BCT and mass remaining, but inboth cases, genotype also interacted with time. N
content was measured only for initial litter and onday 7, but was affected by genotype (p , 0.001),environment (p , 0.001), and the genotype 3
environment interaction (p , 0.001).
Variation explained by genotype, environment, andgenotype 3 environment effects
Variations in ADL, BCT, and litter mass remainingwere differentially explained by genotype, environ-ment, and the genotype 3 environment interaction.Genotype explained little of the variation in ADL earlyin the study, but explained nearly 30% of the variationlater in the study (Fig. 2A). In contrast, nearly 60% ofthe variation in BCT was explained by genotype earlyin the study (Fig. 2B), and very little of the variation inmass remaining was explained by genotype until latein the decomposition process when genotype ex-plained almost 30% of the variation in mass remaining(Fig. 2C). Environment had a much stronger influencethan genotype on ADL and mass remaining (rangingfrom 10–65% and 35–70%, respectively) throughout thestudy but explained very little of the variation in BCTat any point in the study (0–10%). The proportion of thevariation explained by the genotype 3 environmentinteraction was relatively small for both mass remain-ing and ADL, but the genotype 3 environmentinteraction explained almost 40% of the variation inBCT at intermediate stages of decay.
Discussion
Ours is the first study to examine genotype 3
environment interactions as determinants of leaf-litterdecomposition rates in aquatic systems. Our resultssuggest that these effects on decomposition in streamsmight be common, based on the degree of plantvariation controlled by both genetic and environmentalfactors (Whitham et al. 2006, Hughes et al. 2008) andthe degree of connectedness between forest and streamsystems. In our study, the environment effect (nutrientenrichment) was applied in the terrestrial environ-ment and showed ‘‘afterlife’’ effects (Choudhury 1988,
Freshwater Science jnbs-31-04-20.3d 7/10/12 08:11:06 1249 Cust # 12-029R1
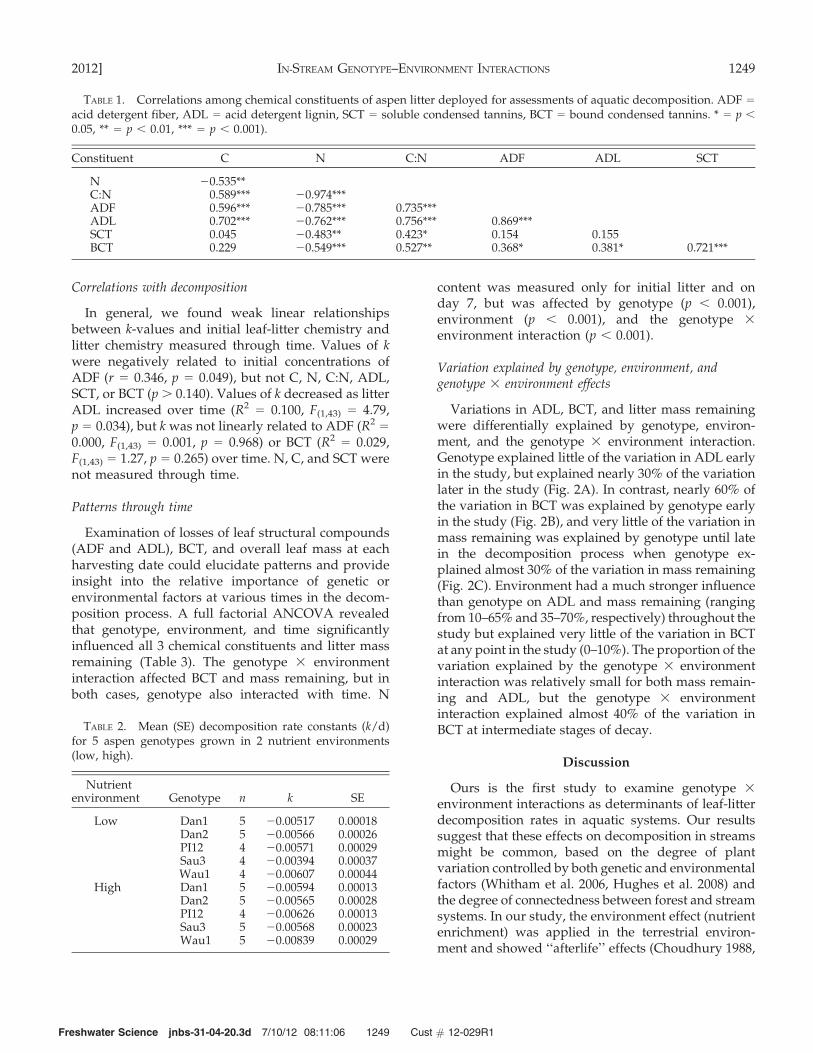
TABLE 1. Correlations among chemical constituents of aspen litter deployed for assessments of aquatic decomposition. ADF =
acid detergent fiber, ADL = acid detergent lignin, SCT = soluble condensed tannins, BCT = bound condensed tannins. * = p ,
0.05, ** = p , 0.01, *** = p , 0.001).
Constituent C N C:N ADF ADL SCT
N 20.535**C:N 0.589*** 20.974***ADF 0.596*** 20.785*** 0.735***ADL 0.702*** 20.762*** 0.756*** 0.869***SCT 0.045 20.483** 0.423* 0.154 0.155BCT 0.229 20.549*** 0.527** 0.368* 0.381* 0.721***
TABLE 2. Mean (SE) decomposition rate constants (k/d)for 5 aspen genotypes grown in 2 nutrient environments(low, high).
Nutrientenvironment Genotype n k SE
Low Dan1 5 20.00517 0.00018Dan2 5 20.00566 0.00026PI12 4 20.00571 0.00029Sau3 4 20.00394 0.00037Wau1 4 20.00607 0.00044
High Dan1 5 20.00594 0.00013Dan2 5 20.00565 0.00028PI12 4 20.00626 0.00013Sau3 5 20.00568 0.00023Wau1 5 20.00839 0.00029
2012] IN-STREAM GENOTYPE–ENVIRONMENT INTERACTIONS 1249
Findlay et al. 1996) mediated by plant litterfall into anaquatic environment.
Aspen genotype, nutrient environment, and geno-type 3 environment interactions influenced leaf-litterchemistry after abscission and leaf-litter decomposi-tion. Nutrient environment tended to influence earlydecomposition more strongly than late decomposi-tion, whereas aspen genotype explained more varia-tion in mass remaining later than earlier in theprocess. Patterns in both litter mass remaining andlitter chemistry through time were differentiallyinfluenced by genotype, environment, and genotype3 environment interactions. Our results are similar insome respects to those of an analogous terrestrialstudy of decomposition on a forest floor (Madritch etal. 2006), but show some significant deviations interms of the strength of the influence of genotype,environment, and genotype 3 environment interac-tion effects on litter decomposition in an adjacentstream.
Detritus is processed differently in aquatic than interrestrial systems. Factors, such as abrasion, frag-mentation, leaching, and shredding by invertebrates,that are thought to be less influential in terrestrialsystems can be major drivers in aquatic systems.Despite these potential ecosystem-level differences inlitter processing, we still saw very strong effects ofgenotype, environment, and genotype 3 environmentinteractions on in-stream leaf-litter decomposition,highlighting the potential for terrestrial nutrientdeposition to influence stream litter processing. Infact, we found a stronger genotype 3 environmentinteraction for aquatic leaf-litter decomposition thanMadritch et al. (2006) found in their terrestrial study.Madritch et al. (2006) reported a marked increase indecomposition rate for all aspen genotypes that hadbeen exposed to the high-nutrient treatment prior toabscission. In contrast, we found that decompositionrates of 3 genotypes (Dan1, Dan2 and PI12) were not
significantly influenced by nutrient enrichment (oronly marginally so), whereas the decomposition ratesof genotypes Sau3 and Wau1 were significantlyaccelerated.
Strong genotype 3 environment interactions sug-gest that environmental gradients may interact withgenetic gradients to yield unique and unpredictableoutcomes in terms of ecosystem function undercurrent and future conditions. Under the GeneticDiffusion Hypothesis, genetic effects are predicted tobe strong in situations where the spatial and temporalextent of genetic variation is small, but our resultsindicate that genetic effects on chemical constituentslike BCT can be maintained throughout the decom-position process. As riparian ecosystems continue tobe developed and managed by humans (Poff et al.2011), the need to understand genotype 3 environ-ment interactions will become increasingly important.
Genetic variation within tree species can be veryhigh, and aspen is a highly variable species (Farmer1996). The variation in decomposition rates found inour study rivals that shown in other studies ofvariation among genotypes of 2 species of cottonwood(Populus fremontii and Populus angustifolia), and acrossmany plant families (LeRoy et al. 2007). In fact, therange in decomposition rates in our study was 0.006 kunits (0.003–0.009/d). This range exceeds the pub-lished ranges for other species of Populus and evenhybrid complexes (LeRoy et al. 2007). This variation inaquatic decomposition rates also exceeds the variationobserved in terrestrial decomposition for the samegenotypes (0.004 k units; 0.0014–0.0054/d; valueswere reported /y by Madritch et al. 2006). Thesecomparisons reveal a large potential for within- andamong-population variation to influence the ecosys-tem function of leaf-litter decomposition, even inadjacent environments. Variation among replicatestypically increases through time in decompositionstudies, so the variation in decomposition rates found
Freshwater Science jnbs-31-04-20.3d 7/10/12 08:11:06 1250 Cust # 12-029R1
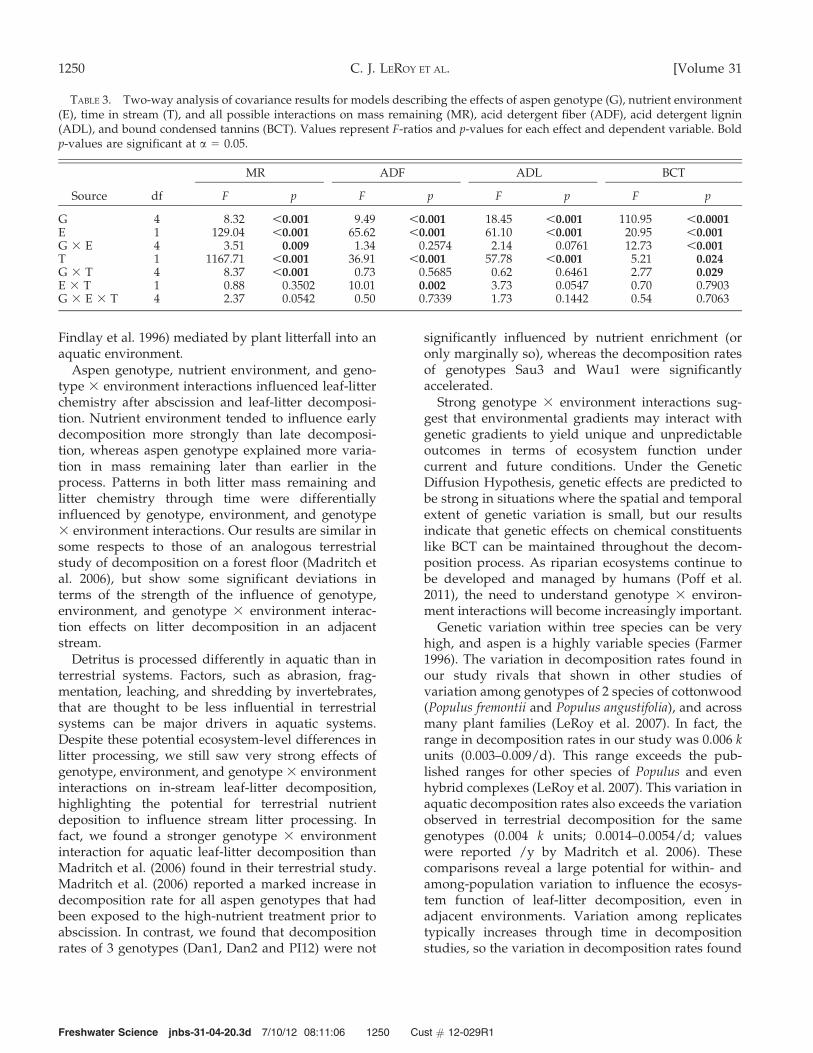
TABLE 3. Two-way analysis of covariance results for models describing the effects of aspen genotype (G), nutrient environment(E), time in stream (T), and all possible interactions on mass remaining (MR), acid detergent fiber (ADF), acid detergent lignin(ADL), and bound condensed tannins (BCT). Values represent F-ratios and p-values for each effect and dependent variable. Boldp-values are significant at a = 0.05.
Source df
MR ADF ADL BCT
F p F p F p F p
G 4 8.32 ,0.001 9.49 ,0.001 18.45 ,0.001 110.95 ,0.0001E 1 129.04 ,0.001 65.62 ,0.001 61.10 ,0.001 20.95 ,0.001G 3 E 4 3.51 0.009 1.34 0.2574 2.14 0.0761 12.73 ,0.001T 1 1167.71 ,0.001 36.91 ,0.001 57.78 ,0.001 5.21 0.024G 3 T 4 8.37 ,0.001 0.73 0.5685 0.62 0.6461 2.77 0.029E 3 T 1 0.88 0.3502 10.01 0.002 3.73 0.0547 0.70 0.7903G 3 E 3 T 4 2.37 0.0542 0.50 0.7339 1.73 0.1442 0.54 0.7063
1250 C. J. LEROY ET AL. [Volume 31
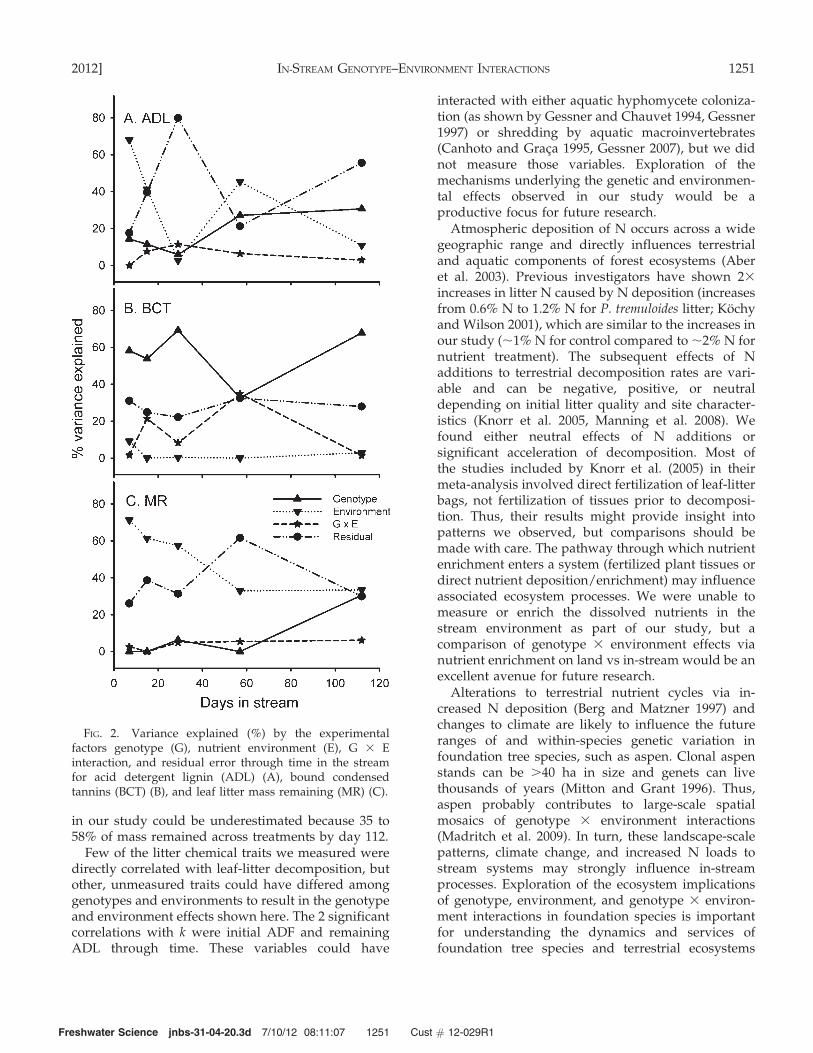
in our study could be underestimated because 35 to58% of mass remained across treatments by day 112.
Few of the litter chemical traits we measured weredirectly correlated with leaf-litter decomposition, butother, unmeasured traits could have differed amonggenotypes and environments to result in the genotypeand environment effects shown here. The 2 significantcorrelations with k were initial ADF and remainingADL through time. These variables could have
interacted with either aquatic hyphomycete coloniza-tion (as shown by Gessner and Chauvet 1994, Gessner1997) or shredding by aquatic macroinvertebrates(Canhoto and Graca 1995, Gessner 2007), but we didnot measure those variables. Exploration of themechanisms underlying the genetic and environmen-tal effects observed in our study would be aproductive focus for future research.
Atmospheric deposition of N occurs across a widegeographic range and directly influences terrestrialand aquatic components of forest ecosystems (Aberet al. 2003). Previous investigators have shown 23
increases in litter N caused by N deposition (increasesfrom 0.6% N to 1.2% N for P. tremuloides litter; Kochyand Wilson 2001), which are similar to the increases inour study (,1% N for control compared to ,2% N fornutrient treatment). The subsequent effects of Nadditions to terrestrial decomposition rates are vari-able and can be negative, positive, or neutraldepending on initial litter quality and site character-istics (Knorr et al. 2005, Manning et al. 2008). Wefound either neutral effects of N additions orsignificant acceleration of decomposition. Most ofthe studies included by Knorr et al. (2005) in theirmeta-analysis involved direct fertilization of leaf-litterbags, not fertilization of tissues prior to decomposi-tion. Thus, their results might provide insight intopatterns we observed, but comparisons should bemade with care. The pathway through which nutrientenrichment enters a system (fertilized plant tissues ordirect nutrient deposition/enrichment) may influenceassociated ecosystem processes. We were unable tomeasure or enrich the dissolved nutrients in thestream environment as part of our study, but acomparison of genotype 3 environment effects vianutrient enrichment on land vs in-stream would be anexcellent avenue for future research.
Alterations to terrestrial nutrient cycles via in-creased N deposition (Berg and Matzner 1997) andchanges to climate are likely to influence the futureranges of and within-species genetic variation infoundation tree species, such as aspen. Clonal aspenstands can be .40 ha in size and genets can livethousands of years (Mitton and Grant 1996). Thus,aspen probably contributes to large-scale spatialmosaics of genotype 3 environment interactions(Madritch et al. 2009). In turn, these landscape-scalepatterns, climate change, and increased N loads tostream systems may strongly influence in-streamprocesses. Exploration of the ecosystem implicationsof genotype, environment, and genotype 3 environ-ment interactions in foundation species is importantfor understanding the dynamics and services offoundation tree species and terrestrial ecosystems
Freshwater Science jnbs-31-04-20.3d 7/10/12 08:11:07 1251 Cust # 12-029R1
FIG. 2. Variance explained (%) by the experimentalfactors genotype (G), nutrient environment (E), G 3 Einteraction, and residual error through time in the streamfor acid detergent lignin (ADL) (A), bound condensedtannins (BCT) (B), and leaf litter mass remaining (MR) (C).
2012] IN-STREAM GENOTYPE–ENVIRONMENT INTERACTIONS 1251

and the aquatic ecosystems to which they are coupledby material transfer.
Acknowledgements
All authors contributed significantly to the studydesign, and the manuscript was written as a collabo-rative effort. Research funding was secured by RLL.Data collection was completed by SCW and dataanalysis was performed by CJL. We thank ErikJohnsen, Adam Gusse, Cassie Mescher, and SarahHopkins for help preparing, retrieving, and analyzingsamples. Mike Madritch and Tim Meehan providedhelpful discussion regarding the experimental designand analysis. We thank Nick Miller and the NatureConservancy for permission to conduct research in theBaxter’s Hollow Nature Conservancy Preserve. Thisresearch was supported by National Science Founda-tion grants DEB-0344019, DEB-0425908, and DEB-0841609. We thank Associate Editor Manuel Graca,Editor Pamela Silver, and 2 anonymous referees forfeedback that significantly improved this manuscript.
Literature Cited
ABER, J. D., C. L. GOODALE, S. V. OLLINGER, M.-L. SMITH, A. H.MAGILL, M. E. MARTIN, R. A. HALLETT, AND J. L. STODDARD.2003. Is nitrogen deposition altering the nitrogen statusof northeastern forests? BioScience 53:375–389.
BAILEY, J. K., J. A. SCHWEITZER, F. UBEDA, J. KORICHEVA, C. J.LEROY, M. D. MADRITCH, B. J. REHILL, R. K. BANGERT, D. G.FISCHER, G. J. ALLAN, AND T. G. WHITHAM. 2009. Fromgenes to ecosystems: a synthesis of the effects of plantgenetic factors across levels of organization. Philosoph-ical Transactions of the Royal Society Series B: BiologicalSciences 364:1607–1616.
BERG, B., AND E. MATZNER. 1997. Effect of N deposition ondecomposition of plant litter and soil organic matter inforest systems. Environmental Reviews 5:1–25.
CANHOTO, C., AND M. A. S. GRACA. 1995. Food value ofintroduced eucalypt leaves for a Mediterranean streamdetritivore: Tipula lateralis. Freshwater Biology 34:209–214.
CARREIRO, M. M., R. L. SINSABAUGH, D. A. REPERT, AND D. F.PARKHURST. 2000. Microbial enzyme shifts explain litterdecay responses to simulated nitrogen deposition.Ecology 81:2359–2365.
CHOUDHURY, D. 1988. Herbivore induced changes in leaf-litter resource quality: a neglected aspect of herbivory inecosystem nutrient dynamics. Oikos 51:389–393.
COLEY, P. D., J. P. BRYANT, AND F. S. CHAPIN. 1985. Resourceavailability and plant antiherbivore defense. Science230:895–899.
CUNNINGHAM, S. A., B. SUMMERHAYES, AND M. WESTOBY. 1999.Evolutionary divergences in leaf structure and chemis-try, comparing rainfall and soil nutrient gradients.Ecological Monographs 69:569–588.
DONALDSON, J. R., AND R. L. LINDROTH. 2004. Cottonwood leafbeetle (Coleoptera: Chrysomelidae) performance inrelation to variable phytochemistry in juvenile aspen(Populus tremuloides Michx.). Environmental Entomolo-gy 33:1505–1511.
DONALDSON, J. R., AND R. L. LINDROTH. 2007. Genetics,environment, and their interaction determine efficacyof chemical defense in trembling aspen. Ecology 88:729–739.
FARMER, R. E. 1996. The genecology of Populus. Pages 33–56in R. F. Stettler, H. D. Bradshaw, P. E. Heilman, andT. M. Hinckley (editors). Biology of Populus and itsimplications for management and conservation. Nation-al Research Council of Canada, Ottawa, Ontario.
FINDLAY, S., M. CARREIRO, V. KRISCHIK, AND C. G. JONES. 1996.Effects of damage to living plants on leaf litter quality.Ecological Applications 6:269–275.
FOG, K. 1988. The effect of added nitrogen on the rate ofdecomposition of organic matter. Biological Reviews 63:433–462.
GESSNER, M. O. 1997. Fungal biomass, production andsporulation associated with particulate organic matterin streams. Limnetica 13:33–44.
GESSNER, M. O. 2007. Proximate lignin and cellulose.Pages 115–120 in M. A. S. Graca, F. Barlocher, and M. O.Gessner (editors). Methods to study decomposition: apractical guide. Springer, Dordrecht, The Netherlands.
GESSNER, M. O., AND E. CHAUVET. 1994. Importance of streammicrofungi in controlling breakdown rates of leaf litter.Ecology 75:1807–1817.
HAGERMAN, A. E., AND L. G. BUTLER. 1989. Choosingappropriate methods and standards for assaying tannin.Journal of Chemical Ecology 15:1795–1810.
HEMMING, J. D. C., AND R. L. LINDROTH. 1999. Effects of lightand nutrient availability on aspen: growth, phytochem-istry, and insect performance. Journal of ChemicalEcology 25:1687–1714.
HOBBIE, S. E. 2000. Interactions between litter lignin and soilnitrogen availability during leaf litter decomposition ina Hawaiian montane forest. Ecosystems 3:484–494.
HUGHES, A. R., B. D. INOUYE, M. T. J. JOHNSON, N. UNDERWOOD,AND M. VELLEND. 2008. Ecological consequences ofgenetic diversity. Ecology Letters 11:609–623.
JENNY, H., S. GESSEL, AND F. BINGHAM. 1949. Comparativestudy of decomposition rates of organic matter intemperate and tropical regions. Soil Science 68:419–432.
KNORR, M., S. D. FREY, AND P. S. CURTIS. 2005. Nitrogenadditions and litter decomposition: a meta-analysis.Ecology 86:3252–3257.
KOCHY, M., AND S. D. WILSON. 2001. Nitrogen deposition andforest expansion in the northern Great Plains. Journal ofEcology 89:807–817.
KYTO, M., P. NIEMELA, AND S. LARSSON. 1996. Insects on trees:population and individual response to fertilization.Oikos 75:148–159.
LECERF, A., AND E. CHAUVET. 2008. Intraspecific variability inleaf traits strongly affects alder leaf decomposition in astream. Basic and Applied Ecology 9:598–605.
Freshwater Science jnbs-31-04-20.3d 7/10/12 08:11:09 1252 Cust # 12-029R1
1252 C. J. LEROY ET AL. [Volume 31

LEDIG, F. T. 1992. Human impacts on genetic diversity inforest ecosystems. Oikos 63:87–108.
LEROY, C. J., T. G. WHITHAM, P. KEIM, AND J. C. MARKS. 2006.Plant genes link forests and streams. Ecology 87:255–261.
LEROY, C. J., T. G. WHITHAM, S. C. WOOLEY, AND J. C. MARKS.2007. Within-species variation in foliar chemistryinfluences leaf-litter decomposition in a Utah river.Journal of the North American Benthological Society 26:426–438.
LINDROTH, R. L., T. L. OSIER, H. R. H. BARNHILL, AND S. A.WOOD. 2002. Effects of genotype and nutrient availabil-ity on phytochemistry of trembling aspen (Populustremuloides Michx.) during leaf senescence. BiochemicalSystematics and Ecology 30:297–307.
MADRITCH, M., J. R. DONALDSON, AND R. L. LINDROTH. 2006.Genetic identity of Populus tremuloides litter influencesdecomposition and nutrient release in a mixed foreststand. Ecosystems 9:528–537.
MADRITCH, M. D., S. L. GREENE, AND R. L. LINDROTH. 2009.Genetic mosaics of ecosystem functioning across aspen-dominated landscapes. Oecologia (Berlin) 160:119–127.
MAGILL, A. H., AND J. D. ABER. 1998. Long-term effects ofexperimental nitrogen additions on foliar litter decayand humus formation in forest ecosystems. Plant andSoil 203:301–311.
MANNING, P., M. SAUNDERS, R. D. BARDGETT, M. BONKOWSKI,M. A. BRADFORD, R. J. ELLIS, E. KANDELER, S. MARHAN, AND
D. TSCHERKO. 2008. Direct and indirect effects of nitrogendeposition on litter decomposition. Soil Biology andBiochemistry 40:688–698.
MARKS, J. C., G. A. HADEN, B. L. HARROP, E. G. REECE, J. L.KEAMS, M. E. WATWOOD, AND T. G. WHITHAM. 2009.Genetic and environmental controls of microbial com-munities on leaf litter in streams. Freshwater Biology 54:2616–2627.
MITTON, J. B., AND M. C. GRANT. 1996. Genetic variation andthe natural history of quaking aspen. BioScience 46:25–31.
NORBY, R. J. 1998. Nitrogen deposition: a component ofglobal change analyses. New Phytologist 139:189–200.
OSIER, T. L., AND R. L. LINDROTH. 2001. Effects of genotype,nutrient availability, and defoliation on aspen phyto-chemistry and insect performance. Journal of ChemicalEcology 27:1289–1313.
PETERSEN, R. C., AND K. W. CUMMINS. 1974. Leaf processing ina woodland stream. Freshwater Biology 4:343–368.
POFF, B., K. A. KOESTNER, D. G. NEARY, AND V. HENDERSON.2011. Threats to riparian ecosystems in western NorthAmerica: An analysis of existing literature. Journal
of the American Water Resources Association 47:1241–1254.
PORTER, L. J., L. N. HRSTICH, AND B. G. CHAN. 1986. Theconversion of procyanidins and prodelphinidins tocyanidin and delphinidin. Phytochemistry 25:223–230.
SCHWEITZER, J. A., J. K. BAILEY, S. C. HART, G. M. WIMP, S. K.CHAPMAN, AND T. G. WHITHAM. 2005. The interaction ofplant genotype and herbivory decelerate leaf litterdecomposition and alter nutrient dynamics. Oikos 110:133–145.
SCHWEITZER, J. A., J. K. BAILEY, B. J. REHILL, G. D. MARTINSEN,S. C. HART, R. L. LINDROTH, P. KEIM, AND T. G. WHITHAM.2004. Genetically based trait in a dominant tree affectsecosystem processes. Ecology Letters 7:127–134.
SELLMER, J. C., B. H. MCCOWN, AND B. E. HAISSIG. 1989. Shootculture dynamics of six Populus clones. Tree Physiology5:219–227.
SILFVER, T., J. MIKOLA, M. ROUSI, H. ROININEN, AND E. OKSANEN.2007. Leaf litter decomposition differs among genotypesin a local Betula pendula population. Oecologia (Berlin)152:707–714.
TANK, J. L., E. J. ROSI-MARSHALL, N. A. GRIFFITHS, S. A.ENTREKIN, AND M. L. STEPHEN. 2010. A review ofallochthonous organic matter dynamics and metabolismin streams. Journal of the North American BenthologicalSociety 29:118–146.
WEBSTER, J. R., AND E. F. BENFIELD. 1986. Vascular plantbreakdown in freshwater ecosystems. Annual Review ofEcology and Systematics 17:567–594.
WHITHAM, T. G., J. K. BAILEY, J. A. SCHWEITZER, S. M. SHUSTER,R. K. BANGERT, C. J. LEROY, E. V. LONSDORF, G. J. ALLAN,S. P. DIFAZIO, B. M. POTTS, D. G. FISCHER, C. A. GEHRING,R. L. LINDROTH, J. C. MARKS, S. C. HART, G. M. WIMP, AND
S. C. WOOLEY. 2006. A framework for community andecosystem genetics: from genes to ecosystems. NatureReviews Genetics 7:510–523.
WHITHAM, T. G., S. P. DIFAZIO, J. A. SCHWEITZER, S. M. SHUSTER,G. J. ALLAN, J. K. BAILEY, AND S. A. WOOLBRIGHT. 2008.Extending genomics to natural communities and eco-systems. Science 320:492–495.
WHITHAM, T. G., W. P. YOUNG, G. D. MARTINSEN, C. A. GEHRING, J.A. SCHWEITZER, S. M. SHUSTER, G. M. WIMP, D. G. FISCHER, J. K.BAILEY, R. L. LINDROTH, S. WOOLBRIGHT, AND C. R. KUSKE. 2003.Community and ecosystem genetics: a consequence of theextended phenotype. Ecology 84:559–573.
Received: 24 February 2012Accepted: 15 August 2012
Freshwater Science jnbs-31-04-20.3d 7/10/12 08:11:10 1253 Cust # 12-029R1
2012] IN-STREAM GENOTYPE–ENVIRONMENT INTERACTIONS 1253